黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47及其在抗瓜类疫病中的应用
技术领域
1.本发明属于植物分子生物学与植物遗传工程领域,具体涉及黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47及其在抗瓜类疫病中的应用。
背景技术:
2.瓜类疫病是一种世界性卵菌病害,病原菌为瓜类疫霉(phytophthora melonis),引起黄瓜、南瓜、冬瓜、苦瓜等瓜类叶、茎、果变褐致死或死苗。该病在我国种植瓜类地区均有发生。在北方以夏秋露地栽培及春季保护地瓜类发生较重,在南方则以春季及夏秋高温多湿的季节发生较重。瓜类整个生长期及各个部位均可受害,幼苗被害生长点及嫩茎基部呈水渍状缢缩,萎蔫死亡。成株期茎被害,基部和节部呈水渍状、黑褐色病斑,逐渐软化缢缩,叶片下垂、植株萎蔫、死亡;叶片多在近叶缘处出现暗绿色、边缘不清晰的大型病斑;瓜果在受害处呈现水渍状、近圆形的凹陷斑,迅速腐烂,表面长出白色或灰白色稀疏霉层。一旦发病,化学药剂控制效果不理想,造成经济损失。控制该病害的最根本途径是培育抗疫病瓜类新品种,要培育高抗疫病瓜类新品种,就需要挖掘瓜类疫病抗性基因并进行应用。
技术实现要素:
3.本发明的第一方面的目的,在于提供黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47。
4.本发明的第二方面的目的,在于提供编码黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47的核酸分子。
5.本发明的第三方面的目的,在于提供包含编码黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47 的核酸分子的载体、表达盒、转基因细胞系、重组菌或重组病毒。
6.本发明的第四方面的目的,在于提供黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47的应用。
7.本发明的第五方面的目的,在于提供编码黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47的核酸分子的应用。
8.本发明的第六方面的目的,在于提供包含编码黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47 的核酸分子的载体、表达盒、转基因细胞系、重组菌或重组病毒的应用。
9.本发明的第七方面的目的,在于提供靶向上调黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47 的表达量或增强黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47活性的试剂的应用。
10.本发明的第八方面的目的,在于提供一种培育抗瓜类疫病转基因植物的方法。
11.本发明的第九方面的目的,在于提供一种产品。
12.为了实现上述目的,本发明所采取的技术方案是:
13.本发明的第一个方面,提供一种黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47,所述黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47的氨基酸序列为:
14.(a)mglpwyrvhtvvlndpgrllsvhimhtalvagwagsmalyelavfdpsdpvldp mwrqdffhsfisgcfgfgafhvtglygpgiwvsdpygltgrvqavnpawgvsgfdpfvpgg iashhiaagtlgilaglfhlslrppqrlykglrmgnietvlsssidavffaafvvaepyhivpa ttpislnylgplarnisffecrvsrkekfl(seq id no.1);或
