苏云金芽孢杆菌菌株
发明领域
1.本发明属于害虫防治领域,具体涉及苏云金芽孢杆菌(bacillus thuringiensis)的害虫防治。更具体地,本发明涉及苏云金芽孢杆菌(bt)的新型细菌菌株svbs-1801,其已证明具有对草地贪夜蛾(spodoptera frugiperda)的杀虫活性。目前市场上可用的基于bt的产品在控制草地贪夜蛾害虫方面要么无效要么效率低下。
2.发明背景
3.苏云金芽孢杆菌(bt)是一种需氧和革兰氏阳性土壤细菌,由于其在缺乏营养物质的情况下具有孢子形成能力,因此在分类学上被归类为蜡状芽孢杆菌(bacillus cereus)。bt以合成多种毒素而闻名,这些毒素靶向破坏栽培植物并显著降低其产量的的昆虫害虫和其他生物(http://www.lifesci.sussex.ac.uk/home/neil_crickmore/bt/toxins2.ht ml)。几种bt毒素是细菌合成的蛋白质聚集的结果。这发生在孢子形成阶段期间,并导致形成伴孢体(晶体),它构成了bt与同一分类群的其他物种相比的差异特征。该晶体包含-内毒素cry和cyt,它们分别是昆虫靶向毒素和溶细胞蛋白。后者具有相当非特异性的活性并仅在少数bt菌株中发现。形成晶体的蛋白质在其结构中含有二硫桥和疏水键,使它们具有蛋白酶抗性。因此,一旦孢子形成阶段结束,这些蛋白质可以在bt营养细胞的细胞质内积累并促进高浓缩的活性化合物的释放。
4.bt自其表征以来一直被认为是作物和储存产品的有效害虫防治剂。因此,过去几十年见证了全球范围内采样计划的开始,导致发现了具有不同杀虫特性的各种菌株。这些微生物最初是根据它们的鞭毛抗原进行分类的(lecadet等人,1999)。然而,现在人们普遍认为,某种菌株的杀虫特性与其抗原h类型之间没有联系。每个bt菌株都显示出独特的毒性模式,这取决于它产生的毒素的相对比例。一些bt菌株可产生热稳定的分泌外毒素或β-外毒素,其与atp竞争rna聚合酶结合位点,因此,使其对多种物种有毒并阻止其被用作杀虫剂(sebesta&horsk
á
,1970)。
5.schnepf和whiteley首先通过在大肠杆菌中克隆和表达cry基因证明了bt晶体的杀虫特性(schnepf和whiteley,1981)。从那时起,许多其他的cry和cyt基因已被证明对分类为鳞翅目(baig等人,2010)、双翅目(roh等人,2010)、鞘翅目(l
ó
pez-pazos等人,2010)和膜翅目(sharma等人,2008)的昆虫物种具有特定的杀虫活性。其他植物噬菌体(phytophage),如螨虫(frankenhuyzen,2009)和线虫(hu等人,2010)也对此类bt毒素易感。
6.有一些值得注意的毒素变体没有整合到bt晶体中,但也表现出令人感兴趣的杀虫特性。蛋白质cry1i、vip(营养期杀虫蛋白)(estruch等人,1996)和sip(分泌的杀虫蛋白)(donovan等人,2006)就是这种情况,它们在细胞营养生长期间分泌到培养基中。有趣的是,所有具有所述杀虫活性的毒素变体(cry、cyt、vip、sip)通常都编码在大约40kbp的质粒中,尽管在某些情况下,相应的基因也可能存在于细胞染色体中。
7.自从carozzi及其同事(carozzi等人,1991)首次引入聚合酶链反应(pcr)作为检测bt杀虫基因的方法以来,它一直是对bt集合进行分类的最广泛方法,直到最近(cer
ó
n等人,1995;ferrandis等人,1999)。然而,高通量测序的最新进展使得以合理的时间/成本比
实现对某种bt菌株的完整遗传表征成为可能。了解bt菌株的完整染色体和质粒序列使得可以检测负责其杀虫特性的基因簇,这已经用作确定此类特定菌株的宿主的指南。尽管如此,这样的估计结果是不准确的,因为它没有考虑每个基因的表达模式,而这最终决定了在晶体中发现的或分泌到培养基中的毒素的相对比例。
8.可用杀生物剂基因的完整列表由1000多个不同的序列组成(crickmore,n.等人,2018),并按照crickmore提出的标准(crickmore等人1998)进行分类,这些标准基于核苷酸和氨基酸序列。该分类系统的工作方式如下:具有相同编号的所有cry基因(例如cry1)共享45%或更高的同一性百分比。如果它们也共享相同的大写字母(例如,cry1a),则此类同一性必须为78%或更高。此外,如果后跟相同的小写字母(例如,cry1aa),则认为它们之间的同一性至少为95%。最后,为了区分编码具有95%或更高同一性的蛋白质的两个基因,可以在小写字母后添加另外的数字(例如,cry1aa1)。
9.所描述的cry基因的数量在不断增长,其中一些基因显示出同一性百分比低于45%,表明它们是具有潜在独特杀虫特性的新候选基因。因此,对bt毒素易感的宿主数量显著增加,包括新的昆虫、螨和线虫物种。尽管bt的兴趣主要集中在害虫防治上,但新基因的发现使得发现其他研究领域的蛋白质成为可能。副孢菌素就是这样一个例子,它是被证明具有抗肿瘤药物的功效的蛋白质家族(ohba等人,2009)。
10.1938年,法国制造了第一种基于bt的配方产品,并且60年代,usa、urss、法国和德国配制和生产了大量bt工业产品(milner,1994)。80年代,由于昆虫种群中出现了抗药性个体,化学杀虫剂已经开始显示出缺陷,人们对bt衍生产品的兴趣急剧增加。然而,随着合成拟除虫菊酯的出现,这种兴趣很快就消失了。人们越来越有兴趣将化学合成的杀虫化合物替换为天然化合物,或甚至是能够控制害虫同时对农作物和/或脊椎动物(如人类)无害的活生物体,这使得人们对基于bt的产品产生了新的兴趣。
11.如今,由于全球各地的采样计划,对bt多样性及其杀虫基因分布的了解,将这种细菌描述为产生新杀虫剂的来源。对于要在发达国家使用这些类型的产品,它们必须适应当前的法规,这在大多数情况下要求bt菌株不含β-外毒素。原因是这些外毒素表现出相当不特异性的广泛作用,可能对包括人类在内的其他生物体有害(sebesta yhorsk
á
,1970)。
12.bt菌株的杀虫特征在传统上是通过执行生物测定法来阐明的。然而,由于它们耗费资源时间的性质,它们已经被通过pcr鉴定cry基因得到部分地取代。然而,基因含量只能用作估计,因为它没有显示每个基因表达量的信息(masson等人,1998)。晶体内的蛋白质相互作用也可能影响其杀虫特征,因此功能多样的cry和/或cyt蛋白质的混合物可能比预期的更有效(协同作用)或更无效(拮抗作用)(tabashnik,1992);因此,无法根据对单个内毒素的了解预测bt蛋白混合物(通常,bt菌株产生一组cry蛋白)的毒性(hilbeck和otto,2015)。因此,仍然需要生物测定作为信息的补充来源,以充分表征bt菌株。
13.因此,寻找针对特定昆虫或线虫害虫具有活性的新bt菌株不仅因为每个bt菌株表现出取决于其产生的毒素、它们的相对比例以及产生的毒素之间可能出现的不可预测的协同或拮抗作用的独特毒性模式而受到阻碍,也因为具有高度同源性的cry蛋白可能在特异性和功能性方面表现出重要差异,因此很难尝试推断其活性来预测可由其他bt菌株产生的同源蛋白的活性,产生与另一种cry蛋白非常相似的cry蛋白的bt菌株可具有对同一组昆虫的活性或活性程度(同源cry蛋白在与bt菌株产生的蛋白质混合物中的其他bt蛋白相互作
用时可表现出)并非明显。
14.使寻找和鉴定对特定昆虫具有高活性的bt菌株变得困难和不明显的附加复杂因素是,关于某些cry蛋白的活性和特异性或关于一些已知bt菌株的杀虫潜力的现有信息不完整,得不到实验数据支持或者基于测定,结果以相对不同于本领域技术人员通常处理的量值或单位表示,因此降低了已发布信息的实用性和适用性。
15.例如,在控制草地贪夜蛾的特殊情况下,可以找到一些不同的出版物,这些出版物涉及据说具有抗草地贪夜蛾活性的bt菌株,但不包括确认其的测定法,或包括以不同于本领域技术人员用于表达杀虫活性的标准单位的单位表示的结果,使得难以评估其有效性程度。人们还可以找到一些出版物,这些出版物报告了一些cry蛋白针对草地贪夜蛾的活性,这些活性不是基于对鳞翅目昆虫的分析,也不是基于对单个蛋白质进行的分析,这不允许推断所测定的蛋白质在与bt菌株产生的其他蛋白质相互作用时可能具有的活性,或氨基酸序列变化可能导致的活性和特异性的差异,即使与其他cry蛋白质保持高度同源,如上所述。
16.例如,欧洲专利申请ep-0366397-a1描述了一种据说具有针对鳞翅目害虫活性的bt菌株(ps814)。然而,没有显示针对草地贪夜蛾的活性测定法。该菌株在其基因组中包含与称为cry1ea7的基因序列具有100%的同一性并完全重叠的基因(数据库btnomenclature,http://www.btnomenclature.info中的登录号ay894137),该基因在文件中以δ内毒素基因的形式呈现,该基因针对鳞翅目害虫具有活性,但没有测定法证明相应的cry蛋白对任何昆虫害虫具有任何活性。
17.美国申请us2010/160231-a1公开了包括编码据说对应于杀虫多肽的氨基酸序列的编码序列的组合物,以及用于将杀虫活性赋予细菌和其他细胞和组织(包括种子)的方法。尤其是命名为axmi-150的δ-内毒素的编码序列,显示出与已知为cry1nb1的基因序列的完全重叠和100%同一性(数据库btnomenclature,http://www.btnomenclature.info中的登录号kc156678)。尽管该文件包括可由包含该基因的重组菌株或包含该多肽基因产物的组合物控制的昆虫长列表,但仅对蛋白质axmi-150的杀虫活性进行了测定并显示对四种害虫的杀虫活性,四种害虫中没有一种是灰翅夜蛾属(spodoptera)的成员,尤其是四种害虫中没有一种是草地贪夜蛾:欧洲玉米螟(ostrinia nubilalis)、天鹅绒豆毛虫(anticarsia gemmatalis)、小菜蛾(plutella xylostella)、西南玉米螟(diatrella grandiosella)。
18.polanczyk及其同事(polanczyk等人,2000)报告了几种苏云金芽孢杆菌菌株针对草地贪夜蛾的测定法。上述文章的图1显示了每种受试菌株的草地贪夜蛾死亡率。效果最好的菌株属于aizawai血清型,与商用产品中存在的菌株相同。结果以每单位体积的细菌(细胞数/ml)表示,这与制备商用杀虫剂中活性成分的其他标准单位(微克/ml)不同,因此很难评估和比较这种菌株与目前使用的商用杀虫剂的杀虫能力。提供的关于该菌株的数据并没有明显表明寻找具有更好杀虫能力的替代天然菌株来对抗草地贪夜蛾。
19.capalbo及其同事(capalbo等人,2001)公开了一种用于发酵bt菌株bt变种tolworthi的方法,该菌株先前被描述为具有针对草地贪夜蛾的活性。发酵是在固态中进行的,而不是在液体介质中。没有显示关于该菌株本身的具体数据,例如其cry基因含量或蛋白质。包括关于其有效性的测定法,显示0.37mg的生物量/ml为能够杀死50%的昆虫的致死浓度,即lc
50
,但该文献的材料和方法部分没有确定该值的方法,这使得难以评估和比较该
菌株与其他bt菌株的杀虫活性。
