谷氨酸脱氢酶变体及其在制备l-氨基酸中的应用
技术领域
1.本发明涉及生物技术领域,尤其涉及一种谷氨酸脱氢酶变体及其在制备l-氨基酸中的应用。
背景技术:
2.氨基酸(amino acid)是一类含有氨基和羧基的有机化合物的统称。其中,α-氨基酸是生物功能大分子蛋白质的基本组成单位,构成了蛋白质分子的基本骨架(一级结构)。目前研究发现有22种氨基酸可以构成蛋白质,包括1986年发现的硒半胱氨酸(selenocysteine,sec)及2002年报道的吡咯赖氨酸(pyrrolysine,pyl),其均为α-氨基酸,通常被称为蛋白质氨基酸。在这22种蛋白质氨基酸之外的氨基酸则被称为非蛋白质氨基酸(non-proteinogenic amino acid)。
3.蛋白质氨基酸可运用于医药、食品、饲料等与人类生活密切相关的行业,例如作为营养品维持人体的氮平衡,作为食品的风味添加剂,作为饲料添加剂用于养殖行业等。而非蛋白质氨基酸,由于其结构和功能的多样性,在化工、医药、农药等领域有着更广泛的用途,例如苯基异丝氨酸(β-氨基酸,抗癌药紫杉醇的侧链部分)、l-正缬氨酸(降血压药培哚普利中间体)、l-高苯丙氨酸(降血压药依那普利中间体)、l-草铵膦(广谱灭生性除草剂)等。
4.蛋白质氨基酸通常通过发酵法或从廉价原料中提取等具有成本效益的方法进行生产,但酶法生产同样具有其优势,例如通过酶法能获得更高浓度的产品,且后续分离工艺相对简单。在非蛋白质氨基酸方面,与蛋白质氨基酸不同,其尚不能通过发酵法进行工业化生产,现在主要依靠有机合成法进行制备。有机合成法制备手性非蛋白质氨基酸的缺点是反应条件苛刻、产品收率和光学纯度不高、以及过程对环境不友好等,几乎没有相关工业化的报导。生物酶催化法具有反应条件温和、立体选择性高、过程绿色环保等特点,是制备手性非蛋白质氨基酸的潜在优势方法。例如,谷氨酸脱氢酶在辅酶的存在下,能够催化酮酸还原胺化生成对应的氨基酸,且立体选择性严格。用酶法生产可以避免传统化学法的高压、高温、高能耗以及高污染的缺点,是一种符合绿色生产理念的先进制造模式。
5.谷氨酸脱氢酶(glutamate dehydrogenase,ec 1.4.1.2-4)是合成谷氨酸的关键酶,参与谷氨酸的合成与分解代谢,广泛存在于所有生命体中,根据辅酶特异性可分为nad(h),nadp(h)以及双辅酶特异性三类。谷氨酸脱氢酶由于其来源广,立体选择性严格等特点,在手性氨基酸合成领域具有极大的应用价值和潜力。但野生的谷氨酸脱氢酶具有很强的底物专一性,对于除天然底物α-酮戊二酸以外的其他酮酸几乎没有酶活或者酶活极低、无法满足于工业应用。因此,通过分子改造提高野生型谷氨酸脱氢酶对非天然底物的催化能力,有助于其更好的进行工业应用。
6.专利pct/cn2018/105158针对多个来源的谷氨酸脱氢酶,将其底物结合口袋中特定位置的丙氨酸变为甘氨酸和/或缬氨酸变为丙氨酸,提高了多个来源的谷氨酸脱氢酶对l-草铵膦以及其他l-氨基酸的生产能力。其中,突变体lsgludh-a175g对2-羰基-4-(羟基甲基膦酰基)丁酸(ppo)的酶活达34.47u/ml,突变体ppgludh-a167g和ppgludh-v378a对ppo的
酶活分别达到了14.85u/ml和13.3u/ml。但上述谷氨酸脱氢酶均为nadp(h)特异性,在工业生产方面辅酶nadp(h)的价格是辅酶nad(h)的2~3倍,nadp(h)特异性的谷氨酸脱氢酶并不具有生产优势。虽然专利中也报道了部分nad(h)特异性的谷氨酸脱氢酶,但对ppo和其他α-酮酸的催化效率并不理想。
7.鉴于此,还需要进一步深入研究,通过分子改造,来解决nad(h)特异性的谷氨酸脱氢酶对非天然底物没有酶活或酶活较低的问题。
技术实现要素:
8.本发明要解决来源于clostridium difficile的原始谷氨酸脱氢酶(氨基酸序列如seq id no.1所示)对非天然底物没有酶活或酶活较低的问题,因此,提供了多种谷氨酸脱氢酶突变体以及这些突变体在制备不同l-氨基酸中的应用。
9.具体技术方案如下:
10.本发明提供了一种谷氨酸脱氢酶变体,所述谷氨酸脱氢酶变体由seq id no.1所示氨基酸序列的第71位,第92位,第143位,第144位,第145位,第146位,第175位,第187位,第352位,第355位进行单点突变或多点组合突变获得。
11.本发明对来源于clostridium difficile的谷氨酸脱氢酶(cdgludh,ncbi登录号:yp_001086649.1,氨基酸序列如seq id no.1所示,核苷酸序列如seq id no.2所示)进行同源建模,用天然底物α-酮戊二酸进行分子对接,选取α-酮戊二酸侧链基团范围内的10个关键氨基酸残基,将其突变为丙氨酸或更小的甘氨酸,以扩大其口袋体积或改变口袋静电属性,通过单点突变、基于位点与催化活性中心距离的顺序迭代突变以及优势位点组合突变的方式构建谷氨酸脱氢酶突变库,筛选谷氨酸脱氢酶突变体对催化制备l-草铵膦、l-正缬氨酸、l-高苯丙氨酸3种氨基酸的阳性突变株。
