用于治疗史密斯-马吉利综合征的方法和组合物
1.相关申请的交叉引用本技术根据35 u.s.c.
§
119(e)要求2019年6月26日提交的美国序列号62/867068的优先权的权益,其全部内容通过引用并入本文。
2.授权信息本发明是在国立卫生研究院授予的mh099082号政府支持下完成的。政府对本发明拥有一定的权利。
3.序列表本技术包括序列表,该序列表已经以ascii格式电子提交,并通过引用整体并入本技术。该ascii副本创建于2020年5月14日,命名为20378-202575_sl.txt,大小为61 kb。
4.发明背景。
发明领域
5.本发明大体上涉及行为障碍,更具体地,涉及用nmda受体拮抗剂治疗此类障碍的方法。
6.背景资料谷氨酸能突触是哺乳动物中枢神经系统中兴奋性突触的主要类别,并且大多数投射神经元是谷氨酸能神经元(collingridge et al., 1983; monaghan et al., 1989; watkins and evans, 1981)。谷氨酸能突触的正常发育和可塑性对行为功能是必需的,其破坏会导致各种疾病。最近的一项研究表明,平面细胞极性(pcp)信号传导通路的组分是谷氨酸能突触形成的关键调节因子(thakar et al., 2017)。celsr3对谷氨酸能突触形成是必需的,而vangl2负调控谷氨酸能突触形成。因此,pcp信号传导组分可以正调控和负调控谷氨酸能突触数量。
7.幼虫巨大致死性基因(lgl1)是高度保守的顶端-基底极性信号传导通路的关键组分,其沿顶端和基底外侧轴极化上皮细胞和组织(karner et al., 2006)。染色体17 p11.2微缺失疾病中lgl1经常缺失,被称为史密斯-马吉利综合征(smith-magenis syndrome,sms)。史密斯-马吉利综合征(sms)是一种通过同源重组在胚胎发育极早期出现的新发(de novo)遗传疾病(chen et al., 1997)。大约70%的患者出现3.5 mb的缺失间隔(gropman et al., 2007)。具有所述缺失的个体经常被诊断为自闭症谱系障碍(asd)、注意缺陷/多动障碍(adhd)、强迫症(ocd)或其他行为障碍(dykens et al., 1997; dykens and smith, 1998; laje et al., 2010; martin et al., 2006; smith ac et al., 1998)。尽管常见的缺失(edelman et al., 2007; potocki et al., 2003)和多个基因可能导致该综合征(girirajan et al., 2006),但个体之间的症状有所不同。最近的研究表明rai1与sms的非asd症状有关(huang et al., 2016)。对于更好地理解lgl1介导的兴奋性毒性神经退行性变以及随后的病状和疾病的分子机制,以及基于这些机制鉴定和设计用于研究和治疗用途的有效调节剂的方法,存在未满足的需求。对于提供用于预防、控制和治疗与lgl1介导的兴奋性毒性相关的病状和疾病(例如突触损失导致的神经退行性疾病)的治疗方法和药剂,也
存在未满足的需求。本发明满足了这些需求。
发明概要
8.因此,本发明提供了一种用于减少或预防神经元群体中幼虫巨大致死性基因(lgl1)诱导的突触增加的方法,其包含使神经元与亚麻醉量的n-甲基-d-天冬氨酸(nmda)受体拮抗剂接触。在各种实施方案中,nmda受体拮抗剂增加神经元中ampa/nmda比例。在各种实施方案中,nmda受体拮抗剂选自氯胺酮(ketamine)、美金刚(memantine)、地佐环平(dizocilpine,mk-801)、右美沙芬(dextromethorphan,dxm)、苯环己哌啶(pcp)、甲氧西敏(mxe)、一氧化二氮(n2o)及其任何组合。
9.在各种实施方案中,该方法还可以包括使神经元群体与非典型蛋白激酶c(apkc)抑制剂接触。在各种实施方案中,apkc抑制剂是小分子化合物、核酸分子或多肽。在各种实施方案中,apkc抑制剂是与一种或多种蛋白激酶c ζ(apkcζ)和蛋白激酶c ι/λ(apkcι/λ)同工型(isoform)结合的抗体或抗原结合片段。在各种实施方案中,apkc抑制剂减少或抑制神经元中apkcζ和apkcι/λ的表达。
10.在各种实施方案中,该方法还可以包括使神经元群体与lgl1激动剂接触。在各种实施方案中,lgl1激动剂是诱导lgl1的表达的小分子化合物、核酸分子或多肽。在各种实施方案中,lgl1激动剂是诱导信号传导通路的小分子化合物、核酸分子或多肽,该信号传导通路诱导lgl1的表达。
11.在各种实施方案中,神经元群体在对象中,并且其中接触步骤通过向对象施用nmda受体拮抗剂和/或apkc抑制剂和/或lgl1激动剂进行。在各种实施方案中,神经元群体在对象的脑中。在各种实施方案中,对象患有或有风险发展神经退行性疾病或行为障碍,例如史密斯-马吉利综合征(sms)、自闭症或注意缺陷多动障碍(adhd)。在各种实施方案中,nmda受体拮抗剂缓解与行为障碍相关的社会交互缺陷。
12.另一方面,本发明提供了一种控制、预防或治疗对象中神经退行性疾病或行为障碍的方法,其包含向对象施用亚麻醉量的n-甲基-d-天冬氨酸(nmda)受体拮抗剂。在各种实施方案中,nmda受体拮抗剂增加神经元中ampa/nmda比例。在各种实施方案中,nmda受体拮抗剂选自氯胺酮、美金刚、地佐环平(mk-801)、右美沙芬(dxm)、苯环己哌啶(pcp)、甲氧西敏(mxe)、一氧化二氮(n2o)及其任何组合。
13.在各种实施方案中,该方法还可以包括向对象施用非典型蛋白激酶c(apkc)抑制剂。在各种实施方案中,apkc抑制剂是小分子化合物、核酸分子或多肽。在各种实施方案中,apkc抑制剂是与一种或多种蛋白激酶c ζ(apkcζ)和蛋白激酶c ι/λ(apkcι/λ)同工型结合的抗体或抗原结合片段。在各种实施方案中,apkc抑制剂减少或抑制神经元中apkcζ和apkcι/λ的表达。
14.在各种实施方案中,该方法还可以包括向对象施用lgl1激动剂。在各种实施方案中,lgl1激动剂是诱导lgl1的表达的小分子化合物、核酸分子或多肽。在各种实施方案中,lgl1激动剂是诱导信号传导通路的小分子化合物、核酸分子或多肽,该信号传导通路诱导lgl1的表达。
15.在各种实施方案中,对象中神经元群体中的突触数量减少。在各种实施方案中,突触是兴奋性突触。在各种实施方案中,突触是谷氨酸能突触。在各种实施方案中,nmda受体
拮抗剂缓解与行为障碍相关的社会交互缺陷。在各种实施方案中,神经元群体包含小脑颗粒神经元、背根神经节神经元、皮质神经元、交感神经元或海马神经元。
16.另一方面,本发明提供了一种通过lgl1的调控治疗行为障碍(如sms)的方法。在各种实施方案中,该方法包括以亚麻醉量向有此需要的对象施用n-甲基-d-天冬氨酸(nmda)受体拮抗剂(如氯胺酮),从而缓解与行为障碍相关的社会交互缺陷。通过阻断nmda受体,氯胺酮可以减轻过度连接(over connectivity)和nmdar活性增加引起的异常活性模式。
17.用于本文所提供的方法的示例性nmda受体拮抗剂包括但不限于氯胺酮、美金刚(namzaric
®
)、地佐环平(mk-801)、右美沙芬(dxm)、苯环己哌啶(pcp)、甲氧西敏(mxe)和一氧化二氮(n2o)。
18.另一方面,本发明提供了一种选择突触形成的调节剂或突触维持的调节剂的方法。在各种实施方案中,该方法包括提供具有lgl1的杂合或纯合缺失、失活或敲除的神经元群体;测量神经元中ampa/nmda比例的第一水平;使候选药剂与神经元群体接触;测量神经元中ampa/nmda比例的第二水平;以及如果联系(association)的第二水平与联系的第一水平不同,选择候选药剂作为调节剂。在各种实施方案中,候选药剂是小分子化合物、核酸或肽。在各种实施方案中,核酸是microrna、sirna或基于crispr基因编辑的构建体。在各种实施方案中,候选药剂是抗体或其抗原结合片段。在各种实施方案中,该方法在apkc的存在下进行。在各种实施方案中,细胞群体在非人类哺乳动物中,并且其中接触步骤通过向非人类哺乳动物施用候选药剂进行。
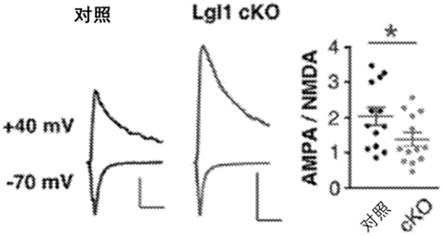
19.附图简要说明图1a-1j是示出了锥体神经元中lgl1条件性敲除导致非对称性突触数量增加的图示和示意图。图1a示出了p14小鼠的schaffer侧支区域中ca1锥体神经元层腹侧150-200 μm(顶部)或50 μm(底部)的电子显微照片。浅色箭头表示非对称性突触。黑色箭头表示对称性突触。比例尺:500 nm。对应每个区域的突触数量的量化:n = 6只对照动物、6只lgl1 cko动物。图1b示出了8周龄小鼠的schaffer侧支区域中ca1锥体神经元层腹侧150-200 μm(顶部)或50 μm(底部)的电子显微照片。浅色箭头表示非对称性突触。黑色箭头表示对称性突触。对应每个区域的突触数量的量化:n = 5只对照动物、5只lgl1 cko动物。图1c示出了来自p13-p15对照和lgl1 cko小鼠的急性切片的mepsc记录的代表性痕迹。mepsc频率和振幅的量化:n = 22个对照神经元、19个lgl1 cko神经元。图1d示出了来自p14对照和lgl1 cko小鼠的急性切片的mipsc记录的代表性痕迹。mipsc频率和振幅的量化:n = 20个对照神经元、19个lgl1 cko神经元。图1e示出了代表性示出充满alexa fluor 555酰肼的斜ca1树突的代表性共焦图像。比例尺:10 μm。图1f示出了树突棘密度的量化,以及图1g示出了树突棘形态的分布。图1h示出了p14对照和cko动物中突触超微结构测量累积分布的量化:n = 194个对照突触、174个lgl1 cko突触。8周龄对照和cko动物中突触超微结构测量累积分布的量化:n = 194个对照突触、208个lgl1 cko突触。图1i示出了野生型p14小鼠的生物化学分级分离。p2,粗制突触体;smf,突触膜级分(fraction);psd,突触后密集区。图1j示出了来自p14对照和在p7缺失lgl1后的lgl1 cko小鼠的急性切片的nmdar电流和ampar/nmdar电流组合的代表性痕迹。比例尺,100 pa(垂直);100 ms(水平)。计算的ampar电流与nmdar电流比例的量化:n = 13个对照神经元、13个lgl1 cko神经元。* p《0.05;** p《0.01;*** p《0.001;**** p《0.0001。
20.图2a-2g是示出了lgl1通过抑制apkc抑制突触形成并促进vangl2的表达的图示和示意图。图2a示出了p14小鼠的schaffer侧支区域中ca1锥体神经元层腹侧150
–
200 mm(顶部)或50 mm(底部)的电子显微照片。浅色箭头表示非对称性突触。黑色箭头表示对称性突触。比例尺,500 nm。对应每个区域的突触数量的量化:n = 8只对照动物、5只apkc dcko动物。图2b示出了8周龄小鼠的schaffer侧支区域中ca1锥体神经元层腹侧150
–
200 mm(顶部)或50 mm(底部)的电子显微照片。浅色箭头表示非对称性突触。黑色箭头表示对称性突触。对应每个区域的突触数量的量化:n = 5只对照动物、8只apkc dcko动物。图2c示出了p14对照和cko动物中突触超微结构测量累积分布的量化:n = 194个对照突触、129个apkc dcko突触。8周龄对照和cko动物中突触超微结构测量累积分布的量化:n = 87个对照突触、168个apkc dcko突触。图2d示出了p14小鼠的schaffer侧支区域中ca1锥体神经元层腹侧150-200 mm(顶部)或50 mm(底部)的电子显微照片。浅色箭头表示非对称性突触。黑色箭头表示对称性突触。比例尺,500 nm。对应每个区域的突触数量的量化:n = 7只对照动物、6只lgl1:pkcι/λ:pkcλ tcko动物。图2e示出了8周龄小鼠的schaffer侧支区域中ca1锥体神经元层腹侧150-200 mm(顶部)或50 mm(底部)的电子显微照片。浅色箭头表示非对称性突触。黑色箭头表示对称性突触。对应每个区域的突触数量的量化:n = 8只对照动物、10只lgl1:pkcι/λ:pkcξ tcko动物。图2f示出了通过蛋白质印迹的p2级分中vangl2和jam-c蛋白的水平。图2g示出了p2级分中vangl2和jam-c蛋白水平的量化。vangl2的n = 5。jam-c的n = 4。* p《0.05;** p《0.01;*** p《0.001;**** p《0.0001。
21.图3a-3n是示出了成年lgl1条件性敲除中突触数量增加、ampa/nmda比例改变和可塑性受损的图示和示意图。图3a示出了来自10周龄对照和6周龄开始缺失lgl1的lgl1 cko小鼠的切片的ca1锥体细胞层腹侧schaffer侧支(sc)150-200 mm的电子显微照片。浅色箭头表示非对称性突触。比例尺,500 nm。n=6只对照动物、8只lgl1 cko动物。图3b示出了图5a所述区域的非对称性突触密度的量化。图3c示出了10周龄动物中ca1锥体细胞层腹侧sc50 mm的电子显微照片。图3d示出了近端区域中非对称性突触的量化。图3e示出了sc的近端区域中对称性突触(黑色箭头)的显微照片。图3f示出了对称性突触的量化。图3g示出了突触后致密区(psd)长度累积频率的量化。图3h示出了psd宽度的量化。图3i示出了突触间隙距离的量化。n = 180个lgl1对照突触、242个lgl1 cko突触。图3j示出了来自6周龄对照和在p28开始缺失lgl1后的lgl1 cko小鼠的急性切片的nmdar电流和ampar/nmdar电流组合的代表性痕迹。比例尺:50 pa(垂直);80 ms(水平)。图3k示出了计算的ampar与nmdar电流比例的量化:n = 5个对照神经元、7个lgl1 cko神经元。图3l示出了tbs刺激被递送至来自对照和lgl1 cko小鼠的急性切片前后epsp的代表性痕迹。比例尺,0.2 mv,10 ms。图3m示出了θ短阵快速脉冲刺激(tbs)前后epsp斜率的量化。n = 5个lgl1对照、4个lgl1 cko。图3n示出了来自对照和在6周时缺失的lgl1 cko动物的配对脉冲比例的量化。n = 6个lgl1对照、6个lgl1 cko。* p《0.05;** p《0.01;**** p《0.0001。
22.图4a-4p示出了在p7 lgl1缺失产生sms样行为表型子集的图示和示意图。图4a示出了p7 lgl1的缺失后的旷场分析结果,示出了对照和lgl1 cko动物的代表性轨迹(左)和热图(右)。图4b示出了测试期间移动距离的量化。n = 30 对照、27 lgl1 cko。图4c示出了在第一个和第二个5分钟周期内,在场地外部区域(趋触性)花费的时间的量化。对在新物体识别(nor)测试的样品阶段期间动物对物体偏好,图4d示出了代表性热图,以及图4e示出了
量化。n = 23只对照动物、19只lgl1 cko动物。对在新物体识别(nor)测试的样品阶段期间动物对物体偏好,图4f示出了代表性热图,以及图4g示出了量化。白色星号表示新物体的位置。图4h示出了社会交互(social interaction,si)测试的习惯化阶段的代表性热图。图4i示出了花费的时间的量化,以及图4j示出了对代表新小鼠和物体的未来位置的感兴趣区域(roi)的偏好。n = 25只对照动物、24只lgl1 cko动物。图4k示出了si测试的社会交互阶段期间的代表性热图。白色星号表示新小鼠的位置。对面的腔室包含该物体。图4l示出了花费的时间的量化,以及图4m示出了对包含新小鼠或新物体的roi的偏好。正值表示对新小鼠的偏好。n = 25只对照动物、22只lgl1 cko动物。图4n示出了si测试的社会新奇阶段的代表性热图。白色星号表示新小鼠的位置。对面的腔室包含之前探索过的小鼠。图4o示出了交互花费的时间的量化,以及图4p示出了在社会新奇阶段期间对目标小鼠的偏好。正值表示对新小鼠的偏好。n = 20只对照动物、14只lgl1 cko动物。ns,非社交;s,社交。* p《0.05;** p《0.01。
23.图5a-5w是示出了lgl1、pkcι/λ和pkcξ的条件性三重敲除挽救非对称性突触数量和认知缺陷的图示和示意图。图5a示出了p7 lgl1的缺失后的旷场分析结果,示出了对照和apkc dcko动物的代表性轨迹(左)和热图(右)。图5b示出了测试期间移动距离的量化。n = 32只对照动物、26只apkc dcko动物。图5c示出了在第一个和第二个5分钟周期内,在场地外部区域(趋触性)花费的时间的量化。图5d示出了在新物体识别(nor)测试的样品阶段期间动物对物体偏好的代表性热图和量化。n=26个对照组,15只apkc dcko动物。图5e示出了在nor测试的测试阶段期间动物对物体偏好的代表性热图和量化。白色星号表示新物体的位置。图5f示出了花费的时间的量化,以及图5g示出了对代表新小鼠和物体的未来位置的感兴趣区域(roi)的偏好。n = 35只对照动物、26只apkc dcko动物。图5h示出了花费的时间的量化,以及图5i示出了对包含新小鼠或新物体的roi的偏好。正值表示对新小鼠的偏好。n = 35只对照动物、27只apkc dcko动物。* p《0.05;** p《0.01。图5j示出了交互花费的时间的量化,以及图5k示出了在社会新奇阶段期间对目标小鼠的偏好。正值表示对新小鼠的偏好。n = 35只对照动物、27只apkc dcko动物。*p《0.05。图5l示出了p7缺失后的旷场分析结果,示出了对照和lgl1:pkcι/λ:pkcξ tcko动物的代表性轨迹(左)和热图(右)。图5m示出了测试期间移动距离的量化。n = 20只对照动物、14只lgl1:pkcι/λ:pkcξ tcko动物。对在新物体识别(nor)测试的样品阶段期间动物对物体偏好,图5n示出了代表性热图,以及图5o示出了量化。n = 21只对照动物、15只lgl1:pkcι/λ:pkcξ tcko动物。对在nor测试的识别测试阶段期间动物对物体偏好,图5p示出了代表性热图,以及图5q示出了量化。白色星号表示新物体的位置。图5r示出了花费的时间的量化,以及图5s示出了对代表新小鼠和物体的未来位置的roi的偏好。图5t示出了花费的时间的量化,以及图5u示出了对包含新小鼠或新物体的roi的偏好。正值表示对新小鼠的偏好。图5v示出了交互花费的时间的量化,以及图5w示出了在社会新奇阶段期间对目标小鼠的偏好。正值表示对新小鼠的偏好。n = 25只对照动物、19只lgl1:pkcι/λ:pkcξ tcko动物。ns,非社交;s,社交。* p《0.05;** p《0.01;*** p《0.001。
