顺式作用调节元件
1.相关申请的交叉引用
2.本技术要求2020年3月4日提交的美国临时专利申请序列号62/984831的提交目的权益,该临时专利申请的公开内容通过引用以其全文特此并入。
3.序列表的引用
4.序列表的正式副本作为ascii格式的序列表(其文件名称是“cis acting regulatory elements_seq_st25”,创建于2020年2月9日)经由efs-web以电子方式提交,并且与说明书同时提交。包含在该ascii格式的文件中的序列表是本说明书的一部分并且通过引用以其全文并入本文。
技术领域
5.本公开一般涉及分子生物学领域,并且在特定的实施例中,本公开涉及在编码序列附近的可以被工程化从而增加编码序列的表达的顺式作用调节元件。在某些方面,顺式作用元件与调节元件(例如启动子、5’utr、内含子或3’utr)一起被工程化,以产生新颖的嵌合序列。在其他方面,由顺式作用调节元件和另一调节元件(例如启动子、5’utr、内含子或3’utr)组成的新颖的嵌合序列驱动编码序列的更高水平的表达。在进一步的方面,顺式作用调节元件在活细胞(例如植物细胞)内起作用,以驱动编码序列的更高水平的表达。因此,本公开提供了用于鉴定、检测和利用这样的顺式作用调节元件的组合物和方法。
背景技术:
6.许多植物物种能够被转基因转化以引入农学上希望的性状或特征。开发和/或修饰所得植物物种使其具有特定的希望的性状。通常,希望的性状包括例如改善营养价值品质、增加产量、赋予有害生物抗性或疾病抗性、增加干旱和胁迫耐受性、改善园艺品质(例如,色素沉着和生长)、赋予除草剂耐受性、使得能够由植物产生工业上有用的化合物和/或材料和/或使得能够产生药物。
7.经由植物转化技术产生包含堆叠在单个基因组基因座的多个转基因的转基因植物物种。植物转化技术导致将转基因引入植物细胞,回收在植物基因组中包含稳定地整合的转基因拷贝的可育转基因植物,并随后经由转录和翻译的转基因表达得到具有希望的性状和表型的转基因植物。然而,希望允许产生转基因植物物种以高表达多个工程化为性状堆叠的转基因的新颖基因调节元件。
8.同样,希望允许转基因在植物的特定组织或器官内表达的新颖基因调节元件。例如,植物对土壤传播的病原体感染的抵抗力的增加可以通过用病原体抗性基因转化植物基因组,从而使病原体抗性蛋白在植物根部内稳健表达来实现。可替代地,可能希望在处于特定生长或发育阶段(诸如像细胞分裂或伸长)的植物组织中表达转基因。此外,可能希望在植物的叶和茎组织中表达转基因以提供对除草剂的耐受性或对地面上昆虫和有害生物的抗性。
9.因此,需要能够驱动特定植物组织中希望水平的转基因表达的新基因调节元件。
no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的核酸序列。在其他方面,转基因植物细胞是单子叶植物。在进一步的方面,转基因植物细胞是双子叶植物。在另外的方面,基因表达盒为组成型表达。在其他方面,转基因植物用基因表达盒进行稳定转化。在一些方面,转基因植物用基因表达盒进行瞬时转化。在另外的方面,从转基因植物开发包含基因表达盒的种子。在进一步的方面,从转基因植物开发包含基因表达盒的后代植物。
13.本文公开了一种用于抑制在除草剂耐受性转基因作物植物的田间的杂草生长的方法,该方法包括种植用包含嵌合调节分子的基因表达盒转化的转基因植物,其中该分子包含如下核酸序列,该核酸序列与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的同一性,在植物细胞中具有活性并且可操作地连接到编码除草剂耐受性基因的多核苷酸分子;以及以抑制杂草生长的施用量将除草剂施用到田间,其中转基因作物植物的生长和产量基本上不受除草剂施用的影响。本文公开了一种用于在转基因作物植物的田间提供有害生物控制的方法,该方法包括以下步骤:种植用包含嵌合调节分子的基因表达盒转化的转基因植物,其中该分子包含以下核酸序列,该核酸序列与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的同一性,在植物细胞中具有活性并且可操作地连接到赋予有害生物抗性的基因。本文公开了一种用于在转基因作物植物的田间提供疾病控制的方法,该方法包括以下步骤:种植用以下基因表达盒转化的转基因植物,该基因表达盒与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的同一性,在植物细胞中具有活性并且可操作地连接到赋予疾病抗性的基因。本文公开了一种在转基因作物植物的田间为植物提供胁迫耐受性的方法,该方法包括以下步骤:种植用基因表达盒转化的转基因植物,该基因表达盒与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的同一性,在植物细胞中具有活性并且可操作地连接到赋予胁迫耐受性的基因。本文公开了一种在转基因作物植物的田间为植物提供增产的方法,该方法包括以下步骤:种植用基因表达盒转化的转基因植物,该基因表达盒与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的同一性,在植物细胞中具有活性并且可操作地连接到赋予增产的基因。
14.本文公开了一种转化宿主细胞的方法,该方法包括以下步骤:提供包含以下多核苷酸的嵌合调节分子,该多核苷酸与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的同一性,其中所述分子可操作地连接到异源编码序列;并用该核酸分子转化所述细胞。在一些方面,该植物转化方法选自以下:农杆菌属(agrobacterium)介导的转化方法、基因枪转化方法、碳化硅转化方法、原生质体转化方法、和脂质体转化方法。本文公开了一种用于产生转基因植物细胞的方法,该方法包括以下步骤:用包含嵌合调节分子的基因表达盒转化植物细胞,该嵌合调节分子包含以下多核苷酸,该多核苷酸与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的同一性、可操作地连接到至少一个目的多核苷酸序列;分离包含该基因表达盒的经
转化的植物细胞;以及,产生包含该嵌合调节分子的转基因植物细胞,该嵌合调节分子包含以下多核苷酸,该多核苷酸与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的同一性、可操作地连接到至少一个目的多核苷酸序列。在一些方面,用植物转化方法转化植物细胞。在其他方面,植物转化方法选自以下:农杆菌属介导的转化方法、基因枪转化方法、碳化硅转化方法、原生质体转化方法、和脂质体转化方法。在进一步的方面,目的多核苷酸序列在植物细胞中表达。在另外的方面,目的多核苷酸序列被稳定整合到转基因植物细胞的基因组中。在其他方面,目的多核苷酸序列被瞬时整合到转基因植物细胞的基因组中。在进一步的方面,该方法进一步包括以下步骤:将转基因植物细胞再生为转基因植物;以及获得该转基因植物,其中该转基因植物包含基因表达盒,该基因表达盒包含可操作地连接到至少一个目的多核苷酸序列的如权利要求1所述的嵌合调节分子。在另外的方面,转基因植物细胞是单子叶转基因植物细胞或双子叶转基因植物细胞。在仍其他方面,目的多核苷酸序列是选自由以下组成的组的性状:杀昆虫抗性性状、除草剂耐受性性状、氮利用效率性状、水利用效率性状、营养品质性状、dna结合性状、选择性标记性状、小rna性状、或其任何组合。在其他方面,与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的多核苷酸以两个或更多个拷贝提供。
15.本文公开了用多核苷酸分子进行稳定转化的转基因植物,其包含:包含选自由以下组成的组的核酸序列的嵌合调节分子:与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性并且具有启动子活性的核酸序列;包含seq id no:9、seq id no:25-583、seq id no:585-2268的至少4-33个连续核苷酸并具有启动子活性的片段;以及seq id no:9、seq id no:25-583、seq id no:585-2268的核酸序列;异源编码序列,其中该嵌合调节分子可操作地连接到该异源编码序列。在一些方面,用植物转化方法转化转基因植物。示例性植物转化方法包括农杆菌属介导的转化方法、基因枪转化方法、碳化硅转化方法、原生质体转化方法、和脂质体转化方法。在其他方面,异源编码序列在转基因植物的细胞中表达。在进一步的方面,转基因植物是单子叶转基因植物细胞或双子叶转基因植物细胞。示例性异源编码序列包括选自由以下组成的组的性状:杀昆虫抗性性状、除草剂耐受性性状、氮利用效率性状、水利用效率性状、营养品质性状、dna结合性状、选择性标记性状、小rna性状、或其任何组合。在一些方面,转基因植物产生商品产品。示例性商品产品包括蛋白质浓缩物、蛋白质分离物、谷物、粗粉、面粉、油或纤维。
16.本文公开了用于增强调节分子表达的方法,该方法包括以下步骤:获得与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的顺式作用调节元件;将调节分子内与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的顺式作用调节元件进行工程化,以产生嵌合调节分子;将嵌合调节分子与异源编码序列进行连接以产生基因表达盒;在植物内转化该基因表达盒以获得转基因植物,其中该转基因植物表达异源编码序列。在一些方面,调节分子是启动子。在其他方面,该植物转化方法选自下组,该组由以下组成:农杆菌属(agrobacterium)介导的转化方法、基因枪转化方法、碳化硅转化方法、原生质体转化方法、
和脂质体转化方法。在进一步的方面,调节分子在转基因植物的细胞内表达。在另外的方面,目的多核苷酸序列被稳定地整合到植物细胞的基因组中。在随后的方面,转基因植物是单子叶植物或双子叶植物。在另外的方面,顺式作用调节元件被提供为两个或更多个拷贝。
17.本文公开了能够调节目的基因转录的分离的核酸分子,其中该核酸分子与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的同一性。在一些方面,调节导致目的基因的表达增强。在其他方面,调节导致目的基因的表达降低。在另外的方面,在真核细胞中发生目的基因表达的调节。在其他方面,目的基因是选自由以下组成的组的性状:杀昆虫抗性性状、除草剂耐受性性状、氮利用效率性状、水利用效率性状、营养品质性状、dna结合性状、选择性标记性状、小rna性状、或其任何组合。本文公开了通过包括以下步骤的方法产生的嵌合调节分子:在启动子调节元件内引入至少一种顺式作用调节元件以产生嵌合调节分子,其中该顺式作用调节元件包含与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的核酸分子。在其他方面,嵌合调节分子包含可操作地连接到嵌合调节分子的异源编码序列。在其他方面,异源编码序列是选自由以下组成的组的性状:杀昆虫抗性性状、除草剂耐受性性状、氮利用效率性状、水利用效率性状、营养品质性状、dna结合性状、选择性标记性状、小rna性状、或其任何组合。在进一步的方面,将嵌合调节分子引入基因表达盒。在其他方面,该方法包括以下步骤:用基因表达盒转化植物细胞;以及,获得包含该基因表达盒的转基因植物细胞。在另外的方面,该方法包括以下步骤:筛选转基因植物用于表达异源编码序列;检测所表达的异源编码序列的水平以确定嵌合调节分子的表达谱;以及,将该嵌合调节分子的表达谱与表达由启动子调节元件驱动的异源编码序列的转基因植物的表达谱进行比较,其中该启动调节元件不包含顺式作用元件。在另外的方面,与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的核酸序列作为两个或更多个拷贝提供。
18.根据权利要求书和具体实施方式中提供的以下实施例,前述和其他特征将变得更加显而易见,其参考附图和序列表进行。
附图说明
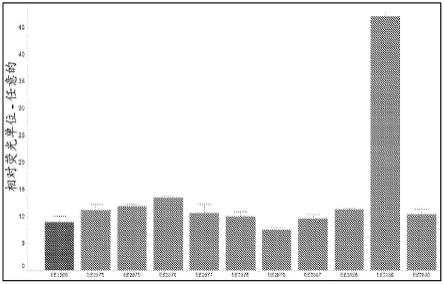
19.图1提供了图,该图显示了从数据库中获得的一组选定的顺式作用调节元件对由camv 35s最小启动子驱动的表达水平的影响。ee1906是背景对照,即没有测试盒的camv 35s最小启动子。误差条如sem所示。
20.图2提供了表,该表显示了seq id no:23-32的序列比对。
21.图3提供了表,该表显示了seq id no:33-36的序列比对。
22.图4提供了图,该图显示了mmv as-1元件对由camv 35s最小启动子驱动的表达水平的影响。ee1906是背景对照,即没有测试盒的camv 35s最小启动子。误差条如sem所示。
23.图5提供了图,该图显示了fmv as-1元件对由camv 35s最小启动子驱动的表达水平的影响。ee1906是背景对照,即没有测试盒的camv 35s最小启动子。误差条如sem所示。
24.图6提供了图,该图显示了mmv as-1元件对由camv 35s最小启动子驱动的表达水平的影响。ee1906是背景对照,即没有测试盒的camv 35s最小启动子。误差条如sem所示。
25.图7和图7.1提供了表,该表显示了seq id no:508-515的序列比对。
26.图8提供了图,该图显示了mmv as-1元件对由camv 35s最小启动子驱动的表达水平的影响。ee1906是背景对照,即没有测试盒的camv 35s最小启动子。误差条如sem所示。
27.图9提供了表,该表显示了seq id no:516-525的序列比对。
28.图10提供了图,该图显示了psgs3af1元件对由camv 35s最小启动子驱动的表达水平的影响。ee1906是背景对照,即没有测试盒的camv 35s最小启动子。误差条如sem所示。
29.图11提供了表,该表显示了seq id no:526-531的序列比对。
30.图12提供了图,该图显示了顺式基因调节元件对由玉蜀黍(zea mays)gos2最小启动子驱动的表达水平的影响。ee1619是背景对照,即没有测试盒的camv 35s最小启动子。误差条如sem所示。
31.图13和图13.1提供了表,该表显示了seq id no:532-550和seq id no:25、28、30和33的序列比对。
32.图14提供了图,该图显示了顺式基因调节元件对由玉蜀黍gos2启动子驱动的表达水平的影响。ee1619是背景对照,即没有测试盒的camv 35s最小启动子。误差条如sem所示。
33.图15、图15.1和图15.2提供了表,该表显示了seq id no:551-583和seq id no:2、3、4、5、8、9的序列比对。
34.图16提供了图,该图显示了mmv-eme 1和gm-psgs3af1-v3元件对由玉蜀黍gos2启动子驱动的表达水平的影响。ee1619是背景对照,即没有测试盒的camv 35s最小启动子。误差条如sem所示。
35.图17提供了表,该表显示了seq id no:516和529的序列比对。
36.图18提供了图,该图显示了从玉蜀黍获得的mmv as-1元件对由camv 35s最小启动子驱动的表达水平的影响。ee3549是背景对照,即没有测试盒的camv 35s最小启动子(yang等人,2000)。误差条如sem所示。
37.图19和图19.1提供了表,该表显示了seq id no:585-601的序列比对。
38.图20提供了图,该图显示了gvbavas1元件对由camv 35s最小启动子驱动的表达水平的影响。ee3549是背景对照,即没有测试盒的camv 35s最小启动子(yang等人,2000)。误差条如sem所示。
39.图21、图21.1和图21.2提供了表,该表显示了seq id no:602-634的序列比对。
40.序列表
41.如37 c.f.r.
