梭菌纲聚生体组合物以及治疗肥胖症、代谢综合征和肠易激疾病的方法
1.相关申请的交叉引用
2.本技术要求2019年7月17日提交的美国临时申请号62/875,194的提交日的权益。这一在先提交的申请的内容特此以引用的方式整体并入。
3.对序列表的引用
4.以2020年7月16日创建的名为“21101_0401p1_sl.txt”并且大小为24,576字节的文本文件在本文提交的序列表根据37c.f.r.
§
1.52(e)(5)特此以引用的方式并入。
背景技术:
5.微生物群影响宿主新陈代谢和肥胖,但保护免于疾病的生物体仍然未知。
技术实现要素:
6.本文公开了细菌聚生体。本文公开了梭菌属聚生体。
7.本文公开了包含来自梭菌纲聚生体的上清液的组合物。
8.本文公开了包含梭菌属聚生体的组合物。
9.本文公开了一种包含梭菌纲anaerovorax菌株、梭菌属xiva、梭菌属iv和毛螺菌科某种的细菌聚生体,其中与未施用聚生体的受试者相比,聚生体遏制受试者的肠上皮细胞内的脂质吸附基因的表达。
10.本文公开了改变受试者中微生物群的相对丰度的方法,所述方法包括向受试者施用有效剂量的本文的任何组合物,从而改变受试者中微生物群的相对丰度。
11.本文公开了治疗患有肥胖症的受试者的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
12.本文公开了治疗患有代谢综合征的受试者的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
13.本文公开了治疗患有肠易激疾病的受试者的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
14.本文公开了减少受试者的体重增加的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
15.本文公开了抑制受试者小肠中的脂质吸收的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
16.本文公开了下调受试者肝脏中的cd36的方法。在一些方面,方法可包括向受试者
施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
附图说明
17.并入本说明书且构成其一部分的附图说明了若干个方面,并连同描述一起用于解释本发明的原理。
18.本发明的额外优势将部分地阐述于随后描述中,并且部分地将根据描述而明显,或者可通过实施本发明来获悉。本发明的优点将借助在所附权利要求书中特别指出的要素和组合来实现和达到。应理解,以上概述和以下详述都仅是示例性和解释性的,并且不限制要求保护的本发明。
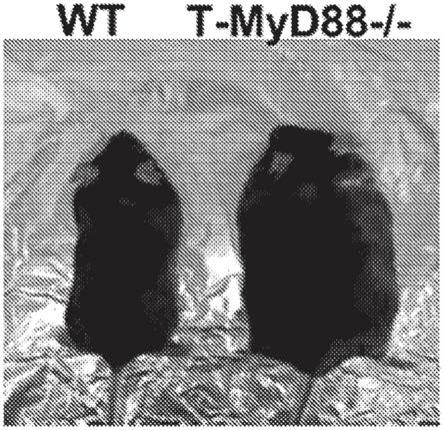
19.图1a-j示出肠道中的缺陷性t细胞信号传导导致年龄相关的肥胖。图1a示出6个月wt和t-myd88-/-小鼠的代表性图像。图1b示出从2月龄开始,随着小鼠年龄增长增加的体重的百分比(绘制wt,n=8;t-myd88-/-,n=7)。代表三个独立实验。图1c示出从2月龄开始,随着小鼠年龄增长的脂肪累积(绘制wt,n=8;t-myd88-/-,n=7)。代表三个独立实验。图1d示出1岁wt和t-myd88-/-小鼠(n=6)的总重量。代表三个独立实验。图1e示出如通过nmr测量的1岁wt和t-myd88-/-小鼠(n=6)的总脂肪百分比。代表三个独立实验。图1f示出来自1岁wt和t-myd88-/-小鼠(wt,n=9;t-myd88-/-,n=10)的空腹血清胰岛素浓度。数据从三个独立的实验汇集。图1g示出1岁wt和t-myd88-/-小鼠的稳态模型评估(homa-ir)。(wt,n=9;t-myd88-/-,n=10)。数据从三个独立的实验汇集。图1h示出在胰岛素抵抗测试期间腹膜内胰岛素(0.75u/kg)注射后随时间推移测量的血糖水平(wt,n=9;t-myd88-/-,n=10)。数据从三个独立的实验汇集。图1i示出来自wt和t-myd88-/-小鼠的肝脏和vat组织的代表性苏木精和伊红染色,放大20倍。比例尺表示100μm。图1j示出用对照或hfd喂养的wt和t-myd88-/-小鼠增加的体重的百分比(wt ctrl,n=8;wt hfd,n=15;t-myd88-/-ctrl,n=9;t-myd88-/-hfd,n=13)。p值<0.05(*);p值<0.01(**);p值<0.001(***);p值<0.0001(****),使用双尾未配对t检验(b-g)和重复测量anova(h,j)。误差线表示sd。
20.图2a-d示出与t-myd88-/-小鼠相关的体重增加需要微生物群。图2a示出随时间推移测量的增加的体重的克数(平均值 /-sd)。图2b示出增加的总重量(auc)。图2c示出vat的克数。图2d示出当wt和t-myd88-/-小鼠在有或没有抗生素的情况下用hfd喂养时的最终体脂百分比(wt ctrl,n=5;tmyd ctrl,n=4;wt abx,n=5,tmyd abx,n=5)。代表两个独立实验。p值<0.05(*);p值<0.01(**);p值<0.001(***);p值<0.0001(****),使用重复测量anova(a)和双尾未配对t检验(b-d)。
21.图3a-f示出多样性丧失和梭菌纲丰度与t-myd88-/-小鼠的体重增加相关。图3a示出pcoa图,并且图3b示出从指定动物的回肠微生物群中观察到的otu的数量(wt,n=8;t-myd88-/-,n=7)。图3c示出影响wt与t-myd88-/-回肠微生物群之间随机森林分类的平均准确性的前10种细菌属。wt动物中相对丰度丰富的属加蓝色阴影,t-myd88-/-动物中相对丰度丰富的属加红色阴影(wt,n=8;t-myd88-/-,n=7)。图3d示出影响体重增加和回肠微生物群的随机森林线性化中的标准误差的前十种细菌属。wt动物中相对丰度丰富的属加蓝色阴影,t-myd88-/-动物中相对丰度丰富的属加红色阴影(wt,n=8;t-myd88-/-,n=7)。图3e示出回肠样品中细菌uniref90基因家族转录物丰度比的火山图(每群组n=6)。图3f示出来自wt和
t-myd88-/-回肠微生物群转录物的每百万显著不同物种的映射读数(每种基因型n=6)。误差线表示sd。图3a、3b、3c和3d中的数据来自一个实验,并且来自图3e和3f的数据来自一个实验。p值<0.05(*);p值<0.01(**);p值<0.001(***);p值<0.0001(****),使用permanova(a)和双尾未配对t检验(b,f)。
22.图4a-g示出对肠道微生物群的操作影响t-myd88-/-相关的体重增加。图4a示出增加的体重的曲线下面积(auc),并且图4b示出维持在单独的笼子中或共同圈养并用hfd喂养的wt和t-myd88-/-小鼠中脱硫弧菌属的相对丰度(每种基因型n=4)。代表两个独立实验。图4c示出来自有或没有脱硫脱硫弧菌定殖的spf小鼠的粪便样品内指定细菌的相对丰度(每种基因型n=5)。图4d示出在单独或与脱硫脱硫弧菌一起用梭菌纲聚生体定殖的无菌小鼠中来自16s测序的指定细菌的相对丰度(每群组n=5)。误差线表示sd。图4e-g)示出用媒介物对照或孢子形成梭菌纲聚生体管饲的t-myd88-/-小鼠(媒介物(ctrl),n=4;梭菌纲聚生体,n=5)。代表两个独立实验。(e)增加的体重的auc。图4f示出如通过nmr测量的总脂肪百分比。(g)vat的克数。p值<0.05(*);p值<0.01(**);p值<0.001(***);p值<0.0001(****),使用双尾未配对的t检验(a,e-g)和曼-惠特尼u检验(b-d)。
23.图5a-h示出微生物群的tfh细胞调控可预防肥胖症。图5a-h示出tcrb
‑′‑
小鼠,在给予wt或t-myd88-/-t细胞前一周向小鼠给予wt和t-myd88-/-微生物群的混合物。然后将小鼠单独圈养8周,并测量体重增加和微生物群组成,同时用正常食物喂养(每群组n=6)。图5a示出增加的体重的曲线下面积(auc)分析。图5b示出代表性流式细胞术图之前在sybr green 细胞上进行了门控,以定量8周时抗体结合的细菌的百分比。rag1
‑′‑
粪便对照(灰色阴影区域);tcrb
‑′‑
粪便(灰线);wt(蓝线);t-myd88-/-(红线)。右侧多个动物的定量。图5c示出在第0、7和28天给予wt或t-myd88-/-t细胞的tcrβ
‑′‑
小鼠的微生物群之间的布雷柯蒂斯(bray-curtis)距离的小提琴图。图5d示出在给予wt或t-myd88-/-cd4 t细胞的tcrb
‑′‑
小鼠中脱硫弧菌科丰度与梭菌科丰度之间的相关性(n=12)。图5e示出梭菌科的相对丰度(4周)。误差线表示sd。图5f示出iga结合和iga未结合的细菌是从给予wt或t-myd88
‑′‑
cd4
t细胞的tcrb
‑′‑
小鼠的盲肠内容
24.物分析的。计算每个otu的iga指数以显示结合的差异。正值表示结合部分的富集,并且负值表示未结合部分的富集。示出所有具有统计学显著差异的otu(p<0.05,wilcoxon秩和检验)。每个图根据其最佳分类等级(属(g)、科(f)或目(o))对具有相同分类学调用的otu进行分组。每个点代表单独的动物,而图中的不同颜色区分分类群中的otu,并且每条线连接来自每个otu的平均值。数据来自一个实验。图5g示出用wt或t-myd88-/-粪便微生物群定殖的rag1
‑′‑
小鼠中增加的体重百分比(每群组n=7)。代表两个独立实验。图5h示出接受供体wt或bcl6
‑′‑
t细胞并用正常食物喂养的t-myd88-/-小鼠中增加的体重的auc(wt供体,n=5;t-myd88-/-供体,n=6)。代表两个独立实验。p值<0.05(*);p值<0.01(**);p值<0.001(***);p值<0.0001(****),使用双尾未配对t检验(a、b、h)、重复测量anova与tukey多重比较(c)、spearman等级次序相关性(d)、重复测量anova sidak多重比较校正(g)和曼-惠特尼u检验(e)。误差线表示sd(d,g)。
25.图6a-m示出梭菌纲抑制肠内的脂质吸收。图6a示出来自1岁wt和t-myd88
‑′‑
小鼠的肝脏中的rna表达的gsea分析,包括显著fdr为.25或更小的途径。图6b示出肝脏转录物比率的火山图。突出显示的基因参与脂质代谢。图6c示出在有或没有抗生素(abx)的情况下用
hfd喂养的wt和t-myd88
‑′‑
小鼠的肝脏内的cd36 rna表达(wt,n=5;t-myd88-/-,n=4;wt abx,n=5,t-myd88-/-abx,n=5)。代表两个独立实验。图6d示出用媒介物对照或孢子形成梭菌纲聚生体管饲的t-myd88
‑′‑
小鼠的肝脏中的cd36 rna表达(对照n=4;梭菌纲聚生体,n=5)。代表两个独立实验。图6e-g示出有或没有梭菌纲聚生体定殖的无菌小鼠(gf,n=8;梭菌纲,n=10)。图6e示出肝脏中的cd36 rna表达。图6f示出小肠(si)中的cd36 rna表达。图6g示出si中的fasn rna表达。图6h示出用培养基或无细菌细胞上清液(cfs)孵育4小时的mode-k细胞中的cd36 rna表达。代表三个独立实验。图6i示出无菌小鼠与梭菌纲聚生体或两种脱硫弧菌属物种(d.piger和脱硫脱硫弧菌)相关。通过nmr分析测量体脂百分比。(无菌小鼠n=12;梭菌纲,n=16,脱硫弧菌属,n=14)。图6j、k示出无菌小鼠与有或没有d.piger和脱硫脱硫弧菌(dsv)的梭菌纲聚生体相关。通过nmr测量体脂百分比(仅梭菌纲n=16;梭菌纲 dsv n=21)(j)并且通过q-pcr测量小肠内的cd36(图6k)。图6l、m示出用hfd喂养的wt和t-myd88-/-小鼠的血清和盲肠内容物内gc-ms检测到的代谢物(每群组n=6)。p值<0.06(φ),p值<0.05(*);p值<0.01(**);p值<0.001(***);p值<0.0001(****),双尾未配对t检验(c-g,i-k)和单因素anova sidak多重比较校正(图6h)。数据表示为平均值 /-sd。