24.图6a-6f是示出了lgl1
/-小鼠的突触数量和sms样行为表型增加的图示和示意图。图6a示出了8周龄的对照和lgl1
/-小鼠的schaffer侧支区域中ca1锥体神经元层腹侧150-200 μm(顶部)或50 μm(底部)的电子显微照片。浅色箭头表示非对称性突触。黑色箭头表示
对称性突触。比例尺,500 nm。对应每个区域的突触数量的量化:n=10只lgl1
/-、7只lgl1
/-动物。图6b示出了8周龄lgl1
/
和lgl1
/
动物中突触超微结构测量累积分布的量化:n = 265个lgl1
/
突触、140个lgl1
/-突触。图6c示出了lgl1
/
和lgl1
/-动物1小时延长持续时间旷场测试的代表性热图,以及延长旷场测试中移动距离的量化。n = 30 lgl1
/
、33 lgl1
/-。图6d示出了窝破坏(nestlet-shredding)活动的量化。n = 19只lgl1
/
、30只lgl1
/-动物。图6e示出了物体的样品阶段24小时后新物体识别(nor)测试的识别测试阶段在的代表性热图。物体偏好的量化在样品阶段24小时后的测试阶段期间。n = 25 lgl1
/
、32 lgl1
/-。图6f示出了社会交互(si)测试中社会交互阶段期间的代表性热图。白色星号表示新小鼠的位置。si测试中交互偏好的量化。n = 25只lgl1
/
动物、33只lgl1
/-动物。* p《0.05;** p《0.01;*** p《0.001;**** p《0.0001。
25.图7a-7e是示出了lgl1
/-小鼠中的癫痫发作(seizure)阈值降低和通过nmdar阻断对社会交互的拯救的图示和示意图。图7a示出了施用50 mg/kg的ptz后lgl1
/
和lgl1
/-动物中出现抽搐/跳跃、管状尾(straub tail)和强直阵挛癫痫发作的量化。fisher精确检验。图7b示出了施用50 mg/kg的ptz后行为的首次观察的潜伏期的量化。n = 17只lgl1
/
动物、15只lgl1
/-动物。mann-whitney u统计检验。图7c示出了腹腔内注射50毫升生理盐水、30 mg/kg氯胺酮、0.3 mg/kg mk-801或20 mg/kg美金刚后,社会交互测试的社会交互阶段期间的代表性热图。白色星号表示新小鼠的位置。图7d示出了在感兴趣的社交或非社交区域花费的时间的量化,以及图7e示出了经治疗的动物的交互偏好。n = (生理盐水)17只lgl1
/
动物、19只lgl1
/-动物;(氯胺酮)14只lgl1
/
动物、17只lgl1
/-动物;(mk-801)13只lgl1
/
动物、4只lgl1
/-动物;(美金刚)19只lgl1
/
动物、17只lgl1
/-动物。* p《0.05;** p《0.01;*** p《0.001;**** p《0.0001。
26.图8a-8i是示出了条件性缺失的评估和当前动力学的特征的图示和示意图。图8a示出了来自在p7天和p8天携带了由yfp(绿色)和rosa26-tdtomato(红色)示出的slick-h的注射他莫昔芬(tamoxifen)的p14小鼠的脑切片的共焦图像。图像示出了包括腹侧海马连合和海马伞、背侧海马体和腹侧海马体的区域。比例尺:1 mm。图8b示出了未注射他莫昔芬但携带指示的slick和rosa26-tdtomato的小鼠的脑切片的共焦图像。比例尺:1 mm。图8c示出了由yfp(绿色)和tdtomato(红色)示出的slick-h和slick-a阳性动物中表达thy-1 cre的大脑区域的图像。比例尺:100 μm。图8d示出了从p14小鼠的schaffer侧支区域中ca1锥体神经元层50 μm拍摄的电子显微照片。浅色箭头表示非对称性突触。非对称性突触数量的量化:n = 6只对照动物、6只lgl1 cko动物。图8e示出了8周龄小鼠的schaffer侧支区域中ca1锥体神经元层50 μm拍摄的电子显微照片。浅色箭头表示非对称性突触。非对称性突触数量的量化:n = 5只对照动物、5只lgl1 cko动物。图8f示出了培养14div的神经元树突(map2;蓝色)的共焦图像,示出了vglut1点(绿色)和psd95点(红色)。白色箭头表示共定位的突触前的点和突触后的点。比例尺:10 μm。图8g示出了roi(包括培养的海马锥体神经元的次级树突)内点的量化。n = 5个对照胚胎、5个ko胚胎。图8h示出了来自p13-15对照和lgl1 cko小鼠的切片的mepsc动力学的量化。n = 22个对照神经元、19个lgl1 cko神经元。图8i示出了来自p13-15对照和lgl1 cko小鼠的切片的mipsc动力学的量化。n = 20个对照神经元、19个lgl1 cko神经元。* p《0.05;** p《0.01。
27.图9a-9c是示出了apkc dcko中lgl1条件性缺失和突触量化后非典型pkc过度激活
的图示和示意图。图9a示出了总细胞裂解物的蛋白质印迹和通过来自用adgfp(ctrl)或adcre(cko)处理的培养的神经元祖细胞的抗apkc抗体免疫沉淀的结果。图9b示出了来自p14对照和apkc dcko动物中ca1神经元层schaffer侧支50 μm区域中非对称性突触数量的量化。图9c示出了来自8周龄对照和apkc dcko动物中ca1神经元层schaffer侧支50μm区域中非对称性突触数量的量化。*** p《0.001。
28.图10a和10b是示出了lgl1:pkcι/λ:pkcξ tcko中突触量化的示意图。图10a示出了来自p14对照和lgl1:pkcι/λ:pkcξ tcko动物中ca1神经元层schaffer侧支50 μm区域中非对称性突触数量的量化。图10b示出了来自8周龄对照和lgl1:pkcι/λ:pkcξ tcko动物中ca1神经元层schaffer侧支50 μm区域中非对称性突触数量的量化。
29.图11a-11g是示出了在p7天lgl1缺失的另外的行为特征的示意图。图11a示出了旷场测试期间自我梳理花费的时间的量化。图11b示出了旷场试验期间直立次数的量化。图11c示出了在p7/p8 slick-h介导的lgl1缺失后y-迷宫测试中自发交替(alternation)的量化。n = 18对照、22 lgl1 cko。图11d示出了y-迷宫测试中的总进臂次数(不显著;p=0.0560)。n = 18对照、22 lgl1 cko。图11e示出了在视动反应任务期间15度头部运动的次数。n = 18对照、22 lgl1 cko。图11f示出了slick-h介导的lgl1缺失后条件性恐惧测试的量化。n = 18对照、22 lgl1 cko。图11g示出了在p7天条件性lgl1缺失后窝破坏活动的量化。n = 12只对照动物、12只lgl1 cko动物。** p《0.01。
30.图12a-12k是示出了lgl1
/-小鼠中行为表型的部分保留的图示和示意图。图12a示出了来自p14 lgl1
/
和lgl1
/-动物中ca1神经元层schaffer侧支50 μm区域中非对称性突触数量的量化。图12b示出了10分钟旷场(of)测试中lgl1
/
和lgl1
/-的热图和轨迹。图12c示出了of测试期间移动距离的量化。n = 34只对照动物、47只lgl1
/-动物。图12d示出了对照(黑色)和lgl1
/-(灰色)小鼠的趋触性的量化。图12e示出了来自新物体识别(nor)测试的样品阶段的对照和lgl1 cko之间相互作用的热图。图12f示出了样品阶段期间物体偏好的量化。n = 17只对照动物、26只lgl1
/-动物。图12g示出了来自样品阶段2分钟后进行的nor测试的物体识别阶段的相互作用的热图。白色星号表示新物体的位置。图12h示出了物体偏好的量化。正值表示对新物体的偏好。n = 17只对照动物、26只lgl1
/-动物。图12i示出了来自社会交互测试的习惯化阶段的热图和量化。n = 25只对照动物、33只lgl1
/-动物。图12j示出了来自si测试的社会新奇阶段的热图和量化。n = 25只对照动物、33只lgl1
/-动物。图12k示出了在发育和成年期早期lgl1
/
和lgl1
/-小鼠的体重增加的对比。注意p28后的间断x轴(n = 9-15只雄性lgl1
/
、9-15只雄性lgl1
/-、8-15只雌性lgl1
/
、9-14只雌性lgl1
/-)。
31.图13a-13e是示出了经生理盐水、氯胺酮和mk-801治疗的动物中社会交互测试的另外的阶段的示意图。图13a-13d示出了来自施用指定的解决方案后社会交互测试的习惯化和社会新奇阶段的交互时间和偏好的量化。白色星号表示新小鼠在社会新奇阶段的位置。n = (生理盐水)27只lgl1
/
动物、30只lgl1
/-动物;(氯胺酮)14只lgl1
/
动物、17只lgl1
/-动物;(mk-801)13只lgl1
/
,14只lgl1
/-动物;(美金刚)19只lgl1
/
动物、17只lgl1
/-动物。图13e示出了注射生理盐水或氯胺酮后窝破坏活动的量化。n = (生理盐水)15只lgl1
/
动物、14只lgl1
/-动物;(氯胺酮)15只lgl1
/
动物、12只lgl1
/-动物。* p《0.05;** p《0.01;**** p《0.0001。
32.本发明的详细说明
本发明基于n-甲基-d-天冬氨酸(nmda)受体拮抗剂以亚麻醉量缓解与行为障碍(如史密斯-马吉利综合征(sms)、自闭症或注意缺陷多动障碍(adhd))相关的社会交互缺陷的发现。
33.除非另有说明,否则本文使用的所有技术和科学术语具有与本领域普通技术人员通常理解的相同含义。出于解释本说明书的目的,将适用以下对术语的描述,并且在任何适当的情况下,以单数形式使用的术语也将包括复数形式,反之亦然。所有专利、申请、公开的申请和其他出版物均通过引用整体并入。如果所述的任何术语描述与通过引用并入本文的任何文件相冲突,应以下述术语描述为准。
34.如在本说明书和所附权利要求书中所使用的,除非上下文另有明确规定,否则单数形式“一”、“一个/一种”和“该”包括复数指代。因此,例如,对“该方法”的引用包括本文所述类型的一个或多个方法和/或步骤,这些将在阅读本公开等内容后对本领域技术人员变得显而易见。
35.术语“约”和“大约”是指在给定值或范围的20%以内、15%以内、10%以内、9%以内、8%以内、7%以内、6%以内、5%以内、4%以内、3%以内、2%以内、1%以内或更小。
36.术语“包含”可与“包括”、“含有”或“特征在于”互换使用,是包容性或开放式语言且不排除另外的、未引用的元素或方法步骤。短语“由
……
组成”不包括权利要求中未指定的任何元素、步骤或成分。短语“基本上由
……
组成”将权利要求的范围限定为具体的材料或步骤和那些不会实质性影响所要求保护的发明的基本和新颖性特征的材料或步骤。本发明考虑了与这些短语中的每一个的范围相对应的本发明的组合物和方法的实施方案。因此,包含所列举的元素或步骤的组合物或方法考虑了基本上由其组成或由其组成的组合物或方法的具体实施方案。
37.如本文所使用的,“治疗”或“处理”或“缓解”或“改善”在本文中可互换使用。这些术语是指获得有益的或预期的结果(包括但不限于治疗益处和/或预防益处)的方法。治疗益处是指根除或改善正在治疗的潜在疾病。此外,通过根除或改善与潜在疾病相关的一种或多种生理症状来实现治疗益处,从而在患者中观察到改善,尽管患者可能仍受潜在疾病的折磨。为了预防益处,可以将组合物施用于有风险发展特定疾病的患者,或者施用于报告了疾病的一种或多种生理症状的患者,即使可能尚未做出对该疾病的诊断。治疗包括:预防疾病,也就是说,通过在诱发疾病之前施用保护性组合物使疾病的临床症状不发展;抑制疾病,也就是说,通过在诱发疾病之后但在疾病的临床出现或再次出现之前施用保护性组合物使疾病的临床症状不发展;阻止疾病,也就是说,通过在最初出现后施用保护性组合物来阻止临床症状的发展;预防疾病的复发生和/或缓解疾病,也就是说,通过在初次出现后施用保护性组合物使临床症状的消退。
38.术语“有效量”或“治疗有效量”是指足以诱导期望的生物学结果的活性剂的量。该结果可以是疾病的病症、症状或病因的减轻,或者是任何其他期望的生物系统的改变。本文中使用术语“治疗有效量”来表示当在一段时间内反复应用于受影响区域时引起疾病状况显著改善的制剂的任何量。量将随治疗情况、病情进展阶段以及所用制剂的类型和浓度而变化。在任何给定情况下的适当量对于本领域技术人员来说将是显而易见的或能够通过常规实验确定。
39.当涉及用于剂量、有效量或治疗有效量时,术语“亚麻醉”是指不足以诱导麻醉的
活性剂的量(或剂量)。例如,与诱导和维持手术麻醉所需的血浆浓度9000 ng/ml至25000 ng/ml相比,氯胺酮的镇痛作用在血浆浓度100 ng/ml至200 ng/ml发生。因此,氯胺酮的示例性亚麻醉量通常为《0.5 mg/kg。
[0040]“预防有效量”是当施用于对象时将具有预期的预防效果(例如,预防、延迟或降低疾病、病症、病状或相关症状(例如sms)发生(或复发)的可能性)的药物组合物的量。通常,但不是必须的,由于预防剂量在疾病、病症或病状之前或早期在对象中使用,因此预防有效量可以小于治疗有效量。完全的治疗或预防效果不一定通过一次剂量的施用而产生,而可以仅在施用一系列剂量后产生。因此,治疗或预防有效量可以在一次或多次施用中被施用。
[0041]“对象”、“个体”或“患者”在本文中可互换使用,指脊椎动物,优选哺乳动物,更优选人类。哺乳动物包括但不限于鼠类、猿猴、人类、农场动物、运动动物和宠物。还包括在体外获得或在体外培养的生物实体的组织、细胞及其后代。
[0042]
如本文所使用的,“促进”或“增加”在本文中可互换使用。这些术语指与未经处理的细胞(组织或对象)相比,经处理的细胞(组织或对象)中测量的参数(例如,活性、表达、信号转导、神经元变性)的增加。也可以在治疗前后对同一细胞、组织或对象进行比较。这种增加足以被检测。在一些实施方案中,与未经处理的细胞相比,经处理的细胞的增加至少约10%、20%、30%、40%、50%、60%、70%、80%、90%、1倍、2倍、3倍、4倍或更多。
[0043]
如本文所使用的,“抑制”、“防止”或“减少”在本文中可互换使用。这些术语指与未经处理的细胞(组织或对象)相比,经处理的细胞(组织或对象)中测量的参数(例如,活性、表达、信号转导、神经元变性)的减少。也可以在治疗前后对同一细胞、组织或对象进行比较。这种减少足以被检测。在一些实施方案中,与未经处理的细胞相比,经处理的细胞的减少至少约10%、20%、30%、40%、50%、60%、70%、80%、90%或完全被抑制。在一些实施方案中,与未经处理的细胞相比,所测量的参数在经处理的细胞中是不可检测的(即,完全抑制)。
[0044]
术语“控制”是指对象从不会导致疾病治愈的疗法(例如,预防剂或治疗剂)中获得的有益效果。在某些实施方案中,向对象施用一种或多种疗法(例如,预防剂或治疗剂)以“控制”神经元疾病、一种或多种其症状,从而 预防疾病的进展或恶化。
[0045]
术语目标分子的“阻断剂”、“拮抗剂”和“抑制剂”在本文中可互换使用,是指例如在体内或体外减少或抑制由目标分子诱导的生物学效应的药剂。该药剂可以是小分子化合物或生物分子,例如核酸或多肽。在评估抑制强度时,可以在候选药剂存在或不存在下测量生物学效应。在某些实施方案中,在拮抗剂存在下测量的生物学效应等于或小于拮抗剂不存在下测量的相同生物学效应的10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%或98%。
[0046]
关于生物活性剂的术语“选择性抑制”或“选择性地抑制”是指与非目标信号传导活性相比,该药剂通过与目标直接或间接相互作用优先降低目标信号传导活性的能力。
[0047]
目标分子的“激动剂”是指例如在体内或体外增加或增强目标分子诱导的生物学效应的药剂。该药剂可以是小分子化合物或生物分子,例如核酸或多肽。在评估增强水平时,可以在候选药剂存在或不存在下测量生物学效应。在某些实施方案中,在激动剂不存在下测量的生物学效应等于或小于激动剂存在下测量的相同生物学效应的10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%或98%。例如,本文所述的lgl的激动剂可以是例如在表达lgl的细胞中能够激活或以其他方式增加lgl的一种或
多种生物活性的分子。在一些实施方案中,lgl的激动剂(例如,本文所述的激动性抗体)可以例如通过激活或以其他方式增加表达lgl蛋白的细胞的激活和/或细胞信号传导通路起作用,从而相对于在激动剂不存在下lgl介导的生物活性增加细胞的lgl介导的生物活性。在一些实施方案中,表达lgl蛋白的细胞是神经元,并且lgl介导的生物活性是lgl介导的神经元突触的形成。
[0048]
