用于检测染色体异常的高分辨率光谱染色体分带的方法
1.相关申请交叉引用
2.本技术要求于2019年12月9日提交的共同未决的美国临时专利申请第62/945,850号的权益,所述美国临时申请通过引用整体并入本文。
技术领域
3.本公开总体上涉及检测染色体中结构变异,并且更具体地涉及用于检测包含但不限于易位、扩增、缺失和倒位的潜在有害结构变异的染色体特异性组合性标记。
背景技术:
4.定向基因组杂交(dgh)是用于在单链中期染色体上绘制基因组的结构的单细胞方法。dgh技术可以促进比先前可能的范围更广范围的基因组结构变体的检测。
5.针对dgh制备染色体的一种方式是co-fish技术。二十世纪90年代开发的co-fish准许荧光探针特异性地靶向任一染色单体上的位点而不是两个染色单体上的位点。在s.m.bailey等人的“链特异性荧光原位杂交:co-fish家族(strand-specific fluorescence in situ hybridization:the co-fish family)”,《(cytogenet.genome res.)》107:11-14(2004)中,使用链特异性fish(荧光(fluorescent)或荧光(fluorescence)原位杂交)[co-fish;染色体定向-fish]研究了染色体组织,其涉及从中期(有丝分裂)染色体的dna中去除新复制的链,导致每个有丝分裂染色单体中存在一个单链靶dna,并且其中每个染色单体中的碱基序列是另一个染色单体的碱基序列的互补序列。这是可实现的,因为存在于新染色单体中的每个新复制的双螺旋体含有一个亲本dna链加新合成的链,并且正是这个新合成的链被去除,因为它在复制期间已经变得光敏。
[0006]
结构变体被广泛定义为与“正常”基因组相比,基因组片段的布置或次序的变化。简单变体包含不平衡易位、平衡易位、同源易位、倒位、重复、插入和缺失的单次发生。复杂变体包含单个细胞中的多个简单变体、与基因组物质的丧失或获得结合的简单变体、全染色体的丧失或获得以及更一般的描述为染色体碎裂的dna损伤。被定义为不同结构变体存在于基因组、相同生物体的单个细胞、细胞培养物、细胞批次中的变体异质性可以涉及简单或复杂的结构变体。当分裂的细胞自发地发展出结构变体并且不含变体的亲本细胞和含有变体的子细胞两者继续繁殖时,发生结构变体的镶嵌。
[0007]
结构变体与如单核苷酸多态性(snip)或短插入和缺失(indel)等碱基水平变化区分开。当多个双链断裂的末端被错误地重新接合或错误修复时,发生结构变体。根据携带重排的细胞的随后生殖活力,所得结构变体的结果可以限于单个细胞,影响生物体中的组织的子集,或者如果在生殖细胞中发生,那么甚至可以被遗传并影响生物体的谱系。
[0008]
每当发生dna双链断裂(dsb)时,就存在导致染色体结构变体的dna错误修复的可能性。dsb可以在正常细胞代谢过程(如复制和转录)期间内源性产生。据估计,dsb在主动代谢细胞中以每细胞周期每细胞约50的速率天然发生,并且在复制期间并且通过不依赖于复制的途径进行修复。当由高于自发速率的外源性因素通过辐射暴露、医疗干预(如用某些药
剂的化学疗法)或在基因编辑过程期间诱导时,双链断裂尤其令人关注。大多数dsb通过在整个细胞周期中起作用的非同源末端接合(nhej)进行修复。在此过程中,检测、处理断裂端并将其连接起来。这是“容易出错”的过程,因为先前存在的碱基配对序列并不总是以高保真度恢复。然而,这种重新接合过程(恢复)恢复染色体的线性连续性,并且不会导致结构异常。然而,如果两个或更多个dsb在足够近的空间和时间接近情况下发生,那么一个断裂配对的断裂末端可能与另一个断裂配对的末端错误重新接合,另外两个松散末端也可能错误重新接合,从而导致交换产生的结构异常。实例包含平衡和不平衡易位、倒位或缺失。还存在涉及同源重组(hr)的dsb修复过程,有时被称为同源定向修复(hdr)。当相同的同源序列的可用性变得可用并且非常接近时,同源定向修复(hdr)在复制后发生。hdr途径在g1或g0细胞中不起作用,其中对于hdr所必需的rad51蛋白水平非常低或不存在。然而,作为基因编辑过程的一部分(如在crispr系统中),待编辑的序列是靶向的,并且引入一个或多个dsb以使用hdr插入期望的序列。因此,每当引入dsb时,自发的或其它dsb之间的错误重新接合总是有可能形成结构异常。结构变体在很大程度上与多种人类疾病相关联,因为它们可以导致拷贝数变异并显著影响基因的功能。估计结构变体对遗传变异的贡献是snip或indel对遗传变异的贡献的10-30倍。因此,需要用于检测结构变体的方法来检测染色体畸变,并将良性遗传变异与有害遗传异常区分开。
[0009]
这些结构变体,无论其如何形成,都可以无害并且没有示出遗传毒性,可以负面地影响细胞功能,可以引起基因组不稳定性,杀死细胞,或者可以形成遗传毒性产物。非无害结构变体通过以下负面地影响细胞并且促成疾病:癌基因的形成;基因失活或敲除;调控元件中断;杂合性丢失;基因或启动子的重复;以及破坏必要的代谢途径或激活惰性代谢途径的其它机制。如果结构变异是先天性的,那么即使它不会导致任何明显的病理,由错位引起的减数交叉错误也可以在受影响的个体的后代中产生遗传异常。以典型的孟德尔方式(mendelian fashion),从父母双方遗传的隐性结构变体可能导致孩子患上在父母一方中不活跃的疾病。x-连锁结构变异选择性地影响雄性后代,因为xy对的y染色体没有补偿性正常基因。
[0010]
单独细胞中由dsb错误修复引起的两个非复发性sv的检测和鉴定,以及单独基因组中存在的sv和其在单独细胞中的表示(异质性/嵌合性)在广泛的人类疾病和病状中是临床相关且重要的。由于细胞死亡和对患者dna的风险两者的可能性,必须测量错误修复和所得结构变体。下一代和桑格测序(sanger sequencing)已尝试通过短读段和长读段全基因组测序和分析来提供此数据,但作为单一方法是不够的。为了检测结构变体,通常采用两种类型的方法,基于阵列的检测/比较基因组杂交(阵列cgh)和基于序列的计算分析。每种方法都可以通过sv检测算法测量一些产物的错误修复,并且当协同使用以交叉验证发现时,可以更有效。当这些技术测量dna碱基的序列而不是单个细胞中的基因、启动子或dna的大片段的关系或结构时,这些技术仅可以用于通过生物信息学重建来假设基因组结构。对于已知结构变体的靶向测量,基于序列的方法可以是足够的,但已证明用基于序列的方法对结构变异进行从头测量产生许多假阳性和假阴性结果,使得所述技术通常不切实际。
技术实现要素:
[0011]
因此,本公开的目的是提供用于染色体结构变体的高分辨率检测的敏感方法。本
公开的另外的目的、优势和新颖特征将部分地在随后的描述中阐明并且部分地在检查以下内容后对本领域技术人员而言变得显而易见,或可以通过所公开的方法的实践来学习。所公开的方法的目的和优势可以借助于所附权利要求书中特别指出的工具和组合实现和获得。
[0012]
以下编号段落[0012]-[00147]含有本文所公开的本发明技术特征的广泛组合的陈述:
[0013]
1.一种用于检测染色体中的至少一个结构变异的方法,所述方法包括以下步骤:
[0014]
a)由所述染色体产生一对单链姐妹染色单体,其中每条姐妹染色单体包括一个或多个靶dna序列;
[0015]
b)使一条或两条单链姐妹染色单体与两个或更多个寡核苷酸探针接触,其中所述探针中的每个探针是单链的、独特的并且与靶dna序列的至少一部分互补,并且其中所述探针中的每个探针包括至少一个标记,并且与所述靶dna序列互补的所述探针中的至少两个探针包括不同颜色的标记,使得通过所述至少两个探针与一条或两条单链姐妹染色单体的杂交模式产生一条或两条单链姐妹染色单体的光谱轮廓;
[0016]
c)检测一条或两条单链姐妹染色单体的所述光谱轮廓;
[0017]
d)将步骤(c)的所述光谱轮廓与表示对照的参考光谱轮廓进行比较;以及
[0018]
e)基于步骤(c)的任一个或两个光谱轮廓与所述参考光谱轮廓之间的至少一个光谱差异,测定所述至少一个结构变异的存在。
[0019]
2.根据方面1所述的方法,其中步骤(c)的所述光谱轮廓属于一条单链姐妹染色单体,并且所述参考光谱轮廓属于另一条单链姐妹染色单体。
[0020]
3.根据方面1或2所述的方法,其中所述结构变异选自由以下组成的组:所述染色体的片段的拷贝数的变化、所述染色体的拷贝数的变化、倒位、易位、姐妹染色单体重组、微核形成、染色体碎裂事件和其任何组合。
[0021]
4.根据方面1到3中任一项所述的方法,其中所述结构变异是所述染色体的片段的拷贝数的变化,并且所述变化选自由以下组成的组:扩增、缺失和其任何组合。
[0022]
5.根据方面1到4中任一项所述的方法,其中所述探针的长度为25到75个核苷酸。
[0023]
6.根据方面1到5中任一项所述的方法,其中所述探针的长度为30到50个核苷酸。
[0024]
7.根据方面1到6中任一项所述的方法,其中所述探针的长度为37到43个核苷酸。
[0025]
8.根据方面1到7中任一项所述的方法,其中所述探针上的所述标记是在所述探针的5'末端处缀合的荧光染料。
[0026]
9.根据方面1到8中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少两种不同颜色的标记。
[0027]
10.根据方面1到9中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少三种不同颜色的标记。
[0028]
11.根据方面1到10中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少四种不同颜色的标记。
[0029]
12.根据方面1到11中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少五种不同颜色的标记。
[0030]
13.根据方面1到12中任一项所述的方法,其中与每条单链姐妹染色单体上的所述
靶dna序列互补的所述探针包括至少六种不同颜色的标记。
[0031]
14.根据方面1到13中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少七种不同颜色的标记。
[0032]
15.根据方面1到14中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少八种不同颜色的标记。
[0033]
16.根据方面1到15中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少九种不同颜色的标记。
[0034]
17.根据方面1到16中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少十种不同颜色的标记。
[0035]
18.根据方面1到17中任一项所述的方法,其中所述至少一个标记选自由以下组成的组:在可见光光谱中可检测到的标记、在红外光光谱中可检测到的标记、在紫外光光谱中可检测到的标记和其任何组合。
[0036]
19.根据方面1到18中任一项所述的方法,其中所述至少一个标记选自由以下组成的组:所述探针的末端上的标记、所述探针的一侧上的标记、所述探针的主体上的一个或多个标记和其任何组合。
[0037]
20.根据方面1到19中任一项所述的方法,其中所述至少一个标记是所述探针的糖或亚酰胺官能团上的主体标记。
[0038]
21.根据方面1到20中任一项所述的方法,其中对所述光谱轮廓的所述检测包括使用窄带滤波器和用软件处理光谱信息。
[0039]
22.根据方面1到21中任一项所述的方法,其中对所述光谱轮廓的所述检测特异性地排除所述光谱轮廓的一个或多个光谱区。
[0040]
23.根据方面1到22中任一项所述的方法,其中步骤(e)在人工智能的帮助下执行。
