tet2基因在调控肝脏损伤修复中的用途
技术领域
1.本发明涉及tet2(ten-eleven-translocation 2)基因的用途,尤其是在调控肝脏损伤修复中的用途。
背景技术:
2.tet2基因(ten-eleven-translocation 2)是tet家族中的一员,人tet2位于染色体4q24,有12个外显子。tet2蛋白在体内广泛表达,表达后主要定位于细胞核中;tet2蛋白被认为是一种α-酮戊二酸(α-kg)依赖的双加氧酶dna羟化酶,能催化5-甲基胞嘧啶(5mc)转化为5-羟甲基胞嘧啶(5hmc),从而使dna去甲基化,发挥表观遗传学作用;影响tet2表达的因素有很多。例如fe
2
,α-酮戊二酸,维生素c,视黄醇、视黄酸等都会对tet2的催化作用产生影响。由于tet2是一种α-酮戊二酸依赖的双加氧酶dna羟化酶,α-酮戊二酸作为tet2的底物直接影响了tet2酶的催化效果。fe
2
和维生素c同样影响底物与tet2的结合。广泛的证据证明,视黄醇、视黄酸可以促进tet2的表达。由于在tet2基因的第一个内含子内含有进化保守的视黄酸受体元件(rare),加入视黄醇或视黄酸后会引起tet2 mrna表达上升。
3.视黄醇(retinol,rol),又称维生素a,是一种机体必需的脂溶性维生素。约80%左右的视黄醇通过视黄酯(主要是棕榈酸视黄酯)的形式存储在肝脏中。全反式视黄酸(all-trans retinoic acid,atra)是视黄醇的生物活性代谢产物。全反式视黄酸是至关重要的信号分子,在发育和成年期间调节基因转录,细胞分裂和细胞凋亡。全反式视黄酸过多和过少都与有害作用有关,包括感染导致死亡的风险增加,生育力降低,阿尔茨海默氏病和发育缺陷。全反式视黄酸可以以配体的形式激活核类视黄醇受体rar和rxr,并以此来调节与细胞生长、分化、发育和体内平衡有关的一系列靶基因的表达。目前的研究认为,健康的机体内存在视黄醇、视黄酸代谢稳态,一旦打破这个平衡,就会激活视黄醇代谢信号通路,并引起肝脏纤维化损伤。
4.肝脏是执行许多重要生理功能的复杂器官。它具有巨大的再生潜力。由于饮食、卫生、环境、传染病等多种因素的作用,使得存在很多种肝脏疾病,给人们的生活造成很大的影响。如今慢性肝病是全球主要的健康负担,全世界每年约有200万人死亡。当肝脏因慢性肝病(如病毒性肝炎、酒精性脂肪肝、非酒精性脂肪肝或非酒精性脂肪性肝炎)而受损时,就会发生纤维化。迄今为止,阻止纤维化的有效治疗手段还很缺乏。肝纤维化共同威胁着人类与动物的生命健康。肝纤维化是各种因素引起慢性肝脏损伤的一种自我修复的病理过程,是肝脏对反复损伤的伤口愈合反应的结果。然而随着细胞外基质(ecm)的持续积累和重塑减少,纤维化最终会破坏肝脏的正常结构。如果得不到有效控制或不加以合理治疗,纤维化可发展为肝硬化、肝癌,最终导致器官衰竭甚至死亡。所以肝纤维化是肝病治疗的关键环节。
5.目前tet2在肝脏中已有部分研究,但是tet2具体在肝脏分化或病变过程中起着什么样的作用目前还不清楚。先前的研究表明,在3种肝损伤动物模型中,包括胆管结扎,四氯化碳注射以及蛋氨酸和胆碱缺乏饮食喂养动物模型中5hmc的降低。并且在肝纤维化动物模
型中tet2和tet3参与5hmc的下调。据报道,肝细胞癌(hcc)的实体癌组织的基因组5hmc含量总体降低,并且大量的数据显示在hcc中tet2无论是在mrna水平还是蛋白水平都是降低的。另外有研究报道乙型肝炎病毒x蛋白通过rela指导的与ezh2和tet2结合的活性dna脱甲基作用诱导epcam表达。据研究,tet2与hdac1相互作用以抑制e-cad表达,促进β-catenin核转位参与肝细胞癌过程。同时还有研究认为tet2通过mir-22-3p的调控抑制肝癌细胞的干性和转移,促进hcc的进展。
技术实现要素:
6.本发明通过研究tet2在肝脏损伤的修复或再生中所发挥的作用,从而为治疗肝脏损伤的药物研发提供新的靶点或理论基础。
7.为实现上述目的,申请人给野生型小鼠及tet2敲除小鼠饲喂ddc饲料,建立小鼠肝损伤模型。结果表明:经饲喂ddc饲料,相比与wt小鼠,tet2基因敲除小鼠肝重/体重比显著升高,h&e染色显示肝脏存在更严重的肝细胞坏死和胆汁淤积现象,且ast和alt水平均明显升高,从而验证了tet2基因敲除小鼠的肝损伤更为严重。
8.接下来申请人通过α-sma,desmin抗体的双标的免疫荧光染色、以及masson染色确定了tet2基因敲除小鼠有更严重的纤维化。
9.随后申请人通过转录组rna-seq分析差异表达基因的变化以及涉及的信号通路的变化,筛选tet2的下游基因。结果显示tet2基因敲除小鼠的视黄醇代谢信号通路显著变化,其中p450家族的视黄酸代谢酶cyp26a1以及cyp26c1的表达量显著下调,由此确定了tet2通过影响视黄醇代谢信号通路来参与肝脏纤维化的修复。
10.进一步,申请人通过q-pcr、免疫组化等实验技术,验证了转录组rna-seq得到的tet2影响cyp26a1的表达的结果。
11.最后申请人通过液相色谱检测,发现ddc损伤后,相比于野生型小鼠,tet2基因敲除小鼠肝脏组织匀浆中视黄酸的含量显著下降,从而使肝脏纤维化程度加重。
12.综上,tet2基因在动物肝脏损伤修复及再生中发挥十分重要的作用,可作为新的靶点,对肝脏纤维化的治疗和药物研发具有重要意义。
13.更详尽的技术方案参见具体实施例。
附图说明
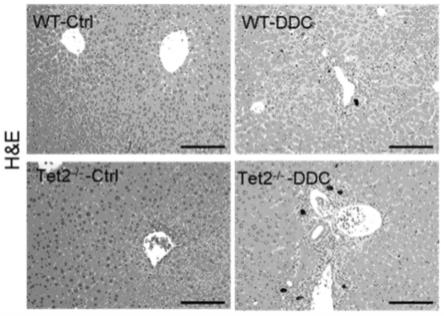
14.图1:正常组和ddc模型组的wt小鼠和tet2基因敲除小鼠肝组织病理变化。
15.图2:正常组和ddc模型组的wt小鼠和tet2基因敲除小鼠alt和ast检测结果。
16.图3:tet2基因敲除小鼠的cyp26a1、cyp26c1 mrna表达情况。
17.图4:tet2基因敲除小鼠的cyp26a1蛋白表达情况。
18.图5:tet2基因敲除小鼠肝脏组织匀浆中视黄酸的含量。
19.图6:免疫荧光双标染色检测tet2基因敲除对小鼠肝脏纤维化的影响。
20.图7:masson染色检测tet2基因敲除对小鼠肝脏纤维化的影响。
具体实施方式