15.(b)seq id no.1所示的氨基酸序列经一个或多个氨基酸的取代、缺失或添加修饰后且功能与氨基酸序列如seq id no.1所示的蛋白相同或相似的氨基酸序列;
16.(c)与seq id no.1所示的氨基酸序列具有80%、85%、90%、95%、98%或99%以上同源性且功能与氨基酸序列如seq id no.1所示的蛋白相同或相似的氨基酸序列。
17.本发明的第二个方面,提供编码本发明第一方面的黄瓜光合系统ii反应中心蛋白cspsii
‑ꢀ
cp47的核酸分子。
18.优选地,所述核酸分子的序列为(d)~(g)中任一种:
19.(d)seq id no.2所示的核苷酸序列;
20.(e)seq id no.3所示的核苷酸序列;
21.(f)与seq id no.2所示的核苷酸序列具有80%、85%、90%、95%、98%或99%以上同源性且功能与核苷酸序列如seq id no.2所示的核酸分子相同或相似的核苷酸序列;
22.(g)与seq id no.3所示的核苷酸序列具有80%、85%、90%、95%、98%或99%以上同源性且功能与核苷酸序列如seq id no.3所示的核酸分子相同或相似的核苷酸序列。
23.本发明的第三个方面,提供包含本发明第二方面的核酸分子的载体、表达盒、转基因细胞系、重组菌或重组病毒。
24.优选地,所述转基因细胞系不包含繁殖材料。
25.本发明的第四个方面,提供本发明第一方面的黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47 在(1)~(3)中任一项中的应用;
26.(1)抗瓜类疫病;
27.(2)制备抗瓜类疫病的产品;
28.(3)培育抗瓜类疫病转基因植物。
29.优选地,所述瓜类疫病为瓜类疫霉菌(phytophthora melonis)引发的病害。
30.优选地,所述植物为黄瓜、南瓜、冬瓜、苦瓜、丝瓜和节瓜中的至少一种;进一步为黄瓜和南瓜中的至少一种。
31.本发明的第五个方面,提供本发明第二方面的核酸分子在(1)~(3)中任一项中的应用;
32.(1)抗瓜类疫病;
33.(2)制备抗瓜类疫病的产品;
34.(3)培育抗瓜类疫病转基因植物。
35.优选地,所述瓜类疫病为瓜类疫霉菌(phytophthora melonis)引发的病害。
36.优选地,所述植物为黄瓜、南瓜、冬瓜、苦瓜、丝瓜和节瓜中的至少一种;进一步为黄瓜和南瓜中的至少一种。
37.本发明的第六个方面,提供本发明第三方面的载体、表达盒、转基因细胞系、重组菌或重组病毒在(1)~(3)中任一项中的应用;
38.(1)抗瓜类疫病;
39.(2)制备抗瓜类疫病的产品;
40.(3)培育抗瓜类疫病转基因植物。
41.优选地,所述瓜类疫病为瓜类疫霉菌(phytophthora melonis)引发的病害。
42.优选地,所述植物为黄瓜、南瓜、冬瓜、苦瓜、丝瓜和节瓜中的至少一种;进一步为黄瓜和南瓜中的至少一种。
43.本发明的第七个方面,提供靶向上调本发明第一方面的黄瓜光合系统ii反应中心蛋白 cspsii
‑
cp47的表达量或增强本发明第一方面的黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47 活性的试剂在(1)~(3)中任一项中的应用;
44.(1)抗瓜类疫病;
45.(2)制备抗瓜类疫病的产品;
46.(3)培育抗瓜类疫病转基因植物。
47.优选地,所述瓜类疫病为瓜类疫霉菌(phytophthora melonis)引发的病害。
48.优选地,所述植物为黄瓜、南瓜、冬瓜、苦瓜、丝瓜和节瓜中的至少一种;进一步为黄瓜和南瓜中的至少一种。
49.本发明的第八个方面,提供一种培育抗瓜类疫病转基因植物的方法,包括如下步骤:向目的植物中导入编码本发明第一方面的黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47的核酸分子,得到抗瓜类疫病转基因植物;
