稳定的crispr复合物
1.交叉引用
2.本技术要求2019年7月19日提交的美国临时专利申请第62/876,204号、2019年7月19日提交的美国临时专利申请第62/876,177号、2019年11月22日提交的美国临时专利申请第62/939,554号、2019年11月22日提交的美国临时专利申请第62/939,553号、2020年1月25日提交的国际专利申请第pct/us20/15127号和2020年4月15日提交的美国临时专利申请第63/010,465号的优先权,这些申请通过引用以其整体并入本文。
3.背景
4.crispr/cas可以用于各种医学、实验室和其他探索性环境。crispr/cas系统可以用作大量不同生物体中的基因编辑工具,以在靶位点处产生断裂并随后在基因座处引入突变。该基因编辑过程可以需要两种主要组分:内切核酸酶样cas酶和用于识别特定dna靶核酸序列的短rna分子。crispr/cas系统可以依赖于定制的短rna分子将cas酶募集至不同的核酸(例如,dna)靶位点,而不是为每种dna靶设计核酸酶。cas酶的实例包括cas9和cpf1。用于形成crispr复合物的合成引导rna,例如,单引导rna(sgrna),当未与cas酶复合时可以经历降解。用于形成crispr复合物的合成引导rna,例如,单引导rna(sgrna),可以诱导免疫应答,这可能限制目前可用的sgrna/cas核酸酶复合物的应用。在体内crispr复合物可以部分或完全解离,这可能降低效率,并且可能引起脱靶裂解事件。由于crispr复合物的不稳定性,它们通常以质粒编码的形式递送,而质粒依赖于靶细胞的转录来产生编码的蛋白和引导序列。对于在任何研究环境中均递送一致的精确比例的crispr cas酶和引导rna分子(诸如以受控的给药方案递送纯试剂)存在需求。此外,对于用于各种环境(例如,需要一种或更多种具有可调活性的外源crispr复合物的精确给药的环境)的具有增强的稳定性的crispr复合物存在需求。
5.概述
6.本文公开了一种crispr复合物,所述crispr复合物包含单引导rna(sgrna),所述单引导rna(sgrna)与crispr效应蛋白在sgrna内的非天然核苷酸处交联,其中sgrna包含crrna区和tracrrna区,并且其中非天然核苷酸位于crrna区的靶结合区之外。非天然核苷酸可以包含尿嘧啶。非天然核苷酸可以位于sgrna的核苷酸位置49处,其中核苷酸位置1位于crrna的靶结合区的5’末端处,并且sgrna的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。非天然核苷酸可以包含糖的修饰。非天然核苷酸可以包含碱基的修饰。非天然核苷酸可以包含马来酰亚胺。马来酰亚胺可以与crispr效应蛋白上的半胱氨酸共价连接。非天然核苷酸可以包括吡啶基二硫化物、烷氧基胺、nhs酯、双吖丙啶、亚氨酸酯、卤代乙酰基基团、酰肼、芳基叠氮化物、异氰酸酯、二硫醇亚磷酰胺dtpa、4
‑
硫代
‑
utp、5
‑
叠氮基
‑
utp、5
‑
溴
‑
utp、8
‑
叠氮基
‑
atp、5
‑
apas
‑
utp或8
‑
n(3)amp。
7.在一些实施方案中,非天然核苷酸可以位于tracrrna区的茎环中。该茎环的结构可以相对于缺少非天然核苷酸的sgrna的茎环的结构被维持。非天然核苷酸可以位于tracrrna区的凸起中。该凸起的结构可以相对于缺少非天然核苷酸的sgrna的凸起的结构被维持。非天然核苷酸可以位于tracrrna区的茎环之间。crispr复合物可以包括核酸酶活
性。
8.在一些实施方案中,crispr复合物的脱靶核酸酶活性等于或小于包含未交联的crispr效应蛋白和sgrna的crispr复合物的脱靶核酸酶活性。非天然核苷酸可以位于crispr效应蛋白的半胱氨酸的20埃以内。在一些实施方案中,非天然核苷酸可以不是4
‑
硫代尿苷或修饰的腺苷。
9.本文还公开了一种crispr复合物,所述crispr复合物包含单引导rna(sgrna),所述单引导rna(sgrna)与crispr效应蛋白在sgrna的核苷酸位置49处的核苷酸处交联,其中核苷酸位置1位于crrna的靶结合区的5’末端处,并且sgrna的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。核苷酸位置49处的核苷酸可以包含尿嘧啶。crispr复合物可以包括核酸酶活性。在一些实施方案中,crispr复合物可以包含单引导rna(sgrna),所述单引导rna(sgrna)与crispr效应蛋白在sgrna内的非天然核苷酸处交联,其中crispr复合物包括核酸酶活性。
10.本文公开了一种药物制剂,所述药物制剂包含crispr复合物和药学上可接受的赋形剂。还公开了一种方法,所述方法包括向受试者施用药物制剂。
11.本文公开了一种方法,所述方法包括将crispr复合物引入细胞中。还公开了一种试剂盒,所述试剂盒包含crispr复合物和说明书。
12.本文公开了一种编辑核酸分子的方法,所述方法包括使crispr复合物与核酸分子接触。crispr复合物可以包括小于2%裂解事件的脱靶裂解活性。
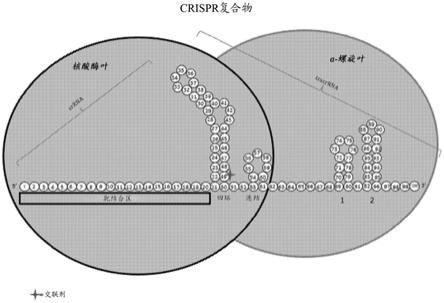
13.本文公开了一种在多于一个细胞中编辑靶基因的方法,所述方法包括将crispr复合物施用至包含靶基因的多于一个细胞中,从而产生包含编辑的靶基因的细胞,其中99%的包含编辑的靶基因的细胞在施用crispr复合物之后保持存活。细胞存活力可以通过刃天青测定来测量。
14.本文公开了一种产生crispr复合物的方法,所述方法包括将包含crrna区和tracrrna区的sgrna与crispr效应蛋白交联,其中交联发生在sgrna的crrna区之外的非天然核苷酸处,其中crispr效应蛋白的核酸酶活性在交联之后被维持。非天然核苷酸可以包含尿嘧啶。非天然核苷酸可以包含马来酰亚胺。交联可以在尿嘧啶和crispr效应蛋白上的半胱氨酸之间。尿嘧啶可以包括含4
‑
硫代尿苷。交联可以在尿嘧啶和crispr效应蛋白上的胺基团之间。尿嘧啶可以包括5
‑
溴尿苷。交联可以在溶液中发生,并且溶液中sgrna与crispr效应蛋白的比例可以是至少9:1。交联可以包括将溶液暴露于uv光。交联可以在sgrna与crispr效应蛋白混合时发生。
15.本文公开了一种方法,所述方法包括将包含含有交联剂的非天然核苷酸的单引导rna(sgrna)与crispr效应蛋白交联,其中交联发生在sgrna的靶结合区之外的非天然核苷酸处,从而产生交联复合物,其中交联复合物包括核酸酶活性。
16.本文公开了一种单引导rna(sgrna),所述单引导rna(sgrna)包含crrna区和tracrrna区以及核苷酸位置49处的非天然核苷酸,其中核苷酸位置1位于crrna区的靶结合区的5’末端处,并且sgrna的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。
17.本文公开了一种单引导rna(sgrna),所述单引导rna(sgrna)包含crrna区和tracrrna区以及核苷酸位置49处的尿嘧啶,其中核苷酸位置1位于crrna区的靶结合区的5’末端处,并且sgrna的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。
18.本文公开了一种crispr复合物,所述crispr复合物包含单引导rna(sgrna),所述单引导rna(sgrna)与crispr效应蛋白交联,其中sgrna包含crrna区、tracrrna区和被配置为调节crispr复合物活性的序列。sgrna可以是crispr on多核苷酸、crispr off多核苷酸、crispr on/off多核苷酸或被修饰以减少脱靶编辑的crispr多核苷酸。sgrna可以在sgrna中包含非天然核苷酸,并且sgrna与crispr效应蛋白在非天然核苷酸处交联。非天然核苷酸可以位于crrna区的靶结合区之外。非天然核苷酸可以位于sgrna的核苷酸位置49处,其中核苷酸位置1位于crrna的靶结合区的5’末端处,并且sgrna的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。非天然核苷酸可以包含糖的修饰。非天然核苷酸可以包含碱基的修饰。非天然核苷酸可以包含马来酰亚胺。马来酰亚胺可以与crispr效应蛋白上的半胱氨酸共价连接。非天然核苷酸可以包括吡啶基二硫化物、烷氧基胺、nhs酯、双吖丙啶、亚氨酸酯、卤代乙酰基基团、酰肼、芳基叠氮化物、异氰酸酯、二硫醇亚磷酰胺dtpa、4
‑
硫代
‑
utp、5
‑
叠氮基
‑
utp、5
‑
溴
‑
utp、8
‑
叠氮基
‑
atp、5
‑
apas
‑
utp或8
‑
n(3)amp。非天然核苷酸可以位于tracrrna区的茎环中。该茎环的结构可以相对于缺少非天然核苷酸的sgrna的茎环的结构被维持。非天然核苷酸可以位于tracrrna区的凸起中。该凸起的结构相对于缺少非天然核苷酸的sgrna的凸起的结构可以被维持。非天然核苷酸可以位于tracrrna区的茎环之间。crispr复合物可以包括核酸酶活性。crispr复合物的脱靶核酸酶活性可以等于或小于包含未交联的crispr效应蛋白和sgrna的crispr复合物的脱靶核酸酶活性。非天然核苷酸可以位于crispr效应蛋白的半胱氨酸的20埃以内。在一些实施方案中,非天然核苷酸可以不是4
‑
硫代尿苷或修饰的腺苷。
19.本文公开了一种包含修饰的多核苷酸,其中所述多核苷酸包含:(i)被配置为与靶核酸分子中的靶序列退火的引导序列,(ii)被配置为与crispr效应蛋白结合并包含修饰的序列,和(iii)被配置为与crispr效应蛋白交联的非天然核苷酸;其中当多核苷酸与crispr效应蛋白复合时,形成第一crispr复合物,与包含与crispr效应蛋白复合的不含修饰的多核苷酸的第二crispr复合物相比,第一crispr复合物具有更低的脱靶核酸分子编辑活性。非天然核苷酸可以位于位置49处。修饰可以包括不包含典型核苷酸碱基的接头。修饰可以包括至少两个不包含典型核苷酸碱基的接头。ii)的序列可以从5’至3’形成四环(tetraloop)、第一茎环、第二茎环和第三茎环。在一些情况下,多核苷酸不包含第四茎环。在一些情况下,多核苷酸在多核苷酸的5’末端处不包含茎环。接头可以包括可裂解接头。接头可以包括3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺。接头可以包括光不稳定接头。光不稳定接头可以被紫外辐射裂解。光不稳定接头可以被可见光裂解。可裂解接头可以包括3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺。可裂解接头可以包括1
‑
(7
‑
(二乙基氨基)
‑2‑
氧代
‑
2h
‑
色烯
‑4‑
基)丙基。可裂解接头可以包括其中*表示与h或第一核苷酸的附接点,并且**表示与oh或第二核苷
酸的附接点。光不稳定接头可以包括亚磷酰胺。光不稳定接头可以包括香豆素。修饰可以位于多核苷酸的位置57或位置74处,其中位置1位于多核苷酸的5’末端处,并且位置从5’至3’计数。修饰可以位于多核苷酸的位置57和位置74处。修饰可以位于环中。修饰可以位于第一茎环或第二茎环中。修饰可以位于第一茎环的环或第二茎环的环中。修饰可以位于位置57和位置74中的一个或两个处,其中位置1位于多核苷酸的5’末端处,并且位置从5’至3’计数。修饰可以包括可光裂解的键。在一些情况下,修饰不位于茎环中。多核苷酸可以在前三个5’末端rna核苷酸和前三个3’末端rna核苷酸处包含2
’‑
o
‑
甲基类似物和3’硫代磷酸酯核苷酸间连接。编辑活性可以以被编辑的脱靶核酸分子的百分比来测量。第一crispr复合物对脱靶核酸分子的编辑活性可以低于第二crispr复合物的编辑活性,其中p值≤0.0001。第一crispr复合物对靶核酸分子的编辑活性和第二crispr复合物对靶核酸分子的编辑活性可以在5%以内。第一crispr复合物对靶核酸分子的编辑活性和第二crispr复合物对靶核酸分子的编辑活性可以以被编辑的靶核酸分子的百分比来测量。本文公开了一种crispr复合物,所述crispr复合物包含任何以上提及的多核苷酸和crispr酶。crispr复合物可以包括核酸酶活性。
[0020]
在另一方面,本文描述了一种核苷酸或寡核苷酸,所述核苷酸或寡核苷酸包含式(i)的接头:
[0021][0022]
其中:r1、r2、r3、r4和r5各自独立地选自h、烷基(alkyl)、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;可选地,r1、r2、r3和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0023]
m可以是选自1至10的整数;x可以选自o、s、=c(cn)2;*可以表示与h或戊糖部分的附接点;并且**可以表示与oh或核苷酸的磷酸基团的附接点。式(i)的接头可以由式(i’)表示:
[0024][0025]
其中:r1、r2、r
3a
、r
3b
、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;可选地,r2、r
2a
、r
3a
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;x可以是氧、s或=c(cn)2。r1、r2、r4和r5可以各自独立地是h或c1‑6烷基;并且r
3a
和r
3b
可以是c1‑6烷基。r1、r2、r4和r5各自可以是h;并且r
3a
和r
3b
各自可以是乙基。
[0026]
在另一方面,本文提供了一种化合物,该化合物包含
[0027]
本文公开了一种多核苷酸,该多核苷酸包含以上提及的化合物。多核苷酸还可以包含被配置为结合crispr酶的序列。多核苷酸还可以包含被配置为与靶核酸分子中的靶序列退火的引导序列。本文公开了一种crispr复合物,该crispr复合物包含crispr酶和以上提及的多核苷酸。
[0028]
在一另方面,本文描述了一种化合物,该化合物包含式(i):
[0029][0030]
其中:r1、r2、r3、r4和r5各自独立地选自h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺
酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;可选地,r1、r2、r3和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0031]
m可以是选自1至10的整数;x可以选自o、s、=c(cn)2;*可以表示与h或戊糖部分的附接点;并且**可以表示与oh或核苷酸的磷酸基团的附接点。式(i)的化合物可以由式(i’)表示:
[0032][0033]
其中:r1、r2、r
3a
、r
3b
、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;可选地,r2、r
2a
、r
3a
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;x可以是氧、s或=c(cn)2。r1、r2、r4和r5可以各自独立地是h或c1‑6烷基;并且r
3a
和r
3b
可以是c1‑6烷基。r1、r2、r4和r5各自可以是h;并且r
3a
和r
3b
各自可以是乙基。
[0034]
通过引用并入
[0035]
本说明书中提及的所有出版物、专利和专利申请通过引用并入本文,其程度如同每个单独的出版物、专利或专利申请被具体和单独地指明通过引用并入的相同程度。
[0036]
附图简述
[0037]
本发明的新颖特征在所附权利要求书中具体地阐述。通过参考以下详细描述和附图将获得对本发明的特征和优点的更好理解,该详细描述阐述了利用本发明的原理的说明性实施方案,在附图中:
[0038]
图1示出了crispr复合物的简化图,其中多核苷酸是单引导rna(sgrna)。星号表示用于将多核苷酸与cas核酸酶交联的非天然核苷酸的示例性位置。条形表示多核苷酸的靶结合区。
[0039]
图2示出了与靶序列结合的sgrna的3
‑
d模型;突出显示了四环和茎环1
‑
3,以将该图与图3
‑
图5相关联(图片修改自nishimasu,h.,ishitani,r.,&nureki,o.(2014).crystal structure of streptococcus pyogenes cas9 in complex with guide rna and target dna.cell.doi:10.2210/pdb4oo8/pdb)。
[0040]
图3示出了连结(nexus)内结合的sgrna核苷酸与相邻的cas9核酸酶的氨基酸的相互作用的图。
[0041]
图4示出了茎环1内结合的sgrna核苷酸与相邻的cas9核酸酶的氨基酸的相互作用的图。
[0042]
图5示出了茎环2内结合的sgrna核苷酸与相邻的cas9核酸酶的氨基酸的相互作用的图。
[0043]
图6示出了显示示例性的待修饰的rna核苷酸和蛋白上的交联位点的晶体结构。
[0044]
图7a
‑
图7d示出了sgrna的野生型构型和对sgrna的三种示例性修饰的图。
[0045]
图8示出了概述可以如何将马来酰亚胺添加至尿嘧啶核苷酸的实例的图。
[0046]
图9示出了crispr复合物的晶体结构,其中突出显示了sgrna中的所有尿嘧啶碱基。
[0047]
图10示出了crispr复合物的晶体结构,聚焦于sgrna的位置44处的尿嘧啶和包含酪氨酸残基的α螺旋之间的空间关系。
[0048]
图11示出了crispr复合物的晶体结构,聚焦于位置59处的尿嘧啶与crispr效应蛋白的二级结构之间的空间关系。
[0049]
图12示出了crispr复合物的晶体结构,聚焦于位置66处的尿嘧啶与crispr效应蛋白的二级结构之间的空间关系。
[0050]
图13示出了crispr复合物的晶体结构,聚焦于位置63处的尿嘧啶与crispr效应蛋白的二级结构之间的空间关系。
[0051]
图14示出了三个示例性交联反应。
[0052]
图15示出了其中使用锁定的crispr复合物来裂解基因序列的实验的结果。x轴表示被引导序列靶向的基因,y轴表示被裂解的含有靶向基因的dna序列的百分比。
[0053]
图16示出了锁定的crispr复合物的纯化的示例性图。
[0054]
图17示出了crispr
‑
on靶向裂解活性的激活的示例性模型。无活性crispr复合物包含crispr
‑
on单引导rna(sgrna),所述crispr
‑
on单引导rna(sgrna)包含位于与crispr效应蛋白cas9复合的典型sgrna的5’末端处的添加的茎环结构。茎环结构可以抑制复合物的活性,产生无活性的复合物。添加裂解媒介(cleavage agent)可以释放茎环结构,产生可以允许基因组编辑发生的活性(on)crispr复合物。
[0055]
图18示出了crispr
‑
off sgrna中可裂解接头的示例性位置。
[0056]
图19示出了可激活的crispr
‑
on sgrna变体的裂解功效。将包含通过uv敏感的可裂解接头与引导序列隔开的5’茎环元件的crispr
‑
on sgrna暴露于uv光0分钟、5分钟、10分钟或15分钟。暴露15分钟后,sgrna展示出与引导序列5’侧序列的裂解一致的条带模式。“对照”泳道是缺少任何另外的引导序列5’侧序列的sgrna,而“无二级(no 2nd)”条件使用具有向引导序列的非茎形成性5’侧添加的sgrna。“3bp茎”和“6bp茎”条件使用被设计为在引导序列的5’末端处分别具有3bp和6bp长度的茎区域的sgrna。
[0057]
图20示出了体外crispr
‑
on sgrna激活靶dna裂解的功效。将具有5’可裂解茎环的crispr
‑
on sgrna与靶dna(人类fancf)一起孵育1小时,并且以规律间隔暴露于裂解媒介uva光(320
‑
390nm)。“mod”是被修饰为在前三个5’末端rna核苷酸和前三个3’末端rna核苷酸处包含2
’‑
o
‑
甲基类似物和3’硫代磷酸酯核苷酸间连接并且缺少向引导序列的任何5’侧添加碱基的sgrna。包括对sgrna的标准修饰。“无二级结构(no secondary)”条件使用具有向引导序列的非茎形成性5’侧添加的sgrna。“3bp茎”和“6bp茎”条件使用在sgrna的5’末端
处具有被设计为分别形成3bp和6bp的茎区域的序列的sgrna。
[0058]
图21示出了可失活的crispr
‑
off sgrna变体的裂解功效。使具有五种不同裂解点的sgrna经历裂解媒介(uv光)0分钟(左图)或5分钟(右图)。
[0059]
图22示出了细胞中基因组编辑效率的时间依赖性crispr
‑
off失活的示意图。将转染了可失活的sgrna变体的细胞在rnp递送后的时间点用uv光处理,并且允许在rnp递送后的总计48小时进行编辑、修复和恢复。在48小时之后,从所有样品收集基因组dna,并且分析插入/缺失(indel)的存在。两种crispr
‑
off sgrna(57和74)展示出基因组编辑活性的时间依赖性增加。
[0060]
图23示出了其中使用crispr off复合物来裂解基因序列的实验的结果。x轴表示被引导序列靶向的基因以及引导序列的形式,并且y轴表示包含被编辑的靶向基因的dna序列的百分比。
[0061]
图24示出了作为对应于图23的实验的对照运行的实验的结果,其中使用包含标准sgrna的crispr复合物来裂解基因序列。x轴表示被引导序列靶向的基因以及引导序列的形式,并且y轴表示包含被编辑的靶向基因的dna序列的百分比。
[0062]
图25示出了其中使用crispr off复合物来裂解基因序列的实验的结果。x轴表示被引导序列靶向的基因以及引导序列的形式,并且y轴表示包含被编辑的靶向基因的dna序列的百分比。
[0063]
图26示出了作为对应于图25的实验的对照运行的实验的结果,其中使用包含标准sgrna的crispr复合物来裂解基因序列。x轴表示被引导序列靶向的基因以及引导序列的形式,并且y轴表示包含被编辑的靶向基因的dna序列的百分比。
[0064]
图27示出了其中使用crispr off复合物来裂解基因序列的实验的结果。x轴表示被引导序列靶向的基因以及引导序列的形式,并且y轴表示包含被编辑的靶向基因的dna序列的百分比。
[0065]
图28示出了作为对应于图27的实验的对照运行的实验的结果,其中使用包含标准sgrna的crispr复合物来裂解基因序列。x轴表示被引导序列靶向的基因以及引导序列的形式,并且y轴表示包含被编辑的靶向基因的dna序列的百分比。
[0066]
图29示出了一系列散点图,这些散点图比较了使用crispr off sgrna或修饰的sgrna在遍及三个基因靶的最高预测的脱靶位点处的脱靶编辑活性。与仅被修饰为在前三个5’末端rna核苷酸和前三个3’末端rna核苷酸处包含2
’‑
o
‑
甲基类似物和3’硫代磷酸酯核苷酸间连接的sgrna相比,crispr off sgrna引起显著更少的脱靶插入/缺失。(****p<0.0001,student非配对t检验,n=24个技术重复)。
[0067]
图30示出了靶向dnmt1的crispr off复合物与包含靶向dnmt1的标准sgrna的crispr复合物相比的时间依赖性编辑活性。
[0068]
图31示出了靶向grk1的crispr off复合物与包含靶向grk1的标准sgrna的crispr复合物相比的时间依赖性编辑活性。
[0069]
图32示出了靶向vegfa的crispr off复合物与包含靶向vegfa的标准sgrna的crispr复合物相比的时间依赖性编辑活性。
[0070]
图33示出了crispr复合物的简化图,其中多核苷酸是单引导rna(sgrna)。四角星号表示用于将多核苷酸与cas核酸酶交联的非天然核苷酸的示例性位置。五角星号表示可
裂解接头的示例性位置。条形表示多核苷酸的靶结合区。
[0071]
图34示出了可以对crispr多核苷酸进行包含香豆素接头的修饰的示例性位置。
[0072]
图35示出了与不含可裂解接头的修饰的sgrna相比,可以对crispr多核苷酸进行包含可裂解接头的修饰的示例性位置。
[0073]
图36a是图34的完整crispr多核苷酸的电喷雾电离(esi)质谱示踪,展示出在不存在光的情况下未观察到片段化。
[0074]
图36b是图34的crispr多核苷酸在光裂解后的电喷雾电离(esi)质谱示踪,展示出多核苷酸在暴露于波长大于420nm的光时在两个可光裂解位点处被裂解。
[0075]
图37是示出多核苷酸片段的比较的凝胶图片,所述多核苷酸片段对应于暴露于uv光之后在位置57和74处的可光裂解接头裂解之后产生的片段、暴露于uv光之后产生的片段和完整的sgrna。
[0076]
图38是比较在hek293细胞中靶向23个不同的靶位点的23种引导rna(包含图34的可光裂解位点)的定量为编辑百分比的性能的图(比较了三种条件:无光、环境光或波长大于345nm的光)。
[0077]
图39是比较在hek293细胞中靶向23个不同的靶位点的23种引导rna(在位置57和74处包含可光裂解接头)与不含可光裂解位点的sgrna相比的定量为编辑百分比的性能的图(比较了三种条件:无光、环境光或波长大于345nm的光)。
[0078]
图40是比较在hep3b细胞中靶向18个不同的靶位点的18种引导rna(在位置57和74处包含可光裂解接头)与不含可光裂解位点的sgrna相比的定量为编辑百分比的性能的图(比较了三种条件:无光、环境光或波长大于345nm的光)。
[0079]
图41是比较在u2os细胞中靶向13个不同的靶位点的13种引导rna(在位置57和74处包含可光裂解接头)与不含可光裂解位点的sgrna相比的定量为编辑百分比的性能的图(比较了三种条件:无光、环境光或波长大于345nm的光)。
[0080]
图42示出了sgrna有活性的时间量和中靶编辑与脱靶编辑的比率之间的关系,展示为以未修饰的sgrna为对照,脱靶编辑随着sgrna被允许有活性的时间加长而增加。
[0081]
图43是示出在细胞中观察到的编辑百分比随着暴露于385nm的光的增加而降低的图。
[0082]
图44是细胞板的图像,该细胞板被选择性掩蔽以防止一些细胞暴露于光,使得在被保持在黑暗中的那些细胞中gfp基因被敲除,而暴露于光的那些细胞表达gfp。
[0083]
图45是示出与不含可光裂解接头的sgrna相比图34的多核苷酸在暴露于光后丰度显著降低的图。
[0084]
图46是示出与不含可光裂解接头的sgrna相比图34的多核苷酸在暴露于光后编辑百分比显著降低的图。
[0085]
图47是示出在hek293细胞中观察到的与cas9核酸酶复合的图34的多核苷酸的编辑活性百分比随时间变化的图,其中每个时间点表示测试的hek293细胞群体暴露于光的时间。
[0086]
图48是示出图34的crispr多核苷酸通过使用与图27相同的方案暴露于具有430
±
23nm波长的光而失活的时间框架的图。
[0087]
图49是示出当位于沿着sgrna的不同位置处的可裂解接头被激活时获得的裂解产
物的凝胶。
[0088]
图50a
‑
图50c示出了当靶向不同基因时各种crispr off可裂解接头位置的编辑活性图。
[0089]
图51示出了光暴露持续时间对编辑消除的影响的图,其中在45
‑
60秒之间实现了完全消除。
[0090]
图52是示出增加细胞暴露于广谱光的时间对细胞存活力的影响的图。
[0091]
图53是靶向camk1的与cas9核酸酶复合的crispr off多核苷酸相比于与cas9核酸酶复合的标准sgrna的插入/缺失谱。
[0092]
图54是细胞培养物的图片,其中与cas9核酸酶复合的图34的多核苷酸用于靶向必需基因。暴露于光( hv)的细胞培养物展示出比未暴露于光的细胞培养物更高的汇合,表明缺乏失活引起高度的细胞死亡。
[0093]
图55是示出在转染后不同时间点的中靶编辑:脱靶编辑的比率的图。
[0094]
图56是施加至图44的细胞培养物的薄膜掩蔽物的图片,使得透明区域允许光通过,使与crispr off复合的cas9核酸酶的编辑活性失活,而黑暗区域是不透明的,以允许编辑不受阻碍地进行。
[0095]
图57a是在位置57和74处具有可光裂解接头的完整crispr多核苷酸的电喷雾电离(esi)质谱示踪,展示出在不存在光的情况下未观察到片段化。
[0096]
图57b是在位置57和74处具有可光裂解接头的crispr off多核苷酸在光裂解后的电喷雾电离(esi)质谱示踪,展示出多核苷酸在暴露于波长大于345nm的光时在两个可光裂解位点处被裂解。
[0097]
图58示出了crispr on v1 sgrna的结构。该sgrna的结构与酿脓链球菌(streptococcus pyogenes)cas9(spcas9)sgrna相同,但包含与前间区互补的20个核苷酸(nt)序列(折回序列(backtrack sequence)),随后是紧接前间区5’侧的4nt环结构。
[0098]
图59示出了使用靶向四个独特基因座的crispr on v1 sgrna的三种变体进行的编辑。三种变体包括:一种预期对光刺激没有响应(非可裂解对照),一种在距sgrna的5’末端的位置24处包含单个可光裂解接头(1个可光裂解),以及一种在位置11和位置24处包含两个可光裂解接头(2个可光裂解)。
[0099]
图60示出了crispr on v2 sgrna的结构。crispr on v2使用与crispr on v1相同的结构,但是crispr on v2用2
’‑
o
‑
甲基(2`o
‑
me)rna替代折回序列(与前间区互补的前20nt的rna)。2`o
‑
me rna与rna更紧密结合,并且在r
‑
环形成期间较不可能被替代。
[0100]
图61示出了标准sgrna(mod)、crispr on v1(rna)和crispr on v2(o
‑
me)之间编辑活性的比较。
[0101]
图62示出了crispr on v3 sgrna的结构。crispr on v3基于crispr on v2构建,但是crispr on v3在前间区折回序列的中间(位置11)和紧接前间区序列的5’侧(位置24)掺入了可光裂解接头。
[0102]
图63示出了与在位置24(k24)处包含单个可光裂解接头的crispr on v2变体、crispr on v2(o
‑
me)和标准sgrna(mod)相比,使用靶向5个独特基因座的crispr on v3 sgrna(k11,24)进行的编辑。
[0103]
图64示出了crispr on v4 sgrna的结构。crispr on v4基于crispr on v3构建,
但引入了另外的可光裂解接头,以确保折回区域被dna靶有效替代。可光裂解残基位于位置23和位置24处,以增加折回序列从sgrna释放的可能性。另外的可光裂解残基位于位置6和位置14处,以辅助解离。
[0104]
图65示出了与5rp(在sgrna的5’侧包含另外的序列5
’‑
ucucccugagcuucagggag
‑3’
)、crispr on v2(me)和标准sgrna(mod)相比,使用crispr on v4 sgrna变体在两个基因座处进行的编辑。crispr on v4 sgrna变体在以下核苷酸处包含可光裂解接头:3、23和24(k3,23,24);6、11、16、23和24(k6,11,16,23,24);6、14、23和24(k6,14,23,24)。
[0105]
详细描述
[0106]
i.概述
[0107]
本文公开了一种多核苷酸(crispr多核苷酸),所述多核苷酸(crispr多核苷酸)包含被设计为与靶核酸序列退火的序列和被设计为结合crispr效应蛋白的序列,其中crispr多核苷酸包含交联剂。交联剂可以在多核苷酸的发夹区。在另一方面,本文提供了一种crispr复合物,所述crispr复合物包含crispr多核苷酸和crispr效应蛋白。crispr多核苷酸可以被设计为与crispr效应蛋白(例如,cas酶)结合,以形成crispr复合物。cas酶可以是cas9、cas12a、cas12b等。本文还提供了用于将crispr多核苷酸与crispr效应蛋白交联以形成交联的crispr复合物的方法。例如,crispr多核苷酸可以与cas酶共价键合,例如,通过交联反应激活来共价键和,交联反应通过暴露于紫外范围内特定波长的光或通过将非天然核苷酸放置在sgrna内来激活,所述非天然核苷酸在非常靠近靶氨基酸侧链时将形成共价键。
[0108]
在另一方面,本文提供了一种crispr复合物,所述crispr复合物包含:a)crispr多核苷酸,所述crispr多核苷酸包含被设计为与靶核酸序列退火的序列、被设计为结合crispr效应蛋白的序列,具有或没有可被调节以影响活性的一个或更多个元件;和b)crispr效应蛋白,其中crispr多核苷酸与crispr效应蛋白结合的平衡解离常数(k
d
)小于8pm。
[0109]
在另一方面,crispr多核苷酸可以包含(i)被配置为与crispr效应蛋白共价结合的序列,(ii)任选地,被配置为与靶分子中的靶序列退火的引导序列,和(iii)可以被调节以影响与crispr多核苷酸复合的crispr效应蛋白的活性的一个或更多个元件。与crispr多核苷酸复合的crispr效应蛋白可以被认为是“可调的”。在一些情况下,一个或更多个元件可以被调节以增加与crispr多核苷酸复合的crispr效应蛋白(例如,crispr“on”复合物)的活性。在一些情况下,一个或更多个元件可以被调节以降低与crispr多核苷酸复合的crispr效应蛋白(例如,crispr“off”复合物)的活性。在一些情况下,crispr多核苷酸中的第一元件可以被调节以增加与crispr多核苷酸复合的crispr效应蛋白的活性,并且第二元件可以被调节以降低与crispr多核苷酸复合的crispr效应蛋白的活性(例如,crispr“on/off”复合物)。
