用于确定生物样本中纤维蛋白原浓度的方法
1.本技术是2015年12月18日提交的申请号为201580068832.x、名称为
ꢀ“
用于确定生物样本中纤维蛋白原浓度的方法”的发明专利申请的分案申 请。
技术领域
2.本发明涉及临床决策支持系统。具体而言,本发明涉及一种用于确定 生物样本中初始纤维蛋白原浓度的方法、以至少一个预定浓度添加到生物 样本中的纤维蛋白原的使用、用于确定生物样本中初始纤维蛋白原浓度的 设备、包括用于令计算机执行根据本发明的方法的至少几个步骤的程序代 码单元的计算机程序、包含用于执行根据本发明的方法的至少一些步骤的 指令的计算机可读非瞬态存储介质,以及用于确定生物样本中初始纤维蛋 白原浓度的套件。
背景技术:
3.纤维蛋白原是参与凝血的一种重要蛋白质。正常血流时,纤维蛋白原 可溶;然而,一旦激活凝血系统,纤维蛋白原最终被凝血酶转化为纤维蛋 白。纤维蛋白随后聚合成不可溶解的纤维蛋白纤维,优选地与活化血小板 一起形成凝块。纤维蛋白原的正常浓度约为2.5g/l,范围约为1.5到3g/l[greer 等人(编辑的)《wintrobe's clinical hematology》第11版第720页]。然而, 在很多情况下,纤维蛋白原浓度可能超出正常范围,这可能与病理性疾病 有关。例如,在遗传性低纤维蛋白原血症患者中,由于持续出血造成纤维 蛋白原浓度低导致的危险状况,可通过添加血液制品来应对。另一方面, 发现纤维蛋白原的浓度升高与心肌梗塞风险升高相关(bom等人, arterioscler.thromb.vasc.biol.(1998)18(4):621
‑
625)、血栓形成(kamath 和lip,q.j.med.(2003)96(10):711
‑
729),以及长期炎症过程,例如类风湿 性关节炎(rooney等人,rheumatology(2011)50(8):1458
‑
1465)有关。由 于这些变化的纤维蛋白原浓度和关联的病理学,纤维蛋白原浓度测试是一 项有价值的临床测试。然而,不幸的是,没有可靠的护理点(poc)纤维 蛋白原测试,并且中心实验室测试预约对于时间紧急的状况(例如,在手 术室中)通常需要太长时间。
[0004]
已经开发了很多方法,旨在准确地检测血浆或血液样本中纤维蛋白原 的水平(例如,参见palarati等人的clinical chemistry(1991)37(5): 714
‑
719,以获取可用技术的概述,或ep 2259069)。
[0005]
遗憾的是,目前所有可用的方法要么是非常劳动密集的,例如,凝血 恢复方法,要么需要从具有已知纤维蛋白原水平的血浆样本导出校准曲线 以推断样本的纤维蛋白原水平,例如clauss化验或凝血酶时间推导方法。 前者涉及大量的手工操作时间,因此难以自动化,而后一种方法需要在测 试规程中包括校准血浆。ep 0059277中公开了需要至少两个血浆参考样本 来校准测量结果的方法。不过,这些方法不适合并入自动化设备中,例如 手持式护理点(poc),以检测纤维蛋白原的水平。
技术实现要素:
[0006]
本发明的目的是提供一种用于确定生物样本中初始纤维蛋白素浓度的 方法,借助于该方法,可以避免现有技术方法的缺点。具体而言,应提供z 这种用于确定生物样本中初始纤维蛋白原浓度的方法,其允许可靠地确定 主体生物样本是否有出血倾向或发生血栓形成或者与主体或患者体液(例 如血液或血浆)中纤维蛋白原水平相关联的其他病理的风险。
[0007]