术语“多肽”、“肽”和“蛋白质”在本文中可互换使用,以指氨基酸残基的聚合物。该术语适用于其中一个或多个氨基酸残基是相应天然存在的氨基酸的人工化学模拟物的氨基酸聚合物,以及天然存在的氨基酸聚合物和非天然存在的氨基酸聚合物。
[0049]
术语“氨基酸”是指天然存在的和合成的氨基酸,以及以类似于天然存在的氨基酸的方式起作用的氨基酸类似物和氨基酸模拟物。天然存在的氨基酸是由遗传密码编码的氨基酸,以及后来被修饰的氨基酸,例如羟脯氨酸、γ-羧谷氨酸和o-磷酸丝氨酸。氨基酸类似物是指与天然存在的氨基酸具有相同基本化学结构的化合物,即α碳与氢、羧基、氨基和r基结合,例如高丝氨酸、去甲亮氨酸、蛋氨酸亚砜、蛋氨酸甲基砜。此类类似物具有经修饰的r基团(例如去甲亮氨酸)或经修饰的肽主干,但保留与天然存在的氨基酸相同的基本化学结构。天然编码的氨基酸是20种常见氨基酸(丙氨酸、精氨酸、天冬酰胺、天冬氨酸、半胱氨酸、谷氨酰胺、谷氨酸、甘氨酸、组氨酸、异亮氨酸、亮氨酸、赖氨酸、蛋氨酸、苯丙氨酸、脯氨酸、丝氨酸、苏氨酸、色氨酸、酪氨酸和缬氨酸)以及吡咯赖氨酸和硒代半胱氨酸。
[0050]
如本文所使用的,术语“抗体”包括天然存在的抗体以及非天然存在的抗体,包括例如单链抗体、嵌合抗体、双功能抗体和人源化抗体,以及其抗原结合片段。此类非天然存在生的抗体可以使用固相肽合成构建、可以重组产生或可以通过例如筛选由可变重链和可变轻链组成的组合文库获得(参见huse et al., science 246:1275-1281, 1989,其通过引用并入本文)。这些和其他制备例如,嵌合、人源化、cdr移植、单链和双功能抗体的方法是众所周知的(winter and harris, immunol. today 14:243-246, 1993; ward et al., nature 341:544-546, 1989; harlow and lane, antibodies: a laboratory manual (cold spring harbor laboratory press, 1999); hilyard et al., protein engineering:a practical approach (irl press 1992); borrabeck, antibody engineering, 2d ed. (oxford university press 1995);其中每一个均通过引用并入本文)。此外,经修饰的或衍生的抗体或抗体的抗原结合片段(例如聚乙二醇化(聚乙二醇修饰)的抗体)可以被用于本发明方法。
[0051]
可以使用本领域已知的各种方法来测试抗体的抗目标多肽活性。各种技术可以被用于筛选以鉴定具有期望的特异性的抗体,包括各种免疫分析,例如酶联免疫吸附分析(elisa)(包括直接和配体捕获elisa)、放射免疫分析(ria)、免疫印迹和荧光激活细胞分选(facs)。使用具有确定特异性的多克隆或单克隆抗体进行竞争性结合或免疫放射分析的许多方案是本领域众所周知的。这种免疫分析通常涉及目标多肽和特定抗体之间复合物形成的测量。优选使用与目标多肽上的两个非干扰表位反应的单克隆抗体的基于双位点单克隆的免疫分析,但也可以使用其他分析,例如竞争性结合分析。参见,例如,maddox et al, 1983, j. exp. med. 158:1211。
[0052]“表位”是抗原分子表面上与单个抗体分子结合的的位点,例如能够与抗体的一个或多个抗原结合区结合、在动物(例如哺乳动物,例如人类)中具有抗原性或免疫原性活性、
能够引发免疫反应的抗原表面上的局部区域。具有免疫原性活性的表位是在动物中引发抗体反应的多肽的一部分。具有抗原活性的表位是如通过本领域公知的任何方法(包括例如通过免疫测定法)确定的抗体结合的多肽的一部分。抗原表位不一定是免疫原性的。表位通常由分子(例如氨基酸或糖侧链)的化学活性表面基团组成,并且具有具体的三维结构特征和具体的电荷特征。抗体表位可以是线性表位或构象表位。线性表位由蛋白质中的连续的氨基酸序列形成。构象表位由蛋白质序列中不连续的氨基酸形成,但其在蛋白质折叠成其三维结构时聚集在一起。当蛋白质的三维结构处于改变的构象时形成诱导表位,例如在另一种蛋白质或配体的激活或结合后。
[0053]
如本文所使用的,术语“特异性结合”、“特异性结合至”或“特异于”特定多肽或特定多肽靶标上的表位指结合,其中分子与特定多肽或特定多肽上的表位结合,而基本上不结合任何其他多肽或多肽表位。例如,优先结合目标分子(例如分子或其抗原或表位)而不是参考分子(例如分子或其抗原或表位)的抗体可以以小于相对于参考分子表现出的kd的kd结合目标分子。在一些实施方案中,抗体以小于相对于参考分子表现出的kd的一半的kd优先结合目标分子。在一些实施方案中,抗体以小于相对于参考分子表现出的kd的至少10倍的kd优先结合目标分子。在一些实施方案中,抗体以kd优先结合目标分子,kd为相对于参考分子表现出的约75%、约50%、约25%、约10%、约5%、约2.5%或约1%。在一些实施方案中,抗体结合参考分子时表现出的kd与结合目标分子时表现出的kd之间的比例为至少2倍、至少3倍、至少4倍、至少5倍、至少10倍、至少20倍、至少100倍、至少500倍、至少103倍、至少104倍、或者至少105倍。优先结合也可以通过结合测定来确定并且通过例如荧光强度(“mfi”)来表示。
[0054]“结合感兴趣的目标分子”的分子(例如,激动剂或拮抗剂)是以足够的亲和力结合目标分子且不与其他分子发生显著交叉反应的分子,使得该分子例如用作靶向表达目标分子的细胞或组织的诊断剂或治疗剂。在此类实施方案中,分子与“非目标”分子的结合程度将小于分子与其特定目标分子结合程度的约10%,例如,如通过荧光激活细胞分选(facs)分析或ria所确定的。
[0055]
关于抗体与目标分子(例如apkc)的结合,术语“特异性结合”、“特异性结合至”或“特异于”特定多肽或特定多肽靶标上的表位是指与非特异性相互作用明显不同的结合。例如,特异性结合可以通过与对照分子的结合相比较确定分子的结合来测量,对照分子通常是不具有结合活性的结构相似的分子。例如,特异性结合可以通过与类似于靶标的对照分子竞争来确定,例如,过量的未标记靶标。在这种情况下,如果标记的靶标与探针的结合被过量的未标记靶标竞争性抑制,则表示特异性结合。如本文所使用的,术语“特异性结合”、“特异性结合至”或“特定于”特定多肽或特定多肽靶标上的表位指结合,其中分子结合与特定多肽或特定多肽上的表位,而基本上不结合任何其他多肽或多肽表位。在某些实施方案中,结合apkc的抗体的解离常数(kd)小于或等于10 nm、5 nm、4 nm、3 nm、2 nm、1 nm、0.9 nm、0.8 nm、0.7 nm、0.6 nm、0.5 nm、0.4 nm、0.3 nm、0.2 nm或0.1 nm。
[0056]
术语“变体”适用于氨基酸序列和核酸序列两者。对于具体的核酸序列,变体是指编码相同或基本相同氨基酸序列的核酸,或在核酸不编码氨基酸序列的情况下,是指基本相同的序列。由于遗传密码的简并性,大量功能相同的核酸编码任何给定的蛋白质。例如,密码子gca、gcc、gcg和gcu都编码氨基酸丙氨酸。因此,在由密码子指定的丙氨酸的每个位置,密码子可以被改变为所述的任何相应密码子,而不改变编码的多肽。这种核酸变异是“沉默变异”,是一种保守修饰变异。本文中编码多肽的每个核酸序列也描述了核酸的每个可能的沉默变异。本领域技术人员将认识到,核酸中的每个密码子(除了通常是蛋氨酸的唯一密码子的aug,以及通常是色氨酸的唯一密码子的tgg)可以被修饰以产生功能相同的分子。因此,编码多肽的核酸的每个沉默变异都隐含在所述的每个序列中。
[0057]
当涉及蛋白质或肽使用时,术语“变体”可以指与天然或未经修饰序列相比包含一个或多个(例如,约1个至约25个、约1个至约20个、约1个至约15个、约1个至约10个或约1个至约5个)氨基酸序列取代、缺失和/或与插入的肽或多肽。变体可以是天然存在的(例如等位基因变体或剪接变体)或者可以是人工构建的。多肽变体可以从编码变体的相应核酸分子制备。在具体实施方案中,蛋白质或肽的变体保留天然蛋白质或肽的功能活性。在某些实施方案中,变体由编码天然蛋白质或肽的核酸分子的单核苷酸多态性(snp)变体编码。肽的功能变体是指保留天然肽的感兴趣的至少一种功能或活性的肽的变体。
[0058]“序列同一性的百分比”通过在比较窗口中比较两个最佳对齐的序列来确定,其中与不包括插入或缺失的参考序列(例如,本发明的多肽)相比,比较窗口中的多核苷酸序列部分可以包含插入或缺失(即,缺口),以实现两个序列的最佳对齐。通过确定两个序列中出现相同核酸碱基或氨基酸残基的位置数量来计算,从而得出匹配位置的数量,将匹配位置的数量除以比较窗口中的位置总数,并将结果乘以100得出序列同一性的百分比。
[0059]
在两个或多个核酸或多肽序列的上下文中,术语“相同的”或百分比“同一性”是指两个或多个序列或子序列是相同序列。当在使用以下序列比较算法之一或通过手动对齐和目视检查测量的比较窗口或指定区域进行比较和对齐以获得最大对应时,如果两个序列具有特定百分比的氨基酸残基或核苷酸是相同的(即在指定区域,或当未指定时在整个序列,60%的同一性,任选地65%、70%、75%、80%、85%、90%或95%的同一性),则两个序列是“基本相同的”。本发明提供分别与本文举例说明的多肽基本相同的多肽及其用途,包括但不限于用于治疗或预防神经疾病或病症,例如神经退行性疾病或病症,和/或治疗sci。任选地,同一性存在于长度至少约为50个核苷酸的区域,或更优选地存在于长度为100至500或1000或更多个核苷酸的区域,或存在于参考序列的整个长度上。
[0060]“基因敲除”是指由细胞中内源性dna序列编码的蛋白质的表达的部分或全部抑制。“敲除”(ko)可以受到全部或部分编码蛋白质的基因的靶向缺失的影响。替代地,转基因生物体可以通过胚胎干细胞中功能蛋白的靶向突变获得。因此,该缺失或突变可以阻止或减少该蛋白质在正常表达的整个动物的任何细胞中的表达,或导致具有不同于正常/野生型蛋白质的生物学功能的突变蛋白质的表达。
[0061]
术语“敲除动物”和“转基因动物”是指通过与靶向载体重组抑制或突变给定基因的转基因动物。需要强调的是,该术语意在包括所有子孙后代。因此,包括亲代动物及其所有f1、f2、f3等及其后代。
[0062]
如本文所使用的,当用于描述非人类转基因哺乳动物(例如小鼠)时,短语“条件性敲除”或“cko”是指在特定组织中含有特定基因敲除的小鼠。基因工程cko小鼠的创建涉及将特定的dna序列(例如敲除构建体/载体)插入到小鼠dna中。插入的序列由通常不存在于小鼠体内的两种dna特异性酶识别,frt重组酶(也称为翻转酶)和cre重组酶。cre重组酶识别位点称为loxp位点,以及翻转酶识别位点称为frt位点。这些酶中的每一种可以切割并去除位于其识别位点两侧的dna序列。如果感兴趣基因的功能性dna序列被去除,这会导致基
因功能的破坏。此外,将可选择的标记基因插入小鼠中,该基因的引入允许选择包含cre重组或翻转酶识别位点的小鼠胚胎细胞(干细胞)。得到的小鼠是条件性敲除小鼠。
[0063]“突触”或是本领域的术语,是指允许信号从神经细胞(即神经元)传递到靶细胞(例如相邻神经元或肌肉细胞)的通信细胞-细胞连接。突触由突触前细胞的突触前膜(例如,神经元的轴突的膜)和突触后细胞的突触后膜(例如,神经元的树突的膜或肌肉或分泌细胞的特定区域的膜)组成,突触前膜和突触后膜通常彼此相对。突触的相对的突触膜之间的间隙称为突触间隙。神经元通常与其相邻的细胞形成多个突触。神经元通常充当在其轴突上形成的突触的突触前细胞,以及充当在其树突上形成的突触的突触后细胞。因此,如本文所使用的“神经元的突触前部位”是指神经元的突触部位,对该突触神经元充当突触前细胞;如本文所使用的“神经元的突触后部位”是指神经元的突出部位,对该突触神经元充当突触后细胞。
[0064]
如本文所使用的,术语“神经元”包括神经元及其部分(例如,神经元胞体、轴突或树突)。如本文所使用的,术语“神经元”表示包括中央细胞体或细胞体和两种类型的延伸或突起(树突,通常大部分神经元信号通过树突传递到细胞体;以及轴突,通常大多数神经元信号通过轴突从细胞体传递到效应细胞,例如靶神经元或肌肉)的神经系统细胞。神经元可以将来自组织和器官的信息传递到中枢神经系统(传入或感觉神经元)中,并将信号从中枢神经系统传递到效应细胞(传出或运动神经元)。其他神经元(称为中间神经元)连接中枢神经系统(大脑和脊柱)内的神经元。可以接受根据本发明的治疗或方法的神经元类型的某些具体实例包括小脑颗粒神经元、背根神经节神经元和皮质神经元。
[0065]
术语“神经元变性”被广泛使用,是指神经元细胞中的任何病理变化,包括但不限于神经元细胞的死亡或损失、细胞死亡之前的任何变化,以及神经元细胞的活性或功能的任何减少或损失。病理变化可以是自发的或可以是由任何事件诱导的,包括例如与细胞凋亡相关的病理变化。神经元可以是任何神经元,包括但不限于感觉神经元、交感神经元、副交感神经元或肠神经元,例如背根神经节神经元、运动神经元和中枢神经元,例如来自脊髓的神经元。神经元变性或细胞损失是各种神经疾病或病症(例如神经退行性疾病或病症)的特征。在一些实施方案中,神经元是感觉神经元。在一些实施方案中,神经元是运动神经元。在一些实施方案中,神经元是受损的脊髓神经元。
[0066]
在一些实施方案中,变性发生在神经元的一部分,例如神经元胞体、轴突或树突。因此,可以在神经元的变性部分中抑制变性。在一些实施方案中,抑制神经元轴突的变性。在一些实施方案中,神经元细胞体的变性被抑制。轴突可以是任何神经元的轴突。例如,在一些实施方案中,轴突是脊髓连合轴突,或上运动神经元轴突,或中枢神经系统轴突。
[0067]“给药”或“施用”是指将存在于体外的物质注射或以其他方式物理递送至患者体内的行为,通过例如粘膜、皮内、静脉内、肌肉递送和/或本文所述的或本领域已知的任何其他物理递送方法。当治疗疾病、病症、病状或其症状时,物质的施用通常发生在疾病、病症、病状或其症状发作之后。当预防疾病、病症、病状或其症状时,物质的施用通常发生在疾病、病症、病状或其症状发作之前。
[0068]
nmda受体是兴奋性毒性(过量细胞外谷氨酸过度刺激谷氨酸受体并损害神经元的过程)进展的关键。因此,nmda受体拮抗剂已被广泛研究用于治疗具有兴奋性毒性组分的疾病,如中风、创伤性脑损伤和神经退行性疾病,如亨廷顿氏病、阿尔茨海默氏症和肌萎缩侧
索硬化症。这些研究已经证明nmda受体拮抗剂在细胞培养和兴奋毒性神经退行性变的动物模型中保护神经元的有效性。然而,nmda拮抗剂在临床试验中基本上没有显示出安全性,这可能是因为抑制了正常神经元功能所必需的nmda受体功能。
[0069]
尽管谷氨酸能突触中的许多蛋白质已经被鉴定,并且它们在突触形成和功能中的作用已经被研究,但将数百种蛋白质组装成高度组织化的和动态的结构的信号传导逻辑仍不清楚。如本文所示,保守的顶端-基底极性信号传导组分lgl1定位于psd中且可能通过与其保守的结合伴侣(特别是膜相关的鸟苷酸激酶(maguk))相互作用调节关键突触蛋白和谷氨酸受体的突触数量和组成。已经研究了非典型pkc(apkc)在记忆形成和巩固中的作用。
[0070]
maguk家族的成员包括discs large同系物1(dlg1),也被称为突触相关蛋白97(sap97)、突触相关蛋白102(sap102)、突触后密集区蛋白95(psd95)和突触后密集区蛋白93(psd93),其被定义为包含pdz、sh3和guk结构域,尽管其中许多还包含与camkii、ww和l27结构域同源的区域(woods df, bryant pj (december 1993). "zo-1, dlga and psd-95/sap90: homologous proteins in tight, septate and synaptic cell junctions". mech. dev. 44 (2-3):85-9)。它们所具有的guk结构域在结构上与鸟苷酸激酶的非常相似,但由于缺乏结合atp的p环,因此已知其是无催化活性的。pdz域拷贝的数量因maguk家族的不同成员而变化。在每个家族成员中发现的pdz结构域通常有不同的结合伴侣,因为它们的氨基酸组成不同。
[0071]
在一些实施方案中,sap97具有以下氨基酸序列:mpvrkqdtqralhlleeyrsklsqtedrqlrssiervinifqsnlfqalidiqefyevtlldnpkcidrskpsepiqpvntweisslpsstvtsetlpsslspsvekyryqdedtppqehispqitnevigpelvhvseknlseienvhgfvshshispikpteavlpspptvpvipvlpvpaentvilptipqanpppvlvntdsletptyvngtdadyeyeeitlergnsglgfsiaggtdnphigddssifitkiitggaaaqdgrlrvndcilrvnevdvrdvthskavealkeagsivrlyvkrrkpvsekimeiklikgpkglgfsiaggvgnqhipgdnsiyvtkiieggaahkdgklqigdkllavnnvcleevtheeavtalkntsdfvylkvakptsmymndgyappditnsssqpvdnhvspssflgqtpasparyspvskavlgddeitreprkvvlhrgstglgfnivggedgegifisfilaggpadlsgelrkgdriisvnsvdlraasheqaaaalknagqavtivaqyrpeeysrfeakihdlreqmmnssissgsgslrtsqkrslyvralfdydktkdsglpsqglnfkfgdilhvinasddewwqarqvtpdgesdevgvipskrrvekkerarlktvkfnsktrdkgeipddmgskglkhvtsnasdsessyrgqeeyvlsyepvnqqevnytrpviilgpmkdrinddlisefpdkfgscvphttrpkrdyevdgrdyhfvtsreqmekdiqehkfieagqynnhlygtsvqsvrevaekgkhcildvsgnaikrlqiaqlypisifikpksmenimemnkrlteeqarktferamkleqeftehftaivqgdtlediynqvkqiieeqsgsyiwvpakekl (seq id no: 7)。genbank