21.下面通过具体实施例对本发明进行详细说明。
22.1.实验材料
23.1.1细胞和菌株
24.大肠杆菌感受态dh5α为本实验室保存菌株。
25.1.2实验动物
26.本研究所涉及的小鼠实验均采用c57bl/6小鼠完成,小鼠购自华中农业大学实验动物中心。实验过程中对动物的饲养和处理方法严格依照《实验动物管理条例》(1988年)、《实验动物管理与使用指南》(1996年)和华中农业大学实验动物管理和伦理委员会规定的步骤进行。
27.tet2基因敲除小鼠服务由上海南方模式动物科技股份有限公司提供。小鼠在华中农业大学试验动物中心饲养,按照规定程序进行扩繁,所有程序均遵守华中农业大学动物饲养和使用委员会准则。
28.手术相关材料包括:皮肤剪、组织剪、线剪、止血钳、持针钳、眼科剪、眼科镊、带线缝合针、医用酒精、无菌纱布、注射器。
29.1.3主要试剂和试剂盒
30.胎牛血清、opti-mem,青霉素-链霉素混合溶液(100
×
双抗)和胰蛋白酶均购自赛默飞世尔科技(中国)科技有限公司(gibco);
31.戊巴比妥钠购自西格玛生物奥利奇(上海)贸易有限公司(sigma-aldrich);
32.trizol及反转录试剂盒购自宝生物工程有限公司(takara);
33.sybr green realtime pcr master mix购自东洋纺(上海)生物科技有限公司(toyobo);
34.pcr引物由生工生物工程(上海)股份有限公司合成;
35.pcr master mix购自北京艾德莱生物科技有限公司产品;
36.dna marker购自北京艾德莱生物科技有限公司产品;
37.lipofectamine 2000购自英潍捷基(上海)贸易有限公司(invitrogen);
38.蛋白酶k、购自美国纽英伦生物技术有限公司(neb);
39.pcr产物回收试剂盒、普通质粒提取试剂盒(plasmid mini kit i)购自omega生物科技公司;
40.无内毒素少量质粒提取试剂盒(endo-free plasmid mini kit ii)购自omega生物科技公司;
41.谷草转氨酶(ast/got)检测试剂盒、谷丙转氨酶(alt/gpt)检测试剂盒购自南京建成生物工程研究所;
42.ripa裂解液、5
×
sds-page蛋白上样缓冲液、bca蛋白浓度测定试剂盒及dapi购自碧云天生物技术有限公司;
43.ddc(msds 137030)、丙烯酰胺、双丙烯酰胺、甘氨酸、三溴乙醇(cas:75-80-9)、trizma base、过硫酸铵、temed和吐温-20均购自西格玛奥德里奇(上海)贸易有限公司(sigma-aldrich);
44.tbs缓冲液、显影液、定影液购自武汉塞维尔科技有限公司;
45.脱脂奶粉和sds购自biosharp公司;
46.pvdf膜购自merck millipore公司;
47.抗tet2抗体(catalog nos:61389)购自active motif公司;
48.抗gapdh抗体(proteintech,sc-293335)购自santa cruz生物公司;
49.辣根过氧化物酶(hrp)标记的羊抗兔igg二抗(sc-2004)购自santa cruz生物公司;
50.鼠igg抗体购自博士德生物技术有限公司;
51.辣根过氧化物酶(hrp)标记的羊抗鼠igg二抗(sc-2005)购自santa cruz生物公司;
52.羊抗兔(h l)二抗af488(a11008),羊抗鼠(h l)二抗af555(a21424)购自赛默飞世尔科技(中国)有限公司;
53.化学发光底物clarity enhanced chemiluminescence,ecl购自武汉聚能译通生物有限公司;
54.甘油、氯化钠(nacl)、乙二胺四乙酸钠(edta)、三羟甲基氨甲烷(tris)、氯仿、异戊醇、甲醇、吐温-20和无水乙醇均购自国药集团化学试剂有限公司;
55.细胞冻存液的配置:
56.将血清,基础培养液和二甲基亚砜按照5:4:1的比例进行混匀,4℃冰箱保存;
57.lb液体培养基配制:
58.取胰蛋白胨1g,酵母提取物0.5g,nacl 1g溶于双蒸水,用5mol/l的naoh调至ph 7.0,定容至100ml,高压灭菌,4℃保存;
59.lb固体培养基配制:
60.取胰蛋白胨1g,酵母提取物0.5g,nacl 1g,琼脂粉1.5g溶于双蒸水,用5mol/l的naoh调至ph 7.0,定容至100ml,高压灭菌,倒入平板冷却至室温,4℃保存;
61.氨苄青霉素溶液:
62.10mg的amp溶解于10ml双蒸水中,一次性过滤器除菌,分装到1.5ml的ep管中,-20℃保存;
63.depc处理水(浓度0.1%):将1ml的depc加入1000ml双蒸水中,在37℃条件下混合过夜后高压灭菌备用;
64.20mg/ml蛋白酶k储存液:
65.200mg蛋白酶k溶于10ml双蒸水,分装到1.5ml的ep管中,-20℃保存;
66.电泳缓冲液1000ml:
67.称取tris-base 3.02g,甘氨酸19g,sds 1g,加水定容到1000ml;
68.电转缓冲液1000ml:
69.称取甘氨酸14.4g,tris-base 3.03g,甲醇200ml,加水定容到1000ml;
70.洗膜溶液tbst 1000ml:
71.称取nacl 8.8g,kcl 0.2g,tris 3g,加吐温-20 500ul,用超纯水先定容到800ml,调ph至7.4,最终定容到1000ml;
72.封闭液:脱脂奶粉5g,用100ml的1
×
tbst混匀后使用;
73.组织裂解液1000ml:
74.称取sds 5g,加热溶解,后加入tris 1.21g,nacl 5.84g,edta.2na 3.72g,加纯水至800ml左右,调ph至8.0,最终定容至1000ml;
75.te溶液1000ml:
76.称取1.21gtris,3.72gedta.2na,加纯水至800ml左右,调ph至8.0,最终定容至1000ml。
77.1.4主要仪器和设备
78.移液器(plus,德国eppendorf公司产品);
79.电子天平称(ax124zh/e,奥豪斯仪器有限公司产品);