本发明的另一个目的是提供:一种用于确定生物样本中初始纤维蛋白 原浓度的设备;一种包括程序代码单元的计算机程序,所述程序代码单元 用于令计算机执行根据本发明的方法的至少一些步骤;一种包含所述计算 机程序的计算机可读非瞬态存储介质;以及一种用于确定生物样本中初始 纤维蛋白原浓度的套件。
[0008]
在本发明的第一方面中,提出了一种用于确定生物样本中初始纤维蛋 白原浓度的方法,所述方法包括以下步骤:
[0009]
(1)提供生物样本;
[0010]
(2)以至少一个预定浓度向所述生物样本添加纤维蛋白原;
[0011]
(3)在所述生物样本中开始凝血过程;
[0012]
(4)确定作为时间的函数的所述生物样本的衰减,以获得衰减曲线;
[0013]
(5)从所述衰减曲线提取指示所述生物样本中纤维蛋白原浓度的特征值, 并且作为在步骤(2)中添加的纤维蛋白原的函数来记录它们;
[0014]
(6)通过步骤(5)的记录的所述特征值来计算拟合函数;
[0015]
(7)根据步骤(6)的所述拟合函数来确定所述生物样本中的所述初始纤 维蛋白原浓度。
[0016]
在本发明的另一个方面中,提供了以至少一个预定浓度添加到生物样 本的纤维蛋白原用于确定所述生物样本中的初始纤维蛋白原浓度的使用。
[0017]
在本发明的又一方面中,提供了一种用于确定生物样本中初始纤维蛋 白原浓度的设备,所述设备包括:
[0018]
(1)能够接收生物样本和至少一个预定浓度的纤维蛋白原的添加的容器;
[0019]
(2)测量单元,其被配置为测量所述生物样本的衰减;
[0020]
(3)计算单元,其被配置为:
[0021]
确定作为时间的函数的所述生物样本的衰减,以获得衰减曲线;
[0022]
从所述衰减曲线提取表示所述生物样本的纤维蛋白原浓度的特征值, 并且作为向所述生物样本添加的纤维蛋白原的函数来记录它们;
[0023]
通过记录的所述特征值来计算拟合函数;
[0024]
根据所述拟合函数来确定所述生物样本中的所述初始纤维蛋白原浓 度。
[0025]
在本发明的又一方面中,提供了一种计算机程序,所述计算机程序包 括程序代码单元,所述代码单元用于令计算机至少执行根据本发明的方法 的步骤(4)、(5)、(6)、(7)。
[0026]
在本发明的又一方面中,提供了一种计算机可读非瞬态存储介质,所 述计算机可读非瞬态存储介质包含用于由处理器执行的指令,其中,所述 指令使处理器至少执行根据本发明的方法的步骤(4)、(5)、(6)、(7)。
[0027]
在本发明的又一方面中,提供了一种用于确定生物样本中初始纤维蛋 白原浓度
的套件,所述套件包括根据本发明的设备,纤维蛋白原,优选为 干的或冻干形式的,还优选地包括预定量的纤维蛋白原,用于执行根据本 发明的方法的手册,以及任选地,凝血触发剂,并且还任选地包括根据本 发明的计算机可读非瞬态存储介质。
[0028]
在本发明的又一个方面中,提供了一种包括程序代码单元的计算机程 序,所述程序代码单元用于,当所述计算机程序在计算机上执行时,令所 述计算机执行所述方法的步骤,并且提供了一种在其中存储有计算机程序 产品非瞬态计算机可读记录介质,所述计算机程序产品当由计算机处理器 运行时,令本文公开的方法的步骤被执行。
[0029]
在从属权利要求中定义了本发明的优选实施例。应当理解,请求保护 的用法、设备、计算机程序、计算机可读非瞬态存储介质和套件与请求保 护的并且如在从属权利要求中定义的方法具有相似和/或相同的优选实施 例。因此,涉及根据本发明的方法的所有从属权利要求也可以与根据本发 明的用法、设备、计算机程序、计算机可读非瞬态存储介质和套件相组合 以及彼此组合。