26.图7a-g示出在t细胞内缺乏myd88信号传导的小鼠发展年龄相关的肥胖症。图7a示出从2月龄开始,随着小鼠年龄增长增加的体重(wt,n=8;t-myd88-/
‑ n=7)。图7b示出从2月龄开始,随着小鼠年龄增长增加的脂肪的百分比(wt,n=8;t-myd88-/-,n=7)。图7c示出在1岁wt和t-myd88-/-小鼠的葡萄糖耐量测试期间腹膜内葡萄糖(1mg/g)注射后随时间推移测量的血液葡萄糖水平(mg/dl)。图7d示出在2个月时用正常食物喂养时每只小鼠的食物摄入克数(每群组n=3)。图7e示出在用正常食物喂养1岁小鼠时每只小鼠的食物摄入克数(每组n=5。图7f示出2月龄的热量、能量消耗和总运动(每组n=3)。图7g示出1岁小鼠的热量、能量消耗和总运动(每组n=5)。统计数据:p值<0.05(*);p值<0.01(**);p值<0.001(***),使用重复测量anova与sidak多重比较校正(图7a、7b、7c),双尾未配对t检验(图7d-g)。误差线表示sd。
27.图8a-b示出t-myd88-/-小鼠的肥胖症因膳食脂肪摄入增加而加速。图8a示出在高脂肪饮食喂养后动物随时间推移的体重。图8b示出对照食物或hfd喂养后16周的年龄匹配的指定动物的内脏脂肪量。统计数据:p值<0.05(*);p值<0.01(**);p值<0.001(***),使用重复测量anova(图8a)和双尾未配对t检验(图8b)。误差线表示sd。
28.图9a-c示出t-myd88-/-小鼠体内微生物组成的变化与自发的体重增加相关。图9a示出来自1岁wt和t-myd88
‑′‑
小鼠的回肠和粪便16s测序样品的β多样性分析,通过未加权的unifrac和加权的unifrac进行测量(wt,n=8;t-myd88
‑′‑
,n=7)。图9b示出微生物群落的随机森林分析。图9c示出粪便和回肠微生物群中梭菌纲otu的数量和相对丰度(wt,n=8;t-myd88
‑′‑
n=7)。统计数据:p值<0.05(*);p<0.01(**);p值<0.001(***),使用permanova(c)。
29.图10a-c示出来自wt或t-myd88-/-小鼠的微生物群的转录组数据。图10a示出粪便样品中细菌uniref90基因家族转录物丰度比的火山图。图10b示出wt和t-myd88
‑′‑
小鼠回肠和粪便中每百万的独特映射读数。图10c示出来自wt和t-myd88
‑′‑
粪便转录物的每百万显著不同物种的映射读数(每种基因型n=6)。统计数据:p值<0.05(*);p值<0.01(**);p值<0.001(***),使用双尾未配对t检验。误差线表示sd。
30.图11a-e示出在共同圈养期间t-myd88-/-小鼠内的生态失调使肥胖症转移至wt动物。图11a示出共同圈养实验的示意图和共同圈养实验的时间线的示意图。图11b示出体重增加百分比,图11c示出脂肪百分比,并且图11d示出用hfd喂养的分开或共同圈养的wt和t-myd88-/-小鼠中的vat克数(每群组n=4)。代表两个独立实验。图11e示出在用hfd喂养的分开或共同圈养的wt和t-myd88-/-小鼠的葡萄糖耐量测试期间腹膜内葡萄糖(1mg/g)注射后随时间推移测量的血液葡萄糖水平(mg/dl)(每群组n=4)。统计数据:p值<0.05(*);p值<0.01(**);p值<0.001(***),重复测量anova(b,e),双尾未配对t检验(c,d)。误差线表示sd。
31.图12a-f示出共同圈养期间可传播生物体的测定。图a和b示出通过用正常食物喂养的分开或共同圈养的wt和t-myd88
‑′‑
小鼠在共同圈养之前和共同圈养后一周(图12a)且然后14周hfd之后(图12b)的未加权unifrac分析测量的β多样性。图12c和d示出在最后时间点来自分开或共同圈养的wt和t-myd88
‑′‑
小鼠的粪便16s测序样品内指定生物体的相对丰度(每群组n=4)。图12e示出仅在共同圈养一周后,来自指定动物的粪便样品内脱硫弧菌属的相对丰度。图12f示出12周共同圈养后指定动物中多尔氏菌属(dorea)的相对丰度。统计数据:p值<0.05(*);p值<0.01(**);p值<0.001(***)permanova(a,b)和曼-惠特尼u检验(c-f)。
32.图13示出脱硫弧菌属的扩增导致梭菌纲的损失。图示出来自有或没有定殖脱硫脱硫弧菌并用hfd喂养的小鼠的粪便16s测序样品内毛螺菌科的相对丰度(每群组n=5)。统计数据:p值<0.05(*);p值<0.01(**);p值<0.001(***)曼-惠特尼u检验。
33.图14示出来自用孢子形成微生物定殖的无菌小鼠的微生物群组成。来自用孢子形成梭菌纲聚生体定殖的无菌小鼠的粪便微生物群的16s测序的全图的部分。
34.图15a-i示出微生物群的t细胞成形与自发的体重增加相关。图15a示出通过多种转移方法(cf=交叉培养)给予wt或t-myd88-/-微生物群的无菌小鼠的体重增加。图15b示出实验策略的示意图。将tcrb-/-动物通过抗生素处理耗尽微生物群,且随后用来自wt或t-myd88-/-动物的1∶1微生物群混合物管饲。将wt或t-myd88-/-t细胞移植到t细胞缺陷型动物中。图15c示出用于定量在第0天、第1周和第8周给予wt或t-myd88-/-细胞的tcrb-/-小鼠内iga结合细菌的百分比的流式细胞术(每群组n=6)。图15d、e示出用于定量在第0天、第1周和第8周给予wt或t-myd88-/-细胞的tcrb-/-小鼠内igg1结合细菌的百分比的流式细胞术。图15f示出用于定量在第0天、第1周和第8周给予wt或t-myd88-/-细胞的tcrb-/-小鼠内igg3结合细菌的百分比的流式细胞术。图15g示出8周后使用elisa测量给予wt或t-myd88-/-细胞的tcrb-/-小鼠中管腔iga的浓度(μg/ml)。误差线表示sd。图15h示出代表性流式细胞术图先前在sybr green
细胞上进行门控,以定量8周后给予wt或t-myd88-/-细胞的tcrb-/-小鼠内igg3结合细菌的百分比。图15i示出用wt或t-myd88-/-粪便微生物群定殖的rag1-/-小鼠的增加的体重的auc(每群组n=7)。统计数据:p值<0.05(*);p值<0.01(**);p值<0.001(***),双尾未配对t检验(a,c-i)。
35.图16a-b示出细菌群落的iga靶向(图16a)。iga结合的和未结合的细菌给予wt或t-myd88-/-t细胞的tcrb-/-的盲肠内容物进行分析。计算每只动物中每种属的iga指数(列)。将气泡根据结合或未结合部分的富集来着色,并根据富集程度(iga指数的绝对值)确定大小。属分类群字符串根据其分类学类别进行着色。指示基因型之间显著差异结合的属(*,p<
0.05;wilcoxon秩和检验)。图16b示出此数据集中的脱硫弧菌属iga靶向。误差线表示sd。
36.图17a-c示出肠道代谢物与体重增加相关(图17a)gc-ms检测到wt和t-myd88-/-小鼠盲肠内容物的scfa(wt,n=3;t-myd88-/-n=5)。图17b示出从2月龄开始,用对照饮食或5-asa饮食喂养的wt和t-myd88-/-小鼠所增加的体重克数(wt ctrl,n=3;wt 5-asa,n=4;t-myd ctrl,n=3;tmyd 5-asa,n=4)。误差线表示sd。图17c示出梭菌纲的相对丰度与脂肪酸和单酰基甘油代谢物之间的spearman等级次序相关性(n=12)。统计数据:p值<0.05(*);p值<0.01(**);p值<0.001(***),双尾未配对t检验(a)和重复测量anova(b)。
37.图18示出4种菌株(即梭菌纲anaerovorax菌株、梭菌属xiva、梭菌属iv和毛螺菌科某种)的精制梭菌纲聚生体(rcc-4)足以减轻肥胖症。雌性(f)和雄性(m)无菌小鼠用从更复杂梭菌纲聚生体培养的4种菌株(rcc)定殖,并在定殖后4周通过nmr进行分析。rcc-4将肥胖症减轻至与雄性而非雌性中的复杂梭菌纲聚生体相同的程度。
38.图19a-g示出梭菌纲治疗改善mets和ibd。图19a-c示出在hfd同时每三天用媒介物对照或孢子形成梭菌纲聚生体管饲持续三个月的t-myd88-/-小鼠。图19a,增加的重量;和图19b,如通过nmr测量的总脂肪百分比。c,内脏脂肪组织。图20d和图20e均示出野生型(wt)和t-myd88-/-(ko)置于hfd并用梭菌纲进行处理,如图19a和图19d,空腹血糖。图19e示出homa-ir值。图19f和图19g示出在dss结肠炎的三个周期(5天dss和10天水)时每隔一天用梭菌纲聚生体口服管饲的动物。f是结肠长度,并且g是来自结肠的h&e染色切片的病理学评分。p值<0.05(*);p值<0.001。
具体实施方式
39.通过参考本发明的以下详细描述、附图和本文所包括的实施例,可以更容易地理解本公开。
40.在公开和描述本发明的方法和组合物之前,应理解它们不限于特定的合成方法(除非另外指明)或特定的试剂(除非另外指明),因此此类方法和组合物当然可以变化。还应理解,本文所使用的术语仅出于描述特定方面的目的,并且不意图是限制性的。尽管与本文中描述的那些方法和材料类似或等效的任何方法和材料均可用于本发明的实践或检验,但现在描述示例性方法和材料。
41.此外,应当理解的是,除非另有明确说明,否则决不意味着在此阐述的任何方法都被解释为要求其步骤以特定次序执行。因此,在方法权利要求项没有实际叙述其步骤所遵循的顺序或在权利要求书或描述中没有另外具体陈述各步骤将限于特定顺序的情况下,决不意图在任何方面推断顺序。这适用于任何可能的非表达解释基础,包括关于步骤排列或操作流程的逻辑事项、从语法组织或标点中得到的明显含义,以及在说明书中所描述的各方面的数量或类型。
42.为了公开和描述公布在引用时所涉及的方法和/或材料,本文中提到的所有公布均以引用的方式并入本文。提供本文中讨论的公布仅仅是针对它们在本技术的提交日期之前的公开。本文中的任何内容均不应解释为承认由于先前发明而使本发明无权先于这些公布。此外,本文中提供的公布日期可能不同于实际的公布日期,它们可能需要单独确认。
43.定义
44.如说明书和所附权利要求书中所使用,除非上下文另外明确规定之外,否则单数
形式“一个/种(a/an)”和“所述”包括复数指示物。
45.如本文所用的字词“或”表示特定列表的任何一个成员,并且还包括列表的成员的任何组合。
46.范围在本文中可表示为“约(about)”或“大约(approximately)”一个特定值和/或至“约”或“大约”另一特定值。当表示这样一个范围时,另外的方面包括从一个特定值和/或至另一个特定值。类似地,当通过使用先行词“约”或“大约”将值表达为近似值时,将理解的是特定值形成了另外方面。将进一步理解,每个范围的端值对于另一端值而言都是重要的,并且独立于另一端值。还应理解,本文公开了多个值,并且本文中每个值除值本身之外还公开为“约”特定值。例如,如果公开了值“10”,则还公开了“约10”。还应理解,还公开了两个特定单位之间的每个单位。例如,如果公开了10和15,那么也公开了11、12、13和14。
47.如本文所使用,术语“任选的”或“任选地”意指随后描述的事件或情况可能发生或可能不发生,并且描述包括所述事件或情况发生的实例和不发生的实例。
48.如本文所用,术语“样品”是指来自受试者的组织或器官;细胞(在受试者体内,直接取自受试者,或维持在培养中或来自培养的细胞系的细胞);细胞裂解物(或裂解物级分)或细胞提取物;或含有一种或多种源自细胞或细胞材料(例如多肽或核酸)的分子的溶液,其如本文所述进行测定。样品也可以是含有细胞或细胞组分的任何体液或排泄物(例如但不限于血液、尿液、粪便、唾液、泪液、胆汁、脑脊髓液)。在一些方面,样品可取自脑、脊髓、脑脊髓液或血液。
49.如本文所用,术语“受试者”是指施用的靶标,例如人。因此所公开方法的受试者可以是脊椎动物,诸如哺乳动物、鱼、鸟、爬行动物或两栖动物。术语“受试者”还包括家养动物(例如,猫、犬等)、家畜(例如,牛、马、猪、绵羊、山羊等)和实验室动物(例如,小鼠、兔、大鼠、豚鼠、果蝇等)。在一个方面中,受试者是哺乳动物。在另一个方面中,受试者是人。术语不表示特定的年龄或性别。因此,旨在涵盖成年、儿童、青春期和新生的受试者,以及胎儿,无论是雄性还是雌性。