[0041]
24.一种用于检测染色体中的至少一个结构变异的方法,所述方法包括以下步骤:
[0042]
a)由所述染色体产生一对单链姐妹染色单体,其中每条姐妹染色单体包括一个或多个靶dna序列;
[0043]
b)在步骤a)之后,使一条或两条单链姐妹染色单体与染色剂接触;
[0044]
c)在步骤a)之后,使一条或两条单链姐妹染色单体与两个或更多个寡核苷酸探针接触,其中所述探针中的每个探针是单链的、独特的并且与靶dna序列的至少一部分互补,并且其中所述探针中的每个探针包括至少一个标记,并且与所述靶dna序列互补的所述探针中的至少两个探针包括不同颜色的标记,使得通过所述至少两个探针与一条或两条单链姐妹染色单体的杂交模式产生一条或两条单链姐妹染色单体的光谱轮廓;
[0045]
d)检测一条或两条单链姐妹染色单体的所述光谱轮廓;
[0046]
e)检测一条或两条单链姐妹染色单体的染色模式;
[0047]
f)将步骤(d)的任一个或两个光谱轮廓与表示对照的参考光谱轮廓进行比较,并且进一步将步骤(e)的任一个或两个染色模式与表示对照的参考染色模式进行比较;以及
[0048]
g)基于步骤(d)的任一个或两个光谱轮廓与所述参考光谱轮廓之间的至少一个光谱差异并且进一步基于步骤(e)的任一个或两个染色模式与所述参考染色模式之间的至少一个染色差异,测定所述至少一个结构变异的存在。
[0049]
25.根据方面24所述的方法,其中步骤(d)的所述光谱轮廓属于一条单链姐妹染色
单体,并且所述参考光谱轮廓属于另一条单链姐妹染色单体。
[0050]
26.根据方面24或25所述的方法,其中步骤(e)的所述染色模式属于一条单链姐妹染色单体,并且所述参考染色模式属于另一条单链姐妹染色单体。
[0051]
27.根据方法24到26中任一项所述的方法,其中所述结构变异选自由以下组成的组:所述染色体的片段的拷贝数的变化、所述染色体的拷贝数的变化、插入、缺失、倒位、平衡易位、不平衡易位、姐妹染色单体重组、微核形成、染色体碎裂事件、遗传物质的损失或增益、一条或多条整个染色体的损失或增益和其任何组合。
[0052]
28.根据方面27所述的方法,其中所述结构变异是所述染色体的片段的拷贝数的变化,并且所述变化选自由以下组成的组:扩增、缺失和其任何组合。
[0053]
29.根据方面24到28中任一项所述的方法,其中所述探针的长度为25到75个核苷酸。
[0054]
30.根据方面24到29中任一项所述的方法,其中所述探针的长度为30到50个核苷酸。
[0055]
31.根据方面24到30中任一项所述的方法,其中所述探针的长度为37到43个核苷酸。
[0056]
32.根据方面24到31中任一项所述的方法,其中所述探针上的所述标记是在所述探针的5'末端处缀合的荧光染料。
[0057]
33.根据方面24到32中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少两种不同颜色的标记。
[0058]
34.根据方面24到33中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少三种不同颜色的标记。
[0059]
35.根据方面24到34中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少四种不同颜色的标记。
[0060]
36.根据方面24到35中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少五种不同颜色的标记。
[0061]
37.根据方面24到36中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少六种不同颜色的标记。
[0062]
38.根据方面24到37中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少七种不同颜色的标记。
[0063]
39.根据方面24到38中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少八种不同颜色的标记。
[0064]
40.根据方面24到39中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少九种不同颜色的标记。
[0065]
41.根据方面24到40中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少十种不同颜色的标记。
[0066]
42.根据方面24到41中任一项所述的方法,其中所述染色剂选自由以下组成的组:dapi、hoechst 33258和放线菌素d。
[0067]
43.根据方面24到42中任一项所述的方法,其中所述至少一个标记选自由以下组成的组:在可见光光谱中可检测到的标记、在红外光光谱中可检测到的标记、在紫外光光谱
中可检测到的标记和其任何组合。
[0068]
44.根据方面24到43中任一项所述的方法,其中所述至少一个标记选自由以下组成的组:所述探针的末端上的标记、所述探针的一侧上的标记、所述探针的主体中的标记和其任何组合。
[0069]
45.根据方面44所述的方法,其中所述至少一个标记是所述探针的糖或亚酰胺官能团上的主体标记。
[0070]
46.根据方面24到45中任一项所述的方法,其中对所述光谱轮廓的所述检测包括使用窄带滤波器和用软件处理光谱信息。
[0071]
47.根据方面24到46中任一项所述的方法,其中对所述光谱轮廓的所述检测特异性地排除所述光谱轮廓的一个或多个光谱区。
[0072]
48.根据方面24到47中任一项所述的方法,其中步骤(e)在人工智能的帮助下执行。
[0073]
49.一种用于检测染色体中的至少一个结构变异的方法,所述方法包括以下步骤:
[0074]
a)由所述染色体产生一对单链姐妹染色单体,其中每条姐妹染色单体包括一个或多个靶dna序列;
[0075]
b)在步骤a)之后,使一条或两条单链姐妹染色单体与和所述单链姐妹染色单体上的重复序列互补的寡核苷酸标志物接触,所述重复序列不是靶dna序列,其中所述标志物中的每种标志物包括至少一个标记;
[0076]
c)在步骤a)之后,使一条或两条单链姐妹染色单体与两个或更多个寡核苷酸探针接触,其中所述探针中的每个探针是单链的、独特的并且与靶dna序列的至少一部分互补,并且其中所述探针中的每个探针包括至少一个标记,并且与所述靶dna序列互补的所述探针中的至少两个探针包括不同颜色的标记,使得通过所述至少两个探针与一条或两条单链姐妹染色单体的杂交模式产生一条或两条单链姐妹染色单体的光谱轮廓;
[0077]
d)检测一条或两条单链姐妹染色单体的所述光谱轮廓;
[0078]
e)检测一条或两条单链姐妹染色单体的标志物杂交模式;
[0079]
f)将步骤(d)的所述光谱轮廓与表示对照的参考光谱轮廓进行比较,并且进一步将步骤(e)的所述标志物杂交模式与表示对照的参考标志物杂交模式进行比较;以及
[0080]
g)基于步骤(d)的任一个或两个光谱轮廓与所述参考光谱轮廓之间的至少一个光谱差异并且进一步基于步骤(e)的任一个或两个标志物杂交模式与所述参考标志物杂交模式之间的至少一个标志物杂交模式差异,测定所述至少一个结构变异的存在。
[0081]
50.根据方面49所述的方法,其中步骤(d)的所述光谱轮廓属于一条单链姐妹染色单体,并且所述参考光谱轮廓属于另一条单链姐妹染色单体。
[0082]
51.根据方面49或50所述的方法,其中步骤(e)的所述标志物杂交模式属于一条单链姐妹染色单体,并且所述参考标志物杂交模式属于另一条单链姐妹染色单体。
[0083]
52.根据方面49到51中任一项所述的方法,其中所述结构变异选自由以下组成的组:所述染色体的片段的拷贝数的变化、所述染色体的拷贝数的变化、倒位、易位、姐妹染色单体重组、微核形成、染色体碎裂事件和其任何组合。
[0084]
53.根据方面49到52中任一项所述的方法,其中所述结构变异是所述染色体的片段的拷贝数的变化,并且所述变化选自由以下组成的组:扩增、缺失和其任何组合。
[0085]
54.根据方面49到53中任一项所述的方法,其中所述探针的长度为25到75个核苷酸。
[0086]
55.根据方面49到54中任一项所述的方法,其中所述探针的长度为30到50个核苷酸。
[0087]
56.根据方面49到55中任一项所述的方法,其中所述探针的长度为37到43个核苷酸。
[0088]
57.根据方面49到56中任一项所述的方法,其中所述探针上的所述标记是在所述探针的5'末端处缀合的荧光染料。
[0089]
58.根据方面49到57中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少两种不同颜色的标记。
[0090]
59.根据方面49到58中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少三种不同颜色的标记。
[0091]
60.根据方面49到59中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少四种不同颜色的标记。
[0092]
61.根据方面49到60中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少五种不同颜色的标记。
[0093]
62.根据方面49到61中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少六种不同颜色的标记。
[0094]
63.根据方面49到62中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少七种不同颜色的标记。
[0095]
64.根据方面49到63中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少八种不同颜色的标记。
[0096]
65.根据方面49到64中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少九种不同颜色的标记。
[0097]
66.根据方面49到65中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少十种不同颜色的标记。
[0098]
67.根据方面49到66中任一项所述的方法,其中所述至少一个标记选自由以下组成的组:在可见光光谱中可检测到的标记、在红外光光谱中可检测到的标记、在紫外光光谱中可检测到的标记和其任何组合。
[0099]
68.根据方面49到67中任一项所述的方法,其中所述至少一个标记选自由以下组成的组:所述探针的末端上的标记、所述探针的一侧上的标记、所述探针的主体中的标记和其任何组合。
[0100]
69.根据方面68所述的方法,其中所述至少一个标记是所述探针的糖或亚酰胺官能团上的主体标记。
[0101]
70.根据方面49到69中任一项所述的方法,其中对所述光谱轮廓的所述检测包括使用窄带滤波器和用软件处理光谱信息。
[0102]
71.根据方面49到70中任一项所述的方法,其中对所述光谱轮廓的所述检测特异性地排除所述光谱轮廓的一个或多个光谱区。
[0103]
72.根据方面49到71中任一项所述的方法,其中步骤(e)在人工智能的帮助下执
行。
[0104]
73.一种用于检测染色体中的至少一个结构变异的计算机实施的方法,所述方法包括以下步骤:
[0105]
a)由所述染色体产生一对单链姐妹染色单体,其中每条姐妹染色单体包括一个或多个靶dna序列;
[0106]