§
1.822中定义,使用核苷酸碱基的标准字母缩写,显示了在所附序列表中列出的核酸序列。仅示出每个核酸序列的一个链,但是通过对展示的链的任何提及,应理解为包括互补链和反向互补链。因为一级核酸序列的互补序列和反向互补序列必然由一级序列公开,所以通过对核酸序列的任何提及,都包括互补序列和反向互补序列,除非另外明确指明(或从其中出现序列的上下文中另外清楚可见)。
具体实施方式
42.转基因植物产品的开发变得越来越复杂。商业上可行的转基因植物现在需要将多个转基因堆叠到单个基因座中。获得最佳水平的转基因/异源编码序列表达对于产生单个多基因性状是必要的。不幸的是,由弱表达启动子驱动的基因表达构建体可能无法以最佳
表达水平表达,从而导致田间的有效转基因产品较少。因此,仍然需要增加植物内转基因/异源编码序列的表达以开发稳健表达转基因/异源编码序列的转基因作物。
43.提供了通过使用顺式作用调节元件植物原位表达转基因来克服此类问题的方法和组合物。
44.定义
45.除非另外定义,本文所用的全部技术术语和科学术语具有与本公开所涉及领域的普通技术人员通常所理解的相同意义。在冲突的情况下,则以包括定义的本技术为准。除非上下文另外要求,否则单数术语应包括复数,并且复数术语应包括单数。本文提及的所有出版物、专利和其他参考文献出于所有目的通过引用以其整体并入,就好像每个单独的出版物或专利申请被具体地和单独地指示以引用方式并入,除非仅指示专利或专利出版物的特定部分通过引用并入。
46.为了进一步阐明本公开,提供以下术语、缩写和定义。
47.如本文所使用的,术语“包含(comprises)”、“包含(comprising)”、“包括(includes)”、“包括(including)”、“具有(has)”、“具有(having)”、“含有(contain)”或“含有(containing)”或其任何其他变型均旨在非排他性或开放性。例如,包含一系列要素的组合物、混合物、工艺、方法、制品、或设备不必仅限于这些要素,而是可以包括其他未明确列出的要素或此类组合物、混合物、工艺、方法、制品或设备固有的要素。此外,除非有相反的明确说明,否则“或”是指包含性的“或”,而不是指排他性的“或”。例如,条件a或b通过以下中的任一项满足:a为真(或存在)且b为假(或不存在),a为假(或不存在)且b为真(或存在),以及a和b两者都为真(或存在)。
48.同样,在本公开的实施例的元素或组分前的不定冠词“一个/一种(a/an)”关于例子(即元素或组分的出现)的数量旨在是非限制性的。因此,“一个”或“一种”应理解为包括一个/一种或至少一个/至少一种,并且元素或组分的单数词语形式还包括复数,除非该数字明显意指单数。
49.如在本文中使用的,术语“发明”或“本发明”是非限制性的术语,并且并不旨在是指特定发明的任何单一实施例,而是涵盖如公开于本技术中的所有可能的实施例。
50.如本文所用的术语“分离的”意指已经从其自然环境中除去,或从首次形成化合物时存在的其他化合物中除去。术语“分离的”涵盖从自然来源分离的材料以及通过在宿主细胞中重组表达制备后回收的材料(例如,核酸和蛋白质),或化学合成的化合物,如核酸分子、蛋白质和肽。
51.如本文所使用的,术语“纯化的”涉及分子或化合物的分离,该分子或化合物以基本上不含通常与在天然或自然环境中的分子或化合物关联的污染物,或者基本上在相对于化合物初次形成时存在的其他化合物的浓缩中富集的,并且意指由于与原始组合物中的其他组分分离而纯度提高了。本文使用的术语“纯化的核酸”来描述已经与其他生物化合物分离的、分离产生的或纯化的核酸序列,这些其他生物化合物包括但不限于多肽、脂质和碳水化合物,同时影响组分中的化学或功能改变组分(例如,可以通过去除蛋白质污染物并断开将核酸与染色体中其余dna连接的化学键,从染色体上纯化核酸)。
52.如本文所使用的,术语“合成的”是指经由化学合成作为体外过程产生的多核苷酸(即,dna或rna)分子。例如,可以在eppendorf
tm
管内的反应期间产生合成dna,使得合成dna
由dna或rna的天然链酶促产生。可以利用其他实验室方法来合成多核苷酸序列。寡核苷酸可以在寡核苷酸合成仪上经由使用亚磷酰胺的固相合成化学合成。合成的寡核苷酸可以作为复合物彼此退火,从而产生“合成的”多核苷酸。化学合成多核苷酸的其他方法是本领域已知的,并且可以容易地实现以用于本公开。
53.如本文所使用的术语“约”意指大于或小于所陈述的值或值的范围的百分之十,但并非旨在仅针对此更宽泛的定义来指定任何值或值的范围。术语“约”之后的每个值或值的范围也旨在涵盖所陈述绝对值或值的范围的实施例。
54.出于本公开的目的,“基因”包括编码基因产物的dna区域(参见下文),以及调节该基因产物的产生的所有dna区域,无论此类调控序列是否与编码和/或转录序列相邻。因此,基因包括但不限于启动子序列、终止子、翻译调节序列(如核糖体结合位点和内部核糖体进入位点)、增强子、沉默子、绝缘子、边界元件、复制起点、基质附着位点、内含子和基因座控制区。
55.如本文所使用的,术语“天然的”或“自然的”定义了天然存在的状况。“天然dna序列”是自然界中存在的dna序列,其是通过自然手段或传统育种技术产生的,但不是通过基因工程(例如,使用分子生物学/转化技术)产生的。
56.如本文所使用的,“转基因”被定义为编码基因产物的核酸序列,该基因产物包括例如但不限于mrna。在一个实施例中,转基因/异源编码序列是外源核酸,其中通过基因工程将转基因/异源编码序列引入宿主细胞(或其后代),其中通常找不到转基因/异源编码序列。在一个实例中,转基因/异源编码序列编码工业上或药学上有用的化合物,或编码希望的农艺性状的基因(例如,除草剂抗性基因)。在又另一个实例中,转基因/异源编码序列是反义核酸序列,其中反义核酸序列的表达抑制靶核酸序列的表达。在一个实施例中,转基因/异源编码序列是内源核酸,其中希望的是内源核酸的另外的基因组拷贝,或相对于宿主生物体中靶核酸的序列处于反义方向的核酸。
57.如本文所使用的,术语“非gmpsid2转基因”或“非gmpsid2基因”是与gmpsid2基因编码序列具有小于80%序列同一性的任何转基因/异源编码序列。
58.如本文所使用的,“异源dna编码序列”是指除自然编码gmpsid2基因的编码序列以外的任何编码序列,或表达的gmpsid2蛋白的任何同源物。在本发明的上下文中,术语“异源的”用于通常发现在自然界中没有紧密联系的核酸序列的任何组合。
59.如本文所定义的“基因产物”是由基因产生的任何产物。例如,基因产物可以是基因(例如,mrna、trna、rrna、反义rna、干扰rna、核酶、结构rna或任何其他类型的rna)或通过mrna翻译产生的蛋白质的直接转录产物。基因产物还包括通过如加帽、聚腺苷酸化、甲基化和编辑等方法修饰的rna,以及通过例如甲基化、乙酰化、磷酸化、泛素化、adp-核糖基化、肉豆蔻化和糖基化修饰的蛋白质。基因表达可能受到外部信号(例如,将细胞、组织或生物体暴露于增加或减少基因表达的药剂)的影响。基因的表达也可以在从dna到rna到蛋白质的途径中的任何地方受到调控。基因表达的调控例如通过对转录、翻译、rna转运和加工,中间分子(如mrna)的降解起作用的控制或通过在制造特定蛋白质分子后将其活化、失活、区室化或降解或者通过其组合而发生。可以通过本领域已知的任何方法(包括但不限于northern印迹、rt-pcr、western印迹或一种或多种体外、原位或体内蛋白活性测定)在rna水平或蛋白质水平上测量基因表达。
60.如本文所使用的,术语“基因表达”涉及这样的过程,通过该过程,核酸转录单位(包括例如基因组dna)的编码信息通常被转化为细胞的可操作的、不可操作的、或结构的部分,经常包括蛋白质的合成。基因表达会受到外部信号的影响;例如,将细胞、组织或生物体暴露于增加或减少基因表达的药剂。基因的表达也可以在从dna到rna到蛋白质的途径中的任何地方受到调控。基因表达的调控例如通过对转录、翻译、rna转运和加工,中间分子(如mrna)的降解起作用的控制或通过在制造特定蛋白质分子后将其活化、失活、区室化或降解或者通过其组合而发生。可以通过本领域已知的任何方法(包括但不限于northern印迹、rt-pcr、western印迹或一种或多种体外、原位或体内蛋白活性测定)在rna水平或蛋白质水平上测量基因表达。
61.如本文所使用的,“基于同源性的基因沉默”(hbgs)是通用术语,其包括转录基因沉默和转录后基因沉默。由于与启动子或转录序列相对应的双链rna(dsrna)的产生,转录抑制(转录基因沉默;tgs)或mrna降解(转录后基因沉默;ptgs)可导致未连锁沉默基因座对靶基因座的沉默。每个方法中不同细胞成分的参与表明,dsrna诱导的tgs和ptgs可能是由古老的共同机制的多样化导致的。但是,很难对tgs和ptgs进行严格比较,因为它通常依赖于不同沉默基因座的分析。在一些情况下,由于产生对应于不同靶基因的启动子和转录序列的dsrna,单个转基因基因座可触发tgs和ptgs。mourrain等人(2007)planta[植物学]225:365-79。sirna可能是在同源序列上触发tgs和ptgs的实际分子:这些sirna在此模型中会通过将转基因序列的甲基化扩散到内源性启动子中引发顺式和反式同源序列的沉默和甲基化。
[0062]
如本文所使用的,术语“核酸分子”(或“核酸”或“多核苷酸”)可以指核苷酸的聚合形式,其可以包括rna、cdna、基因组dna以及上述的合成形式和混合聚合物的有义链和反义链二者。核苷酸可以指核糖核苷酸、脱氧核糖核苷酸或任何一种核苷酸的修饰形式。如本文所使用的,“核酸分子”与“核酸”和“多核苷酸”同义。除非另有说明,否则核酸分子的长度通常至少为10个碱基。术语可以指长度不确定的rna或dna分子。术语包括dna的单链和双链形式。核酸分子可以包括通过自然存在的和/或非自然存在的核苷酸键连接在一起的自然存在的核苷酸和修饰的核苷酸之一或二者。
[0063]
如本领域技术人员将容易理解的,核酸分子可以被化学或生物化学修饰,或可以含有非自然或衍生的核苷酸碱基。此类修饰包括例如标记、甲基化、用类似物取代一个或多个自然存在的核苷酸、核苷酸间修饰(例如,不带电荷的键:例如,甲基膦酸酯、磷酸三酯、亚磷酰胺、氨基甲酸酯等;带电荷的键:例如,硫代磷酸酯,二硫代磷酸酯等;下垂部分:例如,肽;嵌入剂:例如吖啶、补骨脂素等;螯合剂;烷基化剂;和修饰的键:例如,α异头核酸等)。术语“核酸分子”还包括任何拓扑构象,包括单链、双链、部分双链体、三链体、发夹、环状和挂锁构型。
[0064]
转录沿着dna链以5
′
到3
′
的方式进行。这意味着rna是通过在生长链的3
′
末端顺序添加核糖核苷酸5
′‑
三磷酸(必需消除焦磷酸盐)而制备的。在线性或环状核酸分子中,如果离散元件(例如特定的核苷酸序列)在另一元件的5
′
方向上结合同一核酸或将结合同一核酸,则这些离散元件可以称为位于相对于那个元件的“上游”或“5
′”
端。类似地,如果离散元件是或将要从另一个元件在3
′
方向上与相同的核酸结合,则离散元件相对于那个元件可以是“下游”或“3
′”
。
[0065]
如本文所使用的,碱基“位置”是指给定碱基或核苷酸残基在指定核酸内的位置。可以通过与参考核酸比对(参见下文)来定义指定的核酸。
[0066]
杂交涉及经由氢键结合两个多核苷酸链。寡核苷酸及其类似物通过互补碱基之间的氢键合杂交,包括沃森-克里克(watson-crick)、胡斯坦(hoogsteen)或反向胡斯坦(hoogsteen)氢键合。通常,核酸分子由含氮碱基组成,这些含氮碱基是嘧啶(胞嘧啶(c)、尿嘧啶(u)和胸腺嘧啶(t))或嘌呤(腺嘌呤(a)和鸟嘌呤(g))。这些含氮碱基在嘧啶和嘌呤之间形成氢键,并且嘧啶与嘌呤的键合称为“碱基配对”。更具体地说,a将氢键合至t或u,而g将键合至c。“互补的”是指发生在两个不同的核酸序列或同一核酸序列的两个不同的区域之间的碱基配对。
[0067]“特异性可杂交”和“特异性互补”是表示足够程度的互补性的术语,使得在寡核苷酸与dna或rna靶标之间发生稳定且特异性的结合。寡核苷酸不必与其靶序列100%互补即可特异性杂交。当寡核苷酸与靶dna或rna分子的结合干扰靶dna或rna的正常功能时,寡核苷酸可特异性杂交,并且具有足够程度的互补性以避免寡核苷酸与非靶序列在希望特异性结合的条件下(例如在体内测定或系统的情况中在生理条件下)的非特异性结合。这种结合称为特异性杂交。
[0068]
导致特定严格性程度的杂交条件将根据所选择的杂交方法的性质以及杂交核酸序列的组成和长度而变化。通常,杂交温度和杂交缓冲液的离子强度(尤其是na 和/或mg2 浓度)将有助于杂交的严格性,尽管洗涤时间也会影响严格性。sambrook等人(编)molecular cloning:a laboratory manual[分子克隆:实验室手册]第二版第1-3卷,冷泉港实验室出版社,纽约冷泉港,1989年,chs.9和11中讨论了有关获得特定严格性程度所需的杂交条件的计算。
[0069]
如本文所使用的,“严格条件”涵盖仅在杂交分子与dna靶之间的错配小于50%时才发生杂交的条件。“严格条件”包括进一步的严格的特定水平。因此,如本文所使用的,“中严格性”条件是指在这些条件下序列错配率超过50%的分子不会杂交;“高严格性”条件是指在这些条件下错配率超过20%的序列不会杂交;以及“非常高严格性”条件是指在这些条件下错配率超过10%的序列不会杂交。
[0070]
在特定的实施例中,严格条件可以包括在65℃下杂交,然后在65℃下用0.1x ssc/0.1%sds洗涤40分钟。
[0071]
以下是代表性的非限制性的杂交条件:
[0072]
非常高严格性:在65℃下,在5x ssc缓冲液中杂交16小时;在室温下在2x ssc缓冲液中洗涤两次,每次持续15分钟;并且在65℃下在0.5x ssc缓冲液中洗涤两次,每次持续20分钟。
[0073]
高严格性:在65℃-70℃下,在5x-6x ssc缓冲液中杂交16-20小时;在室温下,在2x ssc缓冲液中洗涤两次,每次5-20分钟;并且在55℃-70℃下在1x ssc缓冲液中洗涤两次,每次持续30分钟。
[0074]
中严格性:在室温至55℃下在6x ssc缓冲液中杂交16-20小时;在室温至55℃下在2x-3x ssc缓冲液中洗涤至少两次,每次持续20-30分钟。
[0075]
在特定的实施例中,特异性杂交的核酸分子可以在非常高严格性杂交条件下保持结合。在这些和进一步的实施例中,特异性杂交的核酸分子可以在高严格性杂交条件下保
持结合。在这些和进一步的实施例中,特异性杂交的核酸分子可以在中严格性杂交条件下保持结合。
[0076]
如本文所使用的,术语“寡核苷酸”是指短核酸聚合物。寡核苷酸可以通过切割更长的核酸区段或通过聚合单个核苷酸前体来形成。自动化合成仪允许合成长度长达数百个碱基对的寡核苷酸。因为寡核苷酸可以与互补的核苷酸序列结合,所以它们可以用作检测dna或rna的探针。由dna组成的寡核苷酸(寡脱氧核糖核苷酸)可以用于pcr(扩增小dna序列的技术)。在pcr中,寡核苷酸典型地称为“引物”,它允许dna聚合酶延伸寡核苷酸并复制互补链。
[0077]
术语“百分比序列同一性”或“百分比同一性”或“同一性”可互换使用,是指基于在两个或更多个氨基酸或核苷酸序列之间进行比较的序列中相应同一位置之间的同一匹配的序列比较。百分比同一性是指两个最佳比对的多核苷酸或肽序列在组分例如核苷酸或氨基酸的比对窗口中不变的程度。本领域已知的杂交实验和数学算法可用于确定百分比同一性。存在许多数学算法作为本领域已知的计算序列百分比同一性的序列比对计算机程序。这些程序可以分为全局序列比对程序或局部序列比对程序。
[0078]
全局序列比对程序通过端对端比较比对以找到精确匹配,将精确匹配的数目除以较短序列的长度,然后乘以100,计算出两个序列的百分比同一性。基本上,当两个序列最佳比对(具有适当的核苷酸插入、缺失或缺口)时,参考(“查询”)多核苷酸分子的线性多核苷酸序列与测试(“受试者”)多核苷酸分子相比,同一的核苷酸的百分比。
[0079]
本地序列比对程序在计算上相似,但是仅比较序列的比对片段,而不是利用端到端分析。如blast的局部序列比对程序可用于比较两个序列的特定区域。两个序列的blast比较会产生e值或期望值,该值表示具有得分等于或优于原始比对得分(s)的不同比对的数量,可能偶然在数据库搜索中发生。e值越低,匹配越显著。因为数据库大小是e值计算中的一个元素,所以对于任何给定的查询/条目匹配,通过blasting针对公共数据库(如genbank)获得的e值通常随着时间的推移而增加。在设定多肽功能预测的置信度标准时,“高”blast匹配在本文中被认为具有对于最高blast命中而言的e值小于1e-30;中等的blastx e值为1e-30至1e-8;并且低的blastx e值大于1e-8。使用e值、百分比同一性、查询覆盖率和命中覆盖率的组合来确定本发明中的蛋白质功能分配。查询覆盖率是指查询序列以blast比对表示的百分比。命中覆盖率是指以blast比对方式表示的数据库条目的百分比。在本发明的一个实施例中,从蛋白同源物的功能推断查询多肽的功能,其中(1)hit[命中]_p<1e-30或%identity[同一性]>35%且query[查询]_coverage[覆盖率]>50%且hit[命中]_coverage[覆盖率]>50%,或(2)hit[命中]_p<1e-8且query[查询]_coverage[覆盖率]>70%且hit[命中]_coverage[覆盖率]>70%。在序列的blast分析过程中会产生以下缩写。
[0080]
[0081]
[0082][0083]
用于比较的序列的比对方法是本领域熟知的。描述了各种程序和比对算法。在实施例中,本公开涉及使用vector nti套件的alignx比对程序(英杰公司(invitrogen),卡尔斯巴德,加利福尼亚州)计算两个多核苷酸或氨基酸序列之间的百分比同一性。alignx比对程序是针对多核苷酸或蛋白质的全局序列比对程序。在实施例中,本公开涉及使用lasergene生物信息学计算套件的megalign程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(megalign
tm
dnastar.麦迪逊,威斯康星州)。megalign程序是针对多核苷酸或蛋白质的全局序列比对程序。在实施例中,本公开涉及使用比对程序的clustal套件计算两个多核苷酸或氨基酸序列之间的百分比同一性,该套件包括但不限于clustalw和clustalv(higgins和sharp(1988)gene[基因].12月15日;73(1):237-44;higgins和sharp(1989)cabios[计算机应用生物科学]5:151-3;higgins等人(1992)comput.appl.biosci.[计算机应用生物科学]8∶189-91)。在实施例中,本公开涉及使用程序的gcg套件(威斯康星软件包版本9.0,遗传学计算机组(genetics computer group(gcg)),麦迪逊,威斯康星州)计算两个多核苷酸或氨基酸序列之间的百分比同一性。在实
施例中,本公开涉及使用比对程序(例如包括但不限于blastp、blastn、blastx等)的blast套件计算两个多核苷酸或氨基酸序列之间的百分比同一性(altschul等人(1990)j.mol.biol.[分子生物学杂志]215:403-10)。在实施例中,本公开涉及使用比对程序(包括但不限于fasta、tfastx、tfasty、ssearch、lalign等)的fasta套件计算两个多核苷酸或氨基酸序列之间的百分比同一性(pearson(1994)comput.methods genome res.[基因组研究中的计算方法][proc.int.symp.],会议日期1992(suhai和sandor编辑),普莱南出版公司(plenum):纽约市,纽约州,第111-20页)。在实施例中,本公开涉及使用t-coffee比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(notredame等人(2000)j.mol.biol.[分子生物学杂志]302,205-17)。在实施例中,本公开涉及使用比对程序(包括但不限于dialign、chaos、dialign-tx、dialign-t等)的dialign套件计算两个多核苷酸或氨基酸序列之间的百分比同一性(al ait等人(2013)dialign at gobics[在gobics的dialign]nuc.acids research[核酸研究]41,w3-w7)。在实施例中,本公开涉及使用比对程序的muscle套件计算两个多核苷酸或氨基酸序列之间的百分比同一性(edgar(2004)nucleic acids res.[核酸研究]32(5):1792-1797)。在实施例中,本公开涉及使用mafft比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(katoh等人(2002)nucleic acids research[核酸研究]30(14):3059-3066)。在实施例中,本公开涉及使用genoogle程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(albrecht,felipe.arxiv130702987v1[cs.dc]2015年7月10日)。在实施例中,本公开涉及使用程序的hmmer套件计算两个多核苷酸或氨基酸序列之间的百分比同一性(eddy.(1998)bioinformatics[生物信息学],14:755-63)。在实施例中,本公开涉及使用比对程序(包括但不限于tplastn、plastp、klast、和plastx)的plast套件计算两个多核苷酸或氨基酸序列之间的百分比同一性(nguyen与lavenier.(2009)bmc bioinformatics[bmc生物信息学],10:329)。在实施例中,本公开涉及使用usearch比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(edgar(2010)bioinformatics[生物信息学]26(19),2460-61)。在实施例中,本公开涉及使用比对程序的sam套件计算两个多核苷酸或氨基酸序列之间的百分比同一性(hughey与krogh(1995年1月)technicalreportucsc0crl-95-7[技术报告ucsc0crl-95-7],university of califomia[加利福尼亚大学],圣克鲁兹)。在实施例中,本公开涉及使用idf检索器计算两个多核苷酸或氨基酸序列之间的百分比同一性(o
′
kane,k.c.,the effect of inverse document frequency weights on indexedsequence retrieval[反向文档频率权重对索引序列检索的影响],onlinejournal of bioinformatics[生物信息学在线杂志],第6卷(2)162-173,2005)。在实施例中,本公开涉及使用parasail比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(daily,jeff.parasail:simd c library for global,semi-global,and local pairwise sequence alignments[用于全局、半全局和局部成对序列比对的simd c库].bmc bioinformatics[bmc生物信息学].17:18.2月10日,2016)。在实施例中,本公开涉及使用scalablast比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(oehmen c,nieplocha j.