50.优选地,所述核酸分子的序列如seq id no.2或seq id no.3所示。
51.优选地,所述瓜类疫病为瓜类疫霉菌(phytophthora melonis)引发的病害。
52.优选地,所述植物为黄瓜、南瓜、冬瓜、苦瓜、丝瓜和节瓜中的至少一种;进一步为黄瓜和南瓜中的至少一种。
53.本发明的第九个方面,提供一种产品,含有如下(4)~(7)中任一种物质:
54.(4)本发明第一方面的黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47;
55.(5)本发明第二方面的核酸分子;
56.(6)本发明第三方面的载体、表达盒、转基因细胞系、重组菌或重组病毒;
57.(7)本发明第七方面中所述靶向上调本发明第一方面的黄瓜光合系统ii反应中心蛋白 cspsii
‑
cp47的表达量或增强本发明第一方面的黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47 活性的试剂。
58.优选地,所述产品用于(8)~(9)中任一种:
59.(8)防治瓜类疫病;
60.(9)培育抗瓜类疫病转基因植物。
61.优选地,所述瓜类疫病为瓜类疫霉菌(phytophthora melonis)引发的病害。
62.优选地,所述植物为黄瓜、南瓜、冬瓜、苦瓜、丝瓜和节瓜中的至少一种;进一步为黄瓜和南瓜中的至少一种。
63.本发明的有益效果是:
64.本发明首次发现一种黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47,该黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47对瓜类疫病具有抗性:在过量表达黄瓜光合系统ii反应中心蛋白 cspsii
‑
cp47的植物子叶上接种瓜类疫霉菌,病斑面积明显减小,而在沉默黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47的植物子叶上接种瓜类疫霉菌,病斑面积明显增大;可
见,黄瓜光合系统ii反应中心蛋白cspsii
‑
cp47对植物抗瓜类疫病具有非常重要的作用,可用于抗瓜类疫病、制备抗瓜类疫病的产品、培育抗瓜类疫病转基因植物,具有广泛的应用前景。
附图说明
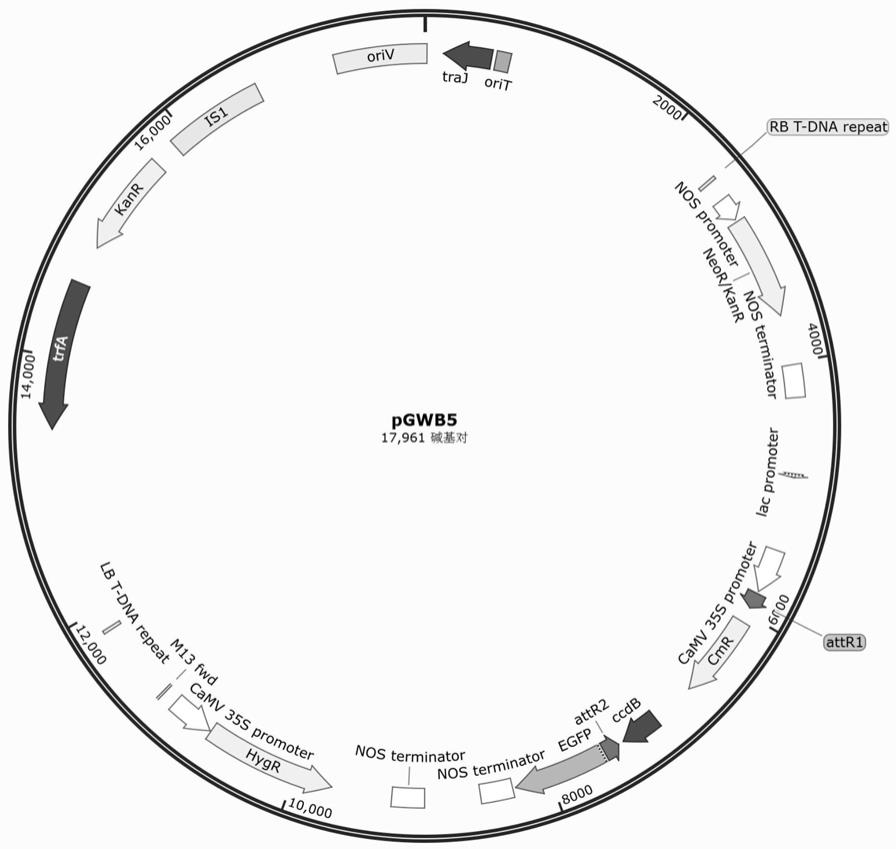
65.图1是pgwb5质粒图谱。
66.图2是pk7gwiw质粒图谱。
67.图3是cspsii
‑
cp47单克隆抗体制备过程图。
68.图4是不同处理的黄瓜子叶中cspsii
‑
cp47蛋白表达量的检测结果图。
69.图5是不同处理的黄瓜对瓜类疫病的抗性功能结果图:其中,a是不同处理的黄瓜对瓜类疫病的抗性功能的直观图;b是不同处理的黄瓜叶片的病斑面积统计结果图;不同字母代表存在显著性差异(p<0.05)。
70.图6是不同处理的南瓜子叶中cspsii
‑
cp47蛋白表达量的检测结果图。
71.图7是不同处理的南瓜对瓜类疫病的抗性功能结果图:其中,a是不同处理的南瓜对瓜类疫病的抗性功能的直观图;b是不同处理的南瓜叶片的病斑面积统计结果图;不同字母代表存在显著性差异(p<0.05)。
具体实施方式
72.以下通过具体的实施例对本发明的内容作进一步详细的说明。
73.这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体条件的实验方法,通常按照常规条件,例如sambrook等人,分子克隆:实验室手册(new york: cold spring harbor laboratory press,1989)中所述的条件,或按照制造厂商所建议的条件。实施例中所用到的各种常用化学试剂,均为市售产品。本实验使用黄瓜材料为本单位林毓娥老师于1995年2月从亚蔬中心(泰国曼谷)引进,自交留种,已于专利文献:cn112175988b 中公开。南瓜材料为广蜜1号,从广东科农蔬菜种业公司购买所得。瓜类疫霉菌为本实验室 2012年从广州白云区黄瓜病株分离所得,已在文献:王瑞,林毓娥,梁肇均,金庆敏,吴廷全.广东地区黄瓜疫病病原菌的分离与鉴定[j].中国农学通报,2016,32(01):76