[0110]
本文还提供了包含与crispr多核苷酸交联的crispr效应蛋白的复合物(例如,crispr on复合物、crispr off复合物或crispr on/off复合物)。在一些情况下,交联可以在crispr多核苷酸中的非天然核苷酸处。本文提供了调节crispr多核苷酸的方法。提供了试剂盒,所述试剂盒包含多核苷酸和例如说明书以及任选地crispr效应蛋白。此外,提供了包含crispr多核苷酸和药学上可接受的赋形剂的药物制剂以及施用药物制剂的方法。本文还提供了将crispr多核苷酸引入细胞中的方法。
[0111]
本文提供了利用crispr多核苷酸和crispr复合物的方法和试剂盒。例如,本文提供了这样的方法,所述方法包括使靶核酸序列与crispr复合物接触。此外,本文提供了一种药物制剂,所述药物制剂包含crispr多核苷酸和/或crispr复合物以及药学上可接受的赋形剂。在另一方面,本文提供了一种方法,所述方法包括向受试者施用药物制剂。此外,本文提供了一种方法,所述方法包括将crispr复合物引入细胞中。
[0112]
本文还提供了试剂盒,所述试剂盒包含crispr多核苷酸和/或crispr复合物。
[0113]
ii.crispr概述
[0114]
本文提供了具有增强的稳定性的crispr/cas复合物。本文提供了具有增强的稳定性和可调的活性的crispr/cas复合物。crispr(成簇规律间隔短回文重复序列)可以是存在于原核生物基因组中的来源于原核生物先前遇到的病毒的dna片段的dna序列家族。crispr效应蛋白(例如,cas核酸酶)可以与来源于该dna序列的crispr多核苷酸(例如,rna)结合,并且也可以与靶区域:与crispr多核苷酸序列互补的(病毒)dna序列结合。结合后,cas核酸酶可以在靶(病毒)dna的靶区域进行双链切割,以便使其失活。靶区域可以包含“前间区(protospacer)”和“前间区邻近基序(protospacer adjacent motif)”(pam),并且这两个结构域都可能是cas酶介导的活性(例如,裂解)所需的。靶位点可以与核酸酶(例如,cas9、c2c1、c2c3或cpf1)的pam位点邻近。cas核酸酶可以是cas9。pam位点可以是被crispr效应蛋白识别的短序列,并且在某些情况下是cas酶活性所需的,例如,pam位点可以是ngg。pam位点的核苷酸序列和数目可以取决于crispr效应蛋白(例如,cas酶)的类型而不同。前间区序列可以称为靶位点(或基因组靶位点)。crispr多核苷酸可以与前间区(结合位点)的相对链配对(或杂交),将cas酶引导至靶区域。
[0115]
a.crispr复合物概述
[0116]
crispr复合物可以是包含一种或更多种dna或rna靶向crispr效应蛋白和一种或更多种crispr多核苷酸的非天然存在的或工程化的dna或rna靶向系统。一种或更多种crispr多核苷酸可以是本文提供的任何crispr多核苷酸。靶序列可以是crispr多核苷酸的引导序列被设计为与其具有互补性的序列,并且“互补性”可以指一种核酸通过常规的沃森
‑
克里克碱基配对或其他非常规类型的碱基配对与另一种核酸序列形成氢键的能力。crispr复合物可以与形成双链结构的两条核酸链、形成多链复合物的三条或更多条链、单个自杂交链或这些链的任何组合相互作用。
[0117]
在crispr复合物与靶序列结合后,与靶序列缔合的序列可以被crispr效应蛋白修饰。crispr效应蛋白可以是融合蛋白的一部分,该融合蛋白可以包含一个或更多个异源蛋白结构域(例如,除了crispr效应蛋白之外,约以下,或多于约以下:1个、2个、3个、4个、5个、6个、7个、8个、9个、10个或更多个结构域)。在一些实例中,crispr复合物的功能由异源蛋白结构域赋予。
[0118]
在一些情况下,crispr系统的一个或更多个元件可以来源于i型、ii型或iii型crispr系统。在crispr ii型系统中,crispr多核苷酸(例如,引导rna)可以与cas内切核酸酶相互作用,并且将cas酶的核酸酶活性引导至靶区域。靶区域可以包含“前间区”和“前间区邻近基序”(pam),并且这两个结构域都可以用于cas酶介导的活性(例如,裂解)。引导序列可以与前间区的相对链(结合位点)配对(或杂交),将cas酶引导至靶区域。pam位点可以指由cas酶识别的短序列,并且在一些情况下,是cas酶活性所需的。pam位点的核苷酸序列
thermophilus)、肉毒杆菌(clostridium botulinum)、解纤维梭菌(clostridium cellulolyticum)、大芬戈尔德菌(finegoldia magna)、直肠真杆菌(eubacterium rectale)、鸡败血支原体(mycoplasma gallisepticum)、运动支原体(mycoplasma mobile)、穿通支原体(mycoplasma penetrans)、滑液囊支原体(mycoplasma synoviae)、念珠状链杆菌(streptobacillus moniliformis)、慢生根瘤菌属(bradyrhizobium)、汉氏硝化细菌(nitrobacter hamburgensis)、沼泽红假单胞菌(rhodopseudomonas palustris)、parvibaculum lavamentivorans、dinoroseobacter shibae、gluconacetobacter diazotrophicus、固氮螺菌属(azospirillum)、深红红螺菌(rhodospirillum rubrum)、acidovorax ebreus、verminephrobacter eiseniae、需盐脱硫弧菌(desulfovibrio salexigens)、空肠弯曲杆菌(campylobacter jejuni)、红嘴鸥弯曲杆菌(campylobacter lari)、肝螺杆菌(helicobacter hepaticus)、产琥珀酸沃廉菌(wolinella succinogenes)、tolumonas auensis、pseudoalteromonas atlantica、shewanella pealeana、嗜肺军团菌(legionella pneumophila)、产琥珀酸放线杆菌(actinobacillus succinogenes)、多杀性巴氏杆菌(pasteurella multocida)、新凶手弗朗西丝氏菌(francisella novicida)、土拉弗朗西斯菌(francisella tularensis)或齿垢密螺旋体(treponema denticola)。
[0122]
cas蛋白可以是来自以下的i型cas7或cas 1:aeropyrum pernix、desulfurococcus kamchatkensis、ignicoccus hospitalis、staphylothermus marinus、hyperthermus butylicus、勤奋金属球菌(metallosphaera sedula)、冰岛硫化叶菌(sulfolobus islandicus)、硫磺矿硫化叶菌(sulfolobus solfataricus)、sulfolobus tokodaii、下垂热丝菌(thermofilum pendens)、caldivirga maquilingensis、pyrobaculum aerophilum、pyrobaculum arsenaticum、pyrobaculum calidifontis、嗜中性热变形菌(thermoproteus neutrophilus)、archaeoglobus fulgidus、ferroglobus placidus、死海盐盒菌(haloarcula marismortui)、halomicrobium mukohataei、halorhabdus utahensis、嗜冷嗜盐菌(halorubrum lacusprofundi)、natronomonas pharaonis、反刍兽甲烷短杆菌(methanobrevibacter ruminantium)、史氏甲烷短杆菌(methanobrevibacter smithii)、methanosphaera stadtmanae、热自养甲烷热杆菌(methanothermobacter thermautotrophicus)、methanocaldococcus fervens、詹氏甲烷暖球菌(methanocaldococcus jannaschii)、甲烷暖球菌属(methanocaldococcus)、methanocaldococcus vulcanius、methanococcus aeolicus、methanococcus maripaludis、万氏甲烷球菌(methanococcus vannielii)、methanocorpusculum labreanum、亨氏甲烷螺菌(methanospirillum hungatei)、methanosphaerula palustris、嗜热甲烷鬃菌(methanosaeta thermophila)、methanococcoides burtonii、乙酸甲烷八叠球菌(methanosarcina acetivorans)、巴氏甲烷八叠球菌(methanosarcina barkeri)、马氏甲烷八叠球菌(methanosarcina mazei)、pyrococcus abyssi、强烈火球菌(pyrococcus furiosus)、掘越氏热球菌(pyrococcus horikoshii)、thermococcus gammatolerans、thermococcus kodakarensis、thermococcus sibiricus、picrophilus torridus、candidatus korarchaeum cryptofilum、nanoarchaeum equitans、acidimicrobium ferrooxidans、catenulispora acidiphila、corynebacterium aurimucosum、白喉棒杆菌、
谷氨酸棒杆菌、杰氏棒杆菌(corynebacterium jeikeium)、解脲棒杆菌(corynebacterium urealyticum)、皮疽诺卡菌、红串红球菌(rhodococcus erythropolis)、弗兰克氏菌(frankia alni)、桤木弗兰克氏菌属(frankia)、nakamurella multipartita、粘滑罗斯菌(rothia mucilaginosa)、xylanimonas cellulosilytica、salinispora arenicola、salinispora tropica、奇迹束丝放线菌(actinosynnema mirum)、绿色糖单孢菌(saccharomonospora viridis)、阿维链霉菌(streptomyces avermitilis)、灰色链霉菌(streptomyces griseus)、褐色嗜热裂孢菌(thermobifida fusca)、弯曲高温单孢菌(thermomonospora curvata)、青春双歧杆菌(bifidobacterium adolescentis)、动物双歧杆菌(bifidobacterium animalis)、齿双歧杆菌、阴道加德纳菌(gardnerella vaginalis)、迟缓埃格特菌(eggerthella lenta)、rubrobacter xylanophilus、aquifex aeolicus、hydrogenobacter thermophilus、hydrogenobaculum、thermocrinis albus、persephonella marina、sulfurihydrogenibium azorense、sulfurihydrogenibium、脆弱拟杆菌、parabacteroides distasonis、牙龈卟啉单胞菌(porphyromonas gingivalis)、spirosoma linguale、海洋红嗜热盐菌(rhodothermus marinus)、chlorobaculum tepidum、chlorobium chlorochromatii、泥生绿菌(chlorobium limicola)、chlorobium phaeobacteroides、chlorobium phaeovibrioides、pelodictyon luteolum、pelodictyon phaeoclathratiforme、chloroherpeton thalassium、prosthecochloris aestuarii、chloroflexus aggregans、chloroflexus aurantiacus、绿曲挠丝状菌属(chloroflexus)、roseiflexus castenholzii、玫瑰弯菌属、橙色滑柱菌(herpetosiphon aurantiacus)、dehalococcoides、sphaerobacter thermophilus、玫瑰红嗜热菌(thermomicrobium roseum)、蓝丝菌属(cyanothece)、铜绿微囊藻(microcystis aeruginosa)、聚球藻属(synechococcus)、集胞藻属、多变鱼腥藻(anabaena variabilis)、点状念珠藻(nostoc punctiforme)、念珠藻属(nostoc)、deinococcus geothermalis、嗜热栖热菌(thermus thermophilus)、嗜热网球菌(dictyoglomus thermophilum)、dictyoglomus turgidum、acidobacterium capsulatum、酸热脂环酸杆菌(alicyclobacillus acidocaldarius)、anoxybacillus flavithermus、bacillus cytotoxicus、克劳氏芽孢杆菌(bacillus clausii)、嗜碱芽孢杆菌(bacillus halodurans)、地芽孢杆菌属(geobacillus)、lysinibacillus sphaericus、exiguobacterium sibiricum、单核细胞增多性李斯特菌(listeria monocytogenes)、斯氏李斯特菌(listeria seeligeri)、干酪乳杆菌、德氏乳杆菌(lactobacillus delbrueckii)、发酵乳杆菌(lactobacillus fermentum)、瑞士乳杆菌(lactobacillus helveticus)、马链球菌、变异链球菌、酿脓链球菌、alkaliphilus metalliredigens、肉毒杆菌、解纤维梭菌、艰难梭菌(clostridium difficile)、克氏梭菌(clostridium kluyveri)、诺氏梭菌(clostridium novyi)、产气荚膜梭菌(clostridium perfringens)、破伤风梭菌(clostridium tetani)、热纤梭菌(clostridium thermocellum)、大芬戈尔德菌、symbiobacterium thermophilum、直肠真杆菌、嗜中温螺旋杆菌(heliobacterium modesticaldum)、candidatus desulforudis audaxviator、desulfitobacterium hafniense、desulfotomaculum acetoxidans、desulfotomaculum reducens、pelotomaculum thermopropionicum、沃氏共养单胞菌(syntrophomonas wolfei)、anaerocellum thermophilum、发酵氨基酸球菌(acidaminococcus fermentans)、
luminescens、肠道沙门氏菌(salmonella enterica)、鲍氏志贺菌(shigella boydii)、弗氏志贺菌(shigella flexneri)、索氏志贺菌(shigella sonnei)、xenorhabdus bovienii、鼠疫杆菌(yersinia pestis)、假结核耶尔森菌(yersinia pseudotuberculosis)、伯氏考克斯体(coxiella burnetii)、嗜肺军团菌(legionella pneumophila)、荚膜甲基球菌(methylococcus capsulatus)、hahella chejuensis、需盐色盐杆菌(chromohalobacter salexigens)、海单胞菌属(marinomonas)、胸膜肺炎放线杆菌(actinobacillus pleuropneumoniae)、产琥珀酸放线杆菌、aggregatibacter actinomycetemcomitans、aggregatibacter aphrophilus、产琥珀酸曼氏杆菌(mannheimia succiniciproducens)、多杀巴斯德菌(pasteurella multocida)、鲍氏不动杆菌(acinetobacter baumannii)、不动杆菌(acinetobacter)、棕色固氮菌(azotobacter vinelandii)、cellvibrio japonicus、铜绿假单胞菌(pseudomonas aeruginosa)、门多萨假单胞菌(pseudomonas mendocina)、施氏假单胞菌(pseudomonas stutzeri)、费氏弧菌(vibrio fischeri)、photobacterium profundum、霍乱弧菌(vibrio cholerae)、哈氏弧菌(vibrio harveyi)、副溶血弧菌(vibrio parahaemolyticus)、黄单胞菌属(xanthomonas)、地毯草黄单孢菌(xanthomonas axonopodis)、水稻白叶枯病(xanthomonas oryzae)、magnetococcus、leptospira borgpetersenii、leptospira interrogans、多节闪烁杆菌(fervidobacterium nodosum)、kosmotoga olearia、petrotoga mobilis、非洲栖热腔菌(thermosipho africanus)、thermosipho melanesiensis、thermotoga lettingae、海栖热袍菌(thermotoga maritima)、那不勒斯栖热袍菌(thermotoga neapolitana)、thermotoga petrophila、热袍菌属(thermotoga)或thermobaculum terrenum。
[0123]
cas蛋白可以是来自以下的iii型cas10:desulfurococcus kamchatkensis、ignicoccus hospitalis、staphylothermus marinus、hyperthermus butylicus、勤奋金属球菌(metallosphaera sedula)、嗜酸热硫化叶菌(sulfolobus acidocaldarius)、冰岛硫化叶菌(sulfolobus islandicus)、硫磺矿硫化叶菌(sulfolobus solfataricus)、sulfolobus tokodaii、下垂热丝菌、caldivirga maquilingensis、pyrobaculum aerophilum、pyrobaculum arsenaticum、pyrobaculum calidifontis、冰岛热棒菌(pyrobaculum islandicum)、嗜中性热变形菌、archaeoglobus fulgidus、natronomonas pharaonis、反刍兽甲烷短杆菌、methanosphaera stadtmanae、热自养甲烷热杆菌、methanocaldococcus fervens、詹氏甲烷暖球菌、甲烷暖球菌属、methanocaldococcus vulcanius、methanococcus aeolicus、万氏甲烷球菌、亨氏甲烷螺菌、嗜热甲烷鬃菌、乙酸甲烷八叠球菌、巴氏甲烷八叠球菌、马氏甲烷八叠球菌、methanopyrus kandleri、强烈火球菌、掘越氏热球菌、thermococcus onnurineus、picrophilus torridus、thermoplasma volcanium、aciduliprofundum boonei、candidatus korarchaeum cryptofilum、牛分枝杆菌(mycobacterium bovis)、结核分支杆菌(mycobacterium tuberculosis)、弗兰克氏菌属、salinispora tropica、绿色糖单孢菌、红色糖多孢菌(saccharopolyspora erythraea)、嗜热裂孢菌、rubrobacter xylanophilus、aquifex aeolicus、thermocrinis albus、sulfurihydrogenibium azorense、sulfurihydrogenibium、牙龈卟啉单胞菌(porphyromonas gingivalis)、海洋红嗜热盐菌、chlorobaculum parvum、chlorobium phaeobacteroides、chlorobium phaeobacteroides、pelodictyon phaeoclathratiforme、
chloroherpeton thalassium、methylacidiphilum infernorum、chloroflexus aggregans、chloroflexus aurantiacus、绿曲挠丝状菌属、roseiflexus castenholzii、玫瑰弯菌属、橙色滑柱菌、玫瑰红嗜热菌、蓝丝菌属、铜绿微囊藻、聚球藻属、集胞藻属、多变鱼腥藻、点状念珠藻、念珠藻属、deinococcus geothermalis、嗜热栖热菌、嗜热网球菌、dictyoglomus turgidum、candidatus solibacter usitatus、产琥珀酸丝状杆菌、酸热脂环酸杆菌、嗜碱芽孢杆菌、地杆菌属、表皮葡萄球菌(staphylococcus epidermidis)、路邓葡萄球菌(staphylococcus lugdunensis)、血链球菌(streptococcus sanguinis)、嗜热链球菌、肉毒杆菌、破伤风梭菌、热纤梭菌、candidatus desulforudis audaxviator、desulfotomaculum acetoxidans、desulfotomaculum reducens、pelotomaculum thermopropionicum、沃氏共养单胞菌、anaerocellum thermophilum、小韦荣球菌(veillonella parvula)、奥氏嗜热盐丝菌、carboxydothermus hydrogenoformans、ammonifex degensii、thermoanaerobacter italicus、thermoanaerobacter pseudethanolicus、高温厌氧杆菌属、thermoanaerobacter tengcongensis、caldicellulosiruptor saccharolyticus、微小脲原体(ureaplasma parvum)、口腔纤毛菌、念珠状链杆菌、thermodesulfovibrio yellowstonii、pirellula staleyi、rhodospirillum centenum、深红红螺菌、欧洲亚硝化单胞菌、nitrosomonas eutropha、candidatus accumulibacter phosphatis、desulfococcus oleovorans、黄色黏球菌、haliangium ochraceum、纤维堆囊菌、syntrophus aciditrophicus、syntrophobacter fumaroxidans、布氏弓形杆菌(arcobacter butzleri)、胎儿弯曲菌、teredinibacter turnerae、allochromatium vinosum、halorhodospira halophila、thioalkalivibrio、dickeya dadantii、pectobacterium carotovorum、海单胞菌属、产琥珀酸曼氏杆菌、创伤弧菌(vibrio vulnificus)、fervidobacterium nodosum、kosmotoga olearia、非洲栖热腔菌、thermosipho melanesiensis、海栖热袍菌、thermotoga naphthophila、那不勒斯栖热袍菌、thermotoga petrophila、热袍菌属或thermobaculum terrenum。
[0124]
cas蛋白可以是cas9。cas9可以包含α螺旋叶和核酸酶叶。α螺旋叶可以包括三个区域:长α螺旋(称为桥螺旋)、rec1结构域和rec2结构域。核酸酶叶可以包含ruvc结构域、hnh结构域和pam相互作用结构域。图3突出显示了与连结非常靠近的不同结构域的氨基酸,所述连结可以是诸如rec1结构域(ser460、leu455、arg467、thr472、ile 473)、桥螺旋(arg69、asn77、arg74、arg70)和pam相互作用结构域(gly1103、phe1105、lys1123、lys1124、phe1105)的交联位点。图4突出显示了与茎环1非常靠近的不同结构域的氨基酸,所述茎环1可以是诸如ruvc结构域(lys33、lys742、lys1097、his721、glu57)、pam相互作用结构域(ser1351、tyr1356、his1349、val1100、thr1102)和桥螺旋结构域(thr62)的交联位点。图5突出显示了与茎环2非常靠近的不同结构域的氨基酸,所述茎环2可以是诸如ruvc结构域(lys30、asn46、arg40、lys44)和pam相互作用结构域(glu1225、ala1227、gln1272)的交联位点。
[0125]
在核酸与crispr多核苷酸(例如,rna)和靶dna分子结合时,核酸酶叶可以相对于α螺旋叶旋转~100
°
。一个或更多个交联基团可以被定位为以便保留crispr效应蛋白的全部活性,并且交联方法可以允许保留crispr效应蛋白(例如,cas核酸酶)的全部活性。
[0126]
c.用于crispr复合物的多核苷酸
[0127]
crispr多核苷酸可以包含rna、dna
‑
rna杂交体或其衍生物。crispr多核苷酸可以包含核苷,所述核苷可以包含共价附接至糖部分(例如,核糖或脱氧核糖)的碱基。核苷可以是核糖核苷或脱氧核糖核苷。核苷可以包含与氨基酸或氨基酸类似物连接的碱基,所述碱基可以包含游离羧基基团、游离氨基基团或保护基团。保护基团可以是例如在p.g.m.wuts和t.w.greene,“protective groups in organic synthesis”第二版,wiley
‑
interscience,new york,1999中描述的保护基团。crispr多核苷酸可以包含典型环核苷酸,例如,camp、cgmp、ccmp、cump、cimp、cxmp或ctmp。典型核苷酸碱基可以是腺嘌呤、胞嘧啶、尿嘧啶、鸟嘌呤或胸腺嘧啶。核苷酸可以包含附接至磷酸基团或磷酸类似物的核苷。
[0128]
crispr多核苷酸可以作为一种或更多种rna或dna分子存在(例如,位于编码所述一种或更多种rna或蛋白分子的一种或更多种载体中)。crispr多核苷酸可以是单链、双链或多链形式的脱氧核糖核酸(dna)、核糖核酸(rna)及其聚合物。crispr多核苷酸可以包含单链、双链或多链的dna或rna、基因组dna、cdna、dna
‑
rna杂交体,或包含嘌呤和/或嘧啶碱基或其他天然、化学修饰、生物化学修饰、非天然、合成或衍生化的核苷酸碱基的聚合物。
[0129]
在crispr
‑
cas系统中使用的多核苷酸(crispr多核苷酸)序列可以包含crrna序列和tracrrna序列。在自然界中,crrna和tracrrna可以以两个单独的rna分子存在。术语“tracrrna”或“tracrrna区段”可以指包含蛋白结合区段(例如,蛋白结合区段能够与crispr效应蛋白诸如cas9相互作用)的多核苷酸分子或其部分。术语“引导rna”和“grna”可以包括单引导rna(sgrna),其中crrna区段和tracrrna区段位于同一rna分子中。
[0130]
在一些情况下,grna可以是crispr rna(crrna)区段和反式激活crrna(tracrrna)区段的复合物(例如,通过氢键)。crrna可以包含杂交多核苷酸序列和tracrrna结合多核苷酸序列。杂交多核苷酸序列可以与靶核酸(例如,选择的外显子)的一部分杂交。crrna的杂交多核苷酸序列的范围可以是17个至23个核苷酸。crrna的杂交多核苷酸序列可以是至少17个、18个、19个、20个、21个、22个、23个或更多个核苷酸。crrna的杂交多核苷酸序列可以是至多23个、22个、21个、20个、19个、18个、17个或更少的核苷酸。在一种实例中,crrna的杂交多核苷酸序列是20个核苷酸。杂交多核苷酸可以是引导序列。引导序列可以包含与靶核酸序列的足够互补性以与靶核酸序列杂交。当使用合适的比对算法进行最佳比对时,互补性程度可以是约以下或多于约以下:50%、60%、75%、80%、85%、90%、95%、97.5%或99%。互补性程度可以是100%。在一些情况下,引导序列的长度可以是,例如,约5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个、20个、25个、30个、40个、50个或更多个核苷酸。引导序列的长度可以是约5个至约40个核苷酸。可以以减少引导序列与其自身碱基配对或与crispr多核苷酸的另一部分碱基配对的可能性的方式设计引导序列。当crispr多核苷酸最佳折叠时,引导序列的约以下或小于约以下的核苷酸可以与引导序列的另一部分或crispr多核苷酸的另一部分形成碱基对:75%、50%、40%、30%、25%、20%、15%、10%、5%、1%或更少。
[0131]
在一些情况下,单个crispr多核苷酸与单个crispr效应蛋白交联。单个crispr多核苷酸可以包含引导序列和与crispr效应蛋白交联的序列。能够交联crispr效应蛋白的序列可以是反式激活rna(tracrrna)。当单个crispr多核苷酸包含引导序列和tracrrna时,该单个crispr多核苷酸可以称为单引导rna(或sgrna)。
[0132]
在一些情况下,两个crispr多核苷酸可以与单个crispr效应蛋白交联。第一
crispr多核苷酸可以包含引导序列,并且第二crispr多核苷酸可以包含tracrrna并且缺少引导序列。
[0133]
在一些情况下,第一crispr多核苷酸包含引导序列和形成crrna的序列的第一部分(可以称为tracr配偶(tracr mate)序列),并且第二crispr多核苷酸包含形成tracrrna的序列的第二部分(可以称为tracr序列)。在一些情况下,tracr序列(或tracrrna)与crrna内的“tracr配偶”序列杂交,从而形成由crispr效应蛋白识别的双链rna双链体蛋白结合区段。包含引导序列(也称为间隔序列)但缺少能够与crispr效应蛋白结合的序列的crispr多核苷酸可以称为引导rna(或grna)。包含引导序列和仅一部分序列能够与crispr效应蛋白结合的序列(例如,tracr配偶序列)(并且缺少tracr序列)的crispr多核苷酸也可以称为引导rna(或grna)或crrna。
[0134]
tracrrna可以与crrna内的“tracr配偶”序列杂交,从而形成由crispr效应蛋白识别的双链rna双链体蛋白结合片段。在一些实例中,两者之间的杂交产生二级结构,诸如发夹。在一些情况下,crispr多核苷酸序列可以包含三个、四个、五个或更多个发夹。tracrrna可以包含一个或更多个发夹或者由一个或更多个发夹组成,并且长度可以是至少10个、20个、30个、40个、50个、60个、70个、80个、90个或100个核苷酸。
[0135]
在一些情况下,第一crispr多核苷酸可以是crrna,并且第二crispr多核苷酸可以是tracrrna,并且第一crispr多核苷酸和第二crispr多核苷酸可以是两个独立的rna分子。在一些情况下,单个crispr多核苷酸可以包含(1)能够与靶序列(例如,真核细胞中的基因组靶基因座)杂交的引导序列(或包含引导序列的crrna)和(2)tracrrna。在一些情况下,第一crispr多核苷酸可以包含(1)引导序列(或包含引导序列的crrna)(例如,能够与真核细胞中的靶序列杂交);和(2)tracr配偶序列(也称为直接重复序列),但缺少tracrrna序列。crispr效应蛋白可以与能够与靶序列杂交的引导序列和tracr配偶序列(直接重复序列)缔合,而不需要tracrrna。