12.最后,针对特定的非天然氨基酸产品进行突变位点的优化;针对l-高苯丙氨酸,对顺序迭代后获得的最优突变体中可能起负作用的突变点进行回复突变,以获得突变点的最佳组合;针对l-草铵膦,对底物ppo有显著催化效果的突变点进行迭代饱和突变,以筛选得到最适合的组合突变体。
13.进一步地,所述谷氨酸脱氢酶变体由单点突变获得,各突变位点及其突变前后的氨基酸单字母简写分别为:k71a、m92a、v143a、p144a、a145g、p146a、t175a、r187a、v352a、s355a。
14.其中,k71a表示:第71位的氨基酸由赖氨酸突变为丙氨酸;m92a表示:第92位的氨基酸由甲硫氨酸突变为丙氨酸;v143a表示:第143位的氨基酸由缬氨酸突变为丙氨酸;p144a表示:第144位的氨基酸由脯氨酸突变为丙氨酸;a145g表示:第145位的氨基酸由丙氨酸突变为甘氨酸;p146a表示:第146位的氨基酸由脯氨酸突变为丙氨酸;t175a表示:第175位的氨基酸由苏氨酸突变为丙氨酸;r187a表示:第187位的氨基酸由精氨酸突变为丙氨酸;v352a表示:第352位的氨基酸由缬氨酸突变为丙氨酸;s355a表示:第355位的氨基酸由丝氨酸突变为丙氨酸。
15.进一步地,所述谷氨酸脱氢酶变体由多点组合突变获得,多点组合突变形式为以下形式中的一种:
16.(1)按照第145位,第355位,第352位,第71位,第187位,第175位,第92位,第146位,
第144位,第143位的排列顺序,依次由相邻的两个或两个以上位点进行顺序迭代突变获得;
17.其中,各突变位点及其突变前后的氨基酸单字母简写分别为:a145g、s355a、v352a、k71a、r187a、t175a、m92a、p146a、p144a、v143a;
18.(2)由第71位,第145位,第143位,第144位中的两个或两个以上位点进行组合突变获得;
19.其中,各突变位点及其突变前后的氨基酸单字母简写分别为:k71a、v143a、v143c、v143g、v143i、v143m、v143s、p144m、p144g、p144a。
20.具体的,v143c表示:第143位的氨基酸由缬氨酸突变为半胱氨酸;v143g表示:第143位的氨基酸由缬氨酸突变为甘氨酸;v143i表示:第143位的氨基酸由缬氨酸突变为异亮氨酸;v143m表示:第143位的氨基酸由缬氨酸突变为硫氨酸;v143s表示:第143位的氨基酸由缬氨酸突变为丝氨酸;p144m表示:第144位的氨基酸由脯氨酸突变为甲硫氨酸;p144g表示:第144位的氨基酸由脯氨酸突变为甘氨酸。
21.更进一步地,所述谷氨酸脱氢酶变体为以下多点突变中的一种:
22.(i)a145g/s355a,a145g/s355a/v352a,a145g/s355a/v352a/k71a,a145g/s355a/v352a/k71a/r187a,a145g/s355a/v352a/k71a/r187a/t175a,a145g/s355a/v352a/k71a/r187a/t175a/m92a,a145g/s355a/v352a/k71a/r187a/t175a/m92a/p146a,a145g/s355a/v352a/k71a/r187a/t175a/m92a/p146a/p144a,a145g/s355a/v352a/k71a/r187a/t175a/m92a/p146a/p144a/v143a;
23.(ii)a145g/k71a,a145g/p144a,a145g/v143a,k71a/p144a,k71a/v143a,v143a/p144a,a145g/k71a/p144a,a145g/k71a/v143a,a145g/v143a/p144a,k71a/v143a/p144a,a145g/k71a/v143a/p144a;
24.(iii)s355a/v352a/k71a/r187a/t175a,a145g/v352a/k71a/r187a/t175a,a145g/s355a/k71a/r187a/t175a,v352a/k71a/r187a/t175a,s355a/k71a/r187a/t175a,a145g/k71a/r187a/t175a,k71a/r187a/t175a;
25.(iv)a145g/v143a,a145g/v143c,a145g/v143g,a145g/v143i,a145g/v143m,a145g/v143s,a145g/v143a/p144m,a145g/v143a/p144g,a145g/v143a/p144a。
26.上文中的“/”表示“且,”即“/”前后两个位点同时突变;例如:a145g/s355a表示第145位丙氨酸突变为甘氨酸,且第355位的氨基酸由丝氨酸突变为丙氨酸;a145g/s355a/v352a/k71a/r187a表示第145位丙氨酸突变为甘氨酸,第355位的氨基酸由丝氨酸突变为丙氨酸;第352位的氨基酸由缬氨酸突变为丙氨酸;第71位的氨基酸由赖氨酸突变为丙氨酸;第187位的氨基酸由精氨酸突变为丙氨酸。
27.本发明还提供如上任一项所述的谷氨酸脱氢酶变体的编码基因。
28.本发明还提供了包含如上所述的编码基因的表达载体。作为优选,原始表达载体为pet-28a( )。
