不直接向其附着的细胞传导信号的非天然nkg2d受体
技术领域
1.本技术总体上涉及附着于哺乳动物细胞的非天然nkg2d受体的非天然胞外域,其中当与经修饰以特异性结合非天然nkg2d受体的非天然nkg2d配体结合时,所述受体不直接激活或直接传导信号给所述哺乳动物细胞,并且涉及附着到nkg2d配体的经修饰的α1
‑
α2结构域的异源分子。
背景技术:
2.nkg2d是在自然杀伤(nk)细胞以及某些t细胞和巨噬细胞表面表达为ii型同型二聚体整合蛋白的活化性受体。当与主要在受损细胞表面表达的其八种天然配体之一结合时,nkg2d激活nk细胞以杀伤受应激的细胞,或者当在t细胞上时,配体占据的nkg2d共刺激激活的t细胞以实现其效应子功能。已经对人类天然nkg2d的胞外域、数种其可溶性天然配体以及在一些情况下可溶性配体和受体胞外域的结合复合物求解了三维结构。nkg2d配体的单体α1
‑
α2结构域特异性结合至天然nkg2d同型二聚体的两个胞外域。
技术实现要素:
3.本公开涉及附着于哺乳动物细胞表面的非天然nkg2d受体,其中当所述非天然受体与经修饰以特异性结合该非天然nkg2d受体的nkg2d配体的同源非天然α1
‑
α2结构域结合时,所述非天然受体不直接传导信号给或直接激活所述细胞。nkg2d配体的非天然α1
‑
α2结构域可附着至异源原子或包括多肽在内的异源分子,在一些实施方式中,所述多肽为细胞因子或经修饰的细胞因子、抗体或抗体片段。细胞的直接活化或向细胞的直接信号传导并非由附着的非天然nkg2d受体所介导,并且即使当免疫突触已经出现时也不发生细胞的直接活化或向细胞的直接信号传导。
附图说明
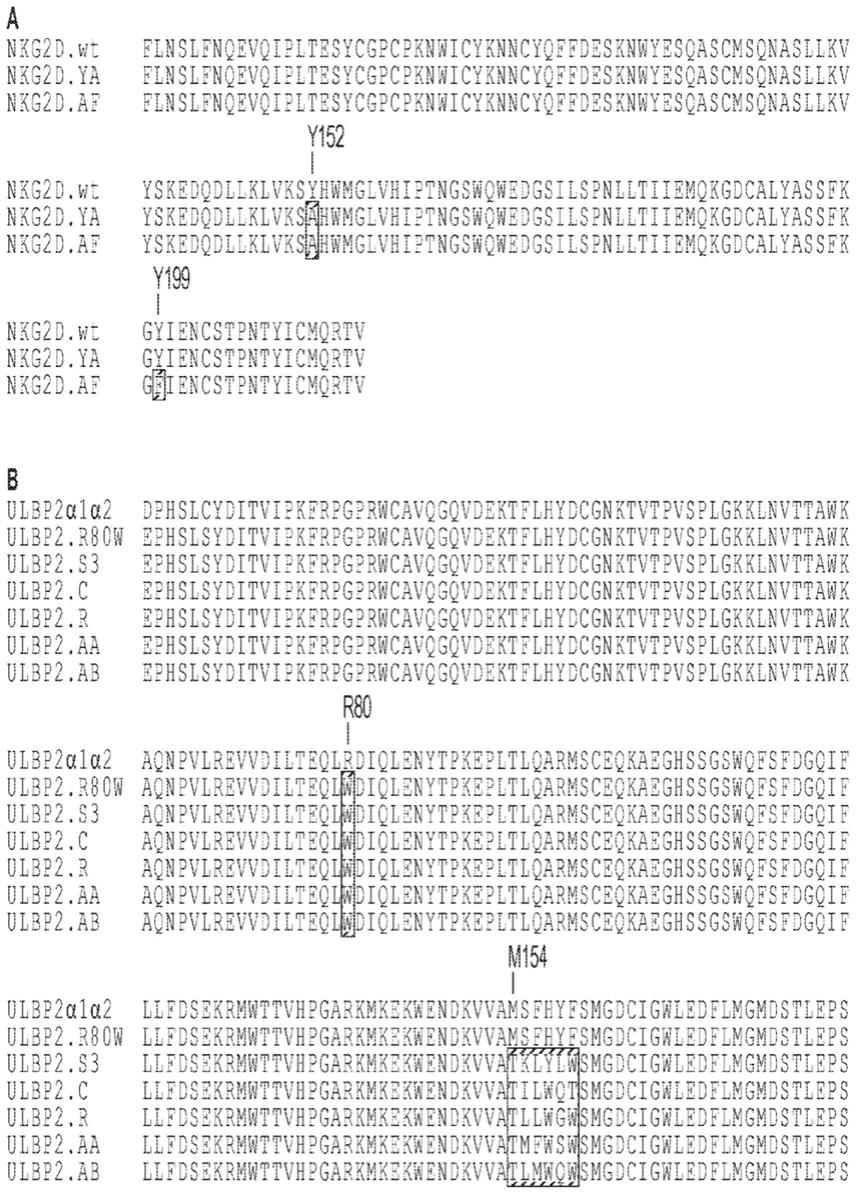
4.图1a
‑
1b:(图1a)天然nkg2d.wt胞外域(seq id no:17)与nkg2d.ya(seq id no:18)和nkg2d.af(seq id no:25)非天然变体的比对。指示了y152和y199的位置,并且灰色突出显示的是非天然变体中存在的突变残基。(图1b)天然/野生型ulbp2(seq id no:4)和ulbp2(包括ulbp2.r80w(seq id no:108))的非天然变体的α1
‑
α2结构域的比对。灰色突出显示的是对非天然ulbp2变体与非天然nkg2d.ya或nkg2d.af受体结合至关重要的残基。指示了残基r80以及m154
‑
f159区域的位置,其用于研究与nkg2d.ya(ulbp2.s3,seq id no.:127)或nkg2d.af(ulbp2.c,seq id no.:111;ulbp2.r,seq id no.:113;ulpb2.aa,seq id no.:115;和ulbp2.ab,seq id no.:117)结合的正交变体。
5.图2:在akta hiload 16/600superdex 200柱上分析的非天然fc
‑
nkg2d融合蛋白的尺寸排阻色谱比较。正确组装的材料的迁移通过在较高体积下洗脱的离散对称峰来例证,而聚集的材料在较低体积下更快地洗脱。通过氨基酸编号y152、y199或两者来指示修饰的位点和性质(从图的顶部开始,seq id no:48,43,42,58,41和40)。
6.图3:在akta superdex 200increase 10/300gl柱上分析了具有一个或两个氨基酸改变的非天然fc
‑
enkg2d变体的尺寸排阻色谱特性。正确组装的材料的迁移通过在较高体积洗脱的离散对称峰来例证,而聚集的材料(特征在于低振幅宽峰或一系列峰)在较低体积下洗脱。括号中的字母分别表示位置152和199处的氨基酸(从顶部开始按顺序为seq id no:57,56,55,54,53,52,51,50,49,48,47,46,45,44和40)。
7.图4:ulbp2野生型、micwed
‑
和mic25
‑
利妥昔单抗micabody与fc
‑
enkg2d候选物的elisa结合。关键字在图的顶部示出,但是由于许多曲线重叠,所以在每个图中也标记了单独的曲线。
8.图5:enkg2d变体与野生型配体的结合。野生型配体(都是fc
‑
融合形式)被octet ahc生物传感器捕获,并且每种天然nkg2d、nkg2d.y152a或enkg2d5(y152a/y199f)作为fc融合体从300nm滴定至0.41nm。通过octet量化最大结合响应。(注意每个图表的不同纵坐标)。
9.图6:各个噬菌体变体的滴定elisa以确认与fc
‑
nkg2d.af的选择性结合和与fc
‑
nkg2d.wt结合的减少或消除。在图22中详细描述了突变。
10.图7:结合nkg2d.wt、nkg2d.ya和nkg2d.af的四种非天然α1
‑
α2ulbp2变体micabody的elisa数据。将fc
‑
nkg2d变体用作捕获剂。用hrp
‑
缀合的抗人κ滴定并检测micabody。
11.图8:用netmhc4.0服务器,针对hla超型代表进行查询,检查了ulbp2.c(seq id no:111)、ulbp2.r(seq id no:113)、ulbp2.aa(seq id no:115)和ulbp2.ab(seq id no:117)相对于野生型ulbp2(seq id no:4)的肽
‑
mhci免疫原性的变化。对于输入序列,输入每个变体的可变区(根据图1b中的比对,残基154
‑
159)以及上游和下游的九个残基(总共24个残基),并检查9
‑
聚体肽窗以预测免疫原性。暗灰色方框对应于强烈预测结合mhci口袋(定义为具有%秩<0.5)并且因此具有强烈被呈递机会的肽。浅灰色方框对应于预测的弱结合剂(%秩<2)。附加细节参见实施例5正文。
12.图9:使用netmhcii 2.3服务器,针对hla
‑
dr、hla
‑
dq、hla
‑
dp进行查询,检查了ulbp2.c(seq id no:111)、ulbp2.r(seq id no:113)、ulbp2.aa(seq id no:115)和ulbp2.ab(seq id no:117)相对于野生型ulbp2(seq id no:4)的肽
‑
mhc ii类免疫原性的变化。对于输入序列,输入每个变体的可变区(根据图1b中的比对,残基154
‑
159)以及上游和下游的15个残基(总共36个残基),并且检查了15
‑
聚体的肽窗以预测免疫原性。暗灰色方框对应于强烈预测结合mhcii口袋并因此可能被呈递且具有免疫原性的肽。浅灰色方框对应于预测的弱结合剂。
13.图10a
‑
10b:elisa
‑
测量的包含ulbp2.wt(野生型)、ulbp2.r80w(其对野生型nkg2d具有增强的亲和力)、ulpb2.s3(nkg2d.ya
‑
选择的正交变体)或ulbp2.r(nkg2d.af
‑
选择的正交变体)的利妥昔单抗
‑
micabody与天然nkg2d.wt、nkg2d.ya和nkg2d.af的结合。(图10a)elisa曲线。对于一些在较高浓度下的测定,458nm吸收的降低是假象,在较高浓度的高亲和力接合体中经常见到这种假象,这是由于tmb
‑
ultra elisa显影试剂的沉淀引起的。(图10b)ec50值(以nm报告),如在graphpad prism中基于(a)中的曲线测定的。nd=未确定,因为增加的浓度和结合之间缺乏关系。
14.图11a
‑
11c:用未转导或用nkg2d.wt、nkg2d.ya或nkg2d.af car构建体转导的cd8效应细胞进行体外细胞溶解测定,所述nkg2d.af car构建体由cd8a铰链/跨膜结构域和细胞内4
‑
1bb和cd3ζ信号传导结构域组成。靶细胞预先负载钙黄绿素,并以递增的效应物与靶
比率(e:t)暴露于效应细胞。五小时后定量释放的钙黄绿素。(图11a)hela细胞的细胞溶解,(图11b)转染过表达表面ulbp1的hela细胞的溶解,和(图11c)在表面上表达非天然ulbp2.r的hela细胞的细胞溶解。误差线对应于实验中的技术重复的标准偏差。
15.图12a
‑
12b:micabody引起nkg2d
‑
car cd8t细胞对肿瘤系的细胞溶解。(图12a)将ramos细胞(其表达cd20,利妥昔单抗的靶标)预先加载钙黄绿素,并且以20:1的e:t比暴露于nkg2d.af
‑
或nkg2d.ya
‑
car细胞,同时增加ulbp2.s3或ulbp2.r利妥昔单抗
‑
micabody的浓度。共孵育两小时后定量细胞溶解水平。(图12b)将转染以表达人类her2的小鼠肿瘤系ct26用作与ramos细胞平行的细胞溶解靶。用饱和浓度(5nm)的利妥昔单抗
‑
ulbp2.r、曲妥珠单抗
‑
ulbp2.r或两种micabody的等摩尔混合物预装备nkg2d.af
‑
car cd8 t细胞。通过洗涤除去未结合的micabody,并将cd8细胞以两个不同的e:t比率添加到靶细胞。两小时后测量细胞溶解。
16.图13a
‑
13d:潜在micabody和micadaptor形式的说明。(图13a)在开发micabody和micadaptor试剂中利用的各种抗体fc变体,并且包括(a)野生型人类igg1 fc,(b)使得fc adcc
‑
缺陷的两个突变,和(c)在每个fc