b)使一条或两条单链姐妹染色单体与两个或更多个寡核苷酸探针接触,其中所述探针中的每个探针是单链的、独特的并且与靶dna序列的至少一部分互补,并且其中所述探针中的每个探针包括至少一个标记,并且与所述靶dna序列互补的所述探针中的至少两个探针包括不同颜色的标记,使得通过所述至少两个探针与一条或两条单链姐妹染色单体的杂交模式产生一条或两条单链姐妹染色单体的光谱轮廓;
[0107]
c)检测一条或两条单链姐妹染色单体的所述光谱轮廓;
[0108]
d)将步骤(c)的所述光谱轮廓与表示对照的参考光谱轮廓进行比较;以及
[0109]
e)基于步骤(c)的任一个或两个光谱轮廓与所述参考光谱轮廓之间的至少一个光谱差异,测定所述至少一个结构变异的存在,其中步骤(d)和(e)是用计算机系统计算的。
[0110]
74.一种可由计算机读取的程序存储装置,所述程序存储装置在用于检测染色体中的至少一个结构变异的方法中具体地体现可由所述计算机执行以执行所述步骤(d)和(e)的指令的程序,所述方法包括以下步骤:
[0111]
a)由所述染色体产生一对单链姐妹染色单体,其中每条姐妹染色单体包括一个或多个靶dna序列;
[0112]
b)使一条或两条单链姐妹染色单体与两个或更多个寡核苷酸探针接触,其中所述探针中的每个探针是单链的、独特的并且与靶dna序列的至少一部分互补,并且其中所述探针中的每个探针包括至少一个标记,并且与所述靶dna序列互补的所述探针中的至少两个探针包括不同颜色的标记,使得通过所述至少两个探针与一条或两条单链姐妹染色单体的杂交模式产生一条或两条单链姐妹染色单体的光谱轮廓;
[0113]
c)检测一条或两条单链姐妹染色单体的所述光谱轮廓;
[0114]
d)将步骤(c)的所述光谱轮廓与表示对照的参考光谱轮廓进行比较;以及
[0115]
e)基于步骤(c)的任一个或两个光谱轮廓与所述参考光谱轮廓之间的至少一个光谱差异,测定所述至少一个结构变异的存在。
[0116]
75.一种用于检测染色体中的至少一个结构变异的方法,所述方法包括以下步骤:
[0117]
a)由所述染色体产生一对单链姐妹染色单体,其中姐妹染色单体包括一个或多个靶dna序列;
[0118]
b)使单链姐妹染色单体与两个或更多个寡核苷酸探针接触,其中所述探针中的每个探针是单链的、独特的并且与靶dna序列的至少一部分互补,并且其中所述探针中的每个探针包括至少一个标记,并且所述探针中的至少两个探针包括不同颜色的标记,使得通过所述至少两个探针与所述单链姐妹染色单体的杂交模式产生所述单链姐妹染色单体的光谱轮廓;
[0119]
c)检测所述单链姐妹染色单体的所述光谱轮廓;
[0120]
d)将步骤(c)的所述光谱轮廓与表示对照的参考光谱轮廓进行比较;以及
[0121]
e)基于步骤(c)的所述光谱轮廓与所述参考光谱轮廓之间的至少一个光谱差异,
测定所述至少一个结构变异的存在。
[0122]
76.根据方面75所述的方法,其中参考光谱轮廓属于另一条单链姐妹染色单体。
[0123]
77.根据方面75或76所述的方法,其中所述结构变异选自由以下组成的组:所述染色体的片段的拷贝数的变化、所述染色体的拷贝数的变化、倒位、易位、姐妹染色单体重组、微核形成、染色体碎裂事件和其任何组合。
[0124]
78.根据方面75到77中任一项所述的方法,其中所述结构变异是所述染色体的片段的拷贝数的变化,并且所述变化选自由以下组成的组:扩增、缺失和其任何组合。
[0125]
79.根据方面75到78中任一项所述的方法,其中所述探针的长度为25到75个核苷酸。
[0126]
80.根据方面75到79中任一项所述的方法,其中所述探针的长度为30到50个核苷酸。
[0127]
81.根据方面75到80中任一项所述的方法,其中所述探针的长度为37到43个核苷酸。
[0128]
82.根据方面75到81中任一项所述的方法,其中所述探针上的所述标记是在所述探针的5'末端处缀合的荧光染料。
[0129]
83.根据方面75到82中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少两种不同颜色的标记。
[0130]
84.根据方面76到83中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少三种不同颜色的标记。
[0131]
85.根据方面76到84中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少四种不同颜色的标记。
[0132]
86.根据方面76到85中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少五种不同颜色的标记。
[0133]
87.根据方面76到86中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少六种不同颜色的标记。
[0134]
88.根据方面76到87中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少七种不同颜色的标记。
[0135]
89.根据方面76到88中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少八种不同颜色的标记。
[0136]
90.根据方面76到89中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少九种不同颜色的标记。
[0137]
91.根据方面76到90中任一项所述的方法,其中与每条单链姐妹染色单体上的所述靶dna序列互补的所述探针包括至少十种不同颜色的标记。
[0138]
92.根据方面76到91中任一项所述的方法,其中步骤(d)和(e)是用计算机系统计算的。
[0139]
93.根据方面76到92中任一项所述的方法,其中所述至少一个标记选自由以下组成的组:在可见光光谱中可检测到的标记、在红外光光谱中可检测到的标记、在紫外光光谱中可检测到的标记和其任何组合。
[0140]
94.根据方面76到93中任一项所述的方法,其中所述至少一个标记选自由以下组
成的组:所述探针的末端上的标记、所述探针的一侧上的标记、所述探针的主体中的标记和其任何组合。
[0141]
95.根据方面94所述的方法,其中所述至少一个标记是所述探针的糖或亚酰胺官能团上的主体标记。
[0142]
96.根据方面76到95中任一项所述的方法,其中对所述光谱轮廓的所述检测包括使用窄带滤波器和用软件处理光谱信息。
[0143]
97.根据方面76到96中任一项所述的方法,其中对所述光谱轮廓的所述检测特异性地排除所述光谱轮廓的一个或多个光谱区。
[0144]
98.根据方面76到97中任一项所述的方法,其中步骤(e)在人工智能的帮助下执行。
[0145]
99.根据方面76到98中任一项所述的方法,其进一步包括:在步骤a)之后,使所述单链姐妹染色单体与染色剂接触;检测所述姐妹染色单体的染色模式;将所述染色模式与表示对照的参考染色模式进行比较;以及部分地基于所述姐妹染色单体的所述染色模式与所述参考染色模式之间的至少一个染色差异来测定所述至少一个结构变异的存在。
[0146]
100.根据方面76到99中任一项所述的方法,其进一步包括:在步骤a)之后,使所述单链姐妹染色单体与和所述单链姐妹染色单体上的重复序列互补的寡核苷酸标志物接触,所述重复序列不是靶dna序列,其中所述标志物中的每种标志物包括至少一个标记;检测所述姐妹染色单体的标志物杂交模式;将所述标志物杂交模式与表示对照的参考标志物杂交模式进行比较;以及部分地基于所述姐妹染色单体的所述标志物杂交模式与所述参考标志物杂交模式之间的至少一个标志物杂交模式差异来测定所述至少一个结构变异的存在。
[0147]
101.根据方面1、24、49、73、74、75、99和100中任一项所述的方法,其中所述参考光谱轮廓缺少所述至少一个结构变异。
[0148]
102.根据方面1、24、49、73、74、75、99和100中任一项所述的方法,其中所述参考光谱轮廓包括所述至少一个结构变异。
[0149]
103.根据方面1、24、49、73、74、75、99和100中任一项所述的方法,其中所述参考光谱轮廓包括标记的探针的有意分布。
[0150]
104.根据方面24或99所述的方法,其中所述参考染色模式缺少所述至少一个结构变异。
[0151]
105.根据方面24或99所述的方法,其中所述参考染色模式包括所述至少一个结构变异。
[0152]
106.根据方面49或100所述的方法,其中所述参考标志物杂交模式缺少所述至少一个结构变异。
[0153]
107.根据方面49或100所述的方法,其中所述参考标志物杂交模式包括所述至少一个结构变异。
[0154]
108.根据方面49或100所述的方法,其中所述参考标志物杂交模式包括标记的探针的有意分布。
[0155]
109.一种鉴定受试dna链的一个或多个结构特征的方法,在处理器中实施的所述方法包括:
[0156]
a)接收表示所述受试dna链上的至少一个碱基对序列的光谱轮廓,所述光谱轮廓
包含对应于所述受试dna链的所述碱基序列的频率数据,所述频率数据包含至少两个颜色通道;
[0157]
b)将所述光谱轮廓转换为所述受试dna链的数据表,所述数据表包括所述碱基序列的所述至少两个颜色通道的定位数据和强度数据;以及
[0158]
c)将所述受试dna链的所述数据表与参考特征查找表进行比较以鉴定所述受试dna链的一个或多个正常和/或异常特征,所述参考特征查找表包括表示对应的对照dna链的正常和/或异常特征的一个或多个特征节点,其中所述一个或多个特征节点中的每个特征节点由表示所述对照dna链的碱基子序列的色带限定,所述碱基子序列在起始碱基处开始并且在末端碱基处结束。
[0159]
110.根据方面109所述的方法,其中所述接收所述光谱轮廓包括:
[0160]
a)从染色体产生一对单链姐妹染色单体,其中所述受试dna链包含单链姐妹染色单体的至少一部分,并且所述受试dna链包括一个或多个靶dna序列;
[0161]
b)使一条或两条单链姐妹染色单体与两个或更多个寡核苷酸探针接触,其中所述探针中的每个探针是单链的、独特的并且与靶dna序列的至少一部分互补,并且其中所述探针中的每个探针包括至少一个标记,并且与所述靶dna序列互补的所述探针中的至少两个探针包括对应于所述至少两个颜色通道的不同颜色的标记,使得通过所述至少两个探针与一条或两条单链姐妹染色单体的杂交模式产生一条或两条单链姐妹染色单体的光谱轮廓,由此产生所述受试dna链上的碱基序列的光谱轮廓;以及
[0162]
c)检测所述受试dna链上的所述碱基序列的所述光谱轮廓。
[0163]
111.根据方面109或110所述的方法,其中转换所述光谱轮廓包含将所述光谱轮廓分割成多个区,所述区中的每个区具有对应于所述至少两个颜色通道中的一个颜色通道的颜色。
[0164]
112.根据方面111所述的方法,其中所述区中的每个区由位置和大小参数限定。
[0165]
113.根据方面109到112中任一项所述的方法,其中所述转换是通过机器学习/ai算法执行的。
[0166]
114.根据方面109到113中任一项所述的方法,其中每个特征节点表示遗传元件、结构变异或其组合的至少一部分。
[0167]
115.根据方面114所述的方法,其中所述遗传元件选自由以下组成的组:蛋白质编码区、影响转录的区、影响翻译的区、影响翻译后修饰的区和其任何组合。
[0168]
116.根据方面114所述的方法,其中所述遗传元件选自由以下组成的组:外显子、内含子、5'非翻译区、3'非翻译区、启动子、增强子、沉默子、操纵子、终止子、poly-a尾部、反向末端重复序列、mrna稳定性元件和其任何组合。
[0169]
117.根据方面114所述的方法,其中所述结构变异选自由以下组成的组:所述染色体的片段的拷贝数的变化、所述染色体的拷贝数的变化、倒位、易位、姐妹染色单体重组、微核形成、染色体碎裂事件和其任何组合。
