
hy5基因在调控植物对病虫害胁迫抗性中的应用
技术领域
1.本发明涉及生物技术领域,具体涉及hy5基因在调控植物对病虫害胁迫尤其是根结线虫的抗性中的应用。
背景技术:
2.近年来,我国设施蔬菜产业迅猛发展,然而,设施复种指数增加造成连作障碍频发,现已成为蔬菜产业可持续发展的瓶颈。土传病虫害是导致连作障碍的重要因素,其中根结线虫因其生活史短和繁殖方式灵活等特点在生产上防治困难,对设施蔬菜发展造成巨大经济损失。
3.根结线虫危害植株根部,使寄主细胞发生病变,形成根结,造成植株吸收水分和营养的能力下降,导致地上部分生长缓慢,产量低下,提前死亡。目前世界范围内已报道的根结线虫种类有90余种,据统计,危害最严重的是南方根结线虫(meloidogyne incognita)、爪哇根结线虫(meloidogyne javanica)、花生根结线虫(meloidogyne arenaria)和北方根结线虫(meloidogyne hapla)4种[moens m等,“meloidogyne species
‑
a diverse group of novel and important plantparasites.”root
‑
knot nematodes,2009,1,1
‑
17],其中南方根结线虫是我国危害面积最大的寄生性线虫。
[0004]
番茄(solanum lycopersicum l.)是世界上消费量最大的茄果类蔬菜之一,种植范围广泛,在设施蔬菜中占有重要地位。随着我国番茄设施栽培面积的不断扩大,重茬生产造成的土传病害日益严重,尤其番茄对根结线虫高度敏感。目前,在设施番茄生产上,综合应用农业、化学、物理和生物等措施进行根结线虫防治,但是不能从根本上制约其危害。
[0005]
基于此,剖析根结线虫的抗性机制,发掘抗性基因对于绿色高效防控技术的研发等具有重大的科学和现实意义。
[0006]
为了应对寄生性病原物的侵染,植物已经进化出两个水平的病原物识别机制以触发相应的免疫反应。其一,病原相关分子模式激发的免疫反应(pti),是指病原物表面的相关分子模式物质(pamps)刺激植物细胞膜上的模式识别受体(prrs),诱导植物产生防卫反应[zipfel和felix,“plants and animals:a different taste for microbes?”curropin plant biol,2005,8,353
‑
360]。蛔苷是迄今为止唯一被发现的线虫相关分子模式物质(namps),其中ascr#18是根结线虫分泌最丰富的蛔苷[jeong py等,“chemical structure and biological activity of the caenorhabditis elegansdauer
‑
inducing pheromone.”nature,2005,433,541
‑
545],可以成功激活番茄的免疫反应,然而,由于ascr#18人工合成困难,成本高昂,在农业生产上并未得到有效推广。
[0007]
其二,由效应因子触发的免疫反应(eti),是指植物细胞中的抗病蛋白r蛋白识别病原物分泌的效应蛋白,特异性地激发植物的防卫反应。番茄中合成抵御根结线虫的r蛋白编码基因是mi
‑
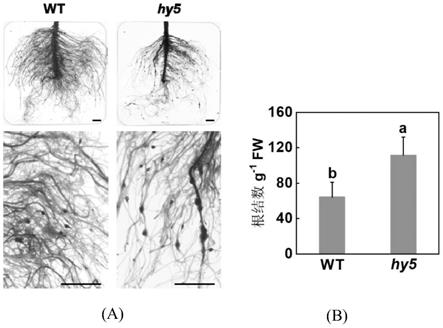
1[vos p等,“the tomato mi
‑
1gene confers resistance to both root
‑
knot nematodes and potato aphids.”nature biotechnology,1999,16(13),1365
‑
1369],但由于mi
‑
1基因存在温敏性,当温度超过28℃时,植株的线虫抗性消失,降低了其在
设施番茄栽培生产中的应用价值。
[0008]
因此,明确宿主植物是如何感知和响应根结线虫侵染,阐明宿主植物的信号转导途径,对了解植物抵御根结线虫的免疫机理具有重要的理论价值。
[0009]
植物激素是低量高效的小分子物质,他们在植物应对环境胁迫时扮演重要角色。其中,茉莉酸(ja)被公认在植物抵御生物胁迫中具有十分关键的作用。ja的合成经过多步酶促反应:叶绿体中的α
‑
亚麻酸在aoc(allene oxide cyclase)基因编码的丙二烯氧化物环化酶等多种酶的参与下,催化生成12
‑