20.barreto及其同事(barreto等人,1999)公开了几种bt菌株的vip蛋白针对草地贪夜蛾的活性测定。该文献摘要提到了所有描述的菌株之间的显著差异,尽管它们都属于同一血清型(berliner)。这突出表明,仅根据遗传和分子特征很难找到针对特定物种(如草地贪夜蛾)具有高活性的bt菌株。在d5中,据说具有较高活性的所测得上清液可能含有β-外毒素(见第680页左栏末尾及其右栏的延续),这是本发明目的不需要的遗传特征,因为它们不适用于生物杀虫剂,并且这在sbvs-1801菌株中不存在。在整个d5中,没有证据表明所述菌株与sbvs-1801、从其培养物中获得的成分或其用作杀虫剂或用于制备杀虫剂有关。
21.其他不同的已发表文献描述了来自bt菌株(从中分离或通过天然蛋白突变获得)的单个δ-内毒素蛋白,据说这些菌株具有针对草地贪夜蛾的一定活性。然而,大多数所述文件教导或讨论了不完整或未经适当测定法证实的信息,这使得这些文献在搜索具有针对草地贪夜蛾具有高活性的新的、天然的bt菌株方面的效用降低:例如,本领域技术人员将发现,例如一些文件没有公开来自几种cry/cyt蛋白组合的实验数据,因此,忽略了杀虫蛋白之间相互作用的影响,这是在天然条件下将由bt菌株产生的结果,或者用于功效测试的蛋白质浓度不是由天然bt菌株产生的,这取决于培养条件。还必须考虑到,bt菌株产生特定多肽并不意味着该菌株将生成具有针对草地贪夜蛾的高活性的晶体。这些文献的某些特定方面如下:
22.以wo 2012/131495 a2公布的国际专利申请公开了天然δ-内毒素cry1ab的几种突变多肽。源自cry1ab的突变体是含有氨基酸取代的突变体。这些重组蛋白显示针对草地贪夜蛾的活性,其中一些甚至比天然多肽更高。虽然进行测定法的条件没有很好的描述,但实验仅涉及蛋白cry1ab及其衍生突变体作为活性成分。因此,wo2012/131495缺乏cry蛋白之间的相互作用测定法,这可能会对整体杀虫效力产生显著影响。此外,使用的浓度是人为的,并因此很难预测它们是否与bt菌株产生的浓度相当。此外,wo 2012/131495中提到的仅有微生物是那些用编码突变多肽的核酸转化的微生物。因此,上述国际申请很难被视为用于识别具有针对草地贪夜蛾的活性的新的天然和替代bt菌株的适当信息来源。
23.中国专利cn 103333230 b描述了一种δ-内毒素,cry1da3,其由可在btnomenclature数据库以登录号hq439784找到的dna序列编码。尽管cry1da3针对小卷心菜蛾(pieris brassicae)和甜菜夜蛾(spodoptera exigua)的活性已被阐明,但并未提及夜蛾属的其他物种,特别是没有提及草地贪夜蛾及其可能的防治效果。
24.国际专利申请wo 95/04146a2公开了一种基因(cryet4)和一种蛋白质(cryet4),该基因和蛋白质与被称为cry1ja1的基因(在btnomenclature数据库中的登录号l32019)及其相应的编码蛋白质具有高同一性。值得注意的是,第5页的第一段强调了具有高度同一性的cry蛋白如何可对个体昆虫物种显示不同量地的毒性。还描述了用cryet4基因转化的bt菌株。wo 95/04146a2的实施例4表明纯化的cryet4蛋白显示出针对草地贪夜蛾的活性。然而,从中分离出该基因的bt菌株的活性及其发酵后获得的晶体组分并未写明,这一信息对于确定其是否构成用于控制草地贪夜蛾的新的、天然和替代bt菌株至关重要。
25.美国专利申请us 2009/005306公开了一种编码蛋白质cry2adshi的bt基因,其具有具有针对几种昆虫害虫的杀虫活性。然而,上述美国申请的表1中显示的结果表明,该蛋白质针对草地贪夜蛾没有活性。这在实施例3的表3中得到进一步证实,表3表明针对秋季粘
虫(草地贪夜蛾)检测到“无活性”。此外,所述美国申请中公开的发明的主要目的似乎是生成表达us 2009/005306中公开的基因的转基因植物,而不是鉴定具有针对草地贪夜蛾的高活性的新的天然bt菌株。
26.美国专利申请us 2018/127771 a1公开了几种源自bt菌株的蛋白质,这些蛋白质具有针对亚洲柑桔木虱(diphorina citri)的活性,亚洲柑桔木虱缩写为acp,属于不同于草地贪夜蛾(鳞翅目)的目(半翅目)。事实上,美国专利申请中甚至没有提到草地贪夜蛾。实施例1说明,该研究的总体目标是鉴定针对acp具有毒性的bt晶体毒素。此外,已鉴定的毒素旨在通过在转基因植物中的实施来控制植物y半翅目昆虫。
27.国际专利申请wo 2013/134734 a2公开了两种与已经提到的基因cry1ab24和cry1ja1编码的蛋白质具有高度同一性的蛋白质。然而,它们不是发明的主要目的。wo 2013/134734 a2重点关注由几种多肽(其中一些具有杀虫特性)组合而成的多肽,以及它们在转基因植物中表达的改善。显示了与草地贪夜蛾控制相关的叶面生物测定法,采用喷雾干燥粉末进行,粉末中含有源自bt蛋白的肽和抑制剂半胱氨酸结肽。但是,只是简单地提到了草地贪夜蛾作为控制所感兴趣的昆虫之一,而没有提供任何针对该物种的测定法。
28.因此,尽管在bt菌株中发现了遗传多样性,但与仅基于遗传和分子特征很难找到针对特定物种(如草地贪夜蛾)具有高活性的bt菌株一致,用于控制鳞翅目昆虫的市售生物杀虫剂仅依赖于来自kurstaki和aizawai血清型的少数。作为最广泛使用的产品之一,采用了由来自菌株abts-351(血清型kurstaki)的孢子和晶体蛋白的组合而制成的活性成分。abts-351菌株产生的晶体包含蛋白质cry1aa、cry1ab、cry1ac和cry2aa,发现它们的相对比例取决于培养基的组成和生长条件。以及其他基于abts-351菌株的商业配方产品可有效控制属于鳞翅目的广谱幼虫物种,包括来自棉铃虫属(heliothis)、菜蛾属(plutella)和花翅小卷蛾属(lobesia)的物种。然而,普遍认为这些产品针对属于灰翅夜蛾属(spodoptera)的昆虫物种效果不佳,包括草地贪夜蛾、海灰翅夜蛾(s.littoralis)、甜菜夜蛾(s.exigua)等。由于这些物种损害作物,对经济造成严重负面影响,因此abts-1857(血清型aizawai)被选为杀虫毒素的新来源。是目前最臭名昭著的基于abts-1857菌株的商用杀虫剂,携带蛋白质cry1ca、cry1da、cry1aa和cry1ab。尽管不能去除由于其他蛋白质的存在而产生的协同效应,但前两种蛋白质是对灰翅夜蛾有效的主要杀虫剂。
29.尽管市场上存在基于bt的产品,但昆虫种群已经对它们产生了不同程度的抗性,寻找用于害虫控制的新bt菌株的努力正在增加。需要鉴定具有更高杀虫效力和更广泛宿主范围的新bt菌株。特别是出于本发明的目的,特别想要找到并鉴定具有针对草地贪夜蛾的改进杀虫活性的新bt菌株。优选地,新的bt菌株应该是从环境样品中分离的天然菌株。同样优选地,为了促进菌株或基于菌株的产品的使用,新的bt菌株不应表达β-外毒素,以避免多种物种的细胞死亡风险。
30.本发明提供了这种问题的解决方案。
31.发明概述
32.本发明涉及一种新的苏云金芽孢杆菌菌株(本发明的苏云金芽孢杆菌菌株),其特征在于是以登录号cect 9753保藏于colecci
ó
n de cultivos tipo(cect)的菌株svbs-1801。
33.第二方面,本发明还涉及一种制备本发明苏云金芽孢杆菌菌株sbvs-1801的孢子和晶体的混合物的方法,包括以下步骤:
34.a)培养sbvs-1801细菌,直到超过95%的细菌细胞已经发生孢子形成、已经裂解并将其晶体和孢子释放到培养基中;
35.b)通过引起晶体和孢子从它们悬浮的培养基中沉降来纯化晶体和孢子,优选地通过离心,并收集沉淀物;
36.c)任选地,用蛋白酶抑制溶液洗涤沉淀物并再次引起晶体和孢子的沉降;
37.d)储存纯化的晶体和孢子:
38.a.在室温,在已经将步骤b)或c)中获得的沉淀物冻干后,或
39.b.在-18
°
至-20℃或更低的温度,在已经将b)或c)中获得的沉淀物重新悬浮在水或水溶液中后。
40.在第三方面,本发明涉及一种获得本发明的苏云金芽孢杆菌菌株svbs-1801的纯化的晶体(伴孢晶体)的方法,该方法包括以下步骤:
41.i)用nacl 1m洗涤svbs-1801的晶体和孢子的混合物,并以9000g离心10min,
42.ii)收集团块并将其重新悬浮在pbs中,
43.iii)向步骤ii)中获得的悬浮液中加入己烷并涡旋;
44.iv)将iii)的悬浮液以6000g,4℃,离心10min,
45.v)重复步骤ii)至iv)至少3次,
46.vi)收集在v)中获得的晶体团块并在冷蒸馏水中洗涤三次。
47.晶体和孢子(用于获得纯化的晶体)的混合物可以是用于制备本发明苏云金芽孢杆菌菌株sbvs-1801孢子和晶体混合物的方法的步骤b)或c)后得到的混合物,这是本发明的第二方面。
48.在第四方面,本发明还涉及一种组合物,其包含以下组中的至少一种:本发明的苏云金芽孢杆菌菌株的a)营养细胞,b)含有孢子的细菌细胞,c)孢子,d)晶体(伴孢晶体),e)孢子、晶体和晶体蛋白的混合物,f)不包含在晶体中的至少一种cry蛋白,g)至少一种vip蛋白,或其组合,其中组合物(如果a)至e)中定义的组成员不存在)i)当至少存在cry蛋白时,至少存在cry1ja1样蛋白(seq id no:8),和/或ii)vip1ca1(seq id no:14)、vip2ac1(seq id no:16)和vip3af3(seq id no:18)组的所有三种蛋白将存在于组合物中。这种组合物被认为是本发明的组合物。任选地,在与任何其他相容的实施方案中,组合物还可以另外包括在执行用于制备本发明的苏云金芽孢杆菌菌株的孢子和晶体的混合物的方法的步骤a)和b)之后获得的部分上清液。本发明组合物的优选实施方案是其中组合物包含至少以下任一项的那些:a)孢子和晶体的混合物;b)通过执行本发明一方面的制备菌株sbvs-1801的孢子和晶体混合物的方法步骤a)和b)获得的孢子、晶体和部分上清液的混合物;c)晶体;d)通过执行本发明一个方面的制备菌株sbvs-1801的孢子和晶体的混合物的方法步骤a)和b)获得的晶体和部分上清液。同样优选的是那些至少包含菌株sbvs-1801的孢子、晶体和晶体蛋白的混合物的组合物(它们是用于从bt菌株的培养基制备杀虫剂的组合物的典型组分)。还优选的是那些包含不包含在晶体中的cry蛋白的组合物,并且其中该组合物包含至少seq id no:4(cry1da3)、seq id no:6(cry1ea7)和seq id no:8(cry1ja1-like)的组中的所有cry蛋白(不包含在晶体中)。本发明的组合物还可另外包含至少一种农业上可接受的赋形剂。
49.本发明的另一方面是本发明的苏云金芽孢杆菌菌株和/或本发明的组合物作为杀虫剂或用于制备杀虫剂的用途。优选地,它将用于防治草地贪夜蛾物种的昆虫害虫。还优选地,该组合物将用于保护植物免受害虫侵害。植物可以是玉米植物,如在实施例8中,或任何其他植物,例如棉花(另一种植物,其中草地贪夜蛾是重要损害的原因,并且之前已经使用表达cry蛋白的转基因植物)、水稻、高粱、甘蔗等。