50.如本文所用,术语“患者”是指患有疾病或病症的受试者。术语“患者”包括人和兽医受试者。在所公开的方法的一些方面中,例如在施用步骤之前,“患者”已经被诊断出需要治疗多发性硬化。在所公开的方法的一些方面中,例如在施用步骤之前,“患者”已经被诊断出需要治疗ii型糖尿病、肥胖症或炎症性肠病。
51.如本文所用,术语“正常”是指不患有ii型糖尿病、肥胖症或炎症性肠病或不具有增加的发展ii型糖尿病、肥胖症或炎症性肠病的易感性的个体、样品或受试者。
52.如本文所用,术语“易感性”是指受试者被临床诊断患有疾病的可能性。例如,对ii型糖尿病、肥胖症或炎症性肠病的易感性增加的人受试者可指受试者被临床诊断为ii型糖尿病、肥胖症或炎症性肠病的可能性增加的人受试者。
53.如本文所用,术语“包含(comprising)”可包括“由......组成”和“基本上由......组成”的方面。
54.如本文所用,“对照”是来自正常受试者或来自未患ii型糖尿病、肥胖症或炎症性肠病的正常受试者的组织的样品。
55.如本文所用,“过表达”意指表达大于在正常样品中检测到的表达。例如,过表达的核酸可表达比正常表达水平高约1个标准差、或比正常表达水平高约2个标准差或比正常表
达水平高约3个标准差。因此,比对照表达水平高约3个标准偏差表达的核酸是过表达的核酸。
56.如本文所用,“治疗”意指将本发明的化合物或分子施用于患有ii型糖尿病、肥胖症或炎症性肠病的受试者,例人或其他哺乳动物(例如,动物模型),以预防或延缓疾病或疾患的影响的恶化,或部分或完全逆转疾病的影响。
57.如本文所用,“预防”意指使具有增加的发展ii型糖尿病、肥胖症或炎性肠病易感性或将发展ii型糖尿病、肥胖症或炎症性肠病的受试者的机会最小化。
58.如本文所用,术语“参考”、“参考表达”、“参考样品”、“参考值”、“对照”、“对照样品”等,当在一种或多种微生物的样品或表达水平背景下使用时是指参考标准,其中参考在不同(即,不是同一组织,而是多个组织)组织之间以恒定水平表达,并且不受实验条件的影响,并且指示样品中预先确定的疾病状态(例如,未患ii型糖尿病、肥胖症或炎症性肠病)的水平。参考值可以是预先确定的标准值或预先确定的标准值的范围,代表没有疾病,或预先确定的疾病类型或严重程度。
59.组合物
60.本公开涉及含有细菌分离物和群落的组合物以及使用细菌分离物和群落的方法。特别地,本公开涉及一种组合物,所述组合物含有一种或多种来自如本文所公开的细菌聚生体的微生物,特别是表1中所公开的微生物或其混合物。在一个优选的实施方案中,组合物将包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株。微生物可通过鉴定对应于seq id no 1-4并且与seq id no 1-4至少98%同一的16s核糖体基因序列来进行表征。
61.本文公开了新鉴定的细菌。发现具有包含seq id no:1、2、3和4的16s rdna序列的细菌属于梭菌属。在一些方面,本文所述的组合物包含至少一种细菌,其中细菌是梭菌属。
62.梭菌纲anaerovorax。本文公开了一种细菌,梭菌纲anaerovorax。如本文所用的梭菌纲anaerovorax是指具有与seq id no:1共享至少98%序列同一性的16s核酸序列的细菌。梭菌纲anaerovorax具有nrrl或atcc登录号____。梭菌纲anaerovorax分离自来自cd4-cre
小鼠(wt)的小肠下部的粪便颗粒和管腔内容物。
63.梭菌属xiva。本文公开了一种细菌,梭菌属xiva。如本文所用的梭菌属xiva是指具有与seq id no:2共享至少98%序列同一性的16s核酸序列的细菌。梭菌属xiva具有nrrl或atcc登录号____。梭菌属xiva分离自来自cd4-cre
小鼠(wt)的小肠下部的粪便颗粒和管腔内容物。
64.梭菌属iv。本文公开了一种细菌,梭菌属iv。如本文所用的梭菌属iv是指具有与seq id no:3共享至少98%序列同一性的16s核酸序列的细菌。梭菌属iv具有nrrl或atcc登录号____。梭菌属iv分离自来自cd4-cre
小鼠(wt)的小肠下部的粪便颗粒和管腔内容物。
65.毛螺菌科某种。本文公开了一种细菌,毛螺菌科某种。如本文所用的毛螺菌科某种是指具有与seq id no:4共享至少98%序列同一性的16s核酸序列的细菌。毛螺菌科某种具有nrrl或atcc登录号____。毛螺菌科某种分离自来自cd4-cre
小鼠(wt)的小肠下部的粪便颗粒和管腔内容物。
66.梭菌纲聚生体。本公开涉及含有细菌分离物和群落的组合物以及使用细菌分离物和群落的方法。本文公开了梭菌纲聚生体(两种或更多种不同细菌菌株的混合物)。特别地,
本公开涉及组合物,所述组合物含有一种或多种来自如本文所公开的细菌聚生体的微生物,特别是表1中所公开的微生物或其混合物。在一些方面,组合物将包含来自表1中列出的那些的两种或更多种细菌菌株。微生物可通过鉴定对应于seq id no:1-4并且与seq id no:1-4至少98%同一的16s核糖体基因序列和/或与分别具有nrrl或atcc登录号:____、____、____和____的细菌比较来进行表征。在一些方面,梭菌纲聚生体包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株。在一些方面,梭菌纲聚生体包含三种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株。在一些方面,梭菌纲聚生体包含四种具有分别包含seq id no:1、2、3和4的16s rdna序列的细菌菌株。
67.在一些方面,梭菌纲聚生体包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株而没有任何其他细菌菌株。在一些方面,梭菌纲聚生体包含三种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,而没有任何其他细菌菌株。在一些方面,梭菌纲聚生体包含四种具有分别包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,而没有任何其他细菌菌株。
68.在一些方面,梭菌纲聚生体包含多达四种表1中列出的细菌菌株。在一些方面,梭菌纲聚生体包含两种或更多种表1的细菌菌株。在一些方面,梭菌纲聚生体包含梭菌纲anaerovorax、梭菌属xiva、梭菌属iv和毛螺菌科某种。在一些方面,梭菌纲聚生体包含梭菌纲anaerovorax、梭菌属xiva、梭菌属iv、毛螺菌科某种和一种或多种表1的细菌菌株。在一些方面,聚生体中的各种细菌可通过它们的16s核糖体基因序列来鉴定。
69.在一些方面,本文公开的梭菌纲聚生体在受试者中定殖时可使受试者的肥胖症降低至与包含更复杂的梭菌纲聚生体的复杂微生物群落相同的程度,所述梭菌纲聚生体包含多于四种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株。
70.在一些方面,分别与未处理的wt小鼠或t-myd88-/-小鼠相比,本文公开的梭菌纲聚生体在喂食高脂肪饮食时可减少wt小鼠或易肥胖t-myd88-/-小鼠的体重增加和脂肪累积。
71.在一些方面,分别与未处理的wt小鼠或t-myd88-/-小鼠相比,本文公开的梭菌纲聚生体可在喂食高脂肪饮食时降低wt小鼠或易肥胖t-myd88-/-小鼠的体脂百分比并减少vat量。
72.在一些方面,分别与未处理的wt小鼠或t-myd88-/-小鼠相比,本文公开的梭菌纲聚生体可在喂食高脂肪饮食时可降低wt小鼠或易肥胖t-myd88-/-小鼠的血糖水平并降低胰岛素抵抗。
73.本文公开了用于减轻受试者的肥胖症、减少受试者的体重增加和/或脂肪累积、降低受试者的体脂肪百分比和/或降低内脏脂肪组织(vat)量、降低受试者的血糖水平和/或降低胰岛素抵抗、抑制受试者小肠中的脂质吸收、下调受试者肝脏中的cd36和遏制受试者肠上皮细胞内脂质吸收基因的表达的梭菌纲聚生体。在一些方面,梭菌纲聚生体包含多达四种表1中列出的细菌菌株。在一些方面,表1的任何两种或更多种细菌菌株的组合可用于梭菌纲聚生体中。在一些方面,梭菌纲聚生体包含梭菌纲anaerovorax、梭菌属xiva、梭菌属iv和毛螺菌科某种。在一些方面,聚生体中的各种细菌可通过它们的16s核糖体基因序列来鉴定。
74.表1.细菌菌株。
75.76.77.78.79.80.81.82.[0083][0084]
本文公开了包含来自梭菌纲聚生体的上清液的组合物。本文还公开了包含梭菌属聚生体的组合物。在一些方面,梭菌纲聚生体包含梭菌纲anaerovorax、梭菌属xiva、梭菌属iv和毛螺菌科某种。在一些方面,梭菌纲聚生体包含梭菌纲anaerovorax、梭菌属xiva、梭菌属iv和毛螺菌科某种中的一种或多种。在一些方面,梭菌纲聚生体包含梭菌纲anaerovorax、梭菌属xiva、梭菌属iv和毛螺菌科某种中的两种或更多种。在一些方面,梭菌纲聚生体包含梭菌纲anaerovorax、梭菌属xiva、梭菌属iv和毛螺菌科某种中的三种或更多种。在一些方面,梭菌纲聚生体由梭菌纲anaerovorax、梭菌属xiva、梭菌属iv和毛螺菌科某种组成。在一些方面,梭菌纲聚生体由梭菌纲anaerovorax、梭菌属xiva、梭菌属iv和毛螺菌科某种中的一种或多种组成。在一些方面,梭菌纲聚生体由梭菌纲anaerovorax、梭菌属xiva、梭菌属iv和毛螺菌科某种中的两种或更多种组成。在一些方面,梭菌纲聚生体由梭菌纲anaerovorax、梭菌属xiva、梭菌属iv和毛螺菌科某种中的三种或更多种组成。
[0085]
在一些方面,本文所述的任何组合物能够遏制受试者肠上皮细胞内脂质吸附基因的表达。在一些方面,本文公开的任何组合物可遏制一种或多种脂质吸收和/或合成基因。在一些方面,脂质吸收基因可以是cd36、fasn、dgat、srepbf1、slc27a1和slc27a4。
[0086]
在一些方面,本文所述的任何组合物能够抑制受试者小肠中的脂质吸收。
[0087]
在一些方面,本文所述的任何组合物能够减少受试者的体重增加。
[0088]
在一些方面,本文所述的任何组合物能够下调受试者肝脏中的cd36。
[0089]
此外,本文公开了包含梭菌纲anaerovorax菌株、梭菌属xiva、梭菌属iv和毛螺菌科某种中的两种或更多种的细菌聚生体,其中与未施用聚生体的受试者相比,聚生体遏制受试者的肠上皮细胞内的脂质吸附基因的表达。
[0090]
在一些方面,本文所述的组合物还可包含一种或多种选自表1的梭菌纲菌株。
[0091]
在一些方面,所公开的组合物包含至少两种或更多种细菌微生物,所述细菌微生物可通过与seq id no:1-4的16s核糖体序列的至少95%、96%、97%、98%、99%或更高百分比同一性的同源性来鉴定。在一些方面,16s序列的量小于约1.2kb、1.1kb、1.0kb、0.9kb、8kb、0.7kb、0.6kb、0.5kb、0.4kb、0.3kb、0.2kb或0.1kb且大于约50nt、0.1kb、2kb、0.3kb、0.4kb、0.5kb、0.6kb、0.7kb、0.8kb、0.9kb、1.0kb或1.1kb。在一些方面,16s核糖体序列同源性的量介于约150nt与500nt之间,例如约250nt。为确定两种核酸的同一性百分比,为了最佳比较目的将序列进行比对(例如,可在第一核酸序列中引入空位,以与第二核酸序列最佳
比对)。然后比较相应核苷酸位置的核苷酸。当第一序列中的位置由与第二序列中的相应位置相同核苷酸占据时,那么在此位置的分子是相同的。两个序列之间的同一性百分比是序列共有的同一位置数量的函数(即,同一性%=同一位置数/位置总数
×
100)。
[0092]
两个序列之间的同源性百分比的确定可使用数学算法来完成。用于比较两个序列的数学算法的优选的非限制性实例是karlin和altschul(1990)proc.nat
′
l acad.sci.usa 87:2264-2268的算法,它在karlin和altschul(1993)proc.nat
′