80.雪花制冰机(xueke,武汉科昊佳生物科技有限公司产品);
81.磁力搅拌器(78-1,金坛市医疗器械厂产品);
82.电热鼓风干燥箱(dgx-9143b-1,上海福玛实验设备有限公司产品);
83.电热恒温培养箱(dnp-9162型,上海精宏实验设备有限公司产品);
84.二氧化碳恒温培养箱(赛默飞世尔科技有限公司产品);
85.优普系列超纯水器(upt-11-20t,四川优普超纯有限公司产品);
86.数显恒温水浴锅(hh-6,国华电器有限公司产品);
87.酶标仪(mk3型,赛默飞世尔科技有限公司产品);
88.涡旋振荡器(mx-s,scilogex产品);
89.超威量分光亮度计(ds-11,denovix产品);
90.普通pcr仪(a200,杭州朗基科学仪器有限公司产品);
91.微孔板迷你离心机(mini-p25,杭州奥盛仪器有限公司产品);
92.荧光定量pcr仪(abi quantstudio3荧光定量pcr仪,美国应用生物系统公司产品);
93.台式高速冷冻离心机(5404,德国eppendorf公司产品);
94.﹣20℃冰箱(dw-yl270,中科美菱产品);
95.﹣80℃超低温冰箱(dw-86l626,青岛海尔特种电器有限公司产品);
96.超净工作台(hdl apparαtus,哈尔滨市东联公司产品);
97.立式自动电热压力蒸汽灭菌器(lx-b50l型,合肥华泰医疗设备有限公司产品);
98.室温水平摇床(sk-r1807-e,scilogex产品);
99.电泳仪及电泳槽(dyy-6c型,北京市六一仪器厂产品);
100.高通量组织研磨器(scientz-48,宁波新芝生物科技股份有限公司产品);
101.旋转式切片机(leica,leicarm2245,德国leica公司产品);
102.凝胶成像系统(newbio indusrty产品);
103.近红外成像系统(g:box,syngene产品);
104.倒置显微镜(奥林巴斯产品);
105.激光共聚焦显微镜(zeiss lsm800)由华中农业大学微生物国家重点实验室公共平台提供。
106.2.实验方法
107.2.1细胞培养
108.2.1.1细胞培养及传代
109.293t细胞用含10%胎牛血清、100iu/ml青霉素-链霉素的h-dmem培养液中培养。所有细胞均在37℃、含5%co2的湿式温箱中孵育。
110.保持细胞室的无菌环境,提前复温培养液,严格执行无菌操作。根据细胞的生长状态及时更换培养液,待细胞生长至80%左右,进行传代。具体操作如下:
111.弃掉旧的培养基,用pbs轻柔清洗细胞2次,以便去除残余培养液和死细胞。依据培养皿规格加入适量0.25%的胰蛋白酶至细胞培养皿底(如10cm皿通常加入1ml胰酶),缓慢摇动细胞培养皿以确保胰酶完全接触细胞层。倾斜培养皿轻轻吸走胰蛋白酶,盖好皿盖,将培养皿置于恒温培养箱消化1-2min,于倒置显微镜下观察,发现细胞逐渐收缩变圆后,迅速向培养皿中加入一定量新鲜的完全培养基,终止胰酶的消化。轻拍细胞培养皿使细胞自然脱落,再用移液枪反复轻吹细胞悬液使之分散成单个细胞均匀分散于培养基中。注意吹打的力度要适宜,尽量不产生气泡。对细胞进行计数,根据需要进行分皿接种,轻轻摇晃使细胞均匀分散于培养皿中,最后将培养皿放回co2培养箱培养。
112.2.1.2细胞冻存
113.取处于对数期的生长状态良好的细胞,用无菌pbs洗两遍,用0.25%的胰蛋白酶将其消化脱落,吹打成单细胞混匀液,对其进行计数并离心。离心后弃上清液,轻弹细胞使之从管壁脱落,按照1*10
^
从管壁脱落,按照1*106个/ml加入相应量的细胞冻存液。随即轻轻混匀冻存液中的细胞,按照1ml/管转移至细胞冻存管中。4℃冰箱垂直放置30min,-20℃冰箱放置30min,-80℃冰箱过夜冷冻,次日转移至液氮罐中长期保存,并记录冻存位置。
114.2.1.3细胞复苏
115.从液氮中迅速取出冻存的细胞,立即放入37℃恒温水浴锅中快速摇晃溶解,解冻时间控制在90s内。将冻存管消毒后在超净工作台内转移至已含预温的培养液的离心管中,1000rpm/min,5min离心后丢弃上清液。轻轻晃动离心管使细胞脱落,加入1ml完全培养基对细胞进行重悬,将细胞悬液均匀滴加至含有完全培养液的培养皿中,轻轻摇晃使细胞均匀分布在细胞培养皿,最后将培养皿放于37℃的培养箱贴壁培养。24h后观察细胞状态,更换新鲜培养液或者进行细胞传代。
116.2.1.4小鼠基因型鉴定
117.将杂合tet2基因敲除小鼠雄鼠与杂合tet2基因敲除小鼠雌鼠进行杂交,获得的纯合tet2基因敲除小鼠子代用于后续实验。
118.小鼠基因组dna提取
119.1)切耳朵的一块小组织(例如:0.5cm2组织)并放置在标记的ep管中。
120.2)添加500ul组织裂解液和10ul proteinase k到每一管,涡旋混匀。
121.3)55℃培养过夜。确保组织被裂解。
122.4)添加250ul 5m nacl到每管。
123.5)剧烈摇晃大于20次。
124.6)最大速度离心10min。
125.7)仔细转移上清液到装有冰冻的100%乙醇700ul的ep管中并涡旋轻轻地混匀,可能会出现一些dna像白色绒毛,如若没有,不用担心。
126.8)最大转速离心ep管5min,应该会看到白色绒毛。
127.9)移除上清液切勿吸取到dna并往里面加1ml 70%乙醇(移除盐remove salt)。
128.10)每个样品都换枪头避免污染。
129.11)上下颠倒混匀(
×
50)。
130.12)最大转速离心5min。
131.13)移除上清液。小心不要吸取到dna沉淀,dna沉淀可能没有贴到管壁。尽可能移除上清液。每个样品都换枪头避免污染。
132.14)重悬之前,在干燥空气中室温放置30min。
133.15)50ul te重悬(可能需要放置在管架上半小时)。最小速度涡旋有助于混匀。
134.保存样品4℃(短期使用)或-20℃(长期使用)。
135.聚合酶链式反应(pcr)鉴定小鼠基因型
136.将提取出的小鼠基因组dna作为模板,taq dna polymerase分别扩增:
137.表1 pcr反应体系(10ul)
[0138][0139]
表2 pcr引物序列
[0140][0141]
将反应体系混匀后瞬离,置于pcr仪中扩增。反应条件如下:
[0142]
表3 pcr扩增条件
[0143][0144]
dna凝胶电泳和成像
[0145]
称取0.48g琼脂糖粉末于烧杯中,加入40ml 1
×
tae buffer,配制1.2%琼脂糖凝胶用于电泳。向10ul反应后的pcr体系里加入2ul 6
×
loading buffer,混匀后点到凝胶孔
中,以4ul dna标准分子量dl2000 marker作为参照,15v/cm电泳,电泳结束后,在凝胶成像系统中观察,拍照保存图片。
[0146]
最终得到野生型小鼠pcr产物长度为1434bp,纯合小鼠pcr产物长度为749bp。
[0147]
2.2动物损伤模型的构建
[0148]
取8周龄健康雄性野生型小鼠和tet2基因敲除小鼠,于12h明暗交替的饲养房饲养,2-3只每笼。将ddc加工为饲料进行ddc饲料饲喂,用于构建胆管细胞损伤为主的肝损伤模型。ddc饲料持续饲喂14d。为保证饲料新鲜,每两天换一次。取材前2h注射brdu(100mg/kg in pbs)。在慢性ddc损伤14d后,摘眼球采血收集血清,随后断颈处死小鼠,采集肝脏组织用于提取rna和蛋白,另外采集部分肝脏放置于福尔马林溶液中固定。
[0149]
2.3生物信息学分析相关软件和数据库
[0150]
2.3.1生物信息学分析相关软件
[0151]
1)ncbi数据库:http://www.ncbi.nlm.nih.gov/用于基因序列查询,其中blast软件可以用来进行多序列对比及同源性分析;
[0152]
2)网站primerbank(https://pga.mgh.harvard.edu/primerbank/index.html)以及软件primer premier 5.0用于引物设计;
[0153]
3)graphpad prism 7.0软件用于图形绘制。
[0154]
2.3.2数据资源
[0155]
本研究所用的tet2 chip-seq数据及相关数据下载自geo(gene expression omnibus)数据库(accession numbers:srr7359640,srr7359641and srr7359644)。本研究所用的5hmc chip-seq数据及相关数据下载自geo数据库(accession number:srr14088633and srr14088619)。
[0156]
2.4rna-seq和chip-seq
[0157]
2.4.1rna-seq和数据分析
[0158]
rna-seq在贝纳生物技术有限公司进行。主要的试验方法为:首先利用trizol法分别提取ddc损伤的wt小鼠和tet2基因敲除小鼠的肝脏rna样本。随后利用琼脂糖凝胶电泳、nanodrop分光光度计、qubit、agilent 2100对rna样品质量进行检测。随后利用ultra
tm
rna library prep kit(neb,美国)进行文库构建。测序在illumina hiseq平台上进行。测序完成后,利用bowtie v2.2.3软件将得到的测序数据在小鼠基因组(ncbi build 38/mm10)进行注释。利用htseq v0.6.1软件对每个基因的reads数进行计数,计算每个基因的fpkm。差异表达基因通过deseq r package(1.18.0)软件以fdr《0.05和fold-change》1.5或《0.7进行鉴定。最后利用goseq和kegg对差异表达基因进行基因功能和通路分析。
[0159]
2.4.2chip-pcr和chip-seq数据分析
[0160]
chip
[0161]
1)称新鲜冰冻组织,切成1-3mm3小块,共80mg;
[0162]
2)将组织块转移到50ml离心管中,加入10ml无菌pbs;
[0163]
3)加入100ul甲醛至终浓度为1%,室温下置于摇床上摇15-20分钟;
[0164]
4)稀释10x glycine至1x(向上步的50ml离心管中加入1ml 10x glycine),4℃转10分钟,然后去离心,100g 4℃5分钟;
[0165]
5)弃上清,洗两次沉淀,用45ml、25ml冰pbs各洗一次,离心,100g 4℃5分钟,弃上
清,取两个2ml破碎管进行标记;
[0166]
6)将组织块转移至2ml破碎管中,再加入约2ml冰pbs,每管加一颗磁珠,之后放入组织破碎仪进行破碎匀浆,离心,100g 4℃5分钟,弃上清,加入pbs,每管分装成两管,1ml/管(1ml用作超声破碎来摸索最佳超声时间,另一管先保存起来做正式实验);
[0167]
7)用700ul 1%sds细胞裂解液重悬细胞,(并加入7ul 100x pmsf与7ul 100
×
pki),冰上孵育20分钟;
[0168]
8)静止完成即可取一套管子去摸索超声条件,另一套放于4℃保存备用;
[0169]
9)超声破碎的时候,破碎时间设置为:2分钟、4分钟、8分钟、12分钟、15分钟以及20分钟,从每个里面分别吸取20ul至离心管中,放在冰上等待跑胶;
[0170]
10)配制1.2%的琼脂糖凝胶,将第9步中的6个pcr管里的样品进行dna电泳,最终选取弥散型亮条带集中在250-700bp那管样品的超声破碎时间来破碎第8步中的备用样品。
[0171]
11)将超声破碎后的样品取至新的1.5ml离心管中,13000g,4℃,5分钟,离心取上清,从每个样品管中取400ul至两个新的2ml离心管中,每管200ul,置于冰上备用(其中一管后续加igg抗体,另一管加h4k16ac抗体);
[0172]
12)提前30分钟将蛋白a/g琼脂糖珠(protein a/g agarose)置于4℃里的旋转摇床上转动,使其混匀;
[0173]
13)配制含有1mm pmsf的chip dilution buffer,向上步含200ul的2mlep管中各加入1.8ml的含有1mm pmsf的chip dilution buffer,使此时终体积为2ml,混匀后取出100ul液体于一新的离心管中,放于-20℃保存,后续用作input