″
scalablast:a scalable implementation of blast for high-performance data-intensive bioinformatics analysis[scalablast:用于高性能数据密集型生物信息学分析的blast的可扩展实现].
″ꢀ
ieee transactions on parallel&distributed systems[ieee并行与分布式系统汇刊]17(8):740-7492006年8
月)。在实施例中,本公开涉及使用swipe比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(rognes,t.faster smilth-waterman database searches with inter-sequence simd parallelization[通过序列间simd并行化,可以更快地进行smilth-waterman数据库搜索].bmc bioiinformatics[bmc生物信息学].12,221(2011))。在实施例中,本公开涉及使用acana比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(weichun huang,david m.umbach和leping li,accurate anchoring alignment of divergent sequences[不同序列的精确锚定比对].bioinformatics[生物信息学]22:29-34,2006年1月1日)。在实施例中,本公开涉及使用dotlet比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(junier,t.和pagni,m.dotlet:diagonal plots in a web browser[dotlet:网络浏览器中的对角线图].bioinformatics[生物信息学]16(2):178-92000年2月)。在实施例中,本公开涉及使用g-pas比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(frohmberg,w.等人g-pas 2.0-an improved version of protein alignment tool with an efficient backtracking routine on multiple gpus[g-pas 2.0-在多个gpu上具有高效的回溯程序的蛋白质比对工具的改进版本].bulletin of the polish academy of sciences technical sciences[波兰科学院技术科学通报],第60卷,4912012年11月)。在实施例中,本公开涉及使用gapmis比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(flouri,t.等人,gap mis:a tool for pairwise sequence alignment with a single gap[gap-mis:单空位成对序列比对工具].recent pat dna gene seq[dna和基因序列最新专利].7(2):84-952013年8月)。在实施例中,本公开涉及使用比对程序的emboss套件计算两个多核苷酸或氨基酸序列之间的百分比同一性,该套件包括但不限于:matcher、needle、streter、water、wordmatch等(rice,p.,longden,i.和bleasby,a.emboss:the european molecular biology open software suite[emboss:欧洲分子生物学开放软件套件].trends in genetics[遗传学趋势]16(6)276-77(2000))。在实施例中,本公开涉及使用ngila比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(cartwright,r.ngila:global pairwise alignments with logarithmic and affine gap costs[ngila:对数和仿射空位成本的全局成对比对].bioinformatics[生物信息学].23(11):1427-28.2007年6月1日)。在实施例中,本公开涉及使用proba(也称为propa)比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(m
ü
ckstein,u.,hofacker,il和stadler,pf.stochastic pairwise alignments[随机成对比对].bioinformatics[生物信息学]18增刊2:s153-60.2002)。在实施例中,本公开涉及使用比对程序的seqaln套件计算两个多核苷酸或氨基酸序列之间的百分比同一性(hardy,p.和waterman,m.the sequence alignment software library at usc[usc的序列比对软件库].1997)。在实施例中,本公开涉及使用比对程序(包括但不限于gap、nap、lap等)的sim套件计算两个多核苷酸或氨基酸序列之间的百分比同一性(huang,x和miller,w.a time-efficient,linear-space local similarity algorithm[省时的线性空间局部相似性算法].advances in applied mathematics[应用数学进展],第12卷(1991)337-57)。在实施例中,本公开涉及使用ugene比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(okonechnikov,k.,golosova,o.和fursov,m.unipro ugene:a unified bioinformatics toolkit[unipro ugene:统一的生物信息学工具包].bioinformatics[生
物信息学].201228:1166-67)。在实施例中,本公开涉及使用bali-phy比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(suchard,ma和redelings,bd.bali-phy:simultaneous bayesian inference of alignment and phylogeny[bali-phy:比对和系统发育的贝叶斯同时推断].bioinformatics[生物信息学].22:2047-48.2006)。在实施例中,本公开涉及使用base-by-base比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(brodie,r.等人,base-by-base:single nucleotide-level analysis of whole viral genome alignments[base-by-base:全病毒基因组比对的单核苷酸水平分析],bmc bioinformatics[bmc生物信息学],5,96,2004)。在实施例中,本公开涉及使用decipher比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(es wright(2015)
″
decipher:harnessing local sequence context to improve protein multiple sequencealignment[decipher:利用局部序列上下文改进蛋白质多序列比对].
″
bmc bioinformatics[bmc生物信息学],doi:10.1186/s12859-015-0749-z.)。在实施例中,本公开涉及使用fsa比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(bradley,rk等人(2009)fast statistical alignment[快速统计比对].plos computationalbiology[plos计算生物学].5:e1000392)。在实施例中,本公开涉及使用geneious比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(kearse,m.等人(2012).geneious basic:an integrated and extendable desktop software platform for the organization and analysis of sequence data[geneious basic:集成和可扩展的用于序列数据组织和分析的桌面软件平台].bioinformatics[生物信息学],28(12),1647-49)。在实施例中,本公开涉及使用kalign比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(lassmann,t.和sonnhammer,e.kalign-an accurate and fast multiple sequence alignment algorithm[kalign-准确和快速的多序列比对算法].bmc bioinformatics[bmc生物信息学]2005 6:298)。在实施例中,本公开涉及使用mavid比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(bray,n.和pachter,l.mavid:constrained ancestral alignment of multiple sequences[mavid:多个序列的约束祖先比对].genome res.[基因组研究]2004年4月;14(4):693-99)。在实施例中,本公开涉及使用msa比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(lipman,dj等人,a tool for multiple sequence alignment[多序列比对工具].proc.nat’l acad.sci.usa.[美国科学院院报]1989;86:4412-15)。在实施例中,本公开涉及使用multalin比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(corpet,f.,multiple sequence alignment with hierarchial clustering[具有系统聚类法的多序列比对].nucl.acids res.[核酸研究],1988,16(22),10881-90)。在实施例中,本公开涉及使用lagan或mlagan比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(brudno等人,lagan and multi-lagan:efficient tools for large-scale multiple alignment of genomic dna[lagan和multi-lagan:基因组dna大规模多重比对的有效工具].genome research[基因组研究]2003年4月;13(4):721-31)。在实施例中,本公开涉及使用opal比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(wheeler,t.j.和kececiouglu,j.d.multiple alignment by aligning alignments[通过对齐比对进行多重比对].proceedings of the 15
th iscb conference on intelligent systems for molecular biology[第15届iscb分
子生物学智能系统会议纪要].bioinformatics[生物信息学].23,i559-68,2007)。在实施例中,本公开涉及使用程序的picxaa套件(包括但不限于picxaa、picxaa-r、picxaa-web等)计算两个多核苷酸或氨基酸序列之间的百分比同一性(mohammad,s.,sahraeian,e.和yoon,b.picxaa:greedy probabilistic construction of maximum expected accuracy alignment of multiple sequences[多个序列的最大预期准确性比对的贪婪概率构建].nucleic acids research[核酸研究].38(15):4917-28.2010)。在实施例中,本公开涉及使用psalign比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(sze,s.-h.,lu,y.与yang,q.(2006)a polynomial time solvable formulation of multiple sequence alignment[多序列比对的多项式时间可解公式]journal of computational biology[计算生物学杂志],13,309-19)。在实施例中,本公开涉及使用statalign比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(nov
á
k,等人(2008)statalign:an extendable software package for joint bayesian estimation of alignments and evolutionary trees[statalign:可扩展的联合贝叶斯比对和进化树估计软件包].bioinformatics[生物信息学],24(20):2403-04)。在实施例中,本公开涉及使用needleman和wunsch的gap比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(needleman和wunsch,journal of molecular biology[分子生物学杂志]48:443-453,1970)。在实施例中,本公开涉及使用smith和waterman的bestfit比对程序计算两个多核苷酸或氨基酸序列之间的百分比同一性(smith和waterman,advances in applied mathematics,[应用数学进展]2:482-489,1981,smith等人,nucleic acids research[核酸研究]11:2205-2220,1983)。这些程序产生发散序列的生物学上有意义的多重序列比对。将针对所选序列计算的最佳匹配比对排列起来,以便可以看到同一性、相似性和差异。
[0084]
术语“相似性”是指氨基酸序列之间的比较,并且不仅考虑对应位置的同一的氨基酸,而且考虑对应位置的功能上相似的氨基酸。因此,除了序列相似性之外,多肽序列之间的相似性还指示功能相似性。
[0085]
术语“同源性”有时用于指两个或更多个核酸或氨基酸序列之间的相似性水平,以位置同一性(即序列相似性或同一性)的百分比表示。同源性也指进化相关性的概念,通常通过共享相似序列的不同核酸或蛋白质之间的相似功能特性来证明。
[0086]
如本文所使用的,术语“变体”是指基本相似的序列。就核苷酸序列来说,可使用本领域熟知的分子生物学技术来鉴定自然存在的变体,诸如像用本文概述的聚合酶链式反应(pcr)和杂交技术来鉴定。
[0087]
对于核苷酸序列,变体包含在天然多核苷酸中的一个或多个内部位点处的一个或多个核苷酸的缺失和/或添加,和/或在天然多核苷酸中的一个或多个位点处的一个或多个核苷酸的取代。如本文所使用的,“天然”核苷酸序列包括自然存在的核苷酸序列。就核苷酸序列来说,可使用本领域熟知的分子生物学技术来鉴定自然存在的变体,例如,用以下概述的聚合酶链式反应(pcr)和杂交技术来鉴定。核苷酸序列变体还包括合成来源的核苷酸序列,如那些采用定点诱变技术产生的核苷酸序列。通常,如通过本文别处所描述的序列比对程序和参数确定的,本发明的特定的核苷酸序列的变体将与该特定的核苷酸序列具有至少约40%、45%、50%>、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更高的序列同一性。本发明的核苷酸序列的生物活性
变体与该序列不同的核酸残基数可能只有1至15个,只有1至10个,如6至10个,只有5个,只有4、3、2个,或甚至只有1个。
[0088]
如本文所使用的,术语“可操作地连接”涉及当第一核酸序列与第二核酸序列具有功能关系时,第一核酸序列与第二核酸序列可操作地连接。例如,当启动子影响编码序列的转录或表达时,该启动子可操作地连接到编码序列。当重组产生时,可操作地连接的核酸序列通常是连续的,并且在需要连接两个蛋白质编码区的情况下,它们在同一阅读框中。然而,元件不必连续地可操作地连接。
[0089]
如本文所使用的,术语“启动子”是指通常位于基因上游(朝向基因的5
′
区域)的dna区域,并且是启动和驱动基因转录所需要的。启动子可以允许其控制的基因的适当激活或抑制。启动子可以含有被转录因子识别的特定序列。这些因子可能与启动子dna序列结合,导致募集rna聚合酶,rna聚合酶是一种从基因编码区合成rna的酶。启动子通常是指位于基因上游的所有基因调节元件,包括上游启动子、5
′
utr、内含子和前导序列。
[0090]
如本文所使用的,术语“上游启动子”是指足以指导转录起始的连续多核苷酸序列。如本文所使用的,上游启动子涵盖具有几个序列基序的转录起始位点,这些序列基序包括tata box、启动子序列、tfiib识别元件和其他启动子基序(jennifer,e.f.等人,(2002)genes&dev.[基因与发育],16:2583-2592)。上游启动子为rna聚合酶ii提供了作用位点,rna聚合酶ii是具有基础或一般的转录因子(例如tfiia、b、d、e、f和h)的多亚基酶。这些因子组装成转录前起始复合物,该转录前起始复合物催化从dna模板合成rna。
[0091]
上游启动子激活是通过各种蛋白质结合并随后与转录起始复合物相互作用以激活基因表达的调控dna序列元件的附加序列完成的。这些基因调控元件序列与特定dna结合因子相互作用。这些序列基序有时可以称为顺式元件。此类顺式元件,结合至其组织特异性或发育特异性的转录因子,单独地或组合,可以在转录水平决定启动子的时空表达模式。这些顺式元件对可操作地连接的基因施加的控制类型差异很大。一些元件的作用是增加可操作连接的基因的转录响应环境响应(例如,温度、湿度和伤害)。其他顺式元件可能对发育线索(例如,发芽、种子成熟和开花)或空间信息(例如,组织特异性)有反应。参见,例如,langridge等人,(1989)proc.natl.acad.sci.usa[美国国家科学院院刊]86:3219-23。这些顺式元件位于距转录起点不同距离的位置,一些顺式元件(称为近端元件)与最小核心启动子区域相邻,而其他元件可位于启动子(增强子)上游或下游几千碱基处。
[0092]
如本文所使用的,术语“5
′
非翻译区”或“5
′
utr”定义为前mrna或成熟mrna的5
′
末端中的非翻译区段。例如,在成熟的mrna上,一个5
′
utr典型地在其5
′
末端带有一个7-甲基鸟苷帽,并参与许多过程,如剪接、聚腺苷酸化、mrna向细胞质的输出,通过翻译机制鉴定mrna的5
′
末端,以及保护mrna免受降解。
[0093]
如本文所使用的,术语“内含子”是指转录但未翻译的基因(或表达的目的多核苷酸序列)中包含的任何核酸序列。内含子包括在表达的dna序列内的未翻译的核酸序列,以及从其转录的rna分子中的相应序列。本文所述的构建体还可含有增强翻译和/或mrna稳定性的序列,如内含子。一个这样的内含子的实例是拟南芥(arabidopsis thaliana)组蛋白h3变体的基因ii的第一内含子或任何其他通常已知的内含子序列。内含子可以与启动子序列组合使用以增强翻译和/或mrna稳定性。
[0094]
如本文所使用的,术语“转录终止子”或“终止子”定义为前mrna或成熟mrna的3
′
末
端中的转录区段。例如,超过“聚腺苷酸化信号”位点的更长的一段dna被转录为前mrna。该dna序列通常含有转录终止信号,用于将前mrna正确加工成成熟的mrna。
[0095]
如本文所使用的,术语“3
′
非翻译区”或“3
′
utr”定义为前mrna或成熟mrna的3
′