氧代植物二烯酸(opda),而后opda转移到过氧化物酶体中继续后半部分的合成,最终形成ja。ja与异亮氨酸(ile)偶联生成复合物ja
‑
ile,该产物被认为是最具生物活性的ja化合物[wasternack和song,“jasmonates:biosynthesis,metabolism,and signaling byproteins activating and repressing transcription.”journal of experimental botany,2017,68(6),1303
‑
1321]。虽然ja信号的转导途径在植物病虫害领域已有广泛研究,但是由根结线虫侵染诱导的植物ja合成的调控机制还有待进一步探究。
[0010]
光不仅是植物光合作用的能量来源,还是参与植物逆境响应的重要信号来源。在植物的光信号调控网络中,hy5(long hypocotyl 5)是位于光受体下游的核心因子,它是c末端具有亮氨酸拉链(bzip)结构域的转录因子,不仅能够调控植物下胚轴伸长、昼夜节律和矿质元素运输等生长发育过程,还以激素依赖的方式参与植物高光胁迫、重金属胁迫和高低温胁迫等逆境响应。然而,hy5基因在植物病虫害领域的作用尚未有研究报道。
技术实现要素:
[0011]
本发明的目的在于发掘植物中参与抵御病虫害胁迫尤其是根结线虫的抗性基因,为研究植物抵御根结线虫的分子机理提供理论依据。
[0012]
为实现上述目的,本发明采用如下技术方案:
[0013]
本发明提供了hy5基因在调控植物对病虫害胁迫抗性中的应用,所述hy5基因的编码序列如seq id no.1所示或与seq id no.1所示序列具有至少70%同源性且编码的蛋白在功能上等价。
[0014]
进一步的,所述hy5基因通过提高植株抗性激素的含量,进而提高植物对病虫害的抗性。
[0015]
所述抗性激素为茉莉酸。
[0016]
本发明研究表明,hy5基因编码的蛋白可以直接结合茉莉酸(ja)合成相关基因aoc(allene oxide cyclase)启动子上的特殊元件,调控ja的合成。aoc基因编码的丙二烯氧化物环化酶为ja合成的关键酶,茉莉酸在植物抵御生物胁迫中具有重要作用。因此,hy5基因在植物抵御生物胁迫中发挥重要作用。
[0017]
进一步的,所述应用包括:利用生物技术手段,使植物体中的hy5基因上调表达,提高其对病虫害的抗性。
[0018]
进一步的,所述植物为番茄。
[0019]
进一步的,所述病虫害为根结线虫。
[0020]
本发明对番茄slhy5基因(solyc08g061130)的dna全长进行序列分析(http://solgenomics.net/),设计特定的向导rna(single guide rna,sgrna)。构建含有sgrna和
cas9蛋白的重组表达载体,接着通过农杆菌侵染的方式转化到番茄子叶中,进行组织培养,筛选得到定点编辑slhy5基因的一种番茄hy5缺失突变体植株;然后,将所述hy5缺失突变体植株进行根结线虫接种处理,观察植株接种根结线虫后的发病情况。研究发现,番茄slhy5基因突变后,根结数目显著增多;与野生型番茄(wt)相比,hy5缺失突变体中抗性激素含量明显降低。因此,番茄slhy5基因有利于提高番茄对根结线虫的抗性。
[0021]
所述slhy5基因的蛋白编码区的核苷酸序列如seq id no.1所示,长度为477bp;该基因编码的蛋白由158个氨基酸残基组成,其序列如seq id no.2所示。
[0022]
slhy5基因编码的蛋白质调控ja合成相关基因aoc的转录,进一步的,所述aoc基因的启动子的核苷酸序列如seq id no.3所示。
[0023]
本发明具备的有益效果:
[0024]
本发明首次公开hy5基因在植物对病虫害尤其是根结线虫抗性中的调控作用,剖析植物防御根结线虫的响应机制,为综合运用生理生化和环境调控等方法提高植物对根结线虫的抗性提供更完善的理论依据,对于研发环境友好型的根结线虫高效防控技术、减少农药使用和保障食品安全等方面具有十分重要的实践价值。
附图说明
[0025]
图1为番茄hy5基因crispr/cas9敲除植株t2代(纯合,不含外源cas9片段)的基因编辑位点和编码氨基酸变化示意图。
[0026]
其中,与野生番茄比较,可知番茄hy5缺失突变体在sgrna的位置发生了5个碱基的缺失突变,导致蛋白翻译提前终止;slhy5代表野生型番茄的基因序列,slhy5代表野生型番茄的氨基酸序列,hy5代表缺失突变体植株,