[0030]
本发明的发明人提出了一种方法,其可能在(例如在poc设备中具有 多个检测室的试剂盒(cartridge)上分配的)并行试验中,向生物样本自身 添加至少一个量但优选多个量的纤维蛋白原,而不是利用根据具有已知纤 维蛋白原水平的若干血浆或具有已知纤维蛋白原水平的血浆的稀释系列构 造的校准曲线。这将得到样本自身内部的内部校准曲线,而不是根据具有 已知纤维蛋白原水平的血浆样本导出的外部校准曲线。根据本发明的方法 可自动用在例如具有多个腔室且具有已知纤维蛋白原含量或浓度的poc平 台上,而不需要校准血浆来确定样本中的纤维蛋白原水平。还可以在中心 实验室设备上实施根据本发明的方法,这导致省略或减少校准血浆的使用。 此外,内部校准的优点是,与外部校准相比,内部校准可以更适当地校正 初始样本的物理性质,例如基质成分。
[0031]
通过向生物样本添加一个或多个纤维蛋白原浓度来使用内部校准曲线 以确定纤维蛋白原的初始浓度在现有技术中既非已知也非显而易见。
[0032]
根据本发明,“生物样本”是指怀疑包含纤维蛋白原的任何样本。生物 样本可以是天然或非天然来源的。优选的样本是体液,尤其是能够经历血 液结块或凝聚的那种,例如血液样本、血浆样本、全血样本等。
[0033]
根据本发明,“初始纤维蛋白原浓度”是指在步骤(1)中,即,在步 骤(2)中添加任何纤维蛋白原之前,提供的生物样本中存在的纤维蛋白原 浓度。
[0034]
根据本发明,“预定浓度”是指用户具有所添加的纤维蛋白原浓度的确 切的知识。“添加纤维蛋白原”是指向生物样本添加纤维蛋白原。“添加
”ꢀ
也可以指向容器中放置的预定浓度纤维蛋白原添加生物样本,例如,以纤 维蛋白原溶液或干的或冻干的形式。
[0035]
根据本发明,“结块过程”是指涉及纤维蛋白原转换成纤维蛋白的过程。 这种转换优选跟随着后续步骤,例如,纤维蛋白聚合成不可溶解纤维蛋白 纤维,它们与优选激活的血小板一起形成凝血。血液或血浆样本中完整的 凝血过程也称为血液凝聚过程。凝血过程例如可以通过向生物样本添加所 谓的凝血触发剂来开始。
[0036]
根据本发明,“衰减”是指例如由于散射和荧光而导致的诸如生物样本 的物质的吸收加损耗的度量。诸如凝聚血浆或血液样本的生物样本的衰减 由于在样本凝血期间形成纤维蛋白网络而随时间改变。纤维蛋白纤维是在 向样本添加凝血触发剂之后形成的。这些纤维蛋白纤维造成入射光散射, 造成更少的光子到达探测器。光子的实际吸收和衰减的
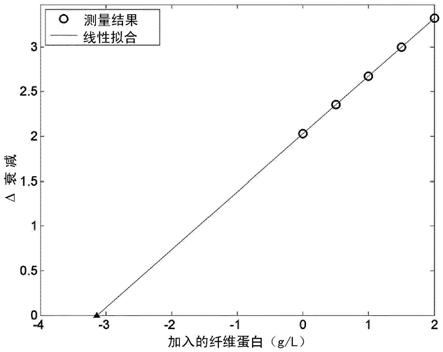
其他原因,例如荧 光,被认为在凝血期间随时间恒定,因此认为衰减并且因此透射的减少和 光密度增大仅仅是因为入射光被样本中形成的纤维蛋白纤维散射造成的。 材料的衰减是log10(p0/p),其中p0是样本上入射的辐射功率,p是由其透 射的功率。这个量还是
‑
log10(t),其中,t是透射率。衰减通常直接与在文 献中诸如“光密度”、“浊度”或“消光”的术语相关或涉及其。而且,常 常针对这个量错误地使用名称“吸收”(符号:a),但是在辐射衰减是由 于散射而非吸收造成时,对于该量而言这是不适当的。该量自身被称为衰 减(符号:d),注意,在散射或反射可忽略时,衰减退化为吸收。发明人 发现,在衰减退化到散射时,科学家可以使用术语“浊度”,一般将其考虑 为
‑
