1.本发明涉及一种蛋白质纤维的制备方法、蛋白质纤维面料的制备方法以及蛋白质纤维的防缩加工方法。
背景技术:
2.以往,丝绸及羊毛等蛋白质纤维发挥其具有优异的肌肤触感及保温性等的特征,被广泛用于服装和床上用品等材料。此外,最近,具有优异强度和高伸缩性的高韧性蜘蛛丝受到关注,并积极对具备该蜘蛛丝特征的人造蜘蛛丝的实用化进行研究。人造蜘蛛丝不仅适用于服装,还正在研究将其应用于需要高强度和高韧性等的工业用材料和医疗用材料等各种材料中。而且,这些蛋白质纤维与合成纤维不同,其均具有生物降解性,并且生产和加工时所需的能量小,因此,随着近年来环保意识的提高,预计需求量会增加。
3.在蛋白质纤维中,存在因与水分接触(例如,浸在水或热水中或暴露在高湿度环境中等)而收缩的蛋白质纤维。特别是在制备后首次与水分接触时发生这种因蛋白质纤维的收缩而引起的未预期的长度变化时,有可能由此引起各种问题。例如,蛋白质纤维在刚刚制备完成后进行保管的过程中暴露于高湿度环境下,或者在制备后首次进行洗涤和湿热定形等水分处理时,发生未预期的长度变化时,不仅可能会降低作业性能和品质,而且还有可能对后加工带来不利影响。
4.在这种情况下,公开了各种蛋白质纤维的防缩方法。例如,在专利文献1中公开了一种蛋白质纤维的制备方法,其包括:伸缩工序,其使含有蛋白质的蛋白质原料纤维与液体或蒸气接触,并使该蛋白质原料纤维收缩或伸长;干燥工序,其经过伸缩工序后的蛋白质原料纤维调整为任意长度并进行干燥。
5.现有技术文献
6.专利文献
7.专利文献1:国际公开第2018/164020号
技术实现要素:
8.发明要解决的问题
9.在一般的在线(连续)纺丝中,为了使蛋白质纤维从上游侧向下游侧移动,不可避免地对蛋白质纤维施加纤维轴方向上的张力。根据本发明人等的研究,当在在线纺丝的中途进行防缩处理时,由于防缩处理后不可避免地产生纤维轴方向上的张力(例如卷绕时的张力),从而在制备后的蛋白质纤维中产生残余应力,由于这种残余应力,即使是实施防缩处理后的蛋白质纤维,在制备后首次与水分接触时也会发生收缩,特别是与高温水分接触时收缩显著。
10.为了更可靠地抑制制备后首次与水分接触时所产生的收缩,考虑对在线纺丝后的蛋白质纤维在尽量不施加纤维轴方向的张力的情况下实施防缩处理,以尽量不产生残余应
力。但是,在这种情况下,蛋白质纤维与水分接触时会发生起皱。因此,所得到的蛋白质纤维存在可使用的用途受限的问题。
11.因此,本发明的目的在于提供一种制备蛋白质纤维的方法,其能够更可靠地抑制制备后首次与水分接触时所产生的收缩,并且还可以抑制起皱的发生。本发明的目的还在于提供一种制备蛋白质纤维面料的方法,其能够更可靠地抑制制备后首次与水分接触时所产生的收缩,并且还可以抑制起皱的发生。本发明的目的还在于提供一种蛋白质纤维的防缩加工方法,其能够更可靠地抑制制备后首次与水分接触时所产生的收缩,并且还可以抑制起皱的发生。
12.用于解决问题的方法
13.本发明涉及例如下述各发明。
14.1.15.一种蛋白质纤维的制备方法,其具备:前收缩工序,其用于使拉伸后且卷绕前的蛋白质原纤维在松弛状态下进行收缩;卷绕工序,其用于将经过上述前收缩工序的蛋白质原纤维卷绕以得到卷绕物;以及后收缩工序,其用于使经过上述卷绕工序的蛋白质原纤维在松弛状态下进行收缩。
16.2.17.根据[1]所述的制备方法,其中,上述后收缩工序是用于使经过上述卷绕工序的蛋白质原纤维在松弛状态下与液体或蒸气相接触后收缩的湿式收缩工序,或者是用于使经过上述卷绕工序的蛋白质原纤维在松弛状态下加热后收缩的干式收缩工序。
[0018]
[3]
[0019]
根据[2]所述的制备方法,其中,上述湿式收缩工序是用于将经过上述卷绕工序的蛋白质原纤维在松弛状态下浸渍于液体中以使其收缩的工序。
[0020]
[4]
[0021]
根据[2]或[3]所述的制备方法,其中,上述液体或蒸气具有极性。
[0022]
[5]
[0023]
根据[4]所述的制备方法,其中,上述液体是水,上述蒸气是水蒸气。
[0024]
[6]
[0025]
根据[1]至[5]中任一项所述的制备方法,其中,上述蛋白质原纤维含有结构蛋白质。
[0026]
[7]
[0027]
根据[6]所述的制备方法,其中,上述结构蛋白质为改造丝心蛋白。
[0028]
[8]
[0029]
根据[7]所述的制备方法,其中,上述改造丝心蛋白是改造蜘蛛丝丝心蛋白。
[0030]
[9]
[0031]
一种蛋白质纤维面料的制备方法,其具备:使用通过[1]~[8]中任一项所述的制备方法得到的蛋白质纤维来制作面料的工序。
[0032]
[10]
[0033]
一种蛋白质纤维的防缩加工方法,其具备:前收缩工序,其用于使拉伸后且卷绕前的蛋白质纤维在松弛状态下进行收缩;卷绕工序,其用于将经过上述前收缩工序的蛋白质
纤维卷绕以得到卷绕物;以及后收缩工序,其用于使经过上述卷绕工序的蛋白质纤维在松弛状态下进行收缩。
[0034]
[11]
[0035]
根据[10]所述的防缩加工方法,其中,上述后收缩工序是用于使经过上述卷绕工序的蛋白质纤维在松弛状态下与液体或蒸气相接触后收缩的湿式收缩工序,或者是用于使经过上述卷绕工序的蛋白质纤维在松弛状态下加热后收缩的干式收缩工序。
[0036]
发明效果
[0037]
根据本发明,可以提供一种制备蛋白质纤维的方法,其能够更可靠地抑制制备后首次与水分接触时所产生的收缩,并且还可以抑制起皱的发生。根据本发明,还可以提供一种制备蛋白质纤维面料的方法,其能够更可靠地抑制制备后首次与水分接触时所产生的收缩,并且还可以抑制起皱的发生。根据本发明,还可以提供一种蛋白质纤维的防缩加工方法,其能够更可靠地抑制制备后首次与水分接触时所产生的收缩,并且还可以抑制起皱的发生。
附图说明
[0038]
图1是示意性地表示一实施方式所涉及的蛋白质原纤维的制备装置的图。
[0039]
图2是示意性地表示一实施方式所涉及的蛋白质原纤维的制备装置的图。
[0040]
图3是表示可设置在图2的高温加热炉中的速度调节单元及温度调节单元的图。
[0041]
图4是示意性地表示一实施方式所涉及的蛋白质纤维的制备装置(后收缩装置)的图。
[0042]
图5是示意性地表示一实施方式所涉及的蛋白质纤维的制备装置(后收缩装置)的图。
[0043]
图6是示意性地表示一实施方式所涉及的蛋白质纤维的制备装置(后收缩装置)的图。(a)是通过蛋白质原纤维的卷绕物得到绞纱的装置的概略图。(b)是湿式收缩装置的概略图。(c)是干式收缩装置的概略图。
[0044]
图7是表示改造丝心蛋白的结构域序列的一例的示意图。
[0045]
图8是表示天然来源的丝心蛋白的z/w(%)的值的分布的图。
[0046]
图9是表示天然来源的丝心蛋白的x/y(%)的值的分布的图。
[0047]
图10是表示改造丝心蛋白的结构域序列的一例的示意图。
[0048]
图11是表示改造丝心蛋白的结构域序列的一例的示意图。
具体实施方式
[0049]
下面,根据情况参照附图对本发明的优选实施方式进行详细说明,但本发明并不限定于以下实施方式。另外,在附图中,对相同或相当部分标注相同符号,并适当省略重复的说明。
[0050]
[蛋白质纤维的制备方法]
[0051]
本实施方式所涉及的蛋白质纤维的制备方法具备:前收缩工序,其用于使拉伸后且卷绕前的蛋白质原纤维在松弛状态下进行收缩;卷绕工序,其用于将经过前收缩工序的蛋白质原纤维卷绕以得到卷绕物;以及后收缩工序,其用于使经过卷绕工序的蛋白质原纤
维在松弛状态下进行收缩。
[0052]
本实施方式所涉及的蛋白质纤维的制备方法通过具备后收缩工序,可以更可靠地抑制所得到的蛋白质纤维在制备后首次与水分接触时所产生的收缩。当所接触的水分的温度高时,该效果尤其显著。此外,本实施方式所涉及的蛋白质纤维的制备方法除了后收缩工序之外还具备前收缩工序,从而可以充分地抑制所得到的蛋白质纤维与水分接触时所产生的起皱。
[0053]
在本说明书中,所谓“使蛋白质原纤维在松弛状态下进行收缩”,是指除了使蛋白质原纤维在不施加任何张力的松弛状态下自然收缩的情况之外,还包括使蛋白质原纤维在施加张力的状态下收缩的情况,该张力使蛋白质原纤维的收缩量与蛋白质原纤维在不施加任何张力的松弛状态下自然收缩时的收缩量实质上相同。另外,本说明书中的“实质上相同”是指等量或者等值,或者与其近似的量或者值,而且,近似的量或者值是指能够起到与等量或者等值时得到的效果相同的效果的量。
[0054]
(前收缩工序)
[0055]
前收缩工序是用于使拉伸后且卷绕前的蛋白质原纤维在松弛状态下进行收缩的工序。前收缩工序可以在卷绕工序之前实施,例如,使用纺液(纺丝原液)通过公知的纺丝方法(干式纺丝、湿式纺丝、干湿式纺丝或熔融纺丝等)进行纺丝后,可以对通过公知的拉伸方法(湿热拉伸或干热拉伸等)拉伸的蛋白质原纤维实施该工序。
[0056]
作为使蛋白质原纤维在松弛状态下进行收缩的方法,存在例如包括用于使蛋白质原纤维在松弛状态下与液体或蒸气相接触后收缩的湿式收缩工序的方法(湿式收缩法)、包括用于使蛋白质原纤维在松弛状态下加热后收缩的干式收缩工序的方法(干式收缩法)等。
[0057]
在湿式收缩法中,通过使蛋白质原纤维与液体或蒸气相接触,以使得蛋白质原纤维在不受外力影响的情况下进行收缩。认为这是在因拉伸而具有残余应力的蛋白质原纤维中,液体或蒸气浸入纤维间或纤维中,缓和了残余应力而产生的。
[0058]
湿式收缩法中使用的液体或蒸气只要是能够通过与蛋白质原纤维接触而使蛋白质原纤维自然收缩的液体或蒸气即可,其种类并没有特别限定。作为液体或蒸气,例如可以列举出具有极性的液体或蒸气。作为具有极性的液体,例如可以列举出水、四氢呋喃、乙腈、丙酮和甲醇等,其中,优选使用水。这是因为水不仅廉价且操作性优异,而且可以通过与蛋白质原纤维的接触来使蛋白质原纤维更迅速且可靠地收缩。作为具有极性的蒸气,例如可以列举出具有上述极性的液体的蒸气,其中,优选使用水蒸气。这是因为水蒸气与水一样,可以通过与蛋白质原纤维接触来使蛋白质原纤维更迅速且可靠地收缩。
[0059]
在湿式收缩法中,优选使液体在加温的状态下与蛋白质原纤维相接触。由此,可以更有利地缩短蛋白质原纤维的收缩时间。从相同的观点出发,湿式收缩法中使用的液体与水相比更优选为热水(加温后的水)。
[0060]
当使用加温后的液体进行湿式收缩法时,液体的温度只要比蛋白质原纤维中所含的蛋白质的分解温度、或蛋白质原纤维受到热损伤的温度低即可。但是,考虑到液体的操作性能和湿式收缩法的作业性能等,液体温度的上限值优选低于沸点。此外,从充分获得缩短收缩时间的效果的观点出发,液体的温度的下限值优选为10℃以上,更优选为40℃以上,进一步优选为70℃以上。作为液体使用水时,水的温度的上限值优选为90℃以下,更优选为80℃以下。水的温度的下限值优选为10℃以上,更优选为40℃以上,进一步优选为70℃以上。
此外,在使液体与蛋白质原纤维接触的期间内,液体的温度可以固定,也可以使液体的温度变动为规定的温度。
[0061]
在湿式收缩法中,使液体或蒸气与蛋白质原纤维相接触的方法并没有特别限定。作为该方法,例如可以列举出以下方法:将蛋白质原纤维浸渍在液体中的方法、将液体在常温或加温后的蒸汽等状态下对蛋白质原纤维进行喷水的方法、以及将蛋白质原纤维暴露在充满蒸气的高湿度环境下的方法等。在这些方法中,由于能够有效地实现收缩时间的缩短,并且能够实现收缩加工设备的简化等,因此优选为将蛋白质原纤维浸渍于液体中的方法。
[0062]
蛋白质原纤维与液体的接触时间并没有特别限制,例如可以为1分钟以上。该时间可以为10分钟以上,也可以为20分钟以上,还可以为30分钟以上。此外,该时间的上限并没有特别限制,但是从缩短制造工序的时间的观点、以及排除蛋白质原纤维分解(水解等)的可能性等观点出发,例如,可以为120分钟以下,也可以为90分钟以下,还可以为60分钟以下。
[0063]
在湿式收缩法中,在湿式收缩工序之后可以进一步包括将蛋白质原纤维进行干燥的工序(干燥工序)。
[0064]
干燥工序中的干燥方法并没有特别限制,例如可以是自然干燥,也可以使用干燥设备强制干燥。作为干燥温度,只要比蛋白质热损伤的温度低即可,并没有任何限定,一般为20~150℃的范围内的温度,优选为40~120℃的范围内的温度,更优选为60~100℃的范围内的温度。在这样的温度范围内,蛋白质不会发生热损伤等,并能够更迅速且高效地对蛋白质原纤维进行干燥。干燥时间根据干燥温度等适当选择,例如,采用能够排除过度干燥蛋白质原纤维对编织体的品质和物理性能等的影响的时间。
[0065]
干式收缩法具备使蛋白质原纤维在松弛状态下加热后收缩的干式收缩工序。干式收缩工序可以是包括加热蛋白质原纤维的步骤(加热步骤)和使处于加热状态的蛋白质原纤维在松弛状态下收缩的步骤(松弛收缩步骤)的工序。
[0066]
在蛋白质原纤维的加热中,加热温度优选为用于蛋白质原纤维的蛋白质的软化温度以上。本说明书中所谓的蛋白质软化温度,是指由于蛋白质原纤维的应力缓和而开始收缩的温度。在蛋白质的软化温度以上的加热松弛收缩中,蛋白质原纤维收缩到无法仅仅通过脱离蛋白质原纤维中的水分来得到的程度。加热温度优选为80℃以上,更优选为180℃~280℃,进一步优选为200℃~240℃,更进一步优选为220℃~240℃。从加热处理后的蛋白质原纤维的伸长率的观点出发,加热时间优选为60秒以下,更优选为30秒以下,进一步优选为5秒以下。认为该加热时间的长度对蛋白质原纤维的应力的影响不大。