l acad.sci.usa 90:5873-5877中进行了修改。这种算法被结合到altschul等人(1990)j.mol.biol.215:403-410的nblast和xblast程序中。blast核苷酸搜索可用nblast程序(得分=100、字长=12)来进行,以获得与本公开的核酸分子相似或同源的核苷酸序列。为了获得用于比较目的的空位比对,可利用空位blast,如altschul等人(1997)nucleic acids res.25:3389-3402中所述。当利用blast和空位blast程序时,可使用相应程序(例如xblast和nblast)的默认参数。这些算法可用于将dna与rna比对,并且在一些情况下可用于将蛋白质与翻译的核苷酸序列比对。
[0093]
在一些方面,至少两种或更多种微生物包含在本公开的组合物中。预期在两种或更多种微生物形成组合物的情况下,所述微生物可共培养以产生所公开的组合物。在一些方面,所公开的组合物可通过合并两种或更多种菌株的单独培养物来形成。微生物可通过本领域已知的方法来繁殖。例如,微生物可在厌氧或需氧条件下在液体培养基中繁殖。用于生长微生物的合适液体培养基包括本领域已知的那些,如营养肉汤和胰蛋白酶大豆琼脂(tsa)等。在一些方面,组合物包含表1中所列菌株的全部列表。在一些方面,组合物包含至少两种或更多种以下菌株:梭菌纲anaerovorax、梭菌属xiva、梭菌属iv和毛螺菌科某种。
[0094]
在一些方面,本文公开的组合物可包含至少1x10-5
个每种梭菌纲菌株的细胞。在一些方面,本文公开的组合物可包含至少1x10-6
个每种梭菌纲菌株的细胞。在一些方面,本文公开的组合物可包含至少1x10-7
个每种梭菌纲菌株的细胞。在一些方面,本文公开的组合物可包含至少1x10-8
个每种梭菌纲菌株的细胞。在一些方面,本文公开的组合物可包含至少1x10-9
个每种梭菌纲菌株的细胞。在一些方面,本文公开的组合物可包含至少1x10-10
个每种梭菌纲菌株的细胞。在一些方面,本文公开的任何组合物的单剂量可包含介于1x10-5
与1x10-10
之间个每种梭菌纲菌株的细胞。在一些方面,聚生体的细胞是活性的。
[0095]
在一些方面,本文公开的组合物能够替代患有与失衡的微生物群相关的疾病或病症的受试者的微生物群。在一些方面,本文公开的组合物能够替代患有与功能失调的微生物群相关的疾病或病症的受试者的微生物群。在一些方面,本文公开的组合物能够替代患有与功能多样性降低的微生物群相关的疾病或病症的受试者的微生物群。在一些方面,失衡的微生物群(或功能失调的微生物群或功能多样性降低的微生物群)可以是脱硫弧菌属的增加和梭菌纲的减少。在一些方面,失衡的微生物群(或功能失调的微生物群或功能多样性降低的微生物群)可以是脱硫弧菌属没有变化(或没有扩增)和梭菌纲的减少。在一些方面,失衡的微生物群(或功能失调的微生物群或功能多样性降低的微生物群)可以是梭菌纲的减少。在一些方面,失衡的微生物群(或功能失调的微生物群或功能多样性降低的微生物群)可以是梭菌纲不存在或缺乏。
[0096]
在一些方面,疾病或病症可以是肥胖症、代谢综合征、胰岛素缺乏、胰岛素抵抗相关病症、葡萄糖耐受不良、糖尿病或炎症性肠病。在一些方面,炎症性肠病可以是克罗恩氏
病或溃疡性结肠炎。在一些方面,胰岛素抵抗相关病症可以是糖尿病、高血压、血脂异常或心血管疾病。在一些方面,糖尿病可以是i型糖尿病。在一些方面,糖尿病可以是ii型糖尿病。
[0097]
在一些方面,本文公开的组合物还可包含药学上可接受的载体。在一些方面,组合物还可包含添加剂。合适的添加剂包括本领域已知的可支持生长、微生物产生特定代谢物、改变ph、富集目标代谢物、增强杀虫效果的物质,以及它们的组合。示例性添加剂包括碳源、氮源、磷源、无机盐、有机酸、生长培养基、维生素、矿物质、乙酸、氨基酸等。
[0098]
合适的碳源的实例包括但不限于淀粉、蛋白胨、酵母提取物、氨基酸、糖类如蔗糖、葡萄糖、阿拉伯糖、甘露糖、葡糖胺、麦芽糖、甘蔗、苜蓿提取物、糖蜜、朗姆酒等;有机酸如乙酸、富马酸、己二酸、丙酸、柠檬酸、葡萄糖酸、苹果酸、丙酮酸、丙二酸等的盐;醇类如乙醇、甘油等;油或脂肪如大豆油、米糠油、橄榄油、玉米油和芝麻油。添加的碳源的量根据碳源的种类而变化,并且通常在每升培养基1至100克之间。组合物中碳源的重量分数可以是组合物的总重量的约98%或更低、约95%或更低、约90%或更低、约85%或更低、约80%或更低、约75%或更低、约70%或更低、约65%或更低、约60%或更低、约55%或更低、约50%或更低、约45%或更低、约40%或更低、约35%或更低、约30%或更低、约25%或更低、约20%或更低、约15%或更低、约10%或更低、约5%或更低、约2%或约1%或更低。优选地,苜蓿包含于培养基中作为主要碳源,浓度为约1%至20%(w/v)。更优选地,苜蓿的浓度为约5至12%(w/v)。
[0099]
合适的氮源的实例包括但不限于氨基酸、酵母提取物、苜蓿提取物、胰蛋白胨、牛肉提取物、蛋白胨、硝酸钾、硝酸铵、氯化铵、硫酸铵、磷酸铵、氨或它们的组合。氮源的量根据氮源而变化,通常在每升培养基0.1至30克之间。组合物中氮源的重量分数可以是组合物的总重量的约98%或更低、约95%或更低、约90%或更低、约85%或更低、约80%或更低、约75%或更低、约70%或更低、约65%或更低、约60%或更低、约55%或更低、约50%或更低、约45%或更低、约40%或更低、约35%或更低、约30%或更低、约25%或更低、约20%或更低、约15%或更低、约10%或更低、约5%或更低、约2%或约1%或更低。
[0100]
合适的无机盐的实例包括但不限于磷酸二氢钾、磷酸氢二钾、磷酸氢二钠、硫酸镁、氯化镁、硫酸铁、硫酸亚铁、氯化铁、氯化亚铁、硫酸锰、氯化锰、硫酸锌、氯化锌、硫酸铜、氯化钙、氯化钠、碳酸钙、碳酸钠或它们的组合。组合物中无机盐的重量分数可以是组合物的总重量的约98%或更低、约95%或更低、约90%或更低、约85%或更低、约80%或更低、约75%或更低、约70%或更低、约65%或更低、约60%或更低、约55%或更低、约50%或更低、约45%或更低、约40%或更低、约35%或更低、约30%或更低、约25%或更低、约20%或更低、约15%或更低、约10%或更低、约5%或更低、约2%或约1%或更低。
[0101]
在一些方面,本公开的组合物还可包含乙酸或羧酸。合适的乙酸包括本领域中已知的任何乙酸,包括但不限于甲酸、乙酸、丙酸、丁酸、异丁酸、3-甲基丁酸、乙酸甲酯、乙酸乙酯、乙酸丙酯、乙酸丁酯、乙酸异丁酯和乙酸2-甲基丁酯。在一些方面,通过使用醋来包括乙酸。组合物中乙酸的重量分数可以是组合物的总重量的约98%或更低、约95%或更低、约90%或更低、约85%或更低、约80%或更低、约75%或更低、约70%或更低、约65%或更低、约60%或更低、约55%或更低、约50%或更低、约45%或更低、约40%或更低、约35%或更低、约30%或更低、约25%或更低、约20%或更低、约15%或更低、约10%或更低、约5%或更
低、约2%或约1%或更低。
[0102]
在一些方面,本文公开的化合物可冷冻。本公开的组合物可呈液体或干燥形式。在一些方面,本文公开的组合物可以是固体。在一些方面,本文公开的组合物可以是液体。在一些方面,组合物可包含组分的水性悬浮液。这种水性悬浮液可作为在施加前稀释的浓缩储备溶液提供,或作为即用型稀释溶液提供。此外,组合物可以是粉末、颗粒、粉剂、丸剂或胶体浓缩物。此类干燥形式可配制成在润湿后立即溶解或以受控释放、持续释放或其他时间依赖性方式溶解。此外,组合物可呈不依赖于润湿或溶解才能有效的干燥形式。
[0103]
在一些方面,本公开的组合物可包含至少一种任选的赋形剂。合适的赋形剂的非限制性实例包括抗氧化剂、添加剂、稀释剂、粘合剂、填充剂、缓冲剂、矿物盐、ph调节剂、崩解剂、分散剂、调味剂、营养剂、膨胀剂和渗透剂、稳定剂、防腐剂、适口性增强剂和着色剂。可根据已知的科学原理选择用于形成组合的赋形剂的量和类型。
[0104]
在一些方面,赋形剂可包括至少一种稀释剂。合适的稀释剂的非限制性实例包括微晶纤维素(mcc)、纤维素衍生物、纤维素粉末、纤维素酯(即乙酸酯和丁酸酯混合酯)、乙基纤维素、甲基纤维素、羟丙基纤维素、羟丙基甲基纤维素、羧甲基纤维素钠、玉米淀粉、磷酸化玉米淀粉、预胶凝玉米淀粉、大米淀粉、马铃薯淀粉、木薯淀粉、淀粉-乳糖、淀粉-碳酸钙、羟基乙酸淀粉钠、葡萄糖、果糖、乳糖、乳糖一水合物、蔗糖、木糖、乳糖醇、甘露醇、麦芽糖醇、山梨糖醇、木糖醇、麦芽糖糊精和海藻糖。
[0105]
在一些方面,赋形剂可包括粘合剂。合适的粘合剂包括但不限于淀粉、预胶凝淀粉、明胶、聚乙烯吡咯烷酮、纤维素、甲基纤维素、羧甲基纤维素钠、乙基纤维素、聚丙烯酰胺、聚乙烯噁唑烷酮、聚乙烯醇、c12-c18脂肪酸醇、聚乙二醇、多元醇、糖、寡糖、多肽、寡肽以及它们的组合。
[0106]
在一些方面,赋形剂可包括填充剂。合适的填充剂包括但不限于碳水化合物、无机化合物和聚乙烯吡咯烷酮。作为非限制性实例,填充剂可以是硫酸钙(二碱式和三碱式)、淀粉、碳酸钙、碳酸镁、微晶纤维素、磷酸氢钙、碳酸镁、氧化镁、硅酸钙、滑石、改性的淀粉、乳糖、蔗糖、甘露糖醇或山梨糖醇。
[0107]
在一些方面,赋形剂可包括缓冲剂。合适的缓冲剂的代表性实例包括但不限于mops、hepes、taps、bicine、tricine、tes、pipes、mes、tris缓冲剂或缓冲盐水盐(例如,tris缓冲盐水或磷酸盐缓冲盐水)。
[0108]
在一些方面,赋形剂可包括崩解剂。合适的崩解剂包括但不限于淀粉(如玉米淀粉、马铃薯淀粉、其预胶凝化和改性淀粉)、甜味剂、粘土(如膨润土)、微晶纤维素、藻酸盐、羟基乙酸淀粉钠、树胶(如琼脂、瓜尔胶、刺槐豆胶、刺梧桐胶、果胶和黄蓍胶)。
[0109]
在一些方面,赋形剂可包括分散增强剂。合适的分散剂可包括但不限于淀粉、海藻酸、聚乙烯吡咯烷酮、瓜尔胶(guar gum)、高岭土、膨润土、纯化木纤维素、羟基乙酸淀粉钠、同形硅酸盐和微晶纤维素。
[0110]
在一些方面,赋形剂可包括润滑剂。合适的润滑剂的非限制性实例包括矿物如滑石或二氧化硅;以及脂肪如植物硬脂、硬脂酸镁或硬脂酸。
[0111]
组合中一种或多种赋形剂的重量分数可以是组合的总重量的约98%或更低、约95%或更低、约90%或更低、约85%或更低、约80%或更低、约75%或更低、约70%或更低、约65%或更低、约60%或更低、约55%或更低、约50%或更低、约45%或更低、约40%或更
低、约35%或更低、约30%或更低、约25%或更低、约20%或更低、约15%或更低、约10%或更低、约5%或更低、约2%或约1%或更低。
[0112]
在一些方面,本公开的组合物在室温下是稳定的。
[0113]
在一些方面,聚生体可保持在降低的温度下用于储存和运输,而不会显著损害活微生物的活力。聚生体或包含所述聚生体的组合物可冷藏、冷冻或冻干。组合物可在32
°
f至44
°
f之间冷藏。
[0114]
在一些方面,聚生体或包含所述聚生体的组合物可以冷冻状态储存和运输。一旦组合物被解冻并达到环境温度,优选通过通气和/或搅拌,活的有益微生物可迅速恢复活力。
[0115]
在一些方面,聚生体可冻干。聚生体首先被冷冻。然后在真空下除去水。这种方法进一步降低用于储存和运输的组合物的重量。可在施加或施用之前使聚生体或包含所述聚生体的组合物重构并恢复活力。
[0116]
在一些方面,浓缩的聚生体或包含所述聚生体的组合物可在施加或施用之前用水稀释。稀释的组合物可储存延长的时间段,例如长达30天,而不会失去活力。为了将活的有益微生物维持在基本上需氧的状态,本公开的稀释组合物中的溶解氧优选保持在最佳水平。优选通过缓慢通气向稀释的组合物提供最佳量的氧气。
[0117]
在一些方面,本文公开的任何组合物可以选自由以下组成的组的形式施用:粉末、颗粒、即用型饮料、食品棒、挤出形式、胶囊、凝胶帽和分散片。
[0118]
保藏信息。本文公开的细菌xxx的保藏将由美国典型培养物保藏中心(atcc),10801university blvd.,manassas,va 20110-2209进行。保藏日期是____,并且保藏的细菌xxx的登录号是atcc登录号
‑‑‑‑‑
。已取消对保藏物的所有限制,并且保藏物旨在满足37c.f.r.