ln(t)。在后一种情况下,“浊度”可以通过测量透射率来估计。为了实际 测量由于溶液中颗粒导致的散射,存在专门的技术,例如小角度散射或浊 度测定法。可以将衰减转换成“浊度”、“透射率”、“光密度”、“吸收率
”ꢀ
和由于溶液中颗粒造成的光衰减或散射的其他度量。因此,根据本发明, 前述尺度或术语可以用于替代衰减。
[0037]
为了建立“衰减曲线”,相对于特定参考点,例如,凝血过程的开始点, 至少针对一个,但优选针对多个时间点,确定生物样本的衰减值。然后随 时间绘制衰减值。血浆样本的典型衰减曲线分别由其“s”形状或乙状曲线 进展来表征。或者,仅在两个时间点处测量衰减,即凝血过程的开始点和 凝血过程(接近)完全发展之后凝血的结束阶段。
[0038]
根据本发明,“特征值”是指可以指,例如,通过使用常见的数学方法, 例如微分、导数、积分等,从衰减曲线提取的这样的信息,其指示所述生 物样本中的纤维蛋白原浓度。
[0039]
在将这样的特征值添加的纤维蛋白原的函数被记录时,根据本发明, 可以通过记录的所述特征值来计算“拟合函数”。这样的流程被称为“曲线 拟合”,即构造最佳拟合到一系列数据点的曲线或数学函数的过程。曲线拟 合可能涉及内插,其中需要精确拟合到数据,或者平滑化,其中构造近似 拟合数据的“平滑”函数。曲线拟合包括回归分析,其聚焦于统计推断的 问题,例如,在拟合到具有随机误差的观测的数据的曲线中存在多大不确 定性。
[0040]
根据本发明,根据拟合函数来“确定”生物样本中的初始纤维蛋白原 浓度是通过利用现有技术中公知的方法,例如外推,包括线性、多项式、 锥形或曲线板外推,来实现的。对于线性拟合,以下方程可以适用:fy=α* fg
add
β,fy为特征值,fg
add
为添加的纤维蛋白原,α和β为拟合参数,分别 是线性拟合的斜率和偏移。那么,纤维蛋白原浓度可以是fg
样本
=
‑
β/α。
[0041]
在根据本发明的设备中,“容器”是指适于接收生物样本和纤维蛋白 原的任何贮存器,例如阱、液槽、试剂盒、试管等,可能分成多个子容器。
[0042]“测量单元”包括适于测量所述生物样本衰减的任何测量工具,例如 光度计。
[0043]“计算单元”是指包括处理信息的硬件和软件部件的设备,例如,处 理器、计算机,例如用于计算机的程序、算法等。
[0044]
作为本发明所有实施例中衰减曲线的替代,可以使用“质量/长度曲线”。 质量/长度是平均纤维蛋白纤维的平均质量除以其长度。质量/长度曲线与衰 减/光密度/浊度曲线通过从纤维蛋白纤维的物理性质提取的方程而彼此相 关。这里,示范性地示出了浊度和平均质量/长度比之间的关系:
[0045][0046]
其中,τ是作为透射率t的负自然对数的浊度,n
a
为将密度变换成da/cm3的阿伏加德罗数,μ为以道尔顿每厘米为单位的纤维的平均质量/长度比, 并且a和b是可以在独立试验中或通过在不同波长处测量已知溶质浓度的 固定质量/长度比光密度而确定的集总参数。在优选的实施例中,在波长 632.8nm处a和b的值可以分别为6.76χ10
22
和1.41χ10
24
。衰减和/或光 密度与质量/长度比之间的关系直接遵循这个方程。
[0047]
在这一替代方式中,指示所述生物样本中的纤维蛋白原浓度的特征值 类似地也是从质量/长度曲线提取的。例如,添加了纤维蛋白原的样本中观 测到的最大比率指示添加的纤维蛋白原和最大比率之间的线性关系。可以 类似地使用这一关系来推断样本的初始纤维蛋白原浓度。可以通过倒数函 数或者替代地通过幂函数或指数函数来最佳地近似从涉及曲线动态的平均 质量/长度比提取的特征值。