‑
80.中公开。
[0074]
实施例1黄瓜cspsii
‑
cp47基因过表达载体的构建
[0075]
黄瓜cspsii
‑
cp47基因属于黄瓜光合系统ii反应中心蛋白基因。将黄瓜cspsii
‑
cp47基因cds全长(seq id no.2)构建到pgwb5(pgwb5质粒图谱如图1所示)上,具体位置为35s启动子后attr1和attr2之间(通过基因重组交换替代了ccdb基因),得到过量表达载体(pgwb5
‑
cspsii
‑
cp47);将黄瓜cspsii
‑
cp47基因cds非保守区段172bp长度的片段 (cds 1
‑
172bp,seq id no.4)及其互补片段构建到pk7gwiw上(pk7gwiw质粒图谱如图2所示),具体位置为35s启动子后两个attr1和attr2之间(通过基因重组交换替代了 ccdb基因),得到基因沉默载体(pk7gwiw
‑
cspsii
‑
cp47)。
[0076]
黄瓜cspsii
‑
cp47蛋白的氨基酸序列为(209aa):
[0077]
mglpwyrvhtvvlndpgrllsvhimhtalvagwagsmalyelavfdpsdpvldpmw rqdffhsfisgcfgfgafhvtglygpgiwvsdpygltgrvqavnpawgvsgfdpfvpggias hhiaagtlgilaglfhlslrppqr
lykglrmgnietvlsssidavffaafvvaepyhivpattp islnylgplarnisffecrvsrkekfl(seq id no.1)。
[0078]
黄瓜cspsii
‑
cp47基因cds(630bp)序列为:
[0079]
atgggtttgccttggtatcgtgttcataccgttgtattgaatgatcccggtcgtttgc tttctgttcatataatgcatacagctctagttgctggttgggccggttcaatggccctata cgaattagcagtttttgatccctctgatcctgttcttgatccaatgtggagacaagatttt tttcattcatttatttcgggttgctttggttttggtgcatttcatgtaaccggattgtatgg tcctggaatatgggtatccgatccttatggattaaccggaagggtacaagctgtaaatcc tgcgtggggtgtctcaggctttgatccttttgttccgggcggaatagcctcgcatcatat tgcagcaggtacattgggtatattagcgggcctattccatcttagtcttcgtccgccccaa cgtctatacaaaggattacgtatgggaaatattgaaaccgtcctttccagtagtatcgatg ctgtcttttttgcagcttttgtagttgctgaaccataccacatagttccagcaactaccc cgatttcattgaattatttgggcccactcgcaagaaatatctctttttttgagtgccgggt tagccgaaaagagaagtttctctga(seq id no.2)。
[0080]
黄瓜cspsii
‑
cp47基因(841bp)全长序列为:
[0081]