“‑‑‑‑‑‑‑‑‑‑”
表示缺失序列。
[0027]
图2为hy5缺失突变体接种根结线虫4周后的根结表型观察。
[0028]
其中,(a)为植株根部酸性品红染色结果,下图为上图的放大图,标尺bar=1cm;(b)为根结数目统计结果;wt代表野生型番茄,hy5代表缺失突变体植株;通过方差分析(anova)对数据进行统计分析,使用tukey检验分析了数据差异的显著性,图中相同字母表示平均值在p<0.05时无显著差异。
[0029]
图3为hy5缺失突变体接种根结线虫24h后的ja含量(a)和ja
‑
ile含量(b)变化;wt代表野生型番茄,hy5代表缺失突变体植株。通过方差分析(anova)对数据进行统计分析,使用tukey检验分析了数据差异的显著性,图中相同字母表示平均值在p<0.05时无显著差异。mock代表未接种线虫;rkn代表接种线虫24h。
[0030]
图4为转录因子slhy5与ja合成相关基因启动子结合能力分析;其中ja合成相关基因为aoc。
具体实施方式
[0031]
下面结合附图和具体实施例,进一步阐述本发明。应理解,以下列举的仅是本发明的具体实施例,本发明的保护范围不仅限于此。
[0032]
若未特别指明,下述实施例中所用实验方法均为常规方法,所用实验材料、试剂等,均可从商业途径得到。下述实施例中采用的番茄品种为番茄常规品种ailsa craig。
[0033]
实施例1番茄hy5缺失突变体植株的制备与鉴定
[0034]
1.1含特异sgrna的crispr/cas9载体的构建
[0035]
在sgn网站(http://solgenomics.net/)找到slhy5的dna全长序列如seq id no.4所示。
[0036]
利用crispr
‑
p网站(http://cbi.hzau.edu.cn/cgi
‑
bin/crispr)设计slhy5基因的sgrna序列,设计引物。
[0037]
sgrna
‑
hy5
‑
f:5
’‑
gattggatgatctctcactacttga
‑3’
(seq id no.5);
[0038]
sgrna
‑
hy5
‑
r:5
’‑
aaactcaagtagtgagagatcatcc
‑3’
(seq id no.6)。
[0039]
将合成的sgrna正反向引物用pcr仪器退火成含有粘性末端接头的双链sgrna,与经过bbsⅰ单酶切过的atu6
‑
sgrna
‑
atubq
‑
cas9中间载体用t4连接酶16℃过夜连接。连接产物热击转化dh5α大肠杆菌感受态细胞,在含有氨苄青霉素的lb固体培养基37℃过夜培养后,挑取单克隆菌落,利用退火时的上游引物sgrna
‑
hy5
‑
f:5
’‑
gattggatgatctctcactacttga
‑3’
(seq id no.5)和下游载体引物:5
’‑
ctacttatcgtcatcgtctttg
‑3’
(seq id no.7),进行pcr验证,阳性单克隆进行测序鉴定,获得阳性sgrna
‑
hy5克隆,提取质粒。
[0040]
质粒sgrna
‑
hy5和载体pcambia1301分别用hindⅲ和ecorⅰ双酶切,二者用t4连接酶16℃过夜连接。连接产物热击转化dh5α感受态细胞,在含有卡那霉素的lb固体培养基37℃过夜培养,挑取单克隆菌落,进行pcr验证鉴定阳性克隆后测序。将测序正确的阳性质粒电击转化gv3101农杆菌感受态细胞,获得含特异sgrna的crispr/cas9载体的农杆菌菌株。
[0041]
1.2 hy5缺失突变体植株材料制备与鉴定
[0042]
利用农杆菌侵染法将实施例1.1制得的农杆菌菌株转化到野生型番茄ailsa craig的子叶中,利用潮霉素初步筛选出候选转基因植株。
[0043]
在slhy5基因的sgrna的序列位置附近设计特异性引物检测靶基因序列的变化情况,检测片段长度为504bp。
[0044]
crispr
‑
hy5
‑
f:5
’‑
tcatgtcccgctatttcttt
‑3’
(seq id no.8);
[0045]
crispr
‑
hy5
‑
r:5
’‑
cttttgttttctttgtcagc
‑3’
(seq id no.9);
[0046]