50.本发明的另一方面是用于鉴定本发明的苏云金芽孢杆菌菌株存在的方法,其包括确定以下任一者的步骤。
51.a)该菌株的基因组包括:
52.i)其序列选自seq id no:1或seq id no:7,或其组合的至少一个基因或dna区域;和/或
53.ii)seq id no:1、seq id no:3、seq id no:5、seq id no:7、seq id no:9和seq id no:11的组中的至少所有基因或dna区域,和/或
54.iii)seq id no:13、seq id no:15和seq id no:17的组中的至少所有基因或dna区域;和/或
55.iv)seq id no:1、seq id no:3、seq id no:5、seq id no:7、seq id no:9、seq id no:11、seq id no:13、seq id no:15和seq id no:17的组中的所有基因或dna区域;
56.或者
57.b)该菌株表达
58.i)其氨基酸序列选自seq id no:2或seq id no:8,或其组合的组的至少一种多肽,和/或
59.ii)其氨基酸序列为seq id no:2、seq id no:4、seq id no:6、seq id no:8、seq id no:10或seq id no:12的多肽的组中的至少所有多肽,或
60.iii)其氨基酸序列为seq id no:14、seq id no:16或seq id no:18的多肽的组中的至少所有多肽,
61.iv)其氨基酸序列为seq id no:2,seq id no:4,seq id no:6,seq id no:8,seq id no:10,seq id no:12、seq id no:14、seq id no:16或seq id no:18的多肽的组中的所有多肽。
62.还可以认为本发明的另一方面是本发明的dna分子,即,包含选自下组的核苷酸序列的核酸分子:
63.a)选自seq id no:1和seq id no:7的组的核苷酸序列;
64.b)编码包含seq id no:2和seq id no:8的组的氨基酸序列的多肽的核苷酸序列;
65.c)编码与seq id no:2和seq id no:8的组的氨基酸序列具有至少99%同一性且具有杀虫活性(例如,杀线虫和/或杀虫活性)的多肽的核苷酸序列。
66.因此,还可以认为本发明的另一方面(与前一方面紧密相连)是包含本发明dna分子的表达载体。
67.同样的,还可以认为另一方面是包含本发明dna分子的重组病毒、细菌、真菌或酵母、植物细胞或整个植物或植物的部分。dna分子将优选地操作性地连接到启动子,并且任选地连接到一个或多个附加控制元件(例如增强子),并且其整合到其基因组中。
68.附图简要说明
id no:8、seq id no:10或seq id no:12的多肽的组的至少所有多肽的表达(分别由seq id no:1、seq id no:3、seq id no:5、seq id no:7、seq id no:9和seq id no:11的基因的组编码的cry蛋白的组,其同时存在于bt基因组中是本发明的bt菌株的特征),和/或其氨基酸序列为seq id no:14、seq id no:16或seq id no:18的多肽的组的至少所有多肽的表达(分别由seq id no:13、seq id no:15和seq id no:17的基因的组编码的vip蛋白的组,其同时存在于bt基因组中是本发明的bt菌株的特征)或其氨基酸序列为seq id no:2、seq id no:4、seq id no:6、seq id no:8、seq id no:10、seq id no:12、seq id no:14、seq id no:16或seq id no:18的多肽的组的所有多肽,可用于鉴定本发明的bt菌株svbs-1801的存在。
82.如在本技术中所使用,“鉴定存在”可理解为确定样品中bt菌株svbs-1801的存在或鉴定特定菌株为本发明的bt菌株svbs-1801。由于基因cry1ab24样和cry1ja1样是新的并且甚至与最相似的已知基因(cry1ab24和cry1ja1)不同,并且之前没有描述过它们同时存在于bt菌株的基因组,它们还可用于定义本发明的新菌株,特别是cry1ja1样或由其编码的蛋白质,因为其表现出针对草地贪夜蛾的活性。如上所述,之前没有报道过bt菌株同时存在seq id no:1、seq id no:3、seq id no:5、seq id no:7、seq id no:9和seq id no:11的cry基因的组,也没有同时存在seq id no:13、seq id no:15或seq id no:17的vip基因的组,使得在其所有基因组中所述两组基因中的任何,或同时存在,也可用于通过所述菌株的特征和不明显或预期在特别是非实验室创建但从环境源分离的菌株中会发现的特征来定义本发明的bt菌株。这一关于通过可发现的cry和vip基因的特定组合来定义菌株的实用性的评论可以扩展到由所述基因编码的蛋白质。
83.于2018年11月14日保藏在colecci
ónꢀꢀ
de cultivos tipo(cect)并获得登录号:cect 9753的菌株svbs-1801是本发明的一方面。
84.关于在svbs-1801bt菌株中发现的cry和/或vip基因编码的蛋白质,所有已鉴定的cry蛋白质可存在于晶体中,而vip蛋白应在生长阶段分泌后在生长培养基的上清液中发现。
85.值得注意的是,该菌株的毒性谱包括草地贪夜蛾,目前市场上的生物制品对其控制效果并不理想。以前没有知识表明显示上述基因组合的菌株可以具有针对草地贪夜蛾的活性,并特别是,以前没有知识表明具有这种特定基因组合的菌株具有针对草地贪夜蛾的活性高于用于制备目前用于针对这种害虫的商业杀虫剂的菌株,如下面的实施例所示。这一点尤其值得注意,因为根据遗传和分子特征发现对特定物种具有高活性的bt菌株是一项困难且不明显的任务。如上所述,这些困难不仅源于具有高度同源性的cry蛋白可能表现出针对害虫的不同活性和特异性的事实,而且尤其是因为无法预测bt菌株产生的不同毒素之间可能会发生哪些组合效应,其类型(协同或拮抗),以及组合效应可能对bt菌株及其发酵产物的杀虫活性的影响程度。事实上,本技术公开了由svbs-1801产生的三种cry蛋白质之间的协同效应,该协同效应导致相对于由所述蛋白质中的每一种的活性的简单组合产生的预期活性的活性增加,这是一种完全出乎意料的组合效应。
86.因此,本发明提供了一种新的bt菌株svbs-1801,该菌株从环境来源分离并且缺乏β-外毒素的表达,可用于控制草地贪夜蛾害虫。本发明还包括制备包含svbs-1801的晶体和孢子的混合物的组合物的方法(通常用于制备商用杀虫剂的组合物种类)、制备菌株的纯化
的晶体的方法,包含由菌株产生或由其生长产生的不同组分的组合物:晶体和孢子的混合物,其中可能还存在其他组分,例如不包含在伴胞晶体中的cry蛋白、营养细菌或vip蛋白、具有不包括在晶体中的cry蛋白的组合物、纯化的晶体或甚至具有纯化的晶体或具有晶体与孢子的混合物的组合物,其中,当培养菌株的液体培养基离心以将晶体和孢子从细菌生长的液体中分离出来时,获得部分上清液。本发明的范围还包括使用该菌株以及包含该菌株或者包含该菌株产生的或其在液体发酵培养基中生长产生的元素的组合物作为杀虫剂或用于制备杀虫剂,尤其是针对草地贪夜蛾的杀虫剂。
87.本发明还涵盖seq id no:1和seq id no:7的新基因、由seq id no:1和seq id no:7编码的蛋白质(即seq id no:2和seq id no:8的蛋白质),以及包含与seq id no:2或seq id no:8具有至少99%同一性的多肽且具有杀虫特性的蛋白质。本发明还涵盖可衍生自svbs-1801的所有突变菌株,尤其是使用所述基因获得的重组细菌菌株(例如,可按照实施例4中所述的程序获得)或甚至酵母、真菌或甚至植物细胞的重组细胞,以及允许在其基因组中表达所述蛋白质或插入seq id no:1或seq id no:7的基因的载体,例如质粒或重组病毒。在植物细胞的情况下,不仅插入seq id no:7的基因,而且插入seq id no:3和seq id no:5的基因,或者,引入所有三个基因的表达载体可能是感兴趣的,因为当单独测试时,由上述三个基因表达的蛋白质显示出针对草地贪夜蛾的活性,但当组合测试时,它们也显示出协同效应。
88.苏云金杆菌菌株svbs-1801具有以下特征:
89.生长:苏云金芽孢杆菌svbs-1801优选在28-30℃的富盐培养基(例如本发明实施例1中描述和使用的ccy培养基)中生长。在这种条件下,孢子萌发并产生双锥晶体,其可以在光学显微镜下观察到(图1)。当在固体培养基(ccy或luria-bertani(lb)肉汤中培养,补充有琼脂)中生长时,菌落的形态与其他bt菌株非常相似,圆形,边缘不规则,外观呈蜡状。当在适当的培养基:富盐培养基,如ccy培养基和温度条件(温度可以在15-45℃之间变化,但最佳范围为28至30℃)中使用时,svbs-1801可以工业规模生长。还需要连续搅拌(例如220rpm)。
90.没有外毒素。如上所述,一些bt菌株可产生分泌性的毒素,β-外毒素,其可以损害rna聚合酶,导致多种物种的细胞死亡,并阻止使用该菌株或基于其的产品作为杀虫剂。如实施例2所示,svbs-1801在发酵过程中不产生β-外毒素。因此,认为它具有广泛而不特定的作用范围。
91.bt毒素/晶体蛋白:由本发明菌株svbs-1801合成的cry蛋白在孢子形成阶段聚集和结晶,产生伴胞晶体,在本技术中简称为“晶体”。它们在sds-page凝胶中的迁移模式可以描述为两条相互接近的带,大小约为120-135kda(cry1ab、cry1da3、cry1ea7和cry1ja1)(图3)。来自基于菌株abts-351(血清型kurstaki的bt菌株)的商业产品的晶体蛋白,(cry1aa,cry1ab,cry1ac y cry2aa)和基于abts-1857株(bt血清型aizawai)的商业产品的晶体蛋白,(cry1aa、cry1ab、cry1ca和cry1da)迁移类似,带的大小与菌株svbs-1801的相当(130-135);然而,总蛋白质库为每个菌株产生独特的迁移模式。众所周知,另外产生一条约70-75kda的条带,对应于蛋白质cry2aa。
92.杀虫能力。如上所述,值得注意的是,svbs-1801晶体蛋白比其和
对应物更有效5.7倍和1235.4倍地针对草地贪夜蛾,如本技术的实施例中所示。因此,基于svbs-1801菌株的产品(即,包含营养细胞、含有孢子的细菌细胞、孢子、晶体、cry蛋白或vip蛋白或其组合的组合物)可用作杀虫剂或用于制备杀虫剂。尤其优选的是,用于制备该杀虫剂组合物的活性成分是孢子和晶体蛋白的组合,其是在对培养物内容物(理解例如培养基和生长的细菌细胞、溶解细胞的残余物和细菌产物,如孢子、晶体、细菌蛋白质(不构成晶体一部分)和其他细菌产生的化合物)进行沉淀处理(优选通过离心)后获得的,其中sbvs-1801细菌一直生长直到形成孢子,其晶体和孢子溶解和释放,如实施例1所示:相当于用于制备基于bt的商业杀虫剂产品(如或)的活性成分。