[0136]
当tracr序列和tracr配偶序列位于单个crispr多核苷酸中时,tracr序列和tracr配偶序列可以共价连接。tracr序列和tracr配偶序列可以通过磷酸二酯键连接。tracr和tracr配偶可以通过非核苷酸环共价连接,所述非核苷酸环包括诸如间隔物、附接物、生物缀合物、发色团、报告物基团、染料标记的rna或非天然存在的核苷酸类似物的部分。间隔物可以是聚醚(例如,聚乙二醇、多元醇、聚丙二醇或乙二醇和丙二醇的混合物)、多胺基团(例如,精胺、亚精胺或其聚合衍生物)、聚酯(例如,聚(丙烯酸乙酯))、聚磷酸二酯、亚烷基及其组合。附接物可以是荧光标记物。生物缀合物可以是例如肽、糖苷、脂质、胆固醇、磷脂、二酰基甘油、二烷基甘油、脂肪酸、烃、酶底物、类固醇、生物素、地高辛、碳水化合物或多糖。发色团、报告物基团或染料标记的rna可以是荧光染料,例如,荧光素或罗丹明、化学发光标志化合物、电化学发光标志化合物或生物发光标志化合物。
[0137]
总体上,crrna的范围可以是35个至45个核苷酸。crrna可以是至少35个、36个、37个、38个、39个、40个、41个、42个、43个、44个、45个或更多个核苷酸。crrna可以是至多45个、44个、43个、42个、41个、40个、39个或更少的核苷酸。tracrrna的范围可以是60个至80个核苷酸。tracrrna可以是至少60个、61个、62个、63个、64个、66个、68个、70个、72个、74个、76个、78个、80个或更多个核苷酸。tracrrna可以是至多80个、79个、78个、77个、76个、74个、72个、70个、68个、66个、64个、62个、60个或更少的核苷酸。在一种实例中,tracrrna可以是72
个核苷酸。在另一实例中,crrna的杂交多核苷酸序列是20个核苷酸,crrna是42个核苷酸,并且相应的tracrrna是72个核苷酸。在另一实例中,crrna的杂交多核苷酸是20个核苷酸,crrna总计是34个核苷酸,并且相应的tracrrna是66个核苷酸。
[0138]
在一些情况下,crrna和tracrrna被连接为单引导rna分子,称为sgrna或“单引导rna”。每个sgrna可以包含长度范围为约20个至约30个、约30个至约40个、约40个至约50个、约50个至约60个、约60个至约70个、约70个至约80个或约80个至约100个核苷酸的恒定区。每个sgrna可以包含至少90个、91个、92个、93个、94个、95个、96个、97个、98个、99个、100个、101个、102个、103个、104个、105个、106个、107个、108个、109个或110个核苷酸。
[0139]
可选地,grna可以是三条或更多条rna链的复合物。三条或更多条rna链的复合物的至少一条rna链可以包含杂交多核苷酸序列。三条或更多条rna链的复合物的至少一条rna链可以包含crispr效应蛋白(例如,cas酶)结合序列。
[0140]
当grna与靶核酸分子杂交时,基因的杂交部分可以是靶区域(或靶基因座),所述靶区域(或靶基因座)包含前间区(靶位点)、被crispr效应蛋白(例如,cas酶)识别的前间区邻近基序(pam)和前间区的相对链(结合位点)。前间区的相对链可以是grna杂交基因组区域(序列)。靶核酸序列中的grna杂交序列范围可以是17个至23个核苷酸。基因中的grna杂交序列可以是至少17个、18个、19个、20个、21个、22个、23个或更多个核苷酸。基因中的grna杂交序列可以是至多23个、22个、21个、20个、19个、18个、17个或更少的核苷酸。
[0141]
crispr效应蛋白(例如,cas蛋白)可以是cas9,其中tracrrna可以通过四个发夹环与cas9的α
‑
螺旋叶和核酸酶叶相互作用;两个发夹环可以分别与每个叶相互作用。crrna可以被设计为与靶核酸(例如,dna)序列互补结合。对于cas9,全长sgrna靶dna结合区可以是20个核苷酸。对于cas9,pam序列可以包括3
’‑
ngg、3
’‑
nggng、3’nnagaaw和3
’‑
acay,其中n是任何核苷酸,w是a或t,并且y是c或t。
[0142]
在一些情况下,为了增加crispr多核苷酸(例如,grna或sgrna)的有效性,可以向crispr多核苷酸(例如,grna或sgrna)添加其他二级结构,以增强crispr多核苷酸的稳定性。在一些情况下,增加的稳定性可以改进核酸编辑。
[0143]
iii.稳定的crispr复合物
[0144]
本公开内容包括与sgrna共价结合的crispr效应蛋白,可称为“锁定的crispr复合物”。本公开内容包括与单独的crrna和/或tracrrna共价结合的crispr效应蛋白。本文还提供了crispr复合物,该crispr复合物中的sgrna和crispr效应蛋白具有增强的结合亲和力。crispr多核苷酸可以包括grna、sgrna、crrna或tracrrna。crispr效应蛋白可以与本文描述的任何crispr多核苷酸(例如,grna、sgrna、crrna、tracrrna、crispr on多核苷酸、crispr off多核苷酸、crispr on/off多核苷酸或被修饰以减少脱靶编辑的crispr多核苷酸)共价结合(例如交联)。
[0145]
crispr
‑
cas系统可以被修饰,以敲除特定基因以及敲入特定基因。crispr介导的敲除可以通过细胞的非同源末端连接修复途径产生。在这种事件下,crispr
‑
cas可以结合与结合的rna互补的靶核酸(例如,dna)区域,并且在靶核酸(例如,dna)区域执行双链切割。当设计sgrna时,可以在sgrna中靠近靶核酸(例如,dna)处为那些需要pam识别位点的cas核酸酶设计独特的3
‑
9个核苷酸pam识别区。并非每种cas核酸酶都需要pam区;例如,cas 14a不需要用于识别的pam区。
[0146]
与未共价结合的crispr复合物相比,通过至少一个共价键增强与crispr效应蛋白复合的sgrna的稳定性,可以减少脱靶裂解事件的数量,降低细胞毒性。相比于与标准sgrna复合的cas9,与sgrna交联(“锁定”)的crispr效应蛋白可以用于降低脱靶编辑。脱靶编辑可以使用以下技术来确定:ice(inference of crispr editing,crispr编辑推理),ice通过分析sanger测序跟踪和提供序列断裂水平的信息以确定插入/缺失形成频率来测量基因编辑的量(如hsiau等人“inference of crispr edits from sanger trace data”,january 14,2019biorxiv中描述的);或深度测序技术(如tsai等人“guide
‑
seq enables genome
‑
wide profiling of off
‑
target cleavage by crispr
‑
cas nucleases”,nature biotechnology 33,187
‑
197(2015)中描述的)。
[0147]
将sgrna与crispr效应蛋白共价锁定可以减少sgrna在细胞内将引起毒性的可能性。将sgrna与crispr效应蛋白锁定可以通过允许人们施用具有为特定靶设计的引导rna的crispr复合物的单一物质来增加给药的准确性。人们可以施用两种或更多种具有彼此独特的靶的锁定的crispr复合物,用于靶向多于一种位点的复合疗法。此外,将sgrna序列与crispr效应蛋白复合配制,使得sgrna序列不能够从复合状态中解离,可以在制剂中和施用之后提供更大的保护,防止降解。
[0148]
a.修饰的crispr多核苷酸
[0149]
本文公开了用至少一个用于交联的非天然核苷酸和用于调节活性的序列修饰的crispr多核苷酸。用于修饰活性的序列可以是crispr on多核苷酸序列和crispr off多核苷酸序列、crispr on/off多核苷酸序列或被修饰以降低脱靶编辑的crispr核苷酸。非天然核苷酸可以位于tracrrna、crrna或crrna的引导序列中。
[0150]
在一些情况下,可以修饰crispr多核苷酸,以改进crispr多核苷酸的核酸酶抗性、血清稳定性、靶特异性、血液系统循环、组织分布、组织渗透、细胞摄取、效力和/或细胞通透性。例如,某些crispr多核苷酸修饰可以增加核酸酶稳定性和/或降低干扰素诱导,而不会显著影响crispr多核苷酸(例如,sgrna)的活性。与具有相同序列的未修饰的crispr多核苷酸相比,修饰的crispr多核苷酸可以具有改进的在血清和/或脑脊液中的稳定性。本文公开的crispr多核苷酸(例如,sgrna)可以在各种位置处(包括糖部分、磷酸二酯连接和/或碱基处)包含一种或更多种修饰。如本文描述的修饰的crispr多核苷酸可以包括sgrna以及单独的crrna和tracrrna。crispr多核苷酸可以通过氢键相互作用与crispr效应蛋白结合。
[0151]
本文提供了crispr多核苷酸,所述crispr多核苷酸可以通过一个或更多个共价键与crispr效应蛋白交联,形成锁定的crispr复合物。crispr多核苷酸和crispr效应蛋白之间可以存在1个至3个、3个至6个、6个至9个、9个至12个、12个至15个、15个至18个、18个至21个、21个至24个、24个至27个、27个至30个、30个至33个、33个至36个、36个至39个、39个至42个、42个至45个、45个至48个、48个至51个、51个至54个、54个至57个、57个至60个、60个至62个、62个至65个、65个至68个、68个至71个、71个至74个、74个至77个、77个至80个、80个至83个、83个至86个、86个至89个、89个至91个、91个至94个、94个至97个、97个至100个共价键。crispr多核苷酸和crispr效应蛋白之间可以存在至少1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个、20个、21个、22个、23个、24个、25个、26个、27个、28个、29个、30个、31个、32个、33个、34个、35个、36个、37个、38个、39个、40个、41个、42个、43个、44个、45个、46个、47个、48个、49个、50个、51个、52个、53个、54
个、55个、56个、57个、58个、59个、60个、61个、62个、63个、64个、65个、66个、67个、68个、69个、70个、71个、72个、73个、74个、75个、76个、77个、78个、79个、80个、81个、82个、83个、84个、85个、86个、87个、88个、89个、90个、91个、92个、93个、94个、95个、96个、97个、98个、99个或100个共价键。crispr多核苷酸和crispr效应蛋白之间可以存在至多100个、99个、98个、97个、96个、95个、94个、93个、92个、91个、90个、89个、88个、87个、86个、85个、84个、83个、82个、81个、80个、79个、78个、77个、76个、75个、74个、73个、72个、71个、70个、69个、68个、67个、66个、65个、64个、63个、62个、61个、60个、59个、58个、57个、56个、55个、54个、53个、52个、51个、50个、49个、48个、47个、46个、45个、44个、43个、42个、41个、40个、39个、38个、37个、36个、35个、34个、33个、31个、30个、29个、28个、27个、26个、25个、24个、23个、22个、21个、20个、19个、18个、17个、16个、15个、14个、13个、12个、11个、10个、9个、8个、7个、6个、5个、4个、3个、2个或1个共价键。crispr多核苷酸和crispr效应蛋白之间可以存在约1个至约10个、约10个至约30个、约30个至约60个或约60个至约80个共价键。
[0152]
crispr多核苷酸可以包含含有磷酰胺、硫代磷酸酯、二硫代磷酸酯、硼烷磷酸酯连接、o
‑
甲基亚磷酰胺连接和/或肽核酸的主链,以便如本文描述的控制crispr复合物的活性。可以修饰单独的核苷酸,以便向crispr多核苷酸添加能够与crispr效应蛋白中的邻近氨基酸形成共价键的交联剂。一个或更多个交联剂的位置可以在crispr多核苷酸的tracrrna区内,例如,如图1中示出的。一个或更多个交联剂可以在sgrna的发夹环内。交联反应可以非常靠近crispr效应蛋白中的受体分子,以便形成共价键。实施方案包括将官能化核苷酸设计为在引发交联之前在crispr效应蛋白的受体分子(例如,crispr效应蛋白中的邻近氨基酸)的20埃、19埃、18埃、17埃、16埃、15埃、14埃、13埃、12埃、11埃、10埃、9埃、8埃、7埃、6埃、5埃、4埃、3埃、2埃或1埃以内。
[0153]
在一些情况下,crispr多核苷酸(例如,grna、sgrna、crrna或tracrrna)可以包含至少1个交联剂、至少2个交联剂、至少5个交联剂、至少12个交联剂、至少15个交联剂、至少20个交联剂、至少25个交联剂、至少30个交联剂、至少35个交联剂、至少40个交联剂、至少50个交联剂、至少55个交联剂、至少60个交联剂、至少65个交联剂、至少70个交联剂、至少75个交联剂或至少80个交联剂。crispr多核苷酸可以包含至多100个、50个、25个、10个、9个、8个、7个、6个、5个、4个、3个、2个或1个交联剂。crispr多核苷酸可以包含约1个至约10个、约10个至约30个、约30个至约60个或约60个至约80个交联剂。
[0154]
可选地或组合地,一个或更多个交联剂的位置可以在tracrrna序列的任何核苷酸处,或者在tracrrna序列的任何两个核苷酸之间,在靶结合crrna区之外。一个或更多个交联剂可以存在于crispr多核苷酸的任何茎区:连结、茎环1、茎环2或四环中(例如,参见,图2)。一个或更多个交联剂可以位于多核苷酸的发夹环或连结茎、茎环1、茎环2或四环或其任何组合中。一个或更多个交联剂可以存在于1个发夹、2个发夹、3个发夹或4个发夹中。发夹的环可以包含1个交联剂、2个交联剂、3个交联剂、4个交联剂、5个交联剂、6个交联剂等。1个、2个、3个、4个、5个或更多个交联剂可以位于四环的凸起中。
[0155]
可选地或与以上组合地,一个或更多个交联剂可以位于sgrna的核苷酸位置49处,其中核苷酸位置1位于crrna的靶结合区的5’末端处,并且sgrna的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。
[0156]
一个或更多个交联剂可以位于sgrna的以下核苷酸位置处:1、2、3、4、5、6、7、8、9、
10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109或110,其中核苷酸位置1位于crrna的靶结合区的5’末端处,并且sgrna的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。
[0157]
一个或更多个交联剂可以位于sgrna的任何尿嘧啶残基处。可以修饰sgrna,使得sgrna的发夹结构相对于缺少交联剂的sgrna的茎环结构被维持。sgrna可以通过发夹结构中互补核苷酸对之间的核苷酸交换来修饰。示例性交换可以是sgrna的位置49处的尿嘧啶
‑
腺嘌呤交换,留下具有腺嘌呤的位置22,如从图7b(从图7a中可见的构型修饰的)中可见的。可选地,脱氧尿苷可以替代位置49处的尿苷,如图7c中可见的。可选地,脱氧腺苷可以替代位置22处的腺苷,如图7d中可见的。作为最后的选择,位置50处的尿苷可以被脱氧尿苷取代。
[0158]
可选地或与以上组合地,一个或更多个交联剂可以位于1个发夹茎、2个发夹茎、3个发夹茎或4个发夹茎中。发夹茎可以包含1个交联剂、2个交联剂、3个交联剂、4个交联剂、5个交联剂、6个交联剂、7个交联剂、8个交联剂等。一个或更多个交联剂可以位于茎之间的非碱基配对(un
‑
base
‑
paired)的核苷酸中。可选地或组合地,一个或更多个交联剂可以位于茎区域之间的一个或更多个核苷酸中。
[0159]
一个或更多个交联剂可以位于crispr多核苷酸(例如,grna、sgrna、crrna或tracrrna)的主链上,或者可以以交联剂修饰的核苷酸被包括。核苷酸修饰可以包括(a)末端修饰,包括5’末端修饰或3’末端修饰;(b)核酸碱基(或“碱基”)修饰,包括碱基的替代或去除;(c)糖修饰,包括2’、3’和/或4’位置处的修饰;和(d)主链修饰,包括磷酸二酯连接的修饰或替代。crispr多核苷酸可以包含2'氟
‑
阿拉伯糖核酸、三环
‑
dna(tc
‑
dna)、肽核酸、环己烯核酸(cena)、亚乙基桥接核酸(ena)、磷酰二亚胺基吗啉代(3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺或其组合。crispr多核苷酸(例如,sgrna)可以包含一个或更多个非天然存在的核苷酸或核苷酸类似物,例如,具有硫代磷酸酯连接、硼烷磷酸酯连接或桥接核酸(bna)的核苷酸。非天然存在的核苷酸或核苷酸类似物可以是2'
‑
o
‑
甲基类似物、2'
‑
脱氧类似物、2
‑
硫代尿苷类似物、n6
‑
甲基腺苷类似物或2'
‑
氟类似物。
[0160]
在一些情况下,多核苷酸可以在5’末端处的前1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个或12个核苷酸处包含修饰的核苷酸和/或修饰的核苷酸间连接。在一些情况下,多核苷酸可以在3’末端处的1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个或12个核苷酸处包含修饰的核苷酸和/或修饰的核苷酸间连接。在一些情况下,多核苷酸可以在5’末端处的前1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个或12个核苷酸处或者在3’末端处的1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个或12个核苷酸处包含修饰的核苷酸和/或修饰的核苷酸间连接。在一些情况下,多核苷酸可以在5’末端处的前1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个或12个核苷酸处以及在3’末端处的前1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个或12个核苷酸处包含修饰的核苷酸和/或修饰的核苷酸间连接。修饰可以是2
’‑
o
‑
甲基类似物和/或3’硫代磷酸酯核苷酸间连接。
[0161]
crispr多核苷酸可以包含一个或更多个修饰的碱基。一个或更多个修饰的碱基可以是2
‑
氨基嘌呤、5
‑
溴尿苷、假尿苷(ψ)、n^甲基假尿苷(n^methylpseudouridine,mel p)、5
‑
甲氧基尿苷(5mou)、肌苷或7
‑
甲基鸟苷。
[0162]
在一些情况下,例如通过修饰3'连接或5'连接,crispr多核苷酸的3'末端和5'末端可以基本上被保护不受核酸酶作用(例如,美国专利第5,849,902号和wo 98/13526)。例如,crispr多核苷酸可以通过包含一个或更多个“封闭基团”而具有抗性。一个或更多个“封闭基团”可以是可以附接至多核苷酸或核酸单体(nucleomonomer)的取代基(例如,除羟基以外的基团),作为用于合成的保护基团或偶联基团(例如,fitc、丙基(
‑
ch2
‑
ch2
‑
ch3)、乙二醇(
‑
o
‑
ch2
‑
ch2
‑
o
‑
)磷酸酯(po3 2
‑
)、氢膦酸酯或亚磷酰胺)。一个或更多个封闭基团可以是可以保护crispr多核苷酸的5
′
末端和3
′
末端的一个或更多个“末端封闭基团”或者一个或更多个“外切核酸酶封闭基团”,包括修饰的核苷酸和非核苷酸外切核酸酶抗性结构。
[0163]
一个或更多个末端封闭基团可以是帽结构(例如,7
‑
甲基鸟苷帽)、反向的核酸单体(例如,具有3
′‑3′
或5
′‑5′
末端反向(参见,例如,ortiagao等人1992.antisense res.dev.2:129))、甲基膦酸酯、亚磷酰胺、非核苷酸基团(例如,非核苷酸接头、氨基接头、缀合物)等。3
′
末端核酸单体可以包含修饰的糖部分。例如,3
′‑
羟基可以通过3
′→3′
核苷酸间连接酯化为核苷酸。例如,烷氧基基团可以是甲氧基、乙氧基或异丙氧基。任选地,在3’末端处的3
′→3′
连接的核苷酸可以通过取代基连接来连接。为了减少核酸酶降解,最5
′
侧的3
′→5′
连接可以是修饰的连接,例如,硫代磷酸酯或对烷氧基磷酸三酯连接。
[0164]
crispr多核苷酸可以包含一个或更多个标记物或标签。一个或更多个“标记物”或“标签”可以是可以附接至另一个分子(例如,crispr多核苷酸或其区段)以提供可以容易地藉以检测该另一个分子的手段的分子。crispr多核苷酸可以包含标记物,该标记物可以是荧光的、发光的、放射性的、酶促活性的等。一个或更多个标记物可以包括荧光染料,例如异硫氰酸荧光素(fitc)、罗丹明、德克萨斯红(texas red)、藻红蛋白、别藻蓝蛋白、6
‑
羧基荧光素(6
‑
fam)、2,7
‑
二甲氧基
‑
4,5
‑
二氯
‑6‑
羧基荧光素(joe)、6
‑
羧基
‑
x
‑
罗丹明(rox)、6
‑
羧基
‑
2,4,7,4,7
‑
六氯荧光素(hex)、5
‑
羧基荧光素(5
‑
fam)或n,n,n,n
‑
四甲基
‑6‑
羧基罗丹明(tamra),放射性标记物,例如,32p、35s、3h等。一个或更多个标记物可以是两级系统,其中crispr多核苷酸与生物素、半抗原等缀合,生物素、半抗原等具有高亲和力结合配偶体,例如,抗生物素蛋白、特异性抗体等,其中结合配偶体与可检测的标记物缀合。
[0165]
crispr多核苷酸(例如,sgrna)可以包含一个或更多个茎环,一种或更多种茎环rna结合蛋白(rbp)能够与所述一个或更多个茎环相互作用。这些茎环可以定位成使得crispr多核苷酸(例如,sgrna)与crispr效应蛋白(例如,crispr酶)的相互作用或crispr复合物与靶dna的结合不受不利影响。一个或更多个茎环可以位于crispr多核苷酸(例如,sgrna)的引导序列之外。一种或更多种茎环rna结合蛋白可以是,例如,ms2、pp7、qp、f2、ga、fr、jp501、m12、r17、bz13、jp34、jp500、ku1、m11、mx1、tw18、vk、sp、fl、id2、nl95、tw19、ap205、s1、s1m、7s或prr1。
[0166]
在一些情况下,茎环rna结合蛋白(rbp)可以充当衔接子蛋白(即中间物),既可以与茎环rna结合,也可以与一种或更多种其他蛋白或多肽或一个或更多个功能结构域结合。衔接子蛋白可以募集可包含一个或更多个功能结构域的效应蛋白或融合蛋白。在一些情况下,rna结合蛋白可以是具有一个或更多个功能结构域的融合蛋白。
[0167]
1.修饰类型
[0168]
在一些情况下,crispr多核苷酸(例如,grna、sgrna、crrna或tracrrna)可以被修饰以促进与crispr效应蛋白的锁定。用于将crispr多核苷酸(例如,sgrna)分子与crispr效应蛋白锁定的修饰可以包括用官能性交联基团修饰sgrna上的核苷酸。
[0169]
修饰的核苷酸可以被引入crispr多核苷酸(例如,sgrna)中。合适的方法是,例如,使用(自动或半自动)寡核苷酸合成装置,例如,以3’至5’方向的合成方法。这样的装置可以包括微阵列、聚合酶循环装置(polymerase cycling assembly,pca)、微芯片等。
[0170]
crispr多核苷酸可以包含糖部分。糖部分可以是天然的、未修饰的糖,例如,单糖(例如,戊糖,例如,核糖、脱氧核糖)、修饰的糖或糖类似物。在一些情况下,糖部分可以具有被卤素、杂原子、脂族基团替代的一个或更多个羟基基团,或者一个或更多个羟基基团可以被官能化为醚、胺、硫醇等。
[0171]
crispr多核苷酸可以在核糖的2’位置处包含一种或更多种修饰。可以在核糖的2’位置处引入一种或更多种修饰,例如,以减少在细胞环境中的免疫刺激。2'部分可以是h、or、r、卤素、sh、sr、h2、hr、r2或on,其中r是c1
‑
c6烷基、烯基或炔基,并且卤素是f、ci、br或i。糖修饰的实例包括2'
‑
脱氧
‑
2'
‑
氟
‑
寡核糖核苷酸(2'
‑
氟
‑
2'
‑
脱氧胞苷
‑
5'
‑
三磷酸、2'
‑
氟
‑
2'
‑
脱氧尿苷
‑
5'
‑
三磷酸)、2'
‑
脱氧
‑
2'
‑
脱氨基寡核糖核苷酸(2'
‑
氨基
‑
2'
‑
脱氧胞苷
‑
5'
‑
三磷酸、2'
‑
氨基
‑
2'
‑
脱氧尿苷
‑
5'
‑
三磷酸)、2'
‑
o
‑
烷基寡核糖核苷酸、2'
‑
脱氧
‑
2'
‑
c
‑
烷基寡核糖核苷酸(2'
‑
o
‑
甲基胞苷
‑
5'
‑
三磷酸、2'
‑
甲基尿苷
‑
5'
‑
三磷酸)、2'
‑
c
‑
烷基寡核糖核苷酸及其异构体(2'
‑
阿糖胞苷
‑
5'
‑
三磷酸、2'
‑
阿糖尿苷
‑
5'
‑
三磷酸)、叠氮三磷酸(2'
‑
叠氮基
‑
2'
‑
脱氧胞苷
‑
5'
‑
三磷酸、2'
‑
叠氮基
‑
2'
‑
脱氧尿苷
‑
5'
‑
三磷酸)及其组合。糖修饰的核糖核苷酸可以具有被以下替代的2’oh基团:h、烷氧基(或or)、r或烷基、卤素、sh、sr、氨基(诸如nh2、nhr、nr2)或cn基团,其中r是低级烷基、烯基或炔基。在2'位置处的修饰可以是甲基基团。
[0172]
多核苷酸可以包含一个或更多个核酸碱基修饰的核糖核苷酸。一种或更多种修饰的核糖核苷酸可以含有非天然存在的碱基(而不是天然存在的碱基),诸如在5'
‑
位置处修饰的尿苷或胞苷,例如5’(2
‑
氨基)丙基尿苷或5'
‑
溴尿苷;在8
‑
位置处修饰的腺苷和鸟苷,例如8
‑
溴鸟苷;脱氮核苷酸,例如7
‑
脱氮
‑
腺苷;和n
‑
烷基化核苷酸,例如n6
‑
甲基腺苷。
[0173]
核酸碱基修饰的核糖核苷酸可以是m5c(5
‑
甲基胞苷)、m5u(5
‑
甲基尿苷)、m6a(n6
‑
甲基腺苷)、s2u(2
‑
硫代尿苷)、um(2'
‑
o
‑
甲基尿苷)、m1a(1
‑
甲基腺苷)、m2a(2
‑
甲基腺苷)、am(2'
‑
o
‑
甲基腺苷)、ms2m6a(2
‑
甲硫基
‑
n6
‑
甲基腺苷)、i6a(n6
‑
异戊烯腺苷)、ms2i6a(2
‑
甲硫基
‑
n6
‑
异戊烯腺苷)、io6a(n6
‑
(顺式
‑
羟基异戊烯基)腺苷)、ms2io6a(2
‑
甲硫基
‑
n6
‑
(顺式
‑
羟基异戊烯基)腺苷)、g6a(n6
‑
甘氨酰氨基甲酰腺苷)、t6a(n6
‑
苏氨酰氨基甲酰腺苷)、ms2t6a(2
‑
甲硫基
‑
n6
‑
苏氨酰氨基甲酰腺苷)、m6t6a(n6
‑
甲基
‑
n6
‑
苏氨酰氨基甲酰腺苷)、hn6a(n6
‑
羟基正缬氨酰氨基甲酰腺苷)、ms2hn6a(2
‑
甲硫基
‑
n6
‑
羟基正缬氨酰氨基甲酰腺苷)、ar(p)(2'
‑
o
‑
核糖腺苷(磷酸))、i(肌苷)、m1i(1
‑
甲基肌苷)、m1im(1,2'
‑
o
‑
二甲基肌苷)、m3c(3
‑
甲基胞苷)、cm(2'
‑
o
‑
甲基胞苷)、s2c(2
‑
硫代胞苷)、ac4c(n4
‑
乙酰胞苷)、f5c(5
‑
甲酰胞苷)、m5cm(5,2
‑
o
‑
二甲基胞苷)、ac4cm(n4
‑
乙酰基
‑
2'
‑
o
‑
甲基胞苷)、k2c(赖胞苷)、m1g(1
‑
甲基鸟苷)、m2g(n2
‑
甲基鸟苷)、m7g(7
‑
甲基鸟苷)、gm(2'
‑
o
‑
甲基鸟苷)、m22g(n2,n2
‑
二甲基鸟苷)、m2gm(n2,2'
‑
o
‑
二甲基鸟苷)、m22gm(n2,n2,2'
‑
o
‑
三甲基鸟苷)、gr
(p)(2'
‑
o
‑
核糖鸟苷(磷酸))、yw(怀丁苷(wybutosine))、o2yw(过氧怀丁苷)、ohyw(羟基怀丁苷)、ohyw*(修饰不足的羟基怀丁苷)、img(怀俄苷(wyosine))、mimg(甲基怀俄苷)、q(辫苷(queuosine))、oq(环氧辫苷)、galq(半乳糖基
‑
辫苷)、manq(甘露糖基
‑
辫苷)、preq0(7
‑
氰基
‑7‑
脱氮鸟苷)、preq1(7
‑
氨基甲基
‑7‑
脱氮鸟苷)、g(古嘌苷(archaeosine))、d(二氢尿苷)、m5um(5,2'
‑
o
‑
二甲基尿苷)、s4u(4
‑
硫代尿苷)、m5s2u(5
‑
甲基
‑2‑
硫代尿苷)、s2um(2
‑
硫代
‑
2'
‑
o
‑
甲基尿苷)、acp3u(3
‑
(3
‑
氨基
‑3‑
羧基丙基)尿苷)、ho5u(5
‑
羟基尿苷)、mo5u(5
‑
甲氧基尿苷)、cmo5u(尿苷5
‑
氧基乙酸)、mcmo5u(尿苷5
‑
氧基乙酸甲酯)、chm5u(5
‑
(羧基羟基甲基)尿苷))、mchm5u(5
‑
(羧基羟基甲基)尿苷甲酯)、mcm5u(5
‑
甲氧基羰基甲基尿苷)、mcm5um(s
‑
甲氧基羰基甲基
‑2‑
o
‑
甲基尿苷)、mcm5s2u(5
‑
甲氧基羰基甲基
‑2‑
硫代尿苷)、nm5s2u(5
‑
氨基甲基
‑2‑
硫代尿苷)、mnm5u(5
‑
甲基氨基甲基尿苷)、mnm5s2u(5
‑
甲基氨基甲基
‑2‑
硫代尿苷)、mnm5se2u(5
‑
甲基氨基甲基
‑2‑
硒尿苷)、ncm5u(5
‑
氨基甲酰甲基尿苷)、ncm5um(5
‑
氨基甲酰甲基
‑
2'
‑
o
‑
甲基尿苷)、cmnm5u(5
‑
羧基甲基氨基甲基尿苷)、cnmm5um(5
‑
羧基甲基氨基甲基
‑
2'
‑
o
‑
甲基尿苷)、cmnm5s2u(5
‑
羧基甲基氨基甲基
‑2‑
硫代尿苷)、m62a(n6,n6
‑
二甲基腺苷)、im(2'
‑
o
‑
甲基肌苷)、m4c(n4
‑
甲基胞苷)、m4cm(n4,2
‑
o
‑
二甲基胞苷)、hm5c(5
‑
羟基甲基胞苷)、m3u(3
‑
甲基尿苷)、cm5u(5
‑
羧基甲基尿苷)、m6am(n6,2'
‑
o
‑
二甲基腺苷)、m6,6am(n6,n6,2'
‑
o
‑
三甲基腺苷)、m2,7g(n2,7
‑
二甲基鸟苷)、m2,2,7g(n2,n2,7
‑
三甲基鸟苷)、m3um(3,2'
‑
o
‑
二甲基尿苷)、m5d(5
‑
甲基二氢尿苷)、f5cm(5
‑
甲酰基
‑
2'
‑
o
‑
甲基胞苷)、m1gm(1,2'
‑
o
‑
二甲基鸟苷)、m1am(1,2'
‑
o
‑
二甲基腺苷)、nm5u(5
‑
氨基甲基尿苷)、tm5s2u(s
‑
牛磺酸甲基
‑2‑
硫代尿苷)、img
‑
14(4
‑
脱甲基怀俄苷)、img2(异怀俄苷)、或ac6a(n6
‑
乙酰腺苷)、次黄嘌呤、肌苷、8
‑
氧代
‑
腺嘌呤、其7
‑
取代的衍生物、二氢尿嘧啶、假尿嘧啶、2
‑
硫代尿嘧啶、4
‑
硫代尿嘧啶、5
‑
氨基尿嘧啶、5
‑
(c1
‑
c6)
‑
烷基尿嘧啶、5
‑
甲基尿嘧啶、5
‑
(c2
‑
c6)
‑
烯基尿嘧啶、5
‑
(c2
‑
c6)
‑
炔基尿嘧啶、5
‑
(羟甲基)尿嘧啶、5
‑
氯尿嘧啶、5
‑
氟尿嘧啶、5
‑
溴尿嘧啶、5
‑
羟基胞嘧啶、5
‑
(c1
‑
c6)
‑
烷基胞嘧啶、5
‑
甲基胞嘧啶、5
‑
(c2
‑
c6)
‑
烯基胞嘧啶、5
‑
(c2
‑
c6)
‑
炔基胞嘧啶、5
‑
氯胞嘧啶、5
‑
氟胞嘧啶、5
‑
溴胞嘧啶、n2
‑
二甲基鸟嘌呤、7
‑
脱氮鸟嘌呤、8
‑
氮杂鸟嘌呤、7
‑
脱氮
‑7‑
取代的鸟嘌呤、7
‑
脱氮
‑7‑
(c2
‑
c6)炔基鸟嘌呤、7
‑
脱氮
‑8‑
取代的鸟嘌呤、8
‑
羟基鸟嘌呤、6
‑
硫代鸟嘌呤、8
‑
氧代鸟嘌呤、2
‑
氨基嘌呤、2
‑
氨基
‑6‑
氯嘌呤、2,4
‑
二氨基嘌呤、2,6
‑
二氨基嘌呤、8
‑
氮杂嘌呤、取代的7
‑
脱氮嘌呤、7
‑
脱氮
‑7‑
取代的嘌呤、7
‑
脱氮
‑8‑
取代的嘌呤及其组合。
[0174]
核酸碱基修饰的核糖核苷酸可以是氨基嘌呤、2,6
‑
二氨基嘌呤(2
‑
氨基
‑
da)、5
‑
溴du、脱氧尿苷、反向dt(inverted dt)、反向双脱氧
‑
t、双脱氧
‑
c、5
‑
甲基dc、超级(t)(super(t))、超级(g)(super(g))、5
‑
硝基吲哚、2'
‑
o
‑
甲基rna碱基、羟基甲基dc、异dg、异dc、氟代c、氟代u、氟代a、氟代g、2
‑
甲氧基乙氧基mec、2
‑
甲氧基乙氧基g或2
‑
甲氧基乙氧基t。
[0175]
a.用于锁定的化学交联剂
[0176]
在一些情况下,crispr效应蛋白可以被修饰以促进与crispr多核苷酸(例如,sgrna)的锁定。crispr多核苷酸(例如,sgrna)可以包含一个或更多个交联剂。一个或更多个交联剂可以是形成共价键(例如,在聚合物诸如异氰酸酯之间)的官能团。一个或更多个交联剂可以是甲醛或戊二醛。交联可以包括生物缀合。生物缀合交联试剂可以包含与官能团(诸如胺和巯基)反应的反应性基团。生物缀合交联剂可以包括巯基反应性基团,诸如马来酰亚胺、卤代乙酰基、氮丙啶、丙烯酰基、烷氧基胺、芳基化剂、乙烯基砜、吡啶基二硫化
物、tnb硫醇、二硫醇亚磷酰胺dtpa等。生物缀合交联剂还可以包括胺反应性交联剂反应性基团,诸如琥珀酰亚胺酯(nhs酯)、磺酰氯、醛、碳二亚胺、酰基叠氮化物、芳基叠氮化物、酸酐、氟苯、碳酸酯、亚氨酸酯、环氧化物、氟苯基酯、亚磷酰胺等。