§
1.801-1.809的所有要求。保藏物将保存在保藏处,为期30年或最近一次请求后的5年或者专利的有效寿命(以较长者为准),在此期间如有必要,将被替换。
[0119]
方法
[0120]
本文公开了改变受试者中微生物群的相对丰度的方法。在一些方面,方法可包括向受试者施用有效剂量的本文公开的任何组合物,从而改变受试者中微生物群的相对丰度。在一些方面,方法可包括向受试者施用有效剂量的组合物,所述组合物包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,从而改变受试者中微生物群的相对丰度。在一些方面,可增加梭菌纲细菌的相对丰度。在一些方面,可替代梭菌纲细菌的相对丰度。在一些方面,本文公开的组合物可用于替代患有与失衡的微生物群(或功能失调的微生物群或功能多样性降低的微生物群)相关的疾病或病症的受试者的微生物群。
[0121]
改变微生物群的方法还可包括测量来自受试者的样品中一种或多种微生物群的相对丰度。如本文所用,术语“相对丰度”是指生物体相对于限定位置或群落中的其他生物体的共性或稀有性。例如,相对丰度可通过与样品中生物体的总存在相比,一般测量特定生物体的存在来确定。
[0122]
可直接或间接地测量微生物群的相对丰度。直接测量可包括基于培养的方法。间接测量可包括比较对与整个样品相关的生物体或一组生物体具有特异性的分子身份指示物如核糖体rna(rrna)基因序列的普遍性。例如,对在从盲肠样品获得的rrna基因序列总数中的脱硫弧菌属和梭菌纲具有特异性的rrna的比率可用于确定盲肠样品中脱硫弧菌属和
梭菌纲的相对丰度。
[0123]
如本文所用,术语“微生物群”用于指可在生物体的胃肠道内发现或存在(定殖)的一种或多种细菌群落。当提到多于一种微生物群时,微生物群可具有同一类型(菌株),或者它也可以是分类群的混合物。在一些方面,本文公开的方法和组合物改变受试者胃肠道中来自诸如梭菌属的属的微生物群的相对丰度。微生物群的相对丰度可通过施用药物组合物或化合物来改变,所述药物组合物包含来自诸如梭菌属的属的微生物群,所述化合物显著增加来自诸如梭菌属的属的微生物群的相对丰度或显著降低来自或诸如脱硫弧菌目的目的微生物群的相对丰度。
[0124]
在一些方面,梭菌纲的相对丰度可在受试者中增加至少约5%。在一些方面,梭菌纲的相对丰度可在受试者中增加至少约10%。在一些方面,梭菌纲的相对丰度可在受试者中增加至少约1%、2%、3%、4%、5%、6%、7%、8%、9%或10%。在一些方面,至少一种梭菌纲物种的相对丰度可增加5%。
[0125]
在一些方面,本文公开的方法还可包括向受试者施用第二治疗剂。在一些方面,第二治疗剂可以是一种或多种噬菌体。在一些方面,一种或多种噬菌体可特异性地靶向并杀死脱硫弧菌属。在一些方面,第二治疗剂可以是一种或多种可商购的治疗剂,可施用所述治疗剂以治疗肥胖症、ii型糖尿病和/或炎症性肠病。在一些方面,第二治疗剂可以是抗炎剂。
[0126]
本文公开了治疗患有肥胖症的受试者的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。在一些方面,方法包括向受试者施用组合物,所述组合物包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
[0127]
本文公开了治疗患有代谢综合征的受试者的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。在一些方面,方法可包括向受试者施用组合物,所述组合物包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
[0128]
本文公开了治疗患有肠易激疾病的受试者的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。在一些方面,方法可包括向受试者施用组合物,所述组合物包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
[0129]
本文公开了减少受试者的体重增加的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。在一些方面,方法可包括向受试者施用组合物,所述组合物包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
[0130]
本文公开了抑制受试者小肠中的脂质吸收的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。在一些方面,方法可包括向受试者施用组合物,所述组合物包含两种或更多种具
有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。在一些方面,本文公开的任何组合物可遏制一种或多种脂质吸收和/或合成基因。在一些方面,脂质吸收基因可以是cd36、fasn、dgat、srepbf1、slc27a1和slc27a4。
[0131]
本文公开了下调受试者肝脏中的cd36的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。在一些方面,方法可包括向受试者施用组合物,所述组合物包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
[0132]
本文公开了减轻受试者的肥胖症的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。在一些方面,方法可包括向受试者施用组合物,所述组合物包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
[0133]
本文公开了减少受试者的体重增加的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。在一些方面,方法可包括向受试者施用组合物,所述组合物包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
[0134]
本文公开了减少受试者的脂肪累积的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。在一些方面,方法可包括向受试者施用组合物,所述组合物包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
[0135]
本文公开了降低受试者的体脂百分比和/或减少内脏脂肪组织(vat)量的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。在一些方面,方法可包括向受试者施用组合物,所述组合物包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
[0136]
本文公开了降低受试者的血糖水平和/或降低胰岛素抵抗的方法。在一些方面,方法可包括向受试者施用本文公开的任何组合物,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。在一些方面,方法可包括向受试者施用组合物,所述组合物包含两种或更多种具有包含seq id no:1、2、3和4的16s rdna序列的细菌菌株,其中与施用前的相对丰度相比,受试者中梭菌纲的相对丰度增加。
[0137]
在一些方面,在本文公开的任何方法中,受试者已被鉴定为需要治疗。在一些方面,受试者患有肥胖症、代谢综合征、胰岛素缺乏、胰岛素抵抗相关病症、葡萄糖耐受不良、糖尿病或炎症性肠病。在一些方面,炎症性肠病可以是克罗恩氏病或溃疡性结肠炎。在一些方面,胰岛素抵抗相关病症可以是糖尿病、高血压、血脂异常或心血管疾病。在一些方面,糖尿病可以是i型糖尿病。在一些方面,糖尿病可以是ii型糖尿病。
[0138]
如本文所用,术语“代谢病症”或“代谢综合征”是指由异常体重增加、能量使用或消耗、对摄入或内源性营养素、能量来源、激素或体内的其他信号传导分子的反应改变或碳水化合物、脂质、蛋白质、核酸或其组合的代谢改变引起或表征的病症、疾病和疾患。代谢病症与导致核酸、蛋白质、脂质和/或碳水化合物代谢失衡的代谢途径的不足或过量相关。影响新陈代谢的因素包括但不限于内分泌(激素)控制系统(例如胰岛素途径、肠内分泌激素包括glp-1、pyy等)、神经控制系统(例如glp-1或脑中的其他神经递质或调控蛋白)等。一些非限制性实例可以是肥胖症、糖尿病(包括ii型糖尿病)、胰岛素缺乏、胰岛素抵抗、胰岛素抵抗相关病症、葡萄糖耐受不良、x综合征、炎症性和免疫性病症、骨关节炎、血脂异常、代谢综合征、非酒精性脂肪肝、脂质代谢异常、癌症、神经变性病症、睡眠呼吸暂停、高血压、高胆固醇、致动脉粥样硬化性血脂异常、高脂血症性疾患如动脉粥样硬化、高胆固醇血症和哺乳动物的其他冠状动脉疾病,以及其他代谢病症。
[0139]
病症还包括发生或聚集在一起并且增加患心脏病、中风、糖尿病和肥胖症的风险的疾患。仅患有这些疾患中的一者如血压升高、胰岛素水平升高、腰部体脂过多或胆固醇水平异常可增加上述疾病的风险。组合起来,患冠心病、中风、胰岛素抵抗综合征和糖尿病的风险甚至更大。
[0140]
在一些方面,施用本文公开的任何组合物的步骤可包括将组合物递送至受试者的至少胃、小肠或大肠。在一些方面,可口服施用组合物。
[0141]
在一些方面,受试者可以是人。
[0142]
在一些方面,聚生体的细胞是活性的。
[0143]
药盒
[0144]
在一些方面,公开了一种药盒,所述药盒包含一种或多种细菌、菌株或微生物,所述一种或多种细菌、菌株或微生物能够用于减轻受试者的肥胖症、减少受试者的体重增加和/或脂肪累积、降低受试者的体脂肪百分比和/或降低内脏脂肪组织(vat)量、降低受试者的血糖水平和/或降低胰岛素抵抗、抑制受试者小肠中的脂质吸收、下调受试者肝脏中的cd36和遏制受试者肠上皮细胞内脂质吸收基因的表达。
[0145]
实施例
[0146]
实施例1:t细胞介导的微生物群调控预防肥胖症
[0147]
摘要:微生物群影响宿主新陈代谢和肥胖,但保护免于疾病的生物体仍然未知。在询问调控微生物群组成的免疫途径的研究中,观察到由微生物群驱动的年龄相关性代谢综合征的发展。在此模型中,脱硫弧菌属的扩增和梭菌纲的损失是与肥胖症相关的重要特征,并且梭菌纲的替代可拯救肥胖症。需要t细胞依赖性事件来防止梭菌纲的损失和脱硫弧菌属的扩增。不适当梭菌纲iga靶向和增加的脱硫弧菌属拮抗有益梭菌纲的定殖。转录和代谢分析揭示肥胖宿主的脂质吸收增强。无菌动物的梭菌纲而非脱硫弧菌属定殖下调了控制脂质吸收和减轻肥胖症的基因的表达。此外,来自梭菌纲的上清液遏制了肠上皮细胞内脂质吸收基因的表达。梭菌纲的减少和脱硫弧菌属的增加是在患有代谢综合征和肥胖症的患者中发现的微生物群特征。因此,微生物群的免疫控制似乎维持有益的微生物群体,所述微生物群体的作用是限制脂质代谢以防止代谢缺陷。
[0148]
引言。在过去的一个世纪里,肥胖症和代谢综合征已经发展成全球流行病。目前,超过19亿人肥胖并有发展代谢功能障碍,如ii型糖尿病、心血管疾病和肝脏疾病的风险
(d.mozaffarian等人,circulation 131,e29-322(2015))。多项研究强调了代谢疾病的免疫系统调控的作用。这些报告主要集中在肥胖症期间炎症性反应的作用。他们报告了体重增加期间脂肪组织内的巨噬细胞浸润增加和调控性t细胞减少(m.f.gregor和g.s.hotamisligil,annu rev immunol 29,415-445(2011);以及f.emanuela等人,journal of nutrition and metabolism 2012,476380(2012))。然而,许多人类研究表明,次优的免疫反应也与代谢综合征和肥胖症相关。事实上,肥胖的成人对免疫接种表现出免疫反应不足、感染发生率增加和粘膜iga水平降低,从而表明这些个体无法获得有效的免疫(a.pallaro等人,j nutr biochem 13,539(2002);a.must等人,jama 282,1523-1529(1999);d.c.nieman等人,j am diet assoc 99,294-299(1999);d.n.mcmurray,p.a.beskitt,s.r.newmark,int j obes 6,61-68(1982);j.hirokawa等人,biochem biophys res commun,235,94-98(1997);以及s.tanaka等人,int j obes relat metab disord 17,631-636(1993))。缺陷性免疫反应影响代谢疾病的机制仍不清楚。
[0149]
微生物群已成为哺乳动物宿主内代谢的重要调节剂,并且肥胖个体中的微生物群组成在转移至动物体内时足以导致代谢缺陷(p.j.turnbaugh等人,nature 444,1027-1031(2006))。特别地,据报告,在代谢疾病期间微生物群的基因丰富度降低,包括丁酸盐和甲烷的产生减少。相反,一些微生物群的功能(如硫化氢和粘液降解)在患有代谢疾病的个体中得到增强(j.qin等人,nature 490,55-60(2012))。最近的研究表明,肠道免疫反应对于调控微生物群的组成是重要的(j.l.kubinak等人,cell host microbe 17,153-163(2015);以及s.kawamoto等人,immunity 41,152-165(2014))。特别地,iga的作用是限制某些微生物的外生长和使微生物群多样化;微生物的iga结合的变化或甚至肠道iga的轻微减少都可对多样性产生负面影响(j.l.kubinak等人,cell host microbe 17,153-163(2015);s.kawamoto等人,immunity 41,152-165(2014);以及s.wang等人,immunity 43,289-303(2015))。因此,微生物群的免疫控制缺陷可导致代谢疾病。
[0150]
结果。最近,鉴定了可指导微生物群的t细胞依赖性iga靶向的适当发展的分子途径。具有先天性衔接分子myd88的t细胞特异性消除的动物(t-myd88-/-小鼠)在肠道内有缺陷性t滤泡辅助(tfh)细胞发育和iga产生。这与肠道微生物的iga靶向失调和基因型之间微生物群内的组成差异相关(j.l.kubinak等人,cell host microbe 17,153-163(2015);和s.wang等人,immunity 43,289-303(2015))。在这些研究中,观察到较老的t-myd88-/-小鼠与其野生型对照相比始终肥胖(图1a)。尽管用标准食物喂养,但t-myd88-/-小鼠随着年龄的增长表现出显著增加的体重增加和脂肪累积(图1b和c以及图7a和b)。到1岁时,雄性t-myd88-/-小鼠重达60g,并且基于nmr分析表现出50%的体脂组成(图1d和e)。