[0048]
根据依据本发明的方法的进一步发展,所述步骤(7)是通过以下步骤 来实现的:
[0049]
(7.1)将步骤(6)的所述拟合函数外推到所述特征值为零之处的添加的纤 维蛋白原的值,以获得外推值;
[0050]
和/或
[0051]
将步骤(6)的所述拟合函数外推到与纤维蛋白原水平为零的假想样 本符合的值之处的添加的纤维蛋白原的值,以获得外推值;
[0052]
(7.2)通过将所述外推值乘以
‑
1来确定所述生物样本中的初始纤维蛋白原 浓度。
[0053]
根据本发明,添加的纤维蛋白原的“值”可以指反映添加的纤维蛋白 原的任何度量,例如,纤维蛋白原的浓度,例如,以g/l表示,或添加的纤 维蛋白原的量等。
[0054]
根据本发明,“假想样本”是指并非实际存在的样本而是作为数学运算 结果的“理论样本”。
[0055]
这样的实施例在已经计算出线性拟合的情况下提供了可靠的结果。将 线性拟合外推到与水平轴的交点,其对应于样本中不存在纤维蛋白原的情 况下的特征值,交点处的这个值指示为了在样本中获得纤维蛋白原零浓度 需要添加的纤维蛋白原浓度。因此,这个交点将为负值(或零),从而必须 要乘以
‑
1以获得初始样本中的纤维蛋白原浓度或水平。
[0056]
因此,在进一步发展中,本发明的计算机程序包括程序代码单元,用 于在计算机上执行所述计算机程序时,令所述计算机至少执行根据本发明 的方法的步骤(4)、(5)、(6)、(7)、(7.1)和(7.2)。
[0057]
因此,在进一步发展中,一种计算机可读非瞬态存储介质包含用于供 处理器执行的指令,其中,所述指令使处理器至少执行根据本发明的方法 的步骤(4)、(5)、(6)、(7)、(7.1)和(7.2)。
[0058]
根据本发明方法的进一步发展,指示所述生物样本中纤维蛋白原浓度 的所述特征值选自以下项:
[0059]
所述生物样本在第一时间点与第二时间点之间的衰减的差异(δ衰减), 优选地,所述生物样本在作为所述第一时间点的凝血过程的开始点与作为 所述第二时间点的凝血过程的结束之间的衰减的差异(δ衰减);
度是合理的,凝血酶浓度的更高值是更加优选的,因为这将缩短化验时间 并将降低系统中可能反馈的效果。对于蛇毒类凝血酶样酶,1bu/ml或更 高的浓度是合理的,对于凝结因子,高于10nm的浓度是优选的。
[0074]
在根据本发明的方法的另一个方面中,在步骤(1)中,所述生物样本 被提供在至少两个等份中,允许在所述至少两个等份中,优选并行地,执 行步骤(2)到(4),从而获得至少两条衰减曲线。
[0075]
即使可以向生物样本的单一等份添加多个预定纤维蛋白原浓度,但将 其分成几个等份,向其每个添加不同的预定纤维蛋白原浓度能够获得更加 可靠的结果。“至少两个等份”和“至少两条衰减曲线”分别包括3、4、5、 6、7、8、9、10、20、30、40、50、100和1000等的等份和衰减曲线甚至 更多。在优选实施例中,可以在板或试剂盒,包括多个阱或腔室中提供生 物样本。阱或腔室中的每个都可以包含已知的纤维蛋白原含量。
[0076]
与此适应,在根据本发明的该设备的另一个方面中,所述容器是能够 接收所述生物样本的至少两个等份的多阱容器,优选为微量滴定板。
[0077]
这样的多阱容器或微量滴定板允许大规模或工业化规模地应用根据本 发明的方法。
[0078]
在本发明设备的又一方面中,所述测量单元是光学测量系统,包括至 少一个光源和至少一个光探测器。