™
登录号nm_001098424提供了示例性人类sap97核酸序列。
[0072]
在一些实施方案中,sap102具有以下氨基酸序列:mhkhqhcckcpecyevtrlaalrrleppgygdwqvpdpygpgggngasagyggyssqtlpsqagatptprtkakliptgrdvgpvppkpvpgkstpklngsgpswwpectctnrdwyeqvngsdgmfkyeeivlergnsglgfsiaggidnphvpddpgifitkiipggaaamdgrlgvndcvlrvnevdvsevvhsravealkeagpvvrlvvrrrqpppetimevnllkgpkglgfsiaggignqhipgdnsiyitkiieggaaqkdgrlqigdrllavnntnlqdvrheeavaslkntsdmvylkvakpgslhlndmyappdyastftaladnhishnsslgylgaveskvsypappqvpptryspiprhmlaeedftreprkiilhkgstglgfnivggedgegifvsfilaggpadlsgelrrgdrilsvngvnlrnatheqaaaalkragqsvtivaqyrpeeysrfeskihdlreqmmnssmssgsgslrtsekrslyvralfdydrtrdsclpsqg
lsfsygdilhvinasddewwqarlvtphgeseqigvipskkrvekkerarlktvkfhartgmiesnrdfpglsddyygaknlkgqedailsyepvtrqeihyarpviilgpmkdrvnddlisefphkfgscvphttrprrdnevdgqdyhfvvsreqmekdiqdnkfieagqfndnlygtsiqsvravaergkhcildvsgnaikrlqqaqlypiaifikpksiealmemnrrqtyeqankiydkamkleqefgeyftaivqgdsleeiynkikqiiedqsghyiwvpspekl (seq id no: 8)。genbank
™
登录号nm_021120提供了示例性人类sap102核酸序列。
[0073]
在一些实施方案中,psd95具有以下氨基酸序列:mdclcivttkkyryqdedtpplehspahlpnqansppvivntdtleapgyvngtegemeyeeitlergnsglgfsiaggtdnphigddpsifitkiipggaaaqdgrlrvndsilfvnevdvrevthsaavealkeagsivrlyvmrrkppaekvmeiklikgpkglgfsiaggvgnqhipgdnsiyvtkiieggaahkdgrlqigdkilavnsvgledvmhedavaalkntydvvylkvakpsnaylsdsyappdittsysqhldneishssylgtdyptamtptsprryspvakdllgeedipreprrivihrgstglgfnivggedgegifisfilaggpadlsgelrkgdqilsvngvdlrnasheqaaialknagqtvtiiaqykpeeysrfeakihdlreqlmnsslgsgtaslrsnpkrgfyiralfdydktkdcgflsqalsfrfgdvlhvidasdeewwqarrvhsdsetddigfipskrrverrewsrlkakdwgsssgsqgredsvlsyetvtqmevhyarpiiilgptkdranddllsefpdkfgscvphttrpkreyeidgrdyhfvssrekmekdiqahkfieagqynshlygtsvqsvrevaeqgkhcildvsanavrrlqaahlhpiaifirprslenvleinkriteeqarkafdratkleqeftecfsaivegdsfeeiyhkvkrviedlsgpyiwvparerl (seq id no: 9)。genbank
™
登录号nm_001128827提供了示例性人类psd95核酸序列。
[0074]
在一些实施方案中,psd93具有以下氨基酸序列:mgifksslfqalldiqefyevtllnsqksceqkieeanqvlqkwektsllapchdrlqksseltdcsgskenascieqnkenqsfenetdetttqnqgrcpaqncsveapawmpvhhctkyryqdedaphdhslprlthevrgpelvhvseknlsqienvhgyvlqshisplkaspapiivntdtldtipyvngteieyefeeitlergnsglgfsiaggtdnphigddpgifitkiipggaaaedgrlrvndcilrvnevdvsevshskavealkeagsivrlyvrrrrpiletvveiklfkgpkglgfsiaggvgnqhipgdnsiyvtkiidggaaqkdgrlqvgdrllmvnnysleevtheeavailkntsevvylkvgkpttiymtdpygppdithsysppmenhllsgnngtleyktslppispgryspipkhmlvdddytrppepvystvnklcdkpasprhyspvecdksfllsapyshyhlgllpdsemtshsqhstatrqpsmtlqravslegeprkvvlhkgstglgfnivggedgegifvsfilaggpadlsgelqrgdqilsvngidlrgasheqaaaalkgagqtvtiiaqyqpedyarfeakihdlreqmmnhsmssgsgslrtnqkrslyvramfdydkskdsglpsqglsfkygdilhvinasddewwqarrvmlegdseemgvipskrrverkerarlktvkfnakpgvidskgsfndkrkksfifsrkfpfyknkeqseqetsdpergqedlilsyepvtrqeinytrpviilgpmkdrinddlisefpdkfgscvphttrpkrdyevdgrdyhfvisreqmekdiqehkfieagqyndnlygtsvqsvrfvaergkhcildvsgnaikrlqvaqlypiaifikprsleplmemnkrlteeqakktydraikleqefgeyftaivqgdtlediynqcklvieeqsgpfiwipskekl (seq id no: 10)。genbank
™
登录号nm_001142699提供了示例性人类psd93核酸序列。
[0075]
蛋白激酶c(pkc)是一个蛋白激酶家族,通过磷酸化蛋白质或该家族的成员上丝氨酸和苏氨酸氨基酸残基的羟基参与控制其他蛋白质的功能。pkc酶反过来被信号激活,如二酰甘油(dag)或钙离子(ca
²⁺
)浓度的增加(wilson ch, ali es, scrimgeour n, martin am, hua j, tallis ga, rychkov gy, barritt gj (2015). "steatosis inhibits liver cell store-operated ca
²⁺ꢀ
entry and reduces er ca
²⁺ꢀ
through a protein kinase c-dependent mechanism". the biochemical journal. 466 (2): 379-90)。在人类中pkc家族由15种同工酶组成。根据它们的第二信使需求,它们被分为三个亚族:传统(或
经典)、新型和非典型(nishizuka y (apr 1995). "protein kinase c and lipid signaling for sustained cellular responses". faseb journal. 9 (7): 484-96)。传统(c)pkc含有同工型α、βi、βii和γ。这些需要ca
²⁺
、dag和磷脂(如磷脂酰丝氨酸)用于激活。新型(n)pkc包括同工型δ、ε、η和θ,以及需要dag但不需要ca
²⁺
用于激活。然而,非典型pkc(apkc)(包括蛋白激酶cζ和ι/λ同工型)既不需要ca
²⁺
也不需要二酰甘油用于激活。
[0076]
在一些实施方案中,apkcι/λ同工型具有以下氨基酸序列:mptqrdsstmshtvagggsgdhshqvrvkayyrgdimithfepsisfeglcnevrdmcsfdneqlftmkwideegdpctvssqleleeafrlyelnkdsellihvfpcvperpgmpcpgedksiyrrgarrwrklycanghtfqakrfnrrahcaictdriwglgrqgykcinckllvhkkchklvtiecgrhslpqepvmpmdqssmhsdhaqtvipynpsshesldqvgeekeamntresgkassslglqdfdllrvigrgsyakvllvrlkktdriyamkvvkkelvnddedidwvqtekhvfeqasnhpflvglhscfqtesrlffvieyvnggdlmfhmqrqrklpeeharfysaeislalnylhergiiyrdlkldnvlldseghikltdygmckeglrpgdttstfcgtpnyiapeilrgedygfsvdwwalgvlmfemmagrspfdivgssdnpdqntedylfqvilekqiriprslsvkaasvlksflnkdpkerlgchpqtgfadiqghpffrnvdwdmmeqkqvvppfkpnisgefgldnfdsqftnepvqltpddddivrkidqsefegfeyinpllmsaeecv (seq id no: 1)。genbank
™
登录号nm_002740提供了示例性人类蛋白激酶c iota核酸序列。
[0077]
在一些实施方案中,apkc ζ同工型1具有以下氨基酸序列:mpsrtgpkmegsggrvrlkahyggdifitsvdaattfeelceevrdmcrlhqqhpltlkwvdsegdpctvssqmeleeafrlarqcrdegliihvfpstpeqpglpcpgedksiyrrgarrwrklyranghlfqakrfnrraycgqcseriwglarqgyrcinckllvhkrchglvpltcrkhmdsvmpsqeppvddknedadlpseetdgiayisssrkhdsikddsedlkpvidgmdgikisqglglqdfdlirvigrgsyakvllvrlkkndqiyamkvvkkelvhddedidwvqtekhvfeqassnpflvglhscfqttsrlflvieyvnggdlmfhmqrqrklpeeharfyaaeicialnflhergiiyrdlkldnvlldadghikltdygmckeglgpgdttstfcgtpnyiapeilrgeeygfsvdwwalgvlmfemmagrspfdiitdnpdmntedylfqvilekpiriprflsvkashvlkgflnkdpkerlgcrpqtgfsdikshaffrsidwdllekkqalppfqpqitddygldnfdtqftsepvqltpddedaikridqsefegfeyinplllsteesv (seq id no: 2)。genbank
™
登录号nm_002744提供了示例性人类蛋白激酶c zeta核酸序列。
[0078]
在一些实施方案中,apkc ζ同工型2具有以下氨基酸序列:mdsvmpsqeppvddknedadlpseetdgiayisssrkhdsikddsedlkpvidgmdgikisqglglqdfdlirvigrgsyakvllvrlkkndqiyamkvvkkelvhddedidwvqtekhvfeqassnpflvglhscfqttsrlflvieyvnggdlmfhmqrqrklpeeharfyaaeicialnflhergiiyrdlkldnvlldadghikltdygmckeglgpgdttstfcgtpnyiapeilrgeeygfsvdwwalgvlmfemmagrspfdiitdnpdmntedylfqvilekpiriprflsvkashvlkgflnkdpkerlgcrpqtgfsdikshaffrsidwdllekkqalppfqpqitddygldnfdtqftsepvqltpddedaikridqsefegfeyinplllsteesv (seq id no: 3)。genbank
™
登录号nm_001033581提供了另一个示例性人类蛋白激酶c zeta核酸序列。
[0079]
vangl家族蛋白是非经典wnt平面细胞极性通路的组分。在人类中,至少有三种vangl蛋白(vangl1和vangl2)属于这个家族。除非另有说明,术语“van gogh样蛋白”、“vang样蛋白”、“vangl平面细胞极性蛋白”或“vangl”包含来自任何脊椎动物来源(包括哺乳动物如灵长类动物(如人类和食蟹猴(猕猴))、狗和啮齿类(如小鼠和大鼠))的多肽(“多肽”和“蛋白质”在本文中可互换使用),包括任何天然多肽。在某些实施方案中,术语包括“相关的vangl多肽”,包括其snp变体。术语“vangl”还包括“全长”、未经处理的vangl以及在细胞中
处理产生的任何形式的vangl。
[0080]
在一些实施方案中,vangl1具有以下氨基酸序列:mdtestysgysyysshskkshrqgertrerhksprnkdgrgseksvtiqpptgepllgndstrteevqddnwgetttaitgtsehsisqediariskdmedsvgldckrylgltvasflgllvfltpiafillppilwrdelepcgticeglfismafkllilligtwalffrkrradmprvfvfralllvliflfvvsywlfygvrildsrdrnyqgivqyavslvdallfihylaivllelrqlqpmftlqvvrstdgesrfyslghlsiqraalvvlenyykdftiynpnlltaskfraakhmaglkvynvdgpsnnatgqsramiaaaarrrdsshnelyyeeaeherrvkkrkarlvvaveeafihiqrlqaeeqqkapgevmdpreaaqaifpsmaralqkylritrqqnyhsmesilqhlafcitngmtpkaflerylsagptlqydkdrwlstqwrlvsdeavtnglrdgivfvlkcldfslvvnvkkipfiilseefidpkshkfvlrlqsetsv (seq id no: 4)。genbank
™
登录号nm_138959提供了另一个示例性人类vangl1核酸序列。
[0081]
在一些实施方案中,vangl2具有以下氨基酸序列:mdtesqysgysyksghsrssrkhrdrrdrhrsksrdggrgdksvtiqapgeplldnestrgderddnwgetttvvtgtsehsishddltriakdmedsvpldcsrhlgvaagatlallsfltplaflllppllwreelepcgtaceglfisvafkllilllgswalffrrpkaslprvfvlrallmvlvfllvvsywlfygvrildarersyqgvvqfavslvdallfvhylavvllelrqlqpqftlkvvrstdgasrfynvghlsiqrvavwilekyyhdfpvynpallnlpksvlakkvsgfkvyslgeenstnnstgqsraviaaaarrrdnshneyyyeeaeherrvrkrrarlvvaveeafthikrlqeeeqknprevmdpreaaqaifasmaramqkylrttkqqpyhtmesilqhlefcithdmtpkaflerylaagptiqyhkerwlakqwtlvseepvtnglkdgivfllkrqdfslvvstkkvpffklseefvdpkshkfvmrlqsetsv (seq id no: 5)。genbank
™
登录号nm_020335提供了另一个示例性人类vangl2核酸序列。
[0082]
幼虫巨大致死性基因(lgl1)是高度保守的顶端-基底极性信号传导通路的关键组分,其沿顶端和基底外侧轴极化上皮细胞和组织。lgl1与scribble和discs large(dlg)形成基底外侧复合物,并且相互排斥和对抗顶端复合物(如apkc/par3/par6复合物)的功能。lgl1与极化胞吐作用有关,并且对建立或维持顶端-基底极性是必需的(betschinger et al., 2003; georgiou et al., 2008; macara, 2004; yamanaka et al., 2006; yamanaka et al., 2003)。dlg同源物是重要的突触后支架蛋白,称为maguk(zhu et al., 2016)。maguk蛋白在突触后密集区组织和谷氨酸受体运输和聚集中起着重要作用。此外,lgl1已被证明与fmrp(许多突触组分的翻译调节因子)相关联并共同运输(zarnescu et al., 2005)。已知顶端-基底极性通路和平面极性通路相互作用。例如,最近已显示顶端-基底极性信号传导可调节pcp信号传导的位置(chuykin et al., 2018)。lgl1基因座位于sms ~950 kb的精确共有缺失位点内,其已在携带染色体缺失的患者的遗传学研究中被报告(vlangos et al., 2003)。