29.本发明还提供了包含如上所述的编码基因的基因工程菌。作为优选,基因工程菌的宿主细胞为e.coli bl21(de3)。
30.本发明还提供了如上所述的谷氨酸脱氢酶变体在制备2-氨基戊酸、l-草铵膦或l-高苯丙氨酸中的应用。
31.本发明还提供了如上所述的基因工程菌在制备2-氨基戊酸、l-草铵膦或l-高苯丙
氨酸中的应用。
32.本发明还提供了一种制备l-正缬氨酸的方法,包括:以2-氧代戊酸为底物,在氨基供体及辅酶再生系统的作用下,利用生物催化剂在缓冲溶液中进行催化反应,得到l-正缬氨酸(即2-氨基戊酸);
33.所述生物催化剂为谷氨酸脱氢酶变体或其固定化酶,或者包含编码所述谷氨酸脱氢酶变体基因的基因工程菌;所述辅酶再生系统所采用的辅酶为醇脱氢酶、葡萄糖脱氢酶或甲酸脱氢酶;所述还原型辅酶为nadh(氧化型辅酶nad
);
34.所述谷氨酸脱氢酶变体由seq id no.1所示氨基酸序列进行单点突变或多点组合突变获得,具体为以下突变中的一种:
35.(a)k71a、m92a、v143a、p144a、a145g、p146a、t175a、r187a、v352a、s355a;
36.(b)a145g/s355a,a145g/s355a/v352a,a145g/s355a/v352a/k71a,a145g/s355a/v352a/k71a/r187a,a145g/s355a/v352a/k71a/r187a/t175a,a145g/s355a/v352a/k71a/r187a/t175a/m92a,a145g/s355a/v352a/k71a/r187a/t175a/m92a/p146a,a145g/s355a/v352a/k71a/r187a/t175a/m92a/p146a/p144a,a145g/s355a/v352a/k71a/r187a/t175a/m92a/p146a/p144a/v143a;
37.(c)a145g/k71a,a145g/p144a,a145g/v143a,k71a/p144a,k71a/v143a,v143a/p144a,a145g/k71a/p144a,a145g/k71a/v143a,a145g/v143a/p144a,k71a/v143a/p144a,a145g/k71a/v143a/p144a。
38.本发明还提供了一种制备l-草铵膦的方法,包括:以2-羰基-4-(羟基甲基膦酰基)丁酸酸为底物,在氨基供体及辅酶再生系统的作用下,利用生物催化剂在缓冲溶液中进行催化反应,得到l-草铵膦;
39.所述生物催化剂为谷氨酸脱氢酶变体或其固定化酶,或者包含编码所述谷氨酸脱氢酶变体基因的基因工程菌;所述辅酶再生系统所采用的辅酶为醇脱氢酶、葡萄糖脱氢酶或甲酸脱氢酶;所述还原型辅酶为nadh(氧化型辅酶nad
);
40.所述谷氨酸脱氢酶变体由seq id no.1所示氨基酸序列进行单点突变或多点组合突变获得,具体为以下突变中的一种:a145g,a145g/s355a,a145g/s355a/v352a,a145g/p144a,a145g/v143a,a145g/v143a/p144a,a145g/v143a,a145g/v143c,a145g/v143g,a145g/v143i,a145g/v143m,a145g/v143s,a145g/v143a/p144m,a145g/v143a/p144g,a145g/v143a/p144a。
41.本发明还提供了一种制备l-高苯丙氨酸的方法,包括:以2-氧代-4-苯基丁酸为底物,在氨基供体及辅酶再生系统的作用下,利用生物催化剂在缓冲溶液中进行催化反应,得到l-高苯丙氨酸;
42.所述生物催化剂为谷氨酸脱氢酶变体或其固定化酶,或者包含编码所述谷氨酸脱氢酶变体基因的基因工程菌;所述辅酶再生系统所采用的辅酶为醇脱氢酶、葡萄糖脱氢酶或甲酸脱氢酶;所述还原型辅酶为nadh(氧化型辅酶nad
);
43.所述谷氨酸脱氢酶变体由seq id no.1所示氨基酸序列进行单点突变或多点组合突变获得,具体为以下突变中的一种:
44.(i)v143a,a145g;
45.(ii)a145g/s355a,a145g/s355a/v352a/k71a,a145g/s355a/v352a/k71a/r187a,
a145g/s355a/v352a/k71a/r187a/t175a,a145g/s355a/v352a/k71a/r187a/t175a/m92a,a145g/s355a/v352a/k71a/r187a/t175a/m92a/p146a,a145g/s355a/v352a/k71a/r187a/t175a/m92a/p146a/p144a,a145g/s355a/v352a/k71a/r187a/t175a/m92a/p146a/p144a/v143a;
46.(iii)a145g/v352a/k71a/r187a/t175a,a145g/s355a/k71a/r187a/t175a,a145g/k71a/r187a/t175a;a145g/k71a,a145g/v143a,v143a/p144a,a145g/k71a/p144a,a145g/k71a/v143a,a145g/v143a/p144a,a145g/k71a/v143a/p144a。
