一种用于在昆虫细胞中产生raav的核酸、vp1衣壳蛋白突变体及应用
技术领域
1.本发明属于基因工程技术领域,更具体地,涉及一种用于在昆虫细胞中产生raav的核酸、vp1衣壳蛋白突变体及应用。
背景技术:
2.腺相关病毒(adeno-associated virus,aav),也称腺伴随病毒,属于微小病毒科依赖病毒属,是目前发现的一类结构最简单的单链dna缺陷型病毒,需要辅助病毒(通常为腺病毒)参与复制。重组腺相关病毒(raav)由于具有宿主范围广、免疫原性低、安全性高、可介导外源基因在动物体内长期稳定表达等特点,是目前基因治疗领域最具应用前景的载体之一。随着第一种重组腺相关病毒(raav)介导的基因治疗药物的批准,对大规模aav载体制造技术的需求不断增加(yla-herttuala s.,2012,mol ther,20:1831-1832)。
3.产生大量重组腺相关病毒(raav)载体的能力是开发基于基因治疗的药物的重要因素。目前,生产raav的体系主要有两大类:一种是利用哺乳动物细胞(如293细胞、cos细胞、hela细胞、kb细胞等)的常规生产体系;另一种是利用昆虫细胞的生产体系。在哺乳动物细胞生产体系中,单个细胞的raav颗粒产量低,并且培养中极可能存在污染的风险,这限制了raav在哺乳动物细胞中的大规模生产及应用。杆状病毒/昆虫细胞表达系统是大规模生产raav的主要系统之一。重组腺相关病毒基因组中cap基因编码病毒vp衣壳蛋白,包括三种结构蛋白,分别为vp1、vp2和vp3,来自野生型病毒的aav中vp1、vp2和vp3的化学计量比约为1:1:10,这样的化学计量比对于重组aav的获得是很重要的。
4.尽管杆状病毒/昆虫细胞表达系统已成功在多种规模下用于raav生产,但是已有多项研究报道相比于利用哺乳动物细胞生产raav,昆虫细胞中生产的raav具有减少的vp1含量,从而导致raav的产率降低。因此,欲实现工业化大规模生产raav,杆状病毒/昆虫细胞系统仍需改善。
技术实现要素:
5.针对现有技术的缺陷,本发明的目的在于提供一种用于在昆虫细胞中产生raav的核酸、vp1衣壳蛋白突变体及应用,旨在解决野生型aav血清8型vp1蛋白不稳定、易降解从而导致raav产率降低的问题。
6.为实现上述目的,第一方面,本发明提供了一种核酸,其包含编码腺相关病毒vp1、vp2和vp3蛋白的核苷酸序列,与野生型aav血清8型的衣壳蛋白相比,所述vp1蛋白的氨基酸序列包含在第84、92和105位中的一个或多个位点的突变,且所述vp2蛋白的氨基酸序列未突变。
7.优选地,所述vp1蛋白的氨基酸序列包含一个或多个选自以下的突变:q84k、r92k和q105k。
8.优选地,除所述突变外,所述vp1蛋白的其它氨基酸序列与野生型aav血清8型的
vp1蛋白的氨基酸序列相同。
9.优选地,所述核酸还包含能够在昆虫细胞中驱动转录的启动子,所述启动子的3’端可操作连接有内含子;所述vp1蛋白的起始密码子atg位于所述内含子的序列之中,或者,所述vp1蛋白对应的cap基因编码区缺失起始密码子且所述内含子位于atg中的任意相邻两个核苷酸之间;所述内含子不包含能够在昆虫细胞中驱动转录的启动子;
10.所述核酸在昆虫细胞中仅能转录得到一种前体mrna,且在转录后加工过程中,仅通过所述内含子的选择性剪接作用,使得所述vp1蛋白的起始密码子atg保留或缺失,或者在vp1蛋白对应的cap基因编码区中形成起始密码子atg,从而实现调控所述vp1、vp2和vp3蛋白的翻译表达。