[0067]
(卷绕工序)
[0068]
卷绕工序是用于将经过前收缩工序的蛋白质原纤维卷绕以得到卷绕物的工序。卷绕工序可以在前收缩工序后立即实施,也可以在前收缩工序后进一步执行其他工序(例如使蛋白质原纤维干燥的干燥工序)后实施。
[0069]
图1是示意性地表示蛋白质原纤维的制备装置(纺丝装置)的一例的图。图1所示的纺丝装置100从上游侧依次具有挤出装置1、凝固浴槽20、洗涤浴槽21、实施前收缩工序的水浴46、干燥装置4。在凝固浴槽20和/或洗涤浴槽21中实施拉伸。
[0070]
挤出装置1具有储槽7,在此储存有纺液(纺丝原液)6。凝固液11(例如甲醇)储存在凝固浴槽20中。纺液6通过安装在储槽7下端部的齿轮泵8,在其与凝固液11之间打开气隙19
并从设置的喷嘴9中挤出。挤出的纺液6经过气隙19供给到凝固液11内。在凝固液11内从纺液6中除去溶剂,蛋白质发生凝固。将凝固后的蛋白质导入至洗涤浴槽21中,通过洗涤浴槽21内的洗涤液12进行洗涤后,由设置在洗涤浴槽21内的第1轧辊13和第2轧辊14将其输送到水浴46。此时,例如,如果将第2轧辊14的转速设定地比第1轧辊13的转速快,则以与转速比相对应的倍率拉伸蛋白质原纤维。洗涤液12中被拉伸的蛋白质原纤维脱离洗涤浴槽21内之后,在通过水浴46内时在松弛状态下收缩(前收缩工序),接下来在通过干燥装置4内时被干燥,之后通过卷线机卷绕(卷绕工序)。如此一来,最终作为卷绕在卷线机上的卷绕物5得到蛋白质原纤维。另外,18a~18g是导丝器。
[0071]
在水浴46的上游侧和下游侧配置有用于转送蛋白质原纤维的转送辊50和52。水浴46具有加热器54,通过该加热器54加热的液体(热水)47容纳在水浴46内。此外,在水浴46内,在浸渍于液体(热水)47中的状态下设置有压辊56。由此,从上游侧送出的蛋白质原纤维在水浴46内被卷绕在压辊56上,并在被按压的状态下浸渍在液体(热水)47中,同时向下游侧移动。蛋白质原纤维在热水47中的浸渍时间根据蛋白质料纤维的移动速度来适当控制。
[0072]
在水浴46中,通过第2轧辊14来控制蛋白质原纤维的送出速度并通过卷线机来控制蛋白质原纤维的卷绕速度,蛋白质原纤维在松弛状态下通过与液体(热水)47相接触而自然地收缩。例如,作为蛋白质原纤维,当使用在松弛状态下通过将其浸渍在液体(热水)47中以使其在纤维轴方向收缩20%左右的纤维时,如果将卷绕速度和送出速度设为后者比前者慢80%的速度,则可以使蛋白质原纤维在液体(热水)47中以松弛的状态自然地收缩。
[0073]
作为凝固液11,只要是能够脱溶剂的溶液即可,可以列举例如甲醇、乙醇和2-丙醇等碳原子数1~5的低级醇、以及丙酮等。凝固液11可以包含适当的水。凝固液11的温度优选为0~30℃。凝固的蛋白质通过凝固液11中的距离(实质上是从导丝器18a到导丝器18b的距离)只要是能够有效地进行脱溶剂的长度即可,例如可以为200~500mm。在凝固液11中的停留时间例如可以为0.01~3分钟,优选为0.05~0.15分钟。此外,可以在凝固液11中进行拉伸(预拉伸)。
[0074]
另外,得到蛋白质原纤维时实施的拉伸,例如除了在上述凝固浴槽20内进行的预拉伸和在洗涤浴槽21内进行的拉伸以外,还可以采用湿热拉伸和干热拉伸。
[0075]
湿热拉伸可以在温水中、向温水中添加有机溶剂等而得到的溶液中、蒸气加热中进行。作为温度,例如可以为50~90℃,优选为75~85℃。在湿热拉伸中,可以将未拉伸丝(或预拉伸丝)拉伸例如1~10倍,优选拉伸2~8倍。
[0076]
干热拉伸可以使用管式电热炉、干热板等来进行。作为温度,例如可以为140℃~270℃,优选为160℃~230℃。在干热拉伸中,可以将未拉伸丝(或预拉伸丝)拉伸例如0.5~8倍,优选拉伸1~4倍。
[0077]
湿热拉伸和干热拉伸可以分别单独进行,另外也可以将它们以多步或组合进行。即,可以以湿热拉伸进行第一步拉伸,以干热拉伸进行第二步拉伸;或者第一步拉伸进行湿热拉伸,第二步拉伸进行湿热拉伸,进一步以干热拉伸进行第三步拉伸等,可以将湿热拉伸和干热拉伸适当组合进行。
[0078]
最终拉伸倍率相对于未拉伸丝(或预拉伸丝),其下限值优选为大于1倍、2倍以上、3倍以上、4倍以上、5倍以上、6倍以上、7倍以上、8倍以上或9倍以上中的任一个,其上限值优选为40倍以下、30倍以下、20倍以下、15倍以下、14倍以下、13倍以下、12倍以下、11倍以下、
10倍以下。
[0079]
图2是示意性地表示蛋白质原纤维的制备装置(纺丝装置)的其他例子的图。图3是表示可设置在图2的高温加热炉中的速度调节单元及温度调节单元的图。图2所示的纺丝装置200用于干式收缩法,其具备用于对蛋白质原纤维进行纺丝的纺丝装置(纺丝单元)25、以及用于使由纺丝装置25纺丝而成的蛋白质原纤维在松弛状态下进行高温加热后收缩的高温加热松弛装置60。在纺丝装置200中,连续对蛋白质原纤维执行纺丝工序和干式收缩工序,该干式收缩工序是用于使蛋白质原纤维在松弛状态下进行高温加热后收缩的工序。
[0080]
纺丝装置25例如是干湿式纺丝用的纺丝装置,从上游侧起依次具备挤出装置1、凝固装置2、洗涤装置3和干燥装置4。挤出装置1具有储槽7,在该储槽7中储存有纺液(纺丝原液)6。凝固装置2具有凝固浴槽20,在该凝固浴槽20中储存有凝固液11(例如甲醇)。纺液6通过安装在储槽7下端部的齿轮泵8,在其与凝固液11之间打开气隙19并从设置的喷嘴9中挤出。挤出的纺液6经过气隙19供给到凝固液11内。在凝固液11内从纺液6中除去溶剂,蛋白质发生凝固。
[0081]
洗涤装置3具有洗涤浴槽21,在该洗涤浴槽21中储存有洗涤液12(例如水)。将在凝固浴槽20内凝固的蛋白质导入至洗涤浴槽21中,并通过洗涤液12进行洗涤。该蛋白质通过设置在洗涤浴槽21内的第1轧辊13和第2轧辊14送出至干燥装置4中。例如,通过将第2轧辊14的转速设定为快于第1轧辊13的转速,从而以与转速比相对应的倍率拉伸蛋白质原纤维。另外,18a~18e是导丝器。
[0082]
得到蛋白质原纤维时,在洗涤浴槽21内实施的拉伸也可以是在温水中进行、或在温水中加入有机溶剂等的溶液中等进行的所谓湿热拉伸。作为湿热拉伸温度,例如可以为0~90℃,优选为20~70℃,更优选为30~60℃。湿热拉伸中的未拉伸丝(或预拉伸丝)的拉伸倍率例如可以为1~10倍,也可以为2~8倍。
[0083]
在洗涤液12中拉伸后的蛋白质原纤维从洗涤浴槽21内脱离后,在通过干燥装置4内时被干燥。干燥装置4具有例如干热式干燥炉17。在干燥炉17内设置有送出辊31和卷绕辊32。蛋白质原纤维通过这些送出辊31和卷绕辊32在干燥炉17内停留预定的停留时间,然后送出至高温加热松弛装置60中。在干燥炉17中,例如也可以通过将卷绕辊32的转速设定为快于送出辊31的转速,从而以与转速比相对应的倍率拉伸蛋白质原纤维。在干燥炉17内设置有未图示的加热器。干燥炉17内的温度、即蛋白质原纤维的干燥温度例如为80℃。另外,也可以在洗涤装置3和干燥装置4之间设置上油装置30。
[0084]
高温加热松弛装置60设置在位于蛋白质原纤维的移动方向上的纺丝装置25的下游侧。高温加热松弛装置60例如具有干热式高温加热炉63。在高温加热炉63内设置有送出辊(送出单元)61和卷绕辊(卷绕单元)62。送出辊61和卷绕辊62均为圆筒状,在它们的周面上卷绕有蛋白质原纤维。蛋白质原纤维通过这些送出辊61和卷绕辊62在高温加热炉63内停留预定的停留时间,然后用卷线机卷绕。
[0085]
在高温加热炉63中,例如通过将卷绕辊62的转速设定为慢于送出辊61的转速,从而以与转速比相对应的倍率使蛋白质原纤维松弛。即,送出辊61构成为以预定的送出速度连续地送出蛋白质原纤维。卷绕辊62构成为以比送出辊61的送出速度慢的卷绕速度连续地卷绕由送出辊61送出的蛋白质原纤维。根据以这种方式构成的送出辊61和卷绕辊62,通过过量供应蛋白质原纤维,从而在送出辊61和卷绕辊62之间产生蛋白质原纤维的松弛状态。
[0086]
参照图3,对高温加热松弛装置60进行更详细的说明。在送出辊61和卷绕辊62上连接有速度控制器66。例如,速度控制器66与分别设置在送出辊61和卷绕辊62上的未图示的驱动马达相连接。速度控制器66是由例如cpu、rom以及ram等硬件和存储在rom中的程序等软件构成的计算机。速度控制器66通过控制送出辊61和卷绕辊62这两者或其中的任意一方,如上所述地调节送出速度和/或卷绕速度。即,速度控制器66构成用于对送出辊61的送出速度和卷绕辊62的卷绕速度中的至少一方进行调节的速度调节单元。速度控制器66例如能够调节送出速度和/或卷绕速度,以将送出辊61的送出速度设定为卷绕辊62的卷绕速度的1倍~3倍的范围内的任意比率(松弛倍率)。
[0087]
在高温加热松弛装置60的高温加热炉63内,例如安装有高温加热器64。在该高温加热器64上连接有温度控制器67。温度控制器67是由例如cpu、rom以及ram等硬件和存储在rom中的程序等软件构成的计算机。温度控制器67通过控制高温加热器64来调节高温加热炉63内的温度。也可以将与高温加热炉63内的温度相关的信息通过设置在高温加热炉63上的未图示的温度传感器输入至温度控制器67中,温度控制器67基于该信息控制高温加热器64。温度控制器67构成用于调节高温加热炉63中的蛋白质原纤维的加热温度的温度调节单元。温度控制器67控制高温加热器64,以将高温加热炉63内的加热温度设定为比干燥装置4的干燥炉17中的加热温度高的温度。温度控制器67可以调节蛋白质原纤维的加热温度,以将其设定为例如80~300℃范围内的任意温度。
[0088]
另外,在本实施方式中,分别表示了速度控制器66和温度控制器67,但并不限于这样的方式。例如,速度控制器66和温度控制器67可以组装在一体式的控制器中,也可以在用于控制整个纺丝装置200的控制器中具备相当于速度控制器66和温度控制器67的功能。
[0089]
高温加热松弛装置60构成用于加热蛋白质原纤维的加热单元、以及用于将处于加热状态的蛋白质原纤维在松弛状态下进行收缩的松弛收缩单元。换言之,高温加热松弛装置60是兼作加热单元和松弛收缩单元的装置。在使用高温加热松弛装置60的干式收缩法中,同时执行加热步骤和松弛收缩步骤。如图2所示,在位于蛋白质原纤维的移动方向上的高温加热松弛装置60的下游侧,例如设置有卷线机。蛋白质原纤维在高温加热松弛装置60中进行收缩处理后,用卷线机卷绕,并得到卷绕物5。
[0090]
在松弛收缩步骤中,松弛倍率优选为大于1倍,更优选为1.4倍以上,进一步优选为1.7倍以上,特别优选为2倍以上。所谓松弛倍率,例如,可以作为送出速度与蛋白质原纤维的卷绕速度的比率来把握。
[0091]
(后收缩工序)
[0092]
在后收缩工序中,使经过卷绕工序的蛋白质原纤维在松弛状态下进行收缩。通过实施后收缩工序,可以更可靠地抑制所得到的蛋白质纤维在制备后首次与水分接触时所产生的收缩。如后述的实施例所示,表明在缺少后收缩工序的情况下,制备后首次与水分接触时产生收缩,特别是与高温水分接触时收缩显著。认为这是由于在卷绕工序中进行卷绕时,不可避免地对蛋白质原纤维施加纤维轴方向的张力,从而在蛋白质原纤维中产生残余应力。后收缩工序是以缓和包括卷绕工序中产生的残余应力在内的蛋白质原纤维的总残余应力为目的而实施的工序。
[0093]
在后收缩工序中,作为使蛋白质原纤维在松弛状态下进行收缩的方法,例如,可以列举出包括用于使蛋白质原纤维在松弛状态下与液体或蒸气相接触后收缩的湿式收缩工
序的方法(湿式收缩法)、包括用于使蛋白质原纤维在松弛状态下加热后收缩的干式收缩工序的方法(干式收缩法)。湿式收缩法和干式收缩法的具体实施方式如前收缩工序中所述。
[0094]
图4是示意性地表示蛋白质纤维的制备装置(后收缩装置)的一例的图。图4所示的后收缩装置300用于湿式收缩法,其构成为具有用于送出蛋白质原纤维的进料辊42、用于卷绕蛋白质纤维的卷线机44、用于实施后收缩工序的水浴46、以及用于实施干燥工序的干燥机48。
[0095]
更详细地说,进料辊42可安装蛋白质原纤维36的卷绕物,通过未图示的电动机等的旋转,可从蛋白质原纤维36的卷绕物连续且自动地送出蛋白质原纤维36。卷线机44能够通过未图示的电动机的旋转连续且自动地卷绕从进料辊42送出后经过后收缩工序和干燥工序后制造的蛋白质纤维38。另外,在此,可以相互独立地控制利用进料辊42送出蛋白质原纤维36的送出速度和利用卷线机44卷绕蛋白质纤维38的卷绕速度。
[0096]
水浴46和干燥机48在进料辊42和卷线机44之间分别并列配置在蛋白质原纤维36的进给方向的上游侧和下游侧。另外,图4所示的后收缩装置300具有用于转送从进料辊42向卷线机44行进的蛋白质原纤维36的转送辊50和52。
[0097]