47.与现有技术相比,本发明具有以下有益效果:
48.(1)本发明以来源于clostridium difficile的谷氨酸脱氢酶(cdgludh,ncbi登录号:yp_001086649.1)为基础,通过理性设计的分子改造方法,来解决其对非天然底物没有酶活或酶活较低的问题,获得可以催化制备除天然产物l-谷氨酸以外的多种l-氨基酸的突变体。
49.(2)本发明所使用的理性设计方法能以较小的突变文库、通过筛选快速获得针对非天然底物具有较高催化活力的谷氨酸脱氢酶突变体。
50.(3)本发明以相应的酮酸为底物,在无机氨与辅酶的存在下,通过谷氨酸脱氢酶催化还原胺化生成l-氨基酸,工艺简单、原料转化率和产品手性纯度均大于99%。
51.(4)本发明方法以nad(h)为辅酶,相较于nadp(h),其价格低廉,大大降低了l-氨基酸的生产成本。
附图说明
52.图1为d,l-草胺膦标品,l-草胺膦标品,反应样品柱前衍生化高效液相图谱。
53.图2为2-羰基-4-(羟基甲基膦酰基)丁酸(ppo)标品的高效液相图谱。
54.图3为d,l-正缬氨酸标品,l-正缬氨酸标品,反应样品柱前衍生化高效液相图谱。
55.图4为2-氧代戊酸标品的高效液相图谱。
56.图5为d,l-高苯丙氨酸酶标品,l-高苯丙氨酸酶标品,反应样品手性柱高效液相图谱。
57.图6为高苯丙氨酸和2-氧代-4-苯基丁酸标品高效液相图谱。
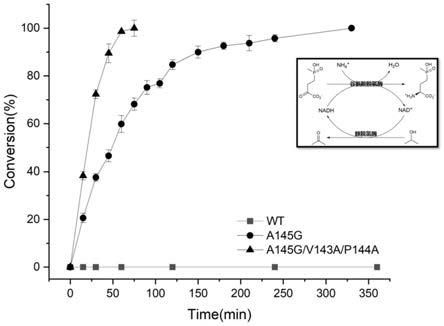
58.图7为实施例8中谷氨酸脱氢酶野生型和突变体不对称合成l-草铵膦的反应式和反应过程曲线。
59.图8为实施例9中谷氨酸脱氢酶野生型和突变体不对称合成l-正缬氨酸的反应式和反应过程曲线。
60.图9为实施例10中谷氨酸脱氢酶野生型和突变体不对称合成l-高苯丙氨酸的反应式和反应过程曲线。
具体实施方式
61.以下结合具体实施例,对本发明做进一步说明。应理解,以下实施例仅用于说明本发明而非用于限制本发明的范围。
62.本发明中的实验方法如无特别说明均为常规方法,基因克隆操作具体可参见j.萨姆布鲁克等编的《分子克隆实验指南》。
63.上游基因工程所用试剂:本发明实施例中使用的dpn i购自takara,宝生物工程(大连)有限公司;质粒提取试剂盒、dna回收纯化试剂盒购自axygen杭州有限公司;e.coli bl21(de3)、质粒pet-28a( )等购自novagen公司;dna marker、低分子量标准蛋白、琼脂糖电泳试剂购自北京全式金生物技术有限公司;引物合成与序列测序工作由擎科生物工程股份有限公司完成。以上试剂使用方法参考商品说明书。
64.下游催化工艺所用试剂:ppo和l-草铵膦为实验室合成;2-氧代戊酸购自tokyo chemical industry,2-氧代-4-苯基丁酸购自上海毕得医药科技股份有限公司,l-正缬氨酸、d-正缬氨酸、l-高苯丙氨酸、d-高苯丙氨酸购自北京百灵威科技有限公司,d,l-草铵膦购自sigma-aldrich公司;α-酮戊二酸、l-谷氨酸、d,l-谷氨酸和其他常用试剂购自国药集团化学试剂有限公司。
65.谷氨酸脱氢酶酶活标准检测体系(hplc法):适量的酶液、80mm底物、40mm nadh、2m nh
4
((nh4)2so4),总体系为500μl,反应介质为ph 7.5 0.1m磷酸盐缓冲液。40℃反应10min,反应结束后加入500μl 1m naoh或1m hcl终止反应。样品中生成的l-氨基酸通过hplc进行定量分析。
66.酶活单位(u)的定义:在标准的反应条件下,每分钟生成1μmol l-氨基酸所需要的酶量。
67.脂肪族氨基酸hplc手性分析方法:采用柱前衍生测定。衍生化试剂:分别称取0.03g邻苯二甲醛与0.1g n-乙酰-l-半胱氨酸,用400μl乙醇助溶,再加入4ml 0.2mol/硼酸缓冲液(ph 9.8),振荡使其充分溶解,4℃冰箱保存备用(不超过4天)。衍生化反应与测定:取100μl样品加入100μl衍生化试剂,混匀后至于25℃保温5min。色谱条件:色谱柱/qs-c18;检测波长/334nm;柱温/30℃;进样量/20μl;流动相a:50mm醋酸钠水溶液,流动相b:乙腈,流动相c:甲醇,流动相配比(v/v)如表1所示。
68.表1 hplc流动相配比
[0069][0070]
高苯丙氨酸hplc手性分析方法:色谱柱/chiralpak zwix(-);检测波长/225nm;流速/0.5ml/min;柱温/40℃;进样量/20μl;流动相/含1.9ml/l甲酸和2ml/l乙二胺的甲醇。
[0071]
ppo的hplc分析方法:色谱柱/qs-c18;检测波长/uv 205nm;柱温/40℃;进样量/20μl;流速/1ml/min;流动相/50mm(nh4)2hpo4水溶液,加入1%的10%四丁基氢氧化铵水溶液,用50%磷酸调ph至3.6,加入8%乙腈。
[0072]
2-氧代戊酸hplc分析方法:色谱柱/qs-c18;检测波长/uv 205nm;柱温/40℃;进样量/20μl;流速/1ml/min;流动相/50mm(nh4)2hpo4水溶液:乙腈=95:5。