末端中的非翻译区段。例如,在成熟的mrna上,该区域带有聚-(a)尾,并且已知在mrna稳定性、翻译起始和mrna输出中具有许多作用。另外,认为3
′
utr包括聚腺苷酸化信号和转录终止子。
[0096]
如本文所使用的,术语“聚腺苷酸化信号”表示存在于mrna转录物中的核酸序列,当存在聚-(a)聚合酶时,其允许转录物在聚腺苷酸化位点上被聚腺苷酸化,例如位于聚-(a)信号下游的10至30个碱基。许多聚腺苷酸化信号是本领域已知的,并且可用于本发明。示例性序列包括aauaaa及其变体,如loke j.等人,(2005)plant physiology[植物生理学]138(3);1457-1468中所述。
[0097]“dna结合转基因”是编码dna结合蛋白的多核苷酸编码序列。dna结合蛋白随后能够结合另一个分子。结合蛋白可以与例如dna分子(dna结合蛋白)、rna分子(rna结合蛋白)和/或蛋白质分子(蛋白质结合蛋白)结合。如果是蛋白质结合蛋白,它可以结合自身(以形成同型二聚体、同型三聚体等)和/或结合不同蛋白质的一个或多个分子。结合蛋白可以具有超过一种类型的结合活性。例如,锌指蛋白具有dna结合、rna结合和蛋白质结合活性。
[0098]
dna结合蛋白的实例包括;大范围核酸酶、锌指、crispr、和talen结合结构域可以被“工程化”以结合预定的核苷酸序列。典型地,工程化的dna结合蛋白(例如,锌指、crispr或talen)是非天然存在的蛋白质。设计和选择用于工程化dna结合蛋白的方法的非限制性实例。设计的dna结合蛋白是自然界中不存在的蛋白,其设计/组成主要来自合理的标准。设计的合理标准包括应用替换规则和计算机算法,以处理数据库中存储现有zfp、crispr和/或talen设计信息和结合数据的信息。参见,例如美国专利6,140,081;6,453,242;和6,534,261;还参见wo 98/53058;wo 98/53059;wo 98/53060;wo 02/016536和wo 03/016496和美国公开号20110301073、20110239315和20119145940。
[0099]“锌指dna结合蛋白”(或结合结构域)是一种蛋白质,或者是较大蛋白质中的一个结构域,其可以通过一个或多个锌指按序列特异性方式结合dna,该锌指是结合结构域内氨基酸序列的区域,其结构通过锌离子的配位而稳定。术语锌指dna结合蛋白通常缩写为锌指蛋白或zfp。锌指结合结构域可以被“工程化”以结合预定的核苷酸序列。设计和选择用于工程化锌指蛋白的方法的非限制性实例。设计的锌指蛋白是自然界中不存在的蛋白,其设计/组成主要来自合理的标准。设计的合理标准包括应用替换规则和计算机算法,以处理数据库中存储现有zfp设计信息和结合数据的信息。参见,例如美国专利号6,140,081;6,453,242;6,534,261和6,794,136;还参见wo 98/53058;wo 98/53059;wo 98/53060;wo 02/016536和wo 03/016496。
[0100]
在其他实例中,一种或多种核酸酶的dna结合结构域包含天然存在或工程化的(非天然存在的)tal效应子dna结合结构域。参见,例如美国专利公开号20110301073,其通过引用以其全文并入本文。已知黄单胞菌属(xanthomonas)的植物致病细菌在重要的作物植物中引起许多疾病。黄单胞菌属的致病性取决于保守的iii型分泌(t3s)系统,该系统向植物细胞中注入的蛋白质超过了不同的效应子蛋白。在这些注射的蛋白质中有转录激活子样(talen)效应子,它们模仿植物转录激活子并操纵植物转录组(参见kay等人,(2007)
science[科学]318:648-651)。这些蛋白质含有dna结合结构域和转录激活结构域。表征最充分的tal效应子之一是来自疱病野油菜黄单胞菌(xanthomonas campestgris pv.vesicatoria)的avrbs3(参见bonas等人,(1989)mol gen genet[分子和普通遗传学]218:127-136和wo 2010079430)。tal效应子含有串联重复的集中结构域,每个重复含有约34个氨基酸,这是这些蛋白质的dna结合特异性的关键。另外,它们含有核定位序列和酸性转录激活结构域(综述参见schornack s等人,(2006)j plant physiol[植物生理学]163(3):256-272)。另外,在植物病原细菌青枯雷尔氏菌(ralstonia solanacearum)两个基因,指定brg11和hpx17已经发现与青枯雷尔氏菌生物变体菌株gmi1000和生物变体4菌株rs1000的黄单胞菌属avrbs3家族同源(参见heuer等人,(2007)appl and enviro micro[应用与环境微生物学]73(13):4379-4384)。这些基因在核苷酸序列上彼此具有98.9%的同一性,但在hpx17的重复结构域中相差1,575bp的缺失。然而,两种基因产物与黄单胞菌属的avrbs3家族蛋白具有少于40%序列同一性。参见,例如美国专利公开号20110301073,其通过引用以其全文并入。
[0101]
这些tal效应子的特异性取决于在串联重复序列中发现的序列。重复序列包含约102bp,重复序列典型地与彼此为91%-100%同源(bonas等人,同上)。重复序列的多态性通常位于位置12和13,并且似乎有位置12和13上的高变二残基的身份与tal效应子的靶序列中连续核苷酸的身份之间的一种一一对应关系(参见moscou和bogdanove,(2009)science[科学]326:1501和boch等人(2009)science[科学]326:1509-1512)。实验上,这些tal效应子的dna识别的自然密码已经确定了使得位置12和13的hd序列导致与胞嘧啶(c)结合,ng与t结合,ni与a、c、g或t结合,nn与a或g结合,而ing与t结合。这些dna结合重复序列已被组装成具有新的重复序列的组合和数量的蛋白质,以制造能够与新序列相互作用并激活植物细胞中的非内源性报告基因表达的人工转录因子(boch等人,同上)。工程化的tal蛋白已与foki切割半结构域连接,以产生tal效应子结构域核酸酶融合蛋白(talen),其在酵母报告基因测定(基于质粒的靶标)中具有活性。
[0102]
crispr(成簇的规律间隔的短回文重复序列)/cas(crispr相关的)核酸酶系统是基于细菌系统的一种最近工程化的核酸酶系统,可用于基因组工程。它基于许多细菌和古细菌的适应性免疫反应的一部分。当病毒或质粒入侵细菌时,入侵者的dna区段会通过“免疫”反应转化为crispr rna(crrna)。然后,该crrna通过部分互补区域与另一种称为tracrrna的rna结合,以将cas9核酸酶引导至与靶dna中与crrna同源的区域,称为“前间区序列”。cas9切割dna以在双链断裂末端(dsb)在由crrna转录物中含有的20核苷酸指导序列指定的位点处产生平末端。cas9需要crrna和tracrrna才能进行位点特异性dna识别和切割。现在已经对该系统进行了工程化,以便可以将crrna和tracrrna组合至一个分子(“单个指导rna”),并且可以对单个指导rna的crrna等效部分进行工程化,以指导cas9核酸酶靶向任何所希望的序列(参见jinek等人,(2012)science[科学]337,第816-821页,jinek等人,(2013),elife 2:e00471和david segal,(2013)elife 2:e00563)。在其他实例中,crrna与tracrrna结合,以将cpf1核酸酶引导至与crrna同源的区域,以切割末端交错的dna(参见zetsche,bernd等人,cell[细胞]163.3(2015):759-771.)。因此,可以对crispr/cas系统工程化以在基因组中的希望的靶标上产生dsb,并且可以通过使用修复抑制剂来影响dsb的修复,从而导致易于出错的修复增加。
[0103]
在其他实例中,dna结合转基因/异源编码序列是位点特异性核酸酶,其包含工程化的(非天然存在的)大范围核酸酶(也称为归巢核酸内切酶)。归巢核酸内切酶或大范围核酸酶的识别序列,如i-scei、i-ceui、pi-pspi、pi-sce、i-sceiv、i-csmi、i-pani、i-sceii、i-ppoi、i-sceiii、i-crei、i-tevi、i-tevii和i-teviii是已知的。还参见美国专利号5,420,032;美国专利号6,833,252;belfort等人,(1997)nucleic acids res.[核酸研究]25:3379-30 3388;dujon等人,(1989)gene[基因]82:115-118;perler等人,(1994)nucleic acids res.[核酸研究]22,11127;jasin(1996)trends genet.[遗传学趋势]12:224-228;gimble等人,(1996)j.mol.biol.[分子生物学杂志]263:163-180;argast等人,(1998)j.mol.biol.[分子生物学杂志]280:345-353和新英格兰生物实验室目录。另外,归巢核酸内切酶和大范围核酸酶的dna结合特异性可以被工程化以结合非天然靶标位点。参见,例如,chevalier等人(2002)molec.cell[分子细胞]10:895-905;epinat等人,(2003)nucleic acids res.[核酸研究]5 31:2952-2962;ashworth等人,(2006)nature[自然]441:656-659;paques等人,(2007)current gene therapy[目前基因疗法]7:49-66;美国专利公开号20070117128。归巢核酸内切酶和大范围核酸酶的dna结合结构域可以在整个核酸酶的背景下改变(即,使得核酸酶包括同源切割结构域),或者可以与异源切割结构域融合。
[0104]
如本文所使用的,术语“转化”涵盖可以将核酸分子引入这种细胞的所有技术。实例包括但不限于:用病毒载体转染;用质粒载体转化;电穿孔;脂质转染;显微注射(mueller等人,(1978)cell[细胞]15:579-85);农杆菌属介导的转移;直接dna摄取;whiskers
tm
介导的转化;和微粒轰击。这些技术可用于植物细胞的稳定转化和瞬时转化。“稳定转化”是指将核酸片段引入宿主生物体的基因组中,导致遗传稳定的遗传。一旦经稳定转化,核酸片段稳定地整合入宿主生物体和任何后代的基因组中。含有经转化的核酸片段的宿主生物体被称为“转基因的”生物体。“瞬时转化”是指将核酸片段引入宿主生物体的细胞核或含dna的细胞器中,导致不具遗传稳定遗传的基因表达。
[0105]
外源核酸序列。在一个实例中,转基因/异源编码序列是基因序列(例如,除草剂抗性基因)、编码工业上或药学上有用的化合物的基因、或编码希望的农艺性状的基因。在又另一个实例中,转基因/异源编码序列是反义核酸序列,其中该反义核酸序列的表达抑制靶核酸序列的表达。转基因/异源编码序列可以含有可操作地连接到该转基因/异源编码序列(例如,启动子)的调节序列。在一些实施例中,目的多核苷酸序列是转基因。然而,在其他实施例中,目的多核苷酸序列是内源核酸序列,其中希望的是内源核酸序列的另外的基因组拷贝,或相对于宿主生物体中靶核酸分子的序列处于反义方向的核酸序列。
[0106]
如本文所使用的,通过以下来产生术语转基因“事件”:用异源dna转化植物细胞,该异源dna为包括目的转基因/异源编码序列的核酸构建体;再生由该转基因/异源编码序列插入该植物的基因组中所产生的植物群体;并且选择表征为插入特定基因组位置的特定植物。术语“事件”是指包括异源dna的原始转化体和该转化体的后代。术语“事件”还指由转化体与包括基因组/转基因dna的另一种变体之间的有性杂交产生的后代。即使在与轮回亲本反复回交后,插入的转基因/异源编码序列dna和来自经转化的亲本的侧翼基因组dna(基因组/转基因dna)也存在于杂交的后代中相同的染色体位置。术语“事件”也指来自原始转化体和其后代的dna,该后代包含插入的dna和与插入的dna直接相邻的侧翼基因组序列,插入的dna预期将被转移到接受包括目的转基因/异源编码序列的插入的dna的后代中,得到
包括插入的dna(例如,自交所产生的原始转化子和后代)的亲本系和不含有插入的dna的亲本系发生性杂交的结果。
[0107]
如本文所使用的,术语“聚合酶链式反应”或“pcr”定义了一种程序或技术,其中如1987年7月28日授权的美国专利号4,683,195中所述扩增了微量的核酸、rna和/或dna。通常,需要从目的区域的末端或以外的区域获得序列信息,使得可以设计寡核苷酸引物;这些引物在序列上与待扩增模板的相反链相同或相似。两个引物的5
′
末端核苷酸可以与扩增的材料的末端一致。pcr可用于从总的基因组dna扩增特定的rna序列、特定的dna序列,以及从总的细胞rna、噬菌体或质粒序列转录的cdna等。通常参见mullis等人,cold spring harbor symp.quant.biol.[冷泉港定量生物学研讨会],51:263(1987);erlich编辑,pcr technology[pcr技术],(stockton press[斯托克顿出版社],纽约州,1989)。
[0108]
如本文所使用的,术语“引物”是指当条件适合于引物延伸产物的合成时能够充当沿着互补链的合成起始点的寡核苷酸。合成条件包括存在四种不同的脱氧核糖核苷酸三磷酸和至少一种聚合诱导剂,如逆转录酶或dna聚合酶。它们存在于合适的缓冲液中,其可以包括作为辅因子的成分或在各种合适的温度下影响如ph等条件的成分。引物优选地是单链序列,使得扩增效率得到优化,但是可以利用双链序列。
[0109]
如本文所使用的,术语“探针”是指与靶序列杂交的寡核苷酸。在或样式的测定程序中,探针与位于两个引物的退火位点之间的靶标的一部分杂交。探针包括约八个核苷酸、约十个核苷酸、约十五个核苷酸、约二十个核苷酸、约三十个核苷酸、约四十个核苷酸或约五十个核苷酸。在一些实施例中,探针包括约八个核苷酸至约十五个核苷酸。探针还可以包括可检测标记,例如荧光团(异硫氰酸荧光素等)。可检测标记可以直接共价附接于探针寡核苷酸,例如位于探针的5
′
末端或探针的3
′
末端。包括荧光团的探针还可以进一步包括淬灭剂,例如black hole quencher
tm
、iowa black
tm
等。
[0110]
如本文所使用的,术语“限制性核酸内切酶”和“限制酶”是指细菌酶,每种这样的酶在特定核苷酸序列处或附近切割双链dna。2型限制酶在同一位点识别并切割dna,包括但不限于xbai、bamhi、hindiii、ecori、xhoi、sali、kpni、avai、psti和smai。
[0111]
如本文所使用的,术语“载体”与术语“构建体”、“克隆载体”和“表达载体”可互换使用,并且意指可将dna或rna序列(例如外源基因)引入宿主细胞,以转化宿主并促进所引入的序列的表达(例如转录和翻译)的载体。“非病毒载体”旨在意指不包含病毒或逆转录病毒的任何载体。在一些实施例中,“载体”是包含至少一个dna复制起点和至少一个选择性标记基因的dna序列。实例包括但不限于将外源dna带入细胞的质粒、粘粒、噬菌体、细菌人工染色体(bac)或病毒。载体还可以包括一个或多个基因、反义分子和/或选择性标记基因以及本领域已知的其他遗传元件。载体可以转导、转化或感染细胞,从而使细胞表达由载体编码的核酸分子和/或蛋白质。
[0112]
术语“质粒”定义了能够在原核或真核宿主细胞中常染色体复制的核酸的环状链。术语包括可以是dna或rna并且可以是单链或双链的核酸。定义的质粒还可以包括对应于细菌复制起点的序列。
[0113]
如本文所使用的,如本文所使用的术语“选择性标记基因”定义了编码蛋白质的基因或其他表达盒,该蛋白质有助于鉴定插入了选择性标记基因的细胞。例如,“选择性标记
基因”涵盖报告基因以及用于植物转化以例如保护植物细胞免于选择剂或对选择剂提供抗性/耐受性的基因。在一个实施例中,仅那些接受功能选择性标记的细胞或植物能够在具有选择剂的条件下分裂或生长。短语“标记阳性”是指已经被转化为包括选择性标记基因的植物。
[0114]
如本文所使用的,术语“可检测标记”是指能够检测的标记,诸如像放射性同位素、荧光化合物、生物发光化合物、化学发光化合物、金属螯合剂或酶。可检测标记的实例包括但不限于以下:荧光标记(例如,fitc、若丹明、镧系元素荧光粉)、酶标记(例如,辣根过氧化物酶、β-半乳糖苷酶、萤光素酶、碱性磷酸酶)、化学发光、生物素基、由二级报告分子识别的预定多肽表位(例如,亮氨酸拉链对序列、二级抗体的结合位点、金属结合结构域、表位标签)。在实施例中,可检测标记可以通过各种长度的间隔臂附接以减少潜在的空间位阻。
[0115]
如本文所使用的,术语“盒”、“表达盒”和“基因表达盒”是指可在特定限制位点处或通过同源重组插入核酸或多核苷酸中的dna区段。如本文所使用的,dna的区段包含编码目的多肽的多核苷酸,并且盒和限制位点被设计为确保将盒插入适当的阅读框中以进行转录和翻译。在实施例中,表达盒可以包括编码目的多肽的多核苷酸,并且除了促进特定宿主细胞转化的多核苷酸外还具有元件。在实施例中,基因表达盒还可以包括允许在宿主细胞中增强表达编码目的多肽的多核苷酸的元件。这些元件可以包括但不限于:启动子、最小启动子、增强子、响应元件、终止子序列、聚腺苷酸化序列等。
[0116]
如本文所使用的,“接头”或“间隔子”是将两个分开的实体彼此结合的键、分子或分子的组。接头和间隔子可以提供两个实体的最佳间隔,或者可以进一步提供允许两个实体彼此分离的不稳定的连接。不稳定的连接包括光可切割基团、酸不稳定部分、碱基不稳定部分和酶可切割基团。如本文所用的术语“多接头”或“多克隆位点”定义了位于核酸序列上彼此10个核苷酸内的三个或更多个2型限制酶位点的簇。在其他情况下,如本文所使用的术语“多接头”是指经由任何已知的无缝克隆方法(即gibsonnebuilderhifidnagolden gate assembly、assembly等)靶向连接两个序列的一段核苷酸。包含多接头的构建体用于核酸序列(如基因的编码区)的插入和/或切除。
[0117]
如本文所使用的,术语“对照”是指在分析程序中用于比较目的的样品。对照可以是“阳性”或“阴性”。例如,在分析程序的目的是检测细胞或组织中差异表达的转录物或多肽的情况下,通常优选包括阳性对照(如来自已知植物的表现出所希望表达的样品)和阴性对照(如来自已知植物的缺少所需表达的样品)。
[0118]
如本文所使用的,术语“植物”包括整株植物以及任何后代,细胞、组织或植物的一部分。可用于本发明的植物种类通常包括适合变异发生的高等及低等植物,包括被子植物(单子叶植物和双子叶植物)、裸子植物、蕨类植物和多细胞藻类。因此,“植物”包括双子叶植物和单子叶植物。术语“植物部位”包括植物的任何部位,包括例如且不限于:种子(包括成熟种子和未成熟种子);植物切段;植物细胞;植物细胞培养物;植物器官(例如花粉、胚、花朵、果实、枝、叶、根、茎和外植体)。植物组织或植物器官可以是种子、原生质体、愈伤组织或组织成结构或功能单元的任何其他组的植物细胞。植物细胞或组织培养物可能能够再生具有从其获得细胞或组织的植物的生理和形态特征的植物,并且能够再生具有与该植物基
02182-9);和meyers(编辑),molecular biology and biotechnology:a comprehensive desk reference,[分子生物学和生物技术:综合办公桌参考]vch publishers,inc.,[vch出版公司]1995(isbn 1-56081-569-8)中找到。
[0125]
实施例
[0126]
提供了使用嵌合调节分子内的顺式作用调节元件在植物内表达异源编码序列的方法和组合物。在实施例中,顺式作用调节元件可以包含seq id no:9、seq id no:25-583、seq id no:585-2268。在另一个实施例中,顺式作用调节元件与seq id no:9、seq id no:25-583、seq id no:585-2268具有80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%同一性。在其他实施例中,嵌合调节分子包含可操作地连接到顺式作用调节元件的启动子、5’utr或内含子。在一些实施例中,顺式作用调节元件作为嵌合调节分子内的多拷贝提供。在进一步的实施例中,嵌合调节分子可操作地连接到异源编码序列/转基因以产生基因表达盒。在另外的实施例中,顺式作用调节元件调节异源编码序列的表达以增强或降低异源编码序列/转基因的表达。
[0127]
