1.本发明涉及大黄素应用技术领域,尤其涉及大黄素在减轻急性低氧诱导的鱼类细胞凋亡中的应用及用于减轻急性低氧诱导的鱼类细胞凋亡的饲料。
背景技术:
2.溶解氧(do)是水产鱼类生存的关键环境变量之一,很容易受到自然和人为环境变化的影响,如高温、水体富营养化、工业废水排放和高密度水产养殖。在水体中,水质不达标或恶化的最重要表现之一是溶解氧水平的变化,因为溶解氧水平低通常会导致大量水生动物死亡。环境低氧应激影响水产动物的行为、生长、繁殖和生存,这对集约化水产养殖的长期发展具有严重的负面影响。
3.低氧应激是水产养殖池塘鱼类死亡的最常见原因之一。在细胞存活和代谢中,自噬和凋亡受同一细胞中相同上游信号通路的调控,自噬发生在凋亡之前。amp活化蛋白激酶(ampk)参与雷帕霉素靶蛋白(mtor)介导的自噬信号转导,形成更复杂的信号级联。作为一种保守的丝氨酸/苏氨酸蛋白激酶,mtor是调节细胞生长、存活和自噬的上游途径的重要节点。目前已经研究表明,低氧会诱导下丘脑内质网应激、自噬损伤和凋亡,从而损害下丘脑的正常生理功能。目前,鱼类急性低氧诱导内质网应激下ampk/mtor信号通路与自噬和凋亡之间的关系尚不明确,有待进一步研究。
4.大黄素是一种具有抗氧化、免疫调节和抗肿瘤功能的天然蒽醌衍生物。目前已有研究表明,大黄素emo参与诱导细胞生长抑制和凋亡。目前,关于大黄素emo在水产养殖中鱼类抗低氧胁迫的研究很少。
5.有鉴于此,有必要设计一种大黄素在减轻急性低氧诱导的鱼类细胞凋亡中的应用及用于减轻急性低氧诱导的鱼类细胞凋亡的饲料,以解决上述问题。
技术实现要素:
6.本发明的目的在于提供大黄素在减轻急性低氧诱导的鱼类细胞凋亡中的应用及用于减轻急性低氧诱导的鱼类细胞凋亡的饲料。
7.为实现上述发明目的,本发明提供了大黄素在减轻急性低氧诱导的鱼类细胞凋亡中的应用。
8.作为本发明的进一步改进,大黄素通过介导鱼类的ampk/mtor信号通路,增强鱼类细胞自噬,从而抑制鱼类细胞凋亡,减轻急性低氧条件对鱼类的损伤。
9.作为本发明的进一步改进,鱼类在急性低氧条件下,ampk的磷酸化水平显著增强,mtor蛋白的磷酸化水平受到显著抑制。
10.作为本发明的进一步改进,对急性低氧诱导的鱼类进行大黄素应用后,大黄素会引起鱼类ampk蛋白的磷酸化水平进一步显著增强,mtor蛋白的磷酸化水平进一步受到抑制。
11.作为本发明的进一步改进,大黄素显著降低急性低氧诱导的鱼类中低氧诱导因子
(hif-1α)蛋白的水平,通过下调低氧诱导因子(hif-1α)的蛋白水平来降低低氧信号传导。
12.作为本发明的进一步改进,鱼类在急性低氧条件下,自噬相关蛋白水平显著升高;对急性低氧诱导的鱼类进行大黄素应用后,自噬相关蛋白水平进一步升高,增强鱼类细胞自噬水平。
13.作为本发明的进一步改进,对急性低氧诱导的鱼类进行大黄素应用后,凋亡相关基因的表达水平显著降低,抗凋亡相关基因的表达水平显著升高。
14.作为本发明的进一步改进,所述鱼类为水产经济鱼类。
15.作为本发明的进一步改进,大黄素在减轻急性低氧诱导的鱼类细胞凋亡中的应用中的应用方式为作为饲料添加剂添加到饲料中。
16.作为本发明的进一步改进,所述大黄素当作饲料添加剂在减轻急性低氧诱导的鱼类细胞凋亡中的应用中的用量为600mg/kg
17.为实现上述发明目的,本发明还提供了一种用于减轻急性低氧诱导的鱼类细胞凋亡的饲料。所述饲料包括基础鱼饲料和饲料添加剂大黄素;所述饲料通过其饲料添加剂大黄素来介导鱼类的ampk/mtor信号通路,增强鱼类细胞自噬,减轻急性低氧诱导的鱼类细胞凋亡。
18.作为本发明的进一步改进,所述大黄素与所述基础鱼饲料的添加比例为(500~700)mg:1kg。