[0073]
高苯丙氨酸非手性及2-氧代-4苯基丁酸hplc分析方法:色谱柱/qs-c18;检测波长/uv 205nm;柱温/40℃;进样量/20μl;流速/1ml/min;流动相/50mm(nh4)2hpo4水溶液:乙腈=85:15。
[0074]
实施例1单点突变库的构建与筛选
[0075]
对cdgludh进行k71a、m92a、v143a、p144a、a145g、p146a、t175a、r187a、v352a、s355a单点突变。具体方法如下:
[0076]
1、全质粒pcr
[0077]
以pet-28a( )-cdgludh质粒为模版,设计覆盖突变点的上下游引物(表2)进行全质粒pcr:
[0078]
表2单点突变库构建所用引物
[0079][0080][0081]
pcr扩增体系:
[0082][0083]
pcr扩增条件:
[0084]
1)预变性:95℃5min;
[0085]
2)变性:98℃10s;退火:58℃15s;延伸:72℃10s;共循环30次;
[0086]
3)后延伸:72℃10min;
[0087]
4)4℃保存。
[0088]
2、模板消化:
[0089]
将pcr产物进行琼脂糖凝胶电泳,回收后用dpnι酶消化其中的质粒模板消化体系为:dpnι酶1μl,pcr产物17μl,buffer 2μl。37℃下2小时可完成模板的消化。
[0090]
3、转化及验证:
[0091]
消化后产物经核酸琼脂糖凝胶电泳验证后,采用42℃热激法转化大肠杆菌bl21(de3)感受态细胞。具体过程如下:
[0092]
(1)将感受态细胞放置在冰上解冻15min;
[0093]
(2)在无菌环境下将10μl dna 加入100μl感受态细胞中并轻轻混匀,冰
[0094]
上放置30min;
[0095]
(3)ep管静置于42℃金属浴中热激90s,结束后置于冰上冷却2min;
[0096]
(4)向ep管中加入800μl lb培养基并用枪头混匀,置于200rpm摇床中37℃孵育40-60min;
[0097]
(5)浓缩后取适量体积凃相应的抗性平板,于37℃培养箱中培养12-16h后可出现菌落。每块平板挑出3~4个单菌落进行培养,并测序验证突变是否成功。
[0098]
4、突变菌株培养及蛋白表达
[0099]
将测序成功的突变株经平板划线活化后,挑单菌落接种至含50μg/ml卡那霉素的5ml lb液体培养基中,37℃震荡培养12h。按2%的接种量转接至50ml同样含50μg/ml卡那霉素的lb液体培养基中,37℃震荡培养至od
600
达到0.6~0.8左右时,加入iptg至其终浓度为0.5mm,18℃下诱导培养16h。培养结束后,将培养液12000g4℃离心10min,弃上清,收集菌体。将收集到的菌体,用50mm ph 7.5的磷酸盐缓冲液洗涤两次后,重悬于磷酸盐缓冲液中,400w功率超声破碎30次,每次超声持续3s,间歇7s。将此细胞破碎液12000g 4℃离心10min去除沉淀,得到的上清为粗酶液。
[0100]
5、酶活测定
[0101]
用hplc分析方法测定上述突变体库对天然底物α-酮戊二酸及3种非天然底物的酶活,测定结果见表3:
[0102]
表3谷氨酸脱氢酶及其单点突变体对不同底物的酶活测定
[0103][0104]
注:n/a表示未检测到酶活。
[0105]
实施例2基于位点与催化活性中心距离的顺序迭代突变库的构建与筛选
[0106]
基于与催化活性中心距离的顺序,对71,92,143,144,145,146,175,187,352,355共10个氨基酸残基位点进行顺序迭代突变,具体地,设计以下突变体:
[0107]
2x:a145g/s355a;
[0108]
3x:a145g/s355a/v352a;
[0109]
4x:a145g/s355a/v352a/k71a;
[0110]
5x:a145g/s355a/v352a/k71a/r187a;
[0111]
6x:a145g/s355a/v352a/k71a/r187a/t175a;
[0112]
7x:a145g/s355a/v352a/k71a/r187a/t175a/m92a;
[0113]
8x:a145g/s355a/v352a/k71a/r187a/t175a/m92a/p146a;
[0114]
9x:a145g/s355a/v352a/k71a/r187a/t175a/m92a/p146a/p144a;
[0115]
10x:a145g/s355a/v352a/k71a/r187a/t175a/m92a/p146a /p144a/v143a。
[0116]
具体实验步骤如下:
[0117]
以pet-28a( )-cdgludh-a145g质粒为模版,设计覆盖突变点的上下游引物2x-f和2x-r(表3),进行全质粒pcr获得突变体2x,然后以2x为模板,以3x-f和3x-r为上下游引物进行全质粒pcr获得突变体3x,以此类推获得上述共九个组合突变体,所用引物见表4,详细pcr操作、化转、菌株培养及蛋白表达步骤与实施例1相同。
[0118]
表4顺序迭代突变库构建所用引物
[0119][0120]