在实施例中,提供了包含顺式作用调节元件的分离的多核苷酸,其中该顺式作用调节元件与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同。在实施例中,嵌合调节分子包含顺式作用调节元件,该顺式作用调节元件包含与多核苷酸seq id no:9、seq id no:25-583、seq id no:585-2268具有至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%同一性的多核苷酸。在实施例中,提供了分离的多核苷酸,其与seq id no:9、seq id no:25-583、seq id no:585-2268的多核苷酸具有至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%、或100%同一性。在实施例中,提供了包含顺式作用调节元件的核酸载体,该顺式作用调节元件与seq id no:9、seq id no:25-583、seq id no:585-2268具有至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%同一性。在实施例中,提供了包含顺式作用调节元件的嵌合调节分子,该顺式作用调节元件与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同,其中多核苷酸可操作地连接到嵌合调节分子。在实施例中,提供了包含顺式作用调节元件的嵌合调节分子,该顺式作用调节元件与可操作地连接到内含子的seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同。在实施例中,提供了包含可操作地连接到5’utr的顺式作用调节元件的嵌合调节分子,该顺式作用调节元件与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同。在实施例中,提供了包含可操作地连接到多接头的顺式作用调节元件的嵌合调节分子,该顺式作用调节元件与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同。在实施例中,提供了包含顺式作用调节元件的基因表达盒,该顺式作用调节元件与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、
99.8%或100%相同。在实施例中,提供了包含嵌合调节分子元件的基因表达盒,该嵌合调节分子元件与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同。在一些情况下,包含顺式作用调节元件的基因表达盒进一步包含启动子、5’utr、内含子、多接头、异源编码序列或转基因,该顺式作用调节元件与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同。在实施例中,提供了包含顺式作用调节元件的载体,该顺式作用调节元件与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同。在实施例中,提供了包含嵌合调节分子元件的载体,该嵌合调节分子元件与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同。在一些情况下,包含顺式作用调节元件的载体进一步包含启动子、5’utr、内含子、多接头、异源编码序列或转基因,该顺式作用调节元件与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同。
[0128]
在实施例中,嵌合调节分子包含至少一个拷贝的顺式作用调节元件,该顺式作用调节元件与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同。在其他实施例中,顺式作用调节元件在嵌合调节分子中提供为单拷贝、双拷贝、三拷贝、四拷贝。在另外的实施例中,顺式作用调节元件在嵌合调节分子中以多个拷贝提供;例如1-100个拷贝。在进一步的实施例中,顺式作用调节元件的多个拷贝可以彼此顺序连接。在其他实施例中,顺式作用调节元件的多个拷贝可以通过间插序列彼此分开。这样的间插序列可以是任何长度,例如长度为1bp至10,000bp。
[0129]
在实施例中,包含顺式作用调节元件的嵌合调节分子可操作地连接到转基因,该顺式作用调节元件与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同,其中转基因/异源编码序列可以是杀昆虫抗性转基因、除草剂耐受性转基因、氮利用效率转基因、水利用效率转基因、营养品质转基因、dna结合转基因、小rna转基因、选择性标记转基因或其组合。
[0130]
在实施例中,核酸载体包含如本文公开的基因表达盒。在实施例中,载体可以是用于直接转化或基因靶向如供体dna的质粒、粘粒、细菌人工染色体(bac)、噬菌体、病毒或切除的多核苷酸片段。
[0131]
本公开的另一方面包含功能变体,其一个或多个核苷酸与本文提供的包含调节元件的核苷酸序列的那些不同。由于包含本文所述的序列的核苷酸序列的一种或多种修饰(例如,缺失、重排、或插入)而产生这种变体。例如,seq id no:9、seq id no:25-583、seq id no:585-2268的片段和变体可以与调节元件融合以产生嵌合调节元件。该嵌合调节元件可以用于dna构建体或基因表达盒中,以驱动异源编码序列的表达。如本文所使用的,术语“片段”是指核酸序列的一部分。seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件的片段可以保留通过引发转录调节表达的生物活性,以驱动增强的表
达。seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件的核苷酸序列的片段可以在至少约4个核苷酸、5个核苷酸、6个核苷酸、7个核苷酸、8个核苷酸、9个核苷酸、10个核苷酸、11个核苷酸、12个核苷酸、13个核苷酸、14个核苷酸、15个核苷酸、16个核苷酸、17个核苷酸、18个核苷酸、19个核苷酸、20个核苷酸、21个核苷酸、22个核苷酸、23个核苷酸、24个核苷酸、25个核苷酸、26个核苷酸、27个核苷酸、28个核苷酸、29个核苷酸、30个核苷酸、31个核苷酸、32个核苷酸、33个核苷酸、或高至顺式作用调节元件的全长核苷酸序列的范围内。
[0132]
seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件的生物活性部分可以通过分离seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件的部分,并评估该顺式作用调节元件的部分调节异源编码序列或转基因转录的生物学活性来制备。作为seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件的片段的核酸分子包含至少约4个核苷酸、5个核苷酸、6个核苷酸、7个核苷酸、8个核苷酸、9个核苷酸、10个核苷酸、11个核苷酸、12个核苷酸、13个核苷酸、14个核苷酸、15个核苷酸、16个核苷酸、17个核苷酸、18个核苷酸、19个核苷酸、20个核苷酸、21个核苷酸、22个核苷酸、23个核苷酸、24个核苷酸、25个核苷酸、26个核苷酸、27个核苷酸、28个核苷酸、29个核苷酸、30个核苷酸、31个核苷酸、32个核苷酸、33个核苷酸、或高至如本文公开的seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件的全长核苷酸序列。
[0133]
seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件的生物活性部分可以通过分离seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件的部分,并评估该顺式作用调节元件的部分增强异源编码序列或转基因表达的生物学活性来制备。作为seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件的片段的核酸分子包含至少约4个核苷酸、5个核苷酸、6个核苷酸、7个核苷酸、8个核苷酸、9个核苷酸、10个核苷酸、11个核苷酸、12个核苷酸、13个核苷酸、14个核苷酸、15个核苷酸、16个核苷酸、17个核苷酸、18个核苷酸、19个核苷酸、20个核苷酸、21个核苷酸、22个核苷酸、23个核苷酸、24个核苷酸、25个核苷酸、26个核苷酸、27个核苷酸、28个核苷酸、29个核苷酸、30个核苷酸、31个核苷酸、32个核苷酸、33个核苷酸、或高至如本文公开的seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件的全长核苷酸序列。
[0134]
核苷酸序列变体还涵盖衍生自诱变和重组发生程序(如dna改组)的序列。通过这种程序,可以操纵seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件以创建新的顺式作用调节元件。以此方式,由相关序列多核苷酸的群体产生重组多核苷酸文库,这些相关序列多核苷酸包含具有基本序列同一性并且能够在体外或体内同源重组的序列区域。这种dna改组的策略在本领域中是已知的。参见,例如,stemmer(1994)proc.natl.acad.sci.usa[美国国家科学院院刊]i:10747-10751;stemmer(1994)nature[自然]570:389-391;crameri等人(1997)nature biotech.[自然生物技术]75:436-438;moore等人(1997)j.mol.biol.[分子生物学杂志]272:336-347;zhang等人(1997)proc.natl.acad.sci.usa[美国国家科学院院刊]£4:4504-4509;crameri等人(1998)nature[自然]527:288-291;以及美国专利号5,605,793和5,837,458。
[0135]
本公开的核苷酸序列可用于分离来自其他生物(特别是其他植物,更特别是其他单子叶植物)的相应序列。以这种方式,可以使用如pcr、杂交等方法来鉴定此类序列(基于
其与本文所示序列的序列同源性)。因此,本发明涵盖基于与本文所示的seq id no:9、seq id no:25-583、seq id no:585-2268的完整顺式作用调节元件或其片段的序列同一性而分离的序列。
[0136]
在pcr方法中,可以设计寡核苷酸引物用于pcr反应,以便从提取自任何目的植物的基因组dna中扩增出相应dna序列。用于设计pcr引物和pcr克隆的方法是本领域通常已知的,并公开于sambrook等人(1989)molecular cloning:a laboratory manual[分子克隆:实验室手册](第2版,cold spring harbor laboratory press[冷泉港实验室出版社],普莱恩维尤,纽约),以下简称为sambrook。还参见innis等人编辑,(1990)pcr protocols:a guide to methods and applications[pcr方案:方法与应用指南](学术出版社,纽约);innis和gelfand编辑,(1995)pcr strategies[pcr策略](学术出版社,纽约);以及innis和gelfand编辑,(1999)pcr methods manual[pcr方法手册](学术出版社,纽约)。已知的pcr方法包括但不限于:使用成对引物、巢式引物、单特异性引物、简并引物、基因特异性引物、载体特异性引物、部分错配引物等的方法。
[0137]
在杂交技术中,将已知核苷酸序列的全部或一部分用作探针,该探针与来自所选生物体的一组克隆的基因组dna片段中存在的其他相应核苷酸序列选择性杂交。杂交探针可以用可检测基团(如p
32
)或任何其他可检测标记进行标记。因此,例如,用于杂交的探针可以通过标记基于本公开的seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件的合成寡核苷酸来制备。制备用于杂交的探针和用于构建基因组文库的方法通常是本领域已知的,并且公开在sambrook中。例如,本文公开的seq id no:9、seq id no:25-583、seq id no:585-2268的整个顺式作用调节元件或其一个或多个部分可用作能够与相应的顺式作用调节元件特异性杂交的探针。要在多种条件下实现特异性杂交,此类探针包括在seq id no:9、seq id no:25-583、seq id no:585-2268的顺式作用调节元件中独特的序列,并且其长度为至少约10个核苷酸或者其长度为至少约20个核苷酸。可以使用此类探针通过pcr从选择的植物扩增相应的顺式作用调节元件。这项技术可以用于来从希望的生物体分离另外的编码序列,或者用作诊断测定以确定编码序列在生物体中的存在。杂交技术包括杂交筛选铺板的dna文库(噬菌斑或菌落;参见,例如sambrook)。
[0138]
根据一个实施例,核酸载体进一步包含编码选择性标记的序列。根据一个实施例,重组基因盒可操作地连接到农杆菌属t-dna边界。根据一个实施例,重组基因盒进一步包含第一和第二t-dna边界,其中第一t-dna边界可操作地连接到基因构建体的一端,而第二t-dna边界可操作地连接到基因构建体的另一端。第一和第二农杆菌属t-dna边界可独立地选自源自细菌菌株的t-dna边界序列,这些细菌菌株选自由以下组成的组:胭脂碱合成的农杆菌属t-dna边界、章鱼碱合成的农杆菌属t-dna边界、甘露碱合成的农杆菌属t-dna边界、农杆碱合成农杆菌属t-dna边界或其任何组合。在一个实施例中,提供了农杆菌属菌株,其选自由以下组成的组:胭脂碱合成菌株、甘露碱合成菌株、农杆碱合成菌株、或章鱼碱合成菌株,其中所述菌株包含质粒,其中该质粒包含可操作地连接到选自顺式作用调节元件的序列的转基因/异源编码序列,该顺式作用调节元件与seq idno:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同。在另一个实施例中,第一和第二农杆菌属t-dna边界可以独立地选自源自细菌菌株的t-dna边界序列,这些细菌菌株选自由以下组成的组:胭脂碱
合成的农杆菌属t-dna边界、章鱼碱合成的农杆菌属t-dna边界、甘露碱合成的农杆菌属t-dna边界、农杆碱合成农杆菌属t-dna边界或其任何组合。在实施例中,提供了农杆菌属菌株,其选自由以下组成的组:胭脂碱合成菌株、甘露碱合成菌株、农杆碱合成菌株、或章鱼碱合成菌株,其中所述菌株包含质粒,其中所述质粒包含可操作地连接到选自顺式作用调节元件或嵌合调节分子的序列的转基因/异源编码序列,该顺式作用调节元件或嵌合调节分子与seq id no:9、seq id no:25-583、seq id no:585-2268至少80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、99.5%、99.8%或100%相同。
[0139]
用于渗入的性状
[0140]
在一些实施例中,包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件可以用于驱动植物中异源编码序列(例如,目的转基因)的表达。
[0141]
目的转基因可以通过包含本公开的seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件表达。适用于本公开的构建体的目的示例性转基因包括但不限于赋予如下的编码序列:(1)有害生物抗性或疾病抗性、(2)对除草剂的耐受性、(3)添加农学性状的价值,如;产量提高、氮利用效率、水利用效率和营养品质,(4)蛋白质以位点特异性方式与dna结合,(5)表达小rna;以及(6)选择性标记。根据一个实施例,包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件用于驱动转基因/异源编码序列的表达,该转基因/异源编码序列编码选择性标记或赋予杀昆虫抗性、除草剂耐受性、小rna表达、氮利用效率、水利用效率、或营养品质的基因产物。
[0142]
1.昆虫抗性
[0143]
各种昆虫抗性基因可以通过包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件进行表达。包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件可以与至少一个含有昆虫抗性基因的其他基因表达盒可操作地连接。然后可将可操作地连接的序列掺入选择的载体中以允许鉴定和选择经转化的植物(“转化体”)。示例性昆虫抗性编码序列是本领域已知的。作为可与本公开的调节元件可操作地连接的昆虫抗性编码序列的实施例,提供以下性状。提供示例性鳞翅目昆虫抗性的编码序列包括:cry1a;cry1a.105;cry1ab;cry1ab(截短的);cry1ab-ac(融合蛋白);cry1ac(作为销售);cry1c;cry1f(作为销售);cry1fa2;cry2ab2;cry2ae;cry9c;mocry1f;pinii(蛋白酶抑制剂蛋白);vip3a(a);和vip3aa20。提供示例性鞘翅目抗昆虫性的编码序列包括:cry34ab1(作为销售);cry35ab1(作为销售);cry3a;cry3bb1;dvsnf7;和mcry3a。提供示例性多重抗昆虫性的编码序列包括ecry31.ab。以上昆虫抗性基因的列表并不意味着具有限制性。本公开涵盖任何昆虫抗性基因。
[0144]
2.除草剂耐受性
[0145]
各种除草剂耐受性基因可以通过包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件进行表达。包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件可以与至少一个含有除草剂耐受性基因的其他基因表达盒可操作地连接。然后可将可操作地连接的序列掺入选择的载体中以允许鉴定和选择经转化的植物(“转化体”)。示例性除草剂耐受性编码序列是本领域已知的。作为可与本公开的调节元
件可操作地连接的除草剂耐受性编码序列的实施例,提供以下性状。草甘膦除草剂通过抑制epsps酶(5-烯醇式丙酮酸莽草酸酯-3-磷酸合酶)而发挥作用。该酶参与植物的生长和发育必不可少的芳香氨基酸的生物合成。本领域已知的各种酶促机制可用于抑制该酶。可以将编码此类酶的基因可操作地连接到本公开的基因调节元件。在实施例中,选择性标记基因包括但不限于编码草甘膦抗性基因的基因,包括:突变的epsps基因,如2mepsps基因、cp4 epsps基因、mepsps基因、dgt-28基因;aroa基因;和草甘膦降解基因,如草甘膦乙酰基转移酶基因(gat)和草甘膦氧化酶基因(gox)。这些性状目前作为gly-tol
tm
、、gt和roundup销售。草铵膦和/或双丙氨磷化合物的抗性基因包括dsm-2、bar和pat基因。bar和pat性状目前作为销售。还包括提供对2,4-d的抗性的耐受性基因,如aad-1基因(应注意aad-1基因对芳氧基苯氧基丙酸酯除草剂具有进一步的活性)和aad-12基因(应注意aad-12基因对乙酰氧基乙酸酯合成植物生长素具有进一步的活性)。这些性状作为作物保护技术销售。als抑制剂(磺酰脲类、咪唑啉酮类、三唑并嘧啶类、嘧啶基硫代苯甲酸酯类、和磺酰基氨基-羰基-三唑啉酮类)的抗性基因是本领域已知的。这些抗性基因最通常是由点突变为als编码基因序列引起的。其他的als抑制剂抗性基因包括hra基因、csr1-2基因、sr-hra基因和surb基因。一些性状以商品名销售。抑制hppd的除草剂包括吡唑啉酮,如苄草唑,吡草酮和苯吡唑草酮;三酮,如硝磺草酮、磺草酮、环磺酮、苯并双环酮;和二酮腈,如异噁唑草酮。已知性状可以耐受这些示例性hppd除草剂可。hppd抑制剂的实例包括hppdpf_w336基因(用于抗异噁唑草酮)和avhppd-03基因(用于抗甲基磺草酮)。奥昔尼除草剂耐受性状的实例包括bxn基因,该基因已被证明对除草剂/抗生素溴苯腈具有抗性。麦草畏的抗性基因包括麦草畏单加氧酶基因(dmo),如国际pct公开号wo 2008/105890中所公开的。ppo或protox抑制剂型除草剂的抗性基因(例如氟锁草醚、氟丙嘧草酯、氟丙草酯、戊基恶唑酮、唑草酮、异丙吡草酯、吡草醚、苯草醚、唑啶草酮、丙炔氟草胺、氟烯草酸、治草醚、乙氧氟草醚、乳氟禾草灵、氟磺胺草醚、乙羧氟草醚、和甲磺草胺)是本领域已知的。赋予对ppo的抗性的示例性基因包括过表达野生型拟南芥ppo酶(lermontova i和grimm b,(2000)overexpression of plastidic protoporphyrinogen ix oxidase leads to resistance to the diphenyl-ether herbicide acifluorfen[质体原卟啉原ix氧化酶的过表达导致了对二苯基醚除草剂酸氟芬的抗性].plant physiol[植物生理学]122:75-83.)、枯草芽孢杆菌(b.subtilis)ppo基因(li,x.和nicholl d.2005.development of ppo inhibitor-resistant cultures and crops[ppo抑制剂抗性培养物和作物的开发].pest manag.sci.[有害生物管理科学]61:277-285以及choi kw,han o,lee hj,yun yc,moon yh,kim mk,kuk yi,han su和guh jo,(1998)generation of resistance to the diphenyl ether herbicide,oxyfluorfen,via expression of the bacillus subtilis protoporphyrinogen oxidase gene in transgenic tobacco plants[经由在转基因烟草植物中表达枯草芽孢杆菌原卟啉原氧化酶基因来产生对二苯基醚除草剂氧氟芬的抗性].biosci biotechnol biochem[生物科学、生物技术和生物化学]62:558-560)。吡啶氧基或苯氧基丙酸和环己酮的抗性基因包括编码accase抑制剂的基因(例如acc1-s1、acc1-s2和acc1-s3)。赋予对环己二酮和/或芳氧基苯氧基丙酸的抗性的示例性基因包括吡氟氯禾灵、禾草灵、精恶唑禾草灵