[0151]
t-myd88-/-动物发展了许多在人中发现的代谢疾病合并症(f.x.pi-sunyer,med sci sports exerc 31,s602-608(1999))。尽管以标准饮食饲养的一岁大的t-myd88-/-小鼠将葡萄糖清除至与wt小鼠相似的水平(图7c),但它们的循环胰岛素水平更高,从而导致homa-ir指数更高(图1f和g)。此外,当用额外的胰岛素激发时,t-myd88-/-小鼠未能以与wt动物相似的动力学清除葡萄糖,从而表明胰岛素抵抗的发展(图1h)。与wt对照相比,食物摄入在两个月大的t-myd88-/-小鼠中减少,但在一岁大的动物中是相当的(图7d和e)。另外,虽然能量消耗在年轻小鼠中减少,但这些变化并没有随时间推移而持续存在(图7d)。运动在两个年龄的wt与t-myd88-/-小鼠之间也相似,并且与wt对照相比,在较老的t-myd88-/-小鼠
中测量到产热的适度增加,从而表明这些不是在其他模型中观察到的体重增加的主要原因(图7f和g)(m.vijay-kumar等人,science 328,228-231(2010))。t-myd88-/-小鼠也发展脂肪性肝病,并在脂肪组织内展现炎症表型,其特征是冠状结构和脂肪细胞大小失调(图1i)。喂养标准小鼠食物饲料的肥胖症需要几个月的时间来发展。相比之下,当动物被置于高脂肪饮食(hfd,45%脂肪)时,早在饮食开始后8周,t-myd88-/-动物就比wt小鼠累积了更多的体重和内脏脂肪组织(vat)量(图1j和图8a和b)。因此,t-myd88-/-动物容易发展代谢综合征和肥胖症,其可能通过增加膳食脂肪的摄入而加速。
[0152]
t-myd88-/-微生物群的组成在年轻动物中与wt不同(j.l.kubinak等人,cell host microbe 17,153-163(2015))。微生物群是代谢功能的已知贡献者,并且与人肥胖症的发展有关(s.ussar等人,cell metab 22,516-530(2015);和e.le chatelier等人,nature 500,541-546(2013))。为了初步确定微生物群是否与t-myd88-/-小鼠中观察到的代谢综合征有关,将wt和t-myd88-小鼠在hfd喂养的同时使用广谱抗生素。wt小鼠在使用抗生素时未表现出体重增加的差异。相比之下,t-myd88-/-动物中通过抗生素治疗完全拯救了体重增加(图2a和b)。这伴随着它们的体脂百分比和vat量降低至类似于在瘦动物中观察到的脂肪累积水平(图2c和d)。
[0153]
为了确定影响t-myd88-/-小鼠的代谢综合征的微生物群特征,对正常食物喂养的年老动物进行16s rrna基因测序,以评估肥胖t-myd88-/-小鼠中微生物群的分类学组成和多样性。在年老wt和t-myd88-/-小鼠的回肠和粪便内容物中存在显著不同的群落(图3a和图9a)。此外,年老小鼠的粪便中的物种丰富度略有降低(图3b)。为了鉴定可解释wt与t-myd88-/-微生物群落之间的主要差异的生物体,对16s rrna数据进行了随机森林分析。粪便微生物群能够以86%的准确率准确分类基因型,而回肠微生物群以100%的准确率预测基因型。对准确性影响最大的微生物群的成员大多属于广泛的分类学类别梭菌纲,并且与t-myd88-/-小鼠相比在wt小鼠中富集(图3c和图9b)。另一种随机森林方法表明,粪便和回肠微生物群可预测总重量,分别r2=0.5和r2=0.76,梭菌纲的许多成员强烈地影响这一预测(图3d和图9b)。与wt小鼠相比,t-myd88-/-小鼠显示出多种梭菌纲分类群(包括多尔氏菌属、smb53、未分类的消化链球菌科和梭菌属)的多样性和总体丰度的广泛降低(图9c)。
[0154]
微生物群的组成变化(包括微生物多样性降低)可对微生物群的功能产生负面影响,并且与包括代谢综合征在内的许多西方生活方式相关疾病相关(m.vijay-kumar等人,science 328,228-231(2010);和s.ussar等人,cell metab 22,516-530(2015))。此外,拥有基因丰富度较低的微生物群的个体更有可能肥胖(e.le chatelier等人,nature 500,541-546(2013))。在粪便和回肠微生物转录组中,来自t-myd88-/-动物中的许多基因家族的转录物的代表性普遍降低。由于通过16srrna基因测序在回肠内检测到相同数量的生物体,这支持了这些位点的微生物群降低了代谢功能的假设(图3e和图10a)。相当比例的总读数在两种基因型之间独特映射至参考基因组,从而表明动物中转录组的覆盖率相同。然而,在t-myd88-/-小鼠的回肠和粪便转录组中,映射至梭菌科参考基因组的读数的比例尤其显著降低(图3f和图10b和c)。因此,肥胖动物中存在的梭菌纲对微生物组的功能贡献降低。此外,肥胖症与梭菌纲内微生物功能多样性的损失相关,这在患有代谢疾病的人中也有类似报告(j.qin等人,nature 490,55-60(2012))。
[0155]
由于重要的梭菌纲生物体的损失可在疾病期间发挥作用,因此进行了共同圈养实
验以确定微生物转移是否能够拯救肥胖症(图11a)。由于小鼠是食粪性的,所以共同圈养允许微生物在基因型之间有效且频繁地转移,并且对微生物群具有已知的均质化作用。wt或t-myd88-/-动物与相同基因型的动物一起圈养,或者在断奶时与相反基因型的动物共同圈养。在共同圈养之前,t-myd88-/-小鼠具有独特的微生物群组成,并且共同圈养一周导致两个群落混合(图12a)。
[0156]
1周后,将动物置于hfd并监测脂肪累积的迹象。与分开的wt小鼠相比,t-myd88-/-小鼠和与它们共同圈养的任何动物体重显著增加,发展胰岛素抵抗,并增加了vat和总体脂肪(图4a和图11b至e)。此外,三个月后,来自共同圈养的wt动物的微生物群与分开圈养的wt小鼠明显不同,并且显示与分开圈养的t-myd88-/-的微生物群更大的相似性(图12b)。因此,微生物群的可转移组分在t-myd88-/-动物中形成,其可导致其他健康wt动物的代谢综合征。
[0157]
由于在前3周内检测到共同圈养动物的体重增加的差异,因此在早期和最终时间点可检测到的微生物组成的差异是焦点。在共同圈养三个月后,脱硫弧菌属、乳杆菌目和假长双歧杆菌在共同圈养的wt小鼠中的丰度更高(图4b和图12c和d)。然而,仅在共同圈养一周后,脱硫弧菌属在分开圈养的t-myd88-/-动物和共同圈养的动物中显示出显著更高的丰度(图12e)。脱硫弧菌属是粘液溶解性d-变形菌门,其产生硫化氢作为粘蛋白内二硫键降解的副产品(g.r.gibson,g.t.macfarlane,j.h.cummings,j appl bacteriol 65,103-111(1988);m.c.pitcher,j.h.cummings,gut 39,1-4(1996);f.e.rey等人,proc natl acad sci u s a 110,13582-13587(2013);以及m.s.desai等人,cell 167,1339-1353(2016))。除了与炎症性肠病(ibd)相关外,在患有ii型糖尿病和肥胖症的患者中检测到脱硫弧菌属的定植增加和与硫化氢产生相关的基因(j.qin等人,nature 490,55-60(2012))。因此,肥胖小鼠的群落变化与人类中观察到的大部分情况相似,并且表明梭菌纲的损失和脱硫弧菌属的增加与代谢疾病高度相关(j.qin等人,nature 490,55-60(2012);j.zierer等人,nat genet 50,790-795(2018);以及s.m.harakeh等人,front cell infect microbiol 6,95(2016))。
[0158]
wt小鼠与导致肥胖症的t-myd88-/-动物的共同圈养也与wt动物中梭菌纲成员的定殖减少相关(图12f)。因此,测试了脱硫弧菌属定殖是否可降低这些生物体的丰度。
[0159]
将无特定病原体的(spf)小鼠用脱硫脱硫弧菌脱硫亚种定殖1周,所述脱硫脱硫弧菌脱硫亚种是与在小鼠中鉴定的脱硫弧菌属具有大于97%的16s rrna基因序列相似性的菌株。结果表明,wt spf动物的梭菌目毛螺菌科和多尔氏菌属显著减少(图4c和图13a)。由于双歧杆菌属显著增加,所以脱硫弧菌属的定殖并未导致所有生物体的总体减少(图4c)。由于群落的这些变化可以是脱硫弧菌属定殖的间接影响,因此测试了脱硫弧菌属是否能够影响梭菌纲成员在无菌系统中的定殖。
[0160]
用氯仿处理的粪便浆液定殖的无菌动物富含梭菌科和毛螺菌科(图14)。然后在存在或不存在脱硫脱硫弧菌的情况下对这一群落进行分析。脱硫弧菌属定殖导致梭菌属的显著减少,梭菌属是强烈地影响基因型和体重的预测准确性的属(图4d)。因此,如在t-myd88-/-小鼠和患有ii型糖尿病的患者中所见,脱硫弧菌属物种的扩增可拮抗与瘦相关的微生物的定殖。
[0161]
试图鉴定重新引入这些瘦相关微生物是否能够预防t-myd88-/-小鼠的肥胖。每隔一天用孢子形成细菌的混合物对容易肥胖的t-myd88-/-动物进行治疗显著降低了体重增加
和脂肪累积(图4e和f)。在3个月结束时,与未处理的t-myd88-/-小鼠相比,用孢子形成微生物处理的t-myd88-/-小鼠的体脂百分比较低,并且vat量减少(图4f和g)。因此,梭菌纲的损失与t-myd88-/-小鼠中的肥胖症和代谢综合征有因果关系。
[0162]
在缺陷型肠道免疫期间形成的微生物群似乎导致代谢综合征。尽管动物共同圈养12周导致肥胖症传播至wt宿主中,但将t-myd88-/-的粪便移植到wt无菌受体中不足以转移肥胖症(图15a)。此外,当spf wt或t-myd88-/-妊娠母鼠与无菌wt妊娠母鼠共同圈养时,在断奶时分开的所得定殖幼崽不会传播肥胖表型(图15a)。因此,测试了t-myd88-/-小鼠的免疫缺陷是否是致胖微生物群持续存在的必要条件。相比之下,在完全完整的免疫系统存在的情况下,微生物群得到了适当的控制。缺乏内源性t细胞的tcrb
‑′‑
小鼠通过广谱抗生素治疗耗尽内源性微生物群,然后在过继转移wt或t-myd88
‑′‑
cd4
t之前用wt和t-myd88-/-微生物群的1∶1混合物定殖(图15b)。将小鼠分开到单独圈养笼中,以便微生物群形成不会受到笼子内其他动物的影响,并且每个微生物群落都将独立地形成。尽管这些小鼠最初定殖了相同的微生物群,但与给予wt cd4
t细胞的tcrb
‑′‑
小鼠相比,给予t-myd88-/-cd4
t细胞的tcrb
‑′‑
小鼠体重显著增加(图5a)。因此,t细胞内myd88信号传导的缺陷驱动动物的代谢缺陷。tcrb
‑′‑
小鼠体内10%的细菌被iga包被,从而证明t细胞对于iga靶向微生物群的重要性(图3b)。然而,在t细胞转移后1周,给予wt t细胞的小鼠显示iga结合微生物增加三倍(图15c)。针对微生物群的igg1或igg3反应需要更长的时间才能形成,但在t细胞转移后8周可检测到(图15d至f)。尽管在这种实验环境中,动物的总iga水平相似,但在接受敲除t细胞的动物中,iga和igg1与微生物群的结合存在缺陷(图5b和图15e和g)。在接受t-myd88-/-t细胞的tcrb
‑′‑
小鼠中,igg3对微生物群的靶向被认为不受t细胞非依赖性机制的控制(图15h)(m.a.koch等人,cell 165,827-841(2016))。随时间推移,微生物群组成在基因型之间的差异增加(图5c)。此外,接受来自致胖小鼠的t细胞的动物群落中的群落变化与在t-myd88-/-动物中观察到的那些相似。实际上,两种基因型中脱硫弧菌科和梭菌科的丰度之间存在显著负相关。
[0163]
尽管从使用相同的微生物群混合物开始,但接受t-myd88-/-t细胞的动物最终定殖显著更少的梭菌科(图5d和e)。属水平的三个分类群被iga不同地靶向,包括梭菌纲的颤螺菌属,而大多数梭菌纲的属在这种分类学分辨率水平上是高度可变的(图16a)。iga结合指数在更精细的otu水平(97%相似性)进行评估,并且发现在接受t-myd88-/-t细胞的动物中富集由iga不同靶向的梭菌纲分类的otu(图5f)。观察到脱硫弧菌属的iga靶向的趋势增加(图16a和b)。因此,梭菌纲及其功能贡献的减少可能是由iga的不适当靶向和脱硫弧菌属的扩增的组合引起的。
[0164]
为了支持抗体反应影响代谢缺陷的假设,将致胖微生物群转移至抗生素处理的wt或rag1
‑′‑
动物中。事实上,致胖微生物群转移至wt小鼠并没有赋予表型,而转移至缺乏抗体的rag1
‑′‑
动物中与接受wt微生物群的动物相比体重增加显著更大(图5g和图15i)。tfh是t细胞,其功能是在生发中心的b细胞内指导抗体类别转换和突变。先前已确定t-myd88-/-小鼠的t细胞的发育缺陷位于tfh细胞内。
[0165]
与接受wt t细胞的动物相比,接受bcl6
‑′‑
t细胞(其不能分化成tfh细胞)的t-myd88-/-动物的体重显著更高(图5h)(s.crotty,annurev immunol 29,621-663(2011))。因此,不具有发育成tfh细胞的能力的t细胞无法拯救肥胖症表型。因此,需要适当的tfh细胞
功能来调控微生物群以预防肥胖症。
[0166]
短链脂肪酸(scfa)是经过充分研究的微生物群依赖性机制,其影响宿主代谢。然而,在wt与t-myd88-/-动物之间scfa产量没有差异(图17a)。还提出了增加肠道渗透性和细菌产物渗漏,所述细菌产物在脂肪组织内诱导低度炎症(f.e.rey等人,proc natl acadsci u s a 110,13582-13587(2013);和p.d.cani等人,diabetes,56,1761-1772(2007))。然而,未检测到t-myd88-/-动物血清中细菌配体的差异。此外,将t-myd88-/-置于注入抗炎5-asa的饮食(h.luck等人,cell metab 21,527-542(2015))未能拯救体重增加(图17b)。肝脏rna-seq和基因集富集分析(gsea)揭示,尽管动物被喂食标准的小鼠食物,但参与脂质代谢的途径(包括甘油磷脂和甘油脂代谢)是与wt对照相比,t-myd88-/-动物中最显著富集的途径(图6a)。特别地,合成脂质所需的基因(包括fasn、dgat2和srebpf1)以及参与脂质吸收的基因(包括slc27a4和cd36)的表达在t-myd88-/-动物的肝脏内高度上调(图6b)。尽管cd36在t-myd88-/-动物中上调,但抗生素治疗显著下调cd36表达(图6c)。此外,肥胖t-myd88-/-动物的梭菌纲治疗产生了cd36的显著下调,从而表明梭菌纲具有减少脂质摄取的功能(图6d)。事实上,与无菌小鼠相比,限菌动物定殖有在肝脏cd36表达中显著降低的梭菌纲聚生体(图6e)。因此,t-myd88-/-中的脂质摄取似乎是微生物群依赖性方式。