19.作为本发明的进一步改进,所述大黄素与所述基础鱼饲料的添加比例为600mg:1kg。
20.作为本发明的进一步改进,所述鱼类为鲤鱼、鲫鱼、团头鲂、草鱼等水产淡水经济鱼类中的一种。
21.本发明的有益效果是:
22.1、本发明提供的大黄素在减轻急性低氧诱导的鱼类细胞凋亡中的应用中,将大黄素应用于水产养殖中鱼类抗低氧胁迫环境中,大黄素emo通过激活ampk/mtor信号通路进一步增强自噬,从而减轻细胞凋亡。大黄素诱导的自噬增强可能是鱼类机体应对急性低氧应激的一种防御策略,增加自噬可以通过自噬的方式来修复细胞,从而减少凋亡,用于有效保护鱼类机体免受低氧诱导的氧化应激和不可恢复的凋亡。其作用机理为:大黄素emo显著增加急性低氧后p-ampk水平,并且显著抑制p-mtor水平,逆转细胞凋亡的诱导从而修复细胞,由此,减轻急性低氧对鱼类的损伤。
23.2、本发明提供的用于减轻急性低氧诱导的鱼类细胞凋亡的饲料,通过其饲料添加剂大黄素,来介导鱼类的ampk/mtor信号通路,增强鱼类细胞自噬,减轻急性低氧诱导的鱼类细胞凋亡,具备巨大的应用潜力。
附图说明
24.图1为本发明提供的在常氧和急性低氧应激两种环境下,对比例及实施例1的对照组和emo实验组肝脏中低氧诱导因子(hif-1α)的蛋白质水平图。
25.图2为本发明提供的在常氧和急性低氧应激两种环境下,对比例及实施例1的对照组和emo实验组鱼类肝脏grp78的蛋白质水平。
26.图3为本发明提供的在常氧和急性低氧应激两种环境下,对比例及实施例1的对照
组和emo实验组鱼类肝脏中内质网应激相关基因的mrna水平。
27.图4为本发明提供的在常氧和急性低氧应激两种环境下,对比例及实施例1的对照组和emo实验组鱼类下丘脑中内质网应激相关基因的mrna水平。
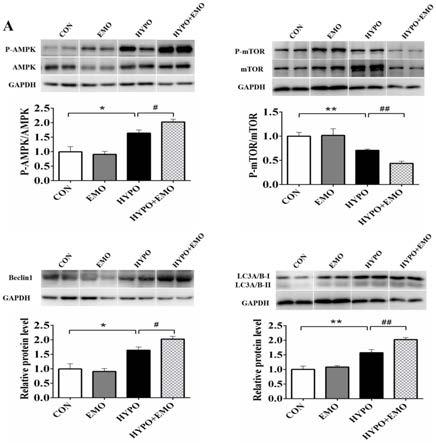
28.图5为本发明提供的在常氧和急性低氧应激两种环境下,对比例及实施例1的对照组和emo实验组鱼类肝脏中ampk、mtor、beclin1和lc3a/b-ii的蛋白质水平。
29.图6为本发明提供的在常氧和急性低氧应激两种环境下,对比例及实施例1的对照组和emo实验组鱼类肝脏中自噬相关关键基因的mrna水平。
30.图7为本发明提供的在常氧和急性低氧应激两种环境下,对比例1及实施例1的对照组和emo实验组鱼类下丘脑中自噬相关关键基因的mrna水平。
31.图8为本发明提供的在常氧和急性低氧应激两种环境下,对比例及实施例1的对照组和emo实验组鱼类肝脏中的caspase 3和caspase 9活性。
32.图9为本发明提供的在常氧和急性低氧应激两种环境下,对比例及实施例1的对照组和emo实验组鱼类肝脏中凋亡相关关键基因的mrna水平。
33.图10为本发明提供的在常氧和急性低氧应激两种环境下,对比例及实施例1的对照组和emo实验组鱼类下丘脑中凋亡相关关键基因的mrna水平。
34.图11为本发明提供的在常氧和急性低氧应激两种环境下,对比例及实施例1的对照组和emo实验组鱼类下丘脑dapi和tunel双重染色及图像定量结果。
具体实施方式
35.为了使本发明的目的、技术方案和优点更加清楚,下面结合附图和具体实施例对本发明进行详细描述。
36.在此,还需要说明的是,为了避免因不必要的细节而模糊了本发明,在附图中仅仅示出了与本发明的方案密切相关的结构和/或处理步骤,而省略了与本发明关系不大的其他细节。