‑
fc1或fc2
‑
(其允许产生异二聚体)
‑
fc分子中的静电转向突变,该分子还含有adcc
‑
缺陷突变。(图13b)正交配体如何可以与(a)重链或(b)轻链的c
‑
末端融合以产生micabody试剂的实例。(图13c)具有直接正交配体融合而没有抗体组分的micadaptor实例。(图13d)说明了多种micadaptor分子,其可以在人类igg1 fc的背景下产生以增强血清稳定性,并且可以(a,b,c;在文本和图例中表示为“fc1/fc2”)或可以不包括(d,e)异二聚体
‑
fc突变,这取决于异源载物或正交配体的所需分子的化合价。这也可以包括根据所需的载物、化合价和功能(f,g)利用完整的抗体结构,并且可以是重链或轻链融合体。
17.图14:car构建体、沉默car和其它car变体的示意图。如所示的每个构建体的seq id no。
18.图15a
‑
15c:选择性递送细胞因子融合体至nkg2d.ya
‑
car表达cd8 t细胞。(图15a)将表达nkg2d.wt
‑
car(seq id no:151)或nkg2d.ya
‑
car seq id no:153)的cd8细胞暴露于30或300iue/ml重组人类il2(rhil2)或il15(rhil15),或变异的突变体
‑
il2/
‑
il15正交配体直接融合体或异源二聚体fc(fc1/fc2)背景下的融合体。三天后,通过wst细胞增殖试剂定量增殖。此处的数据显示为相对于无细胞因子对照归一化。(图15b)将用编码nkg2d.ya
‑
car的载体转导的人类t细胞暴露于各种配体
‑
细胞因子融合分子七天,并随时间跟踪培养物中gfp car
‑
细胞的百分比。(图15c)如wst测定所确定的,正交配体增强了il21和突变体
‑
il21试剂向nkg2d.ya
‑
car细胞的递送,并且在三天的培养中促进了它们相对于未转导的细胞的扩增。横坐标图例的括号中指示了细胞因子或细胞因子
‑
micadaptor的iue/ml。micadaptor seq id no参见图24。
19.图16a
‑
16c:探索单独的nkg2d.ya胞外域促进正交配体
‑
细胞因子
‑
融合体递送至细胞的充分性数据。(图16a)单独的nkg2d.ya胞外域(seq id no:157)不能引起利妥昔单抗
‑
ulbp2.s3 micabody(seq id no:98和129)杀伤ramos细胞。(图16b)暴露于各种细胞因子试剂三天后的wst增殖测定。(图16c)wst增殖测定证明了nkg2d.ya
‑
car与正交配体的接合,但不存在融合的细胞因子或细胞因子突变体时不足以驱动细胞扩增。micadaptor seq id no参见图24。横坐标图例的括号中指示了细胞因子和细胞因子
‑
micadaptor的iue/ml
量。
20.图17a
‑
17c:(图17a)在用指定的细胞因子或细胞因子
‑
micadaptor一起孵育三天后,各种共刺激结构域突变体(seq id no:161,163和165)的wst增殖测定。(图17b)检验了相同的共刺激结构域car突变体在利妥昔单抗
‑
ulbp2.s3 micabody(seq id no:98和129)存在下有效裂解负载钙黄绿素的ramos靶细胞的能力。(图17c)如wst测定法所评估的,在与细胞因子试剂孵育三天后,nkg2d.ya胞外域(nkg2d.ya
‑
ecd)与完整cd19scfv
‑
car的共表达足以促进增殖。micadaptor seq id no参见图24。
21.图18:通过尺寸排阻色谱(sec)确定候选的非天然fc
‑
enkg2d变体突变和蛋白质聚集性质的汇总。
22.图19:通过micwed
‑
micabody或mic25
‑
micabody归一化至野生型nkg2d结合的enkg2d变体的饱和百分比(rmax)。野生型fc
‑
nkg2d和每种fc
‑
enkg2d受体被ahc生物传感器捕获,然后暴露于20nm的曲妥珠单抗
‑
特异性micabody。监测解离动力学,并对fc
‑
enkg2d融合体的rmax值进行排名。那些未测试的样品(nt)是由于sec分级后表达或回收的材料的严重聚集或不足量导致的。
23.图20:图3显示的fc
‑
enkg2d elisa的ec50值(nm)。nt=未测试;nb=即使在300nm下也没有结合或非常低的结合,因此未计算ec50值。
24.图21:通过spot elisa证实了ulbp2内组合突变的亚集,其导致噬菌体克隆与nkg2d.af和天然nkg2d.wt有选择性结合。通过在所选噬菌体中出现的频率对突变体进行排名。
25.图22:通过定量elisa,利妥昔单抗
‑
micabody形式的nkg2d.af选择的ulbp2变体保留了它们与nkg2d.af的结合特异性。显示了每种ulbp2变体的特定氨基酸修饰,均以其与fc
‑
nkg2d.wt融合的结合相对于fc
‑
nkg2d.af融合的比率表示。ulbp2的氨基酸残基位置是图1b的那些。
26.图23:在ulbp2.r80w(图1b;seq id no:108)的指定氨基酸位置处的选择突变,其产生y152a
‑
特异性噬菌体克隆。
27.图24:micadaptor seq id no和从瞬时转染中纯化的方法。
具体实施方式
28.免疫系统的自然杀伤(nk)细胞和某些(cd8 αβ和γδ)t细胞在人类和其它哺乳动物中作为抵抗肿瘤和感染细胞的首要的先天性防御而具有重要作用(cerwenka,a.,and l.l.lanier.2001.nk cells,viruses and cancer.nat.rev.immunol.1:41
‑
49)。nk细胞和某些t细胞在其表面上表现出nkg2d,这是一种突出的同源二聚体表面免疫受体,其负责识别靶细胞并激活抵抗病理细胞的先天性防御(lanier,ll,1998.nk cell receptors.ann.rev.immunol.16:359
‑
393;houchins jp et al.1991.dna sequence analysis of nkg2,a family of related cdna clones encoding type ii integral membrane proteins on human nk cells.j.exp.med.173:1017
‑
1020;bauer,s et al.,1999.activation of nk cells and t cells by nkg2d,a receptor for stress
‑
inducible mica.science 285:727
‑
730)。人类nkg2d分子具有结合其八种不同的同源配体的c型凝集素样细胞外(胞外
‑
)结构域,研究最多的配体是84%序列相同或同源的单体mica
和micb,其为主要组织相容性复合物(mhc)i类链相关糖蛋白(mic)的多态性类似物(weis et al.1998.the c
‑
type lectin superfamily of the immune system.immunol.rev.163:19
‑
34;bahram et al.1994.a second lineage of mammalian mhc class i genes.pnas 91:6259
‑
6263;bahram et al.1996a.nucleotide sequence of the human mhc class i mica gene.immunogenetics 44:80
‑
81;bahram and spies ta.1996.nucleotide sequence of human mhc class i micb cdna.immunogenetics 43:230
‑
233)。mica和micb的非病理性表达限于某些肠上皮、角质形成细胞、内皮细胞和单核细胞,但这些mic蛋白的异常表面表达响应于多种类型的细胞应激(如增殖、氧化和热休克)而发生,并将细胞标记为病理性的(groh et al.1996.cell stress
‑
regulated human mhc class i gene expressed in gi epithelium.pnas 93:12445
‑
12450;groh et al.1998.recognition of stress
‑
induced mhc molecules by intestinalγδt cells.science 279:1737
‑
1740;zwirner et al.1999.differential expression of mica by endothelial cells,fibroblasts,keratinocytes and monocytes.human immunol.60:323
‑
330)。mic蛋白的病理性表达似乎也涉及一些自身免疫疾病(ravetch,jv and lanier ll.2000.immune inhibitory receptors.science 290:84
‑