[0174]
14)向上步的1.9ml液体中各加入70ul蛋白a/g琼脂糖珠,之后在4℃的旋转摇床上缓慢转动30分钟,目的是为了减少蛋白a/g琼脂糖珠与目的蛋白或目的dna序列的非特异性结合;
[0175]
15)1000g,4℃,离心5分钟,将上清转移到一个新的2ml离心管中,并加入一抗(0.5ug),放在4℃旋转摇床上缓慢混匀过夜;
[0176]
16)第二天早上,向每管中加入60ul的蛋白a/g琼脂糖珠,于4℃旋转摇床上缓慢转动1小时,以沉淀一抗识别的蛋白或相应的复合物;
[0177]
17)1000g,4℃,离心1分钟,非常小心地去除上清液体,切勿触碰到沉淀。随后依次用如下溶液对沉淀进行洗涤,每次洗涤液的用量为1ml,每次在4℃旋转摇床上缓慢转动洗5分钟,随后1000g,4℃,离心1分钟,非常小心地去除上清液体,切勿触碰到沉淀。
[0178]
a.low salt immune complex wash buffer洗一次
[0179]
b.high salt immune complex wash buffer洗一次
[0180]
c.licl salt immune complex wash buffer洗一次
[0181]
d.te buffer洗两次
[0182]
18)新鲜配制elution buffer(1%sds,0.1m nahco3),也可直接用试剂盒中的;
[0183]
19)完成上述所有洗涤步骤后,向沉淀中加入250ul的elution buffer并充分涡旋混匀,室温转动洗脱5分钟;
[0184]
20)室温,1000g,离心1分钟,将上清转移到一个新的离心管中,向沉淀中再加入250ul的elution buffer并充分涡旋混匀,室温转动洗脱5分钟;
[0185]
21)室温,1000g,离心1分钟,将上清取出与上步的上清合并,共约500ul;
[0186]
22)在500ul上清中加入20ul 5m nacl,向200ul input中加入10ul 5m nacl,混匀,放于65℃水浴锅孵育4个小时,以去除蛋白和基因组dna之间的交联;
[0187]
23)在约520ul的样品中加入10ul 0.5m edta、20ul 1m tris ph 6.5和1ul 20mg/ml的蛋白酶k(input样品中也按比例加入以上试剂)。混匀后放于65℃水浴锅孵育45分钟,之后进行纯化。
[0188]
chip产物纯化
[0189]
1)将每管样品放入4ml的离心管中并且做好标记;
[0190]
2)向每管中加入6倍体积的cp buffer,充分混匀;
[0191]
3)混匀后取700ul加入dna吸附柱中,15000g,离心1分钟,弃废液,一直重复,直至上步管中的液体耗尽;
[0192]
4)向弃去废液的吸附柱中加入700ul的dna wash buffer,15000g,离心1分钟。
[0193]
5)重复第4步并弃废液;
[0194]
6)将空的管子15000g,离心2分钟,并重复一次;
[0195]
7)将吸附柱放于55-60℃烘箱5-8分钟烘干;
[0196]
8)将吸附柱插入1.5ml离心管中,加入30ul elution buffer,静置10分钟;
[0197]
9)15000rpm,离心1分钟;
[0198]
10)将离得的液体再加入吸附柱中,重复步骤8、9;
[0199]
11)提前30分钟将蛋白a/g琼脂糖珠(protein a/g agarose)置于4℃里的旋转摇床上转动,使其混匀;
[0200]
12)涡旋混匀,测浓度。
[0201]
chip-seq数据分析
[0202]
chip-seq数据分析所用数据来自于geo数据库。采用fastqc对读取序列进行筛选,使用bowtie2与小鼠基因组(ncbi build 38/mm10)进行注释。利用macs version 2.1.0检测峰值。数据集的q值的阈值为0.05。最后利用igv软件分别对tet2结合峰和5hmc结合峰的分布进行可视化分析。
[0203]
2.5qrt-pcr检测基因表达
[0204]
为了研究tet2在肝脏纤维化过程中作用,我们分别对野生型小鼠和tet2基因敲除小鼠用ddc构建胆管细胞损伤为主的肝损伤模型,然后对损伤后的肝脏组织进行总rna和cdna制备,随后用所获得的cdna进行qrt-pcr检测基因表达。
[0205]
2.5.1rna提取
[0206]
1)从-80℃冰箱中取出大鼠肝脏组织约20mg,每管加入1ml trizol裂解液,1颗钢珠,在组织破碎仪中将组织击打呈匀浆状,冰上静置5min;
[0207]
2)每管加入0.2ml氯仿,用力震荡15sec,静置2min;
[0208]
3)4℃离心机,12000rpm/min,离心15min,取上清至新的离心管(注意不要取到中间蛋白层);
[0209]
4)在上清中加入0.5ml异丙醇,将管中的液体轻轻混匀,室温静置10min;
[0210]
5)4℃,12000rpm/min,离心10min,观察沉淀位置,弃尽上清;
[0211]
6)加入1ml 75%乙醇(depc水配制),上下颠倒离心管,轻轻洗涤沉淀;
[0212]
7)4℃,7500rpm/min,离心5min,观察沉淀位置,弃尽上清;
[0213]
8)沉淀自然晾至将干未干,加入适当的depc水溶解;
[0214]
9)取少许检测rna纯度(od260/280)及完整性,记录rna浓度,取部分立即反转录成cdna,剩余于-80℃保存备用。
[0215]
2.5.2cdna制备
[0216]
以提取的rna为模板,进行反转录(两步法)
[0217]
反转录(reverse transcription,rt)的体系(10ul)为:
[0218]
第一步:
[0219]
表4反转录第一步反应体系(10ul)
[0220][0221]
反应条件为:42℃2min,降温至4℃,之后迅速置于冰上。
[0222]
第二步:
[0223]
表5反转录第二步反应体系(20ul)
[0224][0225]