[0079]
该措施的优点是,提供了尤其适于结合本发明工作的这样的测量单元。 不言而喻,例如,根据生物样本的等份数量和设备的具体技术要求,所述 光学测量系统可以具有2、3、4、5和更多的光源和探测器。
[0080]
如上所述,本发明的另一个主题是以至少一个预定浓度添加到生物样 本中的纤维蛋白原来确定所述生物样本中的初始纤维蛋白原浓度的用途。
[0081]
发明人已经认识到,为了确定生物样本中纤维蛋白原的初始浓度,不 需要从具有已知纤维蛋白原水平的若干血浆构造外部校准曲线。相反,可 以使用内部校准曲线,因为这已通过本发明实现。这样的用法可以很容易 实现自动化,例如,以具有已知纤维蛋白原含量的多个腔室的poc平台的 形式,因此不再需要使用校准血浆。
[0082]
应理解,本发明的优选实施例也可以是从属权利要求与各自的独立权 利要求的任意组合。
[0083]
参照下文中描述的实施例,本发明的这些和其他方面将显而易见且得 以阐述。
附图说明
[0084]
在附图中:
[0085]
图1示出了在凝血酶发起的凝血血浆中随时间(水平轴)测量的(垂 直轴上)衰减或光密度或浊度变化的典型范例。在将纤维蛋白原转变成纤 维蛋白的短的滞后时间之后,激活的纤维蛋白单体开始聚合并形成粗的纤 维蛋白纤维,其引起光散射,这可以通过透射光减少或浊度或衰减增大而 检测到;
[0086]
图2示出了在样本以及添加了0.5、1、1.5、2g/l纤维蛋白原(空心圆) 的样本中的δ衰减。可以使用线性函数来拟合测量点。将线性拟合外推到与 水平轴的交点(对应于样本中没有纤维蛋白原情况下的δ衰减),指示样本 的初始纤维蛋白原浓度为3.14159g/l;
[0087]
图3示出了在添加了1、2、3和4g/l的纤维蛋白原并利用0.02(o)、0.06( ) 和1(x)nih u/ml触发的纤维蛋白原不足的血浆中观测到的δ衰减。利用本 文描述的方法,使用线性函数的拟合参数,计算样本中的纤维蛋白原浓度, 分别得到0.12(95%置信度区间:
‑
0.013
–
0.24)、0.16(95%
‑
ci:0.006
–
0.29) 和0.033(95%
‑
ci:
‑
0.074
–
0.13)g/l的纤维蛋白原;
[0088]
图4示出了利用范围从
‑
3.1(对应于纤维蛋白原提取)到0.5g/l的包 含3.14159g/l纤维蛋白原的样本的模拟纤维蛋白原添加试验,观测到的以 秒为单位的滞后时间变化。发现形式为滞后时间(fg
add
)=85.7234/(fg
add
–
3.1416)的倒数拟合紧密拟合观测结果。在
‑
3.1416处发现垂直渐近线,
[0089]
图5示意性示出了根据本发明的方法的示范性实施例的相应流程图。
具体实施方式
[0090]
血浆的凝血可以分别由例如组织因子、高岭土、凝血酶或另一种凝血 剂或触发剂来发起。在这种凝血期间,血浆样本的不透明度会变化。这将 导致光透射的减小,这可以利用样本衰减、光密度或浊度随时间的变化来 测量(carr和gabriel,macromolecules(1980)13(6):1473
‑
1477)。样本 中纤维蛋白原的量对衰减、光密度或浊度曲线的形状有很高影响,并且凝 血化验可以被开发为,使得在充分高浓度的凝血酶或另一种具有与纤维蛋 白原类似活性的类似酶下,血浆样本的衰减或光密度曲线之间的差异主要 因为纤维蛋白原浓度的差异。参见图1,利用凝血酶发起凝血的血浆样本的 典型衰减曲线范例。因此,从衰减曲线提取的特征指示样本中的纤维蛋白 原水平,并可以用于利用向样本添加已知纤维蛋白原水平来构造校准曲线。