89;burgess,sj.2008.immunol.res.40:18
‑
34)。nkg2d配体(如多态性mica和micb)的差异调节,对于为免疫系统提供一种识别和响应各种紧急提示的手段并同时仍然保护健康细胞免受有害攻击是重要的(stephens ha,(2001)mica and micb genes:can the enigma of their polymorphism be resolved?trends immunol.22:378
‑
85;spies,t.2008.regulation of nkg2d ligands:a purposeful but delicate affair.nature immunol.9:1013
‑
1015)。
29.病毒感染是mic蛋白表达的常见诱导剂,可识别病毒感染的细胞以用于nk或t细胞攻击(groh et al.1998;groh et al.2001.co
‑
stimulation of cd8 αβtcells by nkg2d via engagement by mic induced on virus
‑
infected cells.nat.immunol.2:255
‑
260;cerwenka,a.,and l.l.lanier.2001)。事实上,为了避免对其宿主细胞的这种攻击,巨细胞病毒和其它病毒已经进化了防止mic蛋白在它们感染的细胞表面上表达的机制,以逃避被先天免疫系统靶向(lodoen,m.,k.ogasawara,j.a.hamerman,h.arase,j.p.houchins,e.s.mocarski,and l.l.lanier.2003.nkg2d
‑
mediated nk cell protection against cytomegalovirus is impaired by gp40 modulation of rae
‑
1molecules.j.exp.med.197:1245
‑
1253;stern
‑
ginossar et al.,(2007)host immune system gene targeting by viral mirna.science 317:376
‑
381;stern
‑
ginossar et al.,(2008)human micrornas regulate stress
‑
induced immune responses mediated by the receptor nkg2d.nature immunology 9:1065
‑
73;slavuljica,i a busche,m babic,m mitrovic,icekinovic,e markova car,ep pugel,a cikovic,vj lisnic,wj britt,ukoszinowski,m messerle,a krmpotic and s jonjic.2010.recombinant mouse cytomegalovirus expressing a ligand for the nkg2d receptor is attenuated and has improved vaccine properties.j.clin.invest.120:4532
‑
4545)。
30.尽管它们的应激,许多恶性细胞,例如肺癌和成胶质细胞瘤脑癌的那些细胞,也避免mic蛋白的表达,并且结果可能是特别具有侵略性的,因为它们也逃避了先天免疫系统
j.ban,david j.culpepper,luke d.davies,lisa k.sanders,and benjamin j.mcfarland.combining different design strategies for rational affinity maturation of the mica
‑
nkg2d interface.protein science 2012vol 21:1396—1402)。在本文中,我们描述了如下形式的非天然nkg2d受体与哺乳动物细胞表面的附着,其保留经修饰的非天然nkg2d配体与附着的异源分子的特异性结合,但即使在该哺乳动物细胞与异源分子所靶向的细胞或其它表面形成免疫突触时,所述非天然受体也可避免直接或顺式激活哺乳动物细胞或向哺乳动物细胞进行细胞内信号传导。非天然nkg2d受体本身已在一个或两个特定位点发生了突变,其中每一种突变均导致与nkg2d配体的所有天然α1
‑
α2结构域的结合受损或丧失(david j.culpepper,michael k.maddox,andrew b.caldwell,and benjamin j.mcfarland.systematic mutation and thermodynamic analysis of central tyrosine pairs in polyspecific nkg2d receptor interactions.mol immunol.2011january;48(4):516
–
523;uspto申请14/562,534;uspto临时申请62/088,456))。本发明产生了car,当其附着于哺乳动物细胞表面时,提供了沉默受体,其可以作为替代高亲和力受体,用于异源原子或异源分子向细胞表面的附着。因此,通过对非天然修饰的nkg2d受体具有特异性的附着的非天然配体,异源分子(例如包含缺陷性细胞因子)可以被特异性地递送到哺乳动物细胞表面的沉默受体,而不被递送到缺乏同源沉默受体的细胞。一旦结合到带有沉默受体的细胞上,缺陷性异源分子可结合到细胞表面上保留了与其结合性的其各自受体亚基上,从而直接向细胞传导信号,如同细胞被野生型配体刺激一样。
34.当然,在哺乳动物细胞上包含惰性非天然nkg2d、cd3
‑
ζ和共刺激结构域(如cd28、4
‑
1bb、icos或ox40)的car能够在形成免疫突触后直接刺激并激活car
‑
细胞。这种第二或第三代car
‑
t细胞的活化依赖于其cd3
‑
ζ结构域的功能和至少一个共刺激结构域(例如4
‑
1bb或cd28)的功能。然而,作为沉默car,这种car可以作为替代高亲和力受体,用于结合同源非天然配体
‑
附着的异源分子,该异源分子与其各自天然受体或受体亚基具有缺陷性结合。这种高亲和力结合使得附着于非天然配体的异源分子能够通过其各自的其它受体亚基将信号传递至细胞,所述其它受体亚基保留了结合性。
35.重要的是,当选择性失活这种具有直接活化能力的car的cd3
‑
ζ结构域时,其仍可作为沉默car,并使与其各自的天然受体或受体亚基具有缺陷性结合的同源非天然配体
‑
附着的异源分子能够经由它们各自的其它受体亚基向细胞传递信号。当car共刺激结构域(如4
‑
1bb)被失活并且保留活性cd3
‑
ζ结构域时,car不能作为沉默受体。即,尽管cd3
‑
ζ不是必需的,但需要功能性共刺激结构域以使异源分子(如附着于结合至受体的同源非天然配体的缺陷性细胞因子)能够介导其各自向car细胞的信号传导。
36.本发明揭示了为使附着至同源非天然配体的异源缺陷性细胞因子能够介导其各自向car细胞的信号传导,出乎意料的需要共刺激结构域而非cd3
‑
ζ。此外,本发明公开了共刺激结构域可以顺式或反式作用于沉默受体,该沉默受体上附着有融合至缺陷性异源缺陷分子的同源配体。
37.当靶向特定分子的异源分子(如抗体或抗体片段)附着至同源非天然nkg2d配体时(同源非天然nkg2d配体又附着至沉默受体),携带沉默受体的哺乳动物细胞将归巢至靶向异源分子将其导向的表面。即使在携带沉默受体的细胞和靶细胞表面之间产生“突触”时,前者也不会被沉默受体激活。
38.因为在细胞表面上存在本发明的基于非天然nkg2d的沉默受体的许多拷贝,所以在生产过程或治疗方案中,通过异源分子的归巢和/或选择性激活可以是多重的或顺序改变的。
39.携带沉默受体car的细胞也可表达与沉默car正交的另一种受体或car,所述另一种受体或car独立于所述沉默car发挥作用,以在适当刺激时特异性地和直接地激活相同细胞或以其它方式向相同细胞传导信号。其它或“第二”正交car可以是传统的单链fv(scfv)
‑
car或具有其自身同源非天然α1
‑
α2配体的第二正交的、基于非天然修饰的nkg2d的car。(af临时参考)。产生带有多于一种正交非天然car(沉默的或活性的)和(具有附着的异源分子或原子的)多种同源非天然配体的免疫系统的效应细胞的能力,极大地扩展了过继性细胞疗法(act)的效用、灵活性和控制。
40.在表征细胞上的沉默car及其对顺式或反式作用共刺激结构域(如4
‑
1bb)的依赖性的方法中,观察到与未修饰的人类t细胞相比,表达具有共刺激结构域的沉默car的人类t细胞表现出对天然il
‑
2或与天然或突变体il
‑
2融合的同源非天然配体的应答显著增强,其中所述突变体il
‑
2对其受体α
‑
亚基具有低亲和力。这一观察结果在表达car的细胞的离体或体内优先扩增中具有重要效用,所述car包含共刺激结构域且有或没有cd3
‑
ζ结构域。
41.如本文所用,“可溶性mic蛋白”、“可溶性mica”和“可溶性micb”指mic蛋白,其含有mic蛋白的α1