[0083]
在一些实施方案中,lgl1具有以下氨基酸序列:mmkfrfrrqgadpqreklkqelfafnktvehgfpnqpsalafdpelrimaigtrsgavkiygapgveftglhrdaatvtqmhfltgqgrllsllddsslhlweivhhngcahleealsfqlpsrpgfdgasaplsltrvtvvllvaasdiaalgtegssvffldvttltllegqtlapgevlrsvpddyrcgkalgpveslqghlrdptkiligysrgllviwnqasqcvdhiflgnqqleslcwgrdsstvvsshsdgsyavwsvdagsfptlqptvattpygpfpckainkilwrncesgghfiifsggmprasygdrhcvsvlraetlvtldftsriidfftvhstrpedefddpqalavlleeelvvldlqtpgwpavpapylaplhssaitcsahvasvpaklwarivsageqqspqpvssalswpitggrnlaqepsqrgllltghedgtvrfwdasgvalrplyklstaglfqtdcehadslaqaaeddwppfrkvgcfdpysddprlgvqkvalck
ytaqmvvagtagqvlvlelsdvpveqavsvaiidllqdregftwkgherlsprtgplpwpagfqprvlvqclppaavtavtlhtewslvafgtshgfglfdyqrkspvlarctlhpndslamegplsrvkslkkslrqsfrrirksrvsgkkraanassklqeanaqlaeqacphdvemtpvqrrieprsaddslsgvvrclyfadtflrdgahhgptmwagtnsgsvfayalevpaaavggekrpeqaveavlgkevqlmhrapvvaiavldgrgrplpepyeasrdlaqapdmqgghavliaseeqfkvftlpkvsaktkfkltahegcrvrkvalatfasvacedyaetclacltnlgdvhvfsvpglrpqvhyscirkedisgiascvftrhgqgfylispseferfslsarniteplcsldinwprdatqasyrirespklsqangtpsillapqsldgspdpahsmgpdtpeppeaalspmsidsatsadttldttgdvtvedvkdflgsseeseknlrnlaedeahacailik (seq id no: 6)。genbank
™
登录号nm_004140提供了示例性人类lgl1核酸序列。
[0084]
如本文所示,成年期中lgl1的条件性敲除(cko)导致谷氨酸能突触数量增加,ampa/nmda比例降低。此外,成年lgl1的缺失导致ltp诱导和配对脉冲易化受损,表明lgl1是成年突触可塑性所需的。在sms中lgl1经常缺失,因此,进行了大量的行为测试。无论是在p7天还是成年期,lg11的条件性敲除导致行为缺陷,包括多动、认知障碍和社会交互,与sms中自闭症样症状一致。由于sms涉及一条染色体的微缺失,对lgl1
/-小鼠进行了分析,发现lgl1
/-小鼠的突触数量增加、社会交互受损、刻板重复行为增加,表明lgl1是导致sms的自闭症样症状的候选基因。
[0085]
如本文所示,在p7和p8 apkc的两种同工型的条件性缺失导致成年期突触数量减少以及认知和社交缺陷。lgl1或apkc两者的缺失都会导致行为改变的观察表明,突触数量必须针对行为功能进行最佳控制,并且任何方向的改变都会导致损伤。lgl1和apkc同工型的三重条件性敲除挽救了非对称性突触的数量和认知功能,支持它们在突触形成中的拮抗功能。
[0086]
lgl1与discs large形成基础复合物,其是maguk的同源物,突触后密集区中的基本支架蛋白,其调节谷氨酸受体的运输和聚集。发现在lg11 cko中,谷氨酸能突触中maguk的组成发生了改变,sap97(dlg1)在总的和膜表面级分两者中显著增加,并且其他三种maguk(sap102、psd95和psd93)显著减少。
[0087]
因此,lgl1的损失可以导致maguk和谷氨酸受体组成的改变,例如ampa/nmda比例的降低。由于ampa受体水平保持不变,ampa/nmda比例的降低可能是由nmda电流增加引起的。这有点令人惊讶,因为更多的nmdar通常与可塑性增加相关,但发现ltp受损。应该注意的是,在lgl1 cko中也观察到配对脉冲比例的降低,表明也可以是突触前缺陷(图3n)。此处使用的cre系(line)slick-h在ca3和ca1锥体神经元两者中表达creert2。虽然发现lgl1存在于突触后密集区中(图1i),lgl1也可以在突触前侧具有功能。配对脉冲比例的降低可以有助于突触功能的改变。
[0088]
由于lgl1位于sms的微缺失区域,因此使用四种不同的方法来缺失lgl1以测试行为结果,特别是与sms症状相关的行为后果。此外,进行了一些行为测试。来自p7和p8的lg11的cko导致行为缺陷,包括多动、认知障碍和社会交互,与sms中的自闭症样症状一致。由于sms涉及一条染色体的微缺失,对lgl1
/-小鼠进行了分析,发现lgl1
/-小鼠的突触数量增加、社会交互受损、刻板重复行为增加,表明lgl1是导致lgl1缺失的sms的自闭症样症状的候选基因。重复行为涉及纹状体,其中creer
t2
在slick-h系中不表达。在不受理论约束的情况下,这可以解释为什么只在lgl1
/-中观察到重复性行为缺陷。lgl1
/-对旷场的适应速度减慢,并且对新物体的识别也有缺陷。总之,局部损失lgl1的两个拷贝或全局损失一个拷贝
会导致与sms的自闭症样症状子集相关的行为缺陷。有趣的是,lgl1 cko和lgl1
/-动物没有表现出在旷场观察期间梳理行为或在它们的笼子里过度自我梳理的迹象(其他asd样的小鼠模型的特征(pe
ç
a et al., 2011))的增加。即使是在成年期,lgl1对维持正常的突触数量和突触的正常功能似乎是重要的,因为在6周时缺失lgl1仍然会导致突触数量增加和突触结构和功能的改变。因此,lgl1可以是成年期突触可塑性所需的关键分子,因为cko导致长期增强功能受损。因此,sms中lgl1的缺失可能是行为症状的神经生物学基础。这些研究还产生了用于sms的小鼠模型(lgl1
/-小鼠),用于了解疾病机制和治疗进展。事实上,观察到nmdar的阻断挽救社交缺陷,表明nmdar可能是lgl1缺失的sms的有希望的治疗靶标。
[0089]
含有多于一个转基因构建体和/或多于一个转基因表达构建体的动物可以通过几种方式中的任何一种进行制备。一种示例性的制备方式是产生一系列动物,每个动物含有一种期望的转基因表型。这些动物通过一系列杂交、回交和选择在一起繁殖,最终生成一只包含所有期望的转基因性状和/或表达构建体的动物,除存在构建体和/或转基因外,该动物在其他方面与野生型同源(遗传上相同)。
[0090]
通常选择胚胎干细胞(es)是因为它们能够整合并成为发育中胚胎的生殖系的一部分,从而产生转基因的生殖系传递。因此,能够这样做的任何es细胞系都适用于本文。使用本领域技术人员熟知的方法产生和维持es细胞,如doetschman et al. (1985) j. embryol. exp. mol. biol. 87:27-45所述。可以使用任何es细胞系,然而,选择的细胞系通常是根据细胞整合并成为发育中胚胎的生殖系的一部分,从而产生转基因/敲除构建体的生殖系传递的能力来选泽的。因此,被认为具有这种能力的任何es细胞系都适用于本文。一种通常用于生产es细胞的小鼠品系是129j品系。另一种es细胞系是小鼠细胞系d3(american type culture collection, catalog no. ckl 1934)。还有另一种es细胞系是ww6细胞系(ioffe et al. (1995) pnas 92:7357-7361)。使用本领域技术人员熟知的方法培养细胞并准备用于敲除构建体插入,例如robertson(teratocarcinomas and embryonic stem cells: a practical approach, e. j. robertson, ed. irl press, washington, d.c. (1987))和 bradley et al. (1986) current topics in devel. biol. 20:357-371)和hogan et al. (manipulating the mouse embryo: a laboratory manual, cold spring harbor laboratory press, cold spring harbor, n.y. (1986))中所述。
[0091]
可以使用本领域已知的多种方法(包括例如电穿孔、显微注射和磷酸钙处理)将敲除构建体引入es细胞。为了引入dna序列,在选择插入方法的适当条件下,将敲除构建体dna添加到es细胞中。如果要对细胞电穿孔,使用电穿孔机(电穿孔器)并遵循制造商的使用指南,将es细胞和构建体dna暴露于电脉冲中。电穿孔后,细胞在合适的孵育条件下恢复。然后筛选存在敲除构建体的细胞。可以使用多种方法筛选含有转基因(同源重组体)的细胞。例如,如本文所述,可以根据需要对细胞进行处理,以使其中的dna可用于通过聚合酶链反应(pcr)用特定探针进行筛选。
[0092]
一旦确定合适的es细胞,就使用标准方法将其引入胚胎。例如,可以使用显微注射来引入它们。例如通过对怀孕雌性的子宫进行灌注,获得处于适合胚胎干细胞整合发生的发育阶段的胚胎。例如,可以获得发育3-4天的小鼠胚胎,并使用微量移液器注射es细胞。将es细胞引入胚胎后,将胚胎引入假孕雌性小鼠的子宫。选择假孕阶段以增加成功植入的机会。在小鼠中,2-3天的假孕雌性小鼠是合适的。
[0093]
成功地将es细胞并入植入的胚胎会产生被称为嵌合体的后代。通过标准方法鉴定能够对突变等位基因进行种系传播的嵌合体。培育嵌合体,并筛选产生的后代是否存在期望的改变(例如,经修饰的重组ryk等位基因)。例如,这可以基于被毛颜色或通过使用已知方法(例如southern分析、斑点印迹分析、pcr分析)从后代获得dna(例如尾部dna)以评估转基因来完成。转基因表达也可以通过已知的方法(如northern分析或pcr分析)进行评估(例如,确定是否表达了替代构建体)。可以对后代dna(例如尾部dna)进行southern杂交或pcr分析以确定期望的基因型。用于获得完全由es细胞衍生的转基因非人类生物体的合适技术描述于wo 98/06834,其通过引用并入本文。
[0094]
如上所述,上述“敲除”和/或“敲入”构建体的同源重组有时很少见,并且这种构建体可以非同源地插入基因组的随机区域,在该区域中其对已被靶向缺失的基因没有影响,并且其可能会重组从而破坏另一个原本不打算改变的基因。这种非同源重组事件可以通过修饰上述靶向载体来进行选择,从而使其在任一端侧接阴性可选择标记物(特别是通过使用白喉毒素基因、胸腺嘧啶核苷激酶基因,其多肽产物可以在本领域熟知的适当组织培养基(例如,含有如更昔洛韦等药物的培养基)中表达细胞系时被选择。所得的靶向载体(包含阴性可选择标记物)和基因组之间的非同源重组通常会导致这些阴性可选择标记物基因中的一个或两个的稳定整合,因此经过非同源重组的细胞可以通过在适当的选择培养基(例如,含有更昔洛韦等药物的培养基)中生长来选择。同时选择阳性可选择标记物和针对阴性可选择标记物将导致克隆的大量富集,其中构建体已在打算突变的基因的基因座上同源重组。可以通过本领域技术人员熟知的southern印迹分析技术来确认所产生的干细胞系中目标基因位点处预测的染色体改变的存在。或者,可以使用pcr。
[0095]
其他制造转基因动物的方法也广为人知。参见,例如manipulating the mouse embryo, (cold spring harbor laboratory press, cold spring harbor, n.y., 1986)。还可以产生依赖重组酶的转基因生物,例如通过同源重组来插入目标序列,从而通过重组酶序列控制ryk基因失活的组织特异性和/或时序调控。
[0096]
因此,在另一方面,本发明提供了一种转基因非人类哺乳动物,其基因组包括lgl1基因的杂合或纯合缺失、失活或敲除以及pkc的杂合或纯合缺失、失活或敲除。在各种实施方案中,转基因非人类哺乳动物是小鼠。在各种实施方案中,小鼠基因组进一步包含pkcι/λ和pkcξ同工型的杂合或纯合缺失、失活或敲除。
[0097]
如本文所示,在亚麻醉量下使用nmda拮抗剂(如氯胺酮、美金刚或地佐环平(mk-801))也可以缓解社会交互障碍。通过阻断nmda受体(nmda),这种nmda拮抗剂可以减轻因过度连接和nmda活性增加而引起的异常活性模式,这可能是lgl1 cko和lgl1
/-动物中观察到的行为缺陷的基础。了解lgl1的功能也可能通过表达lgl1或其下游效应子,或通过抑制因lgl1缺失而被解除抑制的下游信号传导,在未来产生拯救sms症状的方法。
[0098]
因此,本发明提供了一种用于减少或预防神经元群体中幼虫巨大致死性基因(lgl1)诱导的突触增加的方法,其包含使神经元与亚麻醉量的n-甲基-d-天冬氨酸(nmda)受体拮抗剂接触。在各种实施方案中,nmda受体拮抗剂增加神经元中ampa/nmda比例。在各种实施方案中,nmda受体拮抗剂选自氯胺酮、美金刚、地佐环平(mk-801)、右美沙芬(dxm)、苯环己哌啶(pcp)、甲氧西敏(mxe)、一氧化二氮(n2o)及其任何组合。
[0099]
在各种实施方案中,该方法还可以包括使神经元群体与非典型蛋白激酶c(apkc)
抑制剂接触。在各种实施方案中,apkc抑制剂是小分子化合物、核酸分子或多肽。在各种实施方案中,apkc抑制剂是与一种或多种蛋白激酶c ζ(apkcζ)和蛋白激酶c ι/λ(apkcι/λ)同工型结合的抗体或抗原结合片段。在各种实施方案中,apkc抑制剂减少或抑制神经元中apkcζ和apkcι/λ的表达。
[0100]
在各种实施方案中,该方法还可以包括使神经元群体与lgl1激动剂接触。在各种实施方案中,lgl1激动剂是诱导lgl1的表达的小分子化合物、核酸分子或多肽。在各种实施方案中,lgl1激动剂是诱导信号传导通路的小分子化合物、核酸分子或多肽,该信号传导通路诱导lgl1的表达。
[0101]
在各种实施方案中,神经元群体在对象中,并且其中接触步骤通过向对象施用nmda受体拮抗剂和/或apkc抑制剂和/或lgl1激动剂进行。在各种实施方案中,神经元群体在对象的脑中。在各种实施方案中,对象患有或有风险发展神经退行性疾病或行为障碍,例如史密斯-马吉利综合征(sms)、自闭症或注意缺陷多动障碍(adhd)。在各种实施方案中,nmda受体拮抗剂缓解与行为障碍相关的社会交互缺陷。另一方面,本发明提供了一种控制、预防或治疗对象中神经退行性疾病或行为障碍的方法,其包含向对象施用亚麻醉量的n-甲基-d-天冬氨酸(nmda)受体拮抗剂。在各种实施方案中,nmda受体拮抗剂增加神经元中ampa/nmda比例。在各种实施方案中,nmda受体拮抗剂选自氯胺酮、美金刚、地佐环平(mk-801)、右美沙芬(dxm)、苯环己哌啶(pcp)、甲氧西敏(mxe)、一氧化二氮(n2o)及其任何组合。
[0102]
在各种实施方案中,该方法还可以包括向对象施用非典型蛋白激酶c(apkc)抑制剂。在各种实施方案中,apkc抑制剂是小分子化合物、核酸分子或多肽。在各种实施方案中,apkc抑制剂是与一种或多种蛋白激酶c ζ(apkcζ)和蛋白激酶c ι/λ(apkcι/λ)同工型结合的抗体或抗原结合片段。在各种实施方案中,apkc抑制剂减少或抑制神经元中apkcζ和apkcι/λ的表达。在各种实施方案中,该方法还可以包括向对象施用lgl1激动剂。在各种实施方案中,lgl1激动剂是诱导lgl1的表达的小分子化合物、核酸分子或多肽。在各种实施方案中,lgl1激动剂是诱导信号传导通路的小分子化合物、核酸分子或多肽,该信号传导通路诱导lgl1的表达。
[0103]
在各种实施方案中,对象中神经元群体中的突触数量减少。在各种实施方案中,突触是兴奋性突触。在各种实施方案中,突触是谷氨酸能突触。在各种实施方案中,nmda受体拮抗剂缓解与行为障碍相关的社会互动缺陷。在各种实施方案中,神经元群体包含小脑颗粒神经元、背根神经节神经元、皮质神经元、交感神经元或海马神经元。
[0104]
另一方面,本发明提供了一种通过lgl1的调控治疗行为障碍(如sms)的方法。在各种实施方案中,该方法包括以亚麻醉量向有此需要的对象施用n-甲基-d-天冬氨酸(nmda)受体拮抗剂(如氯胺酮),从而缓解与行为障碍相关的社会交互缺陷。通过阻断nmda受体,氯胺酮可以减轻过度连接和nmdar活性增加引起的异常活性模式。在各种实施方案中,该方法还可以包括向对象单独或与lgl1激动剂组合施用非典型蛋白激酶c(apkc)抑制剂。用于本文所提供的方法的示例性nmda受体拮抗剂包括但不限于氯胺酮、美金刚(namzaric
®
)、地佐环平(mk-801)、右美沙芬(dxm)、苯环己哌啶(pcp)、甲氧西敏(mxe)和一氧化二氮(n2o)。
[0105]