[0177]
交联剂的另外的非限制性实例可以来源于以下化合物:硫醇 硫醇、硫醇 马来酰亚胺、nhs酯 胺、羧酸 nhs 胺、叠氮化物 膦(施陶丁格连接(staudinger ligation))、羰基化合物 胺、羰基化合物 o
‑
取代的羟胺、双吖丙啶 c
‑
h/o
‑
h、n
‑
h、卤乙酸酯 硫醇、叠氮化物 炔烃、硝酮 炔烃、腈氧化物 炔烃、四嗪 烯烃、4
‑
硫代尿苷、5
’‑
叠氮基尿苷、5
‑
溴尿苷、8
‑
叠氮基腺苷、5
‑
((4
‑
叠氮苯甲酰基)硫代)尿苷。
[0178]
crispr多核苷酸(例如,sgrna)可以被修饰以包含一个或更多个非天然核苷酸。非天然核苷酸可以包括包含针对碱基、糖和/或磷酸部分的一个或更多个修饰的核苷酸。一种或更多种修饰可以包括一种或更多种化学修饰。一种或更多种修饰可以是例如3’oh或5’oh基团、主链、糖组分和/或核苷酸碱基(例如,嘌呤或嘧啶)的修饰。一种或更多种修饰可以包括添加用于交联的一个或更多个接头分子。一个或更多个接头分子可以被配置为与氨基酸形成共价键。一个或更多个接头分子可以被配置为与氨基酸形成非共价键。在一方面,修饰的碱基包括除腺嘌呤、鸟嘌呤、胞嘧啶或胸腺嘧啶以外的碱基(在修饰的dna中)或者除腺嘌呤、鸟嘌呤、胞嘧啶或尿嘧啶以外的碱基(在修饰的rna中)。在一些实施方案中,修饰是腺嘌呤、鸟嘌呤、胞嘧啶或胸腺嘧啶的修饰形式(在修饰的dna中)或者腺嘌呤、鸟嘌呤、胞嘧啶或尿嘧啶的修饰形式(在修饰的rna中)。非天然核苷酸可以是在糖、核苷酸间磷酸二酯键、嘌呤或嘧啶残基处共价修饰以包含共价接头的官能团的核苷酸。参见,例如,sletten等人,angew.chem.int.编著(2009)48:6974
‑
6998;manoharan,m.curr.opin.chem.biol.(2004)8:570
‑
9;behlke等人,polynucleotides(2008)18:305
‑
19;watts,等人,drug.discov.today(2008)13:842
‑
55;shukla等人,chemmedchem(2010)5:328
‑
49。非天然核苷酸可以包括在糖、核苷酸间磷酸二酯键、嘌呤或嘧啶残基处被修饰(例如共价修饰)以包含官能团的核苷酸。共价接头可以是选自由以下组成的组的化学部分:氨基甲酸酯、醚、酯、酰胺、亚胺、脒、氨基三嗪(aminotrizine)、腙(hydrozone)、二硫化物、硫醚、硫酯、硫代磷酸酯、二硫代磷酸酯、磺酰胺、磺酸酯、砜、亚砜、脲、硫脲、酰肼、肟、三唑、光不稳定连接、c
‑
c键形成基团如diels
‑
alder环加成对或闭环换位反应对和迈克尔反应对。化学键可以基于、氨基甲酸酯、醚、酯、酰胺、亚胺、脒、氨基三嗪、腙、二硫化物、硫醚、硫酯、硫代磷酸酯、二硫代磷酸酯、磺酰胺、磺酸酯、砜、亚砜、脲、硫脲、酰肼、肟、三唑、光不稳定连接、c
‑
c键形成基团诸如diels
‑
alder环加成对或闭环换位反应对和迈克尔反应对。
[0179]
例如,crispr多核苷酸(例如,sgrna)的非天然核苷酸可以包含马来酰亚胺,以与crispr效应蛋白的邻近半胱氨酸氨基酸交联,形成硫醚键。一种用于整合能够将crispr效应蛋白与crispr多核苷酸(例如,sgrna)交联的非天然核苷酸的技术可以包括用与存在于crispr效应蛋白中的半胱氨酸反应的化学基团(诸如马来酰亚胺)修饰crispr多核苷酸(例如,sgrna)的核苷酸。包含马来酰亚胺的非天然核苷酸可以通过与crispr效应蛋白的半胱氨酸的硫醇侧链反应,将crispr多核苷酸与crispr效应蛋白交联。图6示出了与crispr效应蛋白(包括cas9核酸酶)的邻近半胱氨酸交联的非天然核苷酸的示例性位置的晶体结构。特别地,该结构示出了非天然核苷酸(核苷酸位置22(左侧的白色圈))和crispr效应蛋白上的邻近氨基酸(crispr效应蛋白的位置80处的半胱氨酸(右侧的白色圈))。如以上描述的,
sgrna位置22和49可以用尿苷至腺嘌呤交换来修饰,如图7b中可见的,留下rna核苷酸位置49为尿嘧啶(u49)。u49处的非天然核苷酸可以通过修饰u49的糖分子以包括与spcas9上的cys80相互作用的交联部分来整合。具有被配置为与cas9核酸酶复合的野生型tracr rna序列的sgrna的其他可以用尿苷至腺嘌呤交换来修饰的位置是u72/a77、u71/a78和u94/a84。sgrna的位置77、位置78和位置84处的尿嘧啶核苷酸可以用本文描述的交联基团修饰,以便与cas9核酸酶的邻近氨基酸形成共价键。
[0180]
图8概述了用接头基团修饰核苷酸的示例性方法。亚磷酰胺核苷酸与附接至间隔物和马来酰亚胺基团的亲核体反应。引入的亲核体替代亚磷酰胺核苷酸的接头基团,留下附接至马来酰亚胺的间隔物。当靠近具有硫醇基团的半胱氨酸时,诸如当与crispr效应蛋白复合时,马来酰亚胺可以在生理条件下与半胱氨酸形成共价硫醚键。
[0181]
一种用于将crispr效应蛋白与crispr多核苷酸(例如,sgrna)交联的技术可以包括修饰crispr多核苷酸(例如,sgrna)的核苷酸,以产生具有与存在于crispr效应蛋白中的赖氨酸的侧链上的伯胺反应的化学基团(诸如nhs酯、环氧化物、醛、酰基叠氮化物等)的非天然核苷酸。
[0182]
图9示出了crispr复合物的晶体结构,其中突出显示了sgrna的天然尿嘧啶核苷酸。尿嘧啶核苷酸位于位置22、23、24、25、31、37、44、45、50、56、59、63、64、66、71、72、80、90和94处。这些残基可以被官能化,以与crispr效应蛋白形成交联。
[0183]
图10示出了包含cas9核酸酶的crispr复合物的晶体结构的非限制性实例。sgrna的位置44(尿嘧啶)由于靠近crispr效应蛋白的酪氨酸325而被突出显示为适用于修饰。
[0184]
图11示出了包含cas9核酸酶的crispr复合物的晶体结构的非限制性实例。sgrna的位置59(尿嘧啶)由于靠近crispr效应蛋白的酪氨酸81而被突出显示为适用于修饰。
[0185]
图12示出了包含cas9核酸酶的crispr复合物的晶体结构的非限制性实例。sgrna的位置66(尿嘧啶)由于靠近crispr效应蛋白的精氨酸1171而被突出显示为适用于修饰。
[0186]
图13示出了包含cas9核酸酶的crispr复合物的晶体结构的非限制性实例。sgrna的位置63(尿嘧啶)由于靠近crispr效应蛋白的精氨酸64和赖氨酸65而被突出显示为适用于双吖丙啶修饰。
[0187]
b.用于锁定的光反应性交联剂
[0188]
本文提供的crispr多核苷酸可以包含一个或更多个光不稳定接头,例如,芳基叠氮化物(苯基叠氮化物)和双吖丙啶。一个或更多个光不稳定基团(接头)可以用于光化学交联反应,所述光化学交联反应可以使用来自光的能量来引发。一个或更多个光不稳定基团可以是当暴露于紫外光或可见光时变成有反应性的化学惰性化合物。用于生物缀合技术的掺入到交联化合物中的一个或更多个光不稳定基团可以是芳基叠氮化物、叠氮基
‑
甲基
‑
香豆素、二苯甲酮、蒽醌、重氮化合物、双吖丙啶和补骨脂素衍生物。
[0189]
crispr多核苷酸(例如,sgrna)可以用补骨脂素修饰以用于与crispr效应蛋白的交联反应。补骨脂素可以专一地与rna或dna反应,并且可以用于标记核酸或将crispr效应蛋白与crispr多核苷酸交联。可以掺入crispr多核苷酸的用光不稳定基团修饰的核苷酸可以包括4
‑
硫代
‑
utp、5
‑
叠氮基
‑
upt、5
‑
溴
‑
utp、8
‑
叠氮基
‑
atp、
‑
apas
‑
utp、8
‑
n(3)amp、5
‑
[n
‑
(4
‑
苯甲酰基
‑
苯甲酰基)
‑3‑
氨基烯丙基]
‑
脱氧尿苷三磷酸(bp
‑
dutp,二苯甲酮修饰的)、5
‑
[n
‑
(4
‑
叠氮基
‑
2,3,5,6
‑
四氟苯甲酰基)
‑3‑
氨基烯丙基]
‑
脱氧尿苷三磷酸(fab
‑
dutp,全氟
化芳基叠氮化物修饰的)、5
‑
{n
‑
[4
‑
[3
‑
(三氟甲基)
‑
双吖丙啶
‑3‑
基]苯甲酰基]
‑3‑
氨基烯丙基}
‑
脱氧尿苷三磷酸(db
‑
dutp,双吖丙啶修饰的)和5
‑
[n
‑
(对叠氮苯甲酰基)
‑3‑
氨基烯丙基]
‑
脱氧尿苷三磷酸(ab
‑
dutp,芳基叠氮化物修饰的)。
[0190]
交联可以通过光引发。可以放置产生紫外范围内和紫外范围附近的波长的发光装置,使得携带与crispr效应蛋白结合(例如,通过氢键合)的crispr多核苷酸(例如,sgrna)的溶液在通过发光装置时可以暴露于该波长。如以上描述的,这种暴露可以导致光不稳定基团的光引发和将crispr多核苷酸(例如,sgrna)与crispr效应蛋白连接的共价键的形成。
[0191]
用于光引发的光的波长范围可以是220
‑
465nm。暴露方案中的光强可以是约15mw/cm2、20mw/cm2、25mw/cm2、35mw/cm2、40mw/cm2、50mw/cm2、70mw/cm2、90mw/cm2、110mw/cm2、120mw/cm2、140mw/cm2、160mw/cm2、175mw/cm2、190mw/cm2、200mw/cm2、220mw/cm2、240mw/cm2、260mw/cm2、280mw/cm2、300mw/cm2、320mw/cm2、340mw/cm2、360mw/cm2、380mw/cm2、400mw/cm2、420mw/cm2、440mw/cm2、460mw/cm2、480mw/cm2、500mw/cm2、520mw/cm2、540mw/cm2、560mw/cm2、580mw/cm2、600mw/cm2、620mw/cm2、650mw/cm2、675mw/cm2、700mw/cm2、720mw/cm2、745mw/cm2、765mw/cm2、790mw/cm2、810mw/cm2、830mw/cm2、850mw/cm2、870mw/cm2、900mw/cm2、920mw/cm2、945mw/cm2、965mw/cm2、985mw/cm2、1000mw/cm2、1025mw/cm2、1050mw/cm2、1080mw/cm2、1100mw/cm2、1125mw/cm2、1150mw/cm2、1175mw/cm2、1200mw/cm2、1240mw/cm2、1275mw/cm2、1290mw/cm2、1320mw/cm2、1350mw/cm2、1380mw/cm2、1400mw/cm2、1420mw/cm2、1450mw/cm2、1470mw/cm2、1490mw/cm2、1520mw/cm2、1540mw/cm2、1560mw/cm2、1600mw/cm2、1630mw/cm2、1650mw/cm2、1670mw/cm2、1700mw/cm2、1720mw/cm2或1750mw/cm2。暴露方案中使用的光的功率瓦数可以是如通过oai 306uv功率计测量的约50w、70w、80w、90w、100w、120w、140w、160w、175w、190w、210w、230w、250w、270w、290w、310w、330w、250w、370w、390w、420w、450w、480w、500w、530w、550w、570w、600w、620w、650w、670w、700w、720w、750w、770w、800w、820w、850w、870w、900w、920w、950w、970w、1000w、1020w、1050w、1070w、1100w、1120w、1200w、1300w、1400w、1500w、1600w、1700w、1800w、1900w、2000w、2100w、2200w、2300w、2400w、2500w、2600w、2700w、2800w、2900w、3000w、3100w、3200w、3300w、3400w、3500w、3600w、3700w、3800w、3900w、4000w、4100w、4200w、4300w、44004500w、4600w、4700w、4800w、4900w、5000w、5100w、5200w、5300w、5400w、5500w、5600w、5700w、5800w、5900w或6000w。
[0192]
暴露的持续时间可以是1秒至30分钟。暴露方案可以包括连续暴露或脉冲暴露或两者。脉冲暴露可以是均匀的或具有不同的持续时间。引发交联的暴露时间可以取决于所选择的交联剂。例如,在暴露于uv光时,双吖丙啶可以产生半衰期为纳秒量级的反应性碳烯(carbene)。芳基叠氮化合物在暴露于uv光时可以形成半衰期为毫秒量级的反应性碳烯。暴露时间也可以取决于从用于反应的可用c
‑
h基团至碳烯的距离。
[0193]
用于交联的一个或更多个光不稳定基团可以被一定波长的光激活。该波长可以通过特定频率的光子提供对光不稳定基团的电子壳层的激发。反应可以在激发时发生,这可以提供交联反应关于共价键形成时间的灵活性。可以选择一个或更多个光不稳定基团以便被紫外波长激活。
[0194]
交联可以在体外发生。生理条件可以用于确保crispr效应蛋白与crispr多核苷酸的正确折叠和附接。生理条件可以包括含有试剂(例如,20mm tris,ph 7.5,100mm kcl、5mm mgcl2、1mm dtt和5%(v/v)甘油)的溶液。温度可以是约25℃或37℃。
[0195]
为了促进由单独的crispr多核苷酸(例如,sgrna)和crispr效应蛋白形成crispr复合物,提供的比例(例如,摩尔比)(crispr多核苷酸:crispr效应蛋白)可以是约0.001:1、0.01:1、0.1:1、0.2:1、0.3:1、0.4:1、0.5:1、0.6:1、0.7:1、0.8:1、0.9:1、1:1、2:1、3:1、4:1、5:1、6:1、7:1、8:1、9:1、10:1、11:1、12:1、13:1、14:1、15:1、16:1、17:1、18:1、19:1、20:1、21:1、22:1、25:1、30:1、40:1、50:1、60:1、70:1、80:1、90:1、100:1以及之间的任何变化。比例(例如,摩尔比)(crispr多核苷酸:crispr效应蛋白)可以是约0.001:1至约0.01:1、约0.01至约0.1:1、约0.1:1至约1:1、约1:1至约10:1或约10:1至约100:1或约100:1至约1000:1。
[0196]
交联也可以在体内发生,例如,在细胞与未结合crispr多核苷酸或结合crispr多核苷酸的crispr效应蛋白溶液接触之后。在一些情况下,crispr多核苷酸(例如,sgrna)和/或crispr效应蛋白(例如,cas9)可以由细胞中的核酸表达。可以将细胞暴露于uv光,以便将crispr多核苷酸(例如,sgrna)与crispr效应蛋白(例如,cas9)锁定(例如,共价交联)。细胞可以是外胚层(例如,神经元和成纤维细胞)、中胚层(例如,心肌细胞)、内胚层(例如,胰腺细胞)、上皮(例如,肺道和鼻道)、中性粒细胞、嗜酸性粒细胞、嗜碱性粒细胞、淋巴细胞、破骨细胞、内皮细胞、造血细胞、红细胞等。细胞可以来源于特定细胞系,诸如cho细胞(例如,chokl)、hek293细胞、caco2细胞、u2
‑
os细胞、nih 3t3细胞、nso细胞、sp2细胞、dg44细胞、k
‑
562细胞,u
‑
937细胞、mc5细胞、imr90细胞、jurkat细胞、hepg2细胞、hela细胞、ht
‑
1080细胞、hct
‑
116细胞、hu
‑
h7细胞、huvec细胞和molt 4细胞。适用于本公开内容范围的其他细胞的实例可以包括干细胞、胚胎干细胞(esc)和诱导性多能干细胞(ipsc)、msc
‑
1、k562等。
[0197]
交联的时间可以取决于所选择的交联官能团。在光反应性交联剂的情况下,crispr效应蛋白/crispr多核苷酸复合物暴露于例如光,可以发生在crispr效应蛋白和crispr多核苷酸在溶液中混合在一起之后。
[0198]
暴露于光(例如,uv光)的持续时间可以是1秒至30分钟。暴露于光(例如,uv光)的持续时间可以小于2秒、小于5秒、小于10秒、小于20秒、小于30秒、小于45秒、小于50秒、小于1分钟、小于2分钟、小于5分钟、小于10分钟、小于15分钟、小于20分钟、小于30分钟、小于45分钟。
[0199]
2.调节crispr活性的修饰
[0200]
在一些情况下,例如,为了增加crispr多核苷酸(例如,grna或sgrna)的有效性,可以向crispr多核苷酸(例如,grna或sgrna)添加一种或更多种修饰,所述修饰降低与crispr酶复合的crispr多核苷酸的脱靶编辑活性。一种或更多种修饰可以位于各种位置处,包括糖部分、磷酸二酯连接和/或碱基处。例如,crispr多核苷酸可以包含含有磷酰胺、硫代磷酸酯、二硫代磷酸酯、硼烷磷酸酯连接、o
‑
甲基亚磷酰胺连接和/或肽核酸的主链。一种或更多种修饰可以包括2'氟
‑
阿拉伯核酸、三环
‑
dna(tc
‑
dna)、肽核酸、环己烯核酸(cena)、锁核酸(lna)、在核糖环的2'和4'碳之间包含亚甲基桥的锁核酸(lna)核苷酸、桥接核酸(bna)、亚乙基桥接核酸(ena)、磷酰二亚胺基吗啉代或其组合。
[0201]
用至少一个非天然核苷酸修饰以与crispr效应蛋白交联的crispr多核苷酸可以包含被配置为促进裂解特性的序列。用至少一个非天然核苷酸修饰以与crispr效应蛋白交联的crispr多核苷酸的裂解特性可以通过可裂解元件来改变,该可裂解元件可以在适当的条件下改变crispr多核苷酸在可裂解元件掺入点处的裂解倾向。“可裂解元件”可以包含天
然核苷酸或者一个或更多个修饰的核苷酸。可裂解元件可以在核酸合成期间掺入crispr多核苷酸(例如,sgrna)中。
[0202]
crispr多核苷酸中的两个或更多个可裂解元件可以具有不同的裂解特性,例如,两个或更多个可裂解元件在被掺入crispr多核苷酸(例如,sgrna)中时,可以通过使用不同的媒介和/或反应条件在彼此存在的情况下被选择性地裂解。
[0203]
如本文使用的,术语“裂解(cleaving)”、“裂解(cleaved)”和“裂解(cleavage)”都可以指crispr多核苷酸(例如,sgrna)基本上在crispr多核苷酸(例如,sgrna)中可裂解元件出现的每一点处的切割。
[0204]
裂解可以由媒介引发。媒介可以是例如引起可裂解元件裂解的化学实体或物理力。媒介可以是化学物质或化学物质的组合、生物分子或生物分子的组合、正常或相干(激光)可见光或紫外(uv)光、热或其他形式的电磁能。在一些情况下,可以同时或依次使用媒介的组合,例如两种或更多种媒介,来裂解crispr多核苷酸(例如,sgrna)。同时意指crispr多核苷酸(例如,sgrna)可以在同一时间暴露于两种或更多种媒介,尽管两种或更多种媒介可以一次一种地与crispr多核苷酸(例如,sgrna)反应。依次意指crispr多核苷酸(例如,sgrna)可以与一种媒介接触,并且然后在以后的时间与第二种媒介接触。
[0205]
包含一个或更多个非天然核苷酸以与crispr效应蛋白交联的crispr多核苷酸可以包含多于一种类型的可裂解元件。在一些实例中,第一可裂解元件和第二可裂解元件具有相同的裂解特性。在一些实例中,第二可裂解元件具有与第一可裂解元件不同的裂解特性。例如,第一可裂解元件可以是可光裂解接头,并且第二可裂解元件对由化学核酸酶引起的裂解敏感。在另一实例中,第一可裂解元件可以由化学核酸酶引起的裂解敏感,并且第二可裂解元件可以被工程化为可光裂解,从而允许应用正交处理方案。在一些情况下,相同的可裂解元件可以具有多于一种类型的裂解特性。第一可裂解元件和第二可裂解元件可以是本文描述的任何可裂解元件。
[0206]
可裂解元件(例如,可裂解接头)可以指能够连接crispr多核苷酸(例如,sgrna或crrna)的两种或更多种成分使得crispr多核苷酸(例如,sgrna或crrna)在适当的条件下对裂解敏感的实体。例如,适当条件可以是暴露于uv光。可裂解接头可以包含一个或更多个在适当条件下对切割敏感的修饰或未修饰的核苷酸。
[0207]
可裂解接头可以包含修饰的核苷间连接。修饰的核苷间连接可以是具有磷原子的核苷酸间连接,也可以是没有磷原子的核苷酸间连接。其中包含磷原子的核苷间连接包括,例如,二硫代磷酸酯、磷酸三酯、氨基烷基磷酸三酯、甲基膦酸酯和其他烷基膦酸酯包括3
′‑
亚烷基膦酸酯、5
′‑
亚烷基膦酸酯和手性膦酸酯、亚膦酸酯、磷酰胺酯包括3
′‑
氨基磷酰胺酯和氨基烷基磷酰胺酯、p
‑
乙氧基磷酸二酯(p
‑
ethyoxyphosphodiester)、p
‑
乙氧基磷酸二酯(p
‑
ethoxyphosphodiester)、p
‑
烷氧基磷酸三酯、甲基膦酸酯、硫代磷酰胺酯(thionophosphoramidate)、硫代烷基膦酸酯(thionoalkylphosphonate)、硫代烷基磷酸三酯、硒代磷酸酯和硼磷酸酯,以及包含非磷的连接,例如,诸如本领域已知的缩醛和酰胺,具有正常3
’‑5’
连接、2
’‑5’
连接的类似物的那些以及具有反向极性的那些(其中一个或更多个核苷酸间连接是3’至3’、5’至5’或2’至2’连接)。具有反向极性的多核苷酸可以在最3
′
侧的核苷酸间连接处包含单个3
′
至3
′
连接,即,可以是无碱基的单个反向核苷残基(核酸碱基丢失或具有代替其的羟基基团)。
[0208]
包含非磷的核苷间连接包括短链烷基、环烷基、混合杂原子烷基、混合杂原子环烷基、一个或更多个短链杂原子和一个或更多个短链杂环。这些核苷间连接包括但不限于硅氧烷、硫化物、亚砜、砜、乙酰基、甲酰基(formacetyl)、硫代甲酰基(thioformacetyl)、亚甲基甲酰基(methylene formacetyl)、硫代甲酰基(thioformacetyl)、烯基、氨基磺酸酯;亚甲基亚氨基、亚甲基肼基、磺酸酯、磺酰胺、酰胺和其他具有混合的n、o、s和ch2组分部件的核苷间连接。其他不包含磷原子的修饰的核苷间连接包括
‑
ch2
‑
nh
‑
o
‑
ch2
‑
、
‑
ch2
‑
n(ch3)
‑
o
‑
ch2
‑
(称为亚甲基(甲基亚氨基)主链)、
‑
ch2
‑
o
‑
n(ch3)
‑
ch2
‑
、
‑
ch2
‑
n(ch3)
‑
n(ch3)
‑
ch2
‑
和
‑
o
‑
n(ch3)
‑
ch2
‑
ch2
‑
。
[0209]
可裂解接头本质上可以是非核苷酸的。“非核苷酸”可以指能够在一个或更多个核苷酸单元的位置中(包括糖取代和/或磷酸取代)掺入多核苷酸链的任何基团或化合物。基团或化合物可以是无碱基的,因为它不包含通常公认的核苷酸碱基,诸如腺嘌呤、鸟嘌呤、胞嘧啶、尿嘧啶或胸腺嘧啶,例如在糖的c1位置处。
[0210]
非核苷酸接头可以是例如无碱基残基(d间隔物(dspacer)),多甘醇(oligoethyleneglycol),诸如三甘醇(间隔物9)或六甘醇(间隔物18),或烷烃二醇(alkane
‑
diol),诸如丁二醇。间隔物单元可以优选地通过磷酸二酯键或硫代磷酸酯键连接。接头单元可以仅在分子中出现一次,或者可以被掺入若干次,例如通过磷酸二酯、硫代磷酸酯、甲基膦酸酯或酰胺连接。其他优选的接头是烷基氨基接头,诸如c3、c6、c12氨基接头,以及烷基硫醇接头,诸如c3或c6硫醇接头。在一些实例中,异双官能和同双官能连接部分可以用于将肽和蛋白缀合至核苷酸。实例包括5
′‑
氨基
‑
修饰物c6和3
′‑
氨基
‑
修饰物c6试剂。
[0211]
a.crispr on
[0212]
本文提供了能够与crispr效应蛋白共价交联以形成crispr on复合物的crispr on多核苷酸。crispr on多核苷酸可以包含(i)被配置为与靶分子中的靶序列退火的引导序列,(ii)被配置为与crispr效应蛋白结合的序列(例如,tracrrna序列),和(iii)引导序列5’侧的第一序列元件。引导序列5’侧的第一序列元件可以称为多核苷酸前导序列。第一序列元件可以包含二级结构,例如,茎环。茎环可以包含约3个碱基对(bp)至约30bp。第一序列元件的5’末端可以与序列元件中紧接引导序列5’侧的碱基退火。在一些情况下,第一序列元件的5’末端与引导序列退火。crispr on多核苷酸还可以包含第一可裂解元件,例如,第一非天然存在的可裂解元件,例如,光不稳定接头。可裂解元件可以位于紧接引导序列的5’侧。可裂解元件可以对由光、小分子或一个或更多个细胞过程引起的裂解敏感。多核苷酸前导序列可以干扰引导序列与靶序列退火的能力。
[0213]
可以组装包含crispr效应蛋白和交联的crispr on多核苷酸的复合物(参见,例如,图17a)。包含具有引导序列5’侧的第一序列元件的交联的crispr on多核苷酸和crispr效应蛋白的crispr复合物可以比包含没有第一序列元件的交联的crispr多核苷酸的crispr复合物具有更低的靶特异性活性;例如,活性可以低约2倍至约100倍。本文提供了用于crispr复合物对靶核酸(例如,dna)的可调节靶向的方法。该方法可以包括用裂解媒介裂解可裂解元件(参见,例如,图17a),从而释放引导序列5’侧的第一序列元件(参见,例如,图17c)。例如,可裂解元件可以是光不稳定接头,并且当暴露于光时,光不稳定接头可以被裂解。裂解可裂解接头可以产生比裂解之前的crispr复合物具有更高的靶特异性裂解活性的
crispr复合物。
[0214]
crispr on多核苷酸或crispr on/off多核苷酸可以包含引导序列5’侧的第一序列元件。引导序列5’侧的第一序列元件可以称为多核苷酸前导序列。包含具有多核苷酸前导序列的crispr多核苷酸和与该crispr多核苷酸交联的crispr效应蛋白的crispr复合物可以比包含没有多核苷酸前导序列的crispr多核苷酸的crispr复合物具有更低的活性。多核苷酸前导序列的去除可以产生活性增加的crispr复合物(crispr on)。
[0215]
i.多核苷酸前导序列的长度
[0216]
多核苷酸前导序列的长度范围可以为约1个核苷酸至约50个核苷酸,例如,约5个核苷酸至约30个核苷酸、约10个核苷酸至约20个核苷酸、约15个核苷酸或至少4个核苷酸、3个核苷酸至约15个核苷酸,例如,约5个核苷酸至约15个核苷酸、约3个核苷酸至约10个核苷酸、约3个至约15个核苷酸或约3个核苷酸至约12个核苷酸、约4个核苷酸至约13个核苷酸、约3个核苷酸至约18个核苷酸、约4个核苷酸至约19个核苷酸、4个核苷酸至约30个核苷酸、4个核苷酸至约25个核苷酸、5个核苷酸至约12个核苷酸、5个核苷酸至约至少4个核苷酸或30个或更少的核苷酸。
[0217]
ii.多核苷酸前导序列的组成
[0218]
多核苷酸前导序列可以包含核糖核苷酸和/或脱氧核糖核苷酸。多核苷酸前导序列可以包含非典型核苷酸或核苷酸类似物。多核苷酸前导序列可以包含本文描述的任何核苷酸或修饰的核苷酸或核苷酸间连接。在一些情况下,多核苷酸前导序列可以包含本文描述的任何接头。
[0219]
iii.多核苷酸前导序列中的二级结构
[0220]
多核苷酸前导序列可以形成或被设计为形成二级结构。二级结构可以是例如茎环结构。茎环的茎可以包含至少约3bp,包含互补的x和y序列(其中x表示茎的一条链的序列,并且y表示茎的另一条链的序列)。茎可以包含至少(或至多)2个、3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个、20个、21个、22个、23个、24个、25个、26个、27个、28个、29个、30个、31个、32个、33个、34个、35个、36个、37个、38个、39个、40个碱基对。茎可以包含范围为1
‑
20bp或2
‑
5bp、2
‑
9bp、3
‑
10bp、4
‑
9bp、5
‑
10bp、5
‑
20bp、6
‑
20bp、7
‑
20bp、8
‑
20bp等的双链结构域。在一些情况下,茎的两条链可以共价交联。
[0221]
茎环可以包含单链环。单链环范围可以为1
‑
50个碱基,例如,3
‑
5个碱基、3
‑
7个碱基、4
‑
10个碱基、5
‑
20个碱基、6
‑
25个碱基、3
‑
25个碱基、3
‑
30个碱基、4
‑
30个碱基或4
‑
50个碱基。
[0222]
茎环或多核苷酸前导序列的最5’侧碱基可以与多核苷酸前导序列中紧接引导序列5’侧的碱基退火。在一些情况下,多核苷酸前导序列的最5’侧碱基可以与距引导序列的最5’侧碱基的3’侧1
‑
20个碱基(例如,距引导序列的最5’侧碱基的3’侧2个碱基、3个碱基、4个碱基、5个碱基、6个碱基、7个碱基、8个碱基、9个碱基、10个碱基、11个碱基、12个碱基、15个碱基或20个碱基)的碱基退火。在一些情况下,多核苷酸前导序列不包含与引导序列中的碱基进行碱基配对的碱基。
[0223]
多核苷酸前导序列可以形成包含一个或更多个凸起(bulge,单链序列的区域;这些区域可以对应于二级结构中包含小于100%序列碱基配对的位置)的发夹环或茎环结构。一个或更多个凸起的数目、长度和/或位置可以不同,并且可以影响茎环结构的整体稳定
性。当最佳折叠时,多核苷酸前导序列可以包含2个、3个、4个、5个或更多个凸起。
[0224]
在一些情况下,多核苷酸前导序列可以包含非多核苷酸部分。多核苷酸前导序列中的非核苷酸部分可以是生物素、抗体、肽、亲和部分、报告物或蛋白部分(诸如nhs酯或异硫氰酸酯)、地高辛、酶诸如碱性磷酸酶等。
[0225]
在一些情况下,多核苷酸前导序列缺乏二级结构。多核苷酸前导序列可以包含单链连续核苷酸链段或由单链连续核苷酸链段组成。
[0226]
由多核苷酸前导序列形成的茎环的解链温度可以是约25℃至约60℃、或约30℃至约50℃或约40℃至约50℃。
[0227]
iv.由于多核苷酸前导序列导致的活性降低
[0228]
包含具有多核苷酸前导序列的crispr多核苷酸和与该crispr多核苷酸交联的crispr效应蛋白的crispr复合物可以比包含没有多核苷酸前导序列的crispr多核苷酸的crispr复合物具有更低的活性。在一些情况下,活性低至少(或至多)0.1倍、0.25倍、0.5倍、0.75倍、1倍、2倍、5倍、10倍、50倍、100倍或1000倍。在一些情况下,包含具有多核苷酸前导序列的crispr多核苷酸和crispr效应蛋白的crispr复合物没有活性。活性可以是例如酶促活性或转录激活活性。例如,当crispr效应蛋白是具有催化活性的cas蛋白时,crispr复合物可以不能够裂解靶核酸。在另一种实例中,当crispr效应蛋白是与转录激活结构域融合的没有催化活性的cas蛋白(catalytically dead cas)时,crispr复合物可以不能够激活靶基因的转录。
[0229]
v.去除多核苷酸前导序列
[0230]
crispr多核苷酸可以包含一个或更多个可裂解元件,以允许多核苷酸前导序列的释放。一个或更多个可裂解元件可以位于多核苷酸前导序列和引导序列之间。在一些情况下,一个或更多个可裂解元件位于多核苷酸前导序列内。在一些情况下,至少一个可裂解元件位于多核苷酸前导序列内,并且至少一个可裂解元件位于多核苷酸前导序列和引导序列之间。在一些情况下,一个或更多个可裂解元件位于引导序列的5’侧。一个或更多个可裂解元件可以是至少或至多2个、3个、4个、5个、6个、7个、8个、9个或10个可裂解元件。在一些情况下,一个或更多个可裂解元件被定位为使得裂解后,多核苷酸前导序列的一部分(例如,1个碱基、2个碱基、5个碱基或10个碱基)保持共价连接至引导序列。在一些情况下,一个或更多个可裂解元件被定位为使得裂解后,没有多核苷酸前导序列保持共价附接至引导序列。
[0231]
一个或更多个可裂解元件可以是本文描述的任何可裂解元件。一个或更多个可裂解元件可以是同一类型的可裂解元件或不同类型的可裂解元件。
[0232]
crispr多核苷酸可以在crispr多核苷酸未与crispr效应蛋白结合时在一个或更多个可裂解元件处被裂解。crispr多核苷酸可以在crispr多核苷酸与crispr效应蛋白交联时在一个或更多个可裂解元件处被裂解。crispr多核苷酸可以在crispr多核苷酸与crispr效应蛋白交联并与靶序列结合时在一个或更多个可裂解元件处被裂解。在一些情况下,多核苷酸前导序列阻止crispr多核苷酸与crispr效应蛋白交联或使crispr多核苷酸与crispr效应蛋白交联的能力相对于缺少多核苷酸前导序列的crispr多核苷酸降低;从crispr多核苷酸裂解多核苷酸前导序列可以增加crispr多核苷酸结合crispr效应蛋白的能力。
[0233]
crispr多核苷酸可以在体外在一个或更多个可裂解元件处被裂解。crispr多核苷
酸可以在细胞或生物体(例如,小鼠、兔、山羊、灵长类动物,例如,黑猩猩、大猩猩或人类)中时在一个或更多个可裂解元件处被裂解。
[0234]
crispr多核苷酸在一个或更多个可裂解元件处裂解的时机可以不同。例如,一个或更多个可裂解元件可以在crispr多核苷酸引入细胞或生物体之后立即被裂解,或者在引入细胞或生物体之后至少(或至多)0.25小时、0.5小时、0.75小时、1小时、2小时、3小时、4小时、5小时、6小时、7小时、8小时、9小时、10小时、11小时、12小时、13小时、14小时、15小时、16小时、17小时、18小时、19小时、20小时、21小时、22小时、23小时、24小时、48小时、72小时或96小时被裂解。
[0235]
crispr多核苷酸可以暴露于裂解媒介一次。crispr多核苷酸可以经历裂解媒介多于一次,例如,2次、3次、5次或10次。crispr多核苷酸可以暴露于多于一种类型的裂解媒介,例如,至少(或至多)2种、3种、4种、5种、6种、7种、8种、9种或10种裂解媒介。
[0236]