用hplc分析方法测定上述突变体库对天然底物α-酮戊二酸及3种非天然底物的酶活,测定结果见表5。
[0121]
表5谷氨酸脱氢酶突变体对天然底物α-酮戊二酸及3种非天然底物的酶活测定
[0122][0123]
注:n/a表示未检测到酶活。
[0124]
实施例3基于阳性单突变点的组合突变库的构建与筛选
[0125]
基于实施例1的筛选结果,将对一个或多个底物有正向催化效果的4个突变体
k71a,v143a,p144a,a145g进行两点,三点以及四点组合,构建新的组合突变库。具体构建方法为以a145g,k71a,v143a质粒为模板,使用相应引物构建a145g/k71a,a145g/p144a等两点突变株,再以两点突变株为模板,使用相应引物构建三点突变株,最后以三点突变株为模板构建四点突变株,所涉及引物见表6,其他详细步骤与实施例1相同。
[0126]
表6阳性单突变点组合突变库构建所用引物
[0127][0128]
用hplc分析方法测定上述突变体库对天然底物α-酮戊二酸及3种非天然底物的酶活,测定结果见表7。
[0129]
表7基于阳性单突变点的组合突变体对不同底物的酶活测定
[0130][0131]
注:n/a表示未检测到酶活。
[0132]
实施例4针对底物2-氧代-4-苯基丁酸的回复突变库的构建和筛选
[0133]
由实施例2的结果可知,对于催化底物2-氧代-4-苯基丁酸制备l-高苯丙氨酸的最优突变为a145g/s355a/v352a/k71a/r187a/t175a(6x),但在顺序迭代过程中发现前3个点位a145g,s355a,v352a对高苯丙氨酸的酶活并没有显著提升,推测这三个突变对提高酶活可能不起效果或起到负向效果,因此从6点突变出发将这三个点做单点或组合的回复突变,共构建6个突变体以筛选到最优的突变点位组合,所构建突变体如下:
[0134]
6x-a:s355a/v352a/k71a/r187a/t175a;
[0135]
6x-b:a145g/v352a/k71a/r187a/t175a;
[0136]
6x-c:a145g/s355a/k71a/r187a/t175a;
[0137]
6x-ab:v352a/k71a/r187a/t175a;
[0138]
6x-ac:s355a/k71a/r187a/t175a;
[0139]
6x-bc:a145g/k71a/r187a/t175a;
[0140]
6x-abc:k71a/r187a/t175a。
[0141]
具体构建方法为以6x或6x-a或6x-b突变体为模板,以表8所示引物进行全质粒pcr,详细步骤见实施例1
[0142]
表8回复突变库构建所用引物
[0143][0144]
用hplc分析方法测定上述突变体对2-氧代-4-苯基丁酸的酶活,测定结果见表9。
[0145]
表9谷氨酸脱氢酶回复突变体对2-氧代-4-苯基丁酸的酶活测定
[0146][0147]
注:n/a表示未检测到酶活。
[0148]
实施例5针对底物ppo的迭代饱和突变库的构建和筛选
[0149]
以ppo为模式底物,对第143,144,145三个位点进行迭代饱和突变,筛选出最优的组合突变体。
[0150]
步骤一:对143位、144位、145位进行单点饱和突变
[0151]
对cdgludh的143位、144位、145位分别进行单点饱和突变。具体方法如下:
[0152]
1.饱和突变库构建
[0153]
以pet-28a( )-cdgludh质粒为模版,设计覆盖突变点的上下游引物(表10)进行全质粒pcr,详细操作见实施例1:
[0154]
表10单点饱和突变库构建所用引物
[0155][0156]
2.初筛
[0157]
在灭菌的96深孔板中加入200μl的lb培养基(含50μg/ml的卡那霉素),使用灭菌枪头挑取单菌落至96深孔板。然后将深孔板置于37℃,200rpm培养8h,称之为一级板。在另外的灭菌的96孔板中加入400μl的lb培养基(含50μg/ml的卡那霉素)作为二级板,在一级板中吸取50μl菌液至二级板,一级板加入20%的甘油放置于-80℃冰箱长期保藏。然后将二级板置于37℃进行震荡培养3h,加入终浓度为0.5mm的iptg诱导,然后将二级板置于18℃,200rpm继续培养18h。
[0158]
二级板在4000rpm、4℃下离心20min收集细胞,然后置于-80℃进行冷冻3h以上。将二级板从-80℃取出,室温下放置0.5h解冻,之后在每孔加入300μl溶菌液(10mm ph7.5磷酸盐缓冲液,750mg/l溶菌酶,10mg/l dnaseⅰ),震荡使细胞悬浮,并置于37℃摇床,200rpm孵育1h。孵育结束后,4000rpm、4℃离心20min,取上清进行酶活测定。
[0159]
配制筛选用酶活测定溶液:ph7.5磷酸盐缓冲液(0.1m),包含2mm nadh,10mm底物ppo,1m nh
4
。向新的96孔板(反应板)各孔中加入200μl酶活测定溶液,并置于37℃保温15min。吸取200μl酶液加入反应板中开始反应,分别于20min,40min和60min取样100μl加入到已经预先加好100μl ph7.5磷酸缓冲液(0.1m)的酶标板中,用酶标仪测定340nm处吸光值。吸光值越低代表其催化活力越高,挑选吸光值较对照(野生型)显著降低的突变株作为候选菌株进行复筛。