11.进一步优选地,所述vp1蛋白的起始密码子atg位于所述内含子的序列之中,且所述内含子的3’端可操作连接有编码2a自剪切多肽的核苷酸序列。
12.进一步优选地,所述2a自剪切多肽为t2a肽、p2a肽、e2a肽或f2a肽。
13.第二方面,本发明了提供一种腺相关病毒vp1衣壳蛋白突变体,其由本发明第一方面所述的核酸编码得到。
14.第三方面,本发明提供了一种昆虫细胞,其包含本发明第一方面所述的核酸,所述核酸为杆状病毒载体。
15.优选地,所述昆虫细胞为草地贪夜蛾细胞、粉纹夜蛾细胞、果蝇细胞或蚊子细胞。
16.进一步优选地,所述昆虫细胞还包含另一杆状病毒载体,所述另一杆状病毒载体包含aav的rep基因表达盒、外源基因以及位于所述外源基因两端的aav反向末端重复序列。
17.进一步优选地,所述外源基因为报告基因,所述报告基因为氯霉素乙酰转移酶编码基因、β-半乳糖苷酶编码基因、β-葡萄糖醛酸酶编码基因、海肾荧光素酶编码基因、碱性磷酸酶编码基因、萤火虫荧光素酶编码基因、绿色荧光蛋白编码基因和红色荧光蛋白编码基因中的至少一种。
18.进一步优选地,所述外源基因为编码药物多肽的基因,所述药物多肽为脂蛋白酯酶、载脂蛋白、细胞因子、白细胞介素和干扰素中的至少一种。
19.第四方面,本发明提供了一种重组腺相关病毒粒子,所述重组腺相关病毒粒子的衣壳包含本发明第二方面所述的腺相关病毒vp1衣壳蛋白突变体。
20.第五方面,本发明还提供了一种重组腺相关病毒粒子,其在本发明第三方面所述的昆虫细胞中产生。
21.第六方面,本发明提供了一种生产重组腺相关病毒粒子的方法,其是在能产生重组腺相关病毒粒子的条件下培养第三方面所述的昆虫细胞,然后回收制得的。
22.总体而言,通过本发明所构思的以上技术方案与现有技术相比,具有以下有益效果:本发明通过对野生型aav血清8型的vp1蛋白独有区域的氨基酸序列进行特定位点(第84、92、105位)的突变,而未在现有公开的蛋白质水解区域进行突变,能够减少昆虫细胞中vp1蛋白的降解,得到更合适比例的vp1、vp2和vp3蛋白,使得正确组装成raav的衣壳,从而提高利用杆状病毒/昆虫细胞表达系统生产raav的产率和效力。
附图说明
23.图1为本发明实施例2中重组杆状病毒载体ac-cap8表达vp蛋白的western blot检
测图。
24.图2为本发明实施例2中重组杆状病毒载体ac-cap9表达vp蛋白的western blot检测图。
25.图3为本发明实施例3中提供的aav血清8型和9型的cap基因编码的vp1蛋白n端第1位至137位氨基酸序列比对。
26.图4为本发明实施例3中重组杆状病毒载体ac-cap8、ac-cap8-q84k、ac-cap8-r92k和ac-cap8-q105k表达vp蛋白的western blot检测图。
27.图5为本发明实施例6中aav重组杆状病毒ac-rep-itr分别与ac-cap8、ac-cap8-q84k、ac-cap8-r92k、ac-cap8-q105k共感染宿主细胞后,纯化的重组aav病毒粒子进行sds-page的银染检测图,显示vp1、vp2、vp3三种衣壳蛋白。
具体实施方式
28.为了使本发明的目的、技术方案及优点更加清楚明白,以下结合附图及实施例,对本发明进行进一步详细说明。应当理解,此处所描述的具体实施例仅用以解释本发明,并不用于限定本发明。
29.术语解释