对pcr产物进行测序,将测序结果与野生型番茄的slhy5基因进行序列比对,检测在sgrna的位置是否有碱基突变,筛选出slhy5基因发生改变的t0代基因编辑植株,命名为hy5。
[0047]
将上述t0代植株种植于生长室,自交得到t1种子,并检测基因编辑情况,方法同上。
[0048]
通过pcr方法扩增t1代植株中的crispr/cas9载体序列。
[0049]
cas9载体前引物:5
’‑
agcttcattcggagtttttg
‑3’
(seq id no.10);
[0050]
cas9载体后引物:5
’‑
ctacttatcgtcatcgtctttg
‑3’
(seq id no.11);
[0051]
扩增后进行电泳检测,未观察到cas9条带扩增。从而获得了不含外源cas9片段的缺失了5个碱基的t1代植株,序列比较如图1所示。
[0052]
将该不含外源cas9片段t1代植株进行自交,获得不含外源cas9片段,sgrna发生变异且可稳定遗传的t2代。
[0053]
下述实施例2与实施例3接种线虫实验均采用t2代材料。
[0054]
实施例2番茄slhy5基因对线虫抗性的影响
[0055]
对得到的番茄slhy5基因突变体植株进行接种根结线虫处理,具体如下:
[0056]
将野生型番茄(wt)和突变体植株(hy5)分为两组,一组为对照组,一组为实验组。
[0057]
当番茄长至四叶一心时,对实验组进行根结线虫接种处理,接种的根结线虫为j2期的南方根结线虫。
[0058]
南方根结线虫的培养和接种,具体步骤如下:
[0059]
(1)将感染根结线虫的病根打浆后与沙土充分混匀置于340mm
×
270mm
×
130mm的水培箱中,每盆栽种6株普通栽培番茄,于浙江大学农业试验站温室中繁殖线虫,室温维持在22
‑
26℃。
[0060]
(2)挖取发病严重的根系,将根部沙土用清水缓慢冲洗干净后,浸泡于0.5%的次氯酸钠水溶液中搅拌消毒3min,后用清水冲洗数次。
[0061]
(3)将根剪成1cm的根段,放入搅碎机中打碎。匀浆依次经过80目、200目、325目和500目筛子,最后将500目筛子中的残渣冲洗于烧杯中,获得卵悬浮液。
[0062]
(4)将收集到的卵悬浮液均匀倒入铺有8层吸水纸的13cm
×
13cm方形培养皿中,置于28℃恒温培养箱中孵化2
‑
3d。
[0063]
(5)用洗瓶轻轻冲洗培养皿,使孵化好的根结线虫冲洗进烧杯中,在50倍光学显微镜(bx61;olympus co.,tokyo,japan)下观察记录j2期线虫的数目。
[0064]
(6)将线虫悬浮液接种至待处理植株根部土壤,每株约接种1000条j2期线虫,置于人工气候室培养,期间正常浇水。
[0065]
植株栽培在装有高温灭菌的河沙的塑料杯中,用大量元素水溶肥(杭州康成农业科技有限公司)浇灌。生长条件为:温度25℃/20℃(昼/夜),光周期12h/12h(昼/夜),光通量密度为400μmolm
‑2s
‑1,湿度75%左右。
[0066]
线虫处理4周后,进行根结表型观察和根结数统计。
[0067]
根结表型观察采用酸性品红染色法,具体方法如下:
[0068]
(1)根系洗净,用1.5%
‑
5%的次氯酸钠溶液对根系进行5
‑
10min的光下漂白,以去除根系内影响染色的杂质。
[0069]
(2)将上述根系用清水冲洗并浸泡30min。倒掉浸泡液,吸干根系水分,用3.5%酸性品红溶液将根系完全浸没,加热至沸腾并持续3
‑
5min,室温静置冷却。
[0070]
(3)根系冷却后用清水冲洗,除去表面多余品红液体。
[0071]
(4)加入酸性甘油后加热至沸腾后立即将根系转移至常温酸性甘油中。
[0072]
(5)在酸性甘油中放置24h后进行根结表型拍照和根结数统计。
[0073]
根结数统计的具体方法如下:
[0074]
将待统计的番茄根系洗净,表面水分吸干,称重,仔细记录每棵根的根结数。通过计算每克鲜重根的根结数量来评估植株的线虫敏感性。
[0075]
根结表型观察和根结数统计结果见图2。
[0076]
与wt相比,hy5突变体植株根结表型明显(图2a)且根结数显著增多(图2b),表明植株根结线虫抗性减弱,说明slhy5基因能够提高番茄对南方根结线虫的抗性。
[0077]
实施例3番茄slhy5基因对线虫抗性激素的影响
[0078]
1、由于ja在植物响应生物胁迫中扮演重要角色,故对wt和hy5突变体植株接种根结线虫后,进行ja和ja