酸、吡氟禾草灵和喹禾灵。最后,除草剂可以抑制光合作用,包括三嗪或苄腈,通过psba基因(对三嗪的耐受性)、1s 基因(对三嗪的耐受性)和腈水解酶基因(对苯甲腈的耐受性)提供了耐受性。以上除草剂耐受性基因的列表并不意味着具有限制性。本公开涵盖任何除草剂耐受性基因。
[0146]
3.农学性状
[0147]
各种农学性状基因可以通过包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件进行表达。包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件可以与至少一个含有农学性状基因的其他基因表达盒可操作地连接。然后可将可操作地连接的序列掺入选择的载体中以允许鉴定和选择经转化的植物(“转化体”)。示例性农学性状编码序列是本领域已知的。作为可与本公开的调节元件可操作地连接的农学性状编码序列的实施例,提供以下性状。pg基因提供的延迟的果实软化抑制了导致细胞壁中果胶分子分解的聚半乳糖醛酸酶的产生,从而导致了果实的延迟软化。此外,延迟的acc基因果实成熟/衰老抑制了天然acc合酶基因的正常表达,导致乙烯产量减少和果实成熟延迟。而accd基因代谢果实成熟激素乙烯的前体,导致果实成熟延迟。可替代地,sam-k基因通过减少s-腺苷甲硫氨酸(sam)(乙烯生产的底物)而导致延迟成熟。cspb基因提供的干旱胁迫耐受表型通过保持rna稳定性和翻译来维持水分胁迫条件下的正常细胞功能。另一个实例包括ecbeta基因,其催化渗透保护剂化合物甘氨酸甜菜碱的产生,赋予了对水分胁迫的耐受性。另外,rmbeta基因催化渗透保护剂化合物甘氨酸甜菜碱的产生,赋予了对水分胁迫的耐受性。bbx32基因提供光合作用和增产,该基因表达一种蛋白质,该蛋白质与一种或多种内源性转录因子相互作用以调节植物的昼/夜生理过程。可以通过表达编码热稳定的α-淀粉酶的amy797e基因来增加乙醇产量,该酶可以通过增加用于降解淀粉的淀粉酶的热稳定性来增强生物乙醇的产量。最后,修饰的氨基酸组合物可以通过编码二氢二吡啶甲酸合酶的cordapa基因的表达而产生,该酶增加了氨基酸赖氨酸的产生。农学性状编码序列的列表并不意指是限制性的。本公开涵盖任何农学性状编码序列。
[0148]
4.dna结合蛋白质
[0149]
各种dna结合转基因/异源编码序列可以通过包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件进行表达。包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件可以与至少一个含有dna结合基因的其他基因表达盒可操作地连接。然后可将可操作地连接的序列掺入选择的载体中以允许鉴定和选择经转化的植物(“转化体”)。示例性dna结合蛋白质编码序列是本领域已知的。作为可与本公开的调节元件可操作地连接的dna结合蛋白编码序列的实施例,以下类型的dna结合蛋白可包括:锌指、talen、crispr和大范围核酸酶。dna结合蛋白质编码序列的列表并不意指是限制性的。本公开涵盖任何dna结合蛋白质编码序列。
[0150]
5.小rna
[0151]
各种小rna序列可以通过包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件进行表达。包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件可以与至少一个含有小rna序列的其他基因表达盒可操作地连接。然后可将可操作地连接的序列掺入选择的载体中以允许鉴定和选择经转化的植物(“转化体”)。示例性小rna性状是本领域已知的。作为可与本公开的调节元件可操作地连接的小
rna编码序列的实施例,提供以下性状。例如,通过经由沉默编码乙烯形成酶的aco基因的表达来抑制乙烯的产生,抗efe小rna的延迟果实成熟/衰老延迟了成熟。通过抑制内源性s-腺苷-l-蛋氨酸,改变了ccomt小rna的木质素产生,从而降低了胍基(g)木质素的含量:反式咖啡酰氧基coa 3-o-甲基转移酶(ccomt基因)。此外,可通过ppo5小rna减少疣状茄(solanum verrucosum)中黑斑瘀伤耐受性,该ppo5小rna触发ppo5转录物的降解,从而阻止黑点瘀伤的发展。还包括dvsnf7小rna,其dsrna包含西方玉米根虫(western corn rootworm)snf7基因的240bp片段,可抑制西方玉米根虫。修饰的淀粉/碳水化合物可以由小rna产生,如prhl小rna(降解phl转录物以限制通过淀粉降解形成还原糖)和pr1小rna(降解r1转录物以限制通过淀粉降解形成还原糖)。另外,益处还包括asn1小rna引起的丙烯酰胺含量降低,该asn1小rna触发asn1降解从而损害天冬酰胺的形成并降低了聚丙烯酰胺含量。最后,pgas ppo抑制小rna的非褐色表型导致抑制ppo以产生具有非褐色表型的苹果。以上小rna的列表并不意指是限制性的。本公开涵盖任何小rna编码序列。
[0152]
6.选择性标记
[0153]
各种选择性标记(也称为报告基因)可以通过包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件进行表达。包含seq id no:9、seq id no:25-583、seq id no:585-2268的嵌合调节元件可以与至少一个含有报告基因的其他基因表达盒可操作地连接。然后可将可操作地连接的序列掺入选择的载体中以允许鉴定和选择经转化的植物(“转化体”)。有许多方法可用于确认选择性标记在转化植物中的表达,包括例如dna测序和pcr(聚合酶链式反应)、dna印迹、rna印迹、用于检测从载体表达的蛋白质的免疫学方法。但是,通常通过目测观察蛋白质时观察到的报告基因,这些蛋白质在表达时会产生有色产物。示例性的报告基因是本领域已知的,并编码β-葡萄糖醛酸苷酶(gus)、萤光素酶、绿色荧光蛋白(gfp)、黄色荧光蛋白(yfp、phi-yfp)、红色荧光蛋白(dsrfp、rfp等)、β-半乳糖苷酶等(参见sambrook等人,molecular cloning:a laboratory manual[分子克隆:实验室手册],第三版,冷泉港出版社,纽约,2001,其内容通过引以其全文并入本文)。
[0154]
利用选择性标记基因来选择经转化的细胞或组织。选择性标记基因包括编码抗生素抗性的基因,如编码新霉素磷酸转移酶ii(neo)、壮观霉素/链霉素抗性(aad)和潮霉素磷酸转移酶(hpt或hgr)的基因,以及赋予对除草化合物抗性的基因。除草剂抗性基因通常编码对除草剂不敏感的修饰目标蛋白,或编码在其起作用之前能降解植物中的除草剂或使其解毒的酶。例如,已经通过使用编码突变目标酶5-烯醇式丙酮酸莽草酸酯-3-磷酸合酶(epsps)的基因获得了对草甘膦的抗性。epsps的基因和突变体是熟知的,并且在下面进一步描述。通过使用编码pat或dsm-2、腈水解酶、aad-1或aad-12(这些都是使其对应的除草剂去毒的蛋白质的实例)的细菌基因分别获得了对草铵膦、溴苯腈和2,4-二氯苯氧基乙酸(2,4-d)的抗性。
[0155]
在实施例中,除草剂可抑制生长点或分生组织,包括咪唑啉酮或磺酰脲,并且针对这些除草剂的乙酰羟酸合酶(ahas)和乙酰乳酸合酶(als)抗性/耐受性基因是众所周知的。草甘膦抗性基因包括突变型5-烯醇式丙酮酸莽草酸酯-3-磷酸合酶(epsp)和dgt-28基因(经由引入重组核酸和/或对天然epsp基因进行各种体内诱变)、aroa基因和草甘膦乙酰基转移酶(gat)基因。其他膦酰基化合物的抗性基因包括来自链霉菌种的bar和pat基因,包括吸水链霉菌(streptomyces hygroscopicus)和streptomyces viridichromogenes,以及吡
啶氧基或苯氧基丙酸和环己酮(编码accase抑制剂的基因)。赋予对环己二酮和/或芳氧基苯氧基丙酸的抗性的示例性基因(包括吡氟氯禾灵、禾草灵、精恶唑禾草灵酸、吡氟禾草灵和喹禾灵)包括乙酰辅酶a羧化酶(accase)的基因;acc1-s1、acc1-s2和acc1-s3。在实施例中,除草剂可以抑制光合作用,包括三嗪(psba和1s 基因)或苄腈(硝化酶基因)。此外,此类选择性标记可以包括阳性选择标记,如磷酸甘露糖异构酶(pmi)酶。
[0156]
在实施例中,选择性标记基因包括但不限于编码如下的基因:2,4-d;新霉素磷酸转移酶ii;氰酰胺水合酶;天冬氨酸激酶;二氢吡啶二羧酸合酶;色氨酸脱羧酶;二氢吡啶二羧酸合酶和脱敏天冬氨酸激酶;bar基因;色氨酸脱羧酶;新霉素磷酸转移酶(neo);潮霉素磷酸转移酶(hpt或hyg);二氢叶酸还原酶(dhfr);草丁膦乙酰转移酶;2,2-二氯丙酸脱卤素酶;乙酰羟酸合成酶;5-烯醇式丙酮酸-莽草酸酯-磷酸合酶(aroa);卤代芳基腈水解酶;乙酰辅酶a羧化酶;二氢蝶呤合酶(sul i);和32kd光系统ii多肽(psba)。一个实施例还包括选择性标记基因,其编码对以下的抗性:氯霉素;甲氨蝶呤;潮霉素;壮观霉素;溴草腈;草甘膦;和草丁膦。以上选择性标记基因的列表并不意在是限制性的。本公开涵盖任何报告基因或选择性标记基因。
[0157]
在一些实施例中,合成编码序列以在植物中最佳表达。例如,在实施例中,已经通过密码子优化修饰了基因的编码序列以增强在植物中的表达。可以优化杀昆虫抗性转基因、除草剂耐受性转基因、氮利用效率转基因、水利用效率转基因、营养品质转基因、dna结合转基因、或选择性标记转基因/异源编码序列,以在特定植物物种中表达,或可替代地可以修饰上述转基因/异源编码序列以在双子叶或单子叶植物中最佳表达。植物偏好性密码子可以从特定目的植物物种中以最大量表达的蛋白质中频率最高的密码子确定。在实施例中,编码序列、基因、异源编码序列或转基因/异源编码序列被设计成在植物中以更高的水平表达,从而导致更高的转化效率。植物基因优化的方法是众所周知的。关于合成dna序列的优化和产生的指导可以在例如wo 2013016546、wo 2011146524、wo 1997013402、美国专利号6166302和美国专利号5380831(通过引用并入本文)中找到。
[0158]
分子确认
[0159]
确认植物基因组内存在seq id no:9、seq id no:25-583、seq id no:585-2268的方法是本领域已知的。例如,可以通过例如聚合酶链式反应(pcr)实现对植物基因组内的seq id no:9、seq id no:25-583、seq id no:585-2268的检测。通过使用多态性的多态区段侧翼的两个寡核苷酸引物进行pcr检测,然后进行dna扩增。该步骤包括dna热变性,然后在低温下将引物退火至它们的互补序列,并用dna聚合酶延伸退火的引物的重复循环。扩增后在琼脂糖或聚丙烯酰胺凝胶上对dna片段进行大小分离是该方法的主要部分。此类选择和筛选方法是本领域技术人员熟知的。可用于鉴定转基因植物的分子确认方法是本领域技术人员已知的。下面进一步描述几种示例性方法。
[0160]
已经描述了用于序列检测中的分子信标。简而言之,设计了一种fret寡核苷酸探针,该探针与侧翼基因组和插入dna连接重叠。fret探针的独特结构导致它含有一个二级结构,该二级结构使荧光和猝灭部分保持紧密相邻。fret探针和pcr引物(插入dna序列中的一个引物和侧翼基因组序列中的一个引物)在热稳定的聚合酶和dntp存在下循环。成功进行pcr扩增后,一种或多种fret探针与靶序列的杂交导致探针二级结构的去除以及荧光和淬灭部分的空间分离。荧光信号表明由于成功的扩增和杂交,存在侧翼基因组/转基因插入序
583、seq id no:585-2268衍生序列的顺式作用调节元件包含与seq id no:9、seq id no:25-583、seq id no:585-2268具有80%、85%、90%、95%或99.5%序列同一性的序列。在一个实施例中,提供了植物、植物组织、或植物细胞,其中该植物、植物组织、或植物细胞包含可操作地连接到异源编码序列的顺式作用调节元件,该顺式作用调节元件包含seq id no:9、seq id no:25-583、seq id no:585-2268或与seq id no:9、seq id no:25-583、seq id no:585-2268具有80%、85%、90%、95%或99.5%序列同一性的序列。根据一个实施例,植物、植物组织、或植物细胞包含可操作地连接到异源编码序列的嵌合调节分子,该嵌合调节分子包含seq id no:9、seq id no:25-583、seq id no:585-2268或与seq id no:9、seq id no:25-583、seq id no:585-2268具有80%、85%、90%、95%或99.5%序列同一性的序列。在一个实施例中,植物、植物组织、或植物细胞包含可操作地连接到转基因/异源编码序列的嵌合调节分子,其中该嵌合调节分子由seq id no:2或与seq id no:9、seq id no:25-583、seq id no:585-2268具有80%、85%、90%、95%或99.5%序列同一性的序列组成。根据一个实施例,包含可操作地连接到转基因/异源编码序列的嵌合调节分子的基因构建体被掺入植物、植物组织、或植物细胞的基因组中,该嵌合调节分子包含seq id no:9、seq id no:25-583、seq id no:585-2268。
[0174]
在一个实施例中,植物、植物组织或植物细胞是双子叶植物或单子叶植物或者衍生自双子叶植物或单子叶植物的细胞或组织。在一个实施例中,植物选自由以下组成的组:小麦、水稻、高粱、燕麦、黑麦、香蕉、甘蔗、大豆、棉花、向日葵(sunflower)、玉蜀黍、苜蓿、油菜籽、卡诺拉油菜、印度芥菜、埃塞俄比亚芥菜、豆类、西兰花、卷心菜、花椰菜、芹菜、黄瓜(cucumber)、茄子、莴苣(lettuce);瓜、豌豆、辣椒(pepper)、花生、马铃薯(potato)、南瓜、萝卜、菠菜、甜菜、向日葵、烟草、番茄和西瓜。
[0175]
本领域技术人员将认识到,在将外源序列稳定地掺入转基因植物中并确认是可操作的之后,可以通过有性杂交将其引入其他植物。可以使用许多标准育种技术中的任何一种,这取决于待杂交的物种。
[0176]
本公开还涵盖上述转基因植物的种子,其中所述种子具有含有本公开的基因调控元件的转基因/异源编码序列或基因构建体。本公开进一步涵盖上述转基因植物的后代、克隆、细胞系或细胞,其中所述后代、克隆、细胞系或细胞具有含有本公开的基因调控元件的转基因/异源编码序列或基因构建体。
[0177]
本公开还涵盖上述转基因植物的培养,其中所述转基因植物具有含有本公开的基因调控元件的转基因/异源编码序列或基因构建体。因此,此类转基因植物可被工程化以尤其具有一个或多个所希望的性状或含有本公开的基因调节元件的转基因事件,通过与根据本发明的核酸分子被转化,并且可以通过本领域技术人员已知的任何方法被裁剪或培养。
[0178]
表达转基因的方法
[0179]
在实施例中,在植物中表达至少一个转基因/异源编码序列的方法包括使包含可操作地连接到至少一个转基因/异源编码序列或多接头序列的嵌合调节分子的植物生长,该嵌合调节分子包含seq id no:9、seq id no:25-583、seq id no:585-2268。在实施例中,顺式作用调节元件或嵌合调节元件由选自seq id no:9、seq id no:25-583、seq id no:585-2268的序列或与选自seq id no:9、seq id no:25-583、seq id no:585-2268的序列具有80%、85%、90%、95%或99.5%序列同一性的序列组成。在实施例中,在植物中表达至少
一个转基因/异源编码序列的方法包括使包含可操作地连接到至少一个转基因的嵌合调节分子的植物生长,该嵌合调节分子包含seq id no:9、seq id no:25-583、seq id no:585-2268。在实施例中,在植物组织或植物细胞中表达至少一个转基因/异源编码序列的方法包括培养包含可操作地连接到至少一个转基因的嵌合调节分子的植物组织或植物细胞,该嵌合调节分子包含seq id no:9、seq id no:25-583、seq id no:585-2268。
[0180]
在实施例中,在植物中表达至少一个转基因/异源编码序列的方法包括使包含基因表达盒的植物生长,该基因表达盒包含可操作地连接到至少一个转基因的嵌合调节分子,该嵌合调节分子包含seq id no:9、seq id no:25-583、seq id no:585-2268。在一个实施例中,顺式作用调节元件或嵌合调节元件由选自seq id no:9、seq id no:25-583、seq id no:585-2268的序列或与选自seq id no:9、seq id no:25-583、seq id no:585-2268的序列具有80%、85%、90%、95%或99.5%序列同一性的序列组成。在实施例中,在植物中表达至少一个转基因/异源编码序列的方法包括使包含基因表达盒的植物生长,该基因表达盒包含可操作地连接到至少一个转基因的嵌合调节分子,该嵌合调节分子包含seq id no:9、seq id no:25-583、seq id no:585-2268。在实施例中,在植物中表达至少一个转基因/异源编码序列的方法包括使包含基因表达盒的植物生长,该基因表达盒包含可操作地连接到至少一个转基因的嵌合调节分子,该嵌合调节分子包含seq id no:9、seq id no:25-583、seq id no:585-2268。在实施例中,在植物组织或植物细胞中表达至少一个转基因/异源编码序列的方法包括培养包含基因表达盒的植物组织或植物细胞,该基因表达盒含有可操作地连接到至少一个转基因的嵌合调节分子,该嵌合调节分子包含seq id no:9、seq id no:25-583、seq id no:585-2268。在实施例中,在植物组织或植物细胞中表达至少一个转基因/异源编码序列的方法包括培养包含基因表达盒的植物组织或植物细胞,该基因表达盒含有可操作地连接到至少一个转基因的嵌合调节分子,该嵌合调节分子包含seq id no:9、seq id no:25-583、seq id no:585-2268。
[0181]
本文引用的所有参考文献(包括出版物、专利和专利申请)均以与本公开的明确细节不矛盾的程度通过引用特此并入,并且并入程度如同每个参考文献是单独并且具体地指示通过引用并入并且在本文以其全文阐述一般。本文讨论的参考文献只是为了它们在本技术的提交日之前的公开内容而提供。本文的任何内容不应解释为承认诸位发明人无权因在先发明而享有早于此类公开内容的权利。
[0182]
在以下实例中进一步例示本公开的实施例。应当理解的是,这些实例仅是通过说明的方式给出的。从上述实施例和以下实例中,本领域的技术人员能够确定本公开的本质特性,并且在不脱离本公开的精神和范围的情况下,可进行本公开的实施例的各种变化和修改以使其适应各种用途和条件。因此,从上述说明书来看,除了本文所示出和描述的那些之外,本公开的实施例的各种修改对于本领域技术人员来说是显而易见的。此类修改也旨在落入所附权利要求的范围内。以下内容以说明的方式提供,并不旨在限制本发明的范围。
[0183]
实例
[0184]
实例1:顺式作用调节元件的鉴定
[0185]
顺式作用调节元件获得自place数据库(higo,k.,ugawa,y.,iwamoto,m.和korenaga,t.1999.