[0091]
在根据本发明的方法中,通过感兴趣样本自身通过优选地向初始样本 添加至少一种,但优选几种,已知纤维蛋白原浓度来进行校准替代基于具 有已知纤维蛋白原浓度的外部对照血浆的校准曲线。来自衰减曲线的感兴 趣特征是从多次测量提取的。
[0092]
图2示出了样本衰减(在模型中计算)在测量开始和结束之间的理论 差异,也被称为δ衰减,是添加的纤维蛋白原的浓度的函数(这里是每升血 浆样本添加的以克为单位的纤维蛋白原)。δ衰减是来自本发明中示范性使 用的衰减曲线的优选特征。在图2中可以看出,这一特征随着添加的纤维 蛋白原而线性变化。如果在添加任何纤维蛋白原之前样本中没有纤维蛋白 原,在零添加纤维蛋白原处会没有衰减,因为不会形成纤维蛋白。初始样 本,即在零添加纤维蛋白原处δ衰减的非零值指示样本中纤维蛋白原的初始 浓度,即,要通过根据本发明的方法确定的值。
[0093]
在针对添加的纤维蛋白原的零以及一个或多个非零值处测量凝血特 征,例如δ衰减,因此跟随着涉及通过多个观测的特征值计算拟合的步骤, 有时称为回归。然后,拟合曲线的外推产生血浆样本中的初始纤维蛋白原 浓度。对于δ衰减特征,这以下地工作:
[0094]
估计线性拟合(δ衰减=α*fg
add
β,其中,fg
add
为添加的纤维蛋白原, α和β为拟合参数,分别是线性拟合的斜率和偏移);
[0095]
将拟合的函数外推到δ衰减为零处的添加的纤维蛋白原的值。这将是负 值(或零);
[0096]
样本中的纤维蛋白原水平被计算为外推值加以负号。对于δ衰减所遵循 的添加
的纤维蛋白原的线性关系而言,这等于直线的偏移除以斜率(fg
样本 =
‑
β/α)。
[0097]
将以δ衰减作为观测特征的以上方法应用于纤维蛋白原缺乏的血浆样 本,即,这种样本中纤维蛋白原的水平接近零。在本范例中,发现样本具 有残余纤维蛋白原抗原浓度0.05g/l(利用elisa检测)。接下来,利用多 种凝血酶浓度(0.02、0.06和1nih u/ml)结合多种纤维蛋白原添加(1、 2、3和4g/l)来发起纤维蛋白原缺乏血浆中的凝血。针对(12)试验组合 中的每种,在四次复制中测量每种组合的浊度或衰减。对衰减曲线求平均 值,并针对每种试验条件计算δ衰减,参见图3。
[0098]
下一步是将线性函数拟合到前述凝血酶浓度的每一种(参见图2)。接 下来,利用前述方程推断初始样本的纤维蛋白原浓度。这获得纤维蛋白原 水平为0.12(95%置信度区间;ci:
‑
0.013
‑
0.24),0.16(95%
‑
ci:0.006
‑
0.29) 和0.033(95%
‑
ci:
‑
0.074
‑
0.13)g/l,分别针对0.02、0.06和1nih u/ml 凝血酶,其与预期纤维蛋白原浓度对应得很好。最好的结果,即,最接近 通过elisa确定的实际纤维蛋白原水平以及最小置信度区间的结果,是利 用1nih u/ml的凝血酶获得的,这可能是由于凝聚系统中的反馈对凝血过 程影响最小造成的。
[0099]
替代地,对于衰减曲线的可能展现出针对多次纤维蛋白原添加的非线 性行为的其他提取特征,可以接着进行类似方法。例如,对应于到达最大 衰减(95%)的时间,例如,滞后时间,到达最大速率的时间(即,出现曲 线时间导数最大值的时间)的特征可以与例如以添加的纤维蛋白原作为变 量的指数、幂或倒数函数拟合。这样的特征在样本中真正纤维蛋白原浓度 趋向零时趋向无穷大,如在利用计算机模型模拟的试验中所见,参见图4。 拟合曲线的(垂直)渐近线的水平(x