crispr多核苷酸可以暴露于裂解媒介不同的持续时间。例如,crispr多核苷酸可以暴露于裂解媒介0.1min、0.5min、1min、2min、3min、4min、5min、10min、30min、60min、2hr、4hr、6hr、12hr、24hr、48hr、72hr或96hr。
[0237]
在一些情况下,样品包含多于一个crispr多核苷酸,并且裂解媒介可以用于裂解一定百分比的crispr多核苷酸。例如,裂解媒介可以用于裂解样品中至少(或至多)5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%或99%的crispr多核苷酸。一定剂量的裂解媒介可以用于裂解样品中100%的crispr多核苷酸。裂解量可以在至少(或至多)1min、5min、10min、15min、30min、45min、1hr、2hr、6hr、12hr、24hr、48hr、72hr或96hr内发生。
[0238]
多核苷酸前导序列的释放可以导致与crispr多核苷酸结合的crispr效应蛋白(例如,crispr酶,例如,cas9)的活性增加。在一些情况下,在样品中,多核苷酸前导序列的释放导致活性增加至少0.1倍、0.25倍、0.5倍、0.75倍、1倍、2倍、5倍、10倍、50倍、100倍或1000倍。
[0239]
vi.其他特征
[0240]
包含多核苷酸前导序列的crispr多核苷酸可以包含第二组一个或更多个元件,所述第二组一个或更多个元件可以经历特异性修饰以产生修饰的crispr多核苷酸,当所述修饰的crispr多核苷酸与crispr效应蛋白复合时,形成具有较低靶特异性裂解活性的第二crispr复合物。第二组一个或更多个元件可以是第二组一个或更多个可裂解元件。例如,crispr多核苷酸可以包含多核苷酸前导序列和被配置为允许释放多核苷酸前导序列的第一组一个或更多个可裂解元件以及被配置为允许裂解剩余crispr多核苷酸的第二组一个或更多个可裂解元件;该多核苷酸可以称为crispr on/off多核苷酸。
[0241]
b.crispr off
[0242]
本文提供了能够与crispr效应蛋白交联以形成crispr off复合物的crispr off多核苷酸。crispr off多核苷酸可以包含(i)被配置为结合crispr效应蛋白的序列(例如,tracrrna序列)和(ii)可裂解接头。在一些情况下,crispr off多核苷酸还包含被配置为与靶分子中的靶序列退火的引导序列。可裂解接头可以是非天然存在的可裂解接头。如果crispr off多核苷酸包含引导序列,则可裂解接头可以位于引导序列中最5’侧的碱基的3’侧(参见,例如,图18)。可裂解接头可以位于被配置为交联crispr效应蛋白的序列(例如,
tracrrna序列)内。在一些情况下,紧接可裂解接头3’侧和/或紧接可裂解接头5’侧的碱基不与crispr off多核苷酸中的另一个碱基退火。可裂解接头可以是光不稳定接头。可裂解接头可以对由光、小分子或一个或更多个细胞过程引起的裂解敏感。
[0243]
与crispr off多核苷酸复合的crispr效应蛋白的脱靶编辑活性可以小于与非crispr
‑
off多核苷酸(例如,不含一个或更多个可裂解接头的sgrna)复合的crispr效应蛋白的脱靶编辑活性。脱靶编辑活性(例如,如按本文描述测量的)可以被降低以下倍:约1.1、1.5、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59或60;至少1.1、1.5、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59或60;或至多1.1、1.5、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59或60。在一些情况下,降低发生在不暴露于裂解媒介(例如,uv光)的情况下;在一些情况下,降低发生在暴露于裂解媒介之后。包含与crispr off多核苷酸(在位置57和/或位置74处具有可裂解接头)复合的crispr效应蛋白的复合物可以具有比与不含可裂解接头的sgrna复合的crispr效应蛋白更低的脱靶编辑效率。包含与crispr off多核苷酸复合的crispr效应蛋白的复合物可以具有与与非crispr off多核苷酸复合的crispr效应蛋白的中靶编辑效率相同或在其的1%、2%、3%、4%或5%以内的中靶编辑效率。例如,没有可裂解接头的sgrna。
[0244]
可以组装包含与crispr off多核苷酸交联的crispr效应蛋白的复合物。本文提供了用于crispr复合物对靶dna的可调节靶向的方法。该方法可以包括使可裂解接头裂解。可裂解接头的裂解可以导致crispr复合物具有比裂解之前更低的靶特异性裂解活性。在一些情况下,可裂解接头的裂解可以引起由裂解产生的但未与crispr效应蛋白交联的crispr off多核苷酸片段与crispr效应蛋白解离。在一些情况下,可裂解接头的裂解致使crispr复合物失活。
[0245]
i.一个或更多个可裂解元件的位置
[0246]
一个或更多个可裂解元件可以位于引导序列中最5’侧碱基(或核苷酸)的3’侧或引导序列中最3’侧碱基(或核苷酸)的5’侧。一个或更多个可裂解元件可以位于距crrna或引导序列的5’末端的3'侧约1
‑
30个碱基(例如,2个碱基、3个碱基、4个碱基、5个碱基、6个碱基、7个碱基、8个碱基、9个碱基、10个碱基、11个碱基、12个碱基、13个碱基、14个碱基、15个碱基、16个碱基、17个碱基、18个碱基、19个碱基、20个碱基、21个碱基、22个碱基、23个碱基、24个碱基、25个碱基、26个碱基、27个碱基、28个碱基、29个碱基或30个碱基)处。一个或更多个可裂解元件可以位于距crrna序列或引导序列的3’末端的5’侧约1
‑
30个碱基(例如,2个碱基、3个碱基、4个碱基、5个碱基、6个碱基、7个碱基、8个碱基、9个碱基、10个碱基、11个碱基、12个碱基、13个碱基、14个碱基、15个碱基、16个碱基、17个碱基、18个碱基、19个碱基、20个碱基、21个碱基、22个碱基、23个碱基、24个碱基、25个碱基、26个碱基、27个碱基、28个碱基、29个碱基或30个碱基)处。
[0247]
一个或更多个可裂解元件可以位于crispr多核苷酸的被配置为与crispr效应蛋
白(例如,cas9)结合的序列(例如,tracrrna)序列中。在一些情况下,一个或更多个可裂解元件可以是tracr序列5’末端的3’侧的1
‑
30个碱基,诸如,2个碱基、3个碱基、4个碱基、5个碱基、6个碱基、7个碱基、8个碱基、9个碱基、10个碱基、11个碱基、12个碱基、13个碱基、14个碱基、15个碱基、16个碱基、17个碱基、18个碱基、19个碱基、20个碱基、21个碱基、22个碱基、23个碱基、24个碱基、25个碱基、26个碱基、27个碱基、28个碱基、29个碱基或30个碱基。在一些情况下,一个或更多个可裂解元件可以是tracr序列3’末端的5’侧的1
‑
30个碱基,诸如,2个碱基、3个碱基、4个碱基、5个碱基、6个碱基、7个碱基、8个碱基、9个碱基、10个碱基、11个碱基、12个碱基、13个碱基、14个碱基、15个碱基、16个碱基、17个碱基、18个碱基、19个碱基、20个碱基、21个碱基、22个碱基、23个碱基、24个碱基、25个碱基、26个碱基、27个碱基、28个碱基、29个碱基或30个碱基。
[0248]
在一些实例中,一个或更多个可裂解元件可以位于crispr多核苷酸(例如,sgrna)中紧接碱基(或核苷酸)56和/或核苷酸73的5’侧或3’侧或者替代核苷酸57和/或核苷酸74,其中crispr多核苷酸的引导序列(例如,sgrna)的最5’侧核苷酸是核苷酸1。在一些实例中,一个或更多个可裂解元件可以位于crispr多核苷酸(例如,sgrna)中紧接碱基(或核苷酸)1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99或100的5’侧或3’侧或者替代crispr多核苷酸(例如,sgrna)的碱基1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99或100,其中crispr多核苷酸的引导序列(例如,sgrna)的最5’侧碱基(或核苷酸)是碱基(或核苷酸)1。
[0249]
ii.在暴露于裂解媒介之前一个或更多个裂解元件的影响
[0250]
在一些情况下,相对于不含一个或更多个可裂解元件并与crispr效应蛋白复合的crispr多核苷酸(例如,sgrna),包含一个或更多个可裂解元件并与crispr效应蛋白(例如,cas9)复合的crispr多核苷酸(例如,sgrna)的活性没有降低(例如,在将crispr多核苷酸暴露于裂解媒介之前)。在一些情况下,相对于不含一个或更多个可裂解元件并与crispr效应蛋白复合的crispr多核苷酸(例如,sgrna),包含一个或更多个可裂解元件并与crispr效应蛋白(例如,cas9)复合的crispr多核苷酸(例如,sgrna)确实具有降低的活性(例如,在将crispr多核苷酸暴露于裂解媒介之前)。
[0251]
iii.一个或更多个可裂解元件的裂解
[0252]
一个或更多个可裂解元件可以是本文描述的任何可裂解元件。一个或更多个可裂解元件可以是同一类型的可裂解元件或不同类型的可裂解元件。一个或更多个可裂解元件可以是至少或至多2个、3个、4个、5个、6个、7个、8个、9个或10个可裂解元件。
[0253]
crispr多核苷酸(例如,sgrna)可以在crispr多核苷酸(例如,sgrna)未与crispr效应蛋白(例如,cas9)结合时在一个或更多个可裂解元件处被裂解。crispr多核苷酸(例如,sgrna)可以在crispr多核苷酸(例如,sgrna)与crispr效应蛋白(例如,cas9)复合时在
一个或更多个可裂解元件处被裂解。crispr多核苷酸(例如,sgrna)可以在crispr多核苷酸(例如,sgrna)与crispr效应蛋白(例如,cas9)复合并与靶序列结合时在一个或更多个可裂解元件处被裂解。在一些情况下,crispr多核苷酸(例如,sgrna)的一个或更多个所得片段在裂解后保持与crispr效应蛋白(例如,cas9)结合。在一些情况下,crispr多核苷酸(例如,sgrna)的一个或更多个(或全部)所得片段在裂解后不再结合或不再能够结合crispr效应蛋白(例如,cas9)。
[0254]
crispr多核苷酸可以在体外在一个或更多个可裂解元件处被裂解。crispr多核苷酸可以在体内在一个或更多个可裂解元件处被裂解。crispr多核苷酸可以在细胞或生物体(例如,小鼠、兔、山羊、灵长类动物,例如,黑猩猩、大猩猩或人类)中时在一个或更多个可裂解元件处被裂解。
[0255]
crispr多核苷酸(例如,sgrna)在一个或更多个可裂解元件处裂解的时机可以不同。例如,一个或更多个可裂解元件可以在crispr多核苷酸(例如,sgrna)引入细胞或生物体之后立即被裂解,或者在引入细胞或生物体之后至少(或至多)0.25小时、0.5小时、0.75小时、1小时、2小时、3小时、4小时、5小时、6小时、7小时、8小时、9小时、10小时、11小时、12小时、13小时、14小时、15小时、16小时、17小时、18小时、19小时、20小时、21小时、22小时、23小时、24小时、48小时、72小时或96小时被裂解。
[0256]
crispr多核苷酸(例如,sgrna)可以暴露于裂解媒介一次。crispr多核苷酸(例如,sgrna)可以经历裂解媒介多于一次,例如,2次、3次、5次或10次。crispr多核苷酸(例如,sgrna)可以暴露于多于一种类型的裂解媒介,例如,至少(或至多)2种、3种、4种、5种、6种、7种、8种、9种或10种裂解媒介。
[0257]
crispr多核苷酸(例如,sgrna)可以暴露于裂解媒介不同的持续时间。例如,crispr多核苷酸(例如,sgrna)可以暴露于裂解媒介持续0.1min、0.5min、1min、2min、3min、4min、5min、10min、30min、60min、2hr、4hr、6hr、12hr、24hr、48hr、72hr或96hr。
[0258]
在一些情况下,样品包含多于一个crispr多核苷酸(例如,sgrna),并且裂解媒介可以用于裂解一定百分比的crispr多核苷酸(例如,sgrna)。例如,裂解媒介可以用于裂解样品中至少(或至多)5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%或99%的crispr多核苷酸(例如,sgrna)。一定剂量的裂解媒介可以用于裂解样品中100%的crispr多核苷酸(例如,sgrna)。裂解量可以在至少(或至多)1min、5min、10min、15min、30min、45min、1hr、2hr、6hr、12hr、24hr、48hr、72hr或96hr内发生。
[0259]
裂解可以导致与crispr多核苷酸(例如,sgrna)结合的crispr效应蛋白(例如,crispr酶,例如,cas9)的活性降低。在一些情况下,在样品中,暴露于一种或更多种裂解媒介导致活性降低至少0.1倍、0.25倍、0.5倍、0.75倍、1倍、2倍、5倍、10倍、50倍、100倍或1000倍。在一些情况下,在样品中,暴露于一种或更多种裂解媒介导致活性完全丧失。
[0260]
c.crispr on/off
[0261]
本文提供了能够与crispr效应蛋白交联以形成crispr“on/off”复合物的crispr“on/off”多核苷酸。crispr on/off多核苷酸可以包含被配置为与靶分子中的靶序列退火的引导序列、被配置为与crispr效应蛋白交联的序列(例如,tracrrna序列)和(a)第一元件,所述第一元件被配置为经历第一特异性修饰,产生第一修饰的多核苷酸,当第一修饰的
多核苷酸与crispr效应蛋白交联时,形成第一crispr复合物,与包含未经历第一特异性修饰的多核苷酸的crispr复合物相比,第一crispr复合物具有更高的靶特异性裂解活性;以及(b)第二元件,所述第二元件被配置为经历第二特异性修饰以产生第二修饰的多核苷酸,当第二修饰的多核苷酸与crispr效应蛋白交联时,形成第二crispr复合物,与第一crispr复合物相比,第二crispr复合物具有更低的靶特异性裂解活性。crispr on/off多核苷酸可以包含本文描述的crispr on多核苷酸和crispr off多核苷酸的特征。
[0262]
可以组装包含与crispr on/off多核苷酸交联的crispr效应蛋白的复合物。本文提供了用于crispr复合物对靶dna的可调节靶向的方法。该方法可以包括使crispr on/off多核苷酸的第一元件经历第一特异性修饰,从而产生第一修饰的多核苷酸,当第一修饰的多核苷酸与crispr效应蛋白交联时,形成第一crispr复合物,与包含未经历第一特异性修饰的多核苷酸的crispr复合物相比,第一crispr复合物具有更高的靶特异性裂解活性。该方法还可以包括在使第一元件经历第一修饰之后,使第二元件经历第二特异性修饰,从而形成第二修饰的多核苷酸,当第二修饰的多核苷酸与crispr效应蛋白交联时,形成第二crispr复合物,所述第二crispr复合物具有比第一crispr复合物更低的靶特异性裂解活性。在一些情况下,第二修饰可以引起未与crispr效应蛋白交联的crispr多核苷酸部分片段化和/或与crispr效应蛋白解离。
[0263]
d.crispr off多核苷酸和降低的脱靶编辑
[0264]
crispr多核苷酸可以包含一种或更多种修饰,使得当多核苷酸与crispr效应蛋白(例如,cas9)复合形成crispr复合物时,该crispr复合物在未暴露于光时比具有不含一种或更多种修饰的多核苷酸的crispr复合物具有更低的脱靶裂解活性。一种或更多种修饰可以是本文描述的一种或更多种接头。一种或更多种修饰可以是本文描述的一种或更多种可裂解接头。一种或更多种修饰可以是如本文描述的核糖2’位置处的一种或更多种修饰。一种或更多种修饰可以是一种或更多种可裂解元件。一种或更多种修饰可以包括3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺。crispr off多核苷酸还可以在前三个5’末端rna核苷酸和前三个3’末端rna核苷酸处包含2
’‑
o
‑
甲基类似物和3’硫代磷酸酯核苷酸间连接。
[0265]
a.一种或更多种修饰的位置
[0266]
一种或更多种修饰可以位于引导序列中最5’侧碱基(或核苷酸)的3’侧或引导序列中最3’侧碱基(或核苷酸)的5’侧。一种或更多种修饰可以位于距crrna或引导序列的5’末端的3'侧约1
‑
30个碱基(例如,2个碱基、3个碱基、4个碱基、5个碱基、6个碱基、7个碱基、8个碱基、9个碱基、10个碱基、11个碱基、12个碱基、13个碱基、14个碱基、15个碱基、16个碱基、17个碱基、18个碱基、19个碱基、20个碱基、21个碱基、22个碱基、23个碱基、24个碱基、25个碱基、26个碱基、27个碱基、28个碱基、29个碱基或30个碱基)处。一种或更多种修饰可以位于距crrna序列或引导序列的3’末端的5’侧约1
‑
30个碱基(例如,2个碱基、3个碱基、4个碱基、5个碱基、6个碱基、7个碱基、8个碱基、9个碱基、10个碱基、11个碱基、12个碱基、13个碱基、14个碱基、15个碱基、16个碱基、17个碱基、18个碱基、19个碱基、20个碱基、21个碱基、22个碱基、23个碱基、24个碱基、25个碱基、26个碱基、27个碱基、28个碱基、29个碱基或30个碱基)处。
[0267]
一种或更多种修饰可以位于crispr多核苷酸的被配置为与crispr效应蛋白(例
如,cas9)结合的序列(例如,tracrrna)序列中。在一些情况下,一种或更多种修饰可以位于图1中示出的crispr多核苷酸的四环、连结、茎环1或茎环2中。在一些情况下,一种或更多种修饰可以位于四环的环、四环的凸起、四环的第一茎、四环的第二茎中,位于连结的环结构中,位于连结的茎中,位于茎环1的环结构中,位于茎环1的茎中,位于茎环2的环结构中或在茎环2的茎中;四环、连结、茎环1和茎环2的实例示于图1中。在一些情况下,一种或更多种修饰不包括被配置为形成茎环(例如,与引导序列形成茎环)的引导序列的5’侧序列。在一些情况下,一种或更多种修饰可以是tracr序列5’末端的3’侧的1
‑
30个碱基,诸如,2个碱基、3个碱基、4个碱基、5个碱基、6个碱基、7个碱基、8个碱基、9个碱基、10个碱基、11个碱基、12个碱基、13个碱基、14个碱基、15个碱基、16个碱基、17个碱基、18个碱基、19个碱基、20个碱基、21个碱基、22个碱基、23个碱基、24个碱基、25个碱基、26个碱基、27个碱基、28个碱基、29个碱基或30个碱基。在一些情况下,一种或更多种修饰可以是tracr序列3’末端的5’侧的1
‑
30个碱基,诸如,2个碱基、3个碱基、4个碱基、5个碱基、6个碱基、7个碱基、8个碱基、9个碱基、10个碱基、11个碱基、12个碱基、13个碱基、14个碱基、15个碱基、16个碱基、17个碱基、18个碱基、19个碱基、20个碱基、21个碱基、22个碱基、23个碱基、24个碱基、25个碱基、26个碱基、27个碱基、28个碱基、29个碱基或30个碱基。
[0268]
在一些实例中,一种或更多种修饰可以位于crispr多核苷酸(例如,sgrna)中紧接碱基(或核苷酸)56和/或核苷酸73的5’侧或3’侧或者替代核苷酸57和/或核苷酸74,其中crispr多核苷酸的引导序列(例如,sgrna)的最5’侧核苷酸是核苷酸1。在一些实例中,一个或更多个复合物改变元件可以位于crispr多核苷酸(例如,sgrna)中紧接碱基(或核苷酸)1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99或100的5’侧或3’侧或者替代crispr多核苷酸(例如,sgrna)的碱基1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99或100,其中crispr多核苷酸的引导序列(例如,sgrna)的最5’侧碱基(或核苷酸)是碱基(或核苷酸)1。
[0269]
b.一种或更多种修饰的影响
[0270]
在一些情况下,相对于不含一种或更多种修饰并与crispr效应蛋白复合的crispr多核苷酸(例如,sgrna),包含一种或更多种修饰并与crispr效应蛋白(例如,cas9)复合的crispr多核苷酸(例如,sgrna)在靶序列处的编辑活性没有降低(例如,在将crispr多核苷酸暴露于裂解媒介之前)。在一些情况下,相对于不含一个或更多个复合物改变元件并与crispr效应蛋白复合的crispr多核苷酸(例如,sgrna),包含一种或更多种修饰并与crispr效应蛋白(例如,cas9)复合的crispr多核苷酸(例如,sgrna)在靶序列处确实具有降低的编辑活性(例如,在将crispr多核苷酸暴露于裂解媒介之前)。在一些情况下,在靶序列处的编辑活性相对于标准crispr复合物降低约1%、2%、3%、4%、5%、6%、7%、8%、9%或10%或至多1%、2%或3%、4%、5%、6%、7%、8%、9%或10%。
[0271]
在一些情况下,相对于不含一种或更多种修饰并与crispr效应蛋白复合的crispr多核苷酸(例如,sgrna),包含一种或更多种修饰并与crispr效应蛋白(例如,cas9)复合的crispr多核苷酸(例如,sgrna)在脱靶序列处具有降低的编辑活性(例如,在将crispr多核苷酸暴露于裂解媒介之前)。在脱靶序列处的编辑活性可以描述为脱靶编辑。脱靶编辑可以是在与crispr多核苷酸的引导序列不完全互补的序列处进行编辑。在一些情况下,在脱靶序列处的编辑活性降低约以下、至少以下或至多以下:5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或100%。在一些情况下,脱靶编辑活性为0%、1%、5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或95%。在一些情况下,脱靶编辑活性小于1%、5%、10%、15%、20%、25%、30%、40%、50%、60%、70%、80%、90%或95%。在一些情况下,脱靶编辑活性为0%
‑
5%、5%
‑
10%、10%
‑
25%、25%
‑
50%、50%
‑
75%或75%
‑
95%。
[0272]
在未暴露于光的情况下,与crispr off多核苷酸复合的crispr效应蛋白的脱靶编辑活性可以小于与非crispr off多核苷酸(例如,在前三个5’末端rna核苷酸和前三个3’末端rna核苷酸处仅用2
’‑
o
‑
甲基类似物和3’硫代磷酸酯核苷酸间连接来修饰的sgrna)复合的crispr效应蛋白的脱靶编辑活性。与crispr off多核苷酸复合的crispr效应蛋白在未被裂解时(例如,当未暴露于光时)的脱靶编辑活性在统计学上可以比与非crispr
‑
off多核苷酸复合的crispr效应蛋白更低,其中p值≤0.05、p值≤0.01、p值≤0.005、p值≤0.001、p值≤0.0005或p值≤0.0001。脱靶编辑活性(例如,如按本文描述测量的)可以被降低以下倍:约1.1、1.5、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59或60;至少1.1、1.5、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59或60;或至多1.1、1.5、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59或60。在一些情况下,包含与位置57和/或位置74处具有修饰的crispr off多核苷酸复合的crispr效应蛋白的复合物在未被裂解时(例如,当未暴露于光或另一种裂解诱导处理时)可以比与不含可裂解接头的sgrna复合的crispr效应蛋白具有更低的脱靶编辑效率。包含与crispr off多核苷酸复合的crispr效应蛋白的复合物在未被裂解时(例如,当未暴露于光或另一种裂解诱导处理时)可以具有与与非crispr off多核苷酸(例如,不含可裂解接头的sgrna)复合的crispr效应蛋白的中靶编辑活性相同或在其的1%、2%、3%、4%或5%内的中靶编辑效率。
[0273]
在一些情况下,脱靶编辑活性在一个核酸区域处测量。脱靶编辑活性可以在多于一个基因组区域(例如,基因)处测量。脱靶编辑活性可以在1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、25个、50个、75个或100个基因组区域(例如,基因)处测量。脱靶编辑活性可以在1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、25个、50个、75个、100个、1000个或10,000个基因组区域(例如,基因)处测量。脱靶编辑活性可以在至多1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、25个、50个、75个、100个、1000个或10,000个基因组区域(例如,基因)处测量。
[0274]
脱靶编辑活性可以通过分析来自与crispr复合物接触的细胞的核酸分子来测量。测量可以在转染之后约或至多30分钟、1小时、2小时、5小时、10小时、12小时、24小时、36小时、48小时、60小时、72小时、4天、5天或6天使用从细胞提取的核酸分子进行。crispr复合物可以通过转化引入细胞中。核酸分子可以通过例如,测序、pcr、质谱法、dna印迹等进行分析。脱靶编辑可以被可视化,例如,通过在例如图(例如,散点图)中呈现数据。
[0275]
相比于与不含如本文描述的修饰的sgrna复合的cas9,包含crispr多核苷酸的crispr复合物可以用于降低脱靶编辑。脱靶编辑可以使用以下技术来确定:ice(inference of crispr editing,crispr编辑推理),ice通过分析sanger测序跟踪和提供序列断裂水平的信息以确定插入/缺失形成频率来测量基因编辑的量(如hsiau等人“inference of crispr edits from sanger trace data”,january 14,2019biorxiv中描述的);或深度测序技术(如tsai等人“guide
‑
seq enables genome
‑
wide profiling of off
‑
target cleavage by crispr
‑
cas nucleases”,nature biotechnology 33,187
‑
197(2015)中描述的)。脱靶编辑位点可以具有与靶序列具有高序列同一性百分比的序列。序列同一性可以小于或等于99%、98%、97%、96%、95%、94%、93%、92%、91%、90%、89%、88%、87%、86%、85%、84%、83%、82%、81%、80%、79%、78%、77%、76%、75%、74%、73%、72%、71%、70%、69%、68%、67%、66%、65%、64%、63%、62%、61%、60%、59%、58%、57%、56%、55%、54%、53%、52%、51%、50%、49%、48%、47%、46%、45%、44%、43%、42%、41%、40%、39%、38%、37%、36%、35%、34%、33%、32%、31%或30%。脱靶编辑位点可以具有与pam区域非常接近的序列,例如,在前间区序列的5’末端(pam的远端)处可以容许引导rna和dna之间的错配,以产生脱靶编辑。本领域技术人员容易地理解如何确定两个核酸之间的序列同一性。例如,序列同一性可以在比对两个序列使得序列同一性处于其最高水平之后计算。另一种计算序列同一性的方法可以通过公布的算法来进行。用于比较的最佳序列比对可以通过以下来进行:通过smith和waterman adv.appl.math.2:482(1981)的局部同源性算法、通过needleman和wunsch,j.mol biol.48:443(1970)的同源性比对算法、通过pearson和lipman,proc.natl.acad.sci.u.s.a.85:2444(1988)的相似度检索方法、通过从美国国家生物技术信息中心(the national center for biotechnology information)(http://www.ncbi.nlm.nih.gov/blast/b12seq/b12.html)可用的这些算法的计算机化实施(wisconsin genetics软件包中的gap、bestfit、fasta和tfasta,genetics computer group,575science dr.,madison,wis.;tatusova和madden fems microbiol.lett.174:247
‑
250(1999)的blast算法)或通过目视检查。
[0276]
e.可裂解元件
[0277]
一个或更多个可裂解元件可以是本文描述的任何可裂解元件。
[0278]
i.可裂解元件的类型
[0279]
crispr多核苷酸的裂解特性可以通过可裂解元件来改变,所述可裂解元件可以在适当的条件下改变crispr多核苷酸中掺入该可裂解元件的点处的裂解倾向。“可裂解元件”可以包含天然核苷酸或者一个或更多个修饰的核苷酸。可裂解元件可以在核酸合成期间掺入crispr多核苷酸(例如,sgrna)中。
[0280]
crispr多核苷酸中的两个或更多个可裂解元件可以具有不同的裂解特性,例如,两个或更多个可裂解元件在被掺入crispr多核苷酸(例如,sgrna)中时,可以通过使用不同
的媒介和/或反应条件在彼此存在的情况下被选择性地裂解。
[0281]
如本文使用的,术语“裂解(cleaving)”、“裂解(cleaved)”和“裂解(cleavage)”都可以指crispr多核苷酸(例如,sgrna)基本上在crispr多核苷酸(例如,sgrna)中可裂解元件出现的每一点处的切割。
[0282]
裂解可以由媒介引发。媒介可以是例如引起可裂解元件裂解的化学实体或物理力。媒介可以是化学物质或化学物质的组合、生物分子或生物分子的组合、正常或相干(激光)可见光或紫外(uv)光、热或其他形式的电磁能。在一些情况下,可以同时或依次使用媒介的组合,例如两种或更多种媒介,来裂解crispr多核苷酸(例如,sgrna)。同时意指crispr多核苷酸(例如,sgrna)可以在同一时间暴露于两种或更多种媒介,尽管两种或更多种媒介可以一次一种地与crispr多核苷酸(例如,sgrna)反应。依次意指crispr多核苷酸(例如,sgrna)可以与一种媒介接触,并且然后在以后的时间与第二种媒介接触。
[0283]
crispr多核苷酸(例如,sgrna)可以包含多于一种类型的可裂解元件。在一些实例中,第一可裂解元件和第二可裂解元件具有相同的裂解特性。在一些实例中,第二可裂解元件具有与第一可裂解元件不同的裂解特性。例如,第一可裂解元件可以是可光裂解接头,并且第二可裂解元件由化学核酸酶引起的裂解敏感。在另一实例中,第一可裂解元件可以由化学核酸酶引起的裂解敏感,并且第二可裂解元件可以被工程化为可光裂解,从而允许应用正交处理方案。在一些情况下,相同的可裂解元件可以具有多于一种类型的裂解特性。第一可裂解元件和第二可裂解元件可以是本文描述的任何可裂解元件。
[0284]
可裂解元件(例如,可裂解接头)可以指能够连接crispr多核苷酸(例如,sgrna)的两种或更多种成分使得crispr多核苷酸(例如,sgrna)在适当条件下对裂解敏感的实体。例如,适当条件可以是暴露于uv光。可裂解接头可以包含一个或更多个在适当条件下对切割敏感的修饰或未修饰的核苷酸。
[0285]
可裂解接头可以包含修饰的核苷间连接。修饰的核苷间连接可以是具有磷原子的核苷酸间连接,也可以是没有磷原子的核苷酸间连接。其中包含磷原子的核苷间连接包括,例如,二硫代磷酸酯、磷酸三酯、氨基烷基磷酸三酯、甲基膦酸酯和其他烷基膦酸酯包括3
′‑
亚烷基膦酸酯、5
′‑
亚烷基膦酸酯和手性膦酸酯、亚膦酸酯、磷酰胺酯(包括3
′‑
氨基磷酰胺酯和氨基烷基磷酰胺酯)、p
‑
乙氧基磷酸二酯(p
‑
ethyoxyphosphodiester)、p
‑
乙氧基磷酸二酯(p
‑
ethoxyphosphodiester)、p
‑
烷氧基磷酸三酯、甲基膦酸酯、硫代磷酰胺酯(thionophosphoramidate)、硫代烷基膦酸酯(thionoalkylphosphonate)、硫代烷基磷酸三酯、硒代磷酸酯和硼磷酸酯,以及包含非磷的连接,例如,诸如本领域已知的缩醛和酰胺,具有正常3
’‑5’
连接、2
’‑5’
连接的类似物的那些以及具有反向极性的那些(其中一个或更多个核苷酸间连接是3’至3’、5’至5’或2’至2’连接)。具有反向极性的多核苷酸可以在最3
′
侧的核苷酸间连接处包含单个3
′
至3
′
连接,即,可以是无碱基的单个反向核苷残基(核酸碱基丢失或具有代替其的羟基基团)。
[0286]
包含非磷的核苷间连接包括短链烷基、环烷基、混合杂原子烷基、混合杂原子环烷基、一个或更多个短链杂原子和一个或更多个短链杂环。这些核苷间连接包括但不限于硅氧烷、硫化物、亚砜、砜、乙酰基、甲酰基、硫代甲酰基、亚甲基甲酰基、硫代甲酰基、烯基、氨基磺酸酯;亚甲基亚氨基、亚甲基肼基、磺酸酯、磺酰胺、酰胺和其他具有混合的n、o、s和ch2组分部件的核苷间连接。其中不包含磷原子的修饰的其他核苷间连接包括,
‑
ch2‑
nh
‑
o
‑
ch2‑
、
‑
ch2‑
n(ch3)
‑
o
‑
ch2‑