水浴46具有加热器54,通过该加热器54加热的液体(热水)47容纳在水浴46内。此外,在水浴46内,在浸渍于液体(热水)47中的状态下设置有压辊56。由此,从进料辊42送出的蛋白质原纤维36在水浴46内被卷绕在压辊56上,并在被按压的状态下浸渍在液体(热水)47中,同时向卷线机44侧移动。另外,蛋白质原纤维36浸在液体(热水)47中的浸渍时间根据蛋白质原纤维36的移动速度来适当控制。
[0098]
干燥机48具有一对热辊58。一对热辊58可卷绕从水浴46内脱离并向卷线机44侧行进的蛋白质原纤维36。由此,在水浴46内浸渍于液体(热水)47中的蛋白质原纤维36在干燥机48内通过一对热辊58进行加热,使其干燥后,进一步向卷线机44送出。
[0099]
在使用具有这种结构的后收缩装置300来制备目标蛋白质纤维38时,首先,例如,将使用图1所示的纺丝装置100进行纺丝的蛋白质原纤维36的卷绕物5安装在进料辊42上。然后,从进料辊42连续送出蛋白质原纤维36,并在水浴46内将其浸渍于液体(热水)47中。此时,通过控制进料辊42的送出速度与卷线机44的卷绕速度的比率,以使蛋白质原纤维在松弛的状态下通过与液体(热水)47相接触而自然地收缩。例如,作为蛋白质原纤维,当使用在松弛状态下通过将其浸渍在液体(热水)47中以使其在纤维轴方向收缩20%左右的纤维时,如果将卷绕速度和送出速度设为后者比前者慢80%的速度,则可以使蛋白质原纤维在液体(热水)47中以松弛的状态自然地收缩。
[0100]
其次,通过干燥机48的一对热辊58对在水浴46内的液体(热水)47中收缩后的蛋白质原纤维36进行加热。由此,使收缩后的蛋白质原纤维36干燥,并获得蛋白质纤维38。然后,用卷线机44卷绕所得到的蛋白质纤维38,并得到蛋白质纤维38的卷绕物。另外,也可以代替一对热辊58,使用公知的干热板等仅由热源构成的干燥设备来使蛋白质原纤维36干燥。
[0101]
图5是示意性地表示蛋白质纤维的制备装置(后收缩装置)的其他例子的图。图5所示的后收缩装置400用于干式收缩法,其构成为具有用于送出蛋白质原纤维的进料辊42、用于卷绕蛋白质纤维的卷线机44、以及用于实施后收缩工序的高温加热松弛装置60。
[0102]
进料辊42可以安装蛋白质原纤维36的卷绕物。高温加热松弛装置60具有如图3中所示的结构。即,高温加热松弛装置60例如具有干热式高温加热炉63。在高温加热炉63内设
置有送出辊(送出单元)61和卷绕辊(卷绕单元)62。送出辊61和卷绕辊62均为圆筒状,在它们的周面上卷绕有蛋白质原纤维。蛋白质原纤维通过这些送出辊61和卷绕辊62,在高温加热炉63内停留预定的停留时间,并在松弛状态下收缩后,用卷线机44卷绕。
[0103]
图6是示意性地表示蛋白质纤维的制备装置(后收缩装置)的又一例的图。图6(a)是通过蛋白质原纤维的卷绕物得到绞纱的装置的概略图。图6(b)是湿式收缩装置的概略图。图6(c)是干式收缩装置的概略图。
[0104]
图6所示的后收缩装置具有可安装蛋白质原纤维36的卷绕物的进料辊42。首先,例如将使用图1所示的纺丝装置100进行纺丝的蛋白质原纤维36的卷绕物5安装在进料辊42上。其次,从进料辊42送出蛋白质原纤维36,卷绕在支撑体71上,并得到绞纱状的蛋白质原纤维70(图6(a))。绞纱状的蛋白质原纤维70不施加张力,并处于松弛状态。
[0105]
接下来,通过湿式收缩法或干式收缩法使绞纱状的蛋白质原纤维70在松弛状态下进行收缩。图6(b)是湿式收缩法中使用的装置的概略图,图6(c)是干式收缩法中使用的装置的概略图。
[0106]
图6(b)所示的装置具有水浴46、可加热水浴46中的液体的加热器54、以及容纳在水浴46中的液体(例如,水或热水)47。在图6(b)所示的装置中,将绞纱状的蛋白质原纤维70浸渍在加温后的液体(例如热水)47中。由此,可以使蛋白质原纤维在松弛状态下进行收缩。液体的优选方式(种类、温度等)和浸渍时间的优选方式等如前收缩工序(湿式收缩法)中所述。
[0107]
图6(c)所示的装置具有高温加热器64。在图6(c)所示的装置中,将绞纱状的蛋白质原纤维70保持在高温加热器64的上部。由此,加热蛋白质原纤维,并在松弛状态下进行收缩。高温加热器64的温度优选设定为使蛋白质原纤维的温度(加热温度)大于等于蛋白质原纤维中使用的蛋白质的软化温度。加热温度和加热时间的优选方式等如前收缩工序(干式收缩法)中所述。
[0108]
(蛋白质原纤维和蛋白质纤维)
[0109]
按照本发明所涉及的制备方法制备的蛋白质纤维或作为中间产物的蛋白质原纤维含有蛋白质以作为主成分。作为该蛋白质并没有特别限制,可以使用任意的蛋白质。作为蛋白质,可以列举出天然蛋白质和重组蛋白质(人造蛋白质)。作为重组蛋白质,可以列举出可通过工业规模制备的任意的蛋白质,例如,可以列举出可用于工业的蛋白质、可用于医疗的蛋白质、结构蛋白质等。作为可用于工业或医疗的蛋白质的具体例子,可以列举出酶、调节蛋白、受体、肽类激素、细胞因子、膜或转运蛋白、用于预防接种的抗原、疫苗、抗原结合蛋白、免疫刺激蛋白、过敏源、全长抗体或抗体片段或衍生物。作为结构蛋白质的具体例子,可以列举出蜘蛛丝(蛛丝)、蚕丝、角蛋白、胶原蛋白、弹性蛋白、节肢弹性蛋白、以及来源于它们的蛋白质等。作为使用的蛋白质,优选改造丝心蛋白,更优选改造蜘蛛丝丝心蛋白。
[0110]
(改造丝心蛋白)
[0111]
本实施方式所涉及的改造丝心蛋白为包含式1:[(a)n基序-rep]m或式2:[(a)n基序-rep]m-(a)n基序所表示的结构域序列的蛋白质。改造丝心蛋白可以在结构域序列的n末端侧和c末端侧中的任意一个末端侧,或两个末端侧进一步附加有氨基酸序列(n末端序列和c末端序列)。n末端序列及c末端序列并不限定于此,典型地,不具有丝心蛋白中的特征性的氨基酸基序的重复的区域,并且由100个残基左右的氨基酸构成。
[0112]
在本说明书中,“改造丝心蛋白”是指人工制造的丝心蛋白(人造丝心蛋白)。改造丝心蛋白可以是其结构域序列与天然来源的丝心蛋白的氨基酸序列不同的丝心蛋白,也可以是与天然来源的丝心蛋白的氨基酸序列相同的丝心蛋白。在本说明书中,“天然来源的丝心蛋白”也是包含式1:[(a)n基序-rep]m或式2:[(a)n基序-rep]m-(a)n基序所表示的结构域序列的蛋白质。
[0113]“改造丝心蛋白”可以是直接利用天然来源的丝心蛋白的氨基酸序列的丝心蛋白,也可以是依据天然来源的丝心蛋白的氨基酸序列来改造该氨基酸序列的丝心蛋白(例如,通过对克隆出的天然来源的丝心蛋白的基因序列进行改造来改造氨基酸序列而得到的丝心蛋白),此外也可以为不依赖于天然来源的丝心蛋白而人工设计和合成的丝心蛋白(例如,通过对编码所设计的氨基酸序列的核酸进行化学合成而具有需要的氨基酸序列的丝心蛋白)。
[0114]
在本说明书中,“结构域序列”为生成丝心蛋白特有的结晶区(典型地,相当于氨基酸序列的(a)n基序)和非晶区(典型地,相当于氨基酸序列的rep)的氨基酸序列,是指式1:[(a)n基序-rep]m或式2:[(a)n基序-rep]m-(a)n基序所表示的氨基酸序列。其中,(a)n基序表示以丙氨酸残基为主的氨基酸序列,氨基酸残基数为2~27个。(a)n基序的氨基酸残基数可以是2~20、4~27、4~20、8~20、10~20、4~16、8~16或10~16的整数。此外,只要(a)n基序中的丙氨酸残基数相对于氨基酸残基总数的比例为40%以上即可,可以为60%以上、70%以上、80%以上、83%以上、85%以上、86%以上、90%以上、95%以上或100%(意味着其仅由丙氨酸残基构成)也可以。在结构域序列中存在的多个(a)n基序中的至少7个可以仅由丙氨酸残基构成。rep表示由2~200个氨基酸残基构成的氨基酸序列。rep可以是由10~200个氨基酸残基构成的氨基酸序列。m表示2~300的整数,也可以为10~300的整数。多个存在的(a)n基序彼此可以为相同的氨基酸序列,也可以为不同的氨基酸序列。存在的多个rep可以是彼此相同的氨基酸序列,也可以是彼此不同的氨基酸序列。
[0115]
本实施方式所涉及的改造丝心蛋白例如可以通过对克隆出的天然来源的丝心蛋白的基因序列进行例如与置换、缺失、插入及/或添加一个或多个氨基酸残基相当的氨基酸序列的改造而得到的改造丝心蛋白。氨基酸残基的置换、缺失、插入及/或添加可以通过定点突变方法等本领域技术人员熟知的方法来进行。具体而言,可以根据nucleic acid res.10,6487(1982)、methods in enzymology,100,448(1983)等文献中记载的方法来进行。
[0116]
天然来源的丝心蛋白是包含式1:[(a)n基序-rep]m、或者式2:[(a)n基序-rep]m-(a)n基序所表示的结构域序列的蛋白质,具体而言,例如,可以列举出昆虫或蜘蛛类所产生的丝心蛋白。
[0117]
作为昆虫产生的丝心蛋白,可以列举例如家蚕(bombyx mori)、野桑蚕(bombyx mandarina)、天蚕(antheraea yamamai)、柞蚕(anteraea pernyi)、枫蚕(eriogyna pyretorum)、蓖麻蚕(pilosamia cynthia ricini)、樗蚕(samia cynthia)、樟蚕(caligura japonica)、印度柞蚕(antheraea mylitta)、琥珀蚕(antheraea assama)等蚕产生的蚕丝蛋白及雀蜂(vespa simillima xanthoptera)的幼虫吐出的蜂丝蛋白。
[0118]
作为昆虫产生的丝心蛋白的更具体的示例,可以列举例如蚕
·
丝心蛋白l链(genbank登录号m76430(碱基序列)及aaa27840.1(氨基酸序列))。
[0119]
作为蜘蛛类产生的丝心蛋白,例如,可以列举出属于蜘蛛目(araneae)的蜘蛛产生的蛛丝蛋白。更具体而言,可以列举大腹园蛛、十字园蛛、肥胖园蛛、五纹园蛛和野岛园蛛等属于园蛛属(araneus属)的蜘蛛、青新园蛛、嗜水新园蛛、灌木新园蛛和类青新园蛛等属于新园蛛属(neoscona属)的蜘蛛、小岬蛛等属于岬蛛属(pronus属)的蜘蛛、蟾蜍曲腹蛛和对称曲腹蛛等属于曲腹蛛属(cyrtarachne属)的蜘蛛、库氏棘腹蛛和乳突棘腹蛛等属于棘腹蛛属(gasteracantha属)的蜘蛛、何氏瘤腹蛛和六刺瘤腹蛛等属于瘤腹蛛属(ordgarius属)的蜘蛛、悦目金蛛、小悦目金蛛和横纹金蛛等属于金蛛属(argiope属)的蜘蛛、双峰尾园蛛等属于尾园蛛属(arachnura属)的蜘蛛、褐吊叶蛛等属于吊叶蛛属(acusilas属)的蜘蛛、红云斑蛛、花云斑蛛和全色云斑蛛等属于云斑蛛属(cytophora属)的蜘蛛、丑锥头蛛等属于锥头蛛属(poltys属)的蜘蛛、八瘤艾蛛、四突艾蛛、圆腹艾蛛和黑尾艾蛛等属于艾蛛属(cyclosa属)的蜘蛛和日本壮头蛛等属于壮头蛛属(chorizopes属)的蜘蛛所产生的蛛丝蛋白、以及前齿肖蛸、锥腹肖蛸、直伸肖蛸和鳞纹肖蛸等属于肖蛸属(tetragnatha属)的蜘蛛、纵条银鳞蛛、肩斑银鳞蛛和小肩斑银鳞蛛等属于银鳞蛛属(leucauge属)的蜘蛛、棒络新妇和斑络新妇等属于络新妇属(nephila属)的蜘蛛、美丽麦蛛等属于麦蛛属(menosira属)的蜘蛛、柔弱粗螯蛛等属于锯螯蛛属(dyschiriognatha属)的蜘蛛、红斑寇蛛、哈氏寇蛛、几何寇蛛和间斑寇蛛等属于寇蛛属(latrodectus属)的蜘蛛、以及属于育儿网蛛属(euprosthenops属)的蜘蛛等属于肖蛸科(tetragnathidae科)的蜘蛛所产生的蛛丝蛋白。作为蛛丝蛋白,可以列举例如masp(masp1和masp2)、adf(adf3和adf4)等牵引丝蛋白、misp(misp1和misp2)、acsp、pysp、flag等。
[0120]
作为蜘蛛类所产生的蜘蛛丝蛋白的更具体的例子,例如,可以列举出fibroin-3(adf-3)[来自于araneus diadematus](genbank登录号aac47010(氨基酸序列)、u47855(碱基序列))、fibroin-4(adf-4)[来自于araneus diadematus](genbank登录号aac47011(氨基酸序列)、u47856(碱基序列))、dragline silk protein spidroin 1[来自于nephila clavipes](genbank登录号aac04504(氨基酸序列)、u37520(碱基序列))、major ampullate spidroin 1[来自于latrodectus hesperus](genbank登录号abr68856(氨基酸序列)、ef595246(碱基序列))、dragline silk protein spidroin 2[来自于nephila clavata](genbank登录号aal32472(氨基酸序列)、af441245(碱基序列))、major ampullate spidroin 1[来自于euprosthenops australis](genbank登录号caj00428(氨基酸序列)、aj973155(碱基序列))、以及major ampullate spidroin2[euprosthenops australis](genbank登录号cam32249.1(氨基酸序列)、am490169(碱基序列))、minor ampullate silk protein 1[nephila clavipes](genbank登录号aac14589.1(氨基酸序列))、minor ampullate silk protein 2[nephila clavipes](genbank登录号aac14591.1(氨基酸序列))、minor ampullate spidroin-like protein[nephilengys cruentata](genbank登录号abr37278.1(氨基酸序列)等。