另一方面,本发明提供了一种选择突触形成的调节剂或突触维持的调节剂的方法。在各种实施方案中,该方法包括提供具有lgl1的杂合或纯合缺失、失活或敲除的神经元
群体;测量神经元中ampa/nmda比例的第一水平;使候选药剂与神经元群体接触;测量神经元中ampa/nmda比例的第二水平;以及如果联系的第二水平与联系的第一水平不同,选择候选药剂作为调节剂。在各种实施方案中,候选药剂是小分子化合物、核酸或肽。在各种实施方案中,核酸是microrna、sirna或基于crispr基因编辑的构建体。在各种实施方案中,候选药剂是抗体或其抗原结合片段。在各种实施方案中,该方法在apkc的存在下进行。在各种实施方案中,细胞群体在非人类哺乳动物中,并且其中接触步骤通过向非人类哺乳动物施用候选药剂进行。
[0106]
在体内lgl1条件性敲除增加谷氨酸能突触数量,降低ampa/nmda比例谷氨酸能突触形成在出生后不久开始。lgl1在发育的早期阶段(包括神经发生)中发挥作用。为了避免早期发育缺陷,从出生后第7天(p7)开始,使用可诱导的cre系,slick-h(,在海马锥体神经元中条件性地敲除lgl1图8a-8c)(heimer-mcginn and young, 2011)。在p7和出生后第8天(p8)腹腔内注射他莫昔芬,并在出生后第14天(p14)处死动物并灌注用于电子显微镜观察。辐射层中有非对称性和对称性突触。图像拍摄自脑切片中ca1细胞体层150-200 μm。观察到树突棘上形成的非对称性(兴奋性)突触密度增加了28.7%(图1a)。轴-树突突触的密度没有变化。在这些切片中没有观察到对称性(抑制性)突触密度的显著变化。
[0107]
为了确定增加的突触是否持续存在,将在p7和p8注射他莫昔芬的8周龄成年小鼠切片固定。在这些动物中,观察到ca1细胞体层150-200 μm的非对称性突触密度增加了31.8%(图1b)。与幼年动物相比,成年动物在ca1细胞层50 μm的区域中对称性突触密度显著增加25.5%。对称性突触增加的延迟表明,其不是由lgl1的功能的损失直接引起的,而可能是由海马回路的稳态反应引起的。ca1细胞层50 μm区域中非对称性突触数量也显示出增加(图8d-8e)。
[0108]
为了评估增加的突触的功能,从来自p14、15对照和lgl1 cko动物的急性脑切片中记录了微型兴奋性突触后电流(mepsc)和微型抑制性突触后电流(mipsc)。突触电流的频率和振幅的量化表明,mepsc频率增加了38%,而振幅没有显著变化,表明突触数量增加,但ampa-r组分相似(图1c)。在mipsc电流中未观察到显著变化(图1d),与本文提供的电子显微镜数据一致。与此一致的是,在体外培养14天(div)时,来自携带lgl1种系缺失的小鼠的培养的海马神经元也显示psd95和vglut1 点之间的共定位增加(图8f和8g)。mepsc动力学没有示出显著的变化(图8h),而mipsc动力学仅在衰减时间常数中示出显著降低,但在其他测量中没有(图8i)。
[0109]
为了确定树突棘密度是否受到影响,用alexa555染料填充固定脑切片中的神经元,以观察黄色荧光蛋白(yfp)阳性ca1锥体神经元中的树突棘(图1e)。发现lgl1 cko小鼠中整体树突棘密度确实增加了38%(图1f)。在p14还表征了树突棘的形态。与对照组相比,lgl1 cko小鼠示出了更多的蘑菇树突棘(mushroom spine)且粗短树突棘(stubby spine)的比例减少(图1g)。由于蘑菇树突棘代表稳定的突触,而细树突棘不稳定,这表明lgl1条件性敲除(cko)导致功能性超连接电路。
[0110]
与此一致,然后从电子显微照片中量化超微结构,发现在p14和8周龄时,突触后致密区(psd)的长度增加,突触间隙的间隙减小(图1h)。由于lgl1与maguk蛋白相互作用,亚细胞定位被确定,并且发现lgl1存在于突触后致密区中(图1i)。还测量了ampa/nmda比例,发
现在lgl1 cko中该比例降低,表明谷氨酸受体运输或功能改变(图1j)。
[0111]
lgl1通过抑制非典型pkc负调节突触数量由于lgl1和apkc在细胞极性信号传导中相互拮抗,因此认为lgl1可以通过抑制apkc来调节突触的形成。为了验证lgl1是否也抑制神经元中的apkc,测试了它们在来自e11.5小鼠端脑的神经祖细胞中的相互作用。lgl1 cko和对照细胞分别通过ad5-cmv-cre和ad5-cmv-gfp腺病毒(vector development laboratory, baylor college of medicine)处理培养物生成,并证实lgl1蛋白在cko中完全丢失(图9a)。然后测试lgl1 cko中apkc活性(如与par3的联系所证明的)是否增加。发现在lgl1 cko中活化的apkc(在t555处磷酸化)和apkc与par3的相互作用强烈增加,证实了顶端信号传导的增加和基底侧信号传导的减少。
[0112]
小鼠中有两种apkc同工型。因此,使用slick-h的他莫昔芬诱导的apkc缺失的相同策略用于删除apkc的两种同工型pkcι/λ和pkcξ,以消除补偿的可能性。在该缺失之后,从来自2周龄和8周龄动物的ca1细胞层的schaffer 侧支150
ꢀµ
m计算非对称性和对称性突触。2周时,非对称性或对称性突触的数量没有观察到显著差异(图2a)。然而,在第8周,非对称性突触的数量显著减少(-16.1%)(图2b)。在ca1细胞体层50 mm的近端区域中,在8周时也观察到显著减少,但在2周时没有(图9b和9c)。因此,apkc对初始谷氨酸能突触形成不是必需的,但对其稳定性和维持是必需的。然后分析超微结构,发现psd宽度在2周时减小,突触间隙在2周时增加,并且这种变化的幅度在8周时增加(图2c)。这表明apkc可能对突触的稳定性很重要,与lgl1相反(图1h)。
[0113]
然后被问及lgl1、pkcι/λ和pkcξ同时缺失是否会导致lgl1条件性缺失实验中观察到的影响的减轻。在2周龄的lgl1:pkcι/λ:pkcξtcko动物中,未观察到非对称性突触数量的显著变化,这表明lgl1 cko中突触数量的增加可能部分是由于apkc活性的增加(图2d)。然而,观察到对称性突触的数量显著减少(-25.9%)。在8周龄的lgl1:pkcι/λ:pkcξtcko动物中,未观察到非对称性或对称性突触数量的显著变化(图2e)。在2周和8周时,在近端区域中未观察到非对称性突触密度的显著差异(图10a和10b)。因此,lgl1可能通过抑制apkc负调节谷氨酸能突触的数量,apkc是谷氨酸能突触稳定和维持所必需的。
[0114]
由于pcp蛋白调节突触形成,并且顶端-基底极性信号传导调节定位pcp信号传导组分,使用突触体分级分离测试lgl1是否可以调节pcp组分。在p7 slick-h系(诱导型cre)仅的60%锥体神经元中表达cre。因此,从lgl1 ko、杂合子和野生型培养神经元,并提取突触体组分。发现lgl1
/-和lgl1-/-的级分与野生型相比,在p2中vangl2蛋白水平降低(图2f),而粘附分子jam-c的水平没有受到影响(图2g)。这与vangl2在谷氨酸能突触形成中的抑制功能一致。
[0115]
lgl1控制谷氨酸能突触数量且是成年期中突触可塑性所需的由于lgl1在成年中枢神经系统(包括海马体)中高度表达,因此表征lgl1在成年脑中的作用。与出生后发育相比,在6周的早期成年期,啮齿类海马体中突触形成显著减缓(wang et al., 2007)。在slick-h动物出生后6周,通过注射他莫昔芬条件性敲除lgl1。然后使用电子显微镜评估10周龄动物的schaffer 侧支中非对称性和对称性突触的密度。ca1细胞体层150-200 μm远侧区域的非对称性突触密度在树突棘上再次增加了28.6%(图3a和3b)。在ca1细胞层50 μm的近端区域也观察到类似的增加(图3c和3d)。对称性突触再次不受影响(图3e和3f)。此外,在成年lgl1缺失中,突触超微结构发生改变,psd更长且更宽(图3g
和3h),以及突触间隙更小(图3i),这可能是突触的生化组成改变的结果。用p42切片进行膜片钳,以评估从p28开始lgl1缺失的动物的突触受体表达。在lgl1 cko小鼠的神经元中观察到ampa/nmda比例严重降低(图3j和3k),远远大于在p7/p8缺失lgl1后在p14处观察到的。这些结果表明,lgl1也是控制成年期中突触数量和质量所需的。
[0116]
在6周龄条件性lgl1缺失后,使用来自10周龄小鼠的海马切片测试突触可塑性。来自lgl1 cko动物的切片显示对θ脉冲刺激(tbs)响应的长期增强(ltp)诱导受损(图3l-3m)。此外,当刺激间隔100 ms时,来自lgl1 cko小鼠的切片显示配对脉冲易化受损(图3n),表明突触释放改变。
[0117]
lgl1cko小鼠表现出行为缺陷在史密斯-马吉利综合征(sms)中lgl1经常缺失,并且负责sms行为症状的基因尚未被很好地理解。因此,进行了大量的行为测试,以评估锥体神经元中lgl1的缺失是否可能导致行为缺陷。使用旷场测试评估自主活动(locomotor activity)和探索行为(图4a;gould et al., 2009)。lgl1 cko动物表现出自主活动增加,在10分钟的测试期间,比对照组小鼠多移动20%(图4b)。在趋触性(对旷场外部和中心区域的偏好)中没有观察到显著的变化(图4c)。在旷场测试期间,动物在自我梳理花费的时间上没有表现出差异(图11a),但在直立活动(一种探索性行为)上表现出显著增加(图11b)。
[0118]
为了评估lgl1条件性缺失后的认知功能,在新物体识别(nor)范例中测试了与slick-h杂交的lgl1 cko。nor测试评估动物是否能区分新物体和之前探索过的熟悉物体。通过从新的探索时间中减去与熟悉的物体交互时间,并将其标准化为总探索时间((新-熟悉)/(新 熟悉))来计算每只动物的偏好指数。在样品阶段,当两个物体都是新的时,动物没有表现出偏好,会随机探索这两个物体(图4d和4e)。当新物体在延迟2分钟后被引入时,对照动物表现出对新物体的强烈偏好,而lgl1 cko动物继续随机探索并保持接近零的偏好指数(图4f和4g)。
[0119]
脑超连接与自闭症谱系障碍(asd)有关(dominguez et al., 2013; keown et al., 2013; supekar et al., 2013)。三室社会交互任务中的社交能力已被广泛用于评估小鼠的社交行为(yang et al., 2011)。对与slick-h杂交的lgl1 cko小鼠进行测试,以评估其社交能力(图4h-4p)。在引入目标小鼠之前,动物对任何一方都没有偏好,随机探索该区域(图4h-4j)。当小鼠被引入旷场的一侧,而空的围栏被引入另一侧时,对照动物表现出与新小鼠交互的强烈偏好。lgl1 cko动物没有表现出偏好(图4k-4m),并且花了相似的时间探索新小鼠和新物体。有趣的是,在测试的最后阶段没有观察到统计学上的显著差异,其中受试小鼠在熟悉小鼠的和新目标小鼠之间进行选择(图4n-4p)。与对照组类似,lgl1 cko小鼠似乎仍然表现出对新小鼠的偏好。lgl1 cko小鼠通过在y型迷宫的交替和进入显示出正常的空间记忆(图11c和11d)和正常的视觉表现(图11e)。海马和杏仁核依赖的记忆形成在条件性恐惧任务中得以保留(图11f)。在lgl1条件性缺失后,窝破坏活动没有变化(图11g)。
[0120]
非典型pkc(apkc)缺失部分挽救lgl1 cko的行为缺陷在条件性apkc缺失中观察到突触表型,然后测试这些变化是否会导致行为缺陷。在旷场测试中,pkcι/λ和pkcξ dcko动物在自主活动或趋触性方面没有显著变化(图5a-5c)。还评估了认知功能,发现pkcι/λ:pkcξ dcko在新物体识别测试中受损(图5d和5e)。在社会交互测试中,在引入新小鼠之前,动物没有表现出偏好(图5f和5g)。同窝对照动物更喜
欢社会交互,花费更多时间在新小鼠而不是新物体,而pkcι/λ:pkcξ dcko花费相似的时间与新小鼠和新物体交互,保持接近零的偏好指数(图5h和5i)。apkc缺失不影响对社会新奇的偏好(图5j和5k)。
[0121]
然后测试自主活动,在同窝对照和lgl1:pkcι/λ:pkcξ tcko动物之间没有观察到差异(图5l-5m)。然而,在新物体识别测试中,lgl1、pkcι/λ和pkcξ的三重条件性缺失确实挽救了认知缺陷,在测试中lgl1:pkcι/λ:pkcξ tcko动物的表现与同窝对照相似且优于偶然性(单样本t检验,p=0.0262)(图5n-5q)。这些观察结果表明,lgl1、pkcι/λ和pkcξ的条件性缺失更正观察到的突触密度变化并保留认知功能。在社会交互测试(图5r-5w)中,tcko动物在社会交互阶段(图5t-5u)仍表现出障碍,但在社会新奇阶段(图5v-5w)没有。
[0122]
lgl1杂合子增加突触数量并表现出行为缺陷,表明在史密斯-马吉利综合征中的作用由于sms源于关键区域的杂合性缺失,在从所有神经元和非神经元细胞中移除lgl1的一个拷贝的小鼠模型(klezovitch et al., 2004)中评估lgl1种系杂合性缺失的影响。使用电子显微镜对8周龄对照和生殖系lgl1 ko系lgl1杂合子动物的海马切片进行成像。在这些动物中,来自ca1细胞体层150-200 μm的区域显示非对称性突触密度增加,与在条件性ko中缺失两个拷贝时观察到的情况相比,增加幅度较小但显著增加(22.6%;图6a和图12a所示的近端区域)。对称性突触没有受到影响。突触超微结构的分析显示更长且更宽的psd和更小的突触间隙(图6b)。
[0123]
在旷场试验中,在10分钟后lgl1
/-动物与对照组动物没有显示出显著差异(图12b-12c)。在旷场中60分钟的延长观察期间(图6c),lgl1
/-动物显示出自主活动总体增加12.0%。
[0124]
还测试了刻板重复行为,发现当在新笼子中给予棉花筑巢材料时,lgl1
/-动物表现出窝破坏增加(图6d),在测试期间多破坏59.4%的材料。由于在lgl1 cko中未观察到这种行为,这可能是皮质下区域中杂合子中lgl1缺失的结果,其不受与slick-h杂交的lgl1 cko动物中thy-1依赖性缺失的影响。
[0125]
延迟2分钟后,lgl1
/-动物对新物体识别的偏好得以保留(图12e-12h),表明由于lgl1的剩余拷贝降低对突触密度或突触功能的影响,这种认知任务可能部分得以保留。因此,进行一次另外外的新物体识别测试,这次在初始采样周期和识别测试之间有24小时的延迟。24小时后,对照组动物成功区别新的和熟悉的物体,而lgl1
/-动物则没有(图6e)。sms患者表现出轻度至中度认知障碍或发育迟缓,并且部分保留的对新物体的识别可能反映了小鼠模型中的轻度损伤。
[0126]
与lgl1 cko动物类似,lgl1
/-动物表现出缺乏社会交互,但对社会新奇性没有偏好(图6f和12i-12j),表明损失lgl1的一个拷贝——如sms中所发生的那样——足以导致asd样行为。在年龄依赖性体重增加方面lgl1
/
动物与同窝对照动物没有差异(图12k)。
[0127]
lgl1
/-中的社会交互缺陷可能是由过量的nmda电流引起的因为sms经常表现为癫痫发作或异常eeg但没有明显的癫痫发作(chen et al., 1996; greenberg et al., 1996),测试异常高的突触密度和nmda电流是否会降低响应gabaa阻断药物戊四氮(ptz)的癫痫发作阈值。在注射50 mg/kg ptz后,记录表明癫痫发作的活动的发生和潜伏期。所有测试的动物表现出至少一次跳跃或全身抽搐(图7a),而lgl1
/-动物在ptz施用后表现出较短的行为潜伏期(图7b)。lgl1
/-动物也表现出管状尾的发生率显著更高且潜伏期显著更短,表明持续的肌肉收缩,以及对强直阵挛癫痫发作的发生也也表现出类似的效果。
[0128]
较低的ampa/nmda比例和未受影响的ampa电流表明lgl1突变体可能有过量的nmda电流。因此,有人问,观察到的异常的nmdar依赖性信号传导是否可能是某些行为缺陷(尤其是社会互动)的基础。使用nmdar的药理学阻断剂,并在三室社会交互测试中测试动物。已显示亚麻醉量的氯胺酮可快速阻断nmdar依赖性信号传导。通过注射50 ml无菌生理盐水测试社会交互,观察到对照动物更喜欢社会交互,而lgl1
/-小鼠没有显示出偏好(图7c-7e)。注射30 mg/kg氯胺酮后,lgl1
/-小鼠显示出与对照动物的观察结果类似的对社会交互的明显偏好(图7c-7e)。习惯阶段和社会新奇阶段的偏好不受生理盐水或药物注射的影响(图13a-13d)。氯胺酮注射没有改变窝破坏活动(图13e)。此外,还测试0.3 mg/kg的mk-801(地佐环平)和20 mg/kg的美金刚,观察到lgl1
/-强烈偏好社会交互,和对照组小鼠一样(图7c-7e)。这些结果表明,lgl1缺失导致的过量nmda电流可能导致一些行为缺陷,抑制nmdar可能有助于缓解lgl1缺失的sms患者的一些神经精神症状。
[0129]
以下实施例旨在说明而非限制本发明。
[0130]
实施例1电子显微镜——lgl cko或lgl1