93.此类活性成分(ai)(菌株svbs-1801)以及通常包括在其制备物中的晶体和孢子的混合物,可如实施例1所述或通过任何类似方法获得。作为本发明的一个方面,方法包括以下步骤:
94.a)培养sbvs-1801细菌,直到超过50%的细菌细胞已经发生孢子形成、已经裂解并将其晶体和孢子释放到培养基中;
95.b)通过引起晶体和孢子从它们悬浮的培养基中沉降来纯化晶体和孢子,优选地通过离心,并收集沉淀物;
96.c)任选地,用蛋白酶抑制溶液洗涤沉淀物并再次引起晶体和孢子的沉降;
97.d)储存纯化的晶体和孢子:
98.a.在室温,在已经将步骤b)或c)中获得的沉淀物冻干后,或
99.b.在-18
°
至-20℃或更低的间隔的温度,在已经将b)或c)中获得的沉淀物重新悬浮在水或水溶液中后。
100.此类ai包含晶体和孢子的混合物,其可重新配制为浓缩液体悬浮液形式的组合物或可湿性粉末,该组合物还可包含一种或多种赋形剂或添加剂,尤其是通常用于农业领域的赋形剂或添加剂。任何这种重新配制的形式都可以应用于田间,并可以用作杀虫剂,优选用于保护植物。如上文所述,它可用于控制害虫,例如草地贪夜蛾。
101.但值得注意的是,菌株svbs-1801的优点之一是,本发明的发明人发现,具有由本发明的菌株svbs-1801产生的不同元件的不同组合物也具有针对草地贪夜蛾的活性,在一些处理中观察到了活性,甚至高于通过上述本发明方法获得的晶体和孢子的混合物的活性。如lc
50
值所示,将通过相同方法获得的部分上清液添加到晶体和孢子的混合物中显著提高活性(见表5)。使用不含孢子的纯化的晶体进行的测定法也显示就晶体和孢子的混合物而言活性增加,当晶体与相同上清液混合时,活性甚至更高(lc
50
值更低)。这些结果有助于使用不同的组成来制备ai,每种具有从菌株svbs-1801产生或来源的不同组分,即:a)孢子和晶体的混合物;b)通过执行制备菌株sbvs-1801的孢子和晶体的混合物的方法步骤a)和b)获得的孢子、晶体和部分上清液的混合物,这是本发明的一个方面;c)晶体;d)通过执行制备菌株sbvs-1801的孢子和晶体的混合物的方法步骤a)和b)获得的晶体和部分上清液,这是本发明的一个方面。
102.纯化的晶体可通过实施例8中使用的方法制备,这也是本发明的一个方面,但也可通过本领域技术人员已知的任何其他方法制备,例如蔗糖梯度上的超速离心(thomas和ellar,1983),该方法通常为不连续蔗糖梯度(79-67%),其中晶体被截留在相间,并且孢子
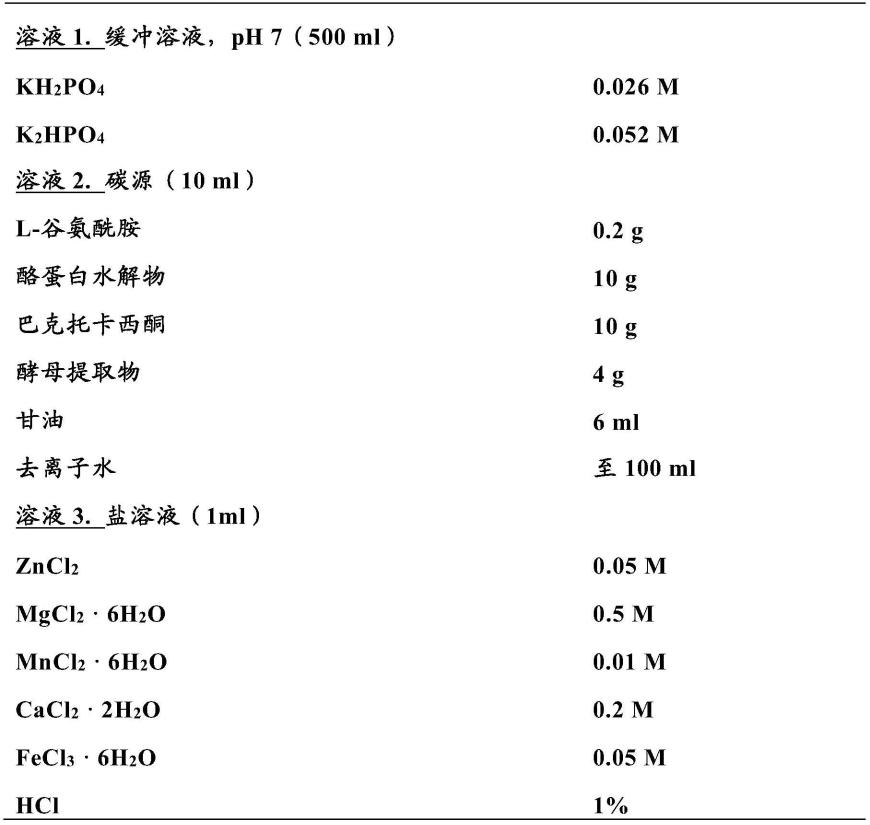
可以从团块中收集。如果如本技术实施例8所述,通过制备物或组合物样品的共焦显微镜,或者通过类似方法检查孢子的存在,观察不到孢子,则晶体制备物或包含sbvs-1801晶体的组合物将被视为不含孢子或基本不含孢子。
103.当组合物中不需要孢子时,可通过添加杀菌剂来代替晶体的纯化。杀菌剂也可添加到具有纯化的晶体的组合物中,以通过消除晶体纯化后可能残留的任何可能的孢子来保证没有孢子。可通过将纯化的晶体悬浮液的稀释液接种到营养琼脂平板中,并验证没有菌落生长,从而检查不存在孢子,并因此晶体完全没有孢子且其纯度最高。如果添加,杀菌剂将优选地选择为与本发明的菌株和组合物的生态特性兼容。
104.在实施例8中获得的结果,其中孢子和晶体的混合物以及纯化的晶体都显示出针对草地贪夜蛾的活性,表明可以设想用所述元素的不同比例(例如,50:50或80:20晶体:孢子,或任何其他)制备孢子和晶体的混合物,可以将它们全部用于制备杀虫剂的活性成分。因此,例如,必须考虑在本发明中包含具有孢子和晶体的那些组合物,其中晶体是通过产生结晶的方法得到的,不是完全提纯的,但它们仅是,例如,80%纯化的(或任何其他百分比,如85%、90%或95%纯化),因此,该制备物实际上将是也包含在本发明范围内的晶体和孢子的混合物。
105.同样在实施例8中,但在第8.3节中,显示了关于由本发明菌株合成的单个cry蛋白的毒性的测定法。根据获得的结果(见表6和表7),菌株svbs-1801的伴胞晶体的杀虫活性可归因于cry蛋白cry1da3、cry1ea7和cry1ja1样,当分别对针对草地贪夜蛾幼虫进行测试时,这些蛋白具有毒性。蛋白质cry1ja1样的活性显著,因为其由本技术中公开的一个基因cry1ja1样(seq id no:7)编码,如上文所述,该基因与已知基因cry1ja1存在一定差异。特别值得注意的是,事实上使用三种蛋白质的混合物进行的测定显示,对于三种蛋白质中任何一种的相同浓度,活性增加,表明三种蛋白质的混合物中的组合协同效应,这已在实施例8的同一部分中得到证实。从先前发表的文献的教导中未预期到所确定的协同效应,并支持没有预期具有svbs-1801基因组合的菌株能够具有针对草地贪夜蛾的高活性(尤其是针对幼虫),其活性高于目前使用的最有效的商用杀虫剂(针对该菌株)(见实施例9至11)和capalbo等人,(2001)描述的最有效菌株,在某种程度上缺乏对该文章中报告的测定法中用于测量lc
50
的方法的解释,可进行比较。
106.在用cry蛋白的测定法中发现的结果使得也可以考虑包含蛋白cry1da3、cry1ea7和cry1ja1样(分别为seq id no:4、seq id no:6和seq id no.8)的混合物的组合物作为本发明的组合物的可能实施方案,特别是如果它们以等摩尔比例。此类组合物还可用作杀虫剂或用于制备针对草地贪夜蛾的杀虫剂。
107.杀虫基因。如表2和表3所示,svbs-1801的基因组(染色体 质粒)分别包含至少6个不同的cry基因和3个不同的vip基因。两个cry基因(cry1ab24样和cry1ja1样)与编码序列公开的最相似参考基因cry1ab24(genbank登录号hq439778)和cry1ja1(genbank登录号l32019)不完全相同。因此,确定至少一个不同于其他已知bt基因的所述基因的存在可足以鉴定菌株svbs-1801。为此,值得注意的是,基因cry1ab24样呈现的编码序列(seq id no:1)比cry1ab24的编码序列短78个核苷酸(见图4),这导致由cry1ab24编码的蛋白质在由seq id no:1编码的多肽的793和794位之间长26个氨基酸(见图6的比对)。除了seq id no:1的906位置存在t而不是c之外,这种大小和序列的差异有助于区分cry1ab24样和cry1ab24
(genbank登录号hq439778)。关于cry1ja1样,如图5和图7所示,其编码序列在长度上与cry1ja1的编码序列(genbank登录号hq439784)相同,但两个序列之间有两个不同的核苷酸。参考基因cry1ja1分别在seq id no:7的位置987处呈现t而不是c,在seq id no:7的位置2345处呈现g而不是a。这些变异可用于区分属于svbs-1801菌株的序列。
108.svbs-1801菌株的鉴定可通过确定本发明人在svbs-1801中发现的六个cry基因的组的存在,如表2所示(cry1ab24、cry1da3、cry1ea7、cry1ja1、cry1nb1、cry2ad1)来完成,或通过本发明人在svbs-1801中发现的三个vip基因的组的存在,如表3所示(vip1ca1、vip2ac1、vip3af3)来完成,或更优选地,通过同时存在至少九个杀虫基因,六个cry基因的组和三个vip基因的组来完成。对所选基因的存在进行鉴定可如下完成:如实施例3所示,可通过完整的dna测序、重叠群组装和组装的重叠群的基因预测,或通过任何其他方法;类似于实施例3中使用的方法,或通过pcr扩增基因片段,如实施例4中所述,使用例如相同的引物对。基因的存在需要与数据库btnomenclature(http://www.btnomenclature.info,该数据库包含的苏云金芽孢杆菌基因也存在于genbank中,并且可以使用相同的登录号访问)中的基因序列至少95%一致,因为它可能对应,但优选的是,当基因cry1ab24或基因cry1ja1是用于将菌株鉴定为svbs-1801菌株的基因之一时,当相应dna片段的序列分别为seq id no:1或seq id no:7中的一个,即在菌株svbs-1801中发现的所述基因的变体时,基因cry1ab24或基因cry1ja的存在被认为是阳性的。
109.基因及其用途。菌株svbs-1801可作为基因源,在医药、兽医、生物技术等其他领域有着广泛的应用,并且它们可以随着它们的生物学特性转移到其他生物体中用于不同的目的。因此,编码序列为seq id no:1、seq id no:3、seq id no:5、seq id no:7、seq id no:9、seq id no:11、seq id no:13、seq id no:15和seq id no:17的基因本身就是感兴趣的产物,因为它们编码杀虫蛋白。例如,可以使用所述基因(或组成它们的dna片段或区域),获得具有昆虫抗性的转基因植物(类似于国际专利申请wo1995/024493或美国专利申请公开us20080016596a1等文献中描述的植物)或线虫抗性的转基因植物(类似于国际专利申请wo2010/027793等文献中描述的植物),例如,通过使用本领域技术人员熟知的此类文献中描述的技术。相同的dna片段或区域可用于获得,例如,非人类转基因动物或重组细胞(动物、植物、真菌或细菌细胞),前提是它们是构建体的部分,在该构建体中它们与适当的控制元件(启动子、增强子
…
)操作性连接,取决于需要它们得到表达的条件,或者对于转基因植物或动物,也取决于需要它们表达的组织。此类控制元件为本领域技术人员所熟知。所述构建物可整合于基因组中,或尤其对于细菌重组细胞而言,可为表达载体(例如质粒)的部分,所述表达载体可通过本领域技术人员熟知的方法引入细胞中。这种转化的细胞或高等生物可用于产生对线虫或昆虫等害虫(例如,灰翅夜蛾属昆虫,如草地贪夜蛾)具有抗性的植物,或可用于任何其他目的(医学、兽医、一般研究
…
)。
110.因此,必须将其视为包含在本发明范围内的任何核酸分子,该核酸分子包含其核苷酸序列为:a)本发明任何新基因的序列(seq id no:1和seq id no:7),b)编码多肽的任何核苷酸序列,该多肽包含为所述基因编码的任何蛋白质的氨基酸序列(seq id no:2和seq id no:8的氨基酸序列,它们是本发明的蛋白质),以及也编码包含seq id no:2或seq id no:8的多肽片段序列的蛋白质但由于遗传密码的兼并性质不同于seq id no:1和seq id no:7的核苷酸序列)或包含在序列和活性上与本发明类似但不能分类为不同蛋白质的
多肽片段的蛋白质,即:c)编码多肽的核苷酸序列,所述多肽具有与seq id no:2的氨基酸序列具有至少98,5%(或至少99%、99,5%、99,90%、99,95%或99,99%)的同一性的序列片段,或与seq id no:8的氨基酸序列具有至少99,85%(或至少99,90%、99,95%或99,99%)的同一性且具有杀害虫活性(杀虫、杀线虫或两者兼有)。杀害虫活性可通过多肽本身或其裂解形式表现,如通常发生在cry蛋白中,其昆虫毒性活性在蛋白质在昆虫肠道中裂解成两个片段后出现。