‑
α2结构域、有或没有mic蛋白的α3结构域、但没有跨膜或细胞内结构域。nkg2d配体
‘
ulbp1
‑6’
天然不具有α3结构域(cerwenka a,lanier ll.2004.nkg2d ligands:unconventional mhc class i
‑
like molecules exploited by viruses and cancer.tissue antigens 61(5):335
–
43.doi:10.1034/j.1399
‑
0039.2003.00070.x.pmid:12753652)。nkg2d配体的“α1
‑
α2结构域”指结合nkg2d受体的配体的蛋白质结构域。
42.在一些实施方式中,本发明的非天然nkg2d配体蛋白的α1
‑
α2结构域与nkg2d配体的原生或天然α1
‑
α2结构域具有至少80%同一性或同源性(seq id no:1
‑
9分别为mica、micb、ulbp1、ulbp2、ulbp3、ulbp4、ulbp5、ulbp6和omcp)。在其它实施方式中,经修饰的α1
‑
α2结构域与nkg2d配体的原生或天然α1
‑
α2结构域具有85%同一性。在另外其它实施方式中,经修饰的α1
‑
α2结构域与天然nkg2d配体蛋白的原生或天然α1
‑
α2结构域具有90%同一性,并且结合非天然nkg2d。
43.优选地,本发明的非天然mic蛋白的经修饰的或非天然α1
‑
α2结构域与8种人类nkg2d配体蛋白(seq id no:1
‑
8)之一的原生或天然α1
‑
α2结构域具有至少80%同一性或同源性,并且结合非天然nkg2d胞外域。在一些实施方式中,所述非天然α1
‑
α2结构域与nkg2d配体蛋白的原生或天然α1
‑
α2结构域具有85%同一性,并且结合非天然nkg2d。在其它实施方式中,所述非天然α1
‑
α2平台结构域与人类天然α1
‑
α2结构域蛋白的原生或天然α1
‑
α2平台具有90%、95%、96%、97%、98%或99%同一性,并且结合非天然nkg2d。
44.在一些实施方式中,可将异源分子标签融合于可溶性mic蛋白的非天然α1
‑
α2结构域的n
‑
末端或c
‑
末端,或融合于附着的异源肽或蛋白的n
‑
末端或c
‑
末端,以协助可溶性配体的纯化。标签序列包括肽,例如聚组氨酸、myc
‑
肽、flag标签、链霉亲和素样标签,或小分子,例如生物素。可在分离mic分子后,通过本领域技术人员已知的方法可以除去这些标签。
45.可以在nkg2d配体的α1
‑
α2结构域中进行特异性突变,以产生结合非天然nkg2d受
体的非天然α1
‑
α2结构域,所述受体本身被工程化以对天然nkg2d配体具有降低的亲和力。这可以通过例如遗传工程来完成。如此修饰的非天然nkg2d受体可用于在nk细胞、t细胞、巨噬细胞或免疫系统的其它细胞的表面上产生基于nkg2d的car,其可结合包含非天然α1
‑
α2结构域的分子。如下所述,与目前的car
‑
t细胞和car
‑
nk细胞相比,这些非天然nkg2d受体及其同源非天然nkg2d配体可提供用于治疗癌症和病毒感染的重要安全性、功效和制造优势。当哺乳动物细胞表面上的非天然nkg2d受体的细胞内信号传导如在本发明中一样已被沉默时,这些发明的car可作为用于原本缺陷性异源分子(例如细胞因子、趋化因子、淋巴因子、细胞毒素和与正交nkg2d配体融合或缀合的原子)的替代高亲和力受体。这提供了异源分子向携带沉默受体的细胞的直接且特异性递送,而沉默受体本身并不直接激活其宿主细胞。此外,结合特定靶标并由此结合携带这些靶标的细胞或其它表面的异源分子,可为携带沉默受体的细胞提供特异性归巢功能,而不会使其非故意地激活或刺激。
46.包含不结合或仅很少结合天然nkg2d配体的非天然nkg2d受体的胞外域的car
‑
t或car
‑
nk细胞,不会受到任何天然配体的激活,因此不会像表达基于天然nkg2d受体的car的细胞那样具有产毒性。此外,细胞上非天然nkg2d受体的胞外域不会被可溶形式的或骨髓衍生抑制细胞(mdsc)上的天然nkg2d配体下调(deng w,gowen bg,zhang l,wang l,lau s,iannello a,xu j,rovis tl,xiong n,raulet dh,2015.antitumor immunity.a shed nkg2dligand that promotes natural killer cell activation and tumor rejection.science.2015apr 3;348(6230):136
‑
9.doi:10.1126/science.1258867.epub 2015mar 5)。然而,当这样的携带非天然nkg2d受体胞外域的car细胞被具有本发明同源非天然α1
‑
α2结构域及其异源靶向基序的双特异性分子接合时,car将被激活并且表达car细胞的效应子功能,其中所述异源靶向基序已经被发现并结合其预期靶标。
47.因为除了在存在接合的双特异性分子时,所述双特异性分子包含同源非天然nkg2d受体胞外域,包含非天然nkg2d受体胞外域的car
‑
t或car
‑
nk细胞不被激活,因此它们的活化可以通过施用的双特异性分子控制,所述双特异性分子作为生物药物将表现出本领域公知的药代动力学和药效学。在不良事件发生的情况下,医师可以简单地改变施用的双特异性分子的给药方案,而不必像目前所做的那样必须使用诱导的自杀机制来破坏输注的car细胞(monica casucci and attilio bondanza.suicide gene therapy to increase the safety of chimeric antigen receptor
‑
redirected t lymphocytes.j cancer.2011;2:378
–
382)。此外,可以同时或顺序施用具有不同特异性靶向基序的此类双特异性分子,以协助解决由于靶抗原丧失所致的肿瘤抗性和逃脱,而不必创建、扩增和输注多种不同的自体car细胞(gill&june,2015)。由于所有car构建体对于所有car细胞而言可以相同,并且靶向特异性简单地由施用的本发明的双特异性分子的靶向基序决定,因此制造过程可被简化并且更便宜。
48.作为附着于nkg2d配体的非天然α1
‑
α2结构域的候选物的亲本或受体蛋白或多肽的实例包括但不限于抗体、包含ig折叠或ig结构域的蛋白(包括募集天然分子或不能募集或结合天然分子的经修饰的fc结构域)、球蛋白、白蛋白、纤连蛋白和纤连蛋白结构域、整联蛋白、荧光蛋白、酶、外膜蛋白、受体蛋白、t细胞受体、嵌合抗原受体、病毒抗原、病毒衣壳、用于细胞受体的病毒配体、激素、细胞因子和经修饰的细胞因子(如白细胞介素、扭结菌素(knottin)、环肽或多肽)、主要组织相容性(mhc)家族蛋白、mic蛋白、凝集素和凝集素的配
体。也可以将非蛋白质分子(如多糖、树枝状聚合物、聚乙二醇、肽聚糖、抗生素和聚酮化合物)连接到nkg2d配体的经修饰的α1
‑
α2结构域。
49.因此,本发明扩大了这种显著的、非常有前景的免疫方法的多样性和实用性,以用car
‑
t细胞、car
‑
nk细胞和car
‑
巨噬细胞样细胞治疗癌症,同时克服了许多这些目前公认的困难。
50.如本文所用,“肽”、“多肽”和“蛋白质”可互换使用;“异源分子”、“异源肽”、“异源序列”或“异源原子”分别为不是在与主题分子物理结合中天然或正常发现的分子、肽、核酸或氨基酸序列或原子。如本文所用,“非天然的”和“修饰的”可互换使用。如本文所用,“天然”、“原生”和“野生型”可互换使用,“nkg2d”和“nkg2d受体”可互换使用。本文的术语“抗体”以最广义使用,并且具体地涵盖单克隆抗体、多特异性抗体(例如双特异性抗体)和抗体片段,只要它们表现出期望的生物学活性即可。“抗体片段”包含抗体的一部分,优选包含其抗原结合区。抗体片段的实例包括fab、fab'、f(ab')2、fv片段和可被插入的fv;双抗体;线性抗体;单链抗体分子;和由抗体片段形成的多特异性抗体。
51.术语“包含”与“包括”、“含有”或“特征在于”可互换使用,是包含性或开放式语言,并且不排除另外的未列举的要素或方法步骤。短语“由
……
组成”排除权利要求中未指定的任何要素、步骤或成分。短语“基本上由
……
组成”将权利要求的范围限制至指定的材料或步骤以及那些实质上不影响所要求保护的发明的基本和新颖特征的材料或步骤。本公开考虑了对应于这些短语中的每一个的范围的本发明组合物和方法的实施方式。因此,包含叙述要素或步骤的组合物或方法考虑了其中组合物或方法基本上由那些要素或步骤组成或由那些要素或步骤组成的特定实施方式。
52.本文引用的所有参考文献都通过引用整体并入本文,无论先前是否具体并入。如本文所用,术语“一(a)”、“一(an)”和“任何”均旨在包括单数和复数形式。
53.现在已经充分描述了本发明,本领域技术人员将理解,在不背离本发明的精神和范围并且在没有过度实验的情况下,可以在宽范围的等效参数、浓度和条件下实施本发明。尽管已经结合具体实施方式描述了本发明,但应当理解的是,能够进一步修改本发明。本技术旨在涵盖通常遵循本发明原理的对本发明的任何变化、用途或改编,并且包括在本发明所属领域内已知或习惯做法范围内并且可以应用于上文所述基本特征的与本公开内容的偏离。