‑
ile含量测定。
[0079]
在根结线虫接种24h后取番茄叶样,提取和测定ja和ja
‑
ile含量,具体方法如下:
[0080]
(1)将100mg液氮冷冻样品在磨样仪中研磨成粉末;
[0081]
(2)加入1ml色谱级乙酸乙酯,该乙酸乙酯已加入d6
‑
ja(olchemim ltd.,czech)和d6
‑
ja
‑
ile(quality control chemicals inc.,usa)作为内标,最终浓度分别为10ng/ml和40ng/ml的标准液。避光,4℃180rpm震荡萃取12h;
[0082]
(3)4℃,12000rpm,离心10min,收集上清液,用1ml色谱级乙酸乙酯重悬沉淀,避光,4℃180rpm再次震荡萃取1h;
[0083]
(4)合并两次上清液于5ml离心管中,n2吹干;加入0.5ml的70%色谱级甲醇(v/v),充分涡旋,将离心管中残余物重新悬浮,4℃,12000rpm,离心10min;
[0084]
(5)将上清液转移至棕色玻璃瓶中,用hplc
‑
ms/ms进行分析测定(agilent technologies,california,america)。ja和ja
‑
ile含量测定结果见图3。
[0085]
根结线虫接种24h后,hy5缺失突变体中由根结线虫诱导的ja和ja
‑
ile含量都显著低于野生型番茄,表明slhy5基因促进ja合成。
[0086]
2、slhy5蛋白被证明可以与基因启动子上核心序列为acgt的顺式作用元件特异性结合,具有转录调控功能。对ja合成相关基因aoc的启动子进行序列分析,结果发现该基因启动子上存在slhy5能够识别的特殊元件,通过emsa实验检测slhy5蛋白与特殊元件的结合能力,具体方法如下:
[0087]
(1)通过pcr方法扩增slhy5基因全长序列。
[0088]
slhy5扩增前引物:
[0089]5’‑
cgggatccatgcaagagcaagcgacgag
‑3’
(seq id no.12);
[0090]
slhy5扩增后引物:
[0091]5’‑
cccaagcttctacttcctcccttcctgtgc
‑3’
(seq id no.13);
[0092]
(2)bamh i和sac i双酶切pet
‑
32a载体,利用同源重组技术与扩增产物连接。重组载体转化到大肠杆菌感受态bl21(de3)中,使用0.1mm的异丙基
‑
β
‑
d
‑
硫代半乳糖吡喃糖苷诱导重组的his
‑
hy5融合蛋白,根据novagenpet纯化系统纯化蛋白;
[0093]
(3)设计合成包含aoc启动子上的g
‑
box(cacgtg)以及将g
‑
box中的acgt序列突变为tttt(cttttg)的引物,序列如表1所示。用biotin3’end dna labeling kit(89818,pierce,usa)对前引物进行3’末端生物素标记,而后退火合成双链探针;
[0094]
表1凝胶电泳迁移实验所用引物
[0095]
引物名称(5
’‑3’
)aoc正向引物gcaggagaaacacgtgttgccactttaoc反向引物aaagtggcaacacgtgtttctc ctgcaoc
‑
mut正向引物gcaggagaaacttttgttgccactttaoc
‑
mut反向引物aaagtggcaacaaaagtttctcc tgc
[0096]
(4)根据light shift chemiluminescent emsa kit(20148,thermo fisher scientific,usa)试剂盒方法,将0.5μg的hy5融合蛋白与2μl标记探针加入20μl反应体系,28℃孵育30min,产物在6%的聚丙烯酰胺凝胶上电泳分离,经过转膜、紫外交联、封闭和孵育后显色拍照。测定结果见图4。
[0097]
emsa实验结果表明,slhy5蛋白可以结合aoc启动子上含有acgt核心序列的顺式作用元件,而当该元件中的acgt序列突变为tttt后,slhy5蛋白则无法与探针结合。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。
![基氮杂双[6]螺烯类化合物及其在空穴传输材料和太阳能电池中的应用的制作方法](http://img.xjishu.com/img/zl/2021/12/14/6cq0lahid.jpg)