atgggtttgccttggtatcgtgttcataccgttgtattgaatgatcccggtcgtttgc tttctgttcatataatgcatacagctctagttgctggttgggccggttcaatggccctata cgaattagcagtttttgatccctctgatcctgttcttgatccaatgtggagacaaggtat gttcgttatacccttcatgactcgtttaggaataaccaattcatggggaggttggagtatc acaggagggactataacgaatccgggtatttggagttacgaaggtgtggccggagcac atattttgttttctggattgtgcttcttagcagctatctggcattgggtgtattgggatcta gaaatattttgtgatgaacaccgaagatttttttcattcatttatttcgggttgctttggt tttggtgcatttcatgtaaccggattgtatggtcctggaatatgggtatccgatccttatg gattaaccggaagggtacaagctgtaaatcctgcgtggggtgtctcaggctttgatcct tttgttccgggcggaatagcctcgcatcatattgcagcaggtacattgggtatattagcgg gcctattccatcttagtcttcgtccgccccaacgtctatacaaaggattacgtatgggaa atattgaaaccgtcctttccagtagtatcgatgctgtcttttttgcagcttttgtagttgc tgaaccataccacatagttccagcaactaccccgatttcattgaattatttgggcccactc gcaagaaatatctctttttttgagtgccgggttagccgaaaagagaagtttctctga (seq id no.3)。
[0082]
黄瓜cspsii
‑
cp47基因cds非保守区段172bp长度的片段如下所示:
[0083]
atgggtttgccttggtatcgtgttcataccgttgtattgaatgatcccggtcgtttgc tttctgttcatataatgcatacagctctagttgctggttgggccggttcaatggccctata cgaattagcagtttttgatccctctgatcctgttcttgatccaatgtggagac(seq idno.4)。
[0084]
实施例2构建瞬时过量表达/基因沉默的黄瓜子叶模型
[0085]
(1)将实施例1中过量表达载体和沉默载体分别转化农杆菌gv3101,在相应抗性的培养基上倒置培养48h。
[0086]
(2)挑取单克隆加入到4ml含有相应抗生素及利福平的lb培养基中,28℃,180rpm 摇菌24h。
[0087]
(3)按1:100比例加入新鲜含有相应抗生素及利福平的lb培养基中,28℃,180rpm摇菌到od600值约3.0。
[0088]
(4)3000rpm离心5分钟收集菌体,用悬浮液(10mm mes,10mm mgcl2)重悬菌体,并调整od600值到0.4左右,加入200mm乙酰丁香酮。
[0089]
(5)室温静置3h。
[0090]
(6)用注射器针头在黄瓜子叶主叶脉两侧各扎一个针孔。
[0091]
(7)用1ml注射器吸取等量静置过后的菌液,注射器对准背面的针孔,正面用手堵住,注射进菌液。
[0092]
(8)注射后的黄瓜幼苗黑暗培养12h后22℃光照培养3天。
[0093]
通过western blot检测cspsii
‑
cp47蛋白瞬时过量表达、沉默及野生型对照的黄瓜离体子叶中检测cspsii
‑
cp47蛋白的表达量。
[0094]
具体步骤如下:
[0095]
(1)将收集的黄瓜子叶液氮速冻后研磨加入蛋白提取液(50mm tris
‑
hcl ph 7.5,150mmnacl,0.1%triton x
‑
100,0.2%乙基苯基聚乙二醇(np
‑
40),1mm苯甲基磺酰氟(pmsf), 1
×
plant protease inhibitor cocktail(sigma,p8340
‑
5ml))混匀置于冰上30分钟。
[0096]
(2)4℃、14000rpm离心10min,收集上清液80ul加入20ul蛋白上样缓冲液均匀混合后沸水浴10分钟。