54.实施例
55.经修饰的nkg2d受体胞外域和nkg2d配体的经修饰的α1
‑
α2结构域
56.实施例1.将人类nkg2d受体的酪氨酸152修改为丙氨酸(y152a)且将酪氨酸199修改为苯丙氨酸(y199f),以产生惰性nkg2d胞外域
57.已由他人证明,人类nkg2d中在酪氨酸152或在酪氨酸199的突变,相当于nkg2d胞外域的位置73和120(图1a,seq id no.:17)可以大大降低与天然配体mica的结合(david j.culpepper,michael k.maddox,andrew b.caldwell,and benjamin j.mcfarland.systematic mutation and thermodynamic analysis of central tyrosine pairs in polyspecific nkg2d receptor interactions.mol immunol.2011january;48(4):516
–
523)。我们推论,鉴于任一酪氨酸残基的突变极大地影响了nkg2d结合其天然配体的能力,则在酪氨酸152(y152)和酪氨酸199(y199)上的同时突
变将实际上消除受体与所有原生配体接合的能力。因此,我们试图探索单个和组合的y152和y199取代,并就其生化行为对其进行表征,目的是识别不能与任何天然配体结合的单突变和双突变变体。还良好表达和组装的那些变体是特别受关注的,因为这些表明了可以更容易产生用于分析的惰性配体。
58.经由短因子xa识别性ile
‑
glu
‑
gly
‑
arg接头(seq id no:38),将天然nkg2d(野生型)胞外域(nkg2d.wt,seq id no:17)和候选非天然nkg2d变体胞外域(seq id no:18
‑
35)——也称为“工程化的nkg2d”或“enkg2d”,作为融合体克隆至人类igg1 fc的c
‑
末端(无fab结构域),并且可互换地称为fc
‑
nkg2d.wt或nkg2d.wt以及fc
‑
enkg2d或enkg2d(seq id no:40
‑
58)。合成了对应于mhci信号序列(seq id no:36和37)、具有接头的人类igg1 fc(seq id no:39)和nkg2d胞外域变体(seq id no:59
‑
77)的dna片段(integrated dna technologies,san diego,ca),并将其插入到pd2610
‑
v12(atum,newark,ca)中。在expi293
tm
细胞(thermofisher scientific,waltham,ma)中瞬时表达在y152、y199或y152/y199突变组合(图18)处探索取代的dna构建体,并通过蛋白a亲和层析(产品编号20334,pierce biotechnology,rockford,il)纯化分泌的蛋白。通过尺寸排阻色谱(sec)在akta pur superdex柱上表征洗脱的物质,并且将正确组装的尺寸合适的材料进行分馏并从聚集体峰中分离出来,然后再纳入测定。
59.纯化的nkg2d.y199a
‑
fc融合体的sec表征显示了主要聚集材料的组成(图2)。相比之下,天然fc
‑
nkg2d融合体和fc
‑
nkg2d.y152a融合材料通过离散的非聚集峰区分开来,该峰容易与更快迁移的聚集体区分。y199a突变对聚集的影响在y152a/y199a双突变体fc
‑
nkg2d融合变体中也是明显的,表明其对蛋白质错误折叠具有最重要的影响(图2)。因此,在nkg2d变体中包括y199a与y152突变的任何组合的这一方面对生产随后工程化工作所需的材料提出了挑战,并引起了关于在细胞表面上组装和呈递的担忧。因此,努力探索在y152和y199处的其它取代,它们可以组合产生更强的分子。将enkg2d组合的y152和y199候选突变体作为fc融合体进行检测并详述于(图18)。此外,通过sec对所有纯化和表达的fc
‑
enkg2d融合候选物进行作图,并且它们的色谱图揭示了不同水平的聚集体形成(图2和3,图18)。在第152位残基处探索的单个氨基酸取代中,丙氨酸、丝氨酸、苏氨酸和缬氨酸都不影响fc
‑
nkg2d分子的组装,但y152
‑
亮氨酸(y152l)产生了高度聚集的材料。与丙氨酸相似,在第199位谷氨酸和天冬氨酸都不耐受,但苯丙氨酸仅适度地增加聚集体形成。在研究的突变组合中,y152a/y199f、y152s/y199f、y152t/y199f和y152f/y199f没有负面地影响所需的二聚体形成,而其它组合导致聚集增加(图18,图2和3)。
60.实施例2:产生具有非天然nkg2d配体变体的基于抗体的双特异性分子“micabody”61.为了产生与人类igg1融合的非天然mica变体,使用引物pcr扩增编码例如micwed(seq id no:79)和mic25(seq id no:81)的α1
‑
α2结构域的dna多核苷酸,所述引物还引入编码用于与人类igg1的c
‑
末端κ轻链(seq id no:84)融合的aptsssggggs接头或用于与c
‑
末端重链(seq id no:82)融合的gggs接头的多核苷酸。此外,将两个突变引入重链的ch2结构域中——d265a/n297a(kabat编号;图13a和13b),这降低了与所有fcγr受体的结合,从而消除了抗体依赖性细胞毒性(adcc)功能(shields et al.,2001jbc,276:6591
‑
6604)。类似地克隆了编码野生型ulbp2(ulbp2.wt)的α1
‑
α2结构域而无其gpi
‑
连接的多核苷酸(seq id no:12),并将其融合到编码接头和igg1重链或轻链的dna多核苷酸上。这些双特异性抗
micabodies孵育两分钟,然后观察解离动力学30秒。然后用fc
‑
enkg2d融合受体作为捕获剂在相同条件下进行结合分析,以fc
‑
nkg2d.wt建立的最大结合响应的百分比对每个enkg2d的结合水平进行排名(图19)。对于micwed,除y199f外,所有单突变fc
‑
enkg2d变体的响应减少至50%。y199f保持了100%结合响应。然而,所有双突变fc
‑
enkg2d变体完全消除了与micwed的结合。对于mic25,相对于野生型fc
‑
nkg2d结合,所有单突变fc
‑
enkg2d变体和y152v/y199f保持了100%结合响应。然而,多种双突变fc
‑
enkg2d变体,包括y152a/y199f、y152s/y199f和y152t/y199f,结合降低至50%。
66.使用fc
‑
enkg2d融合体作为捕获剂的elisa测定用ulbp2.wt、micwed、mic25 micabodies进行,从300nm开始滴定(图4)。当可能时,使用graphpad prism计算ec
50
值(图20)。天然nkg2d结合基于ulbp2、micwed和mic25的micabody,计算的亲和力作为kds值分别为1.4、0.007和0.005nm。尽管ulbp2和micwed micabody与所有单突变enkg2d候选物的亲和力降低,但保留了mic25与enkg2d候选物的结合性。然而,所有双突变enkg2d候选物已消除或显著降低了与micabody形式的所有三种配体
‑
ulbp2、micwed和mic25
‑
的结合。
67.通过octet分析和elisa,enkg2d变体enkg2d5(y152a/y199f)、enkg2d7(y152s/y199f)、enkg2d8(y152t/y199f)和enkg2d9(y152v/y199f)与基于ulbp2、micwed和mic25的micabody的结合降低或消失(图19和20)。此外,enkg2d 5、7和8具有最小量的聚集,表明293t表达时更强的蛋白质组装(图18)。更仔细地检查了enkg2d5(seq id no:48)与野生型配体的结合,如在octet ahc尖端上捕获的micabodies。相对于天然(seq id no:40)nkg2d,单突变体fc
‑
nkg2d.y152a(seq id no:41)与所有天然配体的结合降低(图5)。相对于y152a enkg2d,enkg2d5(y152a/y199f)的结合的响应曲线甚至进一步降低。选择enkg2d5(y152a/y199f,下文称为“af”或“nkg2d.af”)作为先导nkg2d变体,对其工程化同源选择性、正交、非天然的配体。
68.实施例4:构建与非天然nkg2d.af胞外域选择性结合的正交非天然α1
‑
α2结构域
69.我们采用噬菌体展示来工程化正交的非天然α1
‑
α2结构域,其表现出与nkg2d.af(seq id no:48)受体的选择性结合。作为起点,选择对天然、野生型nkg2d(nkg2d.wt)胞外域具有高亲和力的非天然ulbp2.r80wα1
‑
α2结构域(图1b;seq id no:108)作为亲本结构域,用于进一步诱变和通过噬菌体展示筛选。针对ulbp2.r80w(seq id no:108)的α1