30.正如本文所用的,术语“可操作连接”是指多核苷酸(或多肽)序列以功能性关系的连接。当两段核苷酸序列置于功能性关系时,这两段核苷酸序列是“可操作连接”的。例如,转录调控序列(例如启动子)如果影响某基因编码序列的转录,则其与该基因编码序列可操作连接。
31.术语“编码区”,基因包括编码区和非编码区,编码区可作为一个开放阅读框(open reading frame,orf),从起始密码子开始,结束于终止密码子,其可包括内含子和编码蛋白的核苷酸序列;非编码区包括编码区前后的顺式作用元件,包括启动子、增强子等。
32.术语“表达盒”是指包含引入宿主细胞时可操作连接的编码序列和调控序列的核酸构建体,分别导致rna或多肽的转录和/或翻译。表达盒应理解为包括允许转录开始的启动子、目的基因开放阅读框和转录终止子。通常,启动子序列置于目的基因上游,与目的基因的距离与表达控制相容。
33.术语“内含子”又称间隔顺序,指一个基因或mrna分子中无编码作用的片段,是真核生物细胞dna中的间插序列。内含子序列被转录在mrna前体中,经过剪接被去除,最终不存在于成熟mrna分子中。根据剪接过程为自发还是要经过剪接体的加工,将内含子分为自剪接内含子和剪接体内含子。自剪接内含子是一种特殊的内含子,是一种核酶,可以通过自身作用被切除,来离开mrna。本发明涉及的内含子是剪接体内含子,这类内含子的剪除要有剪接体的帮助,内含子序列两端有剪接供体序列和剪接受体序列,是切断和重接位点处两旁的序列。剪接体是由核小rna(small nuclearrna,snrna)和蛋白质因子动态组成的核糖核蛋白复合体,剪接体识别mrna前体的剪接位点并催化剪接反应,将内含子完全剪除,上下游rna序列再重新连接。
34.所有已知腺相关病毒血清型的基因组结构非常相似,aav是单链dna病毒,基因组结构简单,全长约4.7kb,其基因组中包含rep基因表达盒、cap基因表达盒和位于基因组两端的aav反向末端重复序列(inverted terminal repeats,itr)。itr是基因组两端的125个
核苷酸的回文结构,能形成一个自我互补的倒t型发卡结构,是dna复制起始和包装重组aav基因组为感染性的病毒颗粒所需的顺式作用元件。aav作为缺陷型病毒,在没有辅助病毒的存在下不能够独立复制,因此aav只能定点整合在宿主细胞染色体中,呈潜伏状态。在辅助病毒存在的情况下,rep基因表达量增加可以将整合在宿主细胞染色体中的aav基因组拯救出来,大量复制得到aavdna,单链的raav基因组在vp衣壳蛋白的作用下被包装成具有感染性的病毒粒子。cap基因编码结构性的vp衣壳蛋白,其包含3个重叠开放阅读框,分别编码vp1、vp2、vp3三种类型的亚基,vp1、vp2和vp3含有不同的起始密码子,共用一个终止密码子,vp1和vp2共享vp3序列。vp1的n端具有一个保守的磷脂酶a2序列,该序列与病毒从体内逃逸有关,并对其感染性至关重要;vp2蛋白对于组装或者感染并非比不可少;vp3蛋白的核心由保守的β-桶基序组成,决定了不同血清型aav与宿主细胞作用受体的差异。野生型aav中三种蛋白的正确比例为3:3:54,约为1:1:10。rep基因编码rep78、rep68、rep52和rep40四个重叠的多功能蛋白,rep78和rep68蛋白参与aav的复制及整合,可以和itr中的特定序列结合;rep52和rep40蛋白具有解旋酶和atp酶的活性,在参与单链基因组的复制同时也参与病毒的装配。