/-和同窝对照小鼠在室温下用氯胺酮/二甲苯嗪混合物腹腔麻醉,并用改良的karnovsky固定剂(2.5%戊二醛和2%pfa的0.15 m二甲氧基钠缓冲液,ph 7.4)灌注。在4℃下在同一溶液中固定过夜后,用振动切片机将整个脑冠状切割(200μm)。切片在冰上用1%四氧化锇固定1小时。在2%醋酸铀酰的蒸馏水中进行整体染色1-2小时后,切片在分级乙醇系列中脱水,然后在丙酮中处理两次,每次10分钟。切片浸润到durcupan树脂(sigma-aldrich)中,并在60℃下嵌入48小时。超薄切片来自切割ca1区域,并用醋酸铀酰和sato’s lead染色。图像采集使用配有eagle 4k hs数码相机的fei tecnai spirit g2 spirit biotwin透射电子显微镜(fei,hilsboro,or)。在6800倍放大下,从远侧辐射层(ca1锥体细胞体150-200 μm)和近侧辐射层(ca1锥体细胞体40-50 μm)中选择10-15个视野,并在每个视野中计数突触。显示多核糖体和/或肌动蛋白束的突触被确定位于树突轴上,而缺乏这些结构的突触被确定位于树突棘上。对于超微结构分析,来自远端辐射层区域的非对称性突触以18500倍放大成像。仅选择具有明确识别的psd和突触间隙的非对称性突触。使用image j分析bouton面积、间隙距离、储备池(reserve pool)囊泡、对接(docked)囊泡、psd宽度和psd长度。超微结构分析由不了解动物基因型的实验者进行。
[0131]
海马原代细胞培养——如先前所述进行海马神经元培养(meffert et al., 2003)。玻璃盖玻片在硝酸中洗涤一夜且彻底冲洗,并用100%乙醇洗涤,然后放入干燥箱中。在细胞培养前一天,盖玻片在室温下用80mg/ml聚d-赖氨酸(pdl)包被过夜。洗涤盖玻片,并在37℃下用40 mg/ml pdl和2.5mg/ml层粘连蛋白包被3小时。
[0132]
在l15培养基中取出e18.5幼崽的大脑,解剖海马体并将其放置在单独的试管中。用无mg/ca的无菌pbs(cellgro)洗涤海马体两次,并与胰蛋白酶/edta溶液(0.25%胰蛋白酶,2.2mm edta)在37
°
c下孵育15分钟。通过添加10% vol/vol热灭活马血清(life technologies)中和胰蛋白酶。将所得溶液沉淀并重悬于补充有2% b27(invitrogen)、5%胎牛血清(invitrogen)、青霉素/链霉素(cellgro)和谷氨酰胺(invitrogen)的neurobasal培
养液中,并洗涤两次。测定细胞密度,并以2.5x104个细胞/cm2的密度铺板。每3天更换一半的生长培养基。在体外第6天加入阿糖基呋喃胞苷(arabinofuranosyl cytidine)(ara-c,4μm),以防止胶质细胞增殖。培养物在37℃、5%二氧化碳气氛中生长14天。
[0133]
aav注射——用异氟醚麻醉动物,并用70%乙醇和betadine溶液准备手术区域。在整个手术过程中,将动物置于头栏(bar)中并保持在异氟醚麻醉下。在手术开始时,给动物皮下注射buprenex。注射靶向背侧海马的ca1区。使用的立体定向协调是(来自bregma):ap-2.0,ml /-1.4,dv-1.35和-1.25。动物每半球接受2次注射。在回到笼子之前允许动物恢复运动和自我梳理活动。术后,每天给动物注射buprenex两次,持续3天。使用的病毒是aav1.hsyn.hi.egfp-cre.wpre.sv40和aav1.hsyn.egfp.wpre.bgh(upenn vector core)。将病毒稀释至3-5 x 10
12
病毒颗粒/ml用于注射。每个部位注射300 nl病毒溶液。动物在行为实验开始前恢复了14天。
[0134]
行为分析——对于所有行为分析,除非另有说明,否则在光线昏暗的房间(《100 lux)的光照阶段进行测试,测试区域有间接照明。雄性和雌性动物都在行为测试中进行了行为测试,除了在6周时注射aav的动物组,其仅在雄性动物中进行。
[0135]
旷场试验——旷场试验在一个40 cm
×
40 cm的塑料亚克力箱(stoetling)中进行,箱壁为深色,带有非反射性的底板。空间提示被放置在场地的一面墙上。将老鼠放在场地中央,让它们自由探索10分钟。对小鼠的移动距离、在场地内部和外部区域的时间、自我梳理所花费的时间、直立活动和跳跃活动进行评分。外部区域被定义为距场地边缘5厘米以内的区域。
[0136]
60分钟的测试变型应用于lgl1
/-小鼠和同窝对照,测试区域的所有方面与10分钟测试相同。进行10分钟测试的动物不被用于60分钟测试,以避免之前暴露于测试装置的影响。
[0137]
新物体识别测试——在空装置中进行10分钟的探索后,立即按照所述(antunes and biala, 2012)进行新物体识别测试。动物被放置在一个包含两个新物体副本的场地中,并被允许自由探索。在样品阶段之后,将动物从场地移除,并用一个先前探索过的物体的副本和一个新物体替换这些物体,该物体在场地中的位置与样品阶段相同。允许动物自由探索5分钟。为了计算交互时间,在matlab中选择包含目标物体的roi。roi由不了解动物基因型或病毒治疗的实验者选择。
[0138]
社会交互测试——社会交互测试如前所述(yang et al., 2011)进行。社交装置(stoetling)有三个20 cm
×
40 cm的腔室和两个直径7 cm、高15 cm的目标外壳。在测试期间,装置被不透明的面板包围,以防止小鼠看到设备外面。
[0139]
将动物放置在三室社交装置的中心室中,室之间的门关闭10分钟以适应测试条件。移除门,允许小鼠探索整个场地10分钟。在测试的社会交互阶段,引入了来自单独笼子的非同窝的品系和年龄匹配的目标小鼠。对于社会新奇阶段,将来自社会交互阶段的目标小鼠保持在同一位置,并将新的目标小鼠引入对面的腔室。为了计算交互时间,在matlab中选择包含目标小鼠和新物体的roi。roi由不了解动物基因型或病毒治疗的实验者选择。
[0140]
使用matlab 2014b(patel et al., 2014)(mathworks)中的autotyping 15.04,对旷场测试期间的移动距离和趋触性、以及在新物体识别测试中与物体交互花费的时间、以及在社会交互测试中与目标小鼠和物体交互花费的时间进行评分。
[0141]
窝破坏——将小鼠从它们的笼子中移出并单独放入含有一块重量约为2.8g的棉筑巢材料的干净的新笼子中。将小鼠留在笼子中60分钟。在测试期结束时,小鼠被送回它们的笼子。将破坏的材料从未被扰乱的部分分离,并对其进行称重(li et al., 2006)。
[0142]
电生理学——对于p13
–
15的lgl1 cko和同窝对照,用异氟醚麻醉小鼠。将小鼠斩首,迅速移出其大脑并置于含有以下物质(单位:mm)的冰冷解剖缓冲液中:87 nacl、2.5 kcl、1.25 nah2po4、25 nahco3、20葡萄糖、75蔗糖、0.5 cacl2和7 mgcl2,ph 7.35。在振动切片机(letcha vt1200)上将横向海马切片切成300 μm厚的切片。让切片在35℃下恢复20分钟,然后在室温下在碳化的acsf中恢复1至6小时,其含有(单位:mm):124 nacl、2.5 kcl、24 nahco3、1.2 nah2po4、2 cacl2、2 mgcl2和12.5葡萄糖,5 hepes,ph 7.4。将单个切片转移至记录室,然后以2-3 ml/min的速率连续灌注含有1 μm ttx(tocris)和20 μm gabazine或20μm cnqx的acsf,分别用于mepsc或mipsc记录。通过红外干涉差显微镜观察ca1锥体细胞。
[0143]
对于mepsc,使用3-5 mω移液器进行全细胞记录,移液器中填充有内部溶液(单位:mm):145 ch3o3scs、5 nacl、10 hepes、5 egta、0.3 na2gtp和4 mgatp(ph被调整为7.3,以及渗透压保持在280-290 mosm)。对于mipsc记录,使用3-5 mω移液器进行全细胞记录,移液器中填充有内部溶液(单位:mm):135 cscl、4 mgcl2、0.1 egta、10 hepes、2 mgatp、0.3 nagtp、10 na2phosphocreatine(ph值被调整为7.3,以及渗透压保持在280-290 mosm)。
[0144]
细胞被电压钳制在-70 mv。5分钟后开始记录,以允许稳定已建立的全细胞结构。信号用5倍增益记录,在2 khz低通过滤,并在5 khz用pclamp 10软件(molecular devices)数字化(molecular devices axopatch 200b);使用clampfit(pclamp)进行分析。自动检测通过目视检查进行事后验证。值以平均值
±
sem表示。mann-whitney u统计检验用于比较与对照的变化。
[0145]
对于ltp诱导,从9-10周大的小鼠中获取横向海马切片。在移到acsf中的记录室之前,将切片回收至少1小时(单位:mm):nacl 119、kcl 2.5、nah2po
4 1、nahco
3 26.2、葡萄糖11、mgcl
2 1.3和cacl
2 2.5,用95%的o2和5%的co2鼓泡。以0.033 hz给予刺激以激发场电位,玻璃电极的电阻为1-2 mω,内溶液为acsf。通过逐渐增加刺激强度来记录输入输出。在30分钟稳定基线记录后,由hfs或tbs诱导ltp。hfs是100 hz的100个脉冲。tbs包含四列θ脉冲,间隔20秒,每列具有10次脉冲,间隔0.2秒。
[0146]
对于ampa/nmda比例测定,使用碎玻的璃移液器作为刺激电极。膜片钳吸管的电阻为3-5mω。内溶液(单位:mm):csmeso
4 115、cscl 20、na phosphocreatine 10、mgcl
2 2.5、na2atp 4
、na3gtp 0.3、egta 0.6、hepes 10和qx-314-cl 5。在ca1和ca3之间进行切割以避免癫痫事件。轻轻调整刺激电极的位置以引起单峰epsc。对于ampa电流,保持电位设置为-70 mv,对于nmda电流,保持电位更改为 40 mv。nmda电流被量化为刺激伪影(artifact)后峰值100 ms,其中没有ampa电流。
[0147]
生物化学分级分离——亚细胞分级分离如前所述(cohen et al., 1977)进行了修改。来自p14野生型小鼠的前脑在含有1 mm mgcl2、0.5m cacl2、1mm nahco3和蛋白酶抑制剂的0.32 m冰凉蔗糖缓冲液中用玻璃dounce进行16次冲程(stroke)均质化至10%(wt/vol),。匀浆在4℃以710 g旋转30分钟以沉淀出细胞核和大碎片。上清液在4℃以13800 g进一步离心10分钟以获得沉淀(p2)。p2沉淀在蔗糖缓冲液中重悬,并在ph 7.4 含有1.0 m和
1.4 m蔗糖的4 mm hepes缓冲液的不连续蔗糖梯度顶层分层。梯度在4℃以82500 g离心1 h。从1.0 m和1.4 m蔗糖之间的浑浊带中回收突触体,在1 mm nahco3(1:9 vol/vol)中重悬,并通过低渗透性休克用玻璃dounce进行3次冲程进行裂解。然后将裂解物与等体积的含1%triton-x的0.32 m蔗糖缓冲液孵育,在4℃振荡15分钟,并在82500 g旋转1小时。psd级分从产生的沉淀中获得的。使用甲醇/氯仿/h2o方法从上清液中沉淀突触膜级分(smf)(wessel and fl
ü
gge, 1984)。将psd级分和smf两者溶解在3% sds中进行蛋白质印迹分析。
[0148]
为了分离表面定位蛋白,从对照和lgl1 cko动物中分离急性切片。p21小鼠切片厚度为300 μm,p56小鼠切片厚度为400 μm。切片收集于含25 mm nahco3、1.25 mm nah2po4、2.5 mm kcl、7 mm mgso4、25 mm葡萄糖、0.5mm cacl2的碳化分离缓冲液中,使用前添加氯化胆碱(0.11 mm)、抗坏血酸钠(1.3 μm)和丙酮酸钠(2.4 μm)。将切片移至含有10 mm hepes、140 mm nacl、24 mm nahco3、3 mm kcl、1.25 mm nah2po4、1.25 mm mgcl2、2 mm cacl2的hepes缓冲acsf中,并恢复90分钟。表面生物素化如前所述进行了修改(kim and kovacs, 2011)。hepes缓冲acsf用于所有低钙孵育和洗涤。45分钟后停止与nhs-lc-sulfo-biotin(thermo)的孵育。在液氮中快速冷冻之前,将皮质区与海马和皮质下结构分离,并在-80℃下储存,直到蛋白质提取和下拉。向样品中添加含有10 mm tris、150 mm nacl、1% tritonx-100、0.2% sds和0.5%脱氧胆酸钠以及1mm edta的提取缓冲液,并在超声波处理之前,通过在4℃以12000 g离心15分钟将重膜和碎片沉淀。用bca法测定蛋白质浓度。在1 ml提取缓冲液中加入蛋白酶抑制剂混合物(sigma)制备神经亲和素(neuravidin)珠。向样品中添加1 mg蛋白质,并在4℃旋转3小时。对于总蛋白质样品,将200 ug蛋白质在提取缓冲液和sds上样染料中稀释,并在-20℃冷冻直至使用。edta浓度增加到50 mm以分离fmr1。
[0149]
蛋白质印迹——蛋白质样品在5%和8%丙烯酰胺凝胶上通过聚丙烯酰胺凝胶电泳分离,并湿转移到immobilon膜((millipore)上。使用5%脱脂奶粉(apex)的tbst进行封闭,并在封闭缓冲液中稀释一级抗体,在4℃孵育过夜。用tbst洗涤印记,并与hrp偶联的二级抗体在室温孵育2小时。使用west pico chemiluminescent substrate观察条带,并以30秒到2小时的间隔暴露于胶片上。
[0150]
抗体——一级抗体是小鼠抗lgl1、小鼠抗lgl1(novus)、山羊抗vangl2抗体(n-13, santa cruz, sc-46561)、山羊抗jam-c(r&d systems)、小鼠抗gapdh(abcam)、兔抗gfp(invitrogen)和鸡抗map2(abcam)。蛋白质印迹中使用的二级抗体是hrp偶联的驴抗小鼠、抗山羊和抗兔。对于免疫荧光,af568偶联的驴抗鼠(invitrogen)、af488偶联的驴抗鸡(jackson immunoresearch)、af488偶联的驴抗兔(invitrogen)、af647偶联的驴抗兔(invitrogen)。
[0151]
细胞填充和树突棘分析——如前所述,对小鼠进行灌注、切片并用alexafluor酰肼标记(dumitriu et al., 2011)。slick-h:lgl1
flox/flox
幼崽在p7-8腹腔注射他莫昔芬。在p14,用氯胺酮/二甲苯嗪混合物麻醉幼崽,然后在室温用4% pfa穿心灌注(transcardial perfusion),在4% pfa中固定1小时,振动切片机以100 μm切片,然后再次固定10分钟。通过用荧光染料填充细胞,将alexafluor酰肼555(invitrogen,200mm kcl中的10mm)离子电泳注入树突状细胞段。切片短暂固定15分钟,以保存荧光标签。使用gfp抗体(invitrogen)进行免疫标记以确认荧光填充的树突为yfp 。对位于辐射层中ca1锥体神经元胞体100-200 μm的ca1斜顶端树突的30-40 μm片段进行了分析。这些树突棘代表了schaffer 侧支ca1突触
quantal neurotransmitter release and dendritic spine density in ca1 pyramidal neurons. hippocampus 22, 1493-1500.chen, et al. (1997). homologous recombination of a flanking repeat gene cluster is a mechanism for a common contiguous gene deletion syndrome. nature genetics 17, 154
‐
163.chen, et al. (1996). the smith
‑‐
magenis syndrome [del(17)p11.2]: clinical review and molecular advances. mental retardation and developmental disabilities research reviews 2, 122
‐
129.cohen, et al. (1977). the structure of postsynaptic densities isolated from dog cerebral cortex: i. overall morphology and protein composition. journal of cell biology 74, 181
‐
203.collingridge, g., kehl, s., and mclennan, h. (1983). excitatory amino acids in synaptic transmission in the schaffer collateral
‑‐
commissural pathway of the rat hippocampus. journal of physiology 334, 33
‐
46.dominguez, et al. (2013). the imaginary part of coherency in autism: differences in cortical functional connectivity in preschool children. plos one 8, 1
‐
13.dumitriu, d., rodriguez, a., and morrison, j. (2011). high
‐
throughput, detailed, cell
‐
specific neuroanatomy of dendritic spines using microinjection and confocal microscopy. nature protocols 6, 1391