37.另外,还需要说明的是,术语“包括”、“包含”或者其任何其他变体意在涵盖非排他性的包含,从而使得包括一系列要素的过程、方法、物品或者设备不仅包括那些要素,而且还包括没有明确列出的其他要素,或者是还包括为这种过程、方法、物品或者设备所固有的要素。
38.本发明提供了大黄素在减轻急性低氧诱导的鱼类细胞凋亡中的应用。
39.优选的,大黄素通过介导鱼类的ampk/mtor信号通路,增强鱼类细胞自噬,从而抑制鱼类细胞凋亡,减轻急性低氧条件对鱼类的损伤。
40.优选的,鱼类在急性低氧条件下,ampk的磷酸化水平显著增强,mtor蛋白的磷酸化水平显著受到抑制。
41.优选的,对鱼类进行大黄素应用后,急性低氧条件下,大黄素会引起鱼类ampk蛋白的磷酸化水平进一步显著增强,mtor蛋白的磷酸化水平进一步受到抑制。
42.优选的,大黄素显著降低急性低氧诱导的鱼类中低氧诱导因子(hif-1α)蛋白的水平,通过下调低氧诱导因子(hif-1α)的蛋白水平来降低低氧信号传导。
43.优选的,鱼类在急性低氧条件下,自噬相关蛋白水平显著升高;对急性低氧诱导的鱼类进行大黄素应用后,自噬相关蛋白水平进一步升高,由此增强鱼类细胞自噬水平。
44.优选的,对急性低氧诱导的鱼类进行大黄素应用后,凋亡相关基因的表达水平显著降低,抗凋亡相关基因的表达水平显著升高。
45.优选的,所述鱼类为水产淡水经济鱼类。
46.实施例及对比例
47.一、实验过程
48.1.饲料制备
49.实验组饲料:基础鱼饲料中添加大黄素(大黄素与基础鱼饲料添加比例为600mg:1kg)。
50.对照组饲料:基础鱼饲料,具体配方及化学成分见表1。
51.将饲料成分充分混合造粒处理,颗粒在50℃的烘箱中干燥,并在4℃下储存。
52.表1为实验组饲料和对照组饲料的配方及化学成分
[0053][0054]
2.实验动物及饲养试验
[0055]
本发明实施例选用异育银鲫为实验鱼类。
[0056]
实验前,所有鱼类在室内循环水养殖系统中适应对照饲料两周。然后,将规格一致的健康鱼(初始体重:6.66
±
0.01g)随机分配到室内玻璃缸中(每缸30尾,水量:150l,67
×
52
×
46cm),分为实验组和对照组,并每组设置三个平行处理例。
[0057]
每天在8:30、13:30和18:30分别采用实验组饲料和对照组饲料投喂,养殖实验持续8周。水温保持在29~31℃;ph值为7.0-7.4;溶解氧为6.0-7.5mg l-1
,总氨氮《0.1mg l-1
。光照强度为2.34-2.89μmols-1
m-2
,光照时间为12小时,黑暗时间为12小时。
[0058]
3.低氧应激
[0059]
在养殖实验结束时,使用低氧工作站(ruskinn invivo2400)设置低氧浓度值(2%)。从对照组和大黄素喂养的实验组中选择24尾异育银鲫,将它们放入12个透明塑料缸中(水体积:700ml,16
×9×
11cm)。在放置低氧工作站之前,测定水体中的溶氧(6.44
±
0.09mg/l)。随后,将两组塑料缸依次放入低氧工作站,在低氧条件下(2%o2)放置3h,再次使用ldo101探头测量水体中的溶氧(0.06
±
0.01mg/l)。同时,另外24尾异育银鲫放置在常氧条件下。
[0060]
因此,在常氧和急性低氧应激两种条件设置下,实验鱼被分为四组:对比例1的con组(常氧对比例)、对比例2的emo组(常氧 大黄素对比例)、对比例3的hypo(急性低氧对比例)组和实施例1的hypo emo组(急性低氧 大黄素实施例)。
[0061]
4.样本采集
[0062]
使用0.2%肝素钠溶液浸润过的注射器从六尾鱼的尾静脉采集血液。通过在4℃下以3000g离心15min获得血浆,并将血浆快速转移至-80℃保存有待进一步分析。肝脏和下丘脑放在液氮中速冻后移至-80℃保存,用于随后的酶活和蛋白基因表达分析。其余6尾鱼取下丘脑,在4%多聚甲醛中进行固定用于后续组织学检测。