(称为亚甲基(甲基亚氨基)主链)、
‑
ch2‑
o
‑
n(ch3)
‑
ch2‑
、
‑
ch2‑
n(ch3)
‑
n(ch3)
‑
ch2‑
和
‑
o
‑
n(ch3)
‑
ch2‑
ch2‑
。
[0287]
可裂解接头本质上可以是非核苷酸的。“非核苷酸”可以指能够在一个或更多个核苷酸单元的位置中(包括糖取代和/或磷酸取代)掺入多核苷酸链的任何基团或化合物。基团或化合物可以是无碱基的,因为它不包含通常公认的核苷酸碱基,诸如腺嘌呤、鸟嘌呤、胞嘧啶、尿嘧啶或胸腺嘧啶,例如在糖的c1位置处。
[0288]
非核苷酸接头可以是例如无碱基残基(d间隔物),多甘醇,诸如三甘醇(间隔物9)或六甘醇(间隔物18),或烷烃二醇,诸如丁二醇。间隔物单元可以优选地通过磷酸二酯键或硫代磷酸酯键连接。接头单元可以仅在分子中出现一次,或者可以被掺入若干次,例如通过磷酸二酯、硫代磷酸酯、甲基膦酸酯或酰胺连接。其他优选的接头是烷基氨基接头,诸如c3、c6、c12氨基接头,以及烷基硫醇接头,诸如c3或c6硫醇接头。在一些实例中,异双官能和同双官能连接部分可以用于将肽和蛋白缀合至核苷酸。实例包括5
′‑
氨基
‑
修饰物c6和3
′‑
氨基
‑
修饰物c6试剂。
[0289]
ii.裂解可裂解元件的方法
[0290]
可裂解元件可以通过任何合适的方法裂解,包括暴露于酸、碱、亲核体、亲电体、自由基、金属、还原剂或氧化剂、光、温度、酶、小分子、核酸、蛋白等。在一些实例中,可裂解元件(例如,可裂解接头)对由细胞过程或其副产物引起的裂解敏感。细胞过程可以涉及酶、第二信使分子、代谢物、蛋白和自由基。
[0291]
iii.光不稳定基团
[0292]
可裂解元件可以是光不稳定基团。光不稳定基团可以通过亚磷酰胺化学被引入crispr多核苷酸中。如果使用光不稳定基团进行交联,则该光不稳定基团可以与光不稳定可裂解元件相同或不同。如果用于交联的光不稳定基团与用于裂解的光不稳定基团不同,则可以使用与用于激活交联的波长不同的波长来激活裂解。crispr多核苷酸中的两个或更多个光不稳定元件可以具有不同的激活特性,例如,两个或更多个元件在被掺入crispr多核苷酸中时,可以通过使用不同的媒介和/或反应条件在彼此存在的情况下被选择性地激活。
[0293]
pc
‑
氨基标签亚磷酰胺(pc
‑
aminotag phosphoramidite)与生长的寡核苷酸链的游离5
′‑
oh基团的选择性反应,随后从支持物上裂解和脱保护,可以导致磷酸二酯基团的引入,该磷酸二酯基团通过可光裂解接头连接至脂族伯氨基基团。然后,通过与胺反应试剂的合成后修饰反应,该氨基基团可以用于引入各种可光裂解标志物(olejnik j等人,nucleic acids research.1998;26:3572
‑
6)。例如,crispr多核苷酸可以包含连接crispr多核苷酸中两个核苷酸(例如,核苷酸53和核苷酸54)的可光裂解脂族基团,并且crispr多核苷酸可以暴露于uv光,导致crispr多核苷酸的断裂(例如,在核苷酸53和核苷酸54之间)。在其他实例中,可光裂解氨基标签亚磷酰胺可以位于crispr多核苷酸中的多核苷酸前导序列和引导序列之间,并且uv光可以用于在可光裂解氨基标签亚磷酰胺处引发裂解,从而使多核苷酸前导序列分离。可以用于引发crispr多核苷酸裂解的可光裂解接头的实例可以是3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺。例如,crispr多核苷酸可以包含连接crispr多核苷酸中两个核苷酸(例如,核苷酸53和核苷酸54)的可光裂解脂族基团,并且crispr多核苷酸可以暴露于可见光,导致crispr多核
苷酸的断裂(例如,在核苷酸53和核苷酸54之间)。在其他实例中,可光裂解香豆素光接头可以位于crispr多核苷酸中的多核苷酸前导序列和引导序列之间,并且可见光可以用于在可光裂解香豆素光接头处引发裂解,从而使多核苷酸前导序列分离。可以用于引发crispr多核苷酸裂解的可光裂解接头的实例可以是香豆素接头。其他将可光裂解接头引入多核苷酸序列中的方法已描述于以下中:例如,美国专利申请:us20080227742a1、us20100022761a1、us7897737b2,这些申请的内容在此以其整体引用。
[0294]
iv.基于核糖核酸酶的裂解
[0295]
在一些实例中,一个或更多个可裂解元件包含内切核糖核酸酶的裂解位点,例如,在限定的核糖核苷酸序列基序处或限定的核糖核苷酸序列基序内裂解rna的内切核糖核酸酶。例如,可裂解元件可以包含由序列特异性内切核糖核酸酶识别的裂解位点。内切核糖核酸酶可以是天然存在的或工程化的。在一些实例中,内切核糖核酸酶可以对单链rna、双链rna或由dna:rna杂交体形成的核苷酸序列具有特异性。在一些实例中,内切核糖核酸酶的序列特异性可以通过与寡核苷酸融合或通过与其他蛋白结构域融合来工程化。例如,序列特异性内切核糖核酸酶酶可以通过融合两个功能上独立的结构域(以进行性和序列独立的方式水解dna
‑
rna杂交体中的rna的rna酶hi,和识别dna
‑
rna杂交体中序列的锌指)来工程化。在另一种情况下,反义寡脱氧核苷酸与核糖核酸酶h的缀合可以导致序列特异性裂解。参见,例如,sulej等人,nucleic acids research.2012;40(22):11563
‑
70和fukuma等人,bioconjugate chemistry.2003;14(2):295
‑
301。在一些情况下,可裂解元件可以能够募集rna酶h1来裂解crispr多核苷酸的双链区域。(参见,例如,美国专利第5,849,902号)。
[0296]
可裂解元件可以包含由序列特异性ssrna内切核糖核酸酶识别的裂解位点,诸如,例如zaug等人,biochemistry 1988;27,25,8924
‑
8931中描述的嗜热四膜虫(tetrahymena thermophila)的切离的ivs rrna部分。在其他实例中,可裂解元件可以包含一个或更多个由序列特异性ssrna内切核糖核酸酶cas2识别的裂解位点,如描述于以下中的:例如,beloglazova等人,j biol chem.2008;283(29):20361
‑
20371。在其他实例中,可裂解元件可以包含一个或更多个优选的由来自枯草芽孢杆菌(bacillus subtilis)的rna酶mini
‑
iii识别的dsrna位点,例如glow等人,nucleic acids res.2015;43(5)2864
‑
73中讨论的。在其他实例中,短寡核苷酸可以用作外部引导序列(egs)来引导人类rna酶p对crispr多核苷酸的位点特异性裂解。例如,靶向乙型肝炎病毒(hbv)的2.1kb表面抗原mrna的13
‑
mer egs能够诱导rna酶p对hbv rna的裂解(参见werner m等人,rna.1998;4(7):847
‑
55)。内切核糖核酸酶可以是序列或结构特异性内切核糖核酸酶cas6超家族的成员,例如cas6a(例如hong li(2015),structure,january 6;23(1):13
‑
20)。内切核糖核酸酶可以是cs4,也称为cas6f。ssrna内切核糖核酸酶可以属于crispr内切核糖核酸酶的cas13家族或其衍生物。内切核糖核酸酶可以是能够加工前crerna转录物的cpfl或cas5d酶(zetsche,b.等人(2016),“multiplex gene editing by crispr
‑
cpf1 using a single crrna array”,nature biotechnology(2016)doi:10.1038/nbt.3737)。
[0297]
可裂解元件可以是可被核酶(例如锤头状核酶、丁型肝炎病毒核酶等)裂解的元件。核酶可以是天然存在的,或者可以通过分离为“催化剂”和“底物”链而被工程化为反式作用性核酶,例如levy等人,rna 2005.11:1555
‑
1562中讨论的。在一些情况下,两种核酶可以共同使用,以允许在期望的靶序列之后裂解。在一些情况下,在不同细胞区室中发挥功能
的替代的人工核酶
‑
蛋白复合物可以通过使用定位决定簇用于将核酶递送至特定亚细胞位点或靶向特定类型的rna来设计,如samarsky等人,proc natl acad sci u s a.1999;96(12):6609
‑
6614中示出的。在一些情况下,核酶的使用可以包括结合外源小分子以获得活性,例如,glms核酶。
[0298]
在一些实例中,核酶的活性可以通过与适配体偶联而进一步调节为配体控制的。适配体可以基于其结合配体的能力或者另外以通过信息传递结构域直接偶联至环i和/或环ii的方式“感知”环境变化(诸如ph、温度、摩尔渗透压浓度、盐浓度等)的能力来选择。配体可以是,例如,蛋白、核苷酸或小分子配体。配体与适配体的结合可引起信息传递结构域与环、茎或催化核心中的一个或更多个的相互作用的变化,使得核酶活性可以根据配体的存在或不存在而被调节,例如在us8603996b2中描述的。
[0299]
crispr多核苷酸(例如,sgrna)的可裂解元件的裂解可以在期望的时间独立诱导;例如,遗传编码的内切核糖核酸酶可以在宿主细胞内被激活。编码内切核糖核酸酶的载体或质粒可以在期望的时间转染到细胞中。一种或更多种内切核糖核酸酶可以处于一种或更多种独立的启动子的控制下。一种或更多种启动子可以在期望的时间被激活。
[0300]
v.反义寡核苷酸
[0301]
crispr多核苷酸的一个或更多个可裂解元件可以被设计为允许反义寡核苷酸的结合。反义寡核苷酸可以是单链dna(ssdna)寡核苷酸。ssdna寡核苷酸可以与crispr多核苷酸中的单链rna序列杂交,并且rna酶h可以用于裂解dna:rna杂交体的rna。对于反义寡核苷酸可以结合的可裂解元件(例如,crispr多核苷酸中茎环的rna环),可裂解元件(例如,茎环的环)的长度可以是约6个至约40个核苷酸。反义寡核苷酸可以是约12个至约16个核苷酸的长度、或约12个至约25个核苷酸或约10个至约30个核苷酸的长度。反义寡核苷酸和crispr多核苷酸的可裂解元件(例如,茎环的环)之间的互补性程度可以是至少80%、85%、90%、95%、98%、99%或100%。其序列与可裂解元件完全互补或部分互补的反义寡核苷酸可以在宿主细胞内产生或被引入宿主细胞中。反义寡核苷酸可以使用聚乙烯亚胺(pei)或其他已知的转染方法转染到细胞中。
[0302]
crispr多核苷酸的一个或更多个可裂解元件可以包含mirna响应元件。mirna响应元件的长度可以在约15个至约30个核苷酸之间,例如约20个至约25个核苷酸的长度。mirna的长度可以是约20个至约24个核苷酸,例如,约21个至约23个核苷酸,例如,约22个核苷酸的长度。mirna和crispr多核苷酸中的mirna响应元件之间的序列互补性程度可以是至少80%、85%、90%、95%、98%、99%或100%。
[0303]
可裂解元件可以包含mirna响应元件(mre),并且能够结合mre的mirna可以在宿主细胞内产生或被引入宿主细胞中。mirna可以以mirisc复合物的形式存在,该mirisc复合物可以靶向mre并且使第一可裂解元件裂解。
[0304]
vi.位点特异性化学核酸酶
[0305]
crispr多核苷酸的特异性裂解可以通过被设计为具有位点特异性核酸酶活性的化合物来实现。
[0306]
化学核酸酶可以被设计为对本文描述的crispr多核苷酸(例如,crispr on多核苷酸、crispr off多核苷酸或crispr on/off多核苷酸)具有序列特异性亲和力。例如,裂解rna的三(2
‑
氨基苯并咪唑)可以通过二硫键或酰胺键附接至dna寡核苷酸或2
’‑
o
‑
甲基寡核
糖核苷酸,以形成显示出rna底物和位点选择性的有机催化核酸酶(参见,例如,gnaccarini等人,j.am.chem.soc.,2006,128(24),第8063
‑
8067页)。在其他实例中,化学rna酶(例如,1,10
‑
菲咯啉部分、新亚铜(neocuprine)zn(ii)、新霉胺)对crispr多核苷酸的位点特异性可以通过使用肽核酸(pna)例如聚酰胺核酸来实现。
[0307]
化学rna酶(例如,二亚乙基三胺部分)对crispr多核苷酸的位点特异性可以通过组合使用反义寡核苷酸、肽蛋白或pna来实现。在一些实例中,rna结合蛋白可以通过共价附接至配位络合物,诸如1,10
‑
菲咯啉
‑
铜络合物,被化学转化为序列特异性核酸酶。参见,例如,chen等人,sigman ds.science.1987;237(4819):1197
‑
201。在另一种实例中,crispr多核苷酸的位点特异性裂解可以通过博莱霉素
‑
fe(ii)与edta或寡核苷酸的缀合形成对crispr多核苷酸具有特异性的人工核酸酶来实现。
[0308]
化学核酸酶的实例包括1,10
‑
菲咯啉铜(sigman等人,1993)、亚铁
‑
乙二胺四乙酸(edta)、大环镧系络合物、金属卟啉、salen的金属络合物、乙酸铀酰、铑(iii)的八面体金属络合物、四氟硼酸重氮苯(benzene diazonium teetrafluoroborate)、脂族单胺、脂族二胺和脂族多胺、氨基糖苷诸如新霉素b和铜(ii)氨基糖苷络合物等。在一些情况下,化学核酸酶可以靶向核苷的糖部分,并且通过在裂解位点处从糖中夺取氢原子来催化氧化裂解。
[0309]
vii.光化学裂解
[0310]
在一些实例中,光笼基团(photocaging group)可以用于提供对用于裂解crispr多核苷酸的媒介的活性的进一步控制。例如,可光激活或“笼状(caged)”探针的光解可以用于控制本公开内容中描述的位点特异性化学核酸酶的释放。在另一种实例中,光笼基团可以用于阻断核糖核酸酶或限制性内切酶对crispr多核苷酸的裂解,直到通过光解释放,例如,如bohacova等人,biomol.chem.,2018.16,1527中示出的。在另一种实例中,crispr多核苷酸中一个或更多个核苷酸上的光笼基团可以用于掩蔽反义核苷酸的识别序列,直到通过光解释放,从而引发crispr多核苷酸的裂解。在另一种实例中,光笼基团可以附接至裂解媒介诸如反义寡核苷酸,所述裂解媒介在光解时变得可用于与crispr多核苷酸结合并引发risc复合物的形成。在另一种实例中,光笼基团可以用于掩蔽用于裂解crispr多核苷酸的
‘
mirna响应元件’,直到通过光解释放。在其他方面,但不限于这些方面,光笼基团可以以正交处理方案使用,用于裂解具有不同裂解特性的多于一个裂解元件。
[0311]
光笼基团可以用于对裂解反应“加标签”,其中标签可以适合通过一种或更多种方法进行检测和/或定量。例如,基于2
‑
硝基苄基的可光裂解基团可以进一步用染料标记,所述染料在光解时释放,并可用作crispr on多核苷酸激活的“效率”或crispr off多核苷酸失活的“效率”等的可检测标志物。在另一种实例中,与crispr多核苷酸的可裂解元件结合的核糖核酸酶蛋白可以在“裂解事件”开始后通过从掺入可裂解元件的光笼核苷酸中释放“荧光标签”而被加标签(tagged),其中荧光标签的测量可以作为crispr多核苷酸裂解的替代标志物。
[0312]
可以合成掺入crispr多核苷酸的光笼基团的实例包括基于邻硝基苄基的笼基团,所述基于邻硝基苄基的笼基团可以通过本领域已知的方法以醚、硫醚、酯(包括磷酸酯或硫代磷酸酯)、胺或类似官能团的方式连接至杂原子(通常是o、s或n)。基于2
‑
硝基苄基的笼基团的实例包括α
‑
羧基
‑2‑
硝基苄基、1
‑
(2
‑
硝基苯基)乙基、4,5
‑
二甲氧基
‑2‑
硝基苄基、1
‑
(4,5
‑
二甲氧基
‑2‑
硝基苯基)乙基、5
‑
羧基甲氧基
‑2‑
硝基苄基、硝基苯基等。其他可光去除
的保护基团的实例包括苄氧羰基、3
‑
硝基苯基、苯酰基、3,5
‑
二甲氧基苯甲酰基、2,4
‑
二硝基苯磺酰基、单叠氮乙锭、bimane叠氮化物(bimane azide)和它们各自的衍生物。
[0313]
本文描述的光不稳定接头可以表示为若干种内消旋形式。在绘制单个结构时,预期了任何相关的内消旋形式。本文描述的由结构式表示的香豆素接头可以以任何相关的内消旋形式示出。以下示出式(i)的示例性内消旋结构:
[0314][0315]
光不稳定保护基团可以附接至核苷和核苷酸中的羟基和磷酸或核酸碱基。例如,2
′‑
脱氧
‑5‑
(羟甲基)尿苷核苷的光笼衍生物(被2
‑
硝基苄基
‑
、6
‑
硝基胡椒基
‑
和蒽基
‑9‑
甲基基团保护的单磷酸酯和三磷酸酯)可以被酶促掺入多核苷酸中,例如,如bohacova等人,org.biomol.chem.,2018,16,152中描述的。光裂解可以通过各种机制发生,诸如从糖环中夺取氢键,从碱基直接电子转移至光激发的裂解物,或者通过来自光裂解的能量转移和加合物的形成产生单线态氧。
[0316]
viii.可裂解元件的裂解
[0317]
两种或更多种(例如,2种、3种、4种、5种、6种、7种、8种、9种或10种)crispr多核苷酸的可裂解元件可以被相同的裂解部分裂解。两种或更多种(例如,2种、3种、4种、5种、6种、7种、8种、9种或10种)不同的crispr多核苷酸的裂解可以由不同的外部因素诱导。
[0318]
裂解诱导媒介可以是电磁辐射。裂解诱导媒介可以是可见光谱中特定波长的光。裂解元件可以被uv光裂解。
[0319]
光的波长范围可以为220nm
‑
465nm。暴露方案中的光强可以是约5mw/cm2、10mw/cm2、15mw/cm2、20mw/cm2、25mw/cm2、35mw/cm2、40mw/cm2、50mw/cm2、70mw/cm2、90mw/cm2、110mw/cm2、120mw/cm2、140mw/cm2、160mw/cm2、175mw/cm2、190mw/cm2、200mw/cm2、220mw/cm2、240mw/cm2、260mw/cm2、280mw/cm2、300mw/cm2、320mw/cm2、340mw/cm2、360mw/cm2、380mw/cm2、400mw/cm2、420mw/cm2、440mw/cm2、460mw/cm2、480mw/cm2、500mw/cm2、520mw/cm2、540mw/cm2、560mw/cm2、580mw/cm2、600mw/cm2、620mw/cm2、650mw/cm2、675mw/cm2、700mw/cm2、720mw/cm2、745mw/cm2、765mw/cm2、790mw/cm2、810mw/cm2、830mw/cm2、850mw/cm2、870mw/cm2、900mw/cm2、920mw/cm2、945mw/cm2、965mw/cm2、985mw/cm2、1000mw/cm2、1025mw/cm2、1050mw/cm2、1080mw/cm2、1100mw/cm2、1125mw/cm2、1150mw/cm2、1175mw/cm2、1200mw/cm2、1240mw/cm2、
1275mw/cm2、1290mw/cm2、1320mw/cm2、1350mw/cm2、1380mw/cm2、1400mw/cm2、1420mw/cm2、1450mw/cm2、1470mw/cm2、1490mw/cm2、1520mw/cm2、1540mw/cm2、1560mw/cm2、1600mw/cm2、1630mw/cm2、1650mw/cm2、1670mw/cm2、1700mw/cm2、1720mw/cm2或1750mw/cm2。暴露方案中的光强度范围可以为70mw/cm2至100mw/cm2、80mw/cm2至110mw/cm2、90mw/cm2至120mw/cm2、100mw/cm2至130mw/cm2、110mw/cm2至140mw/cm2、120mw/cm2至150mw/cm2、130mw/cm2至160mw/cm2、140mw/cm2至170mw/cm2、150mw/cm2至180mw/cm2、160mw/cm2至190mw/cm2、170mw/cm2至200mw/cm2、180mw/cm2至210mw/cm2、190mw/cm2至220mw/cm2、200mw/cm2至230mw/cm2、210mw/cm2至240mw/cm2、220mw/cm2至250mw/cm2、230mw/cm2至260mw/cm2、240mw/cm2至270mw/cm2、250mw/cm2至280mw/cm2、260mw/cm2至290mw/cm2或270mw/cm2至300mw/cm2。光的波长范围可以为约320nm至约390nm。光的波长范围可以为约320nm至425nm、320nm至420nm、420nm至520nm、520nm至620nm、420nm至700nm。光的波长可以大于约320nm、330nm、340nm、350nm、360nm、370nm、380nm、390nm、400nm、410nm、420nm、430nm、440nm、450nm、460nm、470nm、480nm、490nm、500nm、510nm、520nm、530nm、540nm、550nm、560nm、570nm、580nm、590nm、600nm、610nm、620nm、630nm、640nm、650nm、660nm、670nm、680nm、690nm或700nm。光的波长可以小于约700nm、690nm、680nm、670nm、660nm、650nm、640nm、630nm、620nm、610nm、600nm、590nm、580nm、570nm、560nm、550nm、540nm、530nm、520nm、510nm、500nm、490nm、480nm、470nm、460nm、450nm、440nm、430nm或425nm。光的波长范围可以为420nm至430nm、430nm至440nm、440nm至450nm、450nm至460nm、460nm,至470nm、470nm至480nm、480nm至490nm、490nm至500nm、500nm至510nm、510nm至520nm、520nm至530nm、530nm至540nm、540nm至550nm、550nm至560nm、560nm至570nm、570nm至580nm、580nm至590nm、590nm至600nm、600nm至610nm、610nm至620nm、620nm至630nm、630nm至640nm、640nm至650nm、650nm至660nm、660nm至670nm、670nm至680nm、680nm至690nm或690nm至700nm。暴露方案中使用的光的功率瓦数可以是如通过oai 306uv功率计测量的约50w、70w、80w、90w、100w、120w、140w、160w、175w、190w、210w、230w、250w、270w、290w、310w、330w、250w、370w、390w、420w、450w、480w、500w、530w、550w、570w、600w、620w、650w、670w、700w、720w、750w、770w、800w、820w、850w、870w、900w、920w、950w、970w、1000w、1020w、1050w、1070w、1100w、1120w、1200w、1300w、1400w、1500w、1600w、1700w、1800w、1900w、2000w、2100w、2200w、2300w、2400w、2500w、2600w、2700w、2800w、2900w、3000w、3100w、3200w、3300w、3400w、3500w、3600w、3700w、3800w、3900w、4000w、4100w、4200w、4300w、4400 4500w、4600w、4700w、4800w、4900w、5000w、5100w、5200w、5300w、5400w、5500w、5600w、5700w、5800w、5900w或6000w。
[0320]
暴露的持续时间可以是1秒至30分钟。暴露的持续时间可以是1秒至30秒、30秒至60秒、1min至5min、5min至10min、10min至20min、20min至30min、30min至40min、40min至50min或50min至1hr。暴露的持续时间可以大于约1小时、50min、40min、30min、20min、10min、5min、1min、30秒或1秒。暴露的持续时间可以小于约2秒、30秒、1min、5min、10min、20min、30min、40min、50min或1小时。暴露方案可以包括连续暴露或脉冲暴露或两者。脉冲暴露可以是均匀的或具有不同的持续时间。
[0321]
光源可以是通过带通滤光器过滤的宽谱光。带通滤光器可以是345nm带通滤光器。带通滤光器可以是420nm长通滤光器。光源可以是紫外(uv)光。光源可以是led。led可以发射紫外光。led可以发射可见光。led可以发射红外光。
[0322]
b.用于锁定的修饰的crispr效应蛋白
[0323]
在一些情况下,修饰crispr效应蛋白以促进与crispr多核苷酸的锁定。可以修饰crispr多核苷酸以促进与crispr效应蛋白的锁定。crispr多核苷酸可以包括本文描述的crispr on多核苷酸序列、crispr off多核苷酸序列、crispr on/off多核苷酸序列或当与crispr酶复合时具有降低的脱靶编辑活性的包含一种或更多种修饰的crispr多核苷酸。可以修饰crispr效应蛋白和crispr多核苷酸两者,以促进与crispr效应蛋白的锁定。例如,crispr效应蛋白可以用非天然氨基酸来修饰,以促进与crispr多核苷酸的交联。在一些情况下,修饰crispr多核苷酸(例如,sgrna)和crispr效应蛋白两者以促进锁定。在一些情况下,仅crispr效应蛋白包含交联剂,而crispr多核苷酸不包含交联剂。在一些情况下,crispr多核苷酸包含交联剂,而crispr效应蛋白不包含交联剂。
[0324]
如下文描述的,crispr效应蛋白可以通过包含一个或更多个非天然氨基酸或者通过crispr效应蛋白与被设计为促进与另一种分子的结合的氨基酸序列的融合(诸如snap融合蛋白)(例如,融合体的表达)来修饰。
[0325]
crispr效应蛋白,例如,cas9,可以包含一个或更多个突变(并且因此编码该crispr效应蛋白的一个或更多个核酸分子可以具有一个或更多个突变)。一个或更多个突变可以是人工引入的突变,并且可以是催化结构域中的一个或更多个突变。关于cas9酶的催化结构域的实例可以是ruvc i、ruvc ii、ruvc iii和hnh结构域。一个或更多个突变可以使cas9的一个或更多个催化结构域失活。一个或更多个突变可以使cas9的催化活性降低0.1倍、0.25倍、0.5倍、0.75倍、1倍、2倍、5倍、10倍、50倍、100倍或1000倍。在一些情况下,一个或更多个突变可以使cas9的催化活性增加0.1倍、0.25倍、0.5倍、0.75倍、1倍、2倍、5倍、10倍、50倍、100倍或1000倍。
[0326]
crispr多核苷酸可以用穿透细胞的rna适配体修饰。穿透细胞的rna适配体可以改进crispr多核苷酸向细胞的有效递送。rna适配体可以与细胞表面受体结合,并且促进crispr多核苷酸进入细胞中。穿透细胞的适配体可以被设计成靶向特定的细胞受体,以便介导细胞特异性递送。
[0327]
1.用于锁定的修饰的类型
[0328]
crispr效应蛋白可以通过包含非天然氨基酸来修饰。非天然氨基酸包括光不稳定非天然氨基酸,例如,光不稳定非天然氨基酸交联剂,例如,对叠氮基
‑
l
‑
苯丙氨酸(azf)或对苯甲酰基
‑
l
‑
苯丙氨酸(bzf),可以用于在生物缀合中形成交联。在350
‑
360nm激发时,二苯甲酮,例如,bzf,可以优先与暴露的位于crispr多核苷酸上的官能团上的本来无活性的碳氢键反应。在一些情况下,二苯甲酮不发生光解离,并且二苯甲酮的光激发的三线态在没有合适的碳氢键与之反应的情况下容易弛豫((relax)),因此二苯甲酮可以是比其他交联试剂更宽容的试剂。azf在暴露于紫外光时可以产生可用于与crispr多核苷酸连接的反应性氮烯(nitrene)。
[0329]
crispr效应蛋白可以与另一种蛋白(例如,snap蛋白)融合。配置可以包括通过bg(o6
‑
苄基鸟嘌呤)接头将dna修复模板共价附接至snap蛋白作为保持dna修复模板靠近crispr复合物的方法,和/或通过使用bg接头和rna适配体将crispr多核苷酸(例如,sgrna)与接头核苷酸序列附接以附接至snap蛋白,如下文描述的。
[0330]
crispr效应蛋白可以用snap蛋白融合体修饰,以促进crispr多核苷酸(例如,
sgrna)与crispr效应蛋白的连接。例如,可以使用载体表达包含crispr效应蛋白和snap蛋白以及臂区域的融合蛋白。臂区域还包含一系列为了柔性而配置的氨基酸。臂区域还可以被配置为与苄基鸟嘌呤修饰的多核苷酸连接。snap蛋白区域可以位于crispr效应蛋白的n末端处。臂区域可以位于snap蛋白的n末端处。crispr多核苷酸可以用结合苄基鸟嘌呤的rna适配体修饰(如由以下描述的:例如,carrocci和hoskins,evolution and characterization of a benzylguanine
‑
binding rna aptamer,chem commun(camb).2016january 11;52(3):549
‑
552.doi:10.1039/c5cc07605f)。在crispr多核苷酸与结合苄基鸟嘌呤的rna适配体结后,crispr多核苷酸可以与融合蛋白的臂区域共价键合。臂区域的柔性可以允许crispr多核苷酸与融合蛋白的crispr效应蛋白区域复合。可选地,crispr多核苷酸可以在附接至融合蛋白的臂区域之前复合。
[0331]
c.具有高亲和力结合的crispr效应蛋白和多核苷酸的crispr复合物
[0332]
本文提供了一种crispr复合物,所述crispr复合物包含(a)包含以下的多核苷酸:被设计为与靶核酸序列退火的序列和被设计为结合crispr效应蛋白的序列,以及活性调节性多核苷酸序列(例如本文描述的crispr on、crispr off或crispr on/off);和(b)crispr效应蛋白,其中与crispr效应蛋白结合的多核苷酸的平衡解离常数(kd)小于8pm(pm=皮摩尔/升)。平衡解离常数(kd)可以小于7pm、5pm、4pm、3pm、2pm、1pm、9fm(fm=飞摩尔/升)、8fm、7fm、6fm、5fm、4fm、3fm、2fm、1fm、9am(am=渺摩尔/升(attomolar))、8am、7am、6am、5am、4am、3am、2am或1am。平衡解离常数(kd)可以是约1pm至8pm、约1fm至约10fm或约1am至约10am。crispr效应蛋白可以与crispr多核苷酸共价附接。在一些情况下,crispr效应蛋白与crispr多核苷酸不共价附接。
[0333]
iv.使用稳定的crispr复合物的方法
[0334]
本文提供了使用本文描述的稳定的(例如,锁定的)crispr复合物的方法。
[0335]
a.向细胞施用
[0336]
本发明公开内容的方面包括用于向细胞施用稳定的(例如,锁定的)crispr复合物的方法。稳定的crispr复合物可以包含crispr多核苷酸(包含与crispr效应蛋白交联的非天然核苷酸)和活性调节性序列(例如,本文描述的crispr on、crispr off或crispr on/off)。该方法可以包括使细胞与稳定的(例如,锁定的)crispr复合物的溶液接触。可选地或组合地,该方法可以包括使细胞与包含crispr效应蛋白编码区和/或crispr多核苷酸编码区的载体接触。可选地或组合地,可以使用非病毒介导的技术将crispr多核苷酸引入细胞中。非病毒介导的技术可以包括电穿孔、磷酸钙介导的转移、核转染、声穿孔、热激(heat shock)、磁转染、脂质体介导的转移、显微注射、微弹(microprojectile)介导的转移、纳米颗粒、阳离子聚合物介导的转移(例如,deae
‑
葡聚糖、聚乙烯亚胺、peg、dmso等)或细胞融合。
[0337]
可以使用病毒和非病毒介导的技术将crispr多核苷酸引入细胞中。非病毒介导的技术可以是电穿孔、磷酸钙介导的转移、核转染、声穿孔、热激、磁转染、脂质体介导的转移、显微注射、微弹介导的转移(纳米颗粒)、阳离子聚合物介导的转移(deae
‑
葡聚糖、聚乙烯亚胺、聚乙二醇(peg)等)或细胞融合。
[0338]
用至少一种用于交联的非天然核苷酸修饰的多核苷酸(包括本文描述的crispr on多核苷酸序列、crispr off多核苷酸序列或crispr on/off多核苷酸序列)和相关载体可
以裸递送至细胞(即不含促进转染的剂)。裸crispr多核苷酸可以使用本领域已知的和本文描述的施用途径递送至细胞。
[0339]
在一些情况下,在宿主细胞中靶dna中靶基因编辑的可调的调节包括以下步骤:(i)使用本文描述的本领域已知的病毒或非病毒递送方法或其组合,将以下引入宿主细胞中:(a)crispr多核苷酸,所述crispr多核苷酸包含将crispr多核苷酸与crispr效应蛋白交联的非天然核苷酸以及第一和第二裂解元件,其中裂解元件对裂解敏感,并且其中引导序列的核苷酸序列与靶核酸序列完全或部分互补,其中第一裂解元件位于多核苷酸前导序列和引导序列的5’末端之间;和(b)具有催化活性的crispr效应蛋白(例如,crispr酶,例如,cas9),使得crispr多核苷酸和crispr酶形成crispr复合物;并且(ii)通过暴露于uv光,诱导多核苷酸中第一序列元件的裂解,从而释放多核苷酸前导序列,并且激活crispr复合物对靶基因的更高的靶特异性裂解。随后,该方法可以包括(iii)通过脉冲暴露于uv光,在期望的时间诱导可以位于crispr多核苷酸的支架序列中的第二序列元件的裂解,从而使crispr多核苷酸裂解,并且使crispr复合物对靶基因的靶特异性裂解失活或降低。