‑
α2结构域产生合成的dna文库,该结构域另外具有c8s突变以消除潜在的二硫键。使用nnk密码子替换配体上在结合态下位置与天然nkg2d受体上y152和y199位置紧密相邻的氨基酸残基的密码子;文库由位置154
‑
159处的nnk密码子组成(图1b;seq id no:110)。将文库克隆为与m13噬菌体的piii次要外壳蛋白的融合体,并根据标准方法(andris
‑
widhopf,j.,steinberger,p.,fuller,r.,rader,c.,and barbas,c.f.,3rd.(2011))在ss320大肠杆菌细胞中产生展示诱变的α1
‑
α2结构域变体的噬菌体颗粒。通过在非生物素化的天然fc
‑
nkg2d.wt竞争蛋白存在下,选择性捕获结合生物素化的fc
‑
nkg2d.af蛋白的噬菌体克隆,对这些α1
‑
α2噬菌体展示文库进行分选,以便对非天然nkg2d.af受体具有高结合亲和力。通过使用增加浓度的非生物素化的天然fc
‑
nkg2d进行多轮竞争性选择,从而富集了选择性克隆。
70.四轮选择后,将噬菌体克隆分别排列在96孔格板中,进行spot elisa以验证与板结合的非天然nkg2d.af和nkg2d.wt的优选差异结合。用生物素化的m13噬菌体外壳蛋白单
克隆抗体e1(thermofisher scientific,waltham,ma)、链霉亲和素
‑
hrp检测(r&d systems,minneapolis,mn)和1
‑
step ultra tmb elisa显影(thermofisher scientific,waltham,ma)检测结合的噬菌体。每个克隆的spot elisa信号表达为噬菌体结合nkg2d.af与噬菌体结合nkg2d.wt的比率。对比率大于或等于14的那些噬菌体进行测序,以识别nnk诱变区内的特定突变。图21显示了选择性结合的nkg2d.af的各α1
‑
α2噬菌体变体的所选氨基酸残基。在识别代表相同序列的多个克隆的情况下,绘制elisa信号的比率,并通过数据点的聚类验证噬菌体克隆的一致性(数据未显示)。
71.在单独的单一培养中扩增在elisa中识别的三十种变体,以产生高滴度微批次的噬菌体。将纯化的噬菌体浓度归一化为od
268
=0.5,然后针对板结合的fc
‑
nkg2d.af或fc
‑
nkg2d.wt进行1:3系列稀释,如上所述进行噬菌体检测和elisa显影。以这种方式测定的所有三十种变体一致地证明了与nkg2d.af的选择性结合,与nkg2d.wt结合很少至不结合(图6),即使在测定的最高噬菌体浓度下亦是如此。所选噬菌体还显示两个或多个噬菌体浓度对数的偏移,以实现在nkg2d.af和nkg2d.wt之间的半最大结合。
72.为了确认nkg2d.af
‑
选择性α1
‑
α2结构域变体保留了在抗体融合中的特异性结合特性,将21种变体(图22;例如seq id no:111
‑
118)作为与aptsssggggs接头的c
‑
末端融合克隆到利妥昔单抗抗体的轻链(seq id no:119
‑
126)。将得到的融合体通过gibson克隆(new england biolabs inc.,ipswich,ma)克隆到哺乳动物表达载体pd2610
‑
v12(atum,newark,ca)中,并与亲本抗体的重链(seq id no:99)共表达为成对的全igg抗体。根据制造商的方案在expi293
tm
细胞(thermofisher scientific,waltham,ma)中进行瞬时表达,并使用标准蛋白
‑
a亲和层析(产品编号20334,pierce biotechnology,rockford,il)纯化。elisa测量每个变体ulbp2α1
‑
α2抗体融合体与非天然fc
‑
nkg2d.af和天然fc
‑
nkg2d.wt的结合,显示相对于天然nkg2d.wt,它们对nkg2d.af具有显著更高的结合亲和力(图22)。总之,这些数据证明,本发明的非天然的正交α1
‑
α2结构域对非天然nkg2d.af受体具有高亲和力结合,并且与天然nkg2d受体的结合亲和力显著降低。此外,这些正交α1
‑
α2结构域与抗体多肽的融合体保留了它们的选择性结合特性,并且例如在嵌合抗原受体(car)t细胞的背景下用于将非天然nkg2d.af受体重新导向特异性抗原。
73.实施例5:识别非天然nkg2d配体,其可通过选择性结合一种或另一种区分非天然nkg2d受体变体
74.如上所述,使用非天然ulbp2.r80wα1
‑
α2结构域(seq id no:108)作为起始点,进行了噬菌体展示,以工程化对nkg2d.y152a(此后称为nkg2d.ya)受体具有选择性结合的正交非天然α1
‑
α2结构域。在非生物素化的天然fc
‑
nkg2d.wt(seq id no:40)竞争蛋白存在下,通过选择性捕获结合到生物素化的fc
‑
nkg2d.ya(seq id no:41)蛋白的噬菌体克隆,对α1
‑
α2噬菌体展示文库进行淘选,以获得对非天然fc
‑
nkg2d.ya受体的高结合亲和力。附加的噬菌体克隆验证工作导致识别了与fc
‑
nkg2d.wt相比与fc
‑
nkg2d.ya优先结合的变体(图23)。例如,通过elisa和octet分析(都以单体his
‑
标记的和双特异性抗体融合的形式)一致地证明,相对于天然nkg2d.wt,ulbp2.s3(seq id no:127)与非天然nkg2d.ya选择性结合。这代表了与非天然nkg2d受体具有高亲和力结合的正交α1
‑
α2结构域的本发明的一个独特形式(在这种情况下,nkg2d.ya与示例4中的nkg2d.af相反)。另外,正交α1
‑
α2结构域与抗体多肽的融合体保留了它们的选择性结合特性,并用于选择性地将非天然nkg2d受体重新导
向由融合的异源肽(如抗体)确定的特异性分子。
75.为了确定与nkg2d.ya(ulbp2.s3,seq id no:127)选择性结合的非天然α1
‑
α2结构域和与nkg2d.af选择性结合的非天然α1
‑
α2结构域是否可区分这两种非天然受体变体,进行elisa滴定。直接比较所有21种所选的结合nkg2d.af的α1
‑
α2变体与nkg2d.af和nkg2d.ya的结合。其中,四种表明了不能结合nkg2d.wt、对nkg2d.af具有强亲和力的特性,并且相对于nkg2d.af,大大降低(15
‑
20倍)或消除了与nkg2d.ya的结合(图7)。对于这四种非天然ulbp2α1
‑
α2变体
‑
ulbp2.c、ulbp2.r、ulbp2.aa和ulbp2.ab(seq id no:111,113,115和117),还使用netmhc4.0服务器(对于肽
‑
mhc i类结合,用9
‑
聚体肽分析查询所有hla超型代表;http://www.cbs.dtu.dk/services/netmhc/)和netmhcii 2.3服务器(对于肽
‑
mhc ii类结合,用15
‑
聚体肽分析查询hla
‑
dr、hla
‑
dq、hla
‑
dp单倍型;http://www.cbs.dtu.dk/services/netmhcii/),相对于野生型ulbp2肽序列(seq id no:4)检查了预测的免疫原性谱的改变,前述两种算法都是由丹麦技术大学开发的(http://www.bioinformatics.dtu.dk/;andreatta m and nielsen m,gapped sequence alignment using artificial neural networks:application to the mhc class i system,2016bioinformatics,32:511,pmid:26515819;jensen kk,andreatta m,marcatili p,buus s,greenbaum ja,yan z,sette a,peters b,and nielsen m,improved methods for predicting peptide binding affinity to mhc class i molecules,2018immunology,pmid:29315598)。向ulbp2.c、ulbp2.r和ulbp2.ab中并入突变不增加预测的免疫原性,而对于少数单倍型,ulpb2.aa略微增加(图8和9)。由于ulbp2.r对nkg2d.af的特异性及其缺乏可预测的免疫原性,选择ulbp2.r用于进一步的elisa分析,以直接比较其与ulbp2.s3(nkg2d.ya
‑
选择的非天然正交配体)、ulbp2.r80w(对野生型nkg2d具有增强亲和力的非天然配体)和野生型ulbp2(ulbp2.wt)的结合行为。针对野生型nkg2d(nkg2d.wt)和两种惰性非天然变体nkg2d.ya和nkg2d.af,测定了四种利妥昔单抗
‑
micabody试剂(seq id no:98和121,98和129,131和100,以及98和106分别作为ulbp2.r、ulbp2.s3、ulbp2.r80w和ulbp2.wt的重链和轻链)的结合(图10)。数据表明,nkg2d.ya