[0063]
5.生化分析
[0064]
饲料和鱼样的化学成分(粗蛋白、粗脂肪、水分和灰分)通过aoac方法确定。粗蛋白含量通过凯氏定氮分析法测定。利用索氏抽提仪测定粗脂肪含量。
[0065]
精确称量肝组织,并添加0.9%氯化钠缓冲液以制备10%(w/v)匀浆。离心后,收集上清液进行酶分析,检测血浆应激生物标记物和抗氧化反应的生化指标。
[0066]
6.实时定量聚合酶链式反应(qrt-pcr)
[0067]
对照组低氧应激后,从肝脏和下丘脑中提取总rna。使用延伸因子1α(ef-1α)作为管家基因,qrt-pcr用于检测相关指标,包括参与抗氧化反应、内质网应激、自噬和凋亡的关键基因。
[0068]
7.标记法
[0069]
为了检测低氧应激3h是否诱导下丘脑细胞凋亡,新鲜下丘脑在4%多聚甲醛溶液中固定24h以上,脱水后石蜡切片包埋。用末端脱氧核苷酸转移酶介导的dutp缺口末端标记法(tunel)检测低氧(2%o2)和常氧条件下下丘脑切片中的凋亡细胞。
[0070]
8.统计分析
[0071]
采用spss 19.0软件对con组、emo组、hypo组和hypo emo组进行单因素方差分析比较,p《0.05为差异显著,p《0.01为差异极显著。所有结果以平均值
±
标准误表示。
[0072]
二、实验结果分析
[0073]
1.鱼类生长性能和体成分
[0074]
表2显示了鱼饲料中添加大黄素emo对鱼类生长、饲料利用和体成分的影响。emo处理组的摄食率(fr)显著低于对照组,饲料效率(fe)显著高于对照组,emo处理对鱼的特定生长率(sgr)没有显著影响。在鱼体组成中,粗蛋白质、粗脂肪、灰分和水分含量不受emo饲料的影响。
[0075]
表2为con组和emo组的生长性能和体成分
[0076][0077]
2.低氧诱导因子hif-1α的表达
[0078]
如图1所示,对鱼进行大黄素应用后再进行急性低氧处理,实验鱼肝脏中低氧诱导因子hif-1α的蛋白表达与常氧条件下的对照组相比,低氧处理显著诱导了异育银鲫肝脏中hif-1α的蛋白水平,并激活了低氧信号,而emo处理显著降低了低氧诱导的hif-1α的蛋白水平。
[0079]
注:图1中,在常氧和急性低氧应激下,对照组和emo组肝脏中低氧诱导因子(hif-1α)的蛋白水平。*p《0.05和**p《0.01(con组和hypo组),#p《0.05和##p《0.01(hypo组与hypo emo组)。数据表示为平均值
±
标准误(n=6)。
[0080]
3.内质网应激相关基因的mrna水平
[0081]
为了评估emo是否能减轻急性低氧引起的内质网应激,检测了肝脏中grp78蛋白的水平。结果如图2所示,急性低氧处理显著提高grp78蛋白水平,而emo处理抑制了grp78蛋白水平。grp78蛋白水平与基因表达结果一致,进一步证实emo可以通过抑制grp78蛋白的表达减轻急性低氧引起的内质网应激。
[0082]
急性低氧对内质网应激相关基因表达水平的影响的结果如图3所示。急性低氧处理后,异育银鲫肝脏内质网应激相关基因ire1、bip、chop、perk、atf6和atf4的mrna水平显著升高,但对xbp1或eif2a基因的mrna水平没有显著影响。emo会显著抑制ire1、atf6、atf4、bip和chop的mrna水平,但perk、xbp1和eif2a的mrna水平未受到显著影响。
[0083]