111.因此,含有本发明的任何此类核酸分子的构建体,尤其是表达载体(例如质粒或重组病毒)(其中本发明的核酸分子操作性地连接到启动子和任选地一个或多个控制序列,取决于需要蛋白质得到表达的条件和/或取决于需要的表达水平而选择),也都包含在本发明的范围内。
112.本发明范围的另一部分是包含作为异源dna片段的本发明核酸分子或包含其表达载体的任何细胞(细菌、真菌、酵母或植物细胞)。核酸片段可以整合在基因组中(或者,在细菌的情况下,整合在细菌染色体中),或者可以是其他核酸元件,例如质粒或非整合病毒的部分。细菌细胞甚至可以来自另一种苏云金芽孢杆菌菌株,例如,一个非天然包含以下一个或多个基因的菌株:seq id no:1、seq id no:3、seq id no:5、seq id no:7、seq id no:9、seq id no:11、seq id no:13、seq id no:15或seq id no:17,其杀虫或杀线虫活性可通过表达一种或多种与序列seq id no.2和seq id no:8匹配的蛋白质而得到增加或修改。这可通过使用盒来转化菌株而实现,该盒包含seq id no:1或seq id no:7的dna片段(或编码序列seq id no:2或seq id no:8的多肽的任何其他核苷酸序列)操作性地连接到允许该多肽在菌株中组成型或在所选条件下表达的启动子。此外,那些重组苏云金芽孢杆菌细胞,在其基因组(包括染色体和质粒)修饰后,包含至少一组基因,其编码区匹配序列seq id no:1、seq id no:3、seq id no:5、seq id no:7、seq id no:9和seq id no:11或seq id no:13、seq id no:15和seq id no:17,或者包含这两组基因,因为它将成为具有菌株svbs-1801的特征基因组的bt菌株,这都包括在本发明中。
113.因此,包含本发明的dna分子的任何植物将包含在本发明的范围内,特别是当dna分子或片段是构建体的部分时,其中它与一个或多个控制元件(启动子、增强子、终止序列
…
)操作性地连接,所述控制元件允许或促进dna分子编码的多肽在植物或动物的一个或多个组织中组成型或在一定条件下表达,包括植物或动物发育过程中的不同时期。对于转基因植物,(例如shah等人,2015所综述的),通常使用组成型启动子,如花椰菜花叶病毒(camv)35s,但通过使用可诱导启动子来限制所需转基因基因的表达也很有趣,如番茄hmgr2基因的伤口诱导启动子、pr-1a启动子(由病原体感染和苯并噻二唑等化学诱导子诱导)或玉米in2-2(可诱导基因2-2)启动子(由苯磺酰胺类安全剂诱导,作为提高植物耐除草剂性的农用化学品)。其他感兴趣的可诱导元件可以是空间上调节在不同部位的基因表达的顺式作用调节序列,或由菜豆蛋白基因启动子编码的两个上游激活序列,uas1(-295至-109)和uas2(-468至-391)。此外,针对特定组织的靶向表达对于增值作物的未来发展变得越来越重要,因此,如acc氧化酶基因的启动子(从转录开始范围为21966到21159bp)能够驱动基因在水果(如番茄果实)中的tipening-特异性表达,正如shah等人,2015所综述的那样。其他不同的启动子和控制元件可在所述版本和本领域技术人员可得的其他文献中找到。
114.现在将结合下面包括的实施例和附图进一步详细地解释本发明。
实施例
115.下节提供实施例,说明本发明可用于促进svbs-1801工业开发的过程。
116.实施例1-bt菌株svbs-1801的分离与生长
117.1.1.svbs-1801菌株的分离
118.从来自巴达霍兹市郊区的农业土壤样品(用于该市城市化“圣加布里埃尔住宅”花园的建设)中获得svbs-1801菌株。上述土壤样品于2016年12月采集,并在冷藏条件(4℃)下保存,直至处理完成(2017年1月)。为了进行svbs菌株的分离和鉴定,在去除第一层后,从土壤中收集样品,与蒸馏水混合并在70℃下加热15min。将10倍稀释的体积(15μl)涂布到ccy培养基上并在28℃下培养72h。通过相差显微镜检查阳性bt菌落(iriarte等人,1998)。
119.1.2.在促进孢子形成的培养基中生长
120.svbs-1801样品培养在1l ccy培养基中(stewart等人,1981)。ccy是促进孢子形成的最小培养基,最终导致晶体和孢子的形成(图1)。通过按适当比例混合缓冲成分(溶液1)和碳源成分(溶液2)而制备。混合完成后,在高压灭菌器内对培养基进行灭菌,并补充1ml先前过滤的盐溶液(溶液3)。
121.表1.ccy培养基(stewart等人,1981)
[0122][0123]
通过将100μl的svbs-1801过夜(on)预接种液转移到ccy培养基(1:l)中启动细菌生长。细菌在30℃和220rpm下培养48-72小时。95%的svbs-1801细胞(已经经过孢子形成过程)裂解并释放其晶体和孢子后,发酵过程结束。为验证已达到该阶段,应在光学显微镜下分析1ml培养物样品。
[0124]
为了纯化已产生的晶体和孢子,将培养物的内容物离心,然后用nacl 1m/edta 10mm的10%溶液洗涤沉淀物。该溶液用于防止蛋白酶降解蛋白质晶体。随后,再次离心培养物,并将团块冻干并在室温下储存,直到配制或重新悬浮在kcl 10mm(10ml/l初始培养物)中,并在-20℃下储存。
[0125]
实施例2-菌株svbs-1801的特性:确定菌株svbs-1801是否产生β-外毒素的程序
[0126]
为了确定特定的bt菌株是否为β-外毒素阳性,应分析生长培养基的上清液。为此,采用了hern
á
ndez等人,2003采用的程序。因此,将bt svbs-1801和hd-2菌株(用作阳性对照)的集落在30℃和220rpm在10ml ccy中培养48小时。接下来,培养物在9000xg离心10min,将其上清液转移到新试管中,并在120℃和1atm在高压灭菌器中灭菌20min。
[0127]
为了检测β-外毒素的存在,进行hplc。kh2po4(50mm,ph3)以2ml/min的流速构成流动相,并与灭菌的上清液一起通过0.45μm的尼龙膜进行过滤。在μ
–
bondapak c18柱中,在25-30℃和260nm下分析20μl上清液。如果存在β-外毒素,则应出现两个清晰的峰,表明其磷
酸化和去磷酸化形式。在本情况下,如图2所示,未检测到此类峰。
[0128]
实施例3-菌株svbs-1801的特征:dna提取、序列和基因组序列处理
[0129]
3.1.dna提取、序列和基因组序列处理
[0130]
使用组分并按照基因组dna纯化试剂盒(promega,madison,wi,usa)提供的革兰氏阳性细菌dna分离方案来分离svbs-1801的总dna。提取的dna样品用于制备dna文库,并随后在genomics research hub laboratory(cardiff school of biosciences,uk)使用illumina nextseq500测序仪进行测序。
[0131]
使用clc genomics workbench 10.1.1对生成的原始序列数据进行分析。通过删除低质量或不明确的读数,对原始序列数据进行质量过滤。丢弃短于50bp的读数。使用至少95bp读取重叠和95%同一性的严格标准重头组装读数。使用genemark.hmm原核3.25对组装的重叠群进行基因预测。使用blastp将获得的预测蛋白质序列与根据bt毒素列表建立的数据库http://www.btnomenclature.info(crickmore等人,2018)进行比较。
[0132]
这种生物信息学分析使本发明人能够鉴定杀虫基因的完整基因序列。无论如何,鉴定出一些cry和vip基因片段,如下所述。
[0133]
3.2.鉴定的杀虫基因
[0134]
表2包括在svbs-1801中鉴定的所有cry基因片段(编码序列)的完整列表,表明它们与之前在http://www.btnomenclature.info中描述的密切相关的杀虫基因的同一性程度(%id.)。还指出了比较中包括的已鉴定序列的覆盖率(%cov.),以及svbs-1801中鉴定的序列和数据库中存在的序列的核苷酸数量(ntd.)。
[0135]
表2.svbs-1801cry基因含量
[0136][0137]
*数据库:http://www.btnomenclature.info
[0138]
类似地,表3包含在svbs-1801菌株中鉴定的所有vip或sip基因片段的完整列表,表明它们与之前在http://www.btnomenclature.info中描述的密切相关的杀虫基因的同一性程度。
[0139]
表3.svbs-1801vip基因含量
[0140][0141]
*数据库:http://www.btnomenclature.info
[0142]
3.3.svbs-1801杀虫基因的分子特征
[0143]
使用blast(altschul等人,1990)分析svbs-1801cry基因的核苷酸序列,结果表明它们与先前描述的bt菌株的基因cry1ab24(hq439778)、cry1da3(hq439784)、cry1ea7(ay894137)、cry1ja1(l32109)、crynb1(kc156678)和cry2ad1(af200816)(http://www.btnomenclature.info)具有最高程度的同一性。类似地,发现svbs-1801vip基因序列更接近先前表征的vip1ca1(ay245547)、vip2ac1(ay245547)和vip3af3(hm117634)。如表2和3所示,除了cry1ab24和cry1ja1的情况之外,同一性的百分比是100%,但是同一性的百分比(接近95%)足够高,不考虑svbs-1801基因cry1ab24和cry1ja1样的不同基因(根据苏云金芽孢杆菌基因命名的常用规则)。
[0144]
实施例4-克隆:杀虫cry基因的pcr扩增
[0145]
根据bt菌株的测序结果,设计了包括用于克隆的限制性位点在内的特定寡核苷酸引物对,以扩增cry1ab24、cry1da3、cry1ea7、cry1ja1、cry1nb1和cry2ad1基因的开放阅读框(orf)。所述引物对的序列如下表4所示,其中第二列中的信息表明,对于每个引物,它是正向(fwd)引物还是反向(rev)引物,以及拟用于克隆的限制性内切酶。每个引物的下划线片段对应于酶裂解位点。
[0146]
表4:用于扩增已鉴定的svbs-1801基因的引物对
[0147]
[0148][0149]
将从菌株svbs-1801提取的dna用作模板,在含有10μl反应缓冲液5x hf、1μl dntp混合物100mm、1μl每个正向和反向引物10μm、0.5u高保真dna聚合酶(neb)和100ng总dna的50μl反应混合物中。pcr循环谱为:在c1000 touch热循环仪(bio-rad)中,1个初始变性循环为98℃持续30秒,30个扩增循环为98℃持续10秒、55℃持续1分钟和72℃持续3分钟30秒,然后在72℃持续10分钟的最终延伸。通过在tae缓冲液(40mm tris、20mm乙酸和1mm edta;ph 8)中的1%琼脂糖凝胶上以100v电泳30分钟分析pcr产物。