″
plant cis-acting regulatory dna elements(place)database:1999[植物顺式作用调节dna元件(place)数据库:1999]
″
nar.[核酸研究]27∶297-300)。元件按
大小排序,并基于以下标准选择具有十个元件的子集进行测试:1)长度在18到24个核苷酸之间(包含端值),2)序列不会过于简单,例如
‘
tttttaaaaa’,以及3)不包含模糊核苷酸。获得的顺式作用调节元件如下列出:seq id no:1-10。所选元件及其详细信息示于表1中。
[0186][0187]
实例2:作为调节元件(例如启动子调节元件)内的调控元件的顺式作用调节元件的合成
[0188]
将所选元件作为调节元件(例如启动子调节元件)内的调控元件进行工程化。将调节元件作为具有间插序列(
‘
seq id no:11;cataaacacc’和
‘
seq id no:12;gggcacgcgtc’分别作为5
′
和3
′
位置中的间隔区)的盒的多聚体融合到启动子。在该实例中,将三个盒合成到启动子中。通过南京金斯瑞生物科技有限公司(genscript biotech)(皮斯卡塔韦,新泽西州)将这三个盒与camv35s最小启动子(seq id no:13)和tmv ω 5’utr(seq id no:14)一起合成,并位于其上游。限制性内切酶位点ecori和ncoi分别包含在构建体的5’末端和3’末端,用于克隆到标准化双子叶原生质体测试载体中。包含顺式作用调节元件的最终嵌合启动子作为seq id no:15-24提供。
[0189]
实例3:构建体设计
[0190]
将嵌合顺式作用调节元件和启动子作为嵌合调节元件进行测试,以确定该顺式作用调节元件是否可以增强可操作地连接到该嵌合顺式作用调节元件和启动子的下游编码序列的表达。这样,将嵌合顺式作用调节元件和调节元件(例如启动子调节元件)克隆到编码荧光蛋白的编码序列的上游。标准化双子叶原生质体测试载体含有两个报告蛋白荧光团,即zsgreen(matz,m.v.,fradkov,a.f.,labas,y.a.,zaraisky,a.g.,markelov,m.l.,和lukyanov,s.a.1999.fluorescent proteins from the nonbioluminescent anthozoa species.[来自非生物发光珊瑚虫物种的荧光蛋白]nat biotechnol.[自然生物技术]17:969-973.),用于测量调节元件对转录的影响,以及tagrfp(merzlyak,e.m.,goedhart,j.,shcherbo,d.,bulina,m.e.,shcheglov,a.s.fradkov,a.f.,gaintzeva,a.,lukyanov,k.a.,lukyanov,s.,gadella,t.w.j.和chudakov,d.m.2007.bright monomeric red fluorescent protein with an extended fluorescence lifetime.[荧光寿命延长的明亮单体红色荧光蛋白]nat methods.[自然方法]4∶555-557.),用于归一化。zsgreen编码序列由如上所述的测试盒驱动。tagrfp编码序列由拟南芥泛素启动子at-ubiq10驱动。两个表达构建体由四个终止子pinii(an,g.,mitra,m.,choi,h.k.,costa,m.a.,an,k.,thornburg,r.w.1989.functional analysis of the 3’control region of the potato wound-inducibleproteinase inhibitor ii gene.[马铃薯创伤诱导型蛋白酶抑制剂ii基因3
′
控制区的功能分析]plant cell.[植物细胞]1:115-122.)、w64a(das等人,1991)、ubq14(mayer等人,1999)和in2-1(hershey和stoner,1991)分开,以防止通读转录。载体上包含壮观霉素抗性基因,作为克隆目的的选择性标记。载体的细菌复制起点是puc ori。
[0191]
实例4:顺式作用调节元件的植物原位测试
[0192]
使用大豆下胚轴原生质体测定来测试载体,以确定顺式作用调节元件是否能够增强下游编码序列的表达,该下游编码序列可操作地连接到嵌合顺式作用调节元件和作为嵌合调节元件的调节元件(例如启动子调节元件)。从4-5日龄大豆(基因型93y21)幼苗的经切片(0.5-1mm)的下胚轴组织中分离原生质体,并使用wu和hanzawa(2018)中描述的溶液和方法进行分离,这些溶液和方法具有以下修改:1)在所有相关溶液中使用0.75m甘露醇,2)真空浸润0.5h,3)在没有搅拌的情况下在消化溶液中孵育2.5h,4)离心后,将原生质体重新悬浮在mmg中并使其在冰上静置1h,5)静置后不对原生质体进行离心,而是在不干扰细胞沉淀物的情况下除去上清液并用新鲜mmg代替。按照wu,f.和hanzawa,y.a simple method for isolating of soybean protoplasts and application to transient gene expression analyses.[一种分离大豆原生质体并应用于瞬时基因表达分析的简单方法]j vis exp.
[可视实验杂志]131:e57258的方法,对原生质体进行聚乙二醇(peg)转染。对wu和hanzawa方案进行了一些修改:在转染溶液中使用1)浓度为0.75m的甘露醇和2)浓度为40%的peg,以及3)在转染反应中使用总共2.5nm的质粒dna。所有转染以一式两份进行。转染后,将原生质体铺板至24孔黑色玻璃底板中,并在27℃于黑暗中储存16h。使用cytation5(美国伯腾仪器有限公司(biotek),威努斯基,佛蒙特州)测量原生质体发出的荧光信号。测定中的对照包括空载体,即无测试盒,作为背景对照(ee1906),以及模拟处理的原生质体(无dna)。收集的数据是在细胞(对象)水平上的,其中每个载体计数不少于300个原生质体。除了荧光信号外,还测定了每个原生质体的大小和圆度。
[0193]
实例5:增加编码序列表达的顺式作用调节元件的鉴定
[0194]
为了作为
‘
原生质体’计算,通过cytation5
tm
检测和测量的对象必须1)发出红色荧光,2)在20-150μm之间,3)具有等于或大于0.49的圆度值(滚动球直径为300μm)。对于每个原生质体,将绿色荧光信号数据点(从zsgreen发出)除以其相应的红色荧光数据点(从tagrfp发出)以使转染效率归一化,并计算每个载体的几何平均值(平均值)和平均值的标准误差(sem)。数据以“相对荧光单位”(rfu)报告。活性元件被定义为任何rfu比背景对照高4倍或更多倍。
[0195]
一种构建体ee2889,对应于jerecrestr顺式作用调节元件(seq id no:9),作为嵌合调节元件展示出作为调控元件的增强活性(图1)。该顺式作用调节元件驱动荧光编码序列的稳健表达。其余的元件没有显示活性,甚至没有显示轻微的活性,这表明并非place数据库中列出的所有元件都可以通过仅将它们放置在基因的调节元件(例如启动子调节元件)中而用作调控元件。这样,jerecrestr顺式作用调节元件(seq id no:9)首次被例示为能够在嵌合调节元件内充当调控元件以驱动编码序列的表达的顺式作用调节元件。
[0196]
实例6:增加编码序列表达的另外的顺式作用调节元件的鉴定
[0197]
顺式作用调节元件是从玄参花叶病毒(fmv;sanger,m.,daubert,s.和goodman,r.m.1990.characteristics of a strong promoter from figwort mosaic virus:comparison with the analogous 35s promoter from cauliflower mosaic virus and the regulated mannopine synthase promoter.[玄参花叶病毒强启动子的特征:与来自花椰菜花叶病毒的类似35s启动子和受调控的甘露碱合酶启动子的比较]plant mol biol.[植物分子生物学]14:433-443.)和紫茉莉花叶病毒(mmv;dey,n.和maiti,i.b.1999.structure and promoter/leader deletion analysis of mitabilis virus(mmv)full-length transcription promoter in transgenic plants.[转基因植物中紫茉莉花叶病毒(mmv)全长转录启动子的结构和启动子/前导序列缺失分析]plant mol biol.[植物分子生物学]40:771-782.)的带注释的全长转录启动子中获得的。获得的顺式作用调节元件如下列出:seq id no:25和seq id no:35,并在表2和3中提供。开发了seq id no:25的变体,并在表2中进一步提供为seq id no:26-34。seq id no:25-34的序列比对如图2所提供。同样,开发了seq id no:35的变体,并在表3中进一步提供为seq id no:36-38。seq id no:36-38的序列比对如图3所提供。将这些调节元件工程化于调节元件(例如植物启动子)的上游,并用于产生如实例2和3中所述的标准化双子叶原生质体测试载体。最后,在如实例4中所述的大豆下胚轴原生质体测定中对顺式作用调节元件进行测试,以测试载体来确定顺式作用调节元件是否能够增强下游编码序列的表达,该下游编码序列可操作地
连接到嵌合顺式作用调节元件和充当嵌合调节元件的另一调节元件(例如启动子调节元件)。
[0198]
[0199]
表达谱分析的结果表明,mmv as-1顺式作用调节元件(seq id no:25)及其变体(seq id no:26-34)作为嵌合调节元件(例如顺式作用调节元件和启动子调节元件)内的调控元件展示出优异的活性(图4)。该顺式作用调节元件驱动荧光编码序列的稳健表达。尽管与seq id no:25相比,荧光编码序列的表达存在一定程度的变化,但所有的变体序列的表达均高于camv 35s最小启动子对照。这样,首次将mmv as-1顺式作用调节元件(seq id no:25)和mmv as-1植物衍生变体顺式作用调节元件(seq id no:26-34)例示为能够在嵌合调节元件(例如启动子和顺式作用调节元件)内充当调控元件以驱动编码序列的表达的顺式作用调节元件。
[0200]
表达谱分析的结果表明,fmv as-1顺式作用调节元件(seq id no:35)及其变体(seq id no:36-38)作为嵌合调节元件内的调控元件展示出增强的活性(图5)。该顺式作用调节元件驱动荧光编码序列的稳健表达。尽管与seq id no:33相比,荧光编码序列的表达存在一定程度的变化,但所有的变体序列的表达均高于camv 35s最小启动子对照。这样,首次将fmv as-1顺式作用调节元件(seq id no:35)和fmv as-1植物衍生变体顺式作用调节元件(seq id no:36-38)例示为能够在嵌合调节元件(例如启动子和顺式作用调节元件)内充当调控元件以驱动编码序列的表达的顺式作用调节元件。
[0201]
实例7:基于生物信息学分析表达元件以鉴定推定的顺式基因表达元件。
[0202]
在包括玉蜀黍(探索了源自内部泛基因组研究的各种基因型a63、csb8v、ed85e、ede4n、eecpr、eepar、gh61、gr3kp、gr84z、grw2z、hef3d、hn4cn、hnh9h)、大豆(glycine max)、水稻、高粱(sorghum bicolor)、甘蓝型油菜(brassica napus)在内的多个目的基因组中扫描了经鉴定的公共eme基序的变体(mmv、gvbav、csvmv、mmv-1、gm-psgs3af1-v3和jerecrstr)。这些变体的基因组序列从内部gaia储存库、ncbi和/或ensemblplants下载。
[0203]
首先将从公共eme基序获得的序列转换为表示特定位置碱基概率的位置权重矩阵(pwm)。一旦获得每个eme序列的pwm,然后使用查找出现的单个基序(find individual motif occurrences)(fimo)将pwm用于扫描整个基因组。该程序计算给定序列数据库(在我们的情况下,目的作物的全基因组序列)中每个位置的对数似然比得分,使用已建立的动态规划方法(假设零阶零模型,其中序列以用户指定的每个字母背景频率随机生成)将该得分转换为p值,然后应用错误发现率分析(使用storey jd.a direct approach to false discovery rates.[错误发现率的直接方法]journal ofthe royal statistical society:series b[皇家统计学会杂志:b辑],64:479-498,2002.提出的自举方法)估计给定序列中每个位置的q值(grant ce,bailey tl,noble ws.fimo:scanning for occurrences of a given motif.[fimo:扫描给定基序的出现]bioinformatics[生物信息学],27(7):1017-1018,2011)。输出是扫描的基因组中的基序(eme)出现的排序列表,每个都有相关的对数似然比得分、p值和q值。然后使用排名靠前的变体进行后续表征。
[0204]
实例8:增加编码序列表达的mmv-1大豆变体顺式作用调节元件的鉴定
[0205]
通过对植物基因组序列数据库(例如,水稻、大豆、卡诺拉油菜、玉蜀黍和高粱)进行mmv as-1遗传元件(seq id no:27)筛选,获得顺式作用调节元件。经由meme套件中的fimo算法使用前100次命(冗余)中的上限完成筛选(charles e.grant,timothy l.bailey,和william stafford noble,"fimo:scanning for occurrences of a given motif[fimo:扫描给定基序的出现]
″
,bioinformatics[生物信息学],27(7):1017-1018,2011)。
对植物基因组序列数据库的分析导致鉴定了大量与mmv as-1遗传元件具有相对高水平序列同一性的序列。进一步筛选序列库,以鉴定用于植物原位测试的推定的顺式作用调节元件。基于活性调节元件的预测特征对这些序列进行分选和选择,这些预测特征例如包括完整的bzip结合基序(例如,
‘
acgt’)和与用于测试的亲本元件具有高水平序列同一性的序列。
[0206]
获得的顺式作用调节元件如下列出:seq id no:508-seq id no:515,并在表4中提供。508-seq id no:515的序列比对如图7所提供。将这些调节元件工程化于调节元件(例如植物启动子)的上游,并用于产生如实例2和3中所述的标准化双子叶原生质体测试载体。顺式作用调节元件一式两份地掺入构建体中。本领域技术人员将理解,顺式作用调节元件可以作为单体、二聚体、三聚体或以任何拷贝数被包含以驱动编码序列的稳健表达。最后,在如实例4中所述的大豆下胚轴原生质体测定中对顺式作用调节元件进行测试,以测试载体来确定顺式作用调节元件是否能够增强下游编码序列的表达,该下游编码序列可操作地连接到嵌合顺式作用调节元件和充当嵌合调节元件的另一调节元件(例如启动子调节元件)。
[0207][0208]
表达谱分析的结果表明,mmv as-1顺式作用调节元件(seq id no:27)及从大豆获得的变体(seq id no:508-515)展示出作为嵌合调节元件(例如顺式作用调节元件和启动
子调节元件)内的调控元件的活性(图6)。该顺式作用调节元件驱动荧光编码序列在大豆原生质体中的稳健表达。本领域技术人员将理解,这些顺式作用调节元件还将驱动编码序列在其他植物中的稳健表达。尽管与seq id no:27相比,荧光编码序列的表达存在一定程度的变化,但所有的变体序列的表达均高于camv 35s最小启动子对照。这样,首次将mmv as-1顺式作用调节元件(seq id no:27)和mmv as-1植物衍生变体顺式作用调节元件(seq id no:508-515)例示为能够在嵌合调节元件(例如启动子和顺式作用调节元件)内充当调控元件以驱动编码序列的表达的顺式作用调节元件。
[0209]
实例9:增加编码序列表达的合成mmv-1变体顺式作用调节元件和合成g-盒结合基序顺式作用调节元件的鉴定
[0210]
通过分析mmv-1启动子获得顺式作用调节元件。最初,mmv-1启动子的70碱基对片段彼此重叠以产生共有序列。该序列覆盖了mmv-1启动子核心的360个碱基对,并且这些序列重叠了35个碱基对。对所得的序列进行植物原位测试,以确定是否这些推定的顺式作用调节元件中的任一个可用于稳健地驱动表达。一个片段表现出高于背景表达水平,并将其选择用于进一步表征。将活性区域缩小到29碱基对元件,并将其命名为mmv-eme1(seq id no:516)。使用类似于实例7中所述的生物信息学分析方法,使用经鉴定的mmv-eme1序列从玉蜀黍、大豆、卡诺拉油菜、水稻和高粱基因组中获得变体序列。
[0211]
获得的顺式作用调节元件如下列出:seq id no:516-seq id no:525,并在表5中提供。seq id no:516-seq id no:525的序列比对如图9所提供。将这些调节元件工程化于调节元件(例如植物启动子)的上游,并用于产生如实例2和3中所述的标准化双子叶原生质体测试载体。顺式作用调节元件一式两份地掺入构建体中。本领域技术人员将理解,顺式作用调节元件可以作为单体、二聚体、三聚体或以任何拷贝数被包含以驱动编码序列的稳健表达。最后,在如实例4中所述的大豆下胚轴原生质体测定中对顺式作用调节元件进行测试,以测试载体来确定顺式作用调节元件是否能够增强下游编码序列的表达,该下游编码序列可操作地连接到嵌合顺式作用调节元件和充当嵌合调节元件的另一调节元件(例如启动子调节元件)。
[0212][0213]
表达谱分析的结果表明,mmv-eme1顺式作用调节元件(seq id no:516)和许多其他变体(seq id no:517和seq id no:523-525)展示出作为嵌合调节元件(例如顺式作用调
节元件和启动子调节元件)内的调控元件的活性(图8)。该顺式作用调节元件驱动荧光编码序列在大豆原生质体中的稳健表达。本领域技术人员将理解,这些顺式作用调节元件还将驱动编码序列在其他植物中的稳健表达。尽管与seq id no:516相比,荧光编码序列的表达存在一定程度的变化,但所有的变体序列的表达均高于camv 35s最小启动子对照。这样,首次将mmv-eme1顺式作用调节元件(seq id no:516)和许多其他变体(seq id no:517和seq id no:523-525)例示为能够在嵌合调节元件(例如启动子和顺式作用调节元件)内充当调控元件以驱动编码序列的表达的顺式作用调节元件。
[0214]
实例10:增加编码序列表达的ps-gs3a变体顺式作用调节元件的鉴定
[0215]
通过分析从豌豆谷氨酰胺合成酶gs3a启动子(brears,t.,walker,e.l.,coruzzi,g.m.a promoter sequence involved in cell-specificexpression of the pea gluamine syntetas gs3a gene in organs of transgenic tobacco and alfalfa.[一种涉及转基因烟草和苜蓿的器官中豌豆谷氨酰胺合成酶gs3a基因的细胞特异性表达的启动子序列]plant j.[植物杂志]1(2)235-244)获得的ps-gs3a的调节元件来获得顺式作用调节元件。通过对大豆基因组序列数据库进行ps-gs3a遗传元件(seq id no:526)筛选,获得顺式作用调节元件。使用类似于实例7中所述的生物信息学分析方法。对植物基因组序列数据库的分析导致鉴定了大量与mmv as-1遗传元件具有相对高水平序列同一性的序列。进一步筛选序列库,以鉴定用于植物原位测试的推定的顺式作用调节元件。基于活性调节元件的预测特征对这些序列进行分选和选择,这些预测特征例如包括完整的bzip结合基序(例如,