[0121]
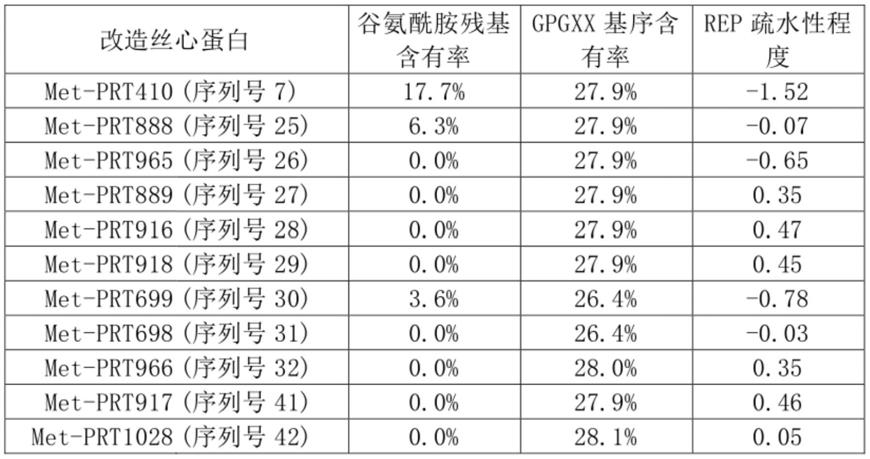
作为天然来源的丝心蛋白的更具体的示例,可以进一步列举在ncbi genbank中登记了序列信息的丝心蛋白。例如,可以通过从ncbi genbank中登记的序列信息中包含inv作为分类码(division)的序列中提取出在定义(definition)中记载了蛛丝蛋白、壶状腺、丝心蛋白、“丝和多肽”或“丝和蛋白质”作为关键词的序列、从cds中提取出指定产物的字符串、从来源(source)中提取出在组织类型(tissue type)中记载了指定字符串的序列来确
认。
[0122]
本实施方式所涉及的改造丝心蛋白可以是改造蚕丝(丝绸)丝心蛋白(改造了蚕所产生的蚕丝蛋白的氨基酸序列的丝心蛋白),也可以是改造蜘蛛丝丝心蛋白(改造了蜘蛛类所产生的蛛丝蛋白的氨基酸序列的丝心蛋白)。
[0123]
作为改造丝心蛋白的具体实施例,可以列举出来自于蜘蛛的大壶状腺所产生的大吐丝管牵引丝蛋白的改造丝心蛋白(第1改造丝心蛋白)、具有降低了甘氨酸残基含量的结构域序列的改造丝心蛋白(第2改造丝心蛋白)、具有降低了(a)n基序含量的结构域序列的改造丝心蛋白(第3改造丝心蛋白)、降低了甘氨酸残基的含量及(a)n基序的含量的改造丝心蛋白(第4改造丝心蛋白)、具有局部包含疏水性指标大的区域的结构域序列的改造丝心蛋白(第5改造丝心蛋白)、及具有降低了谷氨酰胺残基的含量的结构域序列的改造丝心蛋白(第6改造丝心蛋白)。
[0124]
作为第1改造丝心蛋白可以例举包含式1:[(a)n基序-rep]m所示的结构域序列的蛋白质。在第1改造丝心蛋白中,(a)n基序的氨基酸残基数优选为3~20的整数,更优选为4~20的整数,进一步优选为8~20的整数,更进一步优选为10~20的整数,进一步还优选为4~16的整数,特别优选为8~16的整数,最优选为10~16的整数。在第1改造丝心蛋白,式1中构成rep的氨基酸残基数优选为10~200个残基,更优选为10~150个残基,进一步优选为20~100个残基,更进一步优选为20~75个残基。在第1改造丝心蛋白中,式1:[(a)n基序-rep]m所示的氨基酸序列所含的甘氨酸残基、丝氨酸残基及丙氨酸残基的合计残基数相对于氨基酸残基总数优选为40%以上,更优选为60%以上,进一步优选为70%以上。
[0125]
第1改造丝心蛋白可以为以下这种多肽:包含式1:[(a)n基序-rep]m所示的氨基酸序列的单元,并且c末端序列为序列号1~3中任意一个所示的氨基酸序列、或与序列号1~3中任意一个所示的氨基酸序列具有90%以上的同源性的氨基酸序列。
[0126]
序列号1所示的氨基酸序列与由adf3(gi:1263287、ncbi)的氨基酸序列的c末端的50个残基的氨基酸构成的氨基酸序列相同,序列号2所示的氨基酸序列与从序列号1所示的氨基酸序列的c末端除去20个残基后的氨基酸序列相同,序列号3所示的氨基酸序列与从序列号1所示的氨基酸序列的c末端除去29个残基后的氨基酸序列相同。
[0127]
作为第1改造丝心蛋白的更具体的例子,可以列举出包括与(1-i)序列号4(recombinant spider silk protein adf3kailargenrsh1)所示的氨基酸序列、或者(1-ii)序列号4所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列的改造丝心蛋白。序列一致性优选为95%以上。
[0128]
序列号4所示的氨基酸序列为在n末端添加有由起始密码子、his10标签及hrv3c蛋白酶(human rhinovirus 3c蛋白酶)识别位点构成的氨基酸序列(序列号5)的adf3的氨基酸序列中,以使第1~13位的重复区域增加为约2倍、并且在第1154位氨基酸残基处终止翻译的方式突变后得到。序列号4所示的氨基酸序列的c末端的氨基酸序列与序列号3所示的氨基酸序列相同。
[0129]
(1-i)的改造丝心蛋白可以由序列号4所示的氨基酸序列构成。
[0130]
第2改造丝心蛋白具有其结构域序列与天然来源的丝心蛋白相比甘氨酸残基的含量降低的氨基酸序列。第2改造丝心蛋白与天然来源的丝心蛋白相比,可以具有与至少rep中的一个或多个甘氨酸残基被置换为其他氨基酸残基相当的氨基酸序列。
[0131]
第2改造丝心蛋白的结构域序列与天然来源的丝心蛋白相比,在rep中选自ggx及gpgxx(其中,g表示甘氨酸残基,p表示脯氨酸残基,x表示甘氨酸以外的氨基酸残基)中的至少一种基序序列中,可以具有相当于至少一个或多个该基序序列中的一个甘氨酸残基被置换为其他氨基酸残基的氨基酸序列。
[0132]
第2改造丝心蛋白中,上述甘氨酸残基被置换为其他氨基酸残基的基序序列的比例相对于全部基序序列可以为10%以上。
[0133]
第2改造丝心蛋白包括式1:[(a)n基序-rep]m所示的结构域序列并且可以具有下述氨基酸序列:当将从上述结构域序列中除去自位于最靠c末端侧的(a)n基序至上述结构域序列的c末端为止的序列后得到的序列中的全部rep所包含的由xgx(其中,x表示甘氨酸以外的氨基酸残基)构成的氨基酸序列的氨基酸残基总数设为z、并且将从上述结构域序列中除去自位于最靠c末端侧的(a)n基序至上述结构域序列的c末端为止的序列后得到的序列中的氨基酸残基总数设为w时,z/w为30%以上、40%以上、50%以上或50.9%以上。(a)n基序中的丙氨酸残基数相对于氨基酸残基总数可以为83%以上,优选为86%以上,更优选为90%以上,进一步优选为95%以上,更进一步优选为100%(意味着仅由丙氨酸残基构成)。
[0134]
第2改造丝心蛋白优选地通过将ggx基序中的一个甘氨酸残基置换为其他氨基酸残基来提高由xgx构成的氨基酸序列的含有比例。在第2改造丝心蛋白中,结构域序列中,由ggx构成的氨基酸序列的含有比例优选为30%以下,更优选为20%以下,进一步优选为10%以下,更进一步优选为6%以下,进一步还优选为4%以下,特别优选为2%以下。结构域序列中的由ggx构成的氨基酸序列的含有比例可以通过与下述的由xgx构成的氨基酸序列的含有比例(z/w)的计算方法相同的方法来计算。
[0135]
对z/w的计算方法进一步进行详细说明。首先,在包含式1:[(a)n基序-rep]m所示的结构域序列的丝心蛋白(改造丝心蛋白或天然来源的丝心蛋白)中,从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列所含的全部rep中,提取由xgx构成的氨基酸序列。构成xgx的氨基酸残基的总数为z。例如,当提取了50个由xgx构成的氨基酸序列时(没有重复),z为50
×
3=150。此外,例如,如由xgxgx构成的氨基酸序列的情况那样,当包含在2个xgx中的x(中间的x)存在时,通过扣除重复部分来进行计算(当为xgxgx时,为5个氨基酸残基)。w为从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列所含的氨基酸残基总数。例如,当为图7所示的结构域序列时,w为4 50 4 100 4 10 4 20 4 30=230(除去位于最靠c末端侧的(a)n基序)。然后,通过将z除以w,可以计算出z/w(%)。
[0136]
在此,对天然来源的丝心蛋白中的z/w进行说明。首先,如上所述,通过例示在ncbi genbank中登记有氨基酸序列信息的丝心蛋白的方法进行确认,提取出663种丝心蛋白(其中,来源于蜘蛛类的丝心蛋白为415种)。在所提取的所有丝心蛋白中,从包含式1:[(a)n基序-rep]m所表示的结构域序列且由丝心蛋白中的ggx构成的氨基酸序列的含有比例为6%以下的天然来源的丝心蛋白的氨基酸序列中,根据上述计算方法,计算出z/w。其结果如图8所示。图8的横轴表示z/w(%),纵轴表示频率。由图8可知,天然来源的丝心蛋白中的z/w均小于50.9%(最大为50.86%)。
[0137]
在第2改造丝心蛋白中,z/w优选为50.9%以上,更优选为56.1%以上,进一步优选
为58.7%以上,更进一步优选为70%以上,进一步还优选为80%以上。z/w的上限并没有特别限制,例如可以为95%以下。
[0138]
例如可以通过如下方式进行改造而获得第2改造丝心蛋白:从克隆出的天然来源的丝心蛋白的基因序列中,对编码甘氨酸残基的碱基序列的至少一部分进行置换并使其编码其他氨基酸残基。此时,作为改造的甘氨酸残基,可以选择ggx基序及gpgxx基序中的一个甘氨酸残基、此外也可以用将z/w设定为50.9%以上的方式进行置换。此外,例如,也可以通过根据天然来源的丝心蛋白的氨基酸序列设计满足上述方式的氨基酸序列,并通过化学合成编码所设计的氨基酸序列的核酸来获得。在任何一种情况下,除了与在从天然来源的丝心蛋白的氨基酸序列中将rep中的甘氨酸残基置换为其他氨基酸残基相当的改造之外,还可以进一步进行与置换、缺失、插入及/或添加一个或多个氨基酸残基相当的氨基酸序列的改造。
[0139]
作为上述的其他氨基酸残基,只要是除甘氨酸残基以外的氨基酸残基即可,并没有特别限制,优选缬氨酸(v)残基、亮氨酸(l)残基、异亮氨酸(i)残基、蛋氨酸(m)残基、脯氨酸(p)残基、苯丙氨酸(f)残基及色氨酸(w)残基等疏水性氨基酸残基、谷氨酰胺(q)残基、天冬酰胺(n)残基、丝氨酸(s)残基、赖氨酸(k)残基及谷氨酸(e)残基等亲水性氨基酸残基,更优选缬氨酸(v)残基、亮氨酸(l)残基、异亮氨酸(i)残基、苯丙氨酸(f)残基及谷氨酰胺(q)残基,进一步优选谷氨酰胺(q)残基。
[0140]
作为第2改造丝心蛋白的更具体的例子,可以列举出包括与(2-i)序列号6(met-prt380)、序列号7(met-prt410)、序列号8(met-prt525)或序列号9(met-prt799)所示的氨基酸序列、或者(2-ii)序列号6、序列号7、序列号8或序列号9所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列的改造丝心蛋白。
[0141]
对(2-i)的改造丝心蛋白进行说明。序列号6所表示的氨基酸序列是将相当于天然来源的丝心蛋白的序列号10(met-prt313)所表示的氨基酸序列的rep中的ggx全部置换为gqx而得到的氨基酸序列。序列号7所示的氨基酸序列,通过在序列号6所示的氨基酸序列中从n末端侧向c末端侧使(a)n基序每隔2个发生缺失,并进一步在c末端序列的近前插入一个[(a)n基序-rep]而得到。序列号8所示的氨基酸序列,在序列号7所示的氨基酸序列的各个(a)n基序的c末端侧插入2个丙氨酸残基,进一步将部分谷氨酰胺(q)残基置换为丝氨酸(s)残基、并且使c末端侧的部分氨基酸缺失以使得其与序列号7的分子量几乎相同。序列号9所示的氨基酸序列,在将序列号7所示的氨基酸序列中存在的20个结构域序列的区域(其中,该区域的c末端侧的数个氨基酸残基被置换)重复4次后的序列的c末端中添加预定的铰链序列和his标签序列。
[0142]
序列号10所示的氨基酸序列(相当于天然来源的丝心蛋白)中的z/w的值为46.8%。序列号6所示的氨基酸序列、序列号7所示的氨基酸序列、序列号8所示的氨基酸序列及序列号9所示的氨基酸序列中的z/w的值分别为58.7%、70.1%、66.1%及70.0%。此外,序列号10、序列号6、序列号7、序列号8及序列号9所示的氨基酸序列的锯齿比率(在下文中说明)为1:1.8~11.3的x/y的值分别为15.0%、15.0%、93.4%、92.7%及89.8%。
[0143]
(2-i)的改造丝心蛋白可以由序列号6、序列号7、序列号8或序列号9所示的氨基酸序列构成。
[0144]
(2-ii)的改造丝心蛋白包括与序列号6、序列号7、序列号8或序列号9所示的氨基
酸序列具有90%以上的序列一致性的氨基酸序列。(2-ii)的改造丝心蛋白也为包含式1:[(a)n基序-rep]m所示的结构域序列的蛋白质。上述序列一致性优选为95%以上。
[0145]
(2-ii)的改造丝心蛋白与序列号6、序列号7、序列号8或序列号9所示的氨基酸序列具有90%以上的序列一致性,并且当将rep中所含的由xgx(其中,x表示甘氨酸以外的氨基酸残基)构成的氨基酸序列的氨基酸残基总数设为z、将上述结构域序列中的rep的氨基酸残基总数设为w时,z/w优选为50.9%以上。