[0150]
使用nucleospin gel和pcr纯化试剂盒(macherey-nagel inc.,bethlehem,pa)从琼脂糖凝胶中纯化对应于cry1ab、cry1da、cry1ea、cry1ja、cry1nb和cry2ad cds的pcr产物,并钝端克隆到pjet载体中(clonejet pcr cloning kit,fermentas,canada)以获得pjet-protoxin构建体(pjet-1ab、pjet-1da、pjet-1ea、pjet-1ja、pjet-1nb和p-jet-2ad)。大肠杆菌xl1-blue的转化按照标准方案(sambrook等人,1989)进行,用于生产含有每个cry基因cds的重组质粒。通过pcr在20μl反应混合物中检查推定的阳性克隆,该反应混合物含有2μl反应缓冲液10xnh4、1μl mgcl
2 50mm、1μldntp混合物100mm、1μl每个正向和反向引物10μm和0,2μlbiotaq聚合物(bioline)。携带pjet构建体的大肠杆菌xl1-blue细胞在补充有氨苄青霉素(100mg/ml)的5ml lb肉汤中,在37℃和200rpm下培养过夜。根据制造商的方案,使用nucleospin质粒试剂盒(macherey-nagel inc.,bethlehem,pa)提取重组质粒,并通过使用每个基因的中间正向引物和pjet载体的反向引物(stabvida,caparica,portugal)的测序进行验证。使用geneious r8将cry基因核苷酸和蛋白质序列与genbank数据库中可用的毒素序列进行比对。图4和图6分别显示了cry1ab24样和cry1ab24基因的序列与其对应蛋白序列的比对;类似地,cry1ja1样和cry1ja1序列的比对如图5所示,而它们相应蛋白质序列的比对如图7所示。
[0151]
实施例4的重组pjet-原毒素构建体的消化用相应的限制酶组合(thermo scientific)进行,随后的纯化消化产物(之前在琼脂糖凝胶中分离)用nucleospin gel和pcr净化试剂盒(macherey-nagel inc.,bethlehem,pa)进行。之前用相应的限制酶消化的表达pstab载体(park等人,1998)用作插入片段的受体,使用rapid dna连接试剂盒(thermo scientific)以获得具有pstab原毒素构建体的重组质粒。如前所述,用连接产物转化大肠杆菌xl1-blue细胞,并在质粒提取前通过菌落pcr检查推定的阳性克隆。无结晶bt菌株bmb171(li等人,2000)用作在根据先前作者(cucarella等人,2001)进行的转化过程中的受体。通过菌落pcr检查推定的阳性克隆的蛋白质产量。
[0152]
实施例5-蛋白质表达
[0153]
将实施例4的bmb171原毒素构建物在补充有20μg/ml红霉素的50ml ccy培养基中
在旋转摇动器中在28℃和200rpm下培养2-3天,直到观察到95%细胞的孢子形成和裂解。在x1000放大倍数下通过相差显微镜观察包涵体。然后将孢子和晶体混合物在4℃下以9000g离心10min。将团块在1m nacl中洗涤三次,在冷水中洗涤六次,最后在10mm kcl中重新悬浮。
[0154]
对于蛋白质定量,在37℃下,通过温和搅拌2h,将每种重组蛋白质的量(100μl)溶解于1000μl的50mm na2co3(ph 11.3)和10mm二硫苏糖醇(dtt)溶液中。通过在4℃下以9000g离心10min,移除非溶解晶体。等分试样(500μl)用于通过bradford测定法(bradford,1976)(bio-rad)的蛋白质定量,使用牛血清白蛋白(bsa)为标准。
[0155]
实施例6-svbs-1801晶体蛋白的分子量
[0156]
晶体蛋白的分子量通常通过sds-page(laemmli,1970)进行测定。为此,必须首先进行晶体蛋白的纯化。纯化可以通过不同的方法来实现,包括thomas和ellar在1983年提出的梯度方法。一旦回收晶体蛋白,将10μl等分试样与5μl的30
×
还原剂(体积的1/10)和3
×
sds样品缓冲液(1倍体积)(new england biolabs)混合并在100℃下变性5分钟。然后将混合物装入10%聚丙烯酰胺凝胶并在36ma下运行1小时。接下来,使用50%(v/v)乙醇、10%(v/v)乙酸和0.1%考马斯蓝r 250的溶液对凝胶进行染色。
[0157]
如图3所示,svbs-1801呈现出两条约130和133kda的条带,这是cry1蛋白对应的大小。而cry2蛋白的大小为633kda。在对应于这种大小的区域中不存在条带可能表明蛋白质cry2ad1可能不会在实施例1中使用的生长条件下表达。
[0158]
实施例7-获得svbs-1801活性成分(ai)的程序
[0159]
可以通过在促进孢子形成的培养基,例如ccy(stewart等人,1981)中培养菌株svbs-1801获得ai,如实施例1的1.2节所述。当95%的孢子形成细菌细胞天然地裂解后,对培养基的内容物进行离心。产生的团块应包含孢子和晶体的混合物,这些孢子和晶体在孢子形成细胞裂解期间释放到培养基中。物理脱水浓缩可通过使用连续离心以》8.000g运行以产生固体含量为12-15%的浆液,该浆液是体积减少50倍的发酵产物,其中孢子和晶体的混合物更浓缩,但也包括上清液中的部分液体和细菌产物。上清液含有vip3a蛋白但不含cry蛋白,因为分泌到培养基中的唯一cry蛋白是cry1i;由于菌株svbs-1801的基因组中不存在cry1i基因,所述菌株不能合成cry1i蛋白并且它们不存在于上清液。
[0160]
离心以获得浆液之后可以进行配制和喷雾干燥成粉末或直接配制成可流动的。
[0161]
实施例8-从svbs-1801培养物或蛋白质获得的不同制备物对新生草地贪夜蛾幼虫的杀虫特性
[0162]
8.1.用于svbs-1801活性成分体内毒性测定的草地贪夜蛾饮食广泛用于饲养草地贪夜蛾的标准饮食是hoffman和lawson(1964)的烟草天蛾饮食的改良版本。饮食制备如下:将成分加入烧杯中并充分混合,然后加热灭菌(121℃,20分钟)。当混合物冷却至50℃时,必须补充抗生素(链霉素、金霉素)、维生素混合物、抗坏血酸和胆碱。完整的饮食可以用厨房搅拌机混合。最后,将饮食倒入400ml(17
×
10
×
2.5cm)的无菌容器中,使其在室温下固化。
[0163]
8.2.从svbs1801全培养物中获得的不同细菌制备物对新生草地贪夜蛾幼虫的杀虫特性
[0164]
为了确定浓度-死亡率应答,用从bt菌株svbs-1801培养物中获得的不同细菌制备物处理昆虫(表5)。每种处理都制备并测试了七个浓度。处理是:
[0165]-如实施例7所示,包含孢子和晶体(s c)混合物的水溶液通常构成基于bt的产品的活性成分,因此测试了七种浓度的此类水溶液。
[0166]-此外,对存在上清液的制备物测试孢子和晶体混合物与培养基中可能存在的分泌毒素之间可能的相互作用。
[0167]-此外,由于bt菌株的杀虫毒性主要归因于构成伴孢晶体的δ内毒素,因此将晶体从孢子中分离出来,并在回收晶体后,对含有纯化的晶体的处理进行测试,以确定其在没有孢子的情况下的效力。
[0168]-此外,评估了纯化的晶体与上清液之间可能的相互作用
[0169]-还测试了含有孢子、上清液以及两者的混合物、孢子和上清液的制备物。
[0170]
处理的制备
[0171]
对于所有处理,从先前定量的等分试样制备总共七种浓度的杀虫蛋白,范围为0.3至300ng/μl,以确定bt菌株和菌株本身或菌株培养产生的不同产物的毒性。
[0172]
孢子和晶体的混合物(s c)的水溶液。为了确定bt菌株的毒性,从先前定量的等分式样(实施例5)制备了总共七种浓度的杀虫蛋白,范围为0.3至300ng/μl。
[0173]
具有上清液的孢子和晶体的混合物(s c)。离心相应量的孢子和晶体混合物,并在上清液中重新悬浮所得团块。对s c处理所述的相同系列稀释液进行了测试。
[0174]
从孢子中分离伴孢晶体及制备伴有纯化的晶体的处理。为了从样品中获得纯化的晶体,采用了mounsef等人,2014描述的改进方案。将一定体积(200μl)的孢子和晶体混合物用nacl 1m洗涤三次,并以9000g离心10min。然后将团块重新悬浮在900μl pbs 1x中,并添加100μl己烷。将试管涡旋1min,并以6000g,4℃下离心10min。丢弃上清液,并对所得团块进行至少三次相同程序。晶体在冷蒸馏水中清洗三次,并在x1000放大倍数下通过相差显微镜检查不存在孢子。如实施例5末尾所述,对纯化的晶体进行定量。制备含有相当于s c系列稀释液杀虫蛋白浓度(0.3至300ng/μl)的纯化的晶体量的处理。
[0175]
上清液中的纯化的晶体。纯化的晶体和上清液之间可能的相互作用通过在上清液中重新悬浮上一段所述的相应量的伴孢晶体进行评估。
[0176]
孢子的纯化悬浮液。为了获得孢子纯化的悬浮液,通过添加含有50mm na2co3(ph 11.3)和10mm二硫苏糖醇(dtt)的溶液并在37℃下轻轻搅拌2h,对孢子和晶体的混合物的混合物进行溶解。通过在4℃下以9000g离心10min,去除上清液中的溶解晶体。此过程重复五次。在x1000放大倍数下,通过相差显微镜检查不存在晶体。纯化的孢子在petroff-hausser室进行计数。
[0177]
上清液中的孢子。通过添加上清液,从根据前段获得的纯化的孢子制备不同稀释的处理。
[0178]
生物测定表现
[0179]
所有生物测定法,包括下文第8.3节中设置的那些,均使用28个个体化的草地贪夜蛾新孵化(《12h)幼虫的组,在人工饮食表面喷洒相应的细菌制备物来进行。使用独立的制备物在不同的日期重复毒性试验至少三次,并在25 1℃的受控环境室内进行为期5天的16小时光照/8小时暗循环,并检查死亡率。在polo-pc程序(le ora software,1987)中,将浓度-死亡率数据汇总并进行概率回归分析(finney,1971)。如果95%基准限不重叠,则认为平均致死浓度(lc
50
)值存在显著差异。
[0180]
表5显示了在存在和不存在上清液作用的情况下,含有孢子和晶体混合物以及纯化的晶体的不同制备物的浓度-死亡率响应结果。
[0181]
表5.从bt菌株svbs-1801的培养物中获得的不同细菌制备物对新孵化的草地贪夜蛾幼虫的杀虫活性的相对效力。
[0182][0183]
与表5中处理所得回归线对应的参数表明,孢子和晶体混合物的lc
50
值存在显著差异,在存在上清液的情况下毒性增加2.39倍。在没有孢子的情况下,纯化的晶体的活性显著提高,这表明在一起接种时会产生不利影响。然而,当纯化的晶体与上清液组合时,与孢子和晶体混合物的水溶液制备物相比,报告了28.19倍高的有效处理,证明了协同效应。
[0184]