[0160]
3.复筛
[0161]
利用hplc对初筛表现出酶活显著提高的突变体进行复筛。将一级板上相应的突变株经平板划线活化后,挑单菌落接种至含50μg/ml卡那霉素的5ml lb液体培养基中,37℃震荡培养12h。按2%的接种量转接至50ml同样含50μg/ml卡那霉素的lb液体培养基中,37℃震荡培养至od
600
达到0.6~0.8左右时,加入iptg至其终浓度为0.5mm,18℃下诱导培养16h。培养结束后,将培养液12000g、4℃离心10min,弃上清,收集细胞。将收集到的细胞,用50mm ph7.5的磷酸盐缓冲液洗涤两次后,重悬于磷酸盐缓冲液中,400w功率超声破碎30次,每次超声持续3s,间歇7s。将此细胞破碎液12000g、4℃离心10min去除沉淀,得到的上清为粗酶液。按照标准酶活检测体系对各突变株的粗酶液酶活进行测定(以ppo为底物)。最终筛选出a145g突变体酶活显著提高,酶活达到1.06
±
0.04u/mg干细胞重。
[0162]
步骤二:在a145g基础上对143位点进行迭代饱和突变
[0163]
对cdgludh-a145g的143位进行迭代饱和突变。引物设计如下:
[0164]
a145g-143-f:agttgatnnkccggggccggatgtgaatacca
[0165]
a145g-143-r:gccccggmnnatcaactttttcgccaatcagtt
[0166]
按步骤一的方法进行突变库的构建和筛选,最终得到6个酶活显著提高的突变体,结果如表11所示。
[0167]
表11 a145g/143位迭代饱和突变库筛选结果
[0168][0169]
步骤三:在a145g/v143a,a145g/v143g,a145g/v143m的基础上对144位进行迭代饱和突变
[0170]
以cdgludh-a145g/v143a,cdgludh-a145g/v143g,cdgludh-a145g/v143m为模板对144位进行迭代饱和突变,引物见表12。
[0171]
表12单点饱和突变库构建所用引物
[0172][0173]
按步骤一的方法进行突变库的构建和筛选,最终筛选得到3个酶活显著提高的突变体(表13):
[0174]
表13 144位迭代饱和突变库阳性突变筛选结果
[0175][0176]
实施例6谷氨酸脱氢酶突变体(cdgludh-a145g/v143a/p144g),葡萄糖脱氢酶双酶耦联制备l-草铵膦
[0177]
菌体的培养及粗酶液制备:将表达cdgludh-a145g/v143a/p144g谷氨酸脱氢酶突变体基因和葡萄糖脱氢酶基因(ncbi登录号:wp_087960837.1,碱基序列克隆到表达质粒pet-28a( )上,插入酶切位点为bamhⅰ和hindⅲ,转入表达宿主e.coli bl21(de3)中)的工程菌的甘油管经平皿划线活化后,挑单菌落接种至含50μg/ml卡那霉素的50ml lb液体培养基中,37℃震荡培养12h。按2%的接种量转接至1l同样含50μg/ml kan的新鲜lb液体培养基
中,37℃震荡培养至od
600
达到0.6左右时,加入iptg至其终浓度为0.5mm,18℃下诱导培养16h。培养结束后,将培养液12000g 4℃离心10min,收集菌体,超声破胞制备粗酶液。
[0178]
反应体系为50ml,含有200mm底物ppo,300mm葡萄糖,200mm(nh4)2so4与0.2mm nad
;谷氨酸脱氢酶菌体(湿重)浓度为2g/l,葡萄糖脱氢酶菌体(湿重)浓度为4g/l。通过水浴控制反应温度为40℃,反应过程通过滴加氨水控制ph为8.0。反应2h后利用hplc检测ppo的残余浓度,同时利用柱前衍生化高效液相色谱检测l-草铵膦的生成量与ee值。
[0179]
反应结束数据如下:ppo剩余0mm,转化率100%。l-草铵膦的生成浓度为32.3g/l,ee值》99.9%。
[0180]
实施例7谷氨酸脱氢酶突变体(cdgludh-a145g/v143a/p144m),甲酸脱氢酶双酶耦联制备l-草铵膦
[0181]
按实施例6中同样的方法培养表达cdgludh-a145g/v143a/p144m谷氨酸脱氢酶突变体基因和甲酸脱氢酶(ncbi登录号:p33160.3,碱基序列克隆到表达质粒pet-28a( )上,插入酶切位点为bamhⅰ和hindⅲ,转入表达宿主e.coli bl21(de3)中)的工程菌,离心收集细胞,并超声破胞制备粗酶液。
[0182]
反应体系为50ml,含有200mm底物ppo,600mm甲酸铵,100mm(nh4)2so4与0.2mm nad
;谷氨酸脱氢酶菌体(湿重)浓度为2g/l,甲酸脱氢酶菌体(湿重)浓度为4g/l。通过水浴控制反应温度为40℃,反应过程通过滴加氨水控制ph为8.0。反应2h后利用hplc检测ppo的残余浓度,同时利用柱前衍生化高效液相色谱检测l-草铵膦的生成量与ee值。
[0183]
反应结束数据如下:ppo剩余0mm,转化率100%。l-草铵膦的生成浓度为32.1g/l,ee值》99.9%。