[0097]
(3)取20ul样品在sds
‑
page凝胶上进行电泳分离。
[0098]
(4)将电泳后的蛋白样品转到pvdf膜上,用5%pbst牛奶孵育封膜。
[0099]
(5)在封闭液中加入一抗(cspsii
‑
cp47单克隆抗体,制备过程如图3所示)孵育2小时,用pbst洗膜,每次5min,洗4次。
[0100]
(6)随后加入二抗(invitrogen,goat anti
‑
mouse igg,igm,iga(h l)secondary antibody, hrp,a
‑
10668),孵育30min,用pbst洗膜,每次5min,洗4次。
[0101]
(7)对膜进行扫描显色,结果见附图4。
[0102]
结果表明cspsii
‑
cp47基因瞬时过量表达的黄瓜子叶(pgwb5
‑
cspsii
‑
cp47)中cspsii
‑ꢀ
cp47蛋白表达量显著高于野生型对照(control),而cspsii
‑
cp47基因沉默的黄瓜子叶 (pk7gwiw
‑
cspsii
‑
cp47)中cspsii
‑
cp47蛋白表达量明显小于野生型对照(control)。说明cspsii
‑
cp47瞬时过量表达的黄瓜子叶模型以及cspsii
‑
cp47基因沉默的黄瓜子叶模型均构建成功。
[0103]
实施例3黄瓜子叶抗病实验
[0104]
对实施例2中构建成功的cspsii
‑
cp47瞬时过量表达的黄瓜子叶模型、cspsii
‑
cp47基因沉默的黄瓜子叶模型以及野生型的黄瓜子叶,将注射后的黄瓜幼苗黑暗培养12h后22℃光照培养3天后将黄瓜子叶离体,并接种瓜类疫霉菌后将黄瓜子叶于28℃培养皿中,黑暗培养 24h。
[0105]
依据不同处理的黄瓜离体子叶病斑大小来判断目的基因对疫病的抗性功能,结果如附图 5所示。
[0106]
结果显示在黄瓜子叶中瞬时过量表达cspsii
‑
cp47基因(pgwb5
‑
cspsii
‑
cp47)能够使黄瓜子叶对疫病的抗性较对照(control)明显增强,而在黄瓜子叶中瞬时沉默cspsii
‑
cp47 基因(pk7gwiw
‑
cspsii
‑
cp47)则使黄瓜子叶对疫病的抗性较对照(control)明显减弱。对每个处理的50个叶片病斑面积统计取均数(利用photoshop的像素数代替面积),随后,利用过量表达、沉默、对照的叶面积进行显著性分析和比较。结果显示,过量表达cspsii
‑
cp47基因叶片的病斑面积较对照显著减小,而沉默cspsii
‑
cp47基因叶片的病斑面积显著增大(图 5)。
[0107]
实施例4南瓜子叶抗病实验
[0108]
采用与实施例2中方法相同的操作步骤,同样构建了cspsii
‑
cp47瞬时过量表达的南瓜子叶,并通过western blot检测cspsii
‑
cp47蛋白瞬时过量表达及野生型对照的南瓜离体子叶中检测cspsii
‑
cp47蛋白的表达量,结果如附图6所示:cspsii
‑
cp47基因瞬时过量表达的南黄瓜子叶(pgwb5
‑
cspsii
‑
cp47)中cspsii
‑
cp47蛋白表达量显著高于野生型对照(control)。
[0109]
将注射后的南瓜幼苗黑暗培养12h后22℃光照培养3天后将南瓜子叶离体,并接种瓜类疫霉菌后将南瓜子叶于28℃培养皿中,黑暗培养24h。
[0110]
依据不同处理的南瓜离体子叶病斑大小来判断目的基因对疫病的抗性功能,结果如附图 7所示。
[0111]
结果显示:瞬时过量表达cspsii
‑
cp47基因叶片的病斑面积较对照显著减小,抗病性明显增强。
[0112]
上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受上述实施例的限制,其他的任何未背离本发明的精神实质与原理下所作的改变、修饰、替代、组合、简化,均应为等效的置换方式,都包含在本发明的保护范围之内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。