[0340]
细胞可以是外胚层(例如,神经元和成纤维细胞)、中胚层(例如,心肌细胞)、内胚层(例如,胰腺细胞)、上皮(例如,肺道和鼻道)、中性粒细胞、嗜酸性粒细胞、嗜碱性粒细胞、淋巴细胞、破骨细胞、内皮细胞、造血细胞、红细胞等。细胞可以来源于特定细胞系,诸如cho细胞(例如,chokl)、hek293细胞、caco2细胞、u2
‑
os细胞、nih 3t3细胞、nso细胞、sp2细胞、dg44细胞、k
‑
562细胞,u
‑
937细胞、mc5细胞、imr90细胞、jurkat细胞、hepg2细胞、hela细胞、ht
‑
1080细胞、hct
‑
116细胞、hu
‑
h7细胞、huvec细胞和molt 4细胞。适用于本公开内容范围的其他细胞的实例可以包括干细胞、胚胎干细胞(esc)和诱导性多能干细胞(ipsc)、msc
‑
1、k562等。
[0341]
在一些情况下,可以创建掩蔽物来遮蔽细胞培养物。可以使用各种技术(激光切割、3d打印、光刻法等)来创建掩蔽物。掩蔽物可以被设计为让光穿透限定的区域。当与包含可光裂解接头的crispr off复合物结合使用时,光(例如,uv光或可见光)穿透区域中的编辑可以降低,并且未暴露于光的区域中的编辑可以维持。当与crispr on复合物结合使用时,在光(例如,uv光)穿透的区域中可以引发编辑。
[0342]
在一些情况下,crispr off复合物活性可以是时间依赖性的(例如,如实施例6、图30
‑
图32中可见的)。细胞可以在完成编辑之前的时间点暴露于裂解激活因素,诸如uv光或可见光,从而产生杂合克隆。可选地,这样的方法可以用于靶向患者来源的细胞系的病态等位基因。
[0343]
具有接近于零的解离常数的稳定的(例如,锁定的)crispr复合物可以比目前的复合物具有增加的功效,在目前的复合物中,crispr多核苷酸可以在向细胞施用之前或期间与crispr效应蛋白解离。
[0344]
1.多于一种crispr复合物
[0345]
在一些情况下,系统包含本文提供的一种或更多种crispr复合物。第一和第二(或更多)crispr复合物可以用于体外或体内方法。第一和第二crispr复合物中的crispr效应蛋白可以相同或不同。在一种实例中,体外或体内系统可以包含多于一种具有不同引导序列的crispr多核苷酸和相同的crispr效应蛋白(例如,cas9)。在另一种实例中,体外或体内系统可以包含crispr多核苷酸和多于一种不同的crispr效应蛋白(例如,催化活性和催化
无活性的crispr效应蛋白的混合物)。
[0346]
可选地或组合地,宿主细胞可以包含两种或更多种(例如,2种、3种、4种、5种、6种、7种、8种、9种或10种)不同的锁定的crispr复合物,其中不同crispr复合物的引导序列的核苷酸序列独立地与两种或更多种不同靶核酸(例如,dna)的区域完全或部分互补。不同的crispr复合物可以具有不同相对位置的一个或更多个裂解元件,或者相同相对位置的一个或更多个裂解元件。
[0347]
b.裂解靶核酸
[0348]
本发明公开内容的方面包括用于裂解靶核酸的方法。该方法可以包括使核酸序列与稳定的(例如,锁定的)crispr复合物接触。稳定的crispr复合物可以包含crispr多核苷酸(包含与crispr效应蛋白交联的非天然核苷酸)和活性调节性序列(例如,本文描述的crispr on、crispr off或crispr on/off)。例如,具有被选择以便不干扰cas核酸酶的核酸酶活性或crrna区的结合效率的交联位点的锁定的crispr复合物可以在裂解靶核酸方面具有增强的功效。多核苷酸可以包含“前间区”和“前间区邻近基序”(pam),并且这两个结构域都可能是crispr效应蛋白介导的活性(例如,裂解)所需的。
[0349]
crispr效应蛋白的每个催化结构域可以是可选地有活性或组合地有活性的。每个催化域的效率可以是50%至60%、60%至70%、70%至80%、80%至90%或90%至99.9%。
[0350]
裂解之后,dna修复可以通过非同源末端连接(nhej)、微同源介导的末端连接(mmej,替代性非同源末端连接)或同源定向修复(hdr)发生。可以为hdr提供dna模板。
[0351]
在一些实例中,裂解可以通过非同源末端连接(nhej)过程导致插入和/或缺失(“插入/缺失(indel)”)突变或移码,从而导致靶基因特异性敲除(ko)。在一些情况下,crispr/cas复合物通过特定grna(例如,sgrna)可以与共施用的供体多核苷酸(单链或双链)一起被引导至靶基因组区域。靶区域裂解后,同源定向修复(hdr)过程可以使用一种或更多种供体多核苷酸作为一种或更多种模板,用于(a)修复裂解的靶核苷酸序列和(b)将遗传信息从供体多核苷酸转移至靶dna。取决于遗传信息的性质,hdr过程可以产生靶基因特异性的ko或敲入(ki)。hdr介导的基因ki的应用实例包括添加(插入或替代)编码蛋白、mrna、小干扰rna(sirna)、标签(例如,6xhis)、报告物蛋白(例如,绿色荧光蛋白(gfp))的核酸物质,以及基因的调控序列(例如,启动子、聚腺苷酸化信号)。
[0352]
对于hdr过程,供体多核苷酸可以包含期望的编辑(例如,待拷贝的基因编辑(序列)),以及另外的与紧接裂解的靶位点的上游和下游同源的两端的核苷酸序列(同源臂)。在一些情况下,hdr过程的效率可以取决于基因编辑的尺寸和/或同源臂的尺寸。
[0353]
可以提供一种或更多种crispr复合物来靶向一个或更多个裂解位点。例如,可以提供两种crispr复合物来靶向两个裂解位点,可以提供10种crispr复合物来靶向10个裂解位点,可以提供20种crispr复合物来靶向20个裂解位点,等等。可以向细胞提供的不同crispr多核苷酸(例如,sgrna)的数量可以是约1种、2种、3种、4种、5种、6种、7种、8种、9种、10种、11种、12种、13种、14种、15种、25种、50种、100种或1000种,或者1种至3种、1种至5种、1种至10种、10种至50种或50种至100种。
[0354]
本文描述的crispr复合物可以在细胞中诱导一个或更多个编辑或突变。一个或更多个编辑或突变可以包括通过crispr多核苷酸(例如,引导rna或sgrna)在细胞的每个靶序列处引入、缺失或取代一个或更多个核苷酸。一个或更多个编辑或突变可以是在所述细胞
的每个靶序列处引入、缺失或取代约1个至约75个核苷酸。一个或更多个编辑或突变可以是在所述细胞的每个靶序列处引入、缺失或取代1个、5个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个、20个、21个、22个、23个、24个、25个、26个、27个、28个、29个、30个、35个、40个、45个、50个或75个核苷酸。靶序列可以是基因,并且可以包括bub1b、camk1、prkag3、stk3、camk1、chr8q23、cel、irak4、dnmt1、emx1、fancf、grk1、prgn、aavs1、bub1b、cxcr4、fam163a、gaa、crk1、irak4、mapre1、mip、omp、opn1sw、prkag3、stk3和vegfa(如在实施例7、实施例8、实施例11、实施例12中可见的)。
[0355]
多于一个靶位点可以被附接至不同sgrna的crispr复合物的集合靶向。集合中的每个grna可以与距引导rna集合中至少一个其他引导rna的可杂交区域至多170个碱基的区域杂交。靶向感兴趣基因组区域的grna集合中的每个grna可以与距grna集合中至少一个其他grna的可杂交区域约10个至200个碱基(核苷酸)的区域杂交。grna集合中的每个grna可以与距grna集合中至少一个其他grna的可杂交区域至少10个、15个、20个、25个、30个、35个、40个、45个、50个、60个、70个、80个、90个、100个、120个、140个、160个、180个、200个或更多个碱基的区域杂交。grna集合中的每个grna可以与距grna集合中至少一个其他grna的可杂交区域至多200个、180个、160个、140个、120个、100个、90个、80个、70个、60个、50个、45个、40个、35个、30个、25个、20个、15个、10个或更少碱基的区域杂交。在一种实例中,grna集合中的grna的可杂交区域之间的最小距离为距grna集合中至少一个其他grna的可杂交区域至少30个碱基。在另一种实例中,grna集合中的grna的可杂交区域之间的最大距离为距grna集合中至少一个其他grna的可杂交区域至多150个碱基。
[0356]
在一些情况下,crispr/cas活性可以用于其中期望以位点特异性(靶向)方式修饰dna(例如基因敲除(ko)、基因敲入(ki)、基因编辑、基因加标签等)的任何体外或体内应用,如用于例如基因疗法。基因疗法的实例包括治疗疾病或作为抗病毒、抗病原体或抗癌治疗剂;在农业中产生遗传修饰的生物体;通过细胞大规模产生用于治疗、诊断或研究目的的蛋白;诱导诱导性多能干细胞(ips细胞或ipsc);以及靶向病原体基因以用于缺失或替代。
[0357]
c.基因调控
[0358]
本发明公开内容的方面包括通过靶向基因的功能结构域(诸如阻遏因子结构域和激活因子结构域)用于调控基因表达或基因转录mrna的方法,称为敲低方法。与转录阻遏因子(例如,krab、dmt3a和/或lsd1)连接并结合crispr多核苷酸(例如,sgrna)的没有催化活性的cas核酸酶可以与基因中的互补dna区域结合并阻断转录。该方法的实施方案包括使cas核酸酶在光引发sgrna与该cas核酸酶的交联后没有催化活性,同时维持锁定的crispr复合物中的sgrna与靶dna序列的互补结合活性。在一些情况下,没有催化活性的crispr效应蛋白(例如,cas)可以与一种或更多种转录激活因子(例如,vp64、p65和/或rta19)融合,并且可以与crispr多核苷酸(例如,sgrna)形成稳定的(例如,锁定的)复合物。可以将稳定的(例如,锁定的)复合物递送至细胞中的基因来上调基因的转录。
[0359]
在一些情况下,功能结构域可以与没有催化活性的crispr效应蛋白连接(例如,细胞内的没有催化活性的crispr效应蛋白,可以形成crispr复合物)。包含与crispr效应蛋白交联的非天然核苷酸的crispr on多核苷酸还可以包含通过可光裂解元件与引导序列隔开的多核苷酸前导序列。细胞可以暴露于uv辐射,导致裂解元件的裂解和多核苷酸前导序列的释放。然后crispr复合物可以裂解靶序列。在一些情况下,也将供体核酸引入细胞,所述
供体核酸可以用于在裂解位点处的同源重组以将编辑引入核酸。
[0360]
核酸编辑可以靶向内源调控元件(例如,增强子或沉默子)。核酸编辑可以靶向启动子或启动子近侧元件。这些控制元件可以位于转录起始位点(tss)的上游或下游,距tss 200bp开始至100kb远。已知控制元件的靶向可以用于激活或阻遏感兴趣的基因。单个控制元件可以影响多于一种靶基因的转录。因此,单个控制元件的靶向可以用于同时控制多于一种基因的转录。
[0361]
一个或更多个功能结构域可以是核定位序列(nls)或核输出信号(nes)。
[0362]
一个或更多个功能结构域可以是转录激活结构域。转录激活结构域可以是vp64、p65、myod1、hsf1、rta、set7/9或组蛋白乙酰转移酶。crispr效应蛋白可以是与具有转录激活因子或阻遏因子活性的结构域融合的没有催化活性的cas蛋白。与具有转录激活因子或阻遏因子活性的结构域融合的没有催化活性的cas蛋白可以用于研究特定组织中特定基因对之间的上位相互作用。
[0363]
一个或更多个功能结构域可以具有一种或更多种活性,包括甲基化酶活性、脱甲基化酶活性、转录激活活性、转录阻遏活性、转录释放因子活性、组蛋白修饰活性、rna裂解活性、dna裂解活性、dna整合活性或核酸结合活性。
[0364]
一个或更多个功能结构域可以是转座酶结构域、整合酶结构域、重组酶结构域、解离酶结构域、转化酶结构域、蛋白酶结构域、dna甲基转移酶结构域、dna羟甲基化酶结构域、dna脱甲基化酶结构域、组蛋白乙酰化酶结构域、组蛋白脱乙酰化酶结构域、核酸酶结构域、阻遏因子结构域、激活因子结构域、核定位信号结构域、转录调节蛋白(或转录复合物募集)结构域、细胞摄取活性相关结构域、核酸结合结构域、抗体呈递结构域、组蛋白修饰结构酶、组蛋白修饰酶的募集物;组蛋白修饰酶的抑制物、组蛋白甲基转移酶、组蛋白脱甲基化酶、组蛋白激酶、组蛋白磷酸酶、组蛋白核糖基酶、组蛋白脱核糖基酶、组蛋白泛素化酶、组蛋白去泛素化酶、组蛋白生物素酶或组蛋白尾部蛋白酶。
[0365]
在一些情况下,功能结构域可以连接至没有催化活性的crispr效应蛋白(例如,没有催化活性的cas9蛋白)。连接至没有催化活性的crispr效应蛋白(例如,没有催化活性的cas9蛋白)的功能结构域可以用于结合和/或激活启动子或增强子。还可以提供一种或更多种包含能够与启动子或增强子退火的引导序列的crispr多核苷酸,以引导包含crispr效应蛋白(例如,没有催化活性的cas)的crispr复合物与启动子或增强子的结合。crispr on/off多核苷酸可以与crispr效应蛋白共价交联,该crispr效应蛋白可以是没有催化活性的cas9。没有催化活性的cas9可以与转录激活结构域(例如,vp64)融合。并且融合蛋白,例如cas9
‑
vp64融合蛋白,可以用于可调地调节靶基因或染色质区域的表达。例如,crispr on/off多核苷酸的多核苷酸前导序列可以阻止crispr复合物通过引导序列向靶基因的有效定位。多核苷酸前导序列的裂解可以导致crispr复合物通过引导序列对靶序列的有效靶向,这可以导致转录激活。随后,第二裂解媒介可以暴露于crispr多核苷酸,导致crispr多核苷酸的裂解,并且降低或抑制crispr复合物(或crispr效应蛋白,如果裂解的crispr多核苷酸已经与crispr效应蛋白解离)激活基因转录的能力。
[0366]
用本文描述的激活或抑制系统对区域的靶向,可以通过以下的转录读取来跟踪:a)一组推定的靶(例如,一组最接近控制元件定位的基因)或b)通过例如rnaseq或微阵列的全转录物组读取。
[0367]
在另一种实例中,本文提供的crispr复合物可以用于研究宿主细胞中两种或更多种靶基因的上位相互作用。方法可以包括以下步骤:(i)使用病毒或非病毒递送方法或其组合,将以下引入宿主细胞中:(a)包含第一和第二裂解元件的crispr多核苷酸,其中裂解元件对裂解敏感,并且其中引导序列的核苷酸序列与第一靶核酸序列完全或部分互补;(b)具有催化活性的crispr效应蛋白(例如,crispr酶),使得crispr多核苷酸(例如,sgrna)和crispr酶形成crispr复合物;并且(ii)在期望的时间,诱导crispr多核苷酸中第一裂解元件的裂解,并激活crispr复合物对靶基因的更高的靶特异性裂解,并且然后(iii)在期望的时间诱导第二裂解元件的裂解,从而使crispr复合物对靶基因的靶特异性裂解失活或降低。
[0368]
该方法还可以包括(i)使用病毒或非病毒递送方法或其组合,以将以下引入宿主细胞中:(a)包含第一和第二裂解元件的第二crispr多核苷酸,其中裂解元件对裂解敏感,并且其中引导序列的核苷酸序列与第二靶序列(例如,在靶基因中)的区域完全或部分互补;(b)具有催化活性的crispr酶,使得第二crispr多核苷酸(例如,sgrna)和crispr酶形成第二crispr复合物;并且(ii)在期望的时间,诱导第二crispr多核苷酸中第一裂解元件的裂解,并激活靶crispr复合物对基因的更高的靶特异性裂解,并且然后(iii)在期望的时间诱导第二裂解元件的裂解,从而使第二crispr复合物对靶基因的靶特异性裂解失活或降低。
[0369]
此外,第一和第二crispr复合物中第一裂解元件的裂解可以处于组织特异性启动子(例如,肌肉特异性启动子)的控制下。例如,细胞中遗传工程化的内切核糖核酸酶cas6a/csy4的表达可以置于组织特异性启动子(例如,肌肉启动子)的控制下,该启动子可以在特定时间被激活以诱导第一裂解元件的裂解。第一和第二crispr复合物中的第二裂解元件可以通过暴露于特定的序列特异性小分子在期望的时间被诱导裂解。crispr酶可以是与具有转录激活因子或阻遏因子活性的结构域融合的dcas9,并且可以用于研究特定组织中特定基因对之间的上位相互作用。
[0370]
在另一种实例中,本文描述的crispr复合物可以用于诱导宿主细胞中一种或更多种靶dna的两种或更多种靶基因的正交转录。术语“正交(orthogonal)”可以意指独立的,即,两种或更多种靶基因可以被独立地调节或独立地转录。该方法可以包括以下步骤:使用病毒或非病毒递送方法或其组合用于将以下引入宿主细胞中:(a)包含第一和第二裂解元件的两种或更多种不同的诱导型crispr多核苷酸,其中第一和第二序列元件对裂解敏感,并且其中引导序列的核苷酸序列与两种或更多种不同靶基因附近的一种或更多种靶dna完全或部分互补;(b)连接至转录激活因子结构域的无催化活性的crispr酶,使得不同的诱导型crispr多核苷酸和crispr酶形成不同的crispr复合物,其中crispr复合物包含一个或更多个效应物结构域;并且(ii)在期望的时间,诱导第一和第二多核苷酸中第一裂解元件的裂解,并且由此协调靶基因的表达。靶dna可以是在单个基因或控制元件内的邻近区域。
[0371]
d.药物配制
[0372]
本文描述的crispr多核苷酸和crispr复合物可以在体外或体内用于引起细胞或生物体的变化。crisrp多核苷酸和crispr效应蛋白可以作为复合物引入,或者它们可以在细胞内形成复合物。crispr多核苷酸和/或crispr效应蛋白可以被动引入细胞或通过媒介物引入。在引入时,crispr多核苷酸和crispr效应蛋白可以存在于缓冲液中。
[0373]
本发明公开内容的方面包括用于制备用于药物制剂的稳定的(例如,锁定的)crispr复合物的方法。crispr复合物可以被制备成通过例如脂质体和纳米颗粒递送来递送。可选地或组合地,编码crispr效应蛋白和/或crispr多核苷酸的载体可以通过例如显微注射或其他机械、物理或病毒方法递送至患者。crispr复合物和相关载体构建体可以与一种或更多种治疗剂、预防剂、诊断剂或成像剂组合使用。
[0374]
药物制剂可以包含一种或更多种赋形剂,以增加稳定性、增强对细胞的转染、控制释放(诸如从载体例如纳米颗粒的释放)、改变生物分布或改变编码crispr复合物的载体的翻译。药物制剂可以包含交联的crispr复合物、水、共溶剂、缓冲剂和ph调节剂的混合物。共溶赋形剂可以包括油、表面活性剂、乳化剂、稳定剂、螯合剂和防腐剂。稳定剂可以包括糖和氨基酸。糖可以包括蔗糖和乳糖。氨基酸可以包括甘氨酸和谷氨酸一钠。防腐剂可以包括苯酚、苯氧乙醇和硫柳汞。
[0375]
锁定的crispr复合物可以使用一种或更多种赋形剂配制,以便:(1)增加稳定性;(2)增加细胞转染;(3)允许持续释放或延迟释放(例如,从多核苷酸的贮库制剂(depot formulation)释放);(4)改变生物分布(例如,将多核苷酸、初级构建体或mrna靶向特定组织或细胞类型);(5)增加体内编码蛋白的翻译;和/或(6)改变体内编码蛋白的释放谱。
[0376]
赋形剂可以是溶剂、分散介质、稀释剂或其他液体媒介物、分散或悬浮助剂(aids)、表面活性剂、等渗剂、增稠剂或乳化剂、防腐剂和/或乳化剂、防腐剂、缓冲剂、润滑剂和/或油。赋形剂可以是类脂质(lipidoids),脂质体,脂质纳米颗粒,聚合物,复脂质体合物(lipoplex),核
‑
壳纳米颗粒,肽,蛋白,用多核苷酸、初级构建体或cas核酸酶mrna转染的细胞(例如,用于移植到受试者中),透明质酸酶,纳米颗粒模拟物及其组合。
[0377]
药物组合物中的crispr多核苷酸、crispr效应蛋白或编码这两者任一种的核酸以及药学上可接受的赋形剂和/或任何另外成分的相对量可以不同,这取决于所治疗的受试者的身份、尺寸和/或状况,并且还取决于组合物待被施用的途径。组合物可以包含0.1%和100%之间,例如0.5%和50%之间、1%
‑
30%之间、5%
‑
80%之间、至少80%(w/w)的crispr多核苷酸、crispr效应蛋白或编码这两者任一种的核酸。
[0378]
本文描述的包含至少一个用于交联的非天然核苷酸和活性调节性元件的crispr多核苷酸序列(诸如crispr on多核苷酸、crispr off多核苷酸或crispr on/off多核苷酸)可以配制为包含一种或更多种药学上可接受的赋形剂的药物组合物。药物组合物可以包含一种或更多种另外的活性物质,例如,治疗性和/或预防性活性物质。药物组合物的配制和/或制备中的一般考虑可见于例如remington:the science and practice of pharmacy第2版,lippincott williams&wilkins,2005中(通过引用以其整体并入本文)。
[0379]
类脂质的合成已被广泛描述,并且包含这些化合物的制剂特别适于递送本文描述的修饰的crispr on多核苷酸、crispr off多核苷酸或crispr on/off多核苷酸以及初级构建体(参见mahon等人,bioconjug chem.2010 21:1448
‑
1454;schroeder等人,j intern med.2010 267:9
‑
21;akinc等人,nat biotechnol.2008 26:561
‑
569;love等人,proc natl acad sci u s a.2010 107:1864
‑
1869;siegwart等人,proc natl acad sci u s a.2011108:12996
‑
3001;所有文献都以其整体并入本文)。不同比例的类脂质和其他组分(包括但不限于,二硬脂酰磷脂酰胆碱、胆固醇和peg
‑
dmg)可以用于优化多核苷酸、初级构建体或cas核酸酶mrna的制剂,以用于递送至不同细胞类型,包括,但不限于,肝细胞、髓样
细胞、肌细胞等。
[0380]
锁定的crispr复合物可以使用一种或更多种脂质体、脂质体复合物或脂质纳米颗粒(lnp)来配制。药物组合物可以包含脂质体。本文描述的药物组合物可以包含脂质体,诸如由稳定的质粒
‑
脂质颗粒(splp)或稳定的核酸脂质颗粒(snalp)的合成形成的脂质体,所述脂质体先前已被描述并显示适于体外和体内的多核苷酸递送(参见wheeler等人gene therapy.1999 6:271
‑
281;zhang等人gene therapy.1999 6:1438
‑
1447;jeffs等人pharm res.2005 22:362
‑
372;morrissey等人,nat biotechnol.2005 2:1002
‑
1007)。crispr多核苷酸可以被配制在脂质囊泡中,所述脂质囊泡可以在官能化的脂质双层之间具有交联。
[0381]
锁定的crispr复合物可以配制在脂质
‑
聚阳离子复合物中。脂质
‑
聚阳离子复合物的形成可以通过本领域已知的方法和/或如美国公布第20120178702号中描述的方法来完成,所述美国公布通过引用以其整体并入本文。药物组合物可以包含国际公布第2012099755号中描述的peg化脂质中的至少一种,所述国际公布通过引用并入本文。
[0382]
lnp制剂可以通过国际公布第wo2011 127255号或第wo2008 103276号中描述的方法来配制,所述国际公布的每一个都通过引用以其整体并入本文。crispr多核苷酸可以被包封在lnp制剂中,如wo2011 127255和/或wo2008103276中描述的;其中每一项都通过引用以其整体并入本文。
[0383]
锁定的crispr复合物可以配制为固体脂质纳米颗粒。固体脂质纳米颗粒(sln)可以是具有10nm至1000nm之间的平均直径的球形。sln可以具有固体脂质核心基质,所述固体脂质核心基质可以溶解亲脂性分子并且可以用表面活性剂和/或乳化剂稳定。脂质纳米颗粒可以是自组装脂质
‑
聚合物纳米颗粒(参见zhang等人,acs nano,2008,2(8),第1696
‑
1702页;通过引用以其整体并入本文)。
[0384]
锁定的crispr复合物、初级构建体或cas核酸酶mrna可以被包封到脂质纳米颗粒或快速消除性脂质纳米颗粒中,并且然后脂质纳米颗粒或快速消除性脂质纳米颗粒可以被包封到本文描述和/或本领域已知的聚合物、水凝胶和/或手术密封剂中。
[0385]
用于受控释放和/或靶向递送的锁定的crispr复合物制剂还可以包含至少一种受控释放包衣。受控释放包衣包括,但不限于,聚乙烯吡咯烷酮/乙酸乙烯酯共聚物、聚乙烯吡咯烷酮、羟丙基甲基纤维素、羟丙基纤维素、羟乙基纤维素。
[0386]
受控释放和/或靶向递送制剂可以包含至少一种可以包含聚阳离子侧链的可降解聚酯。可降解聚酯可以是聚(丝氨酸酯)、聚(l
‑
丙交酯
‑
共
‑
l
‑
赖氨酸)、聚(4
‑
羟基
‑
l
‑
脯氨酸酯)及其组合。可降解聚酯可以包括peg缀合物以形成peg化聚合物。
[0387]
锁定的crispr复合物可以被包封在治疗性纳米颗粒中。治疗性纳米颗粒可以被配制为用于持续释放。时间段可以包括小时、天、周、月和年。作为非限制性实例,持续释放纳米颗粒可以包含聚合物和治疗剂,例如,本文描述的crispr多核苷酸(参见国际公布第2010075072号和美国公布第us20100216804号和美国公布第us20110217377号,其中每一项通过引用以其整体并入本文)。治疗性纳米颗粒可以被配制为靶特异性的。治疗性纳米颗粒可以包括皮质类固醇(参见国际公布第wo2011084518号)。
[0388]
锁定的crispr复合物可以被包封在合成纳米载体中、与合成纳米载体连接和/或缔合。合成纳米载体可以通过国际公布第wo2010005740号、第wo2010030763号中描述的方法配制。合成纳米载体可以包含反应性基团,以释放本文描述的crispr多核苷酸(参见国际
公布第wo20120952552号和美国公布第us20120171229号,所述公布的每一个通过引用以其整体并入本文)。
[0389]
合成纳米载体可以被配制为用于靶向释放。合成纳米载体可以被配制为在特定的ph和/或在期望的时间间隔之后释放crispr复合物。合成纳米颗粒可以被配制为在24小时之后和/或在4.5的ph释放多核苷酸、初级构建体和/或cas核酸酶mrna(参见国际公布第wo2010138193号和第wo2010138194号以及美国公布第us201 10020388号和第us20110027217号,所述公布的每一个通过引用以其整体并入本文)。
[0390]
合成纳米载体可以被配制为用于本文描述的锁定的crispr复合物的受控释放和/或持续释放。用于持续释放的合成纳米载体可以例如如本文描述的和/或如国际公布第wo2010138192号和美国公布第20100303850号中描述的被配制,所述公布的每一个通过引用以其整体并入本文。
[0391]
锁定的crispr复合物可以用聚合化合物配制或在聚合化合物中配制。聚合物可以包括至少一种聚合物聚乙烯、聚乙二醇(peg)、聚(l
‑
赖氨酸)(pll)、接枝至pll的peg、阳离子脂质聚合物、可生物降解的阳离子脂质聚合物、聚乙烯亚胺(pei)、交联的支化聚(亚烷基亚胺)、聚胺衍生物、改性泊洛沙姆、可生物降解的聚合物、可生物降解的嵌段共聚物、可生物降解的无规共聚物、可生物降解的聚酯共聚物、可生物降解的聚酯嵌段共聚物、可生物降解的聚酯嵌段无规共聚物、线性可生物降解的共聚物、聚[a
‑
(4
‑
氨基丁基)
‑
l
‑
乙醇酸](paga)、可生物降解的交联的阳离子多嵌段共聚物、聚碳酸酯、聚酸酐、聚羟基酸、聚富马酸丙酯、聚己内酯、聚酰胺、聚缩醛、聚醚、聚酯、聚(原酸酯)、聚氰基丙烯酸酯、聚乙烯醇、聚氨酯、聚磷腈、聚丙烯酸酯、聚甲基丙烯酸酯、聚氰基丙烯酸酯、聚脲、聚苯乙烯、聚胺、聚赖氨酸、聚(乙烯亚胺)、聚(丝氨酸酯)、聚(l
‑
丙交酯
‑
共
‑
l
‑
赖氨酸)、聚(4
‑
羟基
‑
l
‑
脯氨酸酯)、丙烯酸聚合物、含胺聚合物或其组合。
[0392]
本文描述的锁定的crispr复合物可以与另一种化合物缀合。crispr多核苷酸也可以使用聚合物、脂质和/或其他可生物降解的剂(例如,磷酸钙)的组合配制为纳米颗粒。组分可以以核
‑
壳、混合和/或逐层(layer
‑
by
‑
layer)结构组合,以允许纳米颗粒的微调,从而可以增强crispr多核苷酸的递送(wang等人,nat mater.2006 5:791
‑
796;fuller等人,biomaterials.2008 29:1526
‑
1532;dekoker等人,adv drug deliv rev.2011 63:748
‑
761;endres等人,biomaterials.2011 32:7721
‑
7731;su等人,mol pharm.2011jun 6;8(3):774
‑
87;通过引用以其整体并入本文)。
[0393]
锁定的crispr复合物可以与肽和/或蛋白一起配制,以增加锁定的crispr复合物对细胞的转染。肽可以是细胞穿透肽和细胞穿透蛋白,并且能够实现细胞内递送的肽可以用于递送药物制剂。
[0394]
锁定的crispr复合物可以离体转染到细胞中,并且随后移植到受试者中。这样的载体的实例包括编码crispr效应蛋白或相关多肽的初级核酸构建体或合成序列。药物组合物可以包括将修饰的rna递送至肝的红细胞和髓样细胞,以病毒样颗粒(vlp)递送修饰的rna的病毒微体(virosome),以及递送修饰的rna的电穿孔细胞,例如来自(gaithersburg,md)和(lyon,france)的电穿孔细胞。
[0395]
本文公开的锁定的crispr复合物或相关的载体构建体的基于细胞的制剂可以用于确保细胞转染(例如,以细胞载体),改变锁定的crispr复合物的生物分布(例如,通过将
细胞载体靶向特定组织或细胞类型)和/或增加编码蛋白的翻译。
[0396]
组合物也可以被配制为直接递送至器官或组织,通过例如直接浸泡或沐浴、通过导管、通过凝胶、粉末、软膏、乳膏、凝胶、洗剂和/或滴剂、通过使用基质诸如涂覆或浸渍了该组合物的织物或可生物降解材料等。
[0397]
本发明公开内容的方面包括用于向患者施用药物制剂的方法。未结合的rna可以通过干扰素γ引发免疫应答。锁定的crispr复合物可以大大降低药物组合物中未结合的sgrna的可能性。连接的crispr复合物和相关序列/多肽可以通过产生有效治疗结果的任何途径施用。这些途径包括肠内、胃肠内、硬膜外、口服、经皮、硬膜外(epidural,硬膜外(peridural))、脑内(进入大脑)、脑室内(进入脑室)、表皮(epicutaneous,应用到皮肤上)、皮内(进入皮肤本身)、皮下(皮肤下)、鼻施用(通过鼻子)、静脉内(进入静脉)、动脉内(进入动脉)、肌内(进入肌肉)、心内(进入心脏)、骨内输注(进入骨髓)、鞘内(进入椎管)、腹膜内(输注或注射到腹膜中)、膀胱内输注、玻璃体内(通过眼睛)、海绵体内注射(进入阴茎根部)、阴道内施用、宫内施用、羊膜外施用、经皮(扩散通过完整皮肤用于全身分布)、经粘膜(扩散通过粘膜)、吹入法(嗅吸法)、舌下、唇下、灌肠、滴眼液(滴到结膜上)或在滴耳液中。组合物可以以允许它们穿过血脑屏障、血管屏障或其他上皮屏障的方式施用。
[0398]
药物制剂可以每天4次、每天3次、每天2次、每天1次、每周3次、每周2次、每周1次、每月4次、每月3次、每月2次、每月1次、每年4次、每年3次、每年2次或每年1次向有相应需要的受试者施用。施用可以持续一定时间段,并且所述时间段可以是至少或多达1周、至少或多达1个月、至少或多达1年、至少或多达10年或受试者的一生。
[0399]
e.状况
[0400]
本发明公开内容的方面包括用crispr复合物治疗疾病状况。疾病状况可以包括癌症、神经状况、自身免疫性紊乱等。疾病状况的治疗可以包括患病组织的治疗。治疗可以包括用crispr复合物处理细胞,随后将所述细胞注射、移植或植入人类患者中。
[0401]
本文描述的crispr on多核苷酸、crispr off多核苷酸或crispr on/off多核苷酸可以用于其中期望将物质(“有效负载”)递送至生物靶的许多不同情况,例如递送用于检测靶的可检测物质或递送治疗剂的情况。crispr多核苷酸和相关载体构建体可以与一种或更多种其他治疗剂、预防剂、诊断剂或成像剂组合使用。
[0402]
本文描述的crispr on多核苷酸、crispr off多核苷酸或crispr on/off多核苷酸和其他初级构建体可以被设计为包括任何有用取向的接头和有效负载两者。例如,具有两个末端的接头可以用于将一个末端附接至有效负载,并且另一个末端附接至核酸碱基,诸如附接至脱氮
‑
腺苷或脱氮
‑
鸟苷的c
‑
7或c
‑
8位置或者胞嘧啶或尿嘧啶的n
‑
3或c
‑
5位置处。有效负载可以是治疗剂,诸如细胞毒素、放射性离子、化学治疗剂或其他治疗剂。
[0403]
本文描述的crispr on多核苷酸、crispr off多核苷酸或crispr on/off多核苷酸可以用于改变细胞的表型。crispr多核苷酸或crispr效应蛋白编码序列可以用于治疗和/或临床和研究环境。本文描述的crispr on多核苷酸、crispr off多核苷酸或crispr on/off多核苷酸和相关载体构建体以及由它们翻译的蛋白可以用作治疗剂或预防剂。例如,本文描述的crispr多核苷酸或cas核酸酶mrna(例如,编码crispr相关多肽或效应蛋白的修饰的mrna)可以被施用至受试者并且在体内翻译,以在受试者中引导治疗相关或预防性多肽的表达。
[0404]