[0170]
118.根据方面109到117中任一项所述的方法,其中所述比较是针对多个特征查找表中的每个特征查找表执行的。
[0171]
119.根据方面118所述的方法,其中所述多个特征查找表中的每个特征查找表对应于不同的所关注遗传元件。
[0172]
120.根据方面109到119中任一项所述的方法,其中所述比较是通过机器学习/ai算法执行的。
[0173]
121.一种处理表示受试dna链的数据的方法,在处理器上实施的所述方法包括:
[0174]
a)接收表示受试dna链上的至少一个碱基序列的光谱轮廓,所述光谱轮廓包含对应于所述受试dna链上的所述碱基序列的频率数据,所述频率数据包含至少两个颜色通道;
[0175]
b)将所述光谱轮廓转换为所述受试dna链的数据表,所述数据表包括所述碱基序列的所述至少两个颜色通道的定位数据和强度数据;以及
[0176]
c)将所述数据表存储到存储器。
[0177]
122.根据方面121所述的方法,其进一步包括:
[0178]
通过将表示所述受试dna链的所述数据表与参考特征查找表进行比较以鉴定所述受试dna链的一个或多个正常和/或异常特征来测定所述受试dna链中的一个或多个正常和/或异常特征,所述参考特征查找表包括表示对应的对照dna链的正常和/或异常特征的一个或多个特征节点,其中所述一个或多个特征节点中的每个特征节点由色带和在起始碱基处开始并且在末端碱基处结束的碱基子序列限定。
[0179]
123.根据方面121或122所述的方法,其进一步包括将所述受试dna链的碱基水平数据合并到所述数据表中。
[0180]
124.根据方面121到123中任一项所述的方法,其进一步包括限定表示对照dna链的正常和/或异常特征的一个或多个特征节点,其中所述一个或多个特征节点中的每个特征节点由色带和在起始碱基处开始并且在末端碱基处结束的碱基子序列限定。
[0181]
125.根据方面124所述的方法,其中所述一个或多个特征节点由经训练的机器学习算法限定。
[0182]
126.根据方面124所述的方法,其进一步包括对多个对照dna链中的每个对照dna链执行所述接收和转换的步骤,并且将多个所得数据表存储到所述存储器。
[0183]
127.根据方面126所述的方法,其中所述多个对照dna链来源于同一基因组区并且从不同患者的样品中获得。
[0184]
128.根据方面126所述的方法,其进一步包括接收关于所述一个或多个限定的特征节点中的特定特征节点的查询,并且使用所述多个所得数据表处理所述查询。
[0185]
129.一种用于鉴定染色体外dna(ecdna)的染色体来源的方法,所述方法包括以下步骤:
[0186]
a)使来自细胞的所述ecdna与两个或更多个寡核苷酸探针接触,其中所述探针中的每个探针是单链的、独特的并且与所述ecdna的至少一部分互补,其中所述探针中的每个探针包括至少一个标记;
[0187]
b)使来自相同细胞的至少一条染色体或染色体的至少一条单链姐妹染色单体与步骤(a)的相同探针接触;
[0188]
c)检测所述ecdna的光谱轮廓,并且检测所述至少一条染色体或染色体的至少一条单链姐妹染色单体的光谱轮廓;
[0189]
d)比较步骤(c)的所述光谱轮廓;以及
[0190]
e)基于所述ecdna的所述光谱轮廓与所述至少一条染色体或染色体的至少一条单链姐妹染色单体的所述光谱轮廓之间的至少一种相似性,鉴定所述至少一条染色体或染色
体的至少一条单链姐妹染色单体作为所述ecdna中的dna来源。
[0191]
130.根据方面129所述的方法,其进一步包括基于步骤d)的所述比较,鉴定所述ecdna中的dna所来源于的所述至少一条染色体或染色体的至少一条单链姐妹染色单体上的定位。
[0192]
131.根据方面130所述的方法,其中所述ecdna来源于所述至少一条染色体或染色体的至少一条单链姐妹染色单体是由所述定位处的dna扩增引起的。
[0193]
132.根据方面130所述的方法,其中在所述ecdna上鉴定出至少一种癌基因。
[0194]
133.根据方面129所述的方法,其中所述ecdna选自由以下组成的组:游离型dna和掺入载体的dna。
[0195]
134.根据方面1到72、75到108和129到133中任一项所述的方法,其中至少一条染色体或染色体的至少一条单链姐妹染色单体上的至少一个靶区域被鉴定用于靶标富集,并且至少一条染色体或染色体的至少一条单链姐妹染色单体与靶标富集探针接触。
[0196]
135.根据方面1到72、75到108和129到134中任一项所述的方法,其中所述光谱轮廓的所述比较包括对相同染色体或相同单链姐妹染色单体上的至少一个带在至少一个其它带上的渗色(bleeding)的光谱分析。
[0197]
136.根据方面1到72、75到108和129到135中任一项所述的方法,其中所述至少一条染色体或染色体的至少一条单链姐妹染色单体与两个或更多个寡核苷酸探针的接触包括将包括所述至少一条染色体或染色体的至少一条单链姐妹染色单体的样品包埋在可溶胀的水凝胶中并且将所述样品与所述水凝胶化学地连接,进一步地其中所述水凝胶溶胀以增加跨x、y和z轴的空间分辨率。
附图说明
[0198]
图1展示了比较分带dgh涂料与单色dgh涂料的染色体内重排的实例。1a:正常染色体2,制备用于dgh,使用具有多色带的染色体2dgh涂料杂交。1b:具有缺失的染色体2,鉴定出带缺失。1c:具有扩增的染色体2,鉴定出具有额外带的区。1d:具有姐妹染色单体重组事件(仅对1个复制周期-完美修复事件可见)的染色体2,由于带顺序正确(未反向)而被鉴定为scr。1e:具有倒位事件的染色体2,通过带的反向顺序鉴定。2a:正常染色体2,制备用于dgh,使用单色染色体2dgh涂料杂交。2b:具有缺失的染色体2,区未知。2c:具有扩增的染色体2,扩增区未知。2d:具有scr或倒位事件的染色体2,特异性变体未知。(scr可能被遗漏,标记为倒位,因为在相对的姐妹染色单体上看到的片段的取向未知。)2e:具有scr或倒位事件的染色体2,特异性变体未知。(倒位可能被遗漏,标记为scr,因为在相对的姐妹染色单体上看到的片段的取向未知。)
[0199]
图2展示了染色体间重排(两条不同染色体之间的易位)的实例,分带dgh涂料对单色dgh涂料。1a:正常染色体2,制备用于dgh,使用具有多色带的染色体2dgh涂料杂交。1b:正常染色体4,出于展示目的未涂染。1c:衍生的染色体a(相互易位的产物),其中来自染色体2的材料(带1-11)与来自染色体4的材料(未涂染)融合。1d:衍生的染色体b(相互易位的其它产物),其中来自染色体2的材料(带12-19)与来自染色体4(未涂染)的材料融合。2a:正常染色体2,制备用于dgh,使用单色染色体2dgh涂料杂交。2b:正常染色体4,出于展示目的未涂染。2c:衍生的染色体a(相互易位的产物),其中来自染色体2的材料与来自染色体4(未涂
染)的材料融合-融合坐标未知。2d:衍生的染色体b(相互易位的其它产物),其中来自染色体2的材料与来自染色体4(未涂染)的材料融合-融合坐标未知。
[0200]
图3展示了染色体间等位基因重排(同一染色体的两个同源物之间的易位)的实例。分带dgh涂料对单色dgh涂料。1a:正常染色体2同源物1,制备用于dgh,使用具有多色带的染色体2dgh涂料杂交。1b:正常染色体2同源物2,制备用于dgh,使用具有多色带的染色体2dgh涂料杂交。1c:衍生的染色体a(同源物之间相互易位的产物),其中来自染色体2同源物1的材料在相同断点(带11与12之间)处与来自染色体2同源物2的材料交换。两个sce在每个同源物上完全相同位置的统计学可能性非常小,与之相对,很可能发生等位基因易位事件-尤其是在单个位置处被编辑的细胞中(两个dsb-每个同源物一个)。1d:衍生的染色体b(同源物之间相互易位的产物),其中来自染色体2同源物1的材料在相同断点(带11与12之间)处与来自染色体2同源物2的材料交换。两个sce在每个同源物上完全相同位置的统计学可能性非常小,与之相对,很可能发生等位基因易位事件-尤其是在单个位置处被编辑的细胞中(两个dsb-每个同源物一个)。2a:正常染色体2同源物1,制备用于dgh,使用单色染色体2dgh涂料杂交。2b:正常染色体2同源物2,制备用于dgh,使用单色染色体2dgh涂料杂交。2c:衍生的染色体a(同源物之间相互易位的产物),其中来自染色体2同源物1的材料在未知断点处与来自染色体2同源物2的材料交换。两个sce在每个同源物上完全相同位置的统计学可能性非常小,与之相对,很可能发生等位基因易位事件-尤其是在单个位置处被编辑的细胞中(两个dsb-每个同源物一个),但由于缺乏基因组坐标特异性,无法用单色涂料确认。2d:衍生的染色体b(同源物之间相互易位的产物),其中来自染色体2同源物1的材料在未知断点处与来自染色体2同源物2的材料交换。两个sce在每个同源物上完全相同位置的统计学可能性非常小,与之相对,很可能发生等位基因易位事件-尤其是在单个位置处被编辑的细胞中(两个dsb-每个同源物一个),但由于缺乏基因组坐标特异性,无法用单色涂料确认。
[0201]
图4展示了复杂染色体重排的实例。在第一个图像中,示出了来自血液衍生淋巴细胞的两个染色体2同源物,所述血液衍生淋巴细胞最近暴露于电离辐射以用于前列腺癌治疗。复杂结构变异存在于右侧同源物上,其可以在使用表1中描述的分带dgh涂料杂交之后可视化。在图像之后提供的图形展示了在使用多色分带dgh涂料对单色dgh涂料的情况下,这种复杂重排将如何呈现。1a:正常染色体2,制备用于dgh,使用具有多色带的染色体2dgh涂料杂交。1b:具有复杂结构重排的染色体2,使用具有多色带的染色体2dgh涂料杂交。存在大的臂间倒位,其中一个断点出现在2p上带1与2之间,并且另一个断点在2q上平分带18。在2q上的着丝粒附近存在另外的较小臂内倒位,其中第一个断点在带9与10之间,并且第二个断点在带10与11之间。还存在带9与11之间与小的臂内倒位共享相同的近端断点的大的姐妹染色单体交换事件,所述姐妹染色单体交换事件也可以用带的顺序来验证,所述带仍然以正确的数字顺序出现,但现在位于与主要涂料(右姐妹染色单体)相反的姐妹染色单体(左姐妹染色单体)上。2a:正常染色体2,制备用于dgh,使用单色染色体2dgh涂料杂交。2b:具有复杂结构重排的染色体2,使用单色染色体2dgh涂料杂交。如果没有色带来提供片段的顺序,那么无法在坐标中鉴定或描述重排。实际上,染色体似乎具有小末端sce或倒位(p臂)和大倒位(q臂),并且存在的结构重排的真正分类将被错误鉴定。
[0202]
图5展示了用于sv检测的靶向探针dgh测定的实例。1a:正常染色体2,制备用于dgh,与围绕所关注基因座的4个靶向探针杂交。1b:具有部分所关注基因座缺失的染色体2
(跨越由靶向探针2覆盖的基因组坐标)。1c:具有姐妹染色单体重组事件的染色体2,在与靶向探针1和4相对的姐妹染色单体上看到靶向探针2和3,其中维持探针的顺序-从端粒到着丝粒为1、2、3、4。1d:具有倒位事件的染色体2,在与靶向探针1和4相对的姐妹染色单体上看到靶向探针2和3,其中探针2和3的顺序颠倒。探针以从端粒到着丝粒为1、3、2、4的顺序出现。
[0203]
图6展示了来自为dgh制备的辐射暴露的血液衍生淋巴细胞样品的重排细胞中单色dgh涂料标记的染色体1、2和3的示例图像。
[0204]
图7的图像a和b示出了来自两个单独的正常中期细胞的染色体2同源物对,不存在结构变异(正常永生化人成纤维细胞系bj-5ta)。图7的图像c和d示出了来自2个单独的中期细胞(正常永生化人成纤维细胞系bj-5ta)的染色体2同源物对,示出姐妹染色单体交换产生的一个同源物中的结构变异(维持了颜色的顺序,但信号存在于相对的姐妹染色单体上)。
[0205]