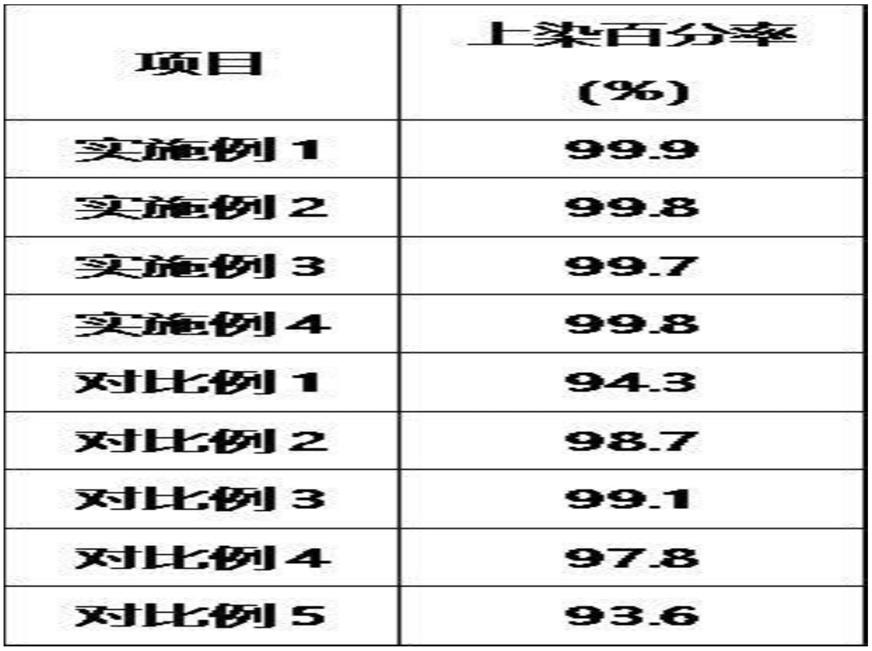
[0146]
第2改造丝心蛋白可以在n末端及c末端中的任意一端或两端包含标签序列。由此,可以实现改造丝心蛋白的分离、固定、检测及可视化等。
[0147]
作为标签序列,例如,可以列举出利用了与其他分子的特异性亲和性(结合性、亲和性)的亲和性标签。作为亲和性标签的具体实施例,可以列举出组氨酸标签(his标签)。his标签是由4至10个左右的组氨酸残基排列而成的短肽,具有与镍等的金属离子特异性结合的性质,因此,可以用于通过金属螯合层析(chelating metal chromatography)进行的改造丝心蛋白的分离。作为标签序列的具体实施例,例如,可以列举出序列号11所示的氨基酸序列(包含his标签序列及铰链序列的氨基酸序列)。
[0148]
此外,也可以利用与谷胱甘肽特异性结合的谷胱甘肽-s-转移酶(gst)、与麦芽糖特异性结合的麦芽糖结合蛋白(mbp)等标签序列。
[0149]
更进一步地,还可以使用利用了抗原抗体反应的“表位标签”。通过添加用于显示抗原性的肽(表位)作为标签序列,可以结合针对该表位的抗体。作为表位标签,可以列举出ha(流感病毒血凝素的肽序列)标签、myc标签、flag标签等。通过利用表位标签,可以很容易地以高特异性对改造丝心蛋白进行纯化。
[0150]
更进一步地,也可以使用利用特定的蛋白酶将标签序列切除而得到的标签。也可以通过对经由该标签序列吸附的蛋白质进行蛋白酶处理,来回收切除标签序列后的改造丝心蛋白。
[0151]
作为含有标签序列的改造丝心蛋白的更具体的例子,可以列举出包括与(2-iii)序列号12(prt380)、序列号13(prt410)、序列号14(prt525)或序列号15(prt799)所示的氨基酸序列、或者(2-iv)序列号12、序列号13、序列号14或序列号15所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列的改造丝心蛋白。
[0152]
序列号16(prt313)、序列号12、序列号13、序列号14及序列号15所示的氨基酸序列分别在序列号10、序列号6、序列号7、序列号8及序列号9所示的氨基酸序列的n末端添加序列号11所示的氨基酸序列(包含his标签序列及铰链序列)。
[0153]
(2-iii)的改造丝心蛋白可以由序列号12、序列号13、序列号14或序列号15所示的氨基酸序列构成。
[0154]
(2-iv)的改造丝心蛋白包括与序列号12、序列号13、序列号14或序列号15所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列。(2-iv)的改造丝心蛋白也为包含式1:[(a)n基序-rep]m所示的结构域序列的蛋白质。上述序列一致性优选为95%以上。
[0155]
(2-iv)的改造丝心蛋白与序列号12、序列号13、序列号14或序列号15所示的氨基酸序列具有90%以上的序列一致性,并且当将rep中所含的由xgx(其中,x表示甘氨酸以外的氨基酸残基)构成的氨基酸序列的氨基酸残基总数设为z、将上述结构域序列中的rep的氨基酸残基总数设为w时,z/w优选为50.9%以上。
[0156]
第2改造丝心蛋白可以含有用于将重组蛋白生产系统所生产的蛋白质释放到宿主外部的分泌信号。分泌信号的序列可以根据宿主的种类来进行适当设定。
[0157]
第3改造丝心蛋白具有其结构域序列与天然来源的丝心蛋白相比(a)n基序的含量降低的氨基酸序列。第3改造丝心蛋白的结构域序列与天然来源的丝心蛋白相比,可以具有相当于缺失了至少一个或多个(a)n基序的氨基酸序列。
[0158]
第3改造丝心蛋白可以具有相当于从天然来源的丝心蛋白中缺失了10~40%的(a)n基序的氨基酸序列。
[0159]
第3改造丝心蛋白的结构域序列与天然来源的丝心蛋白相比,其可以至少从n末端侧向c末端侧具有相当于每1~3个(a)n基序中缺失一个(a)n基序的氨基酸序列。
[0160]
第3改造丝心蛋白的结构域序列与天然来源的丝心蛋白相比,其可以至少从n末端侧向c末端侧具有相当于依次重复发生了两个连续的(a)n基序的缺失及一个(a)n基序的缺失的氨基酸序列。
[0161]
第3改造丝心蛋白的结构域序列可以至少从n末端侧向c末端侧具有相当于每隔2个发生一次缺失(a)n基序的氨基酸序列。
[0162]
第3改造丝心蛋白包括式1:[(a)n基序-rep]m所示的结构域序列并且可以具有下述氨基酸序列:从n末端侧向c末端侧依次比较相邻的2个[(a)n基序-rep]单元的rep的氨基酸残基数,将氨基酸残基数少的rep的氨基酸残基数设为1时,并且将另一个rep的氨基酸残基数的比为1.8~11.3的相邻的2个[(a)n基序-rep]单元的氨基酸残基数相加而得到的合计值的最大值设为x、将结构域序列的氨基酸残基总数设为y时,x/y为20%以上、30%以上、40%以上或50%以上。(a)n基序中的丙氨酸残基数相对于氨基酸残基总数可以为83%以上,优选为86%以上,更优选为90%以上,进一步优选为95%以上,更进一步优选为100%(意味着仅由丙氨酸残基构成)。
[0163]
参照图7进一步地详细说明x/y的计算方法。图7示出了从改造丝心蛋白中除去n末端序列及c末端序列后的结构域序列。该结构域序列从n末端侧(左侧)起具有(a)n基序-第1rep(50个氨基酸残基)-(a)n基序-第2rep(100个氨基酸残基)-(a)n基序-第3rep(10个氨基酸残基)-(a)n基序-第4rep(20个氨基酸残基)-(a)n基序-第5rep(30个氨基酸残基)-(a)n基序的序列。
[0164]
以不重复的方式从n末端侧向c末端侧依次选择相邻的2个[(a)n基序-rep]单元。此时,也可以存在未被选择的[(a)n基序-rep]单元。图7示出了模式1(第1rep与第2rep的比较、以及第3rep与第4rep的比较)、模式2(第1rep与第2rep的比较、以及第4rep与第5rep的比较)、模式3(第2rep与第3rep的比较、以及第4rep与第5rep的比较)、模式4(第1rep与第2rep的比较)。另外,除此之外还存在选择方法。
[0165]
接下来,对于各模式,比较所选择的相邻的2个[(a)n基序-rep]单元中的各rep的氨基酸残基数。比较的方法是确定氨基酸残基数量较少的那一个与氨基酸残基数量较多的那一个的氨基酸残基数量之比。例如,在比较第1rep(50个氨基酸残基)和第2rep(100个氨基酸残基)的情况下,当将氨基酸残基数相对较少的第1rep设为1时,第2rep的氨基酸残基数的比为100/50=2。同样地,在比较第4rep(20个氨基酸残基)和第5rep(30个氨基酸残基)的情况下,当将氨基酸残基数相对较少的第4rep设为1时,第5rep的氨基酸残基数的比为30/20=1.5。
[0166]
图7中,用实线表示当将氨基酸残基数相对较少的一方设为1时,另一方的氨基酸残基数的比为1.8~11.3的[(a)n基序-rep]单元的组。在本说明书中,将该比率称为锯齿比率。用虚线表示当将氨基酸残基数相对较少的一方设为1时,另一方的氨基酸残基数的比小于1.8或大于11.3的[(a)n基序-rep]单元的组。
[0167]
在各个模式中,将实线所示的相邻的2个[(a)n基序-rep]单元的所有氨基酸残基数相加(不仅仅是rep,也是(a)n基序的氨基酸残基数)。而且,将相加后的合计值进行比较,并将该合计值最大的模式的合计值(合计值的最大值)设为x。在图7所示的例子中,模式1的合计值最大。
[0168]
接下来,可以通过将x除以结构域序列的氨基酸残基总数y来计算出x/y(%)。
[0169]
在第3改造丝心蛋白中,x/y优选为50%以上,更优选为60%以上,进一步优选为65%以上,更进一步优选为70%以上,进一步还优选为75%以上,特别优选为80%以上。x/y的上限并没有特别限制,例如可以为100%以下。当锯齿比率为1:1.9~11.3时,x/y优选为89.6%以上,当锯齿比率为1:1.8~3.4时,x/y优选为77.1%以上,当锯齿比率为1:1.9~8.4时,x/y优选为75.9%以上,当锯齿比率为1:1.9~4.1时,x/y优选为64.2%以上。
[0170]
当第3改造丝心蛋白为在结构域序列中存在多个(a)n基序中的至少7个仅由丙氨酸残基构成的改造丝心蛋白时,x/y优选为46.4%以上,更优选为50%以上,进一步优选为55%以上,更进一步优选为60%以上,进一步还优选为70%以上,特别优选为80%以上。x/y的上限并没有特别限制,为100%以下即可。
[0171]
在此,对天然来源的丝心蛋白中的x/y进行说明。首先,如上所述,通过例示在ncbi genbank中登记有氨基酸序列信息的丝心蛋白的方法进行确认,提取出663种丝心蛋白(其中,来源于蜘蛛类的丝心蛋白为415种)。在所提取的所有丝心蛋白中,从由式1:[(a)n基序-rep]m所表示的结构域序列构成的天然来源的丝心蛋白的氨基酸序列中,根据上述计算方法,计算出x/y。图9表示锯齿比率为1:1.9~4.1时的结果。
[0172]
图9的横轴表示x/y(%),纵轴表示频率。由图9可知,天然来源的丝心蛋白中的x/y均小于64.2%(最大为64.14%)。
[0173]
例如可以通过从克隆出的天然来源的丝心蛋白的基因序列中,以将x/y设为64.2%以上的方式使编码(a)n基序的序列中的一个或多个缺失,从而得到第3改造丝心蛋白。此外,例如也可以通过设计出相当于从天然来源的丝心蛋白的氨基酸序列中,以将x/y设为64.2%以上的方式使一个或多个(a)n基序发生缺失的氨基酸序列,并且对编码所设计的氨基酸序列的核酸进行化学合成后得到。在任何情况下,除了与在从天然来源的丝心蛋白的氨基酸序列中缺失(a)n基序相当的改造之外,还可以进一步进行与置换、缺失、插入及/或添加一个或多个氨基酸残基相当的氨基酸序列的改造。
[0174]
作为第3改造丝心蛋白的更具体的例子,可以列举出包括与(3-i)序列号17(met-prt399)、序列号7(met-prt410)、序列号8(met-prt525)或序列号9(met-prt799)所示的氨基酸序列、或者(3-ii)序列号17、序列号7、序列号8或序列号9所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列的改造丝心蛋白。
[0175]
对(3-i)的改造丝心蛋白进行说明。序列号17所示的氨基酸序列,通过在相当于天然来源的丝心蛋白的序列号10(met-prt313)所示的氨基酸序列中,从n末端侧向c末端侧使(a)n基序每隔2个发生缺失,并进一步在c末端序列的近前插入一个[(a)n基序-rep]而得
到。序列号7、序列号8或序列号9所示的氨基酸序列如第2改造丝心蛋白中所说明的那样。
[0176]
序列号10所示的氨基酸序列(相当于天然来源的丝心蛋白)的锯齿比率为1:1.8~11.3中的x/y的值为15.0%。序列号17所示的氨基酸序列及序列号7所示的氨基酸序列中的x/y的值均为93.4%。序列号8所示的氨基酸序列中的x/y的值为92.7%。序列号9所示的氨基酸序列中的x/y的值为89.8%。序列号10、序列号17、序列号7、序列号8及序列号9所示的氨基酸序列中的z/w的值分别为46.8%、56.2%、70.1%、66.1%及70.0%。
[0177]
(3-i)的改造丝心蛋白可以由序列号17、序列号7、序列号8或序列号9所示的氨基酸序列构成。
[0178]
(3-ii)的改造丝心蛋白包括与序列号17、序列号7、序列号8或序列号9所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列。(3-ii)的改造丝心蛋白也为包含式1:[(a)n基序-rep]m所示的结构域序列的蛋白质。上述序列一致性优选为95%以上。
[0179]
(3-ii)的改造丝心蛋白与序列号17、序列号7、序列号8或序列号9所示的氨基酸序列具有90%以上的序列一致性,并且从n末端侧向c末端侧依次比较相邻的2个[(a)n基序-rep]单元的rep的氨基酸残基数,将氨基酸残基数少的rep的氨基酸残基数设为1时,并且将另一个rep的氨基酸残基数的比为1.8~11.3(锯齿比率为1:1.8~11.3)的相邻的2个[(a)n基序-rep]单元的氨基酸残基数相加而得到的合计值的最大值设为x、将结构域序列的氨基酸残基总数设为y时,x/y优选为64.2%以上。
[0180]
第3改造丝心蛋白可以在n末端及c末端中的任意一端或两端包含上述标签序列。
[0181]
作为含有标签序列的改造丝心蛋白的更具体的例子,可以列举出包括与(3-iii)序列号18(prt399)、序列号13(prt410)、序列号14(prt525)或序列号15(prt799)所示的氨基酸序列、或者(3-iv)序列号18、序列号13、序列号14或序列号15所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列的改造丝心蛋白。