如图4所示,急性低氧会诱导激活下丘脑产生内质网应激。急性低氧显著增加内质网应激相关基因如ire1、perk、bip、chop和xbp1的mrna水平,但对atf6、eif2a或atf4的mrna水平没有显著影响。在急性低氧加emo治疗中,ire1、chop、bip和perk基因的mrna水平对atf6、xbp1、eif2a或atf4基因的mrna水平没有显著影响。
[0084]
注:图2至图4所示为在常氧和急性低氧应激下,对照组和emo组肝脏中grp78的蛋白质水平以及肝脏和下丘脑中内质网应激相关基因的mrna水平中,*p《0.05和**p《0.01(con组和hypo组),#p《0.05和##p《0.01(hypo组与hypo emo组)。数据表示为平均值
±
标准误(n=6)。ire1:肌醇需求酶1;perk:真核翻译起始因子2-α激酶3;atf6:激活转录因子6;bip:结合免疫球蛋白;xbp1:x盒结合蛋白1;ei2fa:真核翻译起始因子2a;atf4:激活转录因
子4;chop:dna损伤诱导转录因子3;grp78:葡萄糖调节蛋白78。
[0085]
4.ampk/mtor信号通路与自噬相关基因的表达
[0086]
肝脏ampk/mtor信号通路中关键蛋白的表达的结果如图5所示。在急性低氧条件下,ampk的磷酸化水平显著增强,mtor蛋白的磷酸化水平受到显著抑制。emo应用后低氧条件下ampk的磷酸化水平会进一步显著增强,mtor的磷酸化水平进一步受到抑制。
[0087]
此外,还检测了肝脏中自噬相关蛋白beclin1和lc3a/b-ii的表达水平。结果表明,低氧处理后,beclin1和lc3a/b-ii蛋白水平显著增强。emo应用后低氧条件下beclin1和lc3a/b-ii蛋白水平进一步上调。
[0088]
肝脏中自噬相关基因的mrna水平如图6所示。急性低氧处理后,beclin1、atg5和lc3b的mrna水平显著上调。p62的mrna水平显著降低。然而,emo应用后急性低氧条件下,lc3b、atf5和p62的mrna水平显著升高,而beclin1的mrna水平降低。急性低氧处理和急性低氧加emo处理对atg12基因的mrna水平没有显著影响。
[0089]
下丘脑自噬相关基因的表达情况如图7所示。急性低氧处理后,beclin1、lc3b和atg5的基因mrna水平显著升高。emo应用后会进一步上调beclin1、atg5和lc3b的mrna水平增强。急性低氧处理和急性低氧加emo处理对atg12和p62基因的mrna水平没有显著影响。
[0090]
注:图5至图7中,*p《0.05和**p《0.01(con组和hypo组)#p《0.05和##p《0.01(hypo组与hypo emo组)。数据表示为平均值
±
标准误(n=6)。单因素方差分析用于统计分析(p《0.05)。ampk:amp活化蛋白激酶;mtor:雷帕霉素靶蛋白;beclin1:beclin-1;lc3b:微管相关蛋白1a/1b轻链3b;atg5:自噬蛋白5;atg12:自噬蛋白12;p62:p62。
[0091]
5.凋亡相关基因的表达
[0092]
鱼类肝脏中半胱天冬酶-3和半胱天冬酶-9的活性如图8所示。急性低氧显著激活了鱼类半胱天冬酶-3和半胱天冬酶-9的活性。而emo显著降低了caspase-3和caspase-9的活性。鱼类肝脏中凋亡相关基因的表达水平如图9所示,急性低氧上调了bax、casp9和casp3、ero1α的mrna水平,而抗凋亡基因bcl2的mrna水平显著降低。emo应用后急性低氧条件下bax、casp3、ero1α和casp9的mrna水平降低,bcl2的mrna水平升高。