‐
1411.durand, et al. (2007). mutations in the gene encoding the synaptic scaffolding protein shank3 are associated with autism spectrum disorders. nature genetics 39, 25-27.dykens, e., finucane, b., and gayley, c. (1997). brief report: cognitive and behavioral profiles in persons with smith
‐
magenis syndrome. journal of autism and developmental disorders 27.dykens, e., and smith, a.c.m. (1998). distinctiveness and correlates of maladaptive behaviour in children and adolescents with smith
‐
magenis syndrome. journal of intellectual disability research 42, 481
‐
489.ebnet, et al. (2003). the junctional adhesion molecule (jam) family members jam
‐
2 and jam
‐
3 associate with the cell polarity protein par
‐
3: a possible role for jams in endothelial cell polarity. journal of cell science 116, 3879
‐
3891.edelman, et al. (2007). gender, genotype, and phenotype differences in smith
–
magenis syndrome: a meta
‐
analysis of 105 cases. clinical genetics 71.ehrlich, i., and malinow, r. (2004). postsynaptic density 95 controls ampa receptor incorporation during long
‐
term potentiation and experience
‐
driven synaptic plasticity. j neurosci 24, 916
‐
927.
georgiou, et al. (2008). cdc42, par6, and apkc regulate arp2/3
‐ꢀ
mediated endocytosis to control local adherens junction stability. current biology 18, 1631
–
1638.girirajan, et al. (2006). genotype
‐
phenotype correlation in smith
‐
magenis syndrome: evidence that multiple genes in 17p11.2 contribute to the clinical spectrum. genetics in medicine 8, 417
‐
427.gould, t., dao, d., and kovacsics, c. (2009). the open field test. mood and anxiety related phenotypes in mice 42, 1
‐
20.greenberg, et al. (1996). multi-disciplinary clinical study of smith-magenis syndrome (deletion 17p11.2). am j med genet 62, 247-254.gropman, et al. (2007). new developments in smith-magenis syndrome (del 17p11.2). current opinion in neurology 20, 125-134.harris, et al. (1992). three-dimensional structure of dendritic spines and synapses in rat hippocampus (ca1) at postnatal day 15 and adult ages: implications for the maturation of synaptic physiology and long-term potentiation. journal of neuroscience 12, 2685-2705.heimer-mcginn, v., and young, p. (2011). efficient inducible pan-neuronal cre-mediated recombination in slick-h transgenic mice. genesis 49, 942-949.huang, et al. (2016). molecular and neural functions of rai1, the causal gene for smith-magenis syndrome. neuron 92, 1-15.hung, et al. (2008). smaller dendritic spines, weaker synaptic transmission, but enhanced spatial learning in mice lacking shank1. the journal of neuroscience 28, 1697
–
1708.irifune, m., shimizu, t., and nomoto, m. (1991). ketamine-induced hyperlocomotion associated with alteration of presynaptic components of dopamine neurons in nucleus accumbens of mice. pharmacology biochemistry and behavior 40, 399-407.jeyifous, et al. (2009). sap97 and cask mediate sorting of nmda receptors through a previously unknown secretory pathway. nature neuroscience 12, 1011-1019.karner, c., wharton, k.a., and carroll, t.j. (2006). apical-basal polarity, wnt signaling and vertebrate organogenesis. semin cell dev biol 17, 214-222.keown, et al. (2013). local functional overconnectivity in posterior brain regions is associated with symptom severity in autism spectrum disorders. cell reports 5, 567
–
572.kim, d., and kovacs, d. (2011). surface trafficking of sodium channels in cells and in hippocampal slices. methods mol biol 793, 351-361.
8, 1-16.pe
ç
a, et al. (2011). shank3 mutant mice display autistic like behaviours and striatal dysfunction. nature 472, 437-442.phelan, et al. (2001). 22q13 deletion syndrome. am j med genet 101, 91-99.potocki, et al. (2003). variability in clinical phenotype despite common chromosomal deletion in smith-magenis syndrome[del(17)(p11.2p11.2)]. genet med 5, 430-434.prybylowski, et al. (2005). the synaptic localization of nr2b-containing nmda receptors is controlled by interactions with pdz proteins and ap-2. pnas 109, 7499-7504.sans, et al. (2000). a developmental change in nmda receptor-associated proteins at hippocampal synapses. j neurosci 20, 1260-1271.sato, et al. (2012). shank1 deletions in males with autism spectrum disorder. the american journal of human genetics 90, 879
–
887.schmeisser, et al. (2012). autistic-like behaviours and hyperactivity in mice lacking prosap1/shank2. nature 486, 256
–
260.sheng, m., and kim, e. (2011). the postsynaptic organization of synapses. cold spring harbor perspectives in biology 3.smith ac, dykens e, and f, g. (1998). behavioral phenotype of smith-magenis syndrome (del 17p11.2). am j med genet 81, 179-185.smith, et al. (1986). interstitial deletion of (17)(p11.2p11.2) in nine patients. american journal of medical genetics 24, 393
–
414.soorya, et al. (2013). prospective investigation of autism and genotype-phenotype correlations in 22q13 deletion syndrome and shank3 deficiency. molec autism 4, doi: 10.1186/2040
‐
2392-1184-1118.sudhof, t.c. (2012). the presynaptic active zone. neuron 75, 11-25.supekar, et al. (2013). brain hyperconnectivity in children with autism and its links to social deficits. cell reports 5, 738-747.tang, et al. (2014). loss of mtor-dependent macroautophagy causes autistic-like synaptic pruning deficits. neuron 83, 1131-1143.tao, et al. (2003). impaired nmda receptor
‑‐
mediated postsynaptic function and blunted nmda receptor-dependent persistent pain in mice lacking postsynaptic density-93 protein. j neurosci 23, 6703-6712.thakar, et al. (2017). evidence for opposing roles of celsr3 and vangl2 in glutamatergic synapse formation. pnas 114, e610-e618.vlangos, et al. (2003). refinement of the smith
–
magenis syndrome critical region to 950 kb and assessment of 17p11.2 deletions. are all deletions created equally molecular genetics and metabolism 79, 134-141.
vogel
‐
ciernia, et al. (2013). the neuron-specific chromatin regulatory subunit baf53b is necessary for synaptic plasticity and memory. nat neurosci 16, 552-561.wang, et al. (2007). changes in hippocampal synapses and learning-memory abilities in age-increasing rats and effects of tetrahydroxystilbene glucoside in aged rats. neuroscience 149, 739-746.watkins, j.c., and evans, r.h. (1981). excitatory amino acid transmitters. annu rev pharmacol toxicol 21, 165-204.wessel, d., and fl
ü
gge, u. (1984). a method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. anal biochem 138, 141-143.xiao, et al, (1998). homer regulates the association of group 1 metabotropic receptors with multivalent complexes of homer-related, synaptic proteins. neuron 21, 707-716.yamanaka, et al. (2006). lgl mediates apical domain disassembly by suppressing the par
‐
3-apkc-par-6 complex to orient apical membrane polarity. journal of cell science 119, 2107-2118.yamanaka, et al. (2003). mammalian lgl forms a protein complex with par-6 and apkc independently of par-3 to regulate epithelial cell polarity. current biology 13, 734
–
743.yang, m., silverman, j., and crawley, j. (2011). automated three-chambered social approach task for mice. curr protoc neurosci 8, unit 8.26.young, et al. (2008). single-neuron labeling with inducible cre-mediated knockout in transgenic mice. nature neurscience 11, 721
–
728.zarnescu, et al. (2005). fragile x protein functions with lgl and the par complex in flies and mice. dev cell 8, 43-52.zhu, et al. (2016). mechanistic basis of maguk-organized complexes in synaptic development and signaling. nat rev neurosci 17, 209-223.尽管已经参考上述实施例描述了本发明,但是应当理解,本发明的精神和范围内包含修改和变化。因此,本发明仅受以下权利要求的限制。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。