[0182]
序列号18、序列号13、序列号14及序列号15所示的氨基酸序列分别在序列号17、序列号7、序列号8及序列号9所示的氨基酸序列的n末端添加序列号11所示的氨基酸序列(包含his标签序列及铰链序列)。
[0183]
(3-iii)的改造丝心蛋白可以由序列号18、序列号13、序列号14或序列号15所示的氨基酸序列构成。
[0184]
(3-iv)的改造丝心蛋白包括与序列号18、序列号13、序列号14或序列号15所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列。(3-iv)的改造丝心蛋白也为包含式1:[(a)n基序-rep]m所示的结构域序列的蛋白质。上述序列一致性优选为95%以上。
[0185]
(3-iv)的改造丝心蛋白与序列号18、序列号13、序列号14或序列号15所示的氨基酸序列具有90%以上的序列一致性,并且从n末端侧向c末端侧依次比较相邻的2个[(a)n基序-rep]单元的rep的氨基酸残基数,将氨基酸残基数少的rep的氨基酸残基数设为1时,并且将另一个rep的氨基酸残基数的比为1.8~11.3的相邻的2个[(a)n基序-rep]单元的氨基酸残基数相加而得到的合计值的最大值设为x、将结构域序列的氨基酸残基总数设为y时,x/y优选为64.2%以上。
[0186]
第3改造丝心蛋白可以含有用于将重组蛋白生产系统所生产的蛋白质释放到宿主外部的分泌信号。分泌信号的序列可以根据宿主的种类来进行适当设定。
[0187]
第4改造丝心蛋白是其结构域序列与天然来源的丝心蛋白相比,除了降低(a)n基
序的含量之外,还具有降低甘氨酸残基含量的氨基酸序列的丝心蛋白。第4改造丝心蛋白的结构域序列与天然来源的丝心蛋白相比,其除了至少一个或多个(a)n基序缺失之外,还可以具有相当于至少rep中的一个或多个甘氨酸残基被其他氨基酸残基置换的氨基酸序列。即,第4改造丝心蛋白是同时具有上述的第2改造丝心蛋白和第3改造丝心蛋白的特征的改造丝心蛋白。具体的实施方式等如第2改造丝心蛋白及第3改造丝心蛋白中所说明的那样。
[0188]
作为第4改造丝心蛋白的更具体的例子,可以列举出包括与(4-i)序列号7(met-prt410)、序列号8(met-prt525)、序列号9(met-prt799)、序列号13(prt410)、序列号14(prt525)或序列号15(prt799)所示的氨基酸序列、或者(4-ii)序列号7、序列号8、序列号9、序列号13、序列号14或序列号15所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列的改造丝心蛋白。包含序列号7、序列号8、序列号9、序列号13、序列号14或序列号15所示的氨基酸序列的改造丝心蛋白的具体的实施方式如上所述。
[0189]
第5改造丝心蛋白的结构域序列与天然来源的丝心蛋白相比,其可以具有相当于rep中的一个或多个氨基酸残基被置换为疏水性指标大的氨基酸残基及/或在rep中插入了一个或多个疏水性指标大的氨基酸残基的、且局部包含疏水性指标大的区域的氨基酸序列。
[0190]
局部疏水性指标大的区域优选由连续的2~4个氨基酸残基构成。
[0191]
上述疏水性指标大的氨基酸残基更优选为选自异亮氨酸(i)、缬氨酸(v)、亮氨酸(l)、苯丙氨酸(f)、半胱氨酸(c)、蛋氨酸(m)及丙氨酸(a)中的氨基酸残基。
[0192]
第5改造丝心蛋白与天然来源的丝心蛋白相比,其除了相当于将rep中的一个或多个氨基酸残基置换为疏水性指标大的氨基酸残基及/或在rep中插入一个或多个疏水性指标大的氨基酸残基的改造之外,进一步与天然来源的丝心蛋白相比,还可以进行相当于与置换、缺失、插入及/或添加一个或多个氨基酸残基的氨基酸序列的改造。
[0193]
例如可以通过从克隆出的天然来源的丝心蛋白的基因序列中将rep中的一个或多个亲水性氨基酸残基(例如疏水性指标为负的氨基酸残基)置换为疏水性氨基酸残基(例如疏水性指标为正的氨基酸残基)、及/或在rep中插入一个或多个疏水性氨基酸残基,从而得到第5改造丝心蛋白。此外,例如也可以通过设计出相当于从天然来源的丝心蛋白的氨基酸序列中,将rep中的一个或多个亲水性氨基酸残基置换为疏水性氨基酸残基、及/或在rep中插入一个或多个疏水性氨基酸残基的氨基酸序列,并且对编码所设计的氨基酸序列的核酸进行化学合成后得到。在任何情况下,除了相当于从天然来源的丝心蛋白的氨基酸序列中将rep中的一个或多个亲水性氨基酸残基置换为疏水性氨基酸残基、及/或在rep中插入一个或多个疏水性氨基酸残基的改造之外,还可以进一步进行与置换、缺失、插入及/或添加一个或多个氨基酸残基相当的氨基酸序列的改造。
[0194]
第5改造丝心蛋白包含式1:[(a)n基序-rep]m所示的结构域序列,并且可以具有下述氨基酸序列:从上述结构域序列中除去自位于最靠c末端侧的(a)n基序至上述结构域序列的c末端为止的序列后得到的序列所含的所有rep中,将连续的4个氨基酸残基的疏水性指标的平均值为2.6以上的区域所含的氨基酸残基的总数设为p、将从上述结构域序列中除去自位于最靠c末端侧的(a)n基序至上述结构域序列的c末端为止的序列后得到的序列所含的氨基酸残基的总数设为q时,p/q为6.2%以上。
[0195]
}关于氨基酸残基的疏水性指标,使用公知的指标(hydropathy index:kyte j,&
doolittle r(1982)“a simple method for displaying the hydropathic character of a protein”,j.mol.biol.,157,pp.105-132)。具体而言,各氨基酸的疏水性指标(亲水指数,以下也记为“hi”)如下述表1所示。
[0196]
[表1]
[0197]
氨基酸hi氨基酸hi异亮氨酸(ile)4.5色氨酸(trp)-0.9缬氨酸(val)4.2酪氨酸(tyr)-1.3亮氨酸(leu)3.8脯氨酸(pro)-1.6苯丙氨酸(phe)2.8组氨酸(his)-3.2半胱氨酸(cys)2.5天冬酰胺(asn)-3.5蛋氨酸(met)1.9天冬氨酸(asp)-3.5丙氨酸(ala)1.8谷氨酰胺(gln)-3.5甘氨酸(gly)-0.4谷氨酸(glu)-3.5苏氨酸(thr)-0.7赖氨酸(lys)-3.9丝氨酸(ser)-0.8精氨酸(arg)-4.5
[0198]
对p/q的计算方法进一步进行详细说明。在计算中,使用从式1:[(a)n基序-rep]m所示的结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列(以下称为“序列a”)。首先,在序列a所包含的所有rep中,计算连续的4个氨基酸残基的疏水性指标的平均值。疏水性指标的平均值是通过将连续的4个氨基酸残基中所含有的各氨基酸残基的hi的总和除以4(氨基酸残基数)而求出的。疏水性指标的平均值是针对所有连续的4个氨基酸残基而求出的(各氨基酸残基用于1~4次平均值的计算)。接下来,确定连续的4个氨基酸残基的疏水性指标的平均值为2.6以上的区域。即使某个氨基酸残基属于多个“疏水性指标的平均值为2.6以上的连续的4个氨基酸残基”的情况下,也会作为一个氨基酸残基包含在区域中。而且,该区域中所含的氨基酸残基的总数为p。此外,序列a中所含的氨基酸残基的总数为q。
[0199]
例如,当提取到20处“疏水性指标的平均值为2.6以上的连续的4个氨基酸残基”时(没有重复),在连续的4个氨基酸残基的疏水性指标的平均值为2.6以上的区域中含有20处连续的4个氨基酸残基(没有重复),p为20
×
4=80。此外,例如在2处“疏水性指标的平均值为2.6以上的连续的4个氨基酸残基”仅有一个氨基酸残基重复存在的情况下,在连续的4个氨基酸残基的疏水性指标的平均值为2.6以上的区域中,含有7个氨基酸残基(p=2
×
4-1=7。
“‑
1”为重复部分的扣除)。例如,在图10所示的结构域序列的情况下,“疏水性指标的平均值为2.6以上的连续的4个氨基酸残基”不重复且存在7个,因此p为7
×
4=28。此外,例如当为图10所示的结构域序列时,q为4 50 4 40 4 10 4 20 4 30=170(不包含位于c末端侧最后的(a)n基序)。然后,通过将p除以q,可以计算出p/q(%)。在图10的情况下,28/170=16.47%。
[0200]
在第5改造丝心蛋白中,p/q优选为6.2%以上,更优选为7%以上,进一步优选为10%以上,更进一步优选为20%以上,进一步还优选为30%以上。p/q的上限并没有特别限制,例如可以为45%以下。
[0201]
例如,对于克隆出的天然来源的丝心蛋白的氨基酸序列,通过将rep中的一个或多
个亲水性氨基酸残基(例如疏水性指标为负的氨基酸残基)置换为疏水性氨基酸残基(例如疏水性指标为正的氨基酸残基)、及/或在rep中插入一个或多个疏水性氨基酸残基,以使其满足上述p/q的条件,并且通过将其改造成局部含有疏水性指标大的区域的氨基酸序列,从而得到第5改造丝心蛋白。此外,例如,也可以通过根据天然来源的丝心蛋白的氨基酸序列设计满足上述p/q的条件的氨基酸序列,并通过对编码所设计的氨基酸序列的核酸进行化学合成后得到。在任何情况下,与天然来源的丝心蛋白相比,除了进行相当于rep中的一个或多个氨基酸残基被置换为疏水性指标大的氨基酸残基、及/或在rep中插入一个或多个疏水性指标大的氨基酸残基的改造之外,还可以进一步进行相当于置换、缺失、插入及/或添加一个或多个氨基酸残基的改造。
[0202]
作为疏水性指标大的氨基酸残基,并没有特别限制,优选为异亮氨酸(i)、缬氨酸(v)、亮氨酸(l)、苯丙氨酸(f)、半胱氨酸(c)、蛋氨酸(m)及丙氨酸(a),更优选为缬氨酸(v)、亮氨酸(l)及异亮氨酸(i)。
[0203]
作为第5改造丝心蛋白的更具体的例子,可以列举出包括与(5-i)序列号19(met-prt720)、序列号20(met-prt665)或序列号21(met-prt666)所示的氨基酸序列、或者(5-ii)序列号19、序列号20或序列号21所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列的改造丝心蛋白。
[0204]
对(5-i)的改造丝心蛋白进行说明。序列号19所示的氨基酸序列是相对于序列号7(met-prt410)所示的氨基酸序列,除c末端侧的终端的结构域序列之外,每隔一个rep插入2处分别由3个氨基酸残基构成的氨基酸序列(vli),进而将一部分的谷氨酰胺(q)残基置换为丝氨酸(s)残基,并且使c末端侧的一部分氨基酸缺失的氨基酸序列。序列号20所示的氨基酸序列相对于序列号8(met-prt525)所示的氨基酸序列,每隔一个rep插入1处分别由3个氨基酸残基构成的氨基酸序列(vli)。序列号21所示的氨基酸序列相对于序列号8所示的氨基酸序列,每隔一个rep插入2处分别由3个氨基酸残基构成的氨基酸序列(vli)。
[0205]
(5-i)的改造丝心蛋白可以由序列号19、序列号20或序列号21所示的氨基酸序列构成。
[0206]
(5-ii)的改造丝心蛋白包括与序列号19、序列号20或序列号21所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列。(5-ii)的改造丝心蛋白也为包含式1:[(a)n基序-rep]m所示的结构域序列的蛋白质。上述序列一致性优选为95%以上。
[0207]
(5-ii)的改造丝心蛋白与序列号19、序列号20或序列号21所示的氨基酸序列具有90%以上的序列一致性,并且优选地,从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列所含的所有rep中,将连续的4个氨基酸残基的疏水性指标的平均值为2.6以上的区域所含的氨基酸残基的总数设为p、将从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列所含的氨基酸残基的总数设为q时,p/q为6.2%以上。
[0208]
第5改造丝心蛋白可以在n末端及c末端中的任意一端或两端包含标签序列。
[0209]
作为含有标签序列的改造丝心蛋白的更具体的例子,可以列举出包括与(5-iii)序列号22(prt720)、序列号23(prt665)或序列号24(prt666)所示的氨基酸序列、或者(5-iv)序列号22、序列号23或序列号24所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列的改造丝心蛋白。
[0210]
序列号22、序列号23及序列号24所示的氨基酸序列分别在序列号19、序列号20及序列号21所示的氨基酸序列的n末端添加序列号11所示的氨基酸序列(包含his标签序列及铰链序列)。
[0211]
(5-iii)的改造丝心蛋白可以由序列号22、序列号23或序列号24所示的氨基酸序列构成。
[0212]