35.发明人在研究中发现,野生型的aav血清8型表达产生的vp1蛋白在昆虫细胞中容易降解,使得相对含量明显降低。本发明旨在对cap基因的一个或多个密码子进行突变,使得该基因编码的vp1蛋白在生产过程中能够更加稳定,从而提高raav的产率。本发明提供的一种核酸,其包含编码腺相关病毒vp1、vp2和vp3蛋白的核苷酸序列,与野生型aav血清8型的衣壳蛋白相比,所述vp1蛋白的氨基酸序列包含在第84、92和105位中的一个或多个位点的突变,且所述vp2蛋白的氨基酸序列未突变。
36.中国专利文献cn113728108a提出移除aavvp1/vp2独特区域(即vp1和vp2中共有但vp3中缺乏的区域)中易于在昆虫细胞中蛋白质水解切割的位点,可导致较高纯度和均匀性的raav产物以及经改善的衣壳蛋白。还提出vp1独特区域(即vp1中存在但vp2或vp3中不存在的区域)含有pla2结构域,在该区域中的突变可增加pla2结构域在所得vp1蛋白中的酶活性。同时,griroda等人也提出vp1蛋白独特区域的突变会显著降低突变病毒的感染性(jgen virol.2002,83(5):973-978)。但是,现有技术均未提出vp1独特区域某些位点的突变能提高vp1蛋白对蛋白质降解的抗性。发明人意外发现aav血清8型的vp1独特区域(对应于seq id no.3的残基1至137)中第84、92和105位氨基酸中一个或多个发生突变,而vp2蛋白并不发生突变,能够降低vp1蛋白在生产过程中的降解,使得vp1、vp2和vp3蛋白具有更合适的比例,从而提高raav的包装率和滴度。
37.发明人发现相比野生型的aav血清8型,野生型的aav血清9型的衣壳蛋白vp1、vp2和vp3具有合适的比例,优选将aav血清8型的vp1蛋白的第84、92和105位氨基酸中的一个或多个突变成aav血清9型相同的氨基酸,即突变的vp1蛋白的氨基酸序列包含一个或多个选自以下的突变:q84k、r92k和q105k。
38.一些实施例中,所述vp1蛋白的氨基酸序列仅发生以上三个位点(第84、92和105位)中的一个或多个突变,其它序列均未突变。
39.在本发明构思的基础上,还可以引入内含子选择性剪接策略(参照发明人此前申请的专利cn113897396b)以调控aavcap基因重叠开放阅读框中vp1、vp2和vp3蛋白按照正确的化学计量表达。具体地,本发明核酸包含完整的cap基因表达盒,即所述核酸中还包含能
够在昆虫细胞中驱动转录的启动子,所述启动子的3’端可操作连接有内含子;所述vp1蛋白的起始密码子atg位于所述内含子的序列之中,或者,所述vp1蛋白对应的cap基因编码区缺失起始密码子且所述内含子位于atg中的任意相邻两个核苷酸之间;所述内含子不包含能够在昆虫细胞中驱动转录的启动子;
40.所述核酸在昆虫细胞中仅能转录得到一种前体mrna,且在转录后加工过程中,仅通过所述内含子的选择性剪接作用,使得所述vp1蛋白的起始密码子atg保留或缺失,或者在vp1蛋白对应的cap基因编码区中形成起始密码子atg,从而实现调控所述vp1、vp2和vp3蛋白的翻译表达。
41.一些实施例中,所述vp1蛋白的起始密码子atg位于所述内含子的序列之中,且所述内含子的3’端可操作连接有编码2a自剪切多肽的核苷酸序列。