[0184]
实施例8谷氨酸脱氢酶野生型和突变体与醇脱氢酶双酶耦联制备l-草铵膦
[0185]
按实施例6中同样的方法培养表达谷氨酸脱氢酶野生型(cdgludh-wt)和突变体(cdgludh-a145g,cdgludh-a145g/v143a/p144a)以及醇脱氢酶(ncbi登录号:nz_jynw01000069.1,碱基序列克隆到表达质粒pet-28a( )上,插入酶切位点为bamhⅰ和hindⅲ,转入表达宿主e.coli bl21(de3)中)的工程菌,离心收集细胞,并超声破胞制备粗酶液。
[0186]
反应体系为50ml,含有200mm底物ppo,300mm异丙醇,200mm(nh4)2so4与0.2mm nad
;谷氨酸脱氢酶菌体(湿重)浓度为2g/l,醇脱氢酶菌体(湿重)浓度为4g/l。通过水浴控制反应温度为40℃,反应过程通过滴加氨水控制ph为8.0。定时取样,利用hplc检测ppo的残余浓度,转化率随时间变化数据如图7所示。同时利用柱前衍生化高效液相色谱检测l-草铵膦的最终生成量与ee值。
[0187]
cdgludh-wt催化反应:ppo剩余205.3mm,转化率0%,l-草铵膦的生成浓度为0g/l;
[0188]
cdgludh-a145g催化反应330min:ppo剩余0mm,转化率100%,l-草铵膦的生成浓度为32.8g/l,ee值》99.9%;
[0189]
cdgludh-a145g/v143a/p144a催化反应75min:ppo剩余0mm,转化率100%,l-草铵膦的生成浓度为30.9g/l,ee值》99.9%。
[0190]
实施例9谷氨酸脱氢酶野生型和突变体与醇脱氢酶双酶耦联制备l-正缬氨酸
[0191]
按实施例6中同样的方法培养表达谷氨酸脱氢酶野生型(cdgludh-wt)和突变体(cdgludh-a145g/k71a/v143a,cdgludh-a145g/v352a/k71a/r187a/t175a)以及醇脱氢酶(ncbi登录号:nz_jynw01000069.1)的工程菌,离心收集细胞,并超声破胞制备粗酶液。
[0192]
反应体系为50ml,含有200mm底物2-氧代戊酸,300mm异丙醇,200mm(nh4)2so4与0.2mm nad
;谷氨酸脱氢酶菌体(湿重)浓度为2g/l,醇脱氢酶菌体(湿重)浓度为4g/l。通过水浴控制反应温度为40℃,反应过程通过滴加氨水控制ph为8.0。定时取样,利用hplc检测2-氧代戊酸的残余浓度,转化率随时间变化数据如图8。同时利用柱前衍生化高效液相色谱检测l-正缬氨酸的最终生成量与ee值。
[0193]
cdgludh-wt催化反应270min:2-氧代戊酸剩余0mm,转化率100%,l-正缬氨酸的生成浓度为21.8g/l,ee值》99.5%;
[0194]
cdgludh-a145g/k71a/v143a催化反应150min:2-氧代戊酸剩余0mm,转化率100%,l-正缬氨酸的生成浓度为22.1g/l,ee值》99.6%;
[0195]
cdgludh-a145g/v352a/k71a/r187a/t175a催化反应360min:2-氧代戊酸剩余81.2mm,转化率59.4%,l-正缬氨酸的生成浓度为12.7g/l,ee值》99.3%。
[0196]
实施例10谷氨酸脱氢酶野生型和突变体与醇脱氢酶双酶耦联制备l-高苯丙氨酸
[0197]
按实施例6中同样的方法培养表达谷氨酸脱氢酶野生型(cdgludh-wt)和突变体(cdgludh-a145g/s355a/v352a/k71a/r187a/t175a,cdgludh-a145g/v352a/k71a/r187a/t175a)以及醇脱氢酶(ncbi登录号:nz_jynw01000069.1)的工程菌,离心收集细胞,并超声破胞制备粗酶液。
[0198]
反应体系为50ml,含有50mm底物2-氧代-4-苯基丁酸,300mm异丙醇,200mm(nh4)2so4与0.2mm nad
;谷氨酸脱氢酶菌体(湿重)浓度为2g/l,醇脱氢酶菌体(湿重)浓度为4g/l。通过水浴控制反应温度为40℃,反应过程通过滴加氨水控制ph为8.0。定时取样,利用hplc检测2-氧代-4-苯基丁酸的残余浓度,转化率随时间变化数据如图9。同时利用柱前衍生化高效液相色谱检测l-高苯丙氨酸的最终生成量与ee值。
[0199]
cdgludh-wt催化反应:2-氧代-4-苯基丁酸剩余52.8mm,转化率0%,l-高苯丙氨酸的生成浓度为0g/l;
[0200]
cdgludh-a145g/s355a/v352a/k71a/r187a/t175a催化反应180min:2-氧代-4-苯基丁酸剩余0mm,转化率100%,l-高苯丙氨酸的生成浓度为8.8g/l,ee值》99.9%;
[0201]
cdgludh-a145g/v352a/k71a/r187a/t175a催化反应105min:2-氧代-4-苯基丁酸剩余0mm,转化率100%,l-高苯丙氨酸的生成浓度为8.7g/l,ee值》99.9%。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。