(5-iv)的改造丝心蛋白包括与序列号22、序列号23或序列号24所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列。(5-iv)的改造丝心蛋白也为包含式1:[(a)n基序-rep]m所示的结构域序列的蛋白质。上述序列一致性优选为95%以上。
[0213]
(5-iv)的改造丝心蛋白与序列号22、序列号23或序列号24所示的氨基酸序列具有90%以上的序列一致性,并且优选地,从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列所含的所有rep中,将连续的4个氨基酸残基的疏水性指标的平均值为2.6以上的区域所含的氨基酸残基的总数设为p、将从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列所含的氨基酸残基的总数设为q时,p/q为6.2%以上。
[0214]
第5改造丝心蛋白可以含有用于将重组蛋白生产系统所生产的蛋白质释放到宿主外部的分泌信号。分泌信号的序列可以根据宿主的种类来进行适当设定。
[0215]
第6改造丝心蛋白与天然来源的丝心蛋白相比,具有降低谷氨酰胺残基含量的氨基酸序列。
[0216]
第6改造丝心蛋白优选在rep的氨基酸序列中包含选自ggx基序及gpgxx基序中的至少1种基序。
[0217]
第6改造丝心蛋白在rep中包括gpgxx基序时,gpgxx基序含有率通常为1%以上,也可以为5%以上,优选为10%以上。gpgxx基序含有率的上限并没有特别限制,可以为50%以下,也可以为30%以下。
[0218]
在本说明书中,“gpgxx基序含有率”是通过以下方法计算出的值。在包含式1:[(a)n基序-rep]m或式2:[(a)n基序-rep]m-(a)n基序所示的结构域序列的丝心蛋白(改造丝心蛋白或天然来源的丝心蛋白)中,在从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列所含的所有rep中,当将该区域所含的gpgxx基序的个数的总数乘以3而得到的数(即,相当于gpgxx基序中的g及p的总数)设为s,并且将从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列且除去(a)n基序后得到的所有rep的氨基酸残基的总数设为t时,以s/t来计算gpgxx基序含有率。
[0219]
在gpgxx基序含有率的计算中,将“从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列”作为对象是为了排除下述影响:“位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列”(相当于rep的序列)中有时包含与丝心蛋白的特征性序列相关性低的序列,在m较小的情况下(也就是说,结构域序列短的情况下)会影响gpgxx基序含有率的计算结果。另外,在“gpgxx基序”位于rep的c末端的情况下,即使“xx”例如为“aa”时,也作为“gpgxx基序”来进行处理。
[0220]
图11是表示改造丝心蛋白的结构域序列的示意图。参照图11来具体说明gpgxx基序含有率的计算方法。首先,在图11所示的改造丝心蛋白的结构域序列(“[(a)n基序-rep]
m-(a)n基序”类型)中,所有rep都包含在“从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列”(图11中“区域a”所示的序列)中,因此用于计算s的gpgxx基序的个数为7,s为7
×
3=21。同样地,由于所有rep都包含在“从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列”(图11中“区域a”所示的序列)中,因此从该序列中进一步除去(a)n基序后得到的所有rep的氨基酸残基的总数t为50 40 10 20 30=150。接下来,通过将s除以t,可以计算出s/t(%),在图11的改造丝心蛋白的情况下,为21/150=14.0%。
[0221]
在第6改造丝心蛋白中,谷氨酰胺残基含有率优选为9%以下,更优选为7%以下,进一步优选为4%以下,特别优选为0%。
[0222]
在本说明书中,“谷氨酰胺残基含有率”是通过以下方法计算出的值。在包含式1:[(a)n基序-rep]m或式2:[(a)n基序-rep]m-(a)n基序所示的结构域序列的丝心蛋白(改造丝心蛋白或天然来源的丝心蛋白)中,在从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列(相当于图11的“区域a”的序列)所含的所有rep中,当将该区域所含的谷氨酰胺残基的总数设为u,并且将从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列且进一步除去(a)n基序后得到的所有rep的氨基酸残基的总数设为t时,以u/t来计算谷氨酰胺残基含有率。在谷氨酰胺残基含有率的计算中,以“从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列”为对象的理由与上述的理由相同。
[0223]
第6改造丝心蛋白的结构域序列与天然来源的丝心蛋白相比,可以具有相当于使rep中的一个或多个谷氨酰胺残基发生缺失或置换为其他氨基酸残基的氨基酸序列。
[0224]“其他氨基酸残基”只要是谷氨酰胺残基以外的氨基酸残基即可,优选为疏水性指标大于谷氨酰胺残基的氨基酸残基。氨基酸残基的疏水性指标如表1所示。
[0225]
如表1所示,作为疏水性指标大于谷氨酰胺残基的氨基酸残基,可以列举出选自异亮氨酸(i)、缬氨酸(v)、亮氨酸(l)、苯丙氨酸(f)、半胱氨酸(c)、蛋氨酸(m)、丙氨酸(a)、甘氨酸(g)、苏氨酸(t)、丝氨酸(s)、色氨酸(w)、酪氨酸(y)、脯氨酸(p)及组氨酸(h)中的氨基酸残基。其中,更优选为选自异亮氨酸(i)、缬氨酸(v)、亮氨酸(l)、苯丙氨酸(f)、半胱氨酸(c)、蛋氨酸(m)及丙氨酸(a)中的氨基酸残基,进一步优选为选自异亮氨酸(i)、缬氨酸(v)、亮氨酸(l)及苯丙氨酸(f)中的氨基酸残基
[0226]
第6改造丝心蛋白中的rep的疏水性程度优选为-0.8以上,更优选为-0.7以上,进一步优选为0以上,更进一步优选为0.3以上,特别优选为0.4以上。rep的疏水性程度的上限并没有特别限制,可以为1.0以下,也可以为0.7以下。
[0227]
在本说明书中,“rep的疏水性程度”为通过以下方法而计算出的值。在包含式1:[(a)n基序-rep]m或式2:[(a)n基序-rep]m-(a)n基序所示的结构域序列的丝心蛋白(改造丝心蛋白或天然来源的丝心蛋白)中,在从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列后得到的序列(相当于图11的“区域a”的序列)所含的所有rep中,当将该区域的各氨基酸残基的疏水性指标的总和设为v,并且将从结构域序列中除去自位于最靠c末端侧的(a)n基序至结构域序列的c末端为止的序列且进一步除去(a)n基序后得到的所有rep的氨基酸残基的总数设为t时,以v/t来计算rep的疏水性程度。在rep的疏水性程度的计算中,以“从结构域序列中除去自位于最靠c末端侧的(a)n基序至结
构域序列的c末端为止的序列后得到的序列”为对象的理由与上述的理由相同。
[0228]
第6改造丝心蛋白的结构域序列与天然来源的丝心蛋白相比,其除了相当于缺失了rep中的一个或多个谷氨酰胺残基、及/或将rep中的一个或多个谷氨酰胺残基置换为其他氨基酸残基的改造之外,还可以进一步进行相当于置换、缺失、插入及/或添加一个或多个氨基酸残基的氨基酸序列的改造。
[0229]
例如可以通过在克隆出的天然来源的丝心蛋白的基因序列中使rep中的一个或多个谷氨酰胺残基发生缺失、及/或将rep中的一个或多个谷氨酰胺残基置换为其他氨基酸残基,从而得到第6改造丝心蛋白。此外,例如也可以通过设计出相当于在天然来源的丝心蛋白的氨基酸序列中,使rep中的一个或多个谷氨酰胺残基发生缺失、及/或将rep中的一个或多个谷氨酰胺残基置换为其他氨基酸残基的氨基酸序列,并且对编码所设计的氨基酸序列的核酸进行化学合成后得到。
[0230]
作为第6改造丝心蛋白的更具体的例子,可以列举出包括(6-i)序列号25(met-prt888)、序列号26(met-prt965)、序列号27(met-prt889)、序列号28(met-prt916)、序列号29(met-prt918)、序列号30(met-prt699)、序列号31(met-prt698)、序列号32(met-prt966)、序列号41(met-prt917)或序列号42(met-prt1028)所示的氨基酸序列的改造丝心蛋白,或者包括与(6-ii)序列号25、序列号26、序列号27、序列号28、序列号29、序列号30、序列号31、序列号32、序列号41或序列号42所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列的改造丝心蛋白。
[0231]
对(6-i)的改造丝心蛋白进行说明。序列号25所示的氨基酸序列是将序列号7所示的氨基酸序列(met-prt410)中的qq全部置换为vl而得到的氨基酸序列。序列号26所表示的氨基酸序列是将序列号7所表示的氨基酸序列中的qq全部置换为ts、并且将剩余的q置换为a而得到的氨基酸序列。序列号27所表示的氨基酸序列是将序列号7所表示的氨基酸序列中的qq全部置换为vl、并且将剩余的q置换为i而得到的氨基酸序列。序列号28所表示的氨基酸序列是将序列号7所表示的氨基酸序列中的qq全部置换为vi、并且将剩余的q置换为l而得到的氨基酸序列。序列号29所表示的氨基酸序列是将序列号7所表示的氨基酸序列中的qq全部置换为vf、并且将剩余的q置换为i而得到的氨基酸序列。
[0232]
序列号30所示的氨基酸序列是将序列号8所示的氨基酸序列(met-prt525)中的qq全部置换为vl而得到的氨基酸序列。序列号31所表示的氨基酸序列是将序列号8所表示的氨基酸序列中的qq全部置换为vl、并且将剩余的q置换为i而得到的氨基酸序列。
[0233]
序列号32所示的氨基酸序列是将序列号7所示的氨基酸序列(met-prt410)中存在的20个结构域序列的区域重复2次后得到的序列中的qq全部置换为vf,并且将剩余的q置换为i而得到的氨基酸序列。
[0234]
序列号41所表示的氨基酸序列(met-prt917)是将序列号7所表示的氨基酸序列中的qq全部置换为li、并且将剩余的q置换为v而得到的氨基酸序列。序列号42所表示的氨基酸序列(met-prt1028)是将序列号7所表示的氨基酸序列中的qq全部置换为if、并且将剩余的q置换为t而得到的氨基酸序列。
[0235]
序列号25、序列号26、序列号27、序列号28、序列号29、序列号30、序列号31、序列号32、序列号41及序列号42所示的氨基酸序列的谷氨酰胺残基含有率均为9%以下(表2)。
[0236]
[表2]
[0237][0238]
(6-i)的改造丝心蛋白可以由序列号25、序列号26、序列号27、序列号28、序列号29、序列号30、序列号31、序列号32、序列号41或序列号42所示的氨基酸序列构成。
[0239]
(6-ii)的改造丝心蛋白包括与序列号25、序列号26、序列号27、序列号28、序列号29、序列号30、序列号31、序列号32、序列号41或序列号42所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列。(6-ii)的改造丝心蛋白也是包括式1:[(a)n基序-rep]m、或式2:[(a)n基序-rep]m-(a)n基序所示的结构域序列的蛋白质。上述序列一致性优选为95%以上。
[0240]
(6-ii)的改造丝心蛋白的谷氨酰胺残基含有率优选为9%以下。此外,(6-ii)的改造丝心蛋白的gpgxx基序含有率优选为10%以上。
[0241]
第6改造丝心蛋白可以在n末端及c末端中的任意一端或两端包含标签序列。由此,可以实现改造丝心蛋白的分离、固定、检测及可视化等。
[0242]
作为含有标签序列的改造丝心蛋白的更具体的例子,可以列举出包括(6-iii)序列号33(prt888)、序列号34(prt965)、序列号35(prt889)、序列号36(prt916)、序列号37(prt918)、序列号38(prt699)、序列号39(prt698)、序列号40(prt966)、序列号43(prt917)或序列号44(prt1028)所示的氨基酸序列的改造丝心蛋白,或者包括与(6-iv)序列号33、序列号34、序列号35、序列号36、序列号37、序列号38、序列号39、序列号40、序列号43或序列号44所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列的改造丝心蛋白。
[0243]