反应条件为:37℃15min,85℃5sec,降温至4℃。获得的cdna进行pcr或定量pcr,剩余置于-20℃保存备用。
[0226]
2.5.3real-time pcr
[0227]
以cdna为模板进行real-time pcr扩增,引物设计见表8。在检测目的基因mrna水平时,设计以鼠源36b4为内参基因,同时每个样品做2个重复孔。反应体系如下:
[0228]
表6反转录第二步反应体系(10ul)
[0229]
[0230][0231]
反应扩增条件如下:
[0232]
表7 real-time pcr扩增条件
[0233][0234]
整个反应过程中的荧光信号的变化由abi quantstudio3实时荧光定量pcr仪检测。
[0235]
利用读取的荧光ct值,使用mrna表达=2
‑△△
ct方法计算基因相对表达水平(livak and schmittgen,2001)。
[0236]
表8 real-time pcr引物
[0237][0238]
2.6western blot检测细胞/组织中蛋白表达
[0239]
2.6.1总蛋白提取
[0240]
细胞或组织总蛋白提取按照western细胞裂解液说明书进行。将细胞用冰冻的pbs
洗2遍,吸干残液,置于冰上。根据细胞量配制含有1mmol/l的pmsf及1
×
蛋白酶抑制剂的适量细胞裂解液并加入细胞培养皿中,裂解15分钟后冰上刮下细胞,收集到离心管中,4℃离心机离心10min,15000rpm/min。吸取上清进行蛋白浓度测定。冰冻组织需加入适量含蛋白酶抑制剂和pmsf的裂解液后进行组织破碎,破碎完冰上裂解15min之后,离心取上清。
[0241]
2.6.2蛋白浓度测定
[0242]
1)按照碧云天bca蛋白浓度测定试剂盒说明书进行,具体步骤如下:
[0243]
2)根据样品数量,按照每孔200ul,将a液和b液按照50:1的比例混合配制成bca工作液,充分混匀。bca工作液室温24h内稳定;
[0244]
3)完全溶解蛋白标准品,取储液适量用细胞裂解液稀释标准品,使终浓度为0.5mg/ml;
[0245]
4)将标准品按0ul、1ul、2ul、4ul、8ul、12ul、16ul、20ul,加到96孔板的标准品孔中,用细胞裂解液补足20ul;
[0246]
5)样品以细胞裂解液为溶剂,进行10倍稀释后加入到样品孔中;
[0247]
6)各孔中加入200ul bca工作液,锡箔纸包裹孔板,37℃温箱放置30min;
[0248]
7)测定a562 nm的波长,根据标准曲线计算出蛋白浓度并按照稀释倍数换算出样品原始浓度;
[0249]
8)取适量体积的蛋白样品,加入5
×
loading buffer,以细胞裂解液为溶剂,将样品配制成40ug/10ul浓度的工作液,100℃煮沸5-10min,混合瞬离,-80℃保存备用。
[0250]
2.6.3western blotting(wb)分析
[0251]
1)配胶:按说明书制备12%分离胶和5%浓缩胶;
[0252]
表9分离胶配制体系(10ml)
[0253][0254]
表10浓缩胶配制体系(4ml)
[0255][0256]
2)倒胶:夹好灌制聚丙烯酰胺凝胶的玻璃板,将制好的12%的sds丙烯酰胺分离胶约5ml迅速灌入两块玻璃板的间隙中,留出灌注浓缩胶所需空间,在分离胶上加入一层无水乙醇,置室温聚合30min,分离胶聚合后,倾出覆盖层液体,用纸巾吸净残留液体,再将刚制好的5%浓缩胶灌入分离胶上,立即插入梳子,置37℃聚合30min,待浓缩胶全部聚合后移出梳子,固定于电泳装置;
[0257]
3)电泳:往电泳装置加入适量1
×
tris-甘氨酸电泳缓冲液,取出处理好的样品上样,每孔上10ul。接通电源,电压调到80v,20min,再将电压调到120v,等溴酚蓝指示剂跑到分离胶底部(约需2h)时断电,结束电泳;
[0258]
4)电转:当sds-page电泳结束后,取sds-page电泳后的凝胶,依照目的蛋白分子量与marker蛋白分离条带,切下含目的蛋白和内参蛋白β-actin蛋白的胶,直接用bio-rad公司的mini trans-blot electrophoretic cell转印装置将蛋白以200ma电流,电转印2h,电转到pvdf膜上。具体操作如下:戴上手套,剪8张1层滤纸和1张pvdf膜,pvdf膜的大小要略大于凝胶。在干净托盘中加入少量转膜缓冲液,把滤纸和海绵浸泡于其中。pvdf膜在甲醇中浸泡2min后,也移入转膜缓冲液中。铺上4张1层海绵,滤纸,用玻璃棒赶走气泡,将电泳后的胶轻放在滤纸上,玻璃棒赶走气泡,贴上pvdf膜,再铺上4张1层滤纸及海绵,玻璃棒赶走气泡,将其整体转移到转膜夹上,在冰水混合物的环境下,200ma恒流,转膜2h;
[0259]
5)封闭:配制封闭液,在30ml 1
×
tbst中加入1.5g脱脂奶粉,在磁力搅拌器上充分混匀。将膜用镊子夹出,平铺在封闭液中,蛋白面朝下,置于摇床上室温孵育2h;
[0260]
6)一抗与靶蛋白结合:将膜从封闭液中夹出剪去边缘,加入用封闭液稀释的一抗溶液,摇床上4℃孵育过夜;
[0261]
7)二抗与靶蛋白结合:在保鲜盒内加入8ml 1
×
tbst洗3次,每次5min,再洗2次,每次10min。倒掉冲洗液,加入用封闭液稀释的二抗溶液,摇床上室温孵育90min;
[0262]
8)显色:在保鲜盒内加入8ml 1
×
tbst洗3次,每次5min,再洗2次,每次10min。将pvdf膜放入化学发光底物的a液和b液的等量混合液中进行避光显色,用近红外成像系统进行曝光。
[0263]
2.7组织染色
[0264]
2.7.1取材、脱水浸蜡、包埋及切片
[0265]
1)取材:小鼠,心脏取血后,立即解剖小心取出肝脏,切取组织时应使用锋利的刀、
剪,切取组织块时,从刀的根部开始向后拉动切开组织;
[0266]
2)固定:将所取组织在10%中性福尔马林中固定1-2天;
[0267]
3)洗涤:自来水冲洗组织块过夜;
[0268]
4)脱水:次日将冲洗好的组织于50%乙醇、70%乙醇、80%乙醇各12h,90%乙醇10h,95%乙醇ⅰ、95%乙醇ⅱ各2h,100%乙醇ⅰ及100%乙醇ⅱ分别1h;
[0269]