图8a示出了正常染色体2的杂交、探针分布和荧光波长强度。图8b示出了正常染色体2中检测到的sce的杂交、探针分布和荧光波长强度。
[0206]
图9展示了与染色体杂交的3个单独的序列梯测定。一个序列梯测量关于对每个信号有贡献的寡核苷酸数量的检测极限,在染色体2的p臂上间隔约20mb(在图像中标记为序列梯1)。第二序列梯(染色体2q)评估靶标大小,固定量的寡核苷酸可以分散在上面,也间隔约20mb,并且还测量检测极限(在图像中标记为序列梯2)。第三序列梯(在下方看到的与染色体1q杂交),具有间隔很近的以及间隔很远的探针,从而允许评估在任何给定中期传播中紧邻的两个点的可分辨性(在图像中标记为序列梯3)。
具体实施方式
[0207]
除非另有说明,否则技术术语均根据常规用法使用。分子生物学中常用术语的定义可以在benjamin lewin,《基因v(genes v)》,牛津大学出版社(oxford university press)出版,1994(isbn 0-19-854287-9);kendrew等人(编辑),《分子生物学百科全书(the encyclopedia of molecular biology)》,布莱克威尔科学有限责任公司(blackwell science ltd.)出版,1994(isbn 0-632-02182-9);以及robert a.meyers(编辑),《分子生物学与生物技术:综合案头参考(molecular biology and biotechnology:a comprehensive desk reference)》,vch出版社有限公司(vch publishers,inc.)出版,1995(isbn 1-56081-569-8)中找到。除非另有解释,否则本文所用的所有技术和科学术语都具有与本公开所属领域的普通技术人员通常所理解的含义相同的含义。除非上下文另外明确指出,否则单数术语“一个/一种(a/an)”和“所述”包含复数指示物。“包括a或b”意指包含a或b或a和b。应进一步理解,针对核酸或多肽给出的所有碱基大小或氨基酸大小以及所有分子量或分子质量值都是近似的并且是为了描述而提供的。
[0208]
进一步地,本文提供的范围被理解为所述范围内所有值的简写。例如,1到50的范围应被理解为包含来自由以下组成的组的任何数字、数字组合或子范围:1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49或50(以及其分数,除非上下文另有明确规定)。除非另有说明,否则任何浓度范围、百分比范围、比率范围或整数范围应被理解
为包含所阐述范围内的任何整数的值,并且在适当时包含其分数(如整数的十分之一和百分之一)。而且,本文中所阐述的与如聚合物亚基、大小或厚度等任何物理特征相关的任何数目范围应被理解为包含所陈述的范围内的任何整数,除非另外指明。如本文所使用的,“约”或“基本上由...组成”意指所指定范围、值或结构的
±
20%,除非另有指示。如本文所使用的,术语“包含”和“包括”是开放式的并且同义地使用。
[0209]
尽管类似或等效于本文所描述的方法和材料的方法和材料可以用于实践或测试本公开,但是下面描述了合适的方法和材料。本文所提及的所有出版物、专利申请、专利和其它参考文献均通过引用整体并入本文。在冲突的情况下,应以本说明书(包含术语的解释)为准。另外,所述材料、方法和实例仅是说明性的并且不意在是限制性的。
[0210]
如本文所使用的,“带”是指与标记有类似光发射特征(例如,相同颜色的探针)的探针杂交的染色体区。
[0211]
如本文所使用的,“渗色”是指在至少一个其它带上部分地重叠或以其它方式部分地出现的一个带的光发射特征。
[0212]
如本文使用的,“颜色”是指可以被检测为单独的以及与其它波长不同的光发射波长。
[0213]
如本文使用的,“染色体片段”是指由基因组中的开始和结束坐标(例如,人染色体2中的bp 12900-14900)或已知序列含量(例如,基因或移动元件的序列)限定的dna区。染色体片段可以像两个碱基对一样小,或者像整个染色体一样大。
[0214]
如本文所使用的,“颜色通道”是指光谱的区,包含可见光、红外光和紫外光。颜色通道可以被指定为对实践本文所公开的方法的个体有用的宽的一组波长或窄的一组波长。
[0215]
如本文所使用的,“定向基因组杂交”或“dgh”是指与探针杂交方法组合的样品制备方法,其中(1)dna类似物(brdu)被提供给活跃分裂的细胞进行一个复制周期,并且选择性地并入到新合成的子链中;(2)制备中期扩散;(3)以光解方式靶向所并入的类似物以实现dna切口,所述dna切口选择性地用于酶促地消化和降解新合成的链;(4)单链中期扩散与针对参考基因组的独特序列设计的单向探针原位杂交,使得中期染色体的仅一个单链姐妹染色单体在独特的一个或多个靶位点处被标记。
[0216]
如本文所使用的,“游离基因”或“游离型dna”是指可以在细胞的细胞质中存在和自主复制的dna片段。
[0217]
如本文所使用的,“染色体外dna”或“ecdna”是指在染色体外发现的任何dna,无论是在细胞的细胞核内部还是外部https://en.wikipedia.org/wiki/dnahttps://en.wikipedia.org/wiki/chromosomehttp s://en.wikipedia.org/wiki/cell_nucleushttps://en.wikipedia.org/wiki/cell_(biolog y)。在某些方面,ecdna可以是有害的,并且可以携带扩增的癌基因。在一些方面,有害的ecdna可以是健康体组织中发现的千碱基大小的环状dna的100-1,000倍。在某些方面,ecdna包含游离型dna和载体并入的dna。
[0218]
如本文所使用的,“特征节点”和“节点”可互换地使用以指表示寡核苷酸或多核苷酸链上的任何分析关注区的数值,包含数值组。节点可以是特定基因座、基因座串、基因、多个基因、带或整个染色体。节点可以是可配置的并且大小可变的,以在分析期间允许不同级别的粒度。作为非限制性实例,节点可以表示受试dna链的正常特征或异常特征。同样,通过
非限制性实例,节点可以提供从标记的探针杂交到对照dna链的光谱轮廓数据的数字值,其中节点表示对照dna链的正常结构特征或异常结构特征。
[0219]
如本文所使用的,“特征查找表”是指表示一个或多个特征节点的数字值的表。
[0220]
如本文所使用的,“探针”是指被设计成与所关注靶dna序列互补的标记的寡核苷酸,使得当与杂交反应组合时,所述探针将与靶标结合并检测所述靶标。
[0221]
如本文所使用的,“单链染色单体”是指其中dna类似物(例如,brdu)被提供给主动分裂细胞用于单个复制周期的过程的产物,所述产物然后选择性地并入到新合成的子链中,制备中期扩散,以光解方式靶向所述并入的类似物以实现dna切口,所述dna切口用于选择性地酶促消化和降解新合成的链,从而产生单链产物。如果使用术语沃森(watson)和克里克(crick)来描述双链dna复合物的5'到3'链和3'到5'链,那么未处理的中期染色体将具有一个带有亲本沃森/子克里克的姐妹染色单体,一个带有子沃森/亲本克里克的姐妹染色单体。在根据上述方法制备的染色体中,一个姐妹染色单体将仅由亲本沃森链组成,并且另一个姐妹染色单体将仅由亲本克里克链组成。
[0222]
如本文所使用的,“姐妹染色单体交换”或“sce”是指精确匹配的和相同的dna链的无误差交换(交叉)。姐妹染色单体交换虽然不是结构变体,但与基因组不稳定性的升高速率相关联,因为替代性模板位点(如与断裂位点相邻的重复元件)产生不等交换的可能性增加,从而产生结构变体。
[0223]
如本文所使用的,“姐妹染色单体重组”或“scr”是指涉及相同的姐妹染色单体的同源重组过程,其产生单向非交叉事件,另外被称为基因转换事件。当被称为双霍利迪连接体(holliday junction)的同源重组中间体以使得其导致非交叉的方式被解析时,即认为发生scr。scr可以被细胞采用以解析单链dna病变(其涉及对应的复制叉塌缩)和双链断裂。姐妹染色单体之间的基因转换通常不与相互交换相关联,并且因此与sce区分开。
[0224]
如本文所使用的,“光谱轮廓”是指一种或多种材料在一个或多个波长处的光强度的变化的图形表示。
[0225]
如本文所使用的,“结构特征”广义地是指寡核苷酸或多核苷酸内的碱基序列的任何方面,包含序列的正常特征或异常特征。例如,结构特征包含但不限于选自以下的遗传元件:蛋白质编码区、影响转录的区、影响翻译的区、影响翻译后修饰的区和其任何组合。作为进一步的非限制性实例,结构特征包含选自以下的遗传元件:外显子、内含子、5'非翻译区、3'非翻译区、启动子、增强子、沉默子、操纵子、终止子、poly-a尾部、反向末端重复序列、mrna稳定性元件和其任何组合。
[0226]
如本文所使用的,“训练的”或“训练”是指创建模型,所述模型在训练数据上进行训练,并且然后可以用于处理另外的数据。可以用于训练的模型类型包含但不限于:人工神经网络、决策树、支持向量机、回归分析、贝叶斯网络(bayesian network)和遗传算法。
[0227]
如本文所使用的,“结构变体”或“染色体结构变体”或“sv”是指已经历基因组改变产生片段大小超过50bp的拷贝、结构和含量变化的dna区。术语sv用作单核苷酸变体/indel与片段拷贝数变体之间的操作分界。当测试基因组与参考基因组进行比较时,这些变化包含所述测试基因组中的缺失、新型序列插入、移动元件插入、串联和散布的片段重复、倒位、截短和易位。
[0228]
如本文所使用的,“靶dna”是指由参考基因组的起始和结束坐标(例如,人染色体2
中的bp 12900-14900)限定的dna区或正在被检测的已知序列含量(例如,基因或移动元件的序列)。
[0229]
如本文所使用的,“靶标富集”是指将除了用于分带的那些探针之外的另外的探针用于所关注靶向区域,以便跟踪所述特定区的任何变化。在某些方面,所关注靶向区域可以比带更小。在某些方面,所关注靶向区域可以限于带的一部分、覆盖一个整个带或跨越两个或更多个带的部分或整体。
[0230]
如本文所使用的,“载体并入的dna”是指充当dna插入物的媒剂的任何载体。这些载体可以是引入到细胞中的克隆载体、表达载体或质粒载体,包含但不限于人工染色体载体、噬菌体和噬菌粒载体、穿梭载体和粘粒载体。
[0231]
公开了用于通过用不同颜色的探针标记单链染色单体来检测染色体中的结构变异的方法。标记的探针的杂交模式产生光谱轮廓,所述光谱轮廓使得能够对结构变异进行高分辨率检测,从而促进区分良性变异与有害结构变异。进一步地,所述光谱轮廓提供关于复杂结构变异的信息,其中可能已发生多于一个染色体片段重排。
[0232]
单链染色单体可以通过本领域已知的任何方式产生,包含但不限于co-fish技术。
[0233]
能够与单链染色单体杂交的探针可以具有任何功能性长度。在不受任何特定实施例的限制的情况下,探针的长度可以为10到100个核苷酸、长度为15到90个核苷酸、长度为25到75个核苷酸、长度为30到50个核苷酸、长度为37到43个核苷酸或其任何组合。
[0234]
在某些方面,用于本文所公开的方法的标记的探针组可以根据一条或多于一条染色体上涉及特定染色体区的在有限数量的染色体区(例如,一个或多个染色体区)上提供基因座特异性分带的较小探针组或提供靶向整个基因组的染色体区的探针阵列的较大探针组在探针数量的范围内变化。
[0235]
在某些方面,用于本文所公开的方法的标记的探针组可以根据涉及一个或多于一个所关注基因的小探针组或靶向所研究生物体中所有已知基因的较大探针组在探针数量的范围内变化。一方面,探针的靶标可以相对相等地分散在整个基因组中。另一方面,探针的靶标可以更多地集中在基因组的某些区中,并且在基因组的其它区中更分散。