‘
acgt’)和与用于测试的亲本元件具有高水平序列同一性的序列。对经鉴定的序列进行植物原位测试,以确定是否这些推定的顺式作用调节元件中的任一个可用于稳健地驱动表达。
[0216]
获得的顺式作用调节元件如下列出:seq id no:526-seq id no:531,并在表6中提供。seq id no:526-seq id no:531的序列比对如图11所提供。将这些调节元件工程化于调节元件(例如植物启动子)的上游,并用于产生如实例2和3中所述的标准化双子叶原生质体测试载体。顺式作用调节元件一式两份地掺入构建体中。本领域技术人员将理解,顺式作用调节元件可以作为单体、二聚体、三聚体或以任何拷贝数被包含以驱动编码序列的稳健表达。最后,在如实例4中所述的大豆下胚轴原生质体测定中对顺式作用调节元件进行测试,以测试载体来确定顺式作用调节元件是否能够增强下游编码序列的表达,该下游编码序列可操作地连接到嵌合顺式作用调节元件和充当嵌合调节元件的另一调节元件(例如启动子调节元件)。
[0217][0218]
表达谱分析的结果表明,ps-gs3a-f1顺式作用调节元件(seq id no:526)和许多大豆变体(seq id no:527-531)展示出作为嵌合调节元件(例如顺式作用调节元件和启动
子调节元件)内的调控元件的活性(图10)。该顺式作用调节元件驱动荧光编码序列在大豆原生质体中的稳健表达。本领域技术人员将理解,这些顺式作用调节元件还将驱动编码序列在其他植物中的稳健表达。尽管与ps-gs3a-f1顺式作用调节元件(seq id no:526)相比,荧光编码序列的表达存在一定程度的变化,但所有的变体序列的表达均高于seq id no:526。这样,首次将大豆变体(seq id no:517和seq id no:527-531)例示为能够在嵌合调节元件(例如启动子和顺式作用调节元件)内充当调控元件以驱动编码序列的表达的顺式作用调节元件。
[0219]
实例11:增加编码序列表达的来自病毒的顺式作用调节元件的鉴定
[0220]
通过使用类似于实例7中所述的生物信息学分析方法对病毒基因组序列数据库进行camv as-1遗传元件筛选获得顺式作用调节元件。对植物基因组序列数据库的分析导致鉴定了大量与camv as-1遗传元件具有相对高水平序列同一性的序列。进一步筛选序列库,以鉴定用于植物原位测试的推定的顺式作用调节元件。
[0221]
获得的顺式作用调节元件如下列出:seq id no:532-seq id no:550,并在表7中提供。seq id no:526-seq id no:531的序列比对如图13所提供。将这些调节元件工程化于调节元件(例如植物启动子)的上游,并用于产生如实例2和3中所述的标准化单子叶原生质体测试载体,其中单子叶启动子(玉蜀黍gos2启动子)被掺入构建体设计以驱动玉蜀黍原生质体中的表达。顺式作用调节元件一式两份地掺入构建体中。本领域技术人员将理解,顺式作用调节元件可以作为单体、二聚体、三聚体或以任何拷贝数被包含以驱动编码序列的稳健表达。最后,如专利申请wo 2018183878 a1的实例1和3中所述,在瞬时基因表达玉蜀黍叶肉原生质体平台中对顺式作用调节元件进行测试。完成分析以测试载体,以确定顺式作用调节元件是否能够增强下游编码序列的表达,该下游编码序列可操作地连接到嵌合顺式作用调节元件和充当嵌合调节元件的另一调节元件(例如启动子调节元件)。
[0222]
[0223]
[0224][0225]
表达谱分析的结果表明,camv as-1(seq id no:532-550)展示出作为嵌合调节元件(例如顺式作用调节元件和启动子调节元件)内的调控元件的活性(图12)。该顺式作用调
节元件驱动荧光编码序列的稳健表达。同样,植物fmv和mmv-1变体(seq id no:28-33和seq id no:37-38)在玉蜀黍原生质体内展示出作为嵌合调节元件(例如顺式作用调节元件和启动子调节元件)内的调控元件的活性(图12)。本领域技术人员将理解,这些顺式作用调节元件还将驱动编码序列在其他植物中的稳健表达。这样,首次将seq id no:532-550、seq id no:28-33和seq id no:37-38例示为能够在嵌合调节元件(例如启动子和顺式作用调节元件)内充当调控元件以驱动编码序列的表达的顺式作用调节元件。
[0226]
实例12:增加编码序列表达的来自place数据库的顺式作用调节元件的鉴定
[0227]
通过对如先前实例1中所述的place数据库进行筛选获得顺式作用调节元件。对植物基因组序列数据库的分析导致鉴定了玉蜀黍原生质体中大量待测试的序列。进一步筛选序列库,以鉴定用于植物原位测试的推定的顺式作用调节元件。
[0228]
获得的顺式作用调节元件如下列出:seq id no:551-seq id no:583,并在表8中提供。seq id no:551-seq id no:583的序列比对如图15所提供。将这些调节元件以及seq id no:2-5和seq id no:8-10工程化于调节元件(例如植物启动子)的上游,并用于产生如实例2和3中所述的标准化单子叶原生质体测试载体,其中单子叶启动子(玉蜀黍gos2启动子)被掺入构建体设计以驱动玉蜀黍原生质体中的表达。顺式作用调节元件一式两份地掺入构建体中。本领域技术人员将理解,顺式作用调节元件可以作为单体、二聚体、三聚体或以任何拷贝数被包含以驱动编码序列的稳健表达。最后,如专利申请wo 2018183878 a1的实例1和3中所述,在瞬时基因表达玉蜀黍叶肉原生质体平台中对顺式作用调节元件进行测试。完成分析以测试载体,以确定顺式作用调节元件是否能够增强下游编码序列的表达,该下游编码序列可操作地连接到嵌合顺式作用调节元件和充当嵌合调节元件的另一调节元件(例如启动子调节元件)。
[0229]
[0230]
[0231][0232]
表达谱分析的结果表明,seq id no:551-seq id no:583展示出作为嵌合调节元件(例如顺式作用调节元件和启动子调节元件)内的调控元件的活性(图14)。该顺式作用调
节元件驱动荧光编码序列在玉蜀黍原生质体中的稳健表达。本领域技术人员将理解,这些顺式作用调节元件还将驱动编码序列在其他植物中的稳健表达。这样,首次将seq id no:551-583、seq id no:2-5和seq id no:8-10例示为能够在嵌合调节元件(例如启动子和顺式作用调节元件)内充当调控元件以驱动编码序列的表达的顺式作用调节元件。
[0233]
实例13:增加编码序列在玉蜀黍原生质体内表达的顺式作用调节元件的鉴定
[0234]
来自实例9的gm-psgs3af1-v3(seq id no:529)和mmv-eme1(seq id no:516)调节元件是在玉蜀黍原生质体中进一步筛选的顺式作用调节元件,并且列于表9中。图17中提供了序列的比对。将每个调节元件工程化于调节元件(例如植物启动子)的上游,并用于产生如实例2和3中所述的标准化单子叶原生质体测试载体,其中单子叶启动子(玉蜀黍gos2启动子)被掺入构建体设计以驱动玉蜀黍原生质体中的表达。顺式作用调节元件一式两份地掺入构建体中。本领域技术人员将理解,顺式作用调节元件可以作为单体、二聚体、三聚体或以任何拷贝数被包含以驱动编码序列的稳健表达。最后,如专利申请wo 2018183878 a1的实例1和3中所述,在瞬时基因表达玉蜀黍叶肉原生质体平台中对顺式作用调节元件进行测试。完成分析以测试载体,以确定顺式作用调节元件是否能够增强下游编码序列的表达,该下游编码序列可操作地连接到嵌合顺式作用调节元件和充当嵌合调节元件的另一调节元件(例如启动子调节元件)。
[0235][0236]
表达谱分析的结果表明,seq id no:516和seq id no:529顺式作用调节元件展示出作为嵌合调节元件(例如顺式作用调节元件和启动子调节元件)内的调控元件的活性(图
16)。该顺式作用调节元件驱动荧光编码序列在玉蜀黍原生质体中的稳健表达。本领域技术人员将理解,这些顺式作用调节元件还将驱动编码序列在其他植物中的稳健表达。这样,首次将seq id no:516和seq id no:529例示为能够在嵌合调节元件(例如启动子和顺式作用调节元件)内充当调控元件以驱动编码序列的表达的顺式作用调节元件。
[0237]
实例14:增加编码序列在玉蜀黍原生质体内表达的顺式作用调节元件的鉴定
[0238]
通过对植物基因组序列数据库(例如,水稻、大豆、卡诺拉油菜、玉蜀黍和高粱)进行mmv as-1遗传元件(seq id no:25)筛选,获得顺式作用调节元件。经由meme套件中的fimo算法使用前100次命(冗余)中的上限完成筛选(charles e.grant,timothy l.bailey,和william stafford noble,
″
fimo:scanning for occurrences of a given motif[fimo:扫描给定基序的出现]
″
,bioinformatics[生物信息学],27(7):1017-1018,2011)。对植物基因组序列数据库的分析导致鉴定了大量与mmv as-1遗传元件具有相对高水平序列同一性的序列。进一步筛选序列库,以鉴定用于植物原位测试的推定的顺式作用调节元件。基于活性调节元件的预测特征对这些序列进行分选和选择,这些预测特征例如包括完整的bzip结合基序(例如,
‘
acgt’)和与用于测试的亲本元件具有高水平序列同一性的序列。
[0239]
获得的顺式作用调节元件如下列出:seq id no:585-seq id no:601,并在表10中提供。seq id no:585-seq id no:601的序列比对如图19所提供。将每个顺式作用调节元件工程化于调节元件(例如植物启动子)的上游,并用于产生如实例2和3中所述的标准化单子叶原生质体测试载体,其中单子叶启动子(yang,y,li,r和qi,m.2000.in vivo analysis of plant promoters and transcription factors by agroinfiltration of tobacco leaves.[通过烟草叶片的农杆菌浸润对植物启动子和转录因子进行体内分析]plant j.[植物杂志]22(6):543-551的35s最小camv启动子)被掺入构建体设计以驱动玉蜀黍原生质体中的表达。顺式作用调节元件一式三份地掺入构建体中。本领域技术人员将理解,顺式作用调节元件可以作为单体、二聚体、三聚体或以任何拷贝数被包含以驱动编码序列的稳健表达。最后,如专利申请wo 2018183878 a1的实例1和3中所述,在瞬时基因表达玉蜀黍叶肉原生质体平台中对顺式作用调节元件进行测试。完成分析以测试载体,以确定顺式作用调节元件是否能够增强下游编码序列的表达,该下游编码序列可操作地连接到嵌合顺式作用调节元件和充当嵌合调节元件的另一调节元件(例如启动子调节元件)。
[0240]
[0241][0242]
表达谱分析的结果表明,mmv as-1顺式作用调节元件(seq id no:27)及从玉蜀黍获得的变体(seq id no:585-seq id no:601)展示出作为嵌合调节元件(例如顺式作用调
节元件和启动子调节元件)内的调控元件的活性(图18)。该顺式作用调节元件驱动荧光编码序列在玉蜀黍原生质体中的稳健表达。尽管与seq id no:27相比,荧光编码序列的表达存在一定程度的变化,但许多变体序列的表达均高于camv 35s最小启动子对照。这样,首次将mmv as-1顺式作用调节元件(seq id no:27)和mmv as-1植物衍生变体顺式作用调节元件(seq id no:585-601)例示为能够在嵌合调节元件(例如启动子和顺式作用调节元件)内充当调控元件以驱动编码序列的表达的顺式作用调节元件。
[0243]
实例15:增加编码序列在玉蜀黍原生质体内表达的顺式作用调节元件的鉴定
[0244]
通过对植物基因组序列数据库(例如,水稻、大豆、卡诺拉油菜、玉蜀黍和高粱)进行筛选,获得顺式作用调节元件。获得的顺式作用调节元件如下列出:seq id no:602-seq id no:634,并在表11中提供。seq id no:602-seq id no:634的序列比对如图21所提供。将每个顺式作用调节元件工程化于调节元件(例如植物启动子)的上游,并用于产生如实例2和3中所述的标准化单子叶原生质体测试载体,其中单子叶启动子(35s最小camv启动子)被掺入构建体设计以驱动玉蜀黍原生质体中的表达。顺式作用调节元件一式三份地掺入构建体中。本领域技术人员将理解,顺式作用调节元件可以作为单体、二聚体、三聚体或以任何拷贝数被包含以驱动编码序列的稳健表达。最后,如专利申请wo 2018183878 a1的实例1和3中所述,在瞬时基因表达玉蜀黍叶肉原生质体平台中对顺式作用调节元件进行测试。完成分析以测试载体,以确定顺式作用调节元件是否能够增强下游编码序列的表达,该下游编码序列可操作地连接到嵌合顺式作用调节元件和充当嵌合调节元件的另一调节元件(例如启动子调节元件)。
[0245]
[0246]
[0247][0248]
表达谱分析的结果表明,顺式作用调节元件(seq id no:602-seq id no:634)展示出作为嵌合调节元件(例如顺式作用调节元件和启动子调节元件)内的调控元件的活性(图20)。尽管与seq id no:584相比,荧光编码序列的表达存在一定程度的变化,但大多数
transgene copy number after biolistic gene transfer and production of(secale cereale l.)plants instantly marker-free transgenic rye[在进行生物射弹基因转移后产生具有低转基因拷贝数的黑麦,并立即产生无标记的转基因黑麦(黑麦(secale cereale l.))植物],”transgenic res.[转基因研究]2003年10月;12(5):587-96.)。对于农杆菌属介导的高粱转化,参见,例如,zhao等人,“agrobacterium-mediated sorghum transformation[农杆菌属介导的高粱转化],”plant mol biol.[植物分子生物学]2000年12月;44(6):789-98。对于农杆菌属介导的大麦转化,参见,例如,tingay等人,“agrobacterium tumefaciens-mediated barley transformation[根癌农杆菌介导的大麦转化],”the plant journal[植物杂志],(1997)11:1369-1376。对于农杆菌属介导的小麦转化,参见,例如,cheng等人,“genetic transformation of wheat mediated by agrobacterium tumefaciens[通过根癌农杆菌介导的小麦基因转化],”plant physiol.[植物生理学]1997年11月;115(3):971-980。对于农杆菌属介导的烟草转化,参见例如美国专利申请公开号us 2013/0157369 a1。对于农杆菌属介导的水稻转化,参见,例如,hiei等人,“transformation of rice mediated by agrobacterium tumefaciens[通过根癌农杆菌介导的水稻转化],”plant mol.biol.[植物分子生物学]1997年9月;35(1-2):205-18。可以实现双子叶植物物种的植物转化。对于农杆菌属介导的拟南芥属转化,参见,例如,clough,s.j.和bent,a.f.(1998).floral dip:a simplified method for agrobacterium-mediated transformation ofarabidopsis thaliana[浸花法:用于农杆菌属介导的拟南芥转化的简化方法].the plant journal[植物杂志],16(6),735-743。对于农杆菌属介导的棉花转化,参见,例如,tohidfar,m.,mohammadi,m.和ghareyazie,b.(2005).agrobacterium-mediated transformation of cotton(gossypium hirsutum)using a heterologous bean chitinase gene[使用异源豆类几丁质酶基因,农杆菌属介导的棉花(陆地棉)转化].plant cell,tissue and organ culture[植物细胞、组织和器官培养],83(1),83-96。对于农杆菌属介导的大豆转化,参见例如美国专利申请公开号us 2014/0173780 a1。对于农杆菌属介导的烟草转化,参见,例如,an,g.(1985).high efficiency transformation of cultured tobacco cells[培养的烟草细胞的高效转化].plant physiology[植物生理学],79(2),568-570。
[0255]
可以通过利用先前在专利申请wo2007/053482的实例#8中描述的相同技术,用含有与嵌合调节元件(例如启动子和顺式作用调节元件)融合的顺式作用调节元件的基因表达盒转化玉米。
[0256]
可以通过利用先前在专利申请wo 2007/053482的实例#11或实例#13中描述的相同技术,用含有与嵌合调节元件(例如启动子和顺式作用调节元件)融合的顺式作用调节元件的基因表达盒转化大豆。
[0257]
可以通过利用先前在美国专利号7,838,733的实例#14或专利申请wo 2007/053482(wright等人)的实例#12中描述的相同技术,用含有与嵌合调节元件(例如启动子和顺式作用调节元件)融合的顺式作用调节元件的基因表达盒转化棉花。
[0258]
可以通过利用先前在美国专利号7,838,733的实例#26或专利申请wo 2007/053482(wright等人)的实例#22中描述的相同技术,用含有与嵌合调节元件(例如启动子和顺式作用调节元件)融合的顺式作用调节元件的基因表达盒转化卡诺拉油菜。
spp.))、桦木(桦木属物种(betula spp.))、樱桃树(李属物种(prunus spp.))、桉树(桉属物种(eucalyptus spp.))、山核桃木(山核桃属物种(carya spp.))、槭树(槭属物种(acer spp.))、栎树(栎属物种(quercus spp.))、和松树(松属物种(pinus spp.))。
[0265]
可以将含有与启动子融合的顺式作用调节元件的基因表达盒的应用部署在观赏物种和挂果物种中。此类应用也在本公开的实施例的范围内。实例包括但不限于:玫瑰(蔷薇属物种(rosa spp.))、紫果卫矛(卫矛属物种(euonymus spp.))、矮牵牛(茄科物种petunia spp.))、秋海棠(秋海棠属物种(begonia spp.))、杜鹃(杜鹃花属物种(rhododendron spp.))、红果或苹果(苹果属物种(malus spp.))、梨(梨属物种(pyrus spp.))、桃(李属物种(prunus spp.))、和万寿菊(万寿菊属物种(tagetes spp.))。
[0266]
分子分析。对获得自用含有与启动子融合的顺式作用调节元件的基因表达盒转化的植物材料的样品进行转化的植物组织的分子分析,从而确认稳定整合的与启动子融合的顺式作用调节元件的存在和拷贝数,并且定量从植物细胞中产生的目的基因表达的蛋白质的表达量。各种测定是本领域已知的,并且可以用于植物材料内与启动子融合的顺式作用调节元件的分子分析。这样,首次将顺式调节元件例示为能够在嵌合调节元件(例如嵌合启动子)内充当调控元件以驱动编码序列的表达的顺式作用调节元件。
[0267]
虽然本公开可能易于进行各种修改和替代形式,但是本文已经通过例示的方式详细描述了特定实施例。然而,应当理解,本公开并不旨在限于所公开的特定形式。而是,本公开将覆盖落入如由以下所附权利要求及其合法等同物所定义的本公开的范围内的所有修改、等同物和替代物。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。