5)透明:将脱水好的组织块置于二甲苯与乙醇混合液(1:1)中30min、二甲苯ⅰ3min、二甲苯ⅱ9min,透明好的组织块应该呈现棕黄色透明态;
[0270]
6)浸蜡:将透明好的组织块在二甲苯和石蜡混合液(1:1)中45℃温箱浸蜡30min;石蜡ⅰ中40min、石蜡ⅱ中1h、石蜡ⅲ中1h;
[0271]
7)包埋和蜡块的修整:将包埋盒在温箱中预热,溶解好的蜡ⅲ倒入包埋盒中,然后将组织块轻轻放入应避免产生气泡。将包埋好的蜡块修整以利于切片;
[0272]
8)切片:切片前将包埋好的组织样本置于-20℃冷冻数分钟,待组织达到适当硬度即可切片,切片时组织的冷冻程度是切片的关键环节,冻得过硬,切片呈碎屑状;冻得不够,切片呈粥糜状或切不成片。切片厚度最好为4-5μm,太厚贴片不牢固,更不利于镜下观察。包埋使组织上、下方有一定的边,有利于连续切片,而且展片时可以避免组织皱缩;
[0273]
9)展片、烤片:水浴展片,捞片,将切片小心贴附于载玻片上,捞片时手要稳,并且手要有一个向下伸展的动作。控片,然后将切片放入展片器进行展片,注意使切片无气泡,贴平粘紧,及时烤片。
[0274]
2.8h&e染色
[0275]
1)将组织切片常规脱蜡至水;
[0276]
2)稍水洗1-2min;
[0277]
3)苏木精液染色3-6min;
[0278]
4)流水洗去苏木精液1-2min;
[0279]
5)1%盐酸酒精1-3sec;
[0280]
6)稍水洗1-2sec;
[0281]
7)促蓝液返蓝5-10sec;
[0282]
8)流水冲洗15-30sec;
[0283]
9)0.5%伊红液染色2-3min;
[0284]
10)蒸馏水稍洗1-2sec;
[0285]
11)80%乙醇15-30sec;
[0286]
12)95%乙醇15-30sec;
[0287]
13)无水乙醇1-2sec;
[0288]
14)二甲苯ⅰ2-3sec;
[0289]
15)二甲苯ⅱ2-3sec;
[0290]
16)中性树胶封固。
[0291]
2.9ihc染色
[0292]
1)脱蜡至水
[0293]
2)二甲苯ⅰ20min;
[0294]
3)二甲苯ⅱ20min;
[0295]
4)3号-8号酒精各2min;
[0296]
5)流水冲洗5min(水流要小);
[0297]
6)3%双氧水修复内源性过氧化氢酶(30%双氧水1:9配制,配100ml),室温30min,注意避光;
[0298]
7)单蒸水洗5min
×
2次,最后一次用pbs洗5min;
[0299]
8)热修复抗原,将切片浸入0.01m枸橼酸盐缓冲液中,微波炉处理高火5min,低火20min,完后自然冷却;
[0300]
9)取出切片放入染色缸中pbs洗5min
×
3次;
[0301]
10)取出切片放入湿盒中,滴加5%bsa封闭液(覆盖满组织为宜),室温30min;
[0302]
11)甩去封闭液滴加一抗(覆盖满组织为宜)注:阴性对照滴加pbs,放入4℃冰箱过夜;
[0303]
12)放入染色缸中pbs洗5min
×
3次;
[0304]
13)取出放入湿盒中滴加二抗(覆盖满组织为宜),室温30min;
[0305]
14)取出置染色缸中pbs洗5min
×
3次,放回湿盒中加sabc(覆盖满组织为宜),室温30min;
[0306]
15)取出置染色缸中pbs洗5min
×
3次;
[0307]
16)dab显色,粉状dab,称量0.02克dab充分溶于100ml单蒸水中后再加100微升双氧水,混匀倒入染色缸中显色,镜下观察是否终止显色;
[0308]
17)终止显色用单蒸水洗5min
×
2次;
[0309]
18)苏木素衬染2min,流水冲洗2min,盐酸分化数秒,流水冲洗10min;
[0310]
19)脱水透明9号-16号酒精各2min,17号二甲苯8min,18号二甲苯5min;
[0311]
20)中性树胶封片,放入37℃烘箱。
[0312]
2.10免疫荧光
[0313]
1)预处理:采样时将新鲜的组织样本包埋于oct中,液氮速冻后转至-80℃保存。实验前预约冰冻切片机,根据组织样本不同设定切片厚度,一般为5-10um。冰冻切片粘附性较好,用普通的载玻片即可。短暂保存放置-20℃,长期保存转至-80℃。
[0314]
2)记录切片编号和预设使用抗体及浓度。确保切片标记清晰,保持组织块面始终朝向自己,以免破坏组织块。将-20℃保存的切片放置室温干燥(时间视水分多少而定),组织块不能干,切片周围干燥即可,冰甲醇固定20min(放置-20℃冰箱)。
[0315]
3)用镊子从甲醇中拿出切片,倾斜在滤纸上尽可能的吸净甲醇,pbs洗涤3次,5min/次。注意每次都把切片盒清洗干净,双蒸水润洗后加入已灭菌过滤的pbs,不用加太满以免溢出。
[0316]
4)将片子用纸擦干,并注意不要干片,不能触碰到组织,使用免疫组化笔(用完放回盒子,以免丢失)在组织周围划圈,尽可能小。10%山羊血清(使用已灭菌过滤的pbs稀释)封闭,不要有气泡。(根据圈的大小确定用量,完全覆盖住组织块即可,不要贪多,以免流出圈外,目前回肠组织25ul足够),室温/37℃90min,封闭液中添加1%triton-x100。triton较粘稠,缓慢吸取,减少壁上残留。
[0317]
5)一抗4℃孵育16h(若信号弱可延长)或者37℃1h。根据样品数配制抗体,一抗用10%山羊血清稀释。滴加时不能产生气泡。
[0318]
6)pbst洗涤3次,每次5min。
[0319]
7)从这一步骤开始后面步骤全程避光。二抗,常温1h,尽量避光;二抗用已灭菌过滤的pbs稀释。滴加时不能产生气泡。
[0320]
8)pbst洗涤3次,pbs洗1次,5min/次;操作同2。
[0321]
9)dapi染核,5min。
[0322]
10)pbs洗涤3次,5min/次。
[0323]
11)抗荧光淬灭剂封片。滴加15ul即可,不能有气泡,倾斜盖玻片缓慢接触液面,与载玻片保持平行放置。不要用手接触组织块上方盖玻片,以免有污渍。
[0324]
12)显微镜下拍摄。
[0325]
2.11血清丙氨酸转氨酶(alt/gpt)
[0326]
试剂的组成与配制:
[0327]