[0236]
在某些方面,标记的探针组可以被设计成靶向基因组内的基因座,所述基因座已知影响或引起疾病状态。一方面,探针组可以被设计成靶向已知与肺癌的发展或存在相关联的基因。类似地,探针组可以与本文所公开的方法一起设计和利用,用于任何所关注疾病或病状。
[0237]
在某些方面,标记的探针组可以被设计成靶向基因组内的基因座,所述基因座已知与特定疾病的不同状态相关。一方面,探针组可以被设计成指示疾病进展的状态,例如,在神经退行性疾病中。
[0238]
在某些方面,标记的探针组可以被设计成靶向基因组内的基因座,所述基因座已知与遗传病症相关。一方面,探针组可以被设计成针对遗传病症的产前诊断工具。
[0239]
在某些方面,标记的探针组可以被设计成靶向基因组内的基因座,以提供针对所关注的任何疾病或健康状况的诊断工具。在某些方面,疾病或病状可以选自呼吸道疾病、肌肉骨骼病症、神经病症、皮肤疾病、胃肠道疾病和各种类型的癌症。
[0240]
在某些方面,标记的探针组可以被设计成靶向基因组内的基因的特定类别。一方面,探针可以被设计成靶向不同类型的激酶的基因。
[0241]
在某些方面,标记的探针组可以被设计成集中于所关注研究领域。一方面,探针可以被设计成测试与生物医学科学中的基因组dna序列有关的几乎任何假设。
[0242]
在某些方面,标记的探针组可以被设计成提供包围一条或多条染色体的着丝粒的带,并且此类探针可以作为单个探针组或多个探针组运行,用于染色体鉴定和计数。在某些方面,可以用不同的颜色标记每条染色体的着丝粒的任一侧上的带,用于进一步分区分p臂和q臂。
[0243]
在某些方面,标记的探针组可以被设计成提供靶向一条或多条染色体的亚端粒区和/或端粒区的带。在一些方面,探针组的p和q臂末端带可以作为单独的探针组或作为多个探针组运行,用于跟踪一条或多条染色体的亚端粒区和/或端粒区。在某些方面,涉及一条或多条染色体的亚端粒区和/或端粒区的探针提供靶染色体的结构信息以及靶染色体的特定臂的结构信息。将用于带的探针应用于亚端粒区和/或端粒区提供了用于检测涉及靶向的亚端粒区和/或端粒区的结构重排事件的信息。
[0244]
任何单独带可以覆盖基因的部分或全部。同样,任何特定基因可以由一个或多于一个带的全部或部分覆盖。
[0245]
在某些方面,可以利用靶标富集策略,其中除了用于分带的那些探针之外,另外的探针还用于所关注靶向区域,从而检测所述所关注靶向区域的特征。在某些方面,所关注靶向区域可以比带更小。在某些方面,所关注靶向区域可以限于带的一部分、覆盖一个整个带或跨越两个或更多个带的部分或整体。在某些方面,可以用与靶标富集探针在其内杂交的带相同或不同的荧光团来标记用于靶标富集的探针。在其中相同的荧光团用于靶标富集探针的方面,荧光信号的强度在这样的通道中增强。在其中在靶标富集探针上使用不同荧光团的方面,产生组合荧光信号。
[0246]
在某些方面,被设计用于靶标富集的寡核苷酸探针具有与用于分带涂料的探针相同或不同的设计参数。使用相同的设计参数产生竞争性杂交,而使用不同的设计参数产生竞争性和非竞争性杂交的混合。靶标富集改进了检测极限并且改进了跟踪特定染色体基因座的能力。
[0247]
任何参考光谱轮廓都可以用作比较所研究染色体的光谱轮廓的基础。参考光谱轮廓可以是以下的光谱轮廓:具有已知异常的染色体、被视为正常的染色体、对应的姐妹染色单体、统计学上测定的正常轮廓、含有被认为具有正常或异常轮廓的染色体的参考数据的数据库或其任何组合。另外,因为针对参考基因组或序列设计的探针分布(即,经由电脑模拟的探针跨独特或重复序列的密度模式)与参考光谱轮廓有关(具有较多探针的区中亮度增加,并且具有较少探针的区中亮度减少),所以当靶标的预期光谱轮廓中存在偏差时,所述探针分布可以用于鉴定和描述测试样品中的结构变异。
[0248]
通过本公开的方法测定的结构变异可以是与正常不同的任何类型的结构变异,包含但不限于染色体片段的拷贝数的变化、倒位、易位、截短、姐妹染色单体重组、微核形成、染色体碎裂或片段化事件或其任何组合。片段的拷贝数的变化可以是缺失、扩增或其任何组合。
[0249]
标记的探针可以通过本领域已知的任何方式进行标记。探针还可以包括任何数量的不同类型的标记。探针的组合还可以具有任何数量的不同类型的标记,从而使一个探针的标记不同于另一个探针的标记。探针上的标记可以是荧光的。由探针上的标记发射的光
可以是在可见光光谱、红外光光谱、紫外光光谱或其任何组合中可检测到的。从探针发射的光可以在伪彩色中检测或以其它方式被分配不同于由探针发射的实际光的颜色的颜色。
[0250]
在一个实施例中,用于杂交的探针组包括其中不同探针被标记不同颜色的探针。所述探针组可以包括不同地标记的探针,其中所述单独探针用两种不同颜色(即,第一颜色的一个探针和第二颜色的第二个探针)、三种不同颜色、四种不同颜色、五种不同颜色、六种不同颜色、七种不同颜色、八种不同颜色、九种不同颜色、十种不同颜色、十一种不同颜色、十二种不同颜色、十三种不同颜色、十四种不同颜色、十五种不同颜色、十六种不同颜色、十七种不同颜色、十八种不同颜色、十九种不同颜色、二十种不同颜色、二十一种不同颜色、二十二种不同颜色、二十三种不同颜色、二十四种不同颜色、二十五种不同颜色、二十六种不同颜色、二十七种不同颜色、二十八种不同颜色、二十九种不同颜色、三十种不同颜色或超过三十种不同颜色标记。
[0251]
杂交探针上的标记的位置可以在探针上可以支持标记附着的任何位置。探针可以在探针的末端上标记、在探针的一侧上标记、在探针的主体中标记或其任何组合。探针主体上的标记可以是在探针的糖或亚酰胺官能团上。
[0252]
探针的检测可以通过本领域已知的任何方式执行。任何构件都可以用于对来自探针的光信号进行滤波,包含但不限于窄带滤波器。任何构件都可以用于处理来自探针的光信号,包含但不限于计算软件。在一些实施例中,仅来自探针的光特征的某些部分用于分析染色体结构变体。
[0253]
本文所公开的方法可以与用于检测染色体异常的其它技术组合实践。在一个实施例中,本文所公开的方法可以与染色体染色技术组合实践,所述染色体染色技术包含但不限于用dapi、hoechst 33258、放线菌素d或其任何组合对染色体进行染色。
[0254]
定向基因组杂交(dgh)是可以应用于测量错误修复比率和某些错误修复的身份的技术。此方法可以用于检测单独细胞中的中期染色体中的从头sv,或者可以用于评估涉及特定基因组基因座的sv。在先前的实施例中,取向变化(倒位)、姐妹染色单体交换和非交叉姐妹染色单体重组以及平衡等位基因易位的检测将通过单一方法可视化为单个细胞中相同的信号模式变化。除了标准的基于染色体的细胞遗传学分析方法可见的sv(不平衡和平衡非等位基因易位、倍性变化、大倒位、大插入和大重复)之外,还检测到这些sv。然而,除非采用靶向方法,否则将取向变化sv(高风险)与由sce和scr事件(低风险)产生的瞬时修复中间体和介于两个同源染色体之间的平衡易位(相对低风险)区分开通常是不可能的。近年来,已经描述了另外的错误修复类型和其对肿瘤发生和基因组不稳定性的相对贡献,这进一步说明除了明显需要更精确地绘制断点图和解释由dgh检测到的sv中涉及的基因组区之外,需要通过dgh可见的事件的更精确的分辨率。此处讨论的关于sce形成的分子机制的大部分工作涉及酵母研究,并且比对哺乳动物的了解要多得多。虽然不声称机制是相同的,但就过程类似而言,本技术中所描述的方法将帮助促进此类知识。
[0255]
因为dna错误修复可能导致细胞死亡或对患者构成风险,所以必须采用新型技术来测量错误修复的比率并提供无假设的sv从头鉴定。本发明将dgh方法与独特的dgh杂交探针设计和独特的图像分析方法组合,以提供具有显著增加的分辨率的sv鉴定和表征。由于这种表征包含位置和取向数据,因此它可以与关于哪些基因、启动子和基因组区的公开可用的生物信息学数据组合,以评估由对单独细胞的一个或多个错误修复引起的基因毒性风
险,以及与蛋白质组数据和转录组数据组合,以影响患者诊断。
[0256]
定向基因组杂交(dgh)可以作为从头方法进行,所述方法可以检测与参考(正常)基因组相比的结构变体,或者作为靶向方法,从而评估特定靶区(如编辑位点)处的结构变体(图5)。在两个实施例中,dgh方法被设计成定性的,并且提供关于单独细胞中一种或多种结构变体的流行或发生的确定性数据。当使用靶向实施例时,可以推断特定靶标的存在,因为测定被设计为针对靶标的二元测试。然而,尽管从头实施例在没有先前靶标假设的情况下能够检测几乎任何sv,但仅提供变体的粗略身份(例如,c3的p臂的推定的端粒倒位,大约为7mb),并且无法提供关于变体的重排类型、取向、大小、位置或序列的确定数据。
[0257]
经由光和暗带或多色带的差异染色的分带染色体是广泛用于区分正常核型与结构重排的核型的技术。每种分带方法都有其优点和缺点。g分带和反向(或使用dapi的r分带)和色霉素染色是产生染色体的差异明和暗分带的最广泛使用的技术,并且足以用于检测简单结构变体的子集,所述简单结构变体包含数字变体(整个染色体或染色体的大部分的数量的变化)、简单易位和一些大倒位(取决于带模式破坏的程度)。这些是快速且成本有效的dna染色方法,并且是临床诊断中用于核型分析的当前行业标准。尽管它们提供了基本核型信息,但这些技术对于检测较小数值变体(缺失和插入)和小倒位的效用非常有限,并且通常无法用于描述复杂的重排。除了描述观察到的涉及通常所关注区的明/暗带破坏外,它们不提供任何特定于基因座的信息。在易位的情况下,这些技术也具有显著的盲点。如果染色体分带模式呈现为交替的“...明-暗-明-暗...”序列,如在g分带中那样,那么交换断点位置的分辨率将固有地次于呈现为交替颜色序列的相同模式,比如,“...r-g-b-y...”。这些基于染色的方法在易位断点定位中经受适用于第一(明-暗)情况的“三带不确定性(three-band uncertainty)”(savage 1977)。另外,这些方法不检测平衡易位,所述平衡易位是在同一基因座处或附近基因座处具有断点的两个同源染色体之间的等价交换,这些方法也不检测姐妹染色单体交换/姐妹染色单体重组(基因转换)事件。
[0258]
如sky和mfish等全染色体fish涂染技术可以用于提供观察到的结构变体的更精确描述,因为每条染色体(每个正常细胞每条染色体的2个拷贝)以不同颜色标记。这些技术鉴定哪些染色体涉及观察到的重排,但这些技术无法提供断点坐标,也无法鉴定作为重排产物而包含或缺失的染色体的基因组片段。例如,与单色dgh涂料很像,缺失或扩增无法通过sky、mfish或类似方法归因于特定染色体的任何具体区或基因座。
[0259]
带特异性多色标记策略(最熟知的方法是mband)可以提供某些复杂事件的更解析的图片,包含鉴定具体染色体的哪些片段涉及重排,限于测定的分辨率。mband测定的分辨率由在任何给定区中的带大小的小巧(小)程度,以及样品用于分辨带的存在和相对顺序(例如,染色体的长度和伸展程度)两者有多合适来确定。但是,像所有其它基于fish的技术一样,mband无法检测介于同源染色体、小倒位或姐妹染色单体交换/姐妹染色单体重组事件(基因转换)之间的平衡易位,无论分辨率有多高。此外,带是通过dop-pcr扩增和差别地标记针显微解剖的染色体的部分以创建重叠的探针库并且针对高分辨率g分带和/或反向dapi分带评估正常核型中的这些带以推断每个带的定位而创建的。因此,每个带的精确起始和结束坐标是未知的,并且仅可以通过与具有正常核型的中期细胞的最高分辨率g分带进行比较来推断。
[0260]
如美国专利申请公开第2010/0304994号中提及的,“寡核苷酸染涂
(oligopainting)”将具有优于mband的优点,因为可以针对具有合成寡核苷酸的已知基因组坐标精确地设计带。每个带的精确开始和结束将是已知的基因组坐标,并且不是基于与正常核型上的明暗带的比较的估计。但是,像所有其它基于fish的技术一样,“寡核苷酸染涂”无法检测介于同源染色体、小倒位或姐妹染色单体交换/姐妹染色单体重组事件(基因转换)之间的平衡易位。