42.该实施例在采用内含子调控策略时,需要对野生型aav血清8型vp1蛋白编码序列自身n端的内含子剪接位点进行突变,使得昆虫细胞中的剪接体无法识别该剪接位点。在转录后加工过程中,若昆虫细胞中的剪接体识别所述内含子的剪接位点并催化剪接反应,则去除mrna最前端aug-起始位点,核糖体从5’至3’识别vp2蛋白的起始密码子时,vp2蛋白得以翻译表达,由于vp2起始密码子是次优密码子acg,会造成核糖体扫描泄露,于是vp3蛋白得以翻译表达;若内含子剪接位点没有被识别,aug-起始位点没有被去除,则mrna从第一个aug处翻译,vp1蛋白表达。通过内含子具有一定概率的剪接作用,控制衣壳蛋白vp1、vp2和vp3的相对表达量。但是,当vp1蛋白翻译表达时,从mrna第一个aug起始密码子到vp1蛋白n端序列之间的一段额外序列(即内含子序列的一部分)也会被翻译,而影响vp1蛋白的正常表达,因此,本发明提供的cap基因表达盒中引入了2a自剪切多肽(2a self-cleaving peptides)的编码序列。
43.2a自剪切多肽是一类长18-22个氨基酸残基的肽片段,能诱导细胞内含有2a肽的重组蛋白自我剪切。这种肽都有一段的序列模体,经常会在最后甘氨酸(g)和脯氨酸(p)连接处导致核糖体无法连接,从而造成“剪切”的效果,使得2a自剪切多肽c端与vp1蛋白n端断开,以得到正常的vp1蛋白。根据不同病毒来源,目前共有四种常用的2a肽:t2a、p2a、e2a和f2a,以上四种2a肽都可以应用于本发明技术方案中,本发明的实施例以t2a为例。
44.本发明提供一种腺相关病毒vp1衣壳蛋白突变体,其由本发明提供的核酸编码得到。
45.本发明还提供一种昆虫细胞,其包含本发明提供的核酸,所述核酸为杆状病毒载体。所述昆虫细胞可以为任意昆虫细胞,例如但不限于草地贪夜蛾细胞(spodoptera frugiperda cell)、粉纹夜蛾细胞、果蝇细胞或蚊子细胞,优选为草地贪夜蛾细胞sf9。
46.本发明构思可应用于现有任何合适的杆状病毒/昆虫细胞表达系统中以生产raav,包括但不限于三杆状病毒系统(urabem等,hum gene ther,2002,13(16):1935-1943)、双杆状病毒(chen等,mol ther,2008,16(5):924-930;smithrh等,mol ther,2009,17(11):1888-1896)、onebac系统(mietzsch m等,hum gene ther,2014,25(3):212-222)和monobac系统(galibertl等,abstract at the esgct meeting,madrid/spain,2013,p074)。本发明实施例将突变引入双杆状病毒系统中,昆虫细胞中包含两种杆状病毒载体,一种包含突变的cap基因表达盒,另一种同时包含aav的rep基因表达盒、外源基因以及位于所述外源基因两端的aav反向末端重复序列。
cap9表达vp蛋白采用相同的调控策略,且使用的是同一内含子调控序列,两者产生的vp蛋白(vp1、vp2和vp3)的比例应该是一致的。发明人推测,可能由于aav血清8型vp1蛋白的结构不稳定,在sf9细胞中容易降解,从而导致vp1蛋白的检出量过低。
59.实施例3构建含有aav血清8型cap基因突变体表达盒的重组杆状病毒载体及检测vp蛋白(vp1、vp2、vp3)表达情况
60.从实施例2的结论看出重组杆状病毒载体ac-cap8异常的vp1蛋白表达含量,于是对aav血清8型和9型的cap基因编码的vp1蛋白氨基酸序列进行比对,aav血清8型的vp1蛋白氨基酸序列如seq id no.3所示,aav血清9型的vp1蛋白氨基酸序列如seq id no.4所示,发现aav血清8型和9型vp1蛋白的n端氨基酸序列存在差异,推断vp1蛋白的n端氨基酸序列的差异导致了重组杆状病毒载体ac-cap8的vp1蛋白表达含量异常。