含有孢子、上清液或两者的组合的细菌制备物没有报告活性。
[0185]
8.3.svbs-1801cry蛋白对新生草地贪夜蛾幼虫的杀虫特性
[0186]
如上所述,bt菌株的杀虫毒性主要归因于构成伴胞晶体的δ内毒素。但并不是所有菌株都对特定的昆虫表现出活性或相同程度的活性。因此,按照第8.2节中描述的相同方法,进行单个浓度(100ng/μl)生物测定法,以确定伴胞晶体中所含单个蛋白质的活性。由于在实施例6的sds-page测定法中获得的结果,蛋白质cry2ad1未包含在测定法中,这似乎表明该蛋白质不能在本技术实施例1中使用的细菌生长条件下表达。
[0187]
表6显示了当以单一浓度的杀虫蛋白(100ng/μl)施用时,每种单独的cry蛋白的单独活性。它表明在五种蛋白质中,由于cry1ab1和cry1nb1对新孵化的草地贪夜蛾幼虫缺乏活性,因此伴孢晶体的杀虫活性应归因于cry1da3、cry1ea7和cry1ja1样。
[0188]
表6.单独蛋白质对草地贪夜蛾新孵化幼虫的毒性。
[0189][0190]
随后,进一步评估了毒性蛋白质并估计了平均致死浓度(lc
50
)。最后,通过测试含有等摩尔浓度的先前重组蛋白的混合物来评估蛋白质相互作用。
[0191]
表7显示了当单独评估时以及在包含所有三种相同比例的等摩尔混合物中进行评估时,对应于三种报告的毒性蛋白质(cry1da3、cry1ea7和cry1ja1样)所获得的回归线的参数。
[0192]
表7.三种单独的蛋白质(cry1da、cry1ea和cry1ja样)和以1:1:1的比例包含所有三种蛋白质的人工等摩尔混合物对草地贪夜蛾新生幼虫的杀虫效力。
[0193][0194][0195]
从表7中可以看出,草地贪夜蛾幼虫对cry1da3和cry1ja1样非常敏感,其毒性分别比cry1ea7高10.34倍和8.25倍。cry1ea7的相对效力的基准限(fl)之间没有重叠清楚地表明,与cry1da3或cry1ja样相比,差异具有统计学显著性。当三种蛋白质在等摩尔混合物中组合时,lc
50
值降低至1.09ng/μl,与三种单独的毒素相比,效力明显更强,表明它们之间存在协同作用。
[0196]
为了计算协同因子,根据混合物中cry1da3(ra)、cry1ea7(ra)和cry1ja1样(rc)的相对比例,混合物相对于三种单一蛋白质的预期平均致死浓度(lc
50
)必须进行计算(tabashnik,1992)并与获得的混合物lc
50
进行比较(表7)。
[0197]
[0198][0199]
对于新孵化的草地贪夜蛾幼虫,cry1da3、cry1ea7和cry1ja1样的混合物获得了3.63的协同因子。
[0200]
实施例9-确定svbs-1801菌株相对于商业菌株dipel(abts-351)和xentari(abts-1875)在草地贪夜蛾二龄幼虫中杀虫效力的毒性试验
[0201]
9.1.获取svbs-1801活性成分(ai)的程序
[0202]
获得svbs-1801活性成分的程序如实施例7所述。
[0203]
9.2.商业产品bt菌株活性成分的制备
[0204]
为了比较从svbs-1801菌株和产生商业产品的bt菌株制备的活性成分的活性(对于为abts-351,对于为abts-1857),苏云金芽孢杆菌菌株abts-351和abts-1857分别直接从在欧盟销售的商业产品df(kenogard,valent bioscience corporation;生产批次261-355-pg;生产日期2016年1月)和gd(kenogard,valent bioscience corporation;生产批次264-637-pg;生产日期2016年4月)中分离。每种细菌菌株在28℃的无菌ccy培养基中培养72h(stewart等人,1981),并且对这些菌株也遵循实施例5中解释的过程,直到为任何菌株获得固体含量为12-15%的浆液,这被认为是ai,以与从菌株svbs-1801获得的ai进行比较。
[0205]
9.3.使用来自abts-351abts-1857和svbs-1801菌株的晶体和孢子混合物的液滴喂食法在草地贪夜蛾二龄幼虫中的毒性测定。
[0206]
为了进行毒性测定,需要以下材料:28孔水泡、人工饮食(见第8.1节)和草地贪夜蛾新蜕皮二龄幼虫。晶体和孢子的混合物应使用3到5的稀释因子稀释五次,这将产生下一步测试ai的6种不同浓度。同样,应准备缺乏ai但保留其余组分的阴性对照。按照液滴喂食方法(hughes和wood,1981),给草地贪夜蛾新蜕皮二龄幼虫喂食6种不同浓度的ai中的一种。将喂食的幼虫放入含有1.5cm3人工饮食的28孔水泡的单个孔中。
[0207]
在这项研究中,共有28个新蜕皮二龄幼虫接受了每种蛋白质浓度的处理,每种bt菌株使用了5种浓度的范围。生物测定法进行了三次。对照昆虫饲喂不含毒素的人工饮食。多孔板在25℃、60%r.h.和14h/10h(光/暗)光周期下孵育。6天后记录死亡率。在polo-pc程序(leora software,1987)中,浓度-死亡率数据进行probit回归分析(finney,1971)。
[0208]
用三种bt菌株(从分离的abts-351、从分离的abts-1857和svbs-180)1在二龄草地贪夜蛾幼虫中获得的浓度-死亡率应答结果见表8、9和10。三个bt分离株的拟合回归线斜率范围为abts-351的0.51到svbs-1801的1.47。斜率值差异很大,不允许平行以共同的斜率调整三条回归线。因此,为了估计三种bt菌株之间的相对效力,有必要对回归线进行二对二的比较。
[0209]
表8显示了对应于从分离得到的bt菌株abts-351、从分离得的abts-1857获得的回归线的参数。在表4中,可以看出,菌株abts-1857对草地贪夜蛾二龄幼虫的效力是菌株abts-351的217.2倍。两种菌株相对效力的基准限(fl)之间没有重叠,这清楚地表明观察到的差异是统计学显著的。
[0210]
表8.abts-1857相对于abts-351对草地贪夜蛾二龄的lc
50
值和相对效力。
[0211][0212]
表9显示了对应于从分离的bt菌株abts-351和svbs-1801获得的回归线的参数。表9反映菌株svbs-1801对草地贪夜蛾二龄幼虫的效力是菌株abts-351的1235.4倍。两种菌株的相对效力的基准限(fl)之间没有重叠,这清楚地表明观察到的差异是统计学显著的。
[0213]
表9.svbs-1801相对于abts-351对草地贪夜蛾二龄的lc
50
值和相对效力。
[0214][0215]
表10显示了对应于从分离的bt菌株abts-1857和svbs-1801获得的回归线的参数。svbs-1801对草地贪夜蛾幼虫的效力是abts-351的5.7倍。两种菌株相对效力的基准限(fl)之间没有重叠,这清楚地表明差异是统计学显著的。
[0216]
表10.svbs-1801与abts-1857相比对草地贪夜蛾二龄的lc
50
值和相对效力。
[0217][0218]
实施例10-svbs-1801活性成分相对于abts-351和abts-1857活性成分对草地贪夜蛾二龄幼虫的杀虫效力
[0219]
如例9.3所示,abts-351abts-1857和svbs-1801 ai的毒性通过液滴喂食法进行评估(hughes andwood,1981)。abts-351(苏云金芽孢杆菌ser.kurstaki)被广泛接受为几种鳞翅目物种的国际参考,并构成商业产品如2x(abbot)的ai。类似地,abts-1857(从分离)通常作为旨在测试产品针对夜蛾属
鳞翅目的杀虫效力的生物测定法中的参考ai。
[0220]
svbs-1801杀虫蛋白相对于abts-351和abts-1857杀虫蛋白的效力是根据dulmage(1981)所述用作参考的商业产品标签中指示的效力(参考效力)计算的。
[0221]
公式
[0222][0223]
iu:国际单位
[0224]
相对于abts-351的svbs-1801效力:效力产生32000iu/mg
[0225][0226]
相对于abts-1857的svbs-1801效力:效力产生15000iu/mg:
[0227][0228]
实施例11-svbs-1801相对于hd1和abts-1857对半田间玉米植株中草地贪夜蛾二龄幼虫的杀虫效力
[0229]
在物候期v3(5-6片,30-35厘米高)的玉米植株上寄生有草地贪夜蛾卵(150个卵/株),这些卵可在24小时内孵化。一旦幼虫出现,就对其进行监测,直到达到二龄。接下来,用晶体和孢子的混合物处理植物(对于abts-1857和svbs-1801菌株,晶体和孢子的浓度相当于100mg/l)。使用压缩空气手动喷雾器(matabiantzuola,guipuzcoa,spain)进行应用,其中包括0.05%agral湿润剂贴纸。排放10ml喷雾处理,相当于该地区玉米作物使用的标准喷雾量(约1000liters/ha)。
[0230]
在施用后20、30和40小时,用手从每个地块的植物中随机采集28只草地贪夜蛾幼虫,并在实验室中在25℃用人工饲料在25ml塑料杯中饲养,直至死亡或化蛹。每天记录死亡率。计算每种处理的死亡率百分比,通过反正弦变换归一化,并在spss ver.12.0(spss,chicago,il)中进行重复测量方差分析(anova)。通过应用mauchly球度检验,对该分析和所有后续多元分析的方差-协方差矩阵的特征进行了检验。结果如图8所示。
[0231]
生物材料的保藏
[0232]
菌株svbs-1801,已保藏在colecci
ónꢀꢀ
de cultivos tipo(cect)(parque cient
í
fico de la universidad de valencia;calle catedr
á
tico agust
í
n escardino,9;46980paterna,valencia,spain),其根据1997年5月12日第664/1997号皇家法令接受细菌菌株保藏。svbs-1801已按照《布达佩斯条约指南》为专利目的进行保藏。
[0233]
该菌株的原始保藏收据(bp/4)和存活证明(bp/9)反映了以下相关数据:
[0234]-最终保藏日期:2018年11月8日,
[0235]-菌株参考号(登录号):cect 9753。
[0236]-保藏人:serena valley biological systems s.l.(paseo universidad 49,31192mutilva,navarra,spain)。保藏人与申请人不同。
[0237]
参考文献
[0238]
altschul,s.f.,gish,w.,miller,w.,myers,e.w.&lipman,d.j.(1990)"basic local alignment search tool."j.mol.biol.215:403-410.
[0239]