[0261]
目前公开的用于检测结构变异的方法提供了单色dgh涂料中缺失的元素:提供特定基因组坐标,并且将真正的倒位事件(涉及基因组片段的重新排序)与姐妹染色单体交换事件(其不改变基因组片段的顺序,但无法使用单色dgh涂料与倒位区分开来)区分开。与这2个事件相关联的风险(倒位风险高,sce风险低,因为它们本质上是“正确的修复”,并且不会导致基因组片段的顺序或拷贝数发生变化)对于临床医生来说很重要。存在与姐妹染色单体交换相关联的杂合性缺失(基因的一个好的拷贝被坏的拷贝取代-产生疾病表型)的风险,但在风险和患者结果的上下文中,姐妹染色单体交换应该与真正的倒位事件区分开。crispr cas9和依赖于dna断裂和dna断裂修复的其它基因编辑系统需要准确的风险概况。使用目前所公开的方法有可能将这些sce/scr“假阳性”与潜在的基因毒性事件(倒位)区分开。基因组片段的次序是可见的,并且信号在主要姐妹染色单体或相对的姐妹染色单体上的取向也是可见的(参见示意图)。由于所需的样品制备方法,k带被区分为与其它多色分带方法不同的技术,所述技术涉及从姐妹染色单体复合物中去除新合成的dna子链,从而提供允许染色单体特异性标记的单链模板。
[0262]
在基因编辑的上下文中,检测和鉴定在基因组的操纵和改变期间产生的结构变体是患者健康的优先事项。诊断界和监管机构广泛地认识到,需要在单个细胞中以高分辨率测量作为修复方程式的重要组成部分的倒位和姐妹染色单体交换以及缺失、扩增和易位。目前所公开的方法能够递送测序错过的以及使用其它差别分带或基于fish的分带方法不可访问的结构变体数据。如先前描述中概述的,测定的样品制备组分与寡核苷酸探针的单向性组合,使得能够评估无法通过其它分带技术检测到的事件,并且提供重要的另外水平的结构变体数据。由于酶导向基因编辑过程劫持并利用细胞合成和修复机制,因此它们为原本非常复杂的过程引入了另外的复杂性水平。用于确认编辑的测序方法以及用于评估基因组的其余部分的非预期效应的测序方法经常依赖于完整靶序列的存在来生成数据。然而,如果在靶序列的区中已发生切除和缺失,那么不可能扩增用于序列分析的区。在汇集的dna格式中,此信息将缺失-在筛选包含拷贝数变异和携带增加的基因毒性风险的结构变异时,这是值得关注的。复杂的结构变体也非常难以通过测序来评估。以此方式,最基因组不稳定且危险的结构变体最有可能被测序错过。在中期传播的上下文中,可测量和评估每个细胞的整个基因组是否存在结构变异,而无需任何扩增和序列分析。可以测量从头重排以及对所关注靶标的重排,并且可以随时间推移监测编辑细胞群中可能引起关注的无意自发和稳定结构变化(像癌症驱动的融合基因),以及期望编辑的随时间推移的稳定性。使用由本公开方法提供的基因组坐标特异性,测序可以用于对观察到的结构变体进行更深度的碱基对特异性查看。这两种技术可以协同使用,以使得能够更精确地检测和表征编辑的基因组。
[0263]
染色体外dna(ecdna)分析
[0264]
制备包括细胞的dna的生物样品以促进样品与寡核苷酸探针接触,所述寡核苷酸
探针是单链的、独特的并且与dna的至少一部分互补。在某些方面,包括细胞dna的生物样品进一步包括ecdna。ecdna和染色体dna两者可以与具有相同核酸序列和荧光特征的探针杂交。在其中ecdna和染色体dna被类似地标记的方面,可以测定ecdna来源于染色体上的哪个位置。
[0265]
可以选择用于在检查中对染色体进行分带的寡核苷酸探针,以特异性地定位在ecdna中发现的dna的染色体来源或起源。在某些方面,寡核苷酸探针与染色体dna的杂交模式的光谱分析允许鉴定ecdna中dna的染色体来源。染色体与ecdna之间的光谱特征的比较,在某些方面,光谱特征相似性的研究,提供了将特定染色体dna鉴定为并入ecdna中的dna的扩增区的来源。在某些方面,分析由探针与染色体dna的杂交产生的分带模式提供了对所研究染色体中所关注基因和区的鉴定。在某些方面,如果将多个带鉴定为并入到ecdna中的来源材料以进一步表征存在于ecdna中的序列,那么被鉴定为所研究的染色体中的所关注的一个或多个带可以用于影响特定探针或探针组的设计。
[0266]
用于分析ecdna的方法可以应用于外源dna、载体并入dna以及不存在于染色体上的细胞内的任何其它dna。
[0267]
光谱分析
[0268]
在某些方面,光谱成像和分析捕获关于一个图像中的所有荧光团的信息。在一些方面,由于带彼此紧密接近,相邻的带似乎渗色到彼此中。渗色可以用作另外的标志物,以基于邻近带的渗色的存在和通过信号与带信号的渗色比率来改进带内事件的定位。
[0269]
定向基因组杂交(dgh)扩增
[0270]
在某些方面,扩增显微术(asano等人(2018)细胞生物学中的当前方案(current protocols in cell biology)e56,第80卷)可以应用于dgh样品以改进dgh的空间分辨率。在某些方面,扩增显微术涉及将样品包埋在可溶胀水凝胶中,然后将样品化学地与水凝胶连接。然后可以对样品进行标记、溶胀和成像。使样品溶胀的过程将空间(x,y,z)分辨率增加到与非扩增样品上的共焦或超分辨率成像相当的水平。因此,实现了对事件,例如,结构变异进行定位的能力的改进。
[0271]
节点分析
[0272]
本文公开了用于鉴定受试dna链的一个或多个结构特征的方法。在某些方面,此类方法在处理器中实施。一方面,用于鉴定一个或多个结构特征的方法包括接收表示受试dna链上的至少一个碱基对序列的光谱轮廓,所述光谱轮廓包含对应于受试dna链的碱基序列的频率数据。所述频率数据可以分为至少两个颜色通道。在不同方面,各种数据包含在颜色通道中,包含但不限于定位数据和强度数据。光谱轮廓可以转换成数据表,所述数据表包括定位数据、强度数据以及被确定为至少两个颜色通道中的所关注的其它数据。可以将由此为受试dna链生成的数据表与包含表示对应的对照dna链的正常和/或异常特征的一个或多个特征节点的参考特征查找表进行比较,以鉴定受试dna链的一个或多个正常和/或异常特征。一方面,特征节点由表示在起始碱基处开始并且在结束碱基处结束的对照dna链的碱基子序列的色带限定。
[0273]
节点分析,其中受试dna序列的光谱轮廓信息被转换为数字形式以与对照或参考dna序列进行比较,可以与本文所公开的定向基因组杂交方法结合进行,或者可以在提供可转换为数字形式的多核苷酸序列数据的其它方法的上下文中使用。在某些方面,参考或对
照查找表是单个值表或多个值表。在一些方面,不同的参考或对照查找表提供对应于不同基因组区的值。在某些方面,通过机器学习和/或ai算法执行来自受试dna的查找表与参考或对照查找表的比较。来自受试dna链的光谱轮廓数据的值可以通过对照或参考查找表的分析与特定节点相关。然后,可以通过节点分析运行节点组,以找到相关途径或效应途径,其中节点之间的关系先前已知或通过分析确定。
[0274]
在某些方面,来自受试dna链的光谱轮廓数据可以存储在存储器中以供稍后比较和分析以确定所关注的结构特征。在一些方面,光谱轮廓数据可以存储在关系数据库、图形数据库、查找表或任何其它生物信息学数据库格式中。
[0275]
在一些方面,受试dna链上的所关注特征可以被表征为对应于健康对照dna链上的特征的正常特征。在一些方面,受试dna链上的所关注特征可以被表征为对应于表示至少一个异常的参考dna链上的特征的异常特征。
[0276]
在某些方面,从在空间上不共定位的dna区分析光谱轮廓数据。在一些方面,光谱轮廓数据来源于空间邻近的dna区。在某些方面,基于探针序列、光谱、寡核苷酸密度、染色体、染色体臂、带id、带取向和带覆盖(例如,基因区)的一系列键连接光谱轮廓数据。在一些方面,基因组特征可以由带、带光谱、带序列、带取向和带最近邻或通过探针、探针光谱、探针取向和探针最近邻来限定。
[0277]
在某些方面,可以通过在探针的5'末端、带或所关注区处开始,然后分析连续朝3'末端移动的每个带的带光谱、大小和覆盖来限定跨特征、染色体臂或染色体的序列。在一些方面,这些特征被转换成图例,所述图例可以与数据库进行比较以确定畸变或异常的位置和特征,并且通过扩展,确定数据库中的哪些节点受那些畸变或异常影响。畸变或异常的一些组合指示特定重排事件,例如,一个区中的截断带与不同区中相同光谱的额外信号组合将指示易位事件。
[0278]
可以使用任何统计分析工具对光谱轮廓数据进行分析或元分析,所述工具包含但不限于:图论、节点分析、人工智能、机器学习(包含k最近邻、主成分分析等)以及神经网络。
[0279]
本文所公开的方法可以与将多种类型的数据并入到数据库以供分析的方法组合。在某些方面,来自其它来源的数据包含但不限于测序、基因组学、转录组学、蛋白质组学和代谢组学。在某些方面,根据测序数据分析倒位、姐妹染色单体交换和其它dgh特异性数据。可以针对已知的公开的测序数据或针对新型或未公开的数据进行比较。
[0280]
在一些方面,由本文所公开的方法产生的数据汇总在报告中,其中自动生成的表意图示出样品水平和队列或实验水平上的独特且反复的重排和分析、元分析或节点分析。
[0281]
实例
[0282]
描述以下实例是为了更全面地展示本发明的一些实施例。然而,所述实例决不应该被解释为限制本发明的广泛范围。本领域普通技术人员可以容易地采用此发现的基本原理来设计各种化合物而不背离本发明的精神。
[0283]
实例1
[0284]
图6提供了来自为dgh制备的辐射暴露的血液衍生淋巴细胞样品的重排细胞中单色dgh涂料标记的染色体1、2和3的示例图像。在asi扫描显微镜系统上获取图像,并使用genasis细胞遗传学软件查看所述图像。由软件将来自所选中期的染色体组织成核型图(以竖直取向显示染色体,并将所述染色体组织成来自全中期扩散的原始图像的同源物对),并
且裁剪标记的染色体1、染色体2和染色体3同源物对,并且从原始中期扩散图像放大。在经裁剪和放大的核型图下方提供的整个中期扩散。在此细胞中,存在涉及涂染的染色体(染色体1、2和3)的明显重排,但是确认存在真实的结构变体对姐妹染色体交换事件在没有所述位置处的分段顺序的参考,即观察到的未涂染的姐妹染色单体的信号开关的情况下是不可能的,也不可能确定在每条染色体上观察到的事件的基因组坐标。
[0285]
实例2
[0286]
使用bj-5ta正常人成纤维细胞细胞系的染色体2dgh多色带试点实验。实验说明:19个独特序列寡核苷酸探针池以交替的颜色模式用5种不同的荧光团进行标记。除了在染色体2的末端处的最后一个探针池,每个探针池具有相同数目的寡核苷酸,所述最后一个探针池中的寡核苷酸的量为所述寡核苷酸的量的大约1.6倍。根据可用的独特序列在染色体2之上的分布,寡核苷酸池跨越较长或较短的dna段扩散,使得对染色体2具有独特的“指纹模式”。参见表1获得每个标记的池(带)的bp位置开始到结束、每个标记的池的总靶标大小(以bp为单位)、每个标记的池的寡核苷酸数量以及荧光团跨dna靶区的密度分布。所述表中还包含每个荧光团的伪色分配(一些荧光在可见光谱之外和/或在叠加中具有视觉上类似的颜色),因此,每个颜色通道都被分配了一种伪色,从而允许将带可视化为彼此不同。描绘了表中颜色的顺序以及模板链分配(沃森和克里克,因为它们对应于每个姐妹染色单体)。分配给“克里克”姐妹染色单体的颜色为蓝色,反映dapi dna染色的颜色,在本实验中未被探针标记的端粒、亚端粒和着丝粒区的颜色也一样。带颜色和链分配反映正常中期染色体2(为dgh制备)的基因组坐标。对于此初步实验,带大小范围为900万-1500万碱基对(mb)。对于此实验,在染色体8和染色体1两者上都包含几个对照探针点,用于确认分辨率和杂交质量。应注意,涉及此多色涂料的所有实验中包含的图像被转换为黑色和白色,并且必须使用表和带的出现顺序来推断全色光谱。