61.为了验证上述推断,本实施例分别对aav血清8型cap基因编码的vp1蛋白的n端第84位、92位和105位氨基酸进行突变,分别突变成与aav血清9型vp1蛋白n端相对应的氨基酸,图3显示aav血清8型和9型的cap基因编码的vp1蛋白n端第1位至137位氨基酸序列,其中黑框标注出突变位点(q84k、r92k和q105k)。
62.参照实施例1,本实施例分别构建了含有aav血清8型cap基因突变体表达盒的重组杆状病毒载体ac-cap8-q84k、ac-cap8-r92k和ac-cap8-q105k。所述重组杆状病毒载体ac-cap8-q84k的cap基因编码的vp1蛋白氨基酸序列中第84位的谷氨酰胺突变成赖氨酸,相应的密码子cag突变成aag;所述重组杆状病毒载体ac-cap8-r92k的cap基因编码的vp1蛋白氨基酸序列中第92位的精氨酸突变成赖氨酸,相应的密码子cgg突变成aag;所述重组杆状病毒载体ac-cap8-q105k的cap基因编码的vp1蛋白氨基酸序列中第105位的谷氨酰胺突变成赖氨酸,相应的密码子caa突变成aaa。
63.参照实施例2,将本实施例制备的重组杆状病毒载体ac-cap8-q84k、ac-cap8-r92k和ac-cap8-q105k分别转染宿主细胞系培养获得重组杆状病毒,检测vp蛋白(vp1、vp2、vp3)表达情况。
64.图4为包含有cap基因表达盒的重组杆状病毒载体ac-cap8、ac-cap8-q84k、ac-cap8-r92k和ac-cap8-q105k的vp蛋白(vp1、vp2和vp3)western blot检测图。由图4看出,相较于vp1蛋白n端没有突变的ac-cap8,ac-cap8-q84k、ac-cap8-r92k和ac-cap8-q105k产生的vp1蛋白含量均有显著提高。由此可见,aav血清8型vp1蛋白n端氨基酸的定点突变能提高vp1蛋白的稳定性,从而显著增加vp1蛋白的相对含量。
65.实施例4构建含有aav包装必需元件rep基因表达盒和itr核心元件(itr-goi)的重组杆状病毒载体ac-rep-itr
66.为了测试重组杆状病毒载体ac-cap8、ac-cap8-q84k、ac-cap8-r92k和ac-cap8-q105k分别生产aav的效果,本发明采用two-bac系统进行aav的包装,其中一个杆粒含有aav的cap表达盒,另一个杆粒含有rep基因表达盒和itr核心元件,用两个重组杆状病毒共感染sf9细胞进行aav的包装。
67.首先,构建aav包装必需元件——rep基因表达盒和itr核心元件(itr-goi)。所述rep基因表达盒参照中国专利cn113897396b实施例2所述的方法构建,其核苷酸序列如seq id no.5所示;所述itr-goi的核苷酸序列如seq id no.6所示,本实施例中itr核心元件中的goi采用了红色荧光蛋白mcherry基因表达盒,即由minief1a启动子控制mcherry表达。
68.然后,将上述rep基因表达盒和itr-goi元件克隆到穿梭载体pfastbacdual上,制备转移质粒;将上述质粒分别转化dh10bac菌株中,通过tn7转座,最终得到包含生产raav所必需的rep基因表达盒和itr-goi元件的重组杆状病毒载体ac-rep-itr。
69.实施例5aav重组杆状病毒的制备
70.将实施例1、3和4中制备的重组杆状病毒载体ac-cap8、ac-cap8-q84k、ac-cap8-r92k、ac-cap8-q105k和ac-rep-itr分别转染宿主细胞系培养获得aav重组杆状病毒。