‑
)坐标涉及样本中纤维蛋白原水平的 负值。在拟合形式为α/(fg
add
‑
β)的倒数函数的情况下(其在特征与时间点相 关的情况下是优选的),其中,fg
add
为添加的纤维蛋白原水平,α,β为利用 算法拟合的未知参数,
‑
β等于样本的纤维蛋白原水平,在β处找到垂直渐 近线时。另一个特征,即衰减曲线的最大速率优选利用形式为α*(fg
add
‑
β)2的函数拟合,fg
add
为添加的纤维蛋白原的水平,α,β为利用算法拟合的未知 参数。在可以在β处发现象限函数的最小值的情况下,因此
‑
β也等于样本 的纤维蛋白原水平。
[0100]
替代地,可以使用从平均质量/长度曲线提取的特征来推断纤维蛋白原 浓度。例如,在添加了纤维蛋白原的样本中观测到的最大速率表示添加的 纤维蛋白原和最大速率(即曲线的时间导数的最大值)之间存在线性关系。 这一关系同样可用于推断样本的纤维蛋白原浓度。从与曲线动力学有关的 平均质量/长度比提取的特征,即,滞后时间、到达最大速率(即,曲线时 间导数的最大值)的时间、到达最大质量/长度比(的95%)的时间,可以 通过倒数函数或幂函数或指数函数进行最佳近似。如前所述,可通过找到 (垂直)渐近线来推断样本的纤维蛋白原水平。
[0101]
图5示意性示出了根据本发明的方法的实施例的相应流程图。在步骤 (s1)中,提供生物样本,例如怀疑具有改变的纤维蛋白原浓度的个人的 血浆样本。在步骤(s2)中,在所述生物样本中,例如通过添加凝血触发 剂,例如分别为凝血酶(f2a)或蛇毒类凝血酶样酶,开始凝血过程。在步 骤(s3)中,以至少一个预定浓度向所述生物样本添加纤维蛋白原。优选 地,在并行试验中向样本的等份添加多种量的纤维蛋白原,例如被划分到 具有多个检测腔室的试剂盒上。在步骤(s4)中,确定作为时间的函数的 包含(一个或多个)添加的纤维蛋白原的浓度的(一个或多个)生物样本 的衰减(
‑
s),并获得一条/几条衰减曲线。
[0102]
在步骤(s5)中,从所述衰减曲线提取指示所述生物样本中纤维蛋白 原浓度的特征值,例如,δ衰减,并记录为在步骤(s3)中添加的纤维蛋白 原的函数。在步骤(s6)中,通过步骤(s5)记录的所述特征值来计算拟 合函数。在步骤(s7.1),将步骤(s6)的所述拟合函数外推到所述特征值 为零时所添加的纤维蛋白原的值,或者如果特征与凝血过程的定时有关, 则外推到垂直渐近线的值,或者在衰减曲线的最大速率的情况下,外推到 二次拟合函数具有最小值之处的值,以获得外推值。在步骤(7.2)中,通 过将所述外推值乘以
‑
1来确定所述生物样本中的初始纤维蛋白原浓度。
[0103]
尽管已在附图和前文的描述中详细说明和描述了本发明,但是这种说 明和描述应被视为说明性或示范性的,而非限制性的;本发明不限于所公 开的实施例。本领域技术人员通过研究附图、公开内容以及权利要求书, 在实践请求保护的发明时能够理解并实现对所公开实施例的其他变型。
[0104]
在权利要求中,词语“包括”不排除其他元件或步骤,且词语“一
”ꢀ
或“一个”并不排除复数。单个处理器或其他单元可以履行权利要求书中 所记载的若干个项目的功能。尽管在互不相同的从属权利要求中记载了特 定措施,但是这并不表面不能有利地使用这些措施的组合。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。