‑
选择的变体ulbp2.s3为一种以高亲和力与nkg2d.ya结合的micabody,但不接合nkg2d.af或天然nkg2d。此外,nkg2d.af
‑
选择的变体ulbp2.r以micabody形式以高亲和力结合nkg2d.af,但不接合nkg2d.ya或天然nkg2d。这些结果证明,探索nkg2d
‑
mic配体轴和开发独特成对的新型选择性非天然nkg2d受体及它们各自的同源非天然mic配体结合配偶体具有巨大的潜力。
76.实施例6:表达非天然nkg2d.af胞外域的car
‑
t细胞的靶向和杀伤活性受融合至异源靶向多肽的正交α1
‑
α2结构域控制
77.强烈寻求选择性控制car
‑
t细胞疗法的方法,以减轻毒性和改善抗肿瘤功效(gill and june,前面所引用的)。先前已经尝试使用cd16的胞外域开发car,然后其可以通过治疗性单克隆抗体的fc结构域接合,允许基于抗体的car
‑
t靶向控制(chang et al.,前面所引用的)。然而,基于cd16的car
‑
t细胞可识别血液和组织中几乎所有的内源性抗体分子,并且用于控制这些细胞的治疗性抗体会遇到来自nk细胞、pmn、单核细胞和巨噬细胞上的内源性cd16受体的竞争。这两个特征均分别造成肿瘤外毒性和不良药物动力学的问题。
78.天然nkg2d配体存在于某些健康组织和许多应激组织上,使用目前的nkg2d car方法(vanseggelen et al.2015)产生了极高的毒性风险。y152a非天然nkg2d受体特异性结合
非天然α1
‑
α2结构域nkg2d配体,构成了一种手段的实例,通过该手段,使用包含所发明的nkg2d配体的非天然α1
‑
α2结构域的双特异性蛋白质,可选择性控制非天然nkg2d car的活性。
79.我们用包含nkg2d的经修饰的y152a/y199f(“af”)胞外域的受体工程化car
‑
t细胞,所述胞外域缺乏与所有天然nkg2d配体的结合,或缺乏与y152a修饰的nkg2d(nkg2d.ya)正交并同源的先前描述的非天然α1
‑
α2结构域。所发明的同源非天然α1
‑
α2结构域以高亲和力与非天然nkg2d.af胞外域结合并避免与天然nkg2d胞外域和与nkg2d.ya胞外域结合。因此,工程化的α1
‑
α2结构域表现出相对于天然nkg2d和非天然nkg2d.ya,对非天然nkg2d.af胞外域具有强选择性,其代表了一种理想的系统,可用于选择性控制非天然nkg2d car受体,或可以被本发明的非天然α1
‑
α2结构域选择性接合的融合到非天然nkg2d胞外域的任何受体或蛋白质。本发明进一步使得单细胞表达两种不同的car——一种包含nkg2d.ya且另一种包含nkg2d.af——每种使用明显不同的胞内结构域进行信号传导。这些不同的car通过细胞外暴露于各自的同源正交micabody或另一种非抗体融合多肽而具有对细胞活性的独立双重控制。
80.为了证明用使用非天然nkg2d.af胞外域的嵌合受体构建的car
‑
t细胞的选择性控制,我们基于先前的工作,使用4
‑
1bb/cd3
‑
ζcar构建体(campana专利8,399,645),将各自的nkg2d胞外域融合到car的cd8铰链区(seq id no:151,153,155),构建了具有nkg2d.wt(seq id no:135)、非天然nkg2d.ya(seq id no:137)或非天然nkg2d.af(seq id no:139)胞外域的car。将这些构建体(seq id no:152,154,156)克隆到慢病毒载体中,并使用慢病毒转导在原代人类cd8
‑
阳性t细胞中表达。hela细胞在其表面具有组成性上调水平的mic配体,包括mica、micb、ulbp3和ulbp2/5/6(用于确定这一点的抗体不能区分这三种ulbp;人类ulbp
‑
2/5/6抗体,r&d systems,minneapolis,mn)。转染hela细胞以在表面上过表达天然ulbp1或nkg2d.af
‑
选择的变体ulbp2.r的细胞,并且这些细胞用作体外杀伤测定的靶标。hela靶细胞预先负载钙黄绿素并在不断增加的效应子与靶标(e:t)比率下,暴露于nkg2d.wt
‑
car、nkg2d.ya
‑
car或nkg2d.af
‑
car cd8细胞中五小时,之后对释放到上清液中的钙黄绿素量进行定量并且归一化至在洗涤剂处理后释放的总钙黄绿素(图11)。由于天然表达在hela细胞表面上的mic配体水平升高,表达天然nkg2d(nkg2d.wt)的cd8细胞作为car通过这种过表达的天然配体与hela细胞接合,并实现细胞溶解。然而,即使在高e:t比率下,nkg2d.ya
‑
和nkg2d.af
‑
car转导的cd8细胞都显示非常少的天然hela细胞裂解,与未转导的cd8t细胞的活性水平相当。当ulbp1在hela细胞表面过表达时,仅nkg2d.wt
‑
car cd8t细胞显著裂解它们。在高e:t比率下,使用nkg2d.ya
‑
car细胞存在一些额外的杀伤,但这在nkg2d.af
‑
car细胞中是不存在的,表明双突变y152a/y199f使得nkg2d比单一y152a突变更惰性。在过表达nkg2d.af
‑
选择性非天然ulbp2.r的hela细胞中,nkg2d.wt
‑
car细胞导向裂解(由于内源mic配体的识别),而nkg2d.af
‑
car细胞导向显着水平的裂解,与受体与其选择性配体的接合一致。
81.为了证明nkg2d.ya
‑
或nkg2d.af
‑
car细胞的裂解只能由适当的同源靶向micabody来引起,将ramos细胞与与非天然ulbp2.s3或ulbp2.r正交配体连接的基于利妥昔单抗的micabodies一起用作细胞溶解的靶标。如图12a所示,利妥昔单抗
‑
ulbp2.s3 micabody可引起nkg2d.ya
‑
car cd8细胞的细胞杀伤活性,但不是nkg2d.af
‑
car细胞,而利妥昔单抗
‑
interleukin 2analogues in cancer immunotherapy,1993cancer res 53:2597,pmid:8495422;k.sauv
é
et al.,localization in human interleukin 2of the binding site to the alpha chain(p55)of the interleukin 2receptor,1991pnas 88:4636,pmid:2052547),融合到在两个残基处被改变以在fc均二聚化的界面上表达带正电荷的赖氨酸残基的fc2的c
‑
末端(seq id no:183)。将两者独立地克隆到哺乳动物表达载体pd2610
‑
v12(atum,newark,ca)中,根据制造商的方案共转染到expi293
tm
细胞(thermofisher scientific,waltham,ma)中,并使用标准蛋白
‑
a亲和层析(产品编号20334,pierce biotechnology,rockford,il)纯化。通过尺寸排阻色谱(sec)在akta pur superdex柱上将纯化的物质分级分离。fc1上带负电荷的残基和fc2上带正电荷的残基提供了静电转向效应(kannan gunasekaran et al.,enhancing antibody fc heterodimer formation through electrostatic steering effects:applications to bispecific molecules and monovalent igg,2010j biol chem 285:19637,pmid:20400508),其促进对于正交u2s3配体和mutil2为单价的分子的异二聚体组装(图13da)。另外,研究了包含ulbp2正交配体与细胞因子融合体的其它组合的micadaptor——作为异二聚体fc1/fc2融合体或作为其中两种组分直接融合的单一多肽——并且如图24中详述和如上所述类似地克隆、共表达和纯化它们的dna。根据需要,在测定中,包括重组人类il2(rhil2,peprotech)和重组人类il15(rhil15,peprotech)作为对照。
85.转导cd8人类t细胞以表达nkg2d.wt
‑
car构建体(seq id no:151)或nkg2d.ya
‑
car构建体(seq id no:153)并暴露于30iue/ml对照细胞因子或各种micadaptor三天,并且使用wst
‑
1细胞增殖试剂(millipore sigma)量化细胞增殖水平。对照rhil2促进了两种car
‑
表达细胞的增殖,而单独的mutil2则不能,如从降低的与il2r
‑
α的结合能力以及因此降低的通过il2r
‑
β/γ
‑
c发出信号的能力所预期的一样(图15a)。当micadaptor包含非选择性u2r80w配体(u2r80w
‑
mutil2,seq id no:177)时,其以高亲和力与野生型和经修饰的nkg2d.ya受体接合,表达任一car的细胞通过增殖响应。然而,当mutil2与正交配体(诸如仅选择性接合nkg2d.ya的ulbp2.s2(seq id no:179)、micadaptor、仅与nkg2d.ya接合的u2s2