71.抽提上述重组杆粒dna分别转染sf9昆虫细胞,制备aav重组杆状病毒(bev)。转染后的sf9昆虫细胞成功产生bev,大量复制增殖的bev进一步感染导致sf9细胞发生明显的细胞病变效应(cpe)。收集发生cpe的sf9细胞培养上清液,其中含有大量bev,即为第0代bev(p0)。将制备得到的bev-p0以感染复数(moi=1)感染悬浮培养的sf9细胞,感染72小时后,细胞活性下降至50%以下,将细胞培养液1000g离心5min,分别收集培养上清和细胞沉淀,上清液标记为第1代bev-p1。
72.实施例6重组aav病毒粒子的纯化及其包装率、病毒滴度的检测
73.按照实施例5的操作继续扩大培养,直至用bev-p2的种毒ac-rep-itr分别与ac-cap8、ac-cap8-q84k、ac-cap8-r92k、ac-cap8-q105k按照感染复数(moi=1)共感染悬浮培养的sf9细胞进行raav的包装,包装体积为300ml~400ml。感染3天后监测细胞活性,活性低于50%,分别离心收获细胞沉淀和上清,将收获的细胞沉淀和上清分别纯化,细胞反复冻融3次裂解,5000rpm离心10min收集上清,在上清中加入核酸酶(benzonase)37℃水浴处理60min,处理后5000rpm离心10min。收集的细胞裂解液和收集的上清液peg沉淀,重悬后用碘克沙醇密度梯度离心分离纯化(方法参见aslanidi等,2009,proc.natl.acad.sci.usa,206:5059-5064)。最终纯化的成品病毒用80μl~190μlpbs重悬,取10μl纯化出的成品病毒跑sds-page胶,银染。
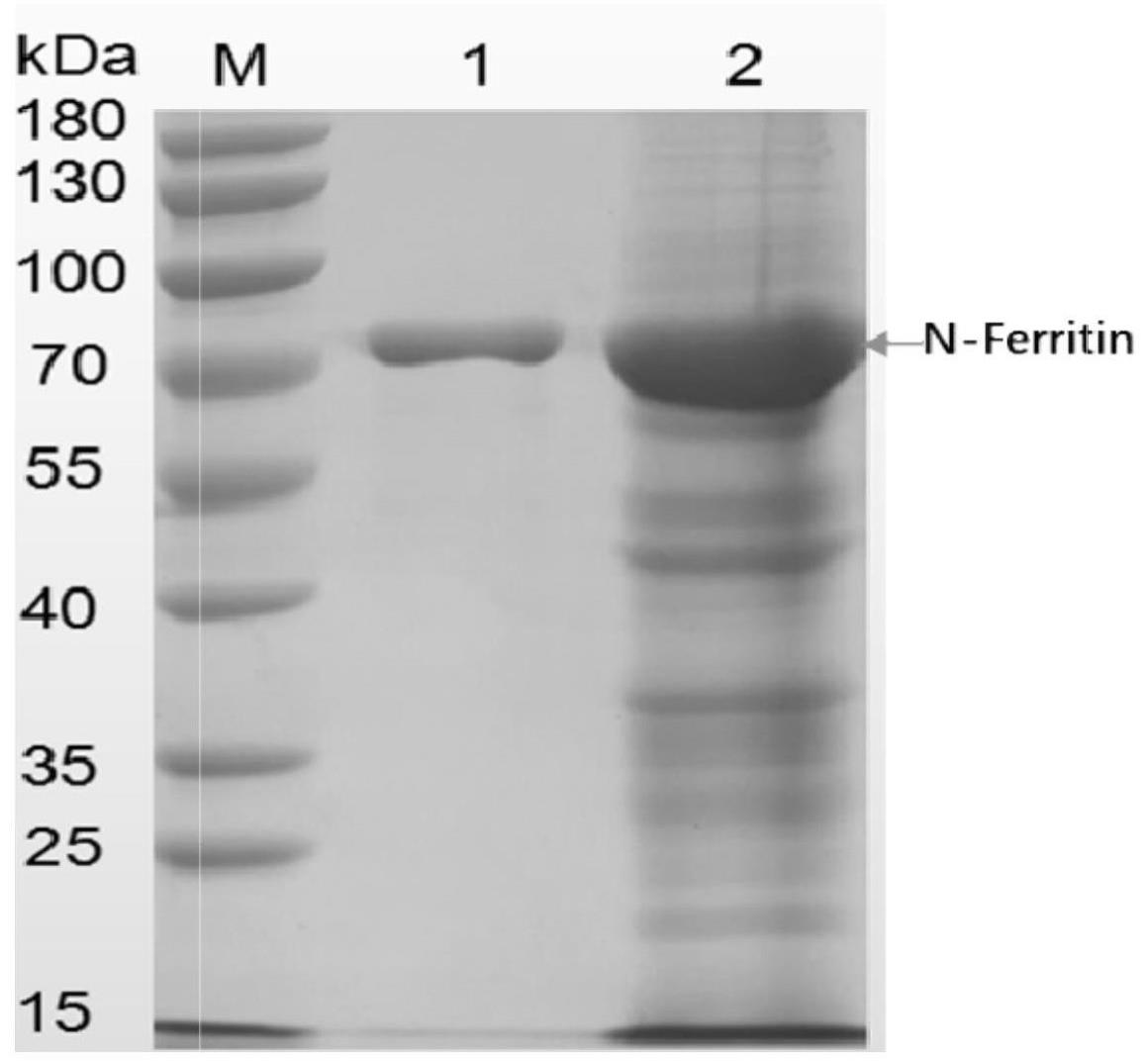
74.图5提供了aav重组杆状病毒ac-rep-itr分别与ac-cap8、ac-cap8-q84k、ac-cap8-r92k、ac-cap8-q105k共感染宿主细胞后,纯化的病毒粒子进行sds-page银染检测。由结果看出,相较于vp1蛋白n端没有突变的ac-cap8,由ac-cap8-q84k、ac-cap8-r92k和ac-cap8-q105k生产的aav颗粒中并入的vp1与vp2蛋白的比例接近1:1。
75.本实施例还采用q-pcr检测收获的重组腺相关病毒(raav)的包装率和上清滴度,滴度单位用vg/l(vg,virus genomes)表示。raav滴度的检测使用靶向itr序列的一对引物(q-itr-f:ggaacccctagtgatggagtt和q-itr-r:cggcctcagtgagcga)或者wpre序列的一对引物(q-wpre-f:ccgttgtcaggcaacgtg和q-wpre-r:agctgacaggtggtggcaat)。检测结果如表1所示。
76.表1不同重组腺相关病毒的包装率和上清滴度
[0077][0078]
由表1可以看出,相较于vp1蛋白n端没有突变的ac-cap8,由ac-cap8-q84k、ac-cap8-r92k和ac-cap8-q105k生产raav,其细胞包装率及产率更高。
[0079]
本领域的技术人员容易理解,以上所述仅为本发明的较佳实施例而已,并不用以限制本发明,凡在本发明的精神和原则之内所作的任何修改、等同替换和改进等,均应包含在本发明的保护范围之内。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。