试剂一:谷丙转氨酶基质液,5ml
×
1瓶,4℃冰箱保存6个月;
[0328]
试剂二:2,4-二硝基苯肼,5ml
×
1瓶,4℃冰箱保存6个月;
[0329]
试剂三:4mol/l氢氧化钠液,5ml
×
1瓶,室温密封保存6个月;
[0330]
注:0.4mol/l氢氧化钠液的配制:按照4mol/l氢氧化钠液:双蒸水=1:9的比例稀释,现配现用,室温封闭保存。
[0331]
试剂四:2μmol/ml丙酮酸钠标准液
×
1支,4℃冰箱保存6个月;
[0332]
试剂五:0.1mol/l磷酸盐缓冲液
×
1支,4℃冰箱保存6个月。
[0333]
操作过程:
[0334]
样本前处理:血清(浆)及其他液体样本测定,取样直接检测;
[0335]
操作流程如下:
[0336][0337]
水平摇动96孔板混匀,室温放置15min,波长510nm,酶标仪测定各孔od值,以(绝对od=测定孔od值-对照孔od值),查标准曲线,求得相应的alt/gpt活力单位。
[0338]
2.12血清天冬氨酸转氨酶(ast/got)
[0339]
试剂的组成与配制(96t):
[0340]
试剂一:谷草转氨酶基质液,5ml
×
1瓶,4℃冰箱保存6个月;
[0341]
试剂二:2,4-二硝基苯肼,5ml
×
1瓶,4℃冰箱保存6个月;
[0342]
试剂三:4mol/l氢氧化钠液,5ml
×
1瓶,室温密封保存6个月;
[0343]
注:0.4mol/l氢氧化钠液的配制:按照4mol/l氢氧化钠液:双蒸水=1:9的比例稀释,现配现用,室温封闭保存。
[0344]
试剂四:2μmol/ml丙酮酸钠标准液
×
1支,4℃冰箱保存6个月;
[0345]
试剂五:0.1mol/l磷酸盐缓冲液
×
1支,4℃冰箱保存6个月。
[0346]
操作过程:
[0347]
样本前处理:血清(浆)及其他液体样本测定,取样直接检测;
[0348]
操作流程如下:
[0349][0350]
轻轻水平摇动96孔板混匀,室温放置15min,波长510nm,酶标仪测定各孔od值,以(绝对od=测定孔od值-对照孔od值),查标准曲线,求得相应的ast/got活力单
[0351]
2.13数据分析
[0352]
数值以均值
±
标准差的方式表示。采用prisem作图软件进行统计性分析,组间差异显着性采用student's t-test进行统计显著性分析,*p<0.05的统计值被认为是差异显着,**p<0.01的统计值被认为是差异极显着。
[0353]
3.实验结果与分析
[0354]
3.1 ddc损伤模型下,tet2-/-小鼠肝脏损伤更严重
[0355]
为检测tet2基因对于肝脏损伤的影响,用含0.1%的3,5-二乙氧基羰基1,4-二氢2,4,6-三甲基吡啶(3,5-diethoxycarbonyl1,4-dihydrocollidine,ddc)饲料饲喂同为8周龄的野生型(wt)小鼠以及tet2基因敲除(tet2-/-)小鼠,构造ddc-14d小鼠肝损伤模型。
[0356]
将wt小鼠(c57bl/6j)和tet2基因敲除小鼠随机分为正常组和模型组,用含ddc饲料喂食2周,2周后处死小鼠。称取体重和肝重,计算肝重/体重比(lw/bw ratio)。分析后发现经过ddc模型处理后,相比与wt小鼠,tet2基因敲除小鼠肝重/体重比显著升高。
[0357]
为了检测tet2基因对于肝脏损伤的影响,分别对正常组和ddc模型组的wt小鼠和
tet2基因敲除小鼠肝组织固定、切片、进行he染色以分析病理变化,同时检测血浆丙氨酸转氨酶(alt)和天冬氨酸转氨酶(ast)水平。结果显示相对于wt小鼠,tet2基因敲除小鼠的肝脏存在更严重的肝细胞坏死和胆汁淤积现象(图1)。检测血浆丙氨酸转氨酶(alt)和天冬氨酸转氨酶(ast)结果显示,在经过ddc模型损伤后,wt小鼠与tet2基因敲除小鼠相较未经ddc损伤前均显著升高;并且ddc模型损伤后,wt小鼠,tet2基因敲除小鼠ast水平升高,alt水平显著升高。alt、ast结果也验证了tet2基因敲除小鼠的损伤更为严重(图2)。
[0358]
3.2ddc损伤后tet2-/-小鼠肝脏视黄酸代谢相关基因表达发生变化
[0359]
接下来,我们提取ddc模型组wt小鼠和tet2基因敲除小鼠肝脏的总rna,并通过转录组rna-seq分析差异表达基因的变化以及涉及的信号通路的变化。我们通过kegg富集分析筛选可能与tet2基因敲除小鼠损伤更严重有关的信号通路,分析结果显示视黄醇代谢信号通路显著变化,其中p450家族的视黄酸代谢酶cyp26a1以及cyp26c1的mrna表达量显著下降。随后,我们通过qrt-pcr在mrna水平上进行验证,结果显示,相比与野生型小鼠,tet2基因敲除小鼠无论是未损伤还是ddc损伤后,cyp26a1、cyp26c1基因表达均下降(图3)。该结果与转录组测序结果一致。
[0360]
紧接着我们又通过免疫组化实验验证cyp26a1蛋白的表达情况,得到与转录组测序、q-pcr一致的结果(图4)。
[0361]
然后我们通过高效液相色谱(hplc)检测小鼠血浆以及组织匀浆中视黄酸的含量。结果显示,相比与野生型小鼠,tet2基因敲除小鼠无论是未损伤还是ddc损伤后,肝脏组织匀浆中视黄酸的含量均显著下降(图5)。
[0362]
3.3ddc损伤后tet2-/-小鼠肝脏纤维化程度加重
[0363]
目前关于视黄酸的研究结果表明,视黄醇代谢通路与肝脏纤维化存在一定的联系。由于ddc模型组的tet2基因敲除小鼠肝脏组织匀浆中视黄酸含量降低,因此我们推测是否tet2基因与纤维化存在一定关联。首先我们通过免疫荧光双标染色α-sma和desmin,通过这两个纤维化指标来说明ddc模型组wt小鼠和tet2基因敲除小鼠的纤维化情况,发现无论是α-sma还是desmin,均在tet2基因敲除小鼠组信号更强(图6)。为了进一步说明这个问题。我们进行了masson染色,其结果也与免疫荧光结果一致(图7)。
[0364]
附:缩略词表
[0365]
[0366]
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。