[0093]
下丘脑的凋亡基因表达水平如图10所示,在急性低氧处理下,下丘脑bax、ero1α和casp9基因的mrna水平显著升高,抗凋亡基因bcl2的mrna水平显著降低。emo显著抑制bax mrna水平,上调bcl2 mrna水平。casp9和ero1α的mrna水平没有受到显著影响。急性低氧处理和急性低氧加emo处理对casp3基因的mrna水平没有显著影响。
[0094]
注:图8至图10中,*p《0.05和**p《0.01(con组和hypo组)#p《0.05和##p《0.01(hypo组与hypo emo组)。数据表示为平均值
±
标准误(n=6)。单因素方差分析用于统计分析(p《0.05)。bcl2:凋亡调节因子bcl-2;bax:bcl2相关x蛋白,凋亡调节因子;casp3:caspase3;casp9:caspase9;ero1α:内质网氧化还原酶1α。
[0095]
综上,大黄素emo显著增加急性低氧后p-ampk水平,抑制p-mtor,逆转内质网应激和细胞凋亡的诱导。大黄素对减轻急性低氧诱导的鱼类细胞凋亡中的保护作用是通过激活ampk/mtor信号通路通过自噬进一步介导的。同时,emo通过下调低氧诱导因子(hif-1α)的蛋白水平降低低氧信号传导。这也保护了肝脏免受急性低氧诱导的氧化损伤。此外,emo通过介导ampk/mtor信号通路增强鱼类细胞自噬并抑制细胞凋亡。由此表明,emo能够通过ampk/mtor途径减轻急性低氧对鲫鱼的损伤。
[0096]
6.下丘脑的tunel分析
[0097]
下丘脑dapi和tunel双荧光染色法对tunel分析结果如图11所示,低氧组的凋亡信号明显高于对照组,emo可显著降低低氧刺激后下丘脑的凋亡率。
[0098]
图11中,在荧光显微镜下,凋亡细胞呈绿色荧光,正常细胞核呈蓝色*p《0.05和**p《0.01(con组和hypo组)#p《0.05和##p《0.01(hypo组与hypo emo组)。数据表示为平均值
±
标准误(n=6)。单因素方差分析用于统计分析(p《0.05)。显微照片放大率为200
×
,比例尺为100μm。
[0099]
需要说明的是,本发明中,大黄素的添加量还可以根据实际情况进行调整,并不限于上述实施例中的添加比例,均能够通过介导ampk/mtor途径减轻急性低氧对鱼类的损伤。
[0100]
本领域技术人员应当理解,本发明中,还可以采用大黄素应用于减轻其他种类淡水经济鱼类,例如:团头鲂、草鱼和鲤鱼等急性低氧诱导下的鱼类细胞凋亡。所述基础鱼饲料也可以根据不同种类的鱼类或者不同生长阶段的鱼类实际情况进行适应性的调整和改变,均不会影响饲料添加剂大黄素在减轻急性低氧诱导的鱼类细胞凋亡中的应用中所起到的作用。
[0101]
综上所述,本发明提供了大黄素在减轻急性低氧诱导的鱼类细胞凋亡中的应用。大黄素通过介导鱼类的ampk/mtor信号通路,增强鱼类细胞自噬,并抑制鱼类细胞凋亡,减轻急性低氧条件对鱼类的损伤。本发明还提供了一种用于减轻急性低氧诱导的鱼类细胞凋亡的饲料,其包括基础鱼饲料和用于减轻急性低氧诱导的鱼类细胞凋亡的饲料添加剂大黄素;饲料添加剂大黄素通过介导ampk/mtor途径减轻急性低氧对鱼类的损伤。所述鱼类为水产经济鱼类。
[0102]
以上实施例仅用以说明本发明的技术方案而非限制,尽管参照较佳实施例对本发明进行了详细说明,本领域的普通技术人员应当理解,可以对本发明的技术方案进行修改或者等同替换,而不脱离本发明技术方案的精神和范围。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。