[0167]
梭菌纲定殖于无菌动物显著下调小肠内的cd36和fasn(图6f和g),从而表明梭菌纲影响肠道内的脂质吸收和代谢。此外,从培养的梭菌纲聚生体收集的无细胞上清液(cfs)显著下调培养的肠上皮细胞(iec)中的cd36(图6h)。相比之下,从培养的脱硫弧菌属物种中收集的cfs直接提高了iec上cd36的表达(图6h)。此外,与脱硫弧菌属或限菌动物单一相关的动物相比,与梭菌纲聚生体相关的无菌动物的体脂百分比显著降低(图6i)。值得注意的是,将脱硫弧菌属添加至仅定殖有梭菌纲聚生体的无菌小鼠中导致体脂百分比和小肠中的cd36表达增加(图6j和k)。因此,微生物群可直接调控肠道上皮细胞内的脂质代谢。
[0168]
支持增加的脂质吸收,hfd喂养的t-myd88-/-在盲肠内的几种长链脂肪酸(lcfa)显著减少,并且血清中随之增加(图6l和m)。管腔脂质谱和16s测序的比较揭示了脱硫弧菌属与梭菌纲的成员之间的相反相关性以及lcfa和其他脂质的丰度。盲肠内容物中lcfa的耗减与脱硫弧菌属的存在显著相关。相比之下,梭菌纲的多个成员(包括smb53和多尔氏菌属)与lcfa累积相关(图17c),从而进一步支持了微生物组成可调控脂质吸收的假设。因此,患有肥胖症和t2d的个体中观察到的特定梭菌纲物种的损失可导致肠道吸收和脂肪代谢增加,从而突出显示适当的微生物群组成对健康的重要性。
[0169]
论述。微生物群一直牵涉于多种自身免疫性和代谢疾患中。然而,这些疾病并不总是与获得病原体生物体相关,并且相反,已提出有益物种的损失是致病因素(i.cho等人,nature 488,621-626(2012))。导致有益细菌损失的机制可包括抗生素使用、环境卫生增加和低纤维饮食(n.m.koropatkin,e.a.cameron,e.c.martens,nat rev microbiol 10,323-335(2012))。本文所述的结果表明,维持健康微生物群落的另一种机制是通过对肠道内这些群体的适当免疫控制。t-myd88-/-动物内形成的微生物群反映了在患有ii型糖尿病和肥胖症的个体中观察到的生态失调,包括脱硫弧菌属的扩增和梭菌纲的损失(j.qin等人,nature 490,55-60(2012))。尽管缺乏全面的人体研究,但据报告患有肥胖症和ii型糖尿病的个体的黏膜iga较低,并且对免疫接种的反应降低。这表明这些个体对肠道微生物群的免疫反应欠佳,但并非完全缺陷,其结合膳食性缺乏导致代谢疾病。这些数据表明,微生物群
的t细胞依赖性靶向对于维持健康群落是重要的。尽管细菌的iga结合通常被认为导致其根除,但iga可调控某些细菌的功能性基因表达,且甚至有助于某些共生体的粘膜缔合(g.p.donaldson等人,science 360,795-800(2018);t.c.cullender等人,cell host microbe 14,571-581(2013);和d.a.peterson,n等人,cell host microbe 2,328-339(2007))。事实上,结果表明,尽管iga在t-myd88-/-动物中的水平较低,但脱硫弧菌属和几种梭菌纲物种显示iga包被增加。因此,iga对梭菌纲的不适当靶向可减少其定殖或改变其代谢功能以影响肥胖症的发展。
[0170]
此外,iga较少靶向几种梭菌纲。令人感兴趣地,最近对患有iga缺乏症的个体内微生物群的评估显示,几种梭菌纲的定殖显著减少(j.fadlallah等人,sci transl med 10,(2018))。因此,iga还可起到增强一些梭菌纲物种定殖的作用,如对于脆弱拟杆菌所显示的那样(g.p.donaldson等人,science 360,795-800(2018))。脱硫弧菌属在此模型和患有代谢综合征的个体中扩增的机制仍不清楚。
[0171]
然而,本文所述的结果表明,这种扩增可直接影响特定梭菌纲成员的定殖,尽管这种情况如何发生仍然是神秘的。了解iga靶向肠道微生物如何影响它们在无菌环境中的定殖和功能可有助于深入了解免疫系统如何影响这种微生物关系。随着梭菌纲的成员越来越多地在多种环境中得到识别(24),确定其他微生物的定殖和免疫系统如何共同影响梭菌纲的功能将是重要的。
[0172]
cd36是肠道内脂质吸收的重要调控因子,并且其缺乏导致在hfd喂养时抵抗肥胖症和代谢综合征的发展(m.buttet等人,plos one 11,e0145626(2016);和m.buttet等人,biochimie 96,37-47(2014))。人肝脏内cd36的表达增加与脂肪性肝病相关。此外,具有cd36多态性的个体(其在肠道内的表达仅降低两倍)对代谢疾病具有抗性(l.love-gregory,n.a.abumrad,curr opin clin nutr metab care 14,527-534(2011))。因此,cd36的相对表达水平对于哺乳动物体内的脂质吸收和体内平衡是重要的。最近的研究表明,微生物群可通过增强cd36表达来上调宿主对肠道内脂质的吸收(y.wang等人,science 357,912-916(2017))。然而,发现细菌也可能能够限制宿主脂质吸收。
[0173]
因此,肠道细菌可差异地调控脂质代谢。事实上,由脱硫弧菌属分泌的产物上调cd36表达,而由梭菌纲产生的产物可下调cd36表达。因此,具有调节cd36表达的功能的生物体的损失可导致脂质的不适当吸收,脂质随着时间推移累积,从而导致肥胖症和代谢综合征。对诸如脱硫弧菌属和梭菌纲的生物体的相互作用的进一步表征以及影响cd36表达的分泌分子的鉴定可为未来的靶向疗法提供信息。
[0174]
材料和方法。小鼠。使c57bl/6 myd88
loxp/loxp
小鼠(jackson laboratories)与c57bl/6 cd4-cre动物(taconic)杂交以产生myd88
/
;cd4-cre
小鼠(wt)和myd88
loxp/loxp
;cd4-cre
(t-myd88-/-)动物。年龄匹配的雄性小鼠用于比较标准饮食下的自发体重表型,包括免疫和微生物群反应。年龄匹配的雄性和雌性小鼠用于比较高脂饮食(hfd)下的体重表型,包括免疫和微生物群反应。为了测量微生物群的t细胞依赖性形成,使用了4周龄的tcrb-/-小鼠(jacksonlaboratories)。为了研究微生物群的脱硫脱硫弧菌依赖性形成,使用了6周龄的wt c57bl/6小鼠(jackson laboratories)或年龄匹配的cd4-cre
(wt)小鼠。为了测量微生物群对免疫缺陷型小鼠的体重增加的影响,使用了4周龄的ragl-/-小鼠(jackson laboratories)。将gf小鼠维持在无菌隔离器中,并且每月通过粪便的涂铺和pcr
验证gf状态。本研究中使用了gf c57bl/6动物。
[0175]
用孢子形成微生物定殖小鼠。从wt小鼠中取出粪便颗粒并在37℃下在厌氧室中在含有3%氯仿(v/v)的还原pbs中孵育1小时。还将含有还原pbs和3%氯仿的对照管在厌氧室中在37℃下孵育1小时。孵育后,轻轻混合试管,并让粪便物质沉降10秒。将上清液转移至新的试管中,并通过将co2压入试管中除去氯仿。对于常规条件下的孢子形成(sf)实验,sf组中的小鼠口服管饲100μl孢子形成粪便部分,并且ctrl组中的小鼠口服管饲100μl pbs对照(其中每隔三天也除去氯仿)。对于与无菌动物的孢子形成关联,将含有管饲材料的管在无菌隔离器的端口中灭菌1小时,然后将它们拉入隔离器进行管饲。然后用100μl的孢子形成混合物对繁殖者对进行口服管饲。他们的后代在8周龄时被处死,用于分析小肠和肝脏。
[0176]
t细胞转移至t-myd88-/-小鼠中。t-myd88-/-小鼠在t细胞转移前一天进行了500雷得的亚致死辐照。使用了来自wt(cd4-cre
)和bcl6ko(bcl6
loxp/loxp cd4-cre
)小鼠的脾脏,并使用cd4
t细胞分离试剂盒(miltenyi)分离t细胞。t-myd88-/-经眶后注射5
×
106个wt或bcl6-/-macs富集的t细胞,并每周称重持续5周。
[0177]
饮食治疗。圈养在spf设施内的动物用标准的辐照2920x食物(envigo)喂养。在hfd实验期间,将小鼠用45kcal%脂肪dio小鼠饲料的高脂肪饮食(研究饮食)或作为对照的10kcal%脂肪dio小鼠饲料的饮食作(研究饮食)喂养。在5-asa炎症实验期间,还将小鼠用含有1%5-asa(envigo)的辐照标准2020食物的定制饮食或缺乏5-asa(envigo)的对照饮食喂养。
[0178]
抗生素治疗。wt和t-myd88-/-小鼠在用hfd喂养时在其饮用水中维持0.5mg/ml氨苄青霉素(fisher scientific)、新霉素(fisher scientific)、红霉素(fisher scientific)和庆大霉素(goldbio)持续14周以确定微生物群对体重增加表型的相对贡献。将tcrb-/-和rag1-/-小鼠置于饮用水中0.5mg/ml的氨苄青霉素(fisher scientific)、新霉素(fisher scientific)、红霉素(fisher scientific)和庆大霉素(goldbio)中持续1周以在被粪便转移重新定殖之前减少内源性微生物群。
[0179]
tcrb-/-小鼠内微生物群的t细胞形成。将三个单独宠子的四只tcrb-/-小鼠置于饮用水中的抗生素混合物持续一周。在进行任何进一步治疗之前,抗生素被除去24小时。将来自wt供体的一个粪便团块和来自t-myd88-/-供体的一个粪便团块在含有0.1%半胱氨酸的还原pbs中捣碎,并立即口服管饲到tcrb-/-小鼠中。这种口服管饲每隔一天重复一次,持续一周。在最后一次管饲后48小时,将小鼠放入单独圈养的笼子中,并用5
×
106个cd4
macs富集的wt或t-myd88-/-细胞进行眶后注射。这被标记为d0。
[0180]
葡葡糖耐量测试。在用葡萄糖激发之前,使小鼠禁食6小时。使用contour血糖仪(bayer)和contour血糖试纸(bayer)检测空腹葡萄糖水平。在零时间点将每克体重一毫克葡萄糖腹膜内注射到动物体内。使用血糖仪和试纸在5分钟、15分钟、30分钟、60分钟和120分钟的时间点测量血糖水平。
[0181]
胰岛素elisa。从禁食6小时的小鼠收集血清,并使用小鼠胰岛素elisa试剂盒(crystal chem)测量胰岛素。根据制造商的说明书,血清样品一式两份运行。
[0182]
胰岛素抵抗测试。在用葡萄糖激发之前,使小鼠禁食6小时。使用contour血糖仪(bayer)和contour血糖试纸(bayer)检测空腹葡萄糖水平。在零时间点将胰岛素(0.75u/kg体重)腹膜内注射到动物体内。使用血糖仪和试纸在5分钟、10分钟、15分钟、20分钟、25分
钟、30分钟、40分钟和60分钟的时间点测量血糖水平。如果血糖水平降至30mg/dl,则在150μl腹膜内注射25%葡萄糖后将动物从实验中移除。
[0183]
使用小鼠肠上皮细胞(mode-k细胞)的体外实验。将小鼠肠上皮细胞维持在含有l-谷氨酰胺和丙酮酸钠的杜尔贝科氏改良的伊格尔氏培养基(dmem)中。dmem补充有10%fbs、1%(v/v)谷氨酰胺、青霉素-链霉素和1%hepes。为了确定细菌是否调控基因表达,将汇合的单层细胞与(1∶1)不含青霉素-链霉素的dmem:从培养的梭菌纲聚生体或脱硫弧菌属物种中收集的cfs一起孵育4小时。然后吸出培养基并将细胞置于600tl ribozol(vwr)中以供以后分析。
[0184]
从小肠、细胞培养物和肝组织中分离rna用于qpcr和rna-seq。将0.5em长或1x105个细胞的组织切片在-70℃下储存在700tl的ribozol(vwr)中。使用direct-zol rna miniprep试剂盒(zymoresearch)分离rna。使用qscript cdna合成试剂盒(quanta biosciences)合成cdna。使用lightcycler480sybr green i master(roche)进行qpcr。qpcr实验在lightcycler lc480仪器(roche)上进行。对于肝脏rna测序,在qc后通过illumina truseq stranded rna sample prep和ribozero处理(人、小鼠、大鼠等)制备rna,并使用illumina hiseq测序进行分析。
[0185]
粪便免疫球蛋白的定量。为了定量管腔iga,将粪便团块收集在1.5ml微量离心管中并称重。将管腔内容物重新悬浮在每毫克粪便重量10tl的无菌1x hbss中,并以100xg旋转5分钟以除去粗粒材料。然后将上清液置于新的1.5ml微量离心管中,以8000xg旋转5分钟以沉淀细菌。
[0186]
然后将上清液(含有iga)置于新的1.5ml微量离心管中,并用作iga特异性elisa试剂盒(ebioscience;根据制造商说明书进行)的样品(1/10和1/100(v/v)稀释液)。在450nm处读取吸光度,并使用标准曲线计算iga浓度。浓度针对粪便重量归一化。
[0187]
将细菌颗粒重新悬浮在500tl的无菌pbs中,并通过以8000xg旋转5分钟洗涤两次。然后将洗涤过的细菌团块重新悬浮在每mg粪便10tl无菌pbs中。将5微升的每种样品涂铺在96孔圆底板上。在室温下用100tl含有10%(v/v)fbs的无菌hbss封闭细菌15分钟。无需洗涤细胞,将100tl的以1∶500稀释在含有10%(v/v)fbs的无菌hbss中的抗iga(ebioscience克隆ma-6e1 pe)、抗igg1(santacruz cruzfluor555)或抗igg3(santacruz cruzfluor555)添加至孔中。将孔在4℃下孵育30分钟。通过以2500x g旋转5分钟将板洗涤两次,然后除去上清液并将细胞重新悬浮在无菌hbss中。最后一次洗涤后,将细菌孔重新悬浮在250tl的含有5tl的1x sybr绿色染色剂(invitrogen目录号s7563)的hbss中。将孔在4℃下孵育20分钟,然后立即在流式细胞仪上计数。rag1-/-粪便团块作为阴性对照包括在所有实验中。
[0188]