‑
mutil2)融合时,仅nkg2d.ya
‑
car细胞通过增殖响应。当融合到u2s2的细胞因子是具有v49d突变的il15的突变形式时(bernard,j.et al.,identification of an interleukin
‑
15αreceptor
‑
binding site on human interleukin
‑
15,2004journal of biological chemistry,279:24313,pmid:15039446)时,获得了类似的结果,所述v49d突变减少了与il15r
‑
α的接合(图15a)。当nkg2d.ya
‑
car
‑
t细胞与这些试剂共培养七天时,%gfp (指示与未转导的gfp
‑
阴性细胞相比存在的car
‑
表达细胞的百分比)增加,但仅当存在正交ulbp2.s2配体(u2s2
‑
mutil2或u2s2
‑
fc1/fc2
‑
mutil2,seq id no见图24)且不存在非选择性u2r80w变体(其结合在人类cd8细胞上组成型存在的天然野生型nkg2d受体)时如此(图15b)。il21也被作为野生型融合(il21.wt)进行了研究,具有影响il21r
‑
α结合的单突变(d18a或e109r),或具有包括所述两个突变的版本(d18a/e109r)(kang,l.et al.,rational design of interleukin
‑
21antagonist through selective elimination of theγc binding epitope,2010journal of biological chemistry,285:12223,pmid:20167599)。比较未转导的cd8细胞或nkg2d.ya
‑
car表达细胞,仅携带nkg2d.ya的细胞对所有版本的il21融合体(seq id no见图24),包括对il21.wt融合体,作出响应(图15c)。有趣的是,尽管
与无细胞因子对照相比,未转导的和nkg2d.ya
‑
car细胞均响应rhil2而扩增,但nkg2d.ya
‑
car细胞的增殖量更高。nkg2d.ya
‑
car表达细胞的增殖响应不依赖于nkg2d.ya结构域的一般化结合或接合,因为u2s3
‑
fc1/fc2(具有单一u2s3结构域且没有附着的细胞因子或细胞因子突变体的异二聚体fc分子)和利妥昔单抗
‑
micabody(对于u2s3结构域是二价的,但缺乏任何细胞因子组分,图13bb)在三天的培养过程中不诱导增殖,甚至在比rhil2对照高得多的iue/ml浓度下亦如此(图16c)。
86.实施例8:顺式或反式细胞内共刺激结构域的存在促进了携带非天然的、经修饰的nkg2d的细胞对细胞因子和细胞因子micadaptor的响应
87.在证明了在嵌合抗原受体构建体的背景下,经修饰的nkg2d.ya结构域实际上确实充当了用于递送与正交配体附着的异源载物的高度选择性对接位点后,我们试图确定nkg2d.ya受体是否不仅对于靶向递送细胞因子是必需的,而且对于靶向递送细胞因子也是足够的,所述细胞因子然后可以作用于接受细胞。nkg2d.ya胞外结构域(nkg2d.ya
‑
ecd)表达为除去了所有胞内组分的跨膜结构域,但保留了胞内egfp标签(图14,seq id no:157)。转导cd8细胞以表达该“沉默car”,并证明在利妥昔单抗
‑
ulbp2.s3 micabody(seq id no:98和129)存在下不能直接杀伤ramos靶细胞,正如没有共刺激结构域所预期的(图16a)。重要的是,表达这种完全惰性或沉默car的细胞在暴露于u2s3
‑
fc1/fc2
‑
mutil2(seq id no:189和193)时不增殖,但在与未转导的细胞相当的水平上响应(图16b)。这一观察结果——除了(a)与未转导的细胞相比,一致观察到rhil2可使nkg2d.ya
‑
car增殖水平更高(图15c、16b、16c)和(b)观察到仅nkg2d.ya
‑
car细胞对包括il21.wt在内的所有形式的il21
‑
micadaptor作出响应之外——导致我们推测nkg2d.ya
‑
car构建体中存在的细胞内结构域增强了对这些细胞因子和细胞因子micadaptor的响应性。
88.为了检测这一点,产生了一系列car构建体,其中car的胞内结构域的信号基序被突变——4
‑
1bb的两个traf2共有结合位点(seq id no:161)、cd3
‑
ζ中的三对itam基序(seq id no:163)或组合的4
‑
1bb/cd3
‑
ζ突变体(seq id no:165)。将这些构建体(图14)转导到cd8细胞中,与所示细胞因子试剂共孵育,三天后量化增殖(图17a)。在所有测试条件下,nkg2d.ya
‑
bb
‑
cd3
δitam
‑
gfp(seq id no:163)保留了与nkg2d.ya
‑
car相当的增殖响应,由此证明cd3
‑
ζ结构域(seq id no:145)对于在car背景下对细胞因子和细胞因子
‑
micadaptor的响应性不是必需的。然而,nkg2d.ya
‑
bb
δtraf2
‑
cd3ζ
‑
gfp受体
‑
表达细胞(seq id no:161)对所有细胞因子和细胞因子
‑
micadaptor的响应性显著降低,在两个细胞内结构域携带突变的nkg2d.ya
‑
bb
δtraf2
‑
cd3
δitam
‑
gfp(seq id no:165)受体也是如此。这表明共刺激4
‑
1bb(seq id no:143)在nkg2d.ya
‑
car细胞针对细胞因子和细胞因子
‑
micadaptor的响应中起作用,并且在细胞表面上表达的沉默car需要还具有细胞内共刺激结构域存在,以促进细胞因子
‑
micadaptor响应性。nkg2d.ya
‑
bb沉默car(seq id no:169,图14)不能引起micabody介导的细胞杀伤(图17b),证明缺乏cd3
‑
ζ的这种car架构适合作为用于micadaptor的沉默对接位点,并且使细胞因子
‑
micadaptor信号传导成为可能,但不能起到介导micabody靶向细胞的细胞溶解。
89.为了进一步探究4
‑
1bb结构域如何促成细胞因子和细胞因子
‑
micadaptor响应性,生成了构建体,其中cd19scfv
‑
car(基于fmc63 fv)含有功能性4
‑
1bb和cd3
‑
ζ结构域(seq id no:173)的完整互补序列。将cd19scfv
‑
car与nkg2d.ya
‑
ecd共表达(图14)。两种组分都
表达为具有t2a自切割肽基序的单一多肽,该t2a自切割肽基序将上游cd19scfv
‑
car构建体与下游nkg2d.ya
‑
ecd分开,其具有独立的gmscfrα链信号序列、cd8a铰链和cd8a跨膜结构域。将该构建体转导入cd8t细胞中,并且通过流式细胞术检查gfp信号和藻红蛋白缀合的micabody染色,分别验证了表面上cd19scfv
‑
car和nkg2d.ya
‑
ecd的共表达。将这些细胞与细胞因子和细胞因子
‑
micadaptor一起孵育三天,并通过wst测定量化增殖。nkg2d.ya
‑
car对rhil2和u2s3
‑
fc1/fc2
‑
mutil2均产生了预期的响应,尽管仅携带cd19scfv
‑
car(seq id no:171)的对照细胞随rhil2扩增,并且以较低程度扩增至u2s3
‑
fc1/fc2
‑
mutil2的最高浓度(300iue/ml)(图17c)。与仅表达cd19scfv
‑
car(seq id no:171)的细胞相比,cd19scfv
‑
car表达细胞(seq id no:173)上nkg2d.ya
‑
ecd的共表达表现出对u2s3
‑
fc1/fc2
‑
mutil2细胞因子
‑
micadaptor更强的增殖响应。在此背景下,4
‑
1bb结构域以反式组成型方式提供给nkg2d.ya
‑
ecd。这些数据证明,顺式或反式4
‑
1bb结构域促进了nkg2d.ya
‑
ecd表达细胞对细胞因子和细胞因子
‑
micadaptor的响应,并且nkg2d.ya
‑
ecd结构域可以与共刺激性含4
‑
1bb的car细胞共表达,以向过继性细胞治疗策略的工程化细胞提供增加的通用功能。这种功能性不仅限于在配体/沉默car接合后接合表面受体,而且可以通过并入胞质序列基序而扩展至细胞内递送(k.n.pandey,functional roles of short sequence motifs in the endocytosis of membrane receptors,2009front biosci,14:5339,pmid:19482617),所述胞质序列基序促进非天然nkg2d变体的周转,使得任何结合的micadaptor都被共内在化并且在细胞内递送异源载物。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。