序列号33、序列号34、序列号35、序列号36、序列号37、序列号38、序列号39、序列号40、序列号43及序列号44所示的氨基酸序列分别在序列号25、序列号26、序列号27、序列号28、序列号29、序列号30、序列号31、序列号32、序列号41及序列号42所示的氨基酸序列的n末端添加序列号11所示的氨基酸序列(包含his标签序列及铰链序列)。由于仅在n末端添加了标签序列,因此谷氨酰胺残基含有率没有变化,序列号33、序列号34、序列号35、序列号36、序列号37、序列号38、序列号39、序列号40、序列号43及序列号44所示的氨基酸序列的谷氨酰胺残基含有率均为9%以下(表3)。
[0244]
[表3]
[0245][0246]
(6-iii)的改造丝心蛋白可以由序列号33、序列号34、序列号35、序列号36、序列号37、序列号38、序列号39、序列号40、序列号43或序列号44所示的氨基酸序列构成。
[0247]
(6-iv)的改造丝心蛋白包括与序列号33、序列号34、序列号35、序列号36、序列号37、序列号38、序列号39、序列号40、序列号43或序列号44所示的氨基酸序列具有90%以上的序列一致性的氨基酸序列。(6-iv)的改造丝心蛋白也是包括式1:[(a)n基序-rep]m、或式2:[(a)n基序-rep]m-(a)n基序所示的结构域序列的蛋白质。上述序列一致性优选为95%以上。
[0248]
(6-iv)的改造丝心蛋白的谷氨酰胺残基含有率优选为9%以下。此外,(6-iv)的改造丝心蛋白的gpgxx基序含有率优选为10%以上。
[0249]
第6改造丝心蛋白可以含有用于将重组蛋白生产系统所生产的蛋白质释放到宿主外部的分泌信号。分泌信号的序列可以根据宿主的种类来进行适当设定。
[0250]
改造丝心蛋白可以为兼具第1改造丝心蛋白、第2改造丝心蛋白、第3改造丝心蛋白、第4改造丝心蛋白、第5改造丝心蛋白及第6改造丝心蛋白所具有的特征中的至少2种以上特征的改造丝心蛋白。
[0251]
本实施方式所涉及的改造丝心蛋白可以使用对该改造丝心蛋白进行编码的核酸,通过常规方法进行制备。编码该改造丝心蛋白的核酸可以根据碱基序列信息进行化学合成,也可以利用pcr法等进行合成。此外,制造的改造丝心蛋白的分离和纯化也可以通过常用方法来进行。
[0252]
[蛋白质纤维面料的制备方法]
[0253]
本实施方式所涉及的蛋白质纤维面料的制备方法包括使用通过本发明所涉及的蛋白质纤维的制备方法得到的蛋白质纤维来制作面料的工序。作为使用蛋白质纤维来制作面料的方法,可以利用公知的方法,并没有特别限制。
[0254]
根据本实施方式所涉及的蛋白质纤维面料的制备方法,通过使用如前所述的本发明所涉及的制备方法而得到的蛋白质纤维,可以很容易地制备蛋白质纤维面料,该面料能够更可靠地抑制制备后首次与水分接触时所产生的收缩,并且还可以抑制起皱的发生。
[0255]
用于制备蛋白质纤维面料的蛋白质纤维可以是短纤维,也可以是长纤维。此外,该蛋白质纤维可以单独使用或与其他纤维组合使用。即,在制备蛋白质纤维面料时,作为材料
丝线,可以分别单独使用或组合使用仅仅由通过本发明所涉及的制备方法而得到的蛋白质纤维构成的单纱、将通过本发明所涉及的制备方法而得到的蛋白质纤维与其他纤维组合而成的复合纱。另外,其他纤维是指通过本发明所涉及的制备方法以外的制备方法而得到的蛋白质纤维,不含蛋白质的纤维等。此外,复合纱例如包括混纺纱、针织纱、包芯纱等。
[0256]
按照本实施方式所涉及的蛋白质纤维面料的制备方法制备的蛋白质纤维面料的种类也并没有特别限定。蛋白质纤维面料例如可以是织物或针织物,也可以是无纺布。此外,对于织物,例如,织物组织也可以是平纹组织、斜纹组织、缎纹组织等,所使用的纱线的种类可以是1种,也可以是多种。针织物也可以是例如特里科经编针织物、拉舍尔经编针织物等经编针织物,也可以是横编、圆编等纬编针织物,所使用的纱线的种类可以是1种,也可以是多种。
[0257]
[蛋白质纤维的防缩加工方法]
[0258]
也可以将以上说明的本发明的蛋白质纤维的制备方法理解为对通过包括拉伸工序和防缩工序在内的公知的制备方法而制备的蛋白质纤维进一步进行防缩加工的防缩加工方法。即,本实施方式所涉及的蛋白质纤维的防缩加工方法具备:前收缩工序,其用于使拉伸后且卷绕前的蛋白质纤维在松弛状态下进行收缩;卷绕工序,其用于将经过前收缩工序的蛋白质纤维卷绕以得到卷绕物;以及后收缩工序,其用于使经过卷绕工序的蛋白质纤维在松弛状态下进行收缩。作为该防缩加工方法的具体实施方式,其与在蛋白质纤维的制备方法中说明的具体实施方式相同。
[0259]
实施例
[0260]
以下,基于实施例更具体地对本发明进行说明。但是,本发明并不限于以下实施例。
[0261]
[试验例1:改造丝心蛋白的制造]
[0262]
(1)表达载体的制备
[0263]
设计具有序列号15所示的氨基酸序列的改造丝心蛋白(prt799)。合成编码所设计的改造丝心蛋白的核酸。该核酸中,在5'末端附加有ndei位点、在终止密码子下游附加有ecori位点。将该核酸克隆至克隆载体(puc118)中。然后,利用ndei和ecori对该核酸进行限制酶处理并将其切出后,重组到蛋白质表达载体pet-22b( )中,得到表达载体。
[0264]
(2)蛋白质的表达
[0265]
用得到的表达载体转化大肠杆菌blr(de3)。将该转化的大肠杆菌在含有氨苄西林的2ml lb培养基中培养15小时。将该培养液添加到含有氨苄西林的100ml的种子培养用培养基(表4)中,以使od600达到0.005。将培养液温度保持于30℃,进行烧瓶培养直至od600达到5为止(约15小时),得到种子培养液。
[0266]
[表4]
[0267]
种子培养用培养基
[0268][0269]
将该种子培养液添加到加有500ml的生产培养基(表5)的发酵罐中,以使od600达到0.05。将培养液温度保持在37℃,将ph恒定控制在6.9下进行培养。此外,将培养液中的溶解氧浓度维持于溶解氧饱和浓度的20%。
[0270]
[表5]
[0271]
生产培养基
[0272][0273]
在生产培养基中的葡萄糖完全消耗后,立即以1ml/分钟的速度添加进料液(葡萄糖455g/1l、酵母提取物120g/1l)。将培养液温度保持在37℃,将ph恒定控制在6.9下进行培养。此外,将培养液中的溶解氧浓度维持在溶解氧饱和浓度的20%,培养20小时。然后,向培养液中添加1m的异丙基-β-硫代半乳糖吡喃糖苷(iptg)以使其最终浓度为1mm,并表达诱导改造丝心蛋白。在添加iptg后经过20小时的时刻,离心分离培养液,回收菌体。使用由添加iptg前和添加iptg后的培养液制备的菌体进行sds-page,根据依赖于iptg添加的目标改造丝心蛋白尺寸的条带的出现,确认到目标改造丝心蛋白的表达。
[0274]
(3)蛋白质的纯化
[0275]
将添加iptg后2小时后回收的菌体用20mm tris-hcl缓冲液(ph7.4)进行清洗。将洗涤后的菌体悬浮在含有约1mm的pmsf的20mm tris-hcl缓冲液(ph7.4)中,用高压均质器(gea niro soavi公司制)来破碎细胞。对破碎后的细胞进行离心分离,得到沉淀物。利用20mm tris-hcl缓冲液(ph7.4)清洗所得到的沉淀物,直至达到高纯度为止。将清洗后的沉淀物以达到100mg/ml的浓度的方式悬浮在8m胍缓冲液(8m胍盐酸盐、10mm磷酸二氢钠、20mm nacl、1mm tris-hcl、ph7.0)中,在60℃下用搅拌器搅拌30分钟,使其溶解。溶解后,使用透析管(三光纯药株式会社制的纤维素管36/32)用水进行透析。通过离心分离来回收透析后得到的白色凝聚蛋白质,用冷冻干燥机去除水分,通过回收冻干粉粉末,而获得改造丝心蛋白(prt799)。
[0276]
(试验例2:蛋白质纤维的制备及评价)
[0277]
(1)蛋白质原纤维的制备
[0278]
准备溶解有licl的二甲基亚砜(dmso)作为溶剂以使其达到4.0质量%,向其中添加改造丝心蛋白(prt799)的冻干粉粉末以使其浓度达到24质量%,并使用振荡器以使其溶解3小时。然后,除去不溶物和气泡,得到改造丝心蛋白溶液(纺丝原液)。
[0279]
作为前收缩工序,在利用湿式收缩法的情况(实施例1和比较例2)下,通过基于图1所示的纺丝装置100的纺丝装置来制备蛋白质原纤维。具体而言,将制备的纺丝原液在60℃用网孔为5μm的金属过滤器过滤,然后将其放入30ml的不锈钢注射器内静置并使其脱泡后,从针头直径为0.2mm的实心喷嘴喷出至100质量%的甲醇凝固浴槽中。喷出温度为60℃。凝固后,经过洗涤,在90℃的热水中浸渍10秒钟后,实施前收缩工序。此时,蛋白质原纤维的卷绕速度相对于送出速度的倍率小于1倍(0.6倍),可称为松弛状态。经过100℃下的干燥,卷绕所得到的原丝(卷绕工序),并得到蛋白质原纤维(改造丝心蛋白原纤维)。
[0280]
作为前收缩工序,在利用干式收缩法的情况下(实施例2和比较例3),通过基于图2所示的纺丝装置200的纺丝装置来制备蛋白质原纤维。具体而言,将制备的纺丝原液在60℃用网孔为5μm的金属过滤器过滤,然后将其放入30ml的不锈钢注射器内静置并使其脱泡后,从针头直径为0.2mm的实心喷嘴喷出至100质量%的甲醇凝固浴槽中。喷出温度为60℃。凝固后,经过洗涤和干燥,在240℃下加热2秒钟后,实施前收缩工序。此时,蛋白质原纤维的卷绕速度相对于送出速度的倍率小于1倍(0.5倍),可称为松弛状态。卷绕所得到的原丝(卷绕工序),并得到蛋白质原纤维(改造丝心蛋白原纤维)。
[0281]
在未实施前收缩工序的情况(比较例1)下,通过现有的干湿式纺丝装置(从图1所示的纺丝装置100中除去水浴46后的装置)来制备蛋白质原纤维。具体而言,将制备的纺丝原液在60℃用网孔为5μm的金属过滤器过滤,然后将其放入30ml的不锈钢注射器内静置并使其脱泡后,从针头直径为0.2mm的实心喷嘴喷出至100质量%的甲醇凝固浴槽中。喷出温度为60℃。凝固后,卷绕所得到的原丝并使其自然干燥,以得到蛋白质原纤维(改造丝心蛋白原纤维)。
[0282]
作为后收缩工序,在利用湿式收缩法的情况(实施例2和比较例1)下,使用基于图6(a)和图6(b)所示装置的装置来实施后收缩工序。具体而言,从蛋白质原纤维的卷绕物中拉出蛋白质原纤维,得到绞纱状的蛋白质原纤维(参照图6(a))。接下来,在90℃的热水中浸渍15分钟后,实施后收缩工序。之后,使其自然干燥,并得到蛋白质纤维(改造丝心蛋白纤维)。
[0283]
作为后收缩工序,在利用干式收缩法的情况(实施例1)下,使用基于图6(a)和图6(c)所示装置的装置来实施后收缩工序。具体而言,从蛋白质原纤维的卷绕物中拉出蛋白质原纤维,得到绞纱状的蛋白质原纤维(参照图6(a))。接下来,使绞纱状的蛋白质原纤维与加热至240℃的高温加热器相接触,并保持10秒钟后,实施后收缩工序。由此,得到蛋白质纤维(改造丝心蛋白纤维)。
[0284]
(2)蛋白质纤维的收缩率评价
[0285]
分别将裁切成长度为300mm的各实施例和比较例中的改造丝心蛋白纤维(试验片)在无负荷状态下浸渍在40℃或90℃的水中10分钟。将从水中取出的各个试验片在室温下干燥2小时。然后,测量试验片的长度(干燥后的纤维长度),并测量收缩率。将结果示于表6。收缩率是通过下式(1)计算出的数值。另外,“浸渍前的纤维长度”为300mm。
[0286]
收缩率=(1-干燥后的纤维长度/浸渍前的纤维长度)
×
100
···
(1)
[0287]
(3)蛋白质纤维的起皱评价
[0288]
通过目视确认(2)中测定收缩率后的试验片(干燥后的试验片)有无起皱。将结果示于表6。
[0289]
[表6]
[0290][0291]
实施前收缩工序和后收缩工序这两者后的蛋白质纤维(实施例1和实施例2)充分抑制了在其制备后首次与水分接触时所产生的收缩,而且也没有发生起皱。仅仅实施后收缩工序后的蛋白质纤维(比较例1)虽然抑制了在其制备后首次与水分接触时所产生的收缩,但是发生了起皱。仅仅实施前收缩工序后的蛋白质纤维(比较例2和比较例3)虽然未发生起皱,但制备后首次与水分接触时所发生的收缩(特别是与高温的水分接触时发生的收缩)较大。
[0292]
符号说明
[0293]1ꢀꢀꢀꢀꢀꢀꢀꢀ
挤出装置
[0294]4ꢀꢀꢀꢀꢀꢀꢀꢀ
干燥装置
[0295]6ꢀꢀꢀꢀꢀꢀꢀꢀ
纺液
[0296]
20
ꢀꢀꢀꢀꢀꢀꢀ
凝固浴槽
[0297]
21
ꢀꢀꢀꢀꢀꢀꢀ
洗涤浴槽
[0298]
36
ꢀꢀꢀꢀꢀꢀꢀ
蛋白质原纤维
[0299]
38
ꢀꢀꢀꢀꢀꢀꢀ
蛋白质纤维
[0300]
42
ꢀꢀꢀꢀꢀꢀꢀ
进料辊
[0301]
44
ꢀꢀꢀꢀꢀꢀꢀ
卷线机
[0302]
46
ꢀꢀꢀꢀꢀꢀꢀ
水浴
[0303]
48
ꢀꢀꢀꢀꢀꢀꢀ
干燥机
[0304]
54
ꢀꢀꢀꢀꢀꢀꢀ
加热器
[0305]
56
ꢀꢀꢀꢀꢀꢀꢀ
压辊
[0306]
58
ꢀꢀꢀꢀꢀꢀꢀ
热辊
[0307]
60
ꢀꢀꢀꢀꢀꢀꢀ
高温加热松弛装置
[0308]
100、200
ꢀꢀ
纺丝装置
[0309]
300、400
ꢀꢀ
后收缩装置。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。