脱硫脱硫弧菌atcc 27774和desulfovibrio piger atcc 29098的生长。细菌物种脱硫脱硫弧菌购自atcc(#27774)。细菌物种desulfovibrio piger购自atcc(#29098)。根据厌氧细菌的atcc说明处理和打开小瓶,并将细胞在之前描述的脱硫弧菌属培养基中生长(f.e.rey等人,proc natl acad sci u s a 110,13582-13587(2013))。培养基由nh4cl(1g/l)(fisher chemical)、na2so4(2g/l)(fisher chemical)、na2s2o3
·
5h2o(1g/l)(sigma)、mgso4
·
7h2o(1g/l)(fisher chemical)、cacl2
·
2h2o(0.1g/l)(fisher chemical)、kh2po4(0.5g/l)(fisher bioreagents)、酵母提取物(1g/l)(amresco)、刃天青(0.5ml/l)(sigma)、半胱氨酸(0.6g/l)(sigma)、dtt(0.6g/l)(sigma)、nahco3(1g/l)
(fisher chemical)、丙酮酸(3g/l)(acros organics)、苹果酸(3g/l)(acros organics)、atcc微量矿物质混合物(10ml/l)、atcc维生素混合物(10ml/l)组成并调节至ph 7.2。使细菌在厌氧室(coy labs)中生长48小时,并在70℃下储存在含有25%甘油的生长培养基中。将2.5
×
108个细菌cfu添加至250μl小鼠饮用水中,持续1周。
[0189]
粪便、回肠和iga结合的微生物dna的分离和16s测序。处死动物,并且取出它们的整个下消化道(从十二指肠至直肠)并纵向切片。从每只动物收集一个粪便团块和来自小肠下部10cm的管腔内容物,以分别表征粪便和回肠微生物群落。将粪便和回肠样品立即在-70℃下冷冻于含有约250mg的0.15mm石榴石珠(mobio,目录号13122-500)的2ml螺旋盖管中。根据制造商的说明书,使用power粪便dna分离试剂盒(mobio)提取dna。将来自t细胞转移实验的iga结合和未结合的细菌从盲肠内容物中分离出来,并在加工前在-70℃下冷冻。使用配对末端300循环miseq读数进行iga结合的细菌分离、16s rdna扩增、测序和序列加工(j.l.kubinak等人,cell host microbe 17,153-163(2015))。计算了iga指数(a.l.kau等人,sci transl med 7,276ra224(2015))。
[0190]
宏转录组学。将粪便团块或回肠腔内容物直接放入trizol中并储存在-20℃直至提取rna。使用direct-zol(zymo research,#r2052)从样品提取总rna,然后制备以供犹他大学高通量基因组学核心设施使用ribo-zero gold rrna(流行病学)除去试剂盒(illumina,#mrze724)进行illumina测序。illumina文库在hiseq 2500上进行多路复用和测序,单端50个循环测序。humann2(v0.9.9)分析框架用于后续的测序处理和数据分析(s.abubucker等人,plos comput biol 8,e1002358(2012))。首先,使用在humann2中实现的knead数据脚本,使用trimmomatic(a.m.bolger,m.lohse,b.usadel,bioinformatics 30,2114-2120(2014))对原始序列进行质量修剪和过滤,然后使用bowtie2(b.langmead,s.l.salzberg,nat methods 9,357-359(2012)过滤以除去针对小家鼠基因组版本grcm38的宿主读数。在基因型之间没有观察到质量过滤读数的显著差异,尽管在整个样品中,来自回肠样品的更多读数映射至小鼠基因,提供了更少的细菌转录物覆盖。然后,为了改进这些短读数的映射,质量过滤读数的映射仅限于具有uniref90基因注释的小鼠分离细菌参考基因组的自定义数据库。此自定义数据库包括最近作为小鼠肠道细菌收集(mibc)的一部分分离和测序的53种生物体(i.lagkouvardos等人,nat microbiol 1,16131(2016)),以及humman2’的chocophlan数据库中包括的9个参考基因组,代表在16s测序中检测到但尚未包括在mibc集合中的物种。这九个基因组是:假长双歧杆菌、动物双歧杆菌、长双歧杆菌、脆弱拟杆菌、柴氏粘螺菌(mucispirillum schaedleri)、罗伊氏乳杆菌、产气荚膜梭菌、脱硫脱硫弧菌和分节丝状菌。为了创建具有uniref90注释的自定义数据库,使用diamond aligner(b.buchfink,c.xie,d.h.huson,nat methods 12,59-60(2015))将来自mibc基因组的氨基酸序列与uniref90数据库进行比对并且需要50%的查询覆盖率和90%同一性。然后,这些uniref90注释的mibc氨基酸序列用于注释每种相应基因的核苷酸序列,并与已注释的九个基因组相结合,以创建含义小鼠特异性细菌基因组的自定义核苷酸作图参考。为了使用humann2将过滤后的序列读数映射至自定义参考,由于读取长度短,使用了核苷酸比对(无翻译比对)。然后,将来自humann2的uniref90基因家族输出的每千碱基比对读数的计数针对每百万计数(样品内)归一化,或重新分组为基因本体论(go)项,然后归一化,用于后续分析。
[0191]
代谢表型分析。在nmr bruker minispec上测量总体脂组成。clams代谢笼用于测量间接量热法。能量消耗(ee)使用以下公式计算。热值(cv)=3.815 (1.232*rer)。ee=cv*vo2。
[0192]
肝脏和脂肪组织显微术。将肝脏和脂肪组织用福尔马林固定,蜡包埋,并进行苏木精和伊红染色。使用来自thermofisher的evos core xl成像系统收集显微镜图像。
[0193]
血清和盲肠内容物代谢组学(不包括scfa测量)。样品提取和制备。盲肠内容物在分析前储存在-70℃。将5毫升含有内部标准品(每个样品1tg的d4-琥珀酸和5tg标记的氨基酸(
13
c、
15
n标记的)混合物)的75%乙醇溶液添加至每个样品中。将样品剧烈涡旋,然后在沸水中孵育10分钟。将冷却的样品以5,000x g旋转5分钟。将上清液转移至新试管中,然后快速抽真空过夜以干燥。
[0194]
gc-ms分析。gc-ms分析使用配备有agilent 6890气相色谱仪gerstel mps2自动进样器的waters gct premier质谱仪进行。将干燥的样品悬浮在40tl的40mg/ml o-甲氧基胺盐酸盐(mox)的吡啶溶液中,并在30℃下孵育1小时。向自动进样器小瓶中添加25tl的这种溶液。通过自动进样器自动添加40微升40tl的n-甲基-n-三甲基甲硅烷基三氟乙酰胺(mstfa),并在37℃下振荡孵育60分钟。孵育后,通过自动进样器添加3tl的脂肪酸甲酯标准(fames)溶液,然后将1μl制备好的样品以分流模式注入气相色谱仪入口,入口温度保持在250℃。使用10∶1的分流比进行分析。气相色谱仪的初始温度为95℃,持续1分钟,然后以40℃/min升温至110℃,保持时间为2分钟。随后第二次5℃/min升温至250℃,第三次升温至350℃,最后保持3分钟。采用具有5-m长保护柱的30-m phenomex zb5-5msi柱进行色谱分离。1ml/min的氦气用作载气。由于几种代谢物的含量高,所以将样品以十倍稀释度再次分析。
[0195]
gc-ms数据的分析。使用masslynx 4.1软件(waters)收集数据。使用quanlynx鉴定代谢物并记录其峰面积。将此数据传输至excel电子表格(microsoft,redmond wa)。使用纯购买的标准品开发的内部代谢物文库和市售nist文库的组合来建立代谢物身份。并非所有代谢物都使用gc-ms进行观察。这是由于几个原因。例如,一些代谢物的浓度非常低。其次,代谢物可能不适合gc-ms,因为要么太大而无法挥发,要么是诸如肉毒碱的季胺,要么就是不能很好地电离。不能很好电离的代谢物包括草酰乙酸、组氨酸和精氨酸。根据细胞条件观察到半胱氨酸,通常与蛋白质形成二硫键,并且在细胞内浓度较低。
[0196]
盲肠内容物的短链脂肪酸检测。样品提取和制备。将样品从冰箱中取出并在室温下解冻5分钟。向这些样品中添加400tl的dd-h2o、10tl的5-磺基水杨酸(1mg/tl)和2tl的内部标准品(1m新戊酸)。将样品涡旋30秒并在冰上放置30分钟。然后将样品在4℃下以2000x g离心10分钟。然后将上清液转移至含有10tl浓hcl的带有ptfe内衬盖的玻璃小瓶中。接下来,添加3ml乙醚并将样品涡旋30秒,然后在4℃下以1,200x g离心10分钟。然后将上清液转移至带有ptfe内衬盖的新玻璃小瓶中,并用50tl的n-甲基-n-(叔丁基二甲基甲硅烷基)三氟乙酰胺、叔丁基二甲基氯硅烷(mtbstfa;thermo scientific)衍生化。将样品涡旋并置于60℃沙浴中30分钟。使样品冷却至室温并在温和的氮气流下部分蒸发至大约250tl的体积,并转移至玻璃gc-ms小瓶中。
[0197]
gc-ms分析。gc/ms分析在与agilent 5793质谱仪和配备有db-1色谱柱(15m x 0.25-mm内径x 0.25-μm薄膜厚度;j&w scientific,folsom,ca,usa)的agilent 7683
(santa clara,ca,usa)自动注射器耦合的agilent 6890气相色谱仪上进行。使用流速为1.0ml/min的氦气载气。将1-μl样品以10∶1的分流比注入保持在250℃的入口。使用的gc柱烘箱升温为40℃(保持1分钟);以5℃/min升温至70℃(保持3分钟);以20℃/min升温至160℃(保持0分钟);以40℃/min升温至330℃(保持6分钟)。以扫描模式采集数据,质量范围为44-200m/z,使用m/z 117.0(对于乙酸)、m/z 131.0(对于丁酸)、m/z 145.0(对于丙酸)和m/z 159.0(新戊酸)对靶标进行定量。
[0198]
表2.引物。
[0199][0200][0201]
实施例2:四种梭菌纲菌株拯救肥胖症、胰岛素抵抗和炎症性肠病
[0202]
由4种梭菌纲成员组成的制剂可减轻小鼠的肥胖症。已采用各种体外技术来缩群落中有助于减轻肥胖症的特定成员的范围。这些实验导致对此群落的4个特定成员的组合进行测试。含有梭菌纲anaerovorax、毛螺菌科某种、梭菌属xiva和梭菌属iv的精制4成员群落(在本文中也称为精制梭菌纲聚生体-rcc-4)。这4种菌株用于定殖于小鼠,并将脂肪累积与更复杂的梭菌纲聚生体(图中称为sf)进行比较。令人感兴趣地,这4种菌株足以将肥胖症(rcc-m)减轻至与复杂微生物群落相同的程度(图18)。
[0203]
替代梭菌纲拯救肥胖症、胰岛素抵抗和炎症性肠病。通过一系列实验,在致胖动物中鉴定了梭菌纲的显著减少。因此,测试了重新引入这些瘦相关微生物是能够预防肥胖症。由于梭菌纲是已知的孢子形成者,因此用氯仿处理粪便以富集这些微生物。将这些动物置于高脂肪饮食(hfd),这已被证明可加速此模型中的体重增加,并更好地模拟西方化的生活方式。用孢子形成细菌的混合物治疗容易肥胖的t-myd88-/-动物显著降低了体重增加和脂肪累积(图19a-b)。在仅仅三个月结束时,与未处理的t-myd88-/-相比,用梭菌纲处理的t-myd88-/-小鼠的体脂百分比较低,并且vat量降低(图19c)。梭菌纲治疗还降低血糖水平并降低胰岛素抵抗(图19d和19e)。重要的是,在置于hfd的wt动物中也观察到梭菌纲治疗对这些代谢参数的改善(图19d和19e;wt),从而支持梭菌纲对mets的改进将与几种不同的mets和tiid模型相关。
[0204]
虽然ibd与糖尿病之间的关系一直存在争议,但现在有几项研究支持这种联系。在对12,601名ibd患者进行的横断面研究中,糖尿病是第三大最常见的合并症。另一项最近对丹麦47,325名患者进行的横断面研究表明,糖尿病与ibd(uc和cd)显著相关。在1200名ibd患者的儿科群组中,uc患者中的糖尿病患病率也高于对照组。2019年使用8070名ibd患者和
40,030名健康对照者的更新研究表明,即使在控制类固醇使用的情况下,患有ibd的个体中的糖尿病发病率也显著更高。最后,从1977年到2014年,由6,028,844名诊断为ibd的个体对没有ibd的个体组成的全国人口群组显示,患有ibd的个体中的糖尿病发病率增加,尤其是在2003年至2014年之间。除了葡萄糖稳态外,据报告ibd患者的脂质代谢也发生了变化,从而支持代谢疾病与ibd之间的联系。这两种疾病的共性包括慢性炎症和对微生物群的干扰,然而,这些疾病之间联系的潜在机制尚不清楚。
[0205]
现在已经有详尽的研究分析了患有糖尿病和ibd的个体与健康对照组相比的微生物群组成,并且出现了一些相似之处。据报告,在患有糖尿病、肥胖症和ibd的个体中微生物群的多样性降低,伴随梭菌目、瘤胃球菌科和毛螺菌科的成员特异性耗减。虽然系统发育水平的微生物群组成在个体之间通常是可变的,但微生物群的功能能力相当稳定。因此,宏基因组研究可提供更多关于微生物群对疾病的贡献的见解。ii型糖尿病和ibd中的宏基因组研究揭示了短链脂肪酸(scfa)的产生减少,这与梭菌纲成员的减少是一致的。除了特定生物体亚群的损失外,在这些病症中还鉴定了类似的致病有机体。对350多名患有tiid的个体进行研究并将微生物群的组成与健康个体的微生物组成进行了比较的研究鉴定与tiid相关的最显著变化是硫酸盐还原生物体脱硫弧菌属的富集。另一项独立研究鉴定了也患有tiid的免疫受损个体中的脱硫弧菌属过度生长。在ibd中,许多研究指出脱硫弧菌属的增加。美沙拉嗪(其是ibd的常见治疗)抑制粪便硫化物,但在未服用此药物的患者中,硫化物含量较高。宏基因组学研究证实了ibd和ii型糖尿病的系统发育研究,并表明参与含硫氨基酸半胱氨酸代谢的基因在患有疾病的个体中增加。因此,在患有ibd和糖尿病的个体中鉴定了类似的微生物群组成变化,从而表明这些共性可能是这些疾病发展的基础。
[0206]
基于糖尿病与ibd之间的联系,测试了这些细菌是否能够拯救或保护ibd小鼠模型。使用葡聚糖硫酸钠(dss)结肠炎的慢性模型,其中dss在饮用水中提供5天,然后是10天的常规水,并重复两个额外的循环。每隔一天口服管饲梭菌纲或pbs并进行组织学检查。事实上,如通过增加的结肠长度和降低的组织病理学评分所确定,用梭菌纲治疗的动物受显著保护免于结肠炎的发展(图19f、19g)。因此,梭菌纲可预防代谢综合征(mets)和ibd的发展。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。