baig,d.n.,bukhari,d.a.和shakoori,a.r.(2010)cry genes profiling and the toxicity of isolates of bacillus thuringiensis from soil samples against american bollworm,helicoverpa armigera.j.appl.microbiol.,109,1967
–
1978.
[0240]
barretto,m.r.等人,(1999).insecticidal activity of culture supernatants from bacillus thuringiensis berliner strains against spodoptera frugiperda smith(lepidoptera.noctuidae)larvae",an.soc.entomol.brasil,28:675-685
[0241]
bradford,m.m.,1976.a rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding.anal.biochem.72,248
–
254
[0242]
capalbo d.等人,(2001)solid-state fermentation of bacillus thuringiensis tolworthi to control fall armyworm in maize,ejb electronic journal of biotechnology 4(2):717-3458
[0243]
carozzi,n.b.,kramer,v.c.,warren,g.w.,evola,s.和koziel,m.g.(1991)prediction of insecticidal activity of bacillus thuringiensis strains by polymerase chain reaction product profiles.appl.environ.microbiol.,57,3057
–
3061.
[0244]
cer
ó
n,j.,ortiz,a.,quintero,r.,g
ü
ereca,i.和bravo,a.(1995)specific pcr primers directed to identify《i》cryi《i》and《i》cryiii《i》genes within a《i》bacillus thuringiensis《i》strain collection.appl.envir.microbiol.,61,3826
–
3831.
[0245]
crickmore,n.,baum,j.,bravo,a.,lereclus,d.,narva,k.,sampson,k.,等人,(2018)bacillus thuringiensis toxin nomenclature.
[0246]
crickmore,n.,zeigler,d.r.,feitelson,j.,schnepf,e.,van rie,j.,lereclus,d.,等人,(1998)revision of the nomenclature for the bacillus thuringiensis pesticidal crystal proteins.microbiol.mol.biol.rev.,62,807
–
13.
[0247]
cucarella,c.,solano,c.,valle,j.,amorena,b.,lasa,i.和penad
é
s,j..(2001)bap,a staphylococcus aureus surface protein involved in biofilm formation.j.bacteriol.,183,2888
–
2896.
[0248]
donovan,w.p.,engleman,j.t.,donovan,j.c.,baum,j.a.,bunkers,g.j.,chi,d.j.,等人,(2006)discovery and characterization of sip1a:a novel secreted protein from bacillus thuringiensis with activity against coleopteran larvae
sheng wu xue bao,40,85
–
90.
[0264]
l
ó
pez-pazos,s.a.,rojas arias,a.c.,ospina,s.a.和cer
ó
n,j.(2010)activity of bacillus thuringiensis hybrid protein against alepidopteran and a coleopteran pest.fems microbiol.lett.,302,93
–
98.
[0265]
masson,l.,erlandson,m.,puzstai-carey,m.,brousseau,r.,ju
á
rez-p
é
rez,v.和frutos,r.(1998)a holistic approach for determining the entomopathogenic potential of bacillus thuringiensis strains.appl.environ.microbiol.,64,4782
–
8.
[0266]
milner,r.j.(1994)history of bacillus thuringiensis.agric.ecosyst.environ.,49,9
–
13.
[0267]
mounsef jr,salameh d,awad m kallassy,chamy l,brandam c,lteif r.2014.a simple method for the separation of bacillus thuringiensis spores and crystals.j microbiol methods 107:147
–
149
[0268]
ohba,m.,mizuki,e.和uemori,a.(2009)parasporin,a new anticancer protein group from bacillus thuringiensis.anticancer res.,29,427
–
33.
[0269]
park,h.w.,ge,b.,bauer,l.s.和federici,b.a.(1998)optimization of cry3a yields in bacillus thuringiensis by use of sporulation-dependent promoters in combination with the stab-sd mrna sequence.appl.environ.microbiol.,64,3932
–
8.
[0270]
polanczyk,r.a.等人,(2000)effectiveness of bacillus thuringiensis strains against spodoptera frugiperda(lepidopera:noctuidae)".brazilian journal of microbiology,31:165-167..
[0271]
roh,j.y.,kim,y.s.,wang,y.,liu,q.,tao,x.,xu,h.g.,等人,(2010)expression of bacillus thuringiensis mosquitocidal toxin in an antimicrobial bacillus brevis strain.j.asia.pac.entomol.,13,61
–
64.
[0272]
sambrook,j.,fritsch,e.f.和maniatis,t.(1989)molecular cloning:a laboratory manual.cold spring harbor laboratory press.
[0273]
schnepf,h.e.和whiteley,h.r.(1981)cloning and expression of the bacillus thuringiensis crystal protein gene in escherichia coli.proc.natl.acad.sci.u.s.a.,78,2893
–
7.
[0274]
k.和horsk
á
,k.(1970)mechanism of inhibition of dna-dependent rna polymerase by exotoxin of bacillus thuringiensis.biochim.biophys.acta-nucleic acids protein synth.,209,357
–
367.
[0275]
shah,s.h.,jan,s.a.,ahmad,n.,khan,s.u.,kumar,t.,iqbal,a.,等人,(2015)use of different promoters in transgenic plant development:current challenges and future perspectives.am.j.agric.environ.sci.,15,664
–
675.
[0276]
sharma,h.c.,dhillon,m.k.,和arora,r.(2008)effects of bacillus thuringiensisδ-endotoxin-fed helicoverpa armigera on the survival and development of the parasitoid campoletis chlorideae.entomol.exp.appl.,126,1
–
8.
[0277]
stewart,g.s.,johnstone,k.,hagelberg,e.和ellar,d.j.(1981)commitment of bacterial spores to germinate a measure of the trigger reaction.biochem.j.,198,101
–
106.
[0278]
tabashnik,b.e.(1992)evaluation of synergism among bacillus thuringiensis toxins.appl.environ.microbiol.,58,3343
–
3346.
[0279]
thomas,w.e.和ellar,d.j.(1983)bacillus thuringiensis var israelensis crystal delta-endotoxin:effects on insect and mammalian cells in vitro and in vivo.j.cell sci.,60,181
–
97。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。