[0287]
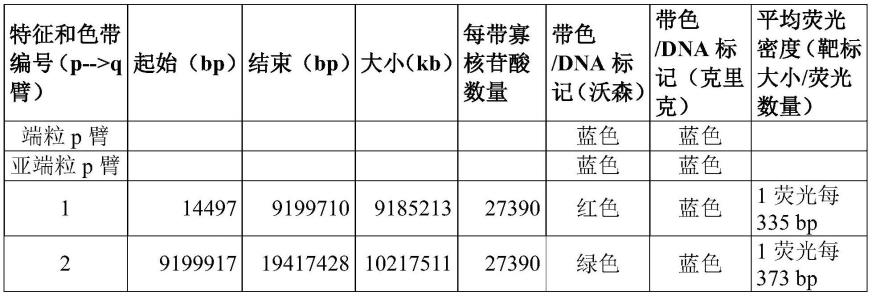
表1:
[0288]
[0289][0290]
图7的图像a和b示出了来自两个单独的正常中期细胞的染色体2同源物对,不存在结构变异(正常永生化人成纤维细胞系bj-5ta)。在asi扫描显微镜系统上获取图像,并使用genasis细胞遗传学软件查看所述图像。由软件将来自所选中期的染色体组织成核型图(以竖直取向显示染色体,并将所述染色体组织成来自全中期扩散的原始图像的同源物对),并且裁剪标记的染色体2同源物对,并且从原始中期扩散图像放大。
[0291]
另外,对显示异常信号模式的2个细胞(来自使用相同细胞系的相同实验)进行成像和分析。图7的图像c和d示出了来自2个单独的中期细胞(正常永生化人成纤维细胞系bj-5ta)的染色体2同源物对,示出姐妹染色单体交换产生的一个同源物中的结构变异(维持了颜色的顺序,但信号存在于相对的姐妹染色单体上)。注意:在使用单色涂料的情况下,端粒或亚端粒探针是区分大倒位(错误修复)和姐妹染色单体交换(完美修复)事件所必需的。如果在端粒或亚端粒区中存在另外的姐妹染色单体重组事件,那么使用单色涂料加端粒/亚
端粒方法可能会混淆此类事件的分类。新颖实施例允许对结构重排事件进行检测和准确分类两者。在图像c中,右边的染色体2同源物具有sce,其中sv的断点平分带#13,并且右边的同源物正常。在图像d中,左边的同源物具有sce,其中断点在带#9与#10之间发生,并且右边的同源物正常。
[0292]
实例3
[0293]
使用最近暴露于电离辐射以用于前列腺癌治疗的血液衍生淋巴细胞的染色体2dgh多色带试点实验。
[0294]
使用实施例1中描述的测定,dgh测定由19个独特序列寡聚物探针池(每个池跨越9mb-15mb)组成,所述池用交替的颜色模式标记,使得颜色的顺序对应于基因组坐标,在为dgh制备的辐射暴露的血液衍生淋巴细胞样品上运行正常的中期染色体2。图4示出了来自中期细胞的染色体2同源物对,其中鉴定出的sv原本是不可能表征的。存在大的臂间倒位(可能可由当前的细胞遗传学技术检测到,但由于发生在染色体最远端处的带破坏的性质,可能会被遗漏),连同着丝粒附近的较小臂内倒位(来自q臂上大多数标记的池的相对的姐妹染色单体的品红色探针乱序),以及非常接近所述较小臂内倒位的较大姐妹染色单体交换事件-所有这些都可以使用交替颜色作为参考框架来描述。难以在彩色组合叠加(示出)中可视化的重排可以通过查看每个单独颜色通道上的信号来确认。注意:如图4(2a、2b)所示,如果此细胞已经用单色染色体2dgh涂料标记,那么此染色体看起来会具有小末端sce或倒位(p臂)以及大倒位(q臂),并且会错过对现有结构重排的真正分类。图4中示出了杂交的染色体对的图像,对应的表意图示出为1a和1b,并且描绘dgh单色涂料的相同表意图在右边。在图4(1a)中,示出了制备用于dgh并且使用具有多色带的染色体2dgh涂料杂交的正常染色体2同源物。在图4(1b)中,示出了来自具有复杂结构重排的相同细胞的第二染色体2同源物。存在大的臂间倒位,其中一个断点出现在2p上带1与2之间,并且另一个断点在2q上平分带18。在2q上的着丝粒附近存在另外的较小臂内倒位,其中第一个断点在带9与10之间,并且第二个断点在带10与11之间。还存在带9与11之间与小的臂内倒位共享相同的近端断点的大的姐妹染色单体交换事件,所述姐妹染色单体交换事件也可以用带的顺序来验证,所述带仍然以正确的数字顺序出现,但现在位于与主要涂料(右姐妹染色单体)相反的姐妹染色单体(左姐妹染色单体)上。
[0295]
如果没有色带来提供片段的顺序,那么无法在坐标中鉴定或描述重排。实际上,使用右边的示意图作为视觉参考,染色体似乎具有小末端sce或倒位(p臂)和大倒位(q臂),并且存在的结构重排的真正分类将被错误鉴定。
[0296]
实例4
[0297]
使用来自实例2的测定、细胞系和成像方法,获取沿每个姐妹染色单体的光谱强度测量结果并连同跨染色体2的寡核苷酸密度分布一起绘制。图8a示出了正常染色体2的杂交、探针分布和荧光波长强度。姐妹染色单体被描绘为“沃森”和“克里克”。测量两个姐妹染色单体的颜色通道。在姐妹染色单体克里克上,显示的信号强度表示每个通道上的背景噪声,其中实际信号强度峰值在沃森上可见。信号强度峰值与寡核苷酸分布图和染色体图像叠加一致。图8a中为基因组背景提供了染色体2的表意图。图8b示出了正常染色体2中检测到的sce的杂交、探针分布和荧光波长强度。姐妹染色单体被描绘为“沃森”和“克里克”。测量两个姐妹染色单体的颜色通道。图8b中为基因组背景提供了染色体2的表意图。在姐妹染
色单体沃森和克里克上,光谱轮廓上存在或不存在信号峰值与每个姐妹染色单体上的可见信号竖直地对应。估计sce的断点平分带14(以橙色示出)。信号强度峰值与寡核苷酸分布图和染色体图像叠加一致。图8b中为基因组背景提供了染色体2的表意图。图8b中估计了断点区id。
[0298]
实例5
[0299]
序列梯图像-简介:中期扩散制剂中的染色体浓缩(紧凑对长)在细胞之间和细胞制剂之间变化。在通过dgh测定确定sv检测的分辨率之前,必须在评估中考虑此材料可变性。例如,在更长、更拉伸的染色质构型中,来自紧密间隔开的探针的杂交信号可以解析为单独的信号,并且在更紧凑和浓缩的染色质中,来自紧密间隔开的探针的杂交信号将表现为单个合并信号。在如图9所示的中期扩散中,3个单独的序列梯测定与染色体杂交。一个序列梯测量关于对每个信号有贡献的寡核苷酸数量的检测极限,在染色体2的p臂上间隔约20mb(在图像中标记为序列梯1)。第二序列梯(染色体2q)评估靶标大小,固定量的寡核苷酸可以分散在上面,也间隔约20mb,并且还测量检测极限(在图像中标记为序列梯2)。第三序列梯(在下方看到的与染色体1q杂交),具有间隔很近的以及间隔很远的探针,从而允许评估在任何给定中期传播中紧邻的两个点的可分辨性(在图像中标记为序列梯3)。这些序列梯针对来自分带涂料的相对的dna链进行设计,并且可以用作每个扩展中的测定分辨率的内部对照。
[0300]
实例6
[0301]
可以在为dgh制备的中期样品上运行包含一种颜色的易碎位点相关联的alu重复序列和其它颜色的多色分带dgh涂料的探针杂交的测定。alu重复序列(已在参考基因组中进行了表征和绘制)可以显示和检测为与已知脆弱位点和已知对基因调控很重要的区密切相关联的独特分带模式,使得就可以将观察到的已知或从头重排的接近程度与已知的脆弱区进行比较。如通过测定可视化的重排的染色体中存在的结构变体可以用于使表型与基因型相关,因为所述结构变体与基因组的已知高风险区相关。
[0302]
实例7
[0303]
多色分带涂料可以与分配给包围所关注靶标的区的两个特定色带相结合,并在为dgh制备的样品中期上运行。在同一视场中,包围所关注靶标的两种颜色可以在间期细胞(核(nuclei))中显示为细胞间靶向探针“分离”测定,所述测定通过对特定颜色通道的选择性分析示出与染色体涂料的剩余部分分离的特定区活性,从而允许分析细胞周期的g1、s和g2相以及通过所有细胞检查点并成功进入中期的细胞。样品中存在的间期核通常多于载玻片制备时中期中存在的间期核,并且存在的任何核将在中期扩散的同时与探针杂交。当各层中的若干种类型的数据与特定的成像方法相结合以单独地使基因组区可视化并且所述数据在含有中期细胞和间期细胞的样品中彼此相关时,可以通过单个测定提供所述数据。
[0304]
实例8a具有未知来源的可见大ecdna的癌细胞系可以与每个人类染色体的具有独特颜色的dgh整个染色体涂料杂交。经扩增且包含在ecdna中的染色体dna将含有与起源染色体相同的一种或多种信号颜色。一旦鉴定,已知包含也存在于ecdna中的遗传物质的特定染色体可以与对应于先前鉴定的起源染色体的一种或多种分带涂料进行连续杂交。可以被鉴定为标记的ecdna的区或dna坐标将对应于分带染色体中的一种或多种特定带颜色。可以针对已鉴定的起源区用特定的靶向探针进行进一步细化坐标,所述坐标将出现在ecdna和
对应的染色体上,并且可以用于跟踪和描述基因组的潜在有害变化。
[0305]
关于通过引用和变更并入的声明
[0306]
贯穿本技术,所有参考文献,例如专利文件,包含已发布或授权的专利或等同物、专利申请出版物和非专利文献文件或其它来源的资料,均通过全文引用的方式并入本文,如同通过引用的方式单独地并入,在一定程度上每个参考文献至少部分地不与本技术中的公开内容不一致(例如,部分不一致的参考文献通过引用的方式并入本文,参考文献的部分不一致的部分除外)。
[0307]
本文已经采用的术语和表达被用作描述性而非限制性术语,并且不旨在使用不包含所示出和描述的特征或其部分的任何等同物的此类术语和表达,但应认识到,在所要求保护的本发明的范围内可以进行各种修改。因此,应当理解,尽管已经通过优选方面、示例性方面和任选的特征具体地公开了本发明,但是本领域的技术人员可以对本文公开的概念进行修改和变更,并且此类修改和变更被认为是在如由所附权利要求书限定的本发明的范围之内。本文提供的具体方面是本发明的有用方面的实例,并且对于本领域技术人员来说将显而易见的是可以使用本说明书中阐述的装置、装置组件、方法步骤的大量变化来实施本发明。如对于本领域技术人员来说将显而易见的是,可用于本发明方法的方法和装置可以包含大量任选的组成以及处理元件和步骤。
[0308]
本说明书中所提及的所有专利和出版物指示本发明所属领域的技术人员的水平。本文引用的参考文献通过全文引用的方式并入本文以指示截至其公开或提交日期的现有技术,并且意图是在必要时,在本文中可以采用此信息,以排除现有技术中的具体方面。例如,当要求保护物质的组合物时,应当理解,在申请人的发明之前本领域已知和可获得的化合物(包含在本文引用的参考文献中提供了启用公开的化合物)并不旨在包含在本文的物质权利要求的组合物中。
[0309]
如本文所用,“包括”与“包含”、“含有”或“特征在于”同义,并且是包含性的或开放式的,并且不排除另外的、未列举的要素或方法步骤。如本文所使用的,“由...组成”排除未在权利要求要素中指明的任何要素、步骤或成分。如本文所使用的,“基本上由...组成”不排除对权利要求书的基本和新颖特征没有实质性影响的材料或步骤。在本文的每种情况下,术语“包括”、“基本上由...组成”和“由...组成”中的任何一个可以用其它两个术语中的任一个代替。可以在不存在本文未具体公开的任何一个或多个要素、一个或多个限制的情况下适当地实践本文说明性地描述的本发明。
[0310]
除非另有解释,否则本文所用的所有技术和科学术语都具有与所公开的公开所属领域的普通技术人员通常所理解的含义相同的含义。除非上下文另外明确指出,否则单数术语“一个/一种(a/an)”和“所述”包含复数指示物。类似地,除非上下文另外明确说明,否则单词“或”旨在包含“和”。“包括”意指“包含”;因此,“包括a或b”意指“包含a”或“包含b”或“包含a和b”。本文引用的所有参考文献通过引用并入。
[0311]
本领域普通技术人员将理解,除了具体举例说明的那些之外的起始材料、生物材料、试剂、合成方法、纯化方法、分析方法、测定方法和生物方法可以用于实践本发明,而无需进行过度的实验。任何此类材料和方法的所有本领域已知的功能等同物都旨在包含在本发明中。已经采用的术语和表达被用作说明性而非限制性术语,并且不旨在使用不包含所示出和描述的特征或其部分的任何等同物的此类术语和表达,但是应认识到,在所要求保
护的本发明的范围内可以进行各种修改。因此,应当理解,尽管已经通过优选方面和任选特征明确地公开了本发明,但是本领域的技术人员可以对本文公开的概念进行修改和变更,并且此类修改和变更被认为是在如由所附权利要求限定的本发明的范围之内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。