引导序列(位于核酸靶向引导rna或sgrna内)引导核酸靶向复合物与靶核酸序列的序列特异性结合的能力可以通过任何合适的测定来评估。例如,核酸靶向crispr系统的足以形成核酸靶向复合物的组分(包括待测试的引导序列)可以被提供至具有对应靶核酸序列的宿主细胞,诸如通过用编码核酸靶向复合物的组分的载体转染,随后评估靶核酸序列内的优先靶向(例如,裂解)。靶核酸序列的裂解可以在试管中通过以下来评估:提供靶核酸序列、核酸靶向复合物的组分(包括待测试的引导序列和不同于测试引导序列的对照引导序列),并且比较测试引导序列和对照引导序列反应之间靶序列处的结合或裂解速率。
[0405]
本文提供的组合物可以用于治疗多种疾病、紊乱和/或状况中的任一种,例如以下一种或更多种:自身免疫性紊乱(例如糖尿病、狼疮、多发性硬化、银屑病、类风湿性关节炎);炎性紊乱(例如关节炎、盆腔炎性疾病);感染性疾病(例如病毒感染(例如,hiv、hcv、rsv)、细菌感染、真菌感染、脓毒症);神经系统紊乱(例如阿尔茨海默病、亨廷顿氏病;孤独症;杜氏肌营养不良症);心血管紊乱(例如动脉粥样硬化、高胆固醇血症、血栓形成、凝血障碍、血管生成紊乱诸如黄斑变性);增生性紊乱(例如癌症、良性赘生物);呼吸系统紊乱(例如慢性阻塞性肺疾病);消化系统紊乱(例如炎性肠病、溃疡);肌肉骨骼紊乱(例如纤维肌痛、关节炎);内分泌、代谢和营养紊乱(例如糖尿病、骨质疏松症);泌尿系统紊乱(例如肾病);精神疾病(例如抑郁症、精神分裂症);皮肤紊乱(例如创伤、湿疹);血液和淋巴紊乱(例如贫血、血友病)等。
[0406]
在一方面,疾病状况可以是心血管疾病、糖尿病、肺部疾病、慢性阻塞性肺病(copd)、哮喘、特发性肺纤维化、慢性支气管炎、囊性纤维化、冠心病、脑血管疾病等。在一方面,疾病状况可以是阿尔茨海默病、肌萎缩性侧索硬化(als)、动静脉畸形、多发性硬化(ms)或帕金森病。在另一方面,疾病状况可以是银屑病、格雷夫斯病、格林
‑
巴利综合征、桥本氏甲状腺炎、血管炎、重症肌无力、慢性炎性脱髓鞘性多神经病、1型糖尿病、多发性硬化、全身性红斑狼疮、类风湿性关节炎等。
[0407]
在一方面,疾病状况可以是胆道癌(例如,腺癌)、肺癌(例如,大细胞癌、非小细胞癌、鳞状细胞癌、赘生物等),结肠直肠癌、前列腺癌、子宫内膜癌、卵巢癌、造血系统癌、白血病、淋巴癌、肾癌、乳腺癌(例如癌(carcinoma))、食管癌、胰腺癌、皮肤癌(例如基底细胞癌、鳞状细胞癌、恶性黑素瘤等)、软组织癌(例如血管肉瘤、平滑肌肉瘤、脂肪肉瘤、横纹肌肉瘤、粘液瘤、恶性纤维组织细胞瘤
‑
多形性肉瘤等)、睾丸癌(例如,生殖细胞瘤、精原细胞瘤等)、甲状腺癌(例如,未分化癌、滤泡性癌、乳头状癌、hurthle细胞癌等)、膀胱癌(例如移行细胞癌)、宫颈癌(例如腺癌)、子宫癌、腹膜癌、脑癌、神经母细胞瘤、间皮瘤、胆管癌、软骨肉瘤、白血病(例如aml、cml、cmml、jmml等)、淋巴瘤(例如all、伯基特淋巴瘤、霍奇金淋巴瘤、浆细胞骨髓瘤等)、肾上腺皮质癌、肛门癌、再生障碍性贫血、胆管癌、骨癌、骨转移、脑癌、中枢神经系统(cns)癌、外周神经系统(pns)癌、宫颈癌、儿童非霍奇金淋巴瘤、胰腺癌(例如导管腺癌、内分泌肿瘤等)、结肠癌(例如腺癌、腺瘤等)、结肠癌和直肠癌、子宫内膜癌、食管癌、尤因氏肿瘤家族(例如尤因氏肉瘤)、眼癌、胆囊癌、胃肠道类癌、胃肠道间质瘤、妊娠性滋养层细胞疾病、毛细胞白血病、霍奇金淋巴瘤、卡波西肉瘤、肾癌、喉癌和咽癌、急性淋巴细胞白血病、急性髓性白血病、儿童白血病、慢性淋巴细胞白血病、慢性髓性白血病、肝癌(例如肝细胞癌)、肺类癌肿瘤、男性乳腺癌、恶性间皮瘤、多发性骨髓瘤、骨髓增生异常综合征、骨髓增生性紊乱、鼻腔和副鼻癌、鼻咽癌、神经母细胞瘤、口腔和口咽癌、骨肉瘤、卵巢
癌、阴茎癌、垂体肿瘤、前列腺癌(例如腺癌)、视网膜母细胞瘤、横纹肌肉瘤、唾液腺癌、肉瘤、黑素瘤皮肤癌、非黑素瘤皮肤癌、胃癌(例如腺癌等)、胸腺癌、甲状腺癌、子宫癌(例如子宫肉瘤)、移行细胞癌、阴道癌、外阴癌、间皮瘤、鳞状细胞癌或表皮样癌、支气管腺瘤、绒毛膜癌、头颈癌、畸胎瘤、瓦尔登斯特伦巨球蛋白血症、神经节癌(ganglia cancer)(例如神经母细胞瘤)或神经鞘癌。
[0408]
f.crispr效应蛋白和crispr多核苷酸的表达
[0409]
在一些情况下,可以将用于表达crispr多核苷酸和crispr效应蛋白(例如,crispr酶)的一种或更多种表达载体转染到宿主细胞中。可以首先将包含编码crispr多核苷酸的dna序列的表达载体转染到宿主细胞中,并且然后将包含编码crispr效应蛋白(例如,crispr酶)的dna序列的表达载体转染到宿主细胞中。可以将包含编码crispr效应蛋白(例如,crispr酶)的dna序列的表达载体和包含编码诱导型crispr多核苷酸的dna序列的表达载体同时转染到宿主细胞中。可以将包含编码crispr效应蛋白(例如,crispr酶)的dna序列和编码诱导型crispr多核苷酸的dna序列的单一(类型)表达载体转染到宿主细胞中。宿主细胞可以是内源表达crispr效应蛋白(例如,crispr酶)的宿主细胞。编码crispr效应蛋白(例如,crispr酶)的信使rna也可以与crispr多核苷酸(例如,sgrna)一起使用以用于基因编辑。当使用载体时,载体可以包含诱导型启动子。条件型启动子和/或诱导型启动子和/或组织特异性启动子可以是rna聚合酶pol i、pol ii、pol iii、t7、u6、h1启动子、逆转录病毒劳斯肉瘤病毒(rsv)ltr启动子、巨细胞病毒(cmv)启动子、sv40启动子、二氢叶酸还原酶启动子、β肌动蛋白启动子、磷酸甘油激酶(pgk)启动子和ef1启动子。在一些情况下,可以将编码crispr效应蛋白(例如,crispr酶)的转基因整合到细胞基因组中。
[0410]
可以将表达crispr效应蛋白(例如,crispr酶)的转基因引入细胞中。可以将crispr效应蛋白(例如,crispr酶,例如,cas9)转基因引入分离的细胞中。crispr复合物转基因细胞可以通过从转基因生物体中分离细胞来获得。也如本文描述的,crispr效应蛋白(例如,crispr酶,例如,cas9)转基因可以通过载体(例如,aav、腺病毒、慢病毒)和/或颗粒和/或纳米颗粒递送的方式递送至真核细胞。
[0411]
在一些情况下,crispr多核苷酸可以被诱导表达。在一些情况下,crispr效应蛋白(例如,crispr酶)可以被诱导表达。诱导crispr多核苷酸和/或crispr效应蛋白(例如,crispr酶)的表达可以导致crispr多核苷酸/crispr效应蛋白(例如,crispr酶)复合物的形成,该复合物可以在期望的时间被“开启”以靶向靶核酸(例如,靶dna)并使该靶核酸(例如,靶dna)裂解。通过限制复合物的活性半衰期或通过在模型生物体或人类细胞中实现组织特异性编辑,诱导型复合物可以用于降低脱靶效应。
[0412]
诱导型crispr多核苷酸和/或crispr效应蛋白(例如,crispr酶)可以在宿主细胞内表达。表达可以是以任何顺序。
[0413]
v.crispr多核苷酸合成
[0414]
本文描述的用用于交联的非天然核苷酸修饰的多核苷酸(还包括crispr on多核苷酸序列、crispr off多核苷酸序列、crispr on/off多核苷酸序列)或者当与crispr酶复合时具有降低的脱靶编辑活性的包含一种或更多种修饰的crispr多核苷酸可以通过本领域普通技术人员已知的任何方法合成。本文描述的用用于交联的非天然核苷酸修饰的多核苷酸(还包括crispr on多核苷酸、crispr off多核苷酸或crispr on/off多核苷酸)可以化
学合成。本文描述的用用于交联的非天然核苷酸修饰的多核苷酸(还包括crispr on多核苷酸、crispr off多核苷酸或crispr on/off多核苷酸)可以使用2'
‑
o
‑
硫代氨基甲酸酯保护的核苷亚磷酰胺合成。多核苷酸的合成方法描述于以下中:例如,dellinger等人,j.american chemical society 133,11540
‑
11556(2011);threlfall等人,organic&biomolecular chemistry 10,746
‑
754(2012);和dellinger等人,j.american chemical society 125,940
‑
950(2003)。本文描述的任何修饰可以组合并掺入本文描述的crispr on多核苷酸、crispr off多核苷酸或crispr on/off多核苷酸中,例如掺入引导序列和/或结合crispr效应蛋白的序列(例如,支架序列)中。可选地,crispr多核苷酸可以通过以下制备:通过beaucage和caruthers(tetrahedron lett.,(1981)22:1859
‑
1862)描述的亚磷酰胺方法或根据matteucci等人,(j.am.chem.soc,(1981)103:3185)通过三酯方法,每个文献通过引用明确并入本文,或者通过使用商业自动化多核苷酸合成仪的其他化学方法。
[0415]
crispr多核苷酸可以化学合成,例如,根据由beaucage和caruthers,tetrahedron lett.22:1859
‑
1862(1981)首次描述的固相亚磷酰胺三酯方法,使用自动化合成仪,如van devanter等人,nucleic acids res.12:6159
‑
6168(1984)中描述的。crispr多核苷酸的合成可以包括在固相合成期间利用特殊的亚磷酰胺试剂引入化学修饰。
[0416]
a.sgrna连接
[0417]
为sgrna的crispr多核苷酸可以包含通过非磷酸二酯键化学连接或缀合的修饰的crrna和tracrrna序列。修饰的crrna和tracrrna序列可以通过非核苷酸环来化学连接或缀合。修饰的crrna和tracrrna可以通过非磷酸二酯共价接头连接。共价接头可以是选自由以下组成的组的化学部分:香豆素、氨基甲酸酯、醚、酯、酰胺、亚胺、脒、氨基三嗪、腙、二硫化物、硫醚、硫酯、硫代磷酸酯、二硫代磷酸酯、磺酰胺、磺酸酯、砜、亚砜、脲、硫脲、酰肼、肟、三唑、光不稳定连接、c
‑
c键形成基团诸如diels
‑
alder环加成对或闭环换位反应对和迈克尔反应对。
[0418]
b.可裂解元件
[0419]
本文的crispr多核苷酸中的可裂解元件可以在每个末端处提供有可以被适当保护或激活的官能团。官能团可以通过醚、酯、氨基甲酸酯、磷酸酯或胺连接共价附接。例如,六甘醇可以在一个末端上用光不稳定保护基团(即,nvoc或menpoc)保护,并且在另一个末端上用2
‑
氰乙基
‑
n,n
‑
二异丙基氨基
‑
氯代亚磷酸酯激活以形成亚磷酰胺。其他形成醚、氨基甲酸酯或胺连接的方法是本领域技术人员已知的,并且特定试剂和参考文献可见于诸如march,advanced organic chemistry,第4版,wiley
‑
interscience,new york,n.y.,1992的教科书中。
[0420]
合成本文描述的接头的方法是本领域熟知的。下文提供了合成本技术的接头的方法的非限制性实例:
[0421][0422][0423]
可裂解接头和合成所述接头的方法的其他非限制性实例描述如下:
[0424][0425][0426]
可裂解接头和合成所述接头的方法的非限制性实例描述如下:
[0427][0428][0429]
c.sgrna合成
[0430]
包含crrna和tracrrna的sgrna可以首先使用亚磷酰胺合成方案合成(herdewijn,p.,编著,methods in molecular biology col 288,polynucleotide synthesis:methods and applications,humana press,new jersey(2012))。包含crrna和tracrrna序列的sgrna可以被官能化,以包含用于连接的适当的官能团(参见,例如,hermanson,g.t.,bioconjugate techniques,academic press(2013))。官能团可以是羟基、胺、羧酸、羧酸卤化物、羧酸活性酯、醛、羰基、氯代羰基、香豆素、补骨脂素、双吖丙啶或叠氮化物。修饰的tracr和tracr配偶序列被官能化后,两个多核苷酸之间可以形成共价化学键或连接。化学键可以基于香豆素、氨基甲酸酯、醚、酯、酰胺、亚胺、脒、氨基三嗪、腙、二硫化物、硫醚、硫酯、硫代磷酸酯、二硫代磷酸酯、磺酰胺、磺酸酯、砜、亚砜、脲、硫脲、酰肼、肟、三唑、光不稳定连接、c
‑
c键形成基团诸如diels
‑
alder环加成对或闭环换位反应对和迈克尔反应对。
[0431]
包含crrna和tracrrna序列的sgrna可以化学合成。sgrna可以以融合的形式一起合成,或者可以单独合成并化学连接。化学合成可以使用自动化固相多核苷酸合成机器与2'
‑
乙酰氧基乙基原酸酯(2'
‑
ace)(scaringe等人,j.am.chem.soc.(1998)120:11820
‑
11821;scaringe,methods enzymol.(2000)317:3
‑
18)或2'
‑
硫代氨基甲酸酯(2'
‑
tc)化学(dellinger等人,j.am.chem.soc.(2011)133:11540
‑
11546;hendel等人,nat.biotechnol.(2015)33:985
‑
989)。
[0432]
sgrna可以通过糖、核苷酸间磷酸二酯键、嘌呤和嘧啶残基的修饰与各种生物缀合反应物、环、桥和非核苷酸连接共价连接。sletten等人,angew.chem.int.编著(2009)48:6974
‑
6998;manoharan,m.curr.opin.chem.biol.(2004)8:570
‑
9;behlke等人,polynucleotides(2008)18:305
‑
19;watts,等人,drug.discov.today(2008)13:842
‑
55;shukla等人,chemmedchem(2010)5:328
‑
49。
[0433]
sgrna可以通过点击化学来组装。crrna tracrrna和/或其中的序列元件可以使用三唑接头通过共价连接来组装。sgrna可以通过连接5
’‑
己炔tracrrna和3
’‑
叠氮化物crrna共价连接。5
’‑
己炔tracrrna和3
’‑
叠氮化物crrna中的任一个或两个可以用2
’‑
乙酰氧基乙基原酸酯(2'
‑
ace)基团来保护,2
’‑
乙酰氧基乙基原酸酯(2'
‑
ace)基团随后可以使用dharmacon方案去除(scaringe等人,j.am.chem.soc.(1998)120:11820
‑
11821;scaringe,methods enzymol.(2000)317:3
‑
18)。
[0434]
vi.试剂盒
[0435]
试剂盒可以包括一种或更多种本文描述的组分。试剂盒可以包括本文描述的crispr多核苷酸。试剂盒可以包括本文描述的crispr效应蛋白(例如,crispr酶,例如cas9)。试剂盒可以包括本文描述的crispr复合物,该crispr复合物包含本文描述的crispr多核苷酸和本文描述的crispr效应蛋白。试剂盒可以包括接头,例如可裂解接头。试剂盒可以包括可光裂解接头。试剂盒可以包括说明书。试剂盒可以包括细胞或生物体,该细胞或生物体包含本文描述的crispr多核苷酸、crispr效应蛋白或crispr复合物。
[0436]
试剂盒可以包括遗传构建体(例如,用于表达一种或更多种crispr多核苷酸和/或一种或更多种crispr效应蛋白的载体系统)和使用该试剂盒的说明书。试剂盒可以包括细胞,该细胞包含一种或更多种用于表达本文描述的crispr多核苷酸和/或crispr效应蛋白的遗传构建体(例如,一种或更多种载体系统)。
[0437]
试剂盒可以包括赋形剂,该赋形剂用于产生适于使核酸靶与例如本文描述的crispr复合物接触的组合物。组合物可以适于接触基因组内的核酸靶序列。组合物可以适于将组合物(例如,crispr多核苷酸(例如,sgrna),例如,与crispr效应蛋白(例如,crispr酶,例如,cas9)复合的crispr多核苷酸)递送至细胞。组合物可以适于将crispr多核苷酸(例如,grna,或其与crispr酶的复合物)递送至受试者。赋形剂可以是药学上可接受的赋形剂。
[0438]
试剂盒可以包括用于裂解本文描述的crispr多核苷酸的一个或更多个可裂解元件的一种或更多种试剂。一种或更多种试剂可以在任何合适的容器中提供。试剂盒可以包括一种或更多种反应缓冲液或储存缓冲液。试剂盒可以包括试剂。试剂可以以可用于特定测定的形式提供,或者以在使用之前需要添加一种或更多种其他组分的形式(例如以浓缩或冻干的形式)提供。反应缓冲液或储存缓冲液可以是任何缓冲液,例如,碳酸钠缓冲液、碳
酸氢钠缓冲液、硼酸盐缓冲液、tris缓冲液、mops缓冲液、hepes缓冲液或其组合。缓冲液可以具有约7至约10的ph。
[0439]
试剂盒可以包括对应于引导序列的一种或更多种多核苷酸,所述一种或更多种多核苷酸用于插入到载体中以便可操作地连接引导序列和调节元件。试剂盒可以包括同源重组模板多核苷酸。
[0440]
本发明公开内容的示例性非限制性方面
[0441]
以上描述的本发明主题的各方面,包括实施方案,单独的或与一个或更多个其他方面或实施方案组合可以是有益的。在不限制以上描述的情况下,下文提供了编号为1
‑
390的公开内容的某些非限制性方面。对于阅读本发明公开内容的本领域技术人员来说将明显的是,每个单独编号的方面可以与任何前面或后面的单独编号的方面一起使用或组合。这意图为所有这样的方面组合提供支持,并且不限于下文明确提供的方面组合。
[0442]
1.一种crispr复合物,所述crispr复合物包含与crispr效应蛋白交联的引导rna(grna),所述引导rna(grna)与所述crispr效应蛋白在所述grna内的核苷酸处交联,其中所述grna包含靶结合区和crispr效应蛋白结合区,并且其中所述核苷酸位于所述grna的靶结合区之外。
[0443]
2.根据实施方案1所述的crispr复合物,其中所述核苷酸包含尿嘧啶。
[0444]
3.根据实施方案1或2所述的crispr复合物,其中所述grna是包含crrna区和tracrrna区的单引导rna(sgrna),所述crrna区包含靶结合区,并且其中所述核苷酸位于所述crrna区的靶结合区之外。
[0445]
4.根据实施方案3所述的crispr复合物,其中所述核苷酸位于所述sgrna的核苷酸位置49处,其中核苷酸位置1位于所述crrna的靶结合区的5’末端处,并且所述sgrna的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。
[0446]
5.根据实施方案3所述的crispr复合物,其中所述核苷酸位于所述sgrna的以下一个或更多个核苷酸位置处:22、23、24、25、31、37、44、49、45、50、56、59、63、64、66、71、72、77、78、80、84、90或94,其中核苷酸位置1位于所述crrna的靶结合区的5’末端处,并且所述sgrna的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。
[0447]
6.根据实施方案1至5中任一项所述的crispr复合物,其中所述核苷酸是非天然核苷酸。
[0448]
7.根据实施方案6所述的crispr复合物,其中所述非天然核苷酸包含糖的修饰。
[0449]
8.根据实施方案6或7所述的crispr复合物,其中所述非天然核苷酸包含碱基的修饰。
[0450]
9.根据实施方案6所述的crispr复合物,其中所述非天然核苷酸包含马来酰亚胺。
[0451]
10.根据实施方案9所述的crispr复合物,其中所述马来酰亚胺与所述crispr效应蛋白上的半胱氨酸共价连接。
[0452]
11.根据实施方案6所述的crispr复合物,其中所述grna通过交联剂与所述crispr效应蛋白交联,其中所述交联剂包括或衍生自吡啶基二硫化物、烷氧基胺、nhs酯、双吖丙啶、亚氨酸酯、卤代乙酰基基团、酰肼、芳基叠氮化物、异氰酸酯、二硫醇亚磷酰胺dtpa、4
‑
硫代
‑
utp、5
‑
叠氮基
‑
utp、5
‑
溴
‑
utp、8
‑
叠氮基
‑
atp、5
‑
apas
‑
utp或8
‑
n(3)amp。
[0453]
12.根据实施方案6所述的crispr复合物,其中所述grna通过交联剂与所述crispr
效应蛋白交联,其中所述交联剂包括二硫化物、酰胺、亚胺、酰肼、o
‑
烷基肟、烷基、胺、醇、三唑、异噁唑啉、异噁唑烷、异噁唑或哒嗪。
[0454]
13.根据实施方案3至12中任一项所述的crispr复合物,其中所述核苷酸位于所述tracrrna区的茎环中。
[0455]
14.根据实施方案13所述的crispr复合物,其中所述茎环的结构相对于缺少所述核苷酸的sgrna的茎环的结构被维持。
[0456]
15.根据实施方案3至14中任一项所述的crispr复合物,其中所述核苷酸位于所述tracrrna区的凸起中。
[0457]
16.根据实施方案15所述的crispr复合物,其中所述凸起的结构相对于缺少所述核苷酸的sgrna的凸起的结构被维持。
[0458]
17.根据实施方案3至16中任一项所述的crispr复合物,其中所述核苷酸位于所述tracrrna区的茎环之间。
[0459]
18.根据实施方案1至17中任一项所述的crispr复合物,其中所述crispr复合物包括核酸酶活性。
[0460]
19.根据实施方案18所述的crispr复合物,其中所述crispr复合物的脱靶核酸酶活性等于或小于包含未交联的所述crispr效应蛋白和所述grna的crispr复合物的脱靶核酸酶活性。
[0461]
20.根据实施方案1至19中任一项所述的crispr复合物,其中所述核苷酸位于所述crispr效应蛋白的半胱氨酸的20埃以内。
[0462]
21.根据实施方案1至20中任一项所述的crispr复合物,其中所述核苷酸不是4
‑
硫代尿苷或修饰的腺苷。
[0463]
22.一种crispr复合物,所述crispr复合物包含与crispr效应蛋白交联的单引导rna(sgrna),所述单引导rna(sgrna)与所述crispr效应蛋白在所述sgrna的核苷酸位置49处的核苷酸处交联,其中所述sgrna包含crrna区和tracrrna区,所述crrna区包含靶结合区,并且其中核苷酸位置1位于所述crrna的靶结合区的5’末端处,并且所述sgrna的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。
[0464]
23.根据实施方案22所述的crispr复合物,其中核苷酸位置49处的核苷酸包含尿嘧啶。
[0465]
24.根据实施方案22或23所述的crispr复合物,其中所述crispr复合物包括核酸酶活性。
[0466]
25.根据实施方案1至24中任一项所述的crispr复合物,所述crispr复合物还包含被配置为调节所述crispr复合物的活性的序列。
[0467]
26.根据实施方案25所述的crispr复合物,其中所述grna包括包含可裂解接头的crispr on多核苷酸、crispr off多核苷酸或crispr on/off多核苷酸。
[0468]
27.根据实施方案26所述的crispr复合物,其中所述crispr on/off多核苷酸包含crispr on多核苷酸和crispr off多核苷酸。
[0469]
28.根据实施方案26或27所述的crispr复合物,其中所述crispr on多核苷酸包含通过可裂解接头与引导序列的5’末端共价连接的序列元件,其中所述引导序列包含被配置为与靶核酸分子中的靶序列退火的靶结合区。
[0470]
29.根据实施方案26或27所述的crispr复合物,其中所述crispr on多核苷酸包含与引导序列的5’末端共价连接的序列元件,其中所述引导序列包含被配置为与靶核酸分子中的靶序列退火的靶结合区。
[0471]
30.根据实施方案29所述的crispr复合物,其中所述序列元件通过可裂解接头与引导序列的5’末端共价连接。
[0472]
31.根据实施方案26或27所述的crispr复合物,其中所述crispr on多核苷酸包含含有一个或更多个可裂解接头的序列元件,其中所述序列元件不通过可裂解接头与引导序列的5’末端共价连接,并且其中所述引导序列包含被配置为与靶核酸分子中的靶序列退火的靶结合区。
[0473]
32.根据实施方案28至29中任一项所述的crispr复合物,其中所述序列元件包含至少15个核苷酸。
[0474]
33.根据实施方案28至29中任一项所述的crispr复合物,其中所述序列元件包含至少20个核苷酸。
[0475]
34.根据实施方案28至30中任一项所述的crispr复合物,其中所述序列元件包含24个核苷酸。
[0476]
35.根据实施方案29至31中任一项所述的crispr复合物,其中所述序列元件包含rna序列。
[0477]
36.根据实施方案32所述的crispr复合物,其中所述rna序列包含修饰的rna碱基。
[0478]
37.根据实施方案33所述的crispr复合物,其中所述修饰的rna碱基是2'
‑
o
‑
甲基rna碱基。
[0479]
38.根据实施方案28至34中任一项所述的crispr复合物,其中所述序列元件形成包含环的茎环。
[0480]
39.根据实施方案35所述的crispr复合物,其中所述环包含至少两个核苷酸。
[0481]
40.根据实施方案35或36所述的crispr复合物,其中所述环包含至少三个核苷酸。
[0482]
41.根据实施方案35至37中任一项所述的crispr复合物,其中所述环包含四个核苷酸。
[0483]
42.根据实施方案35至38中任一项所述的crispr复合物,其中所述序列元件包含与所述引导序列的碱基配对。
[0484]
43.根据实施方案39所述的crispr复合物,其中所述序列元件包含与所述引导序列的靶结合区的碱基配对。
[0485]
44.根据实施方案40所述的crispr复合物,其中所述序列元件与所述引导序列的靶结合区中的至少10个核苷酸碱基配对。
[0486]
45.根据实施方案40所述的crispr复合物,其中所述序列元件与所述引导序列的靶结合区中的至少15个核苷酸碱基配对。
[0487]
46.根据实施方案40所述的crispr复合物,其中所述序列元件与所述引导序列的靶结合区中的20个核苷酸碱基配对。
[0488]
47.根据实施方案35至38中任一项所述的crispr复合物,其中所述序列元件不包含与所述引导序列的碱基配对。
[0489]
48.根据实施方案44所述的crispr复合物,其中所述序列元件的最5’侧碱基与所
述序列元件中紧接所述引导序列5’侧的碱基退火。
[0490]
49.根据实施方案28至45中任一项所述的crispr复合物,所述crispr复合物还包含位于所述序列元件中的一个或更多个可裂解接头。
[0491]
50.根据实施方案28至45中任一项所述的crispr复合物,所述crispr复合物还包含位于所述序列元件中的至少两个可裂解接头。
[0492]
51.根据实施方案28至45中任一项所述的crispr复合物,所述crispr复合物还包含位于所述序列元件中的至少三个可裂解接头。
[0493]
52.根据实施方案28至45中任一项所述的crispr复合物,所述crispr复合物还包含位于所述序列元件中的至少四个可裂解接头。
[0494]
53.根据实施方案28至45中任一项所述的crispr复合物,所述crispr复合物还包含位于所述序列元件中的至少五个或更多个、七个或更多个、十个或更多个、十五个或更多个或者二十个或更多个可裂解接头。
[0495]
54.根据实施方案35至45中任一项所述的crispr复合物,所述crispr复合物还包含位于所述序列元件的环中的一个或更多个可裂解接头。
[0496]
55.根据实施方案35至45中任一项所述的crispr复合物,所述crispr复合物还包含位于所述序列元件的环中的两个或更多个可裂解接头。
[0497]
56.根据实施方案35至45中任一项所述的crispr复合物,所述crispr复合物还包含位于所述序列元件的环中的三个可裂解接头。
[0498]
57.根据实施方案35至45或51至53中任一项所述的crispr复合物,所述crispr复合物还包含位于所述序列元件的中间位置的核苷酸处的一个或更多个可裂解接头。
[0499]
58.根据实施方案35至45或51至53中任一项所述的crispr复合物,所述crispr复合物还包含位于所述序列元件的中间位置的核苷酸处的两个或更多个可裂解接头。
[0500]
59.根据实施方案35至45或51至53中任一项所述的crispr复合物,所述crispr复合物还包含位于所述序列元件的中间位置的核苷酸处的三个或更多个可裂解接头。
[0501]
60.根据实施方案35至45或51至53中任一项所述的crispr复合物,所述crispr复合物还包含位于所述序列元件的中间位置的核苷酸处的四个或更多个可裂解接头。
[0502]
61.根据实施方案46所述的crispr复合物,其中所述复合物包含在所述序列元件中的位置24处的第一可裂解接头和在所述序列元件中的位置11处的第二可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0503]
62.根据实施方案46所述的crispr复合物,其中所述复合物包含在所述序列元件中的位置24处的第一可裂解接头和在所述序列元件中的以下任一位置处的一个或更多个可裂解接头:5、6、10、11、14、15、16、21、22或23,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0504]
63.根据实施方案46所述的crispr复合物,其中所述复合物包含在所述序列元件中的位置24处的第一可裂解接头和在所述序列元件中的位置23处的第二可裂解接头,以及在所述序列元件中的以下任一位置处的一个或更多个可裂解接头:5、6、10、11、14、15或16,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0505]
64.根据实施方案46所述的crispr复合物,其中所述复合物包含在所述序列元件中的位置24处的可裂解接头和在所述序列元件中的以下任一位置处的第一一个或更多个可裂解接头:21、22或23,以及在所述序列元件中的以下任一位置处的第二一个或更多个可裂解接头:5、6、10、11、14、15或16,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0506]
65.根据实施方案46所述的crispr复合物,其中所述复合物包含在所述序列元件中的位置24处的第一可裂解接头、在所述序列元件中的位置23处的第二可裂解接头和在所述序列元件中的位置3处的第三可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0507]
66.根据实施方案46所述的crispr复合物,其中所述复合物包含在所述序列元件中的位置24处的第一可裂解接头、在所述序列元件中的位置23处的第二可裂解接头和在所述序列元件中的位置3处的第三可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0508]
67.根据实施方案46所述的crispr复合物,其中所述复合物包含在所述序列元件中的位置24处的第一可裂解接头、在所述序列元件中的位置23处的第二可裂解接头、在所述序列元件中的位置6处的第三可裂解接头、在所述序列元件中的位置16处的第四可裂解接头和在所述序列元件中的位置11处的第五可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0509]
68.根据实施方案46所述的crispr复合物,其中所述复合物包含在所述序列元件中的位置24处的第一可裂解接头、在所述序列元件中的位置23处的第二可裂解接头、在所述序列元件中的位置6处的第三可裂解接头和在所述序列元件中的位置14处的第四可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0510]
69.根据实施方案58至65中任一项所述的crispr复合物,其中所述序列元件的长度为24个核苷酸,所述序列元件的环包含从位置21至位置24的核苷酸,并且所述序列元件的从位置1至位置20的核苷酸与所述引导序列的靶结合区碱基配对。
[0511]
70.根据实施方案26或27所述的crispr复合物,其中所述crispr off多核苷酸包含引导序列的最5’侧核苷酸的3’侧的可裂解接头,其中所述引导序列包含被配置为与靶核酸分子中的靶序列退火的靶结合区。
[0512]
71.根据实施方案67所述的crispr复合物,其中所述可裂解接头不位于所述多核苷酸的3’末端处,其中所述多核苷酸以从所述多核苷酸的5’末端至3’末端的顺序包含所述引导序列和包含被配置为与crispr效应蛋白结合的crispr效应蛋白结合区的序列。
[0513]
72.根据实施方案26至68中任一项所述的crispr复合物,其中所述可裂解接头位于所述多核苷酸中紧接核苷酸56或73的3’侧,其中所述引导序列的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述引导序列的5’末端至所述多核苷酸的3’末端的顺序编号。
[0514]
73.根据实施方案26至68中任一项所述的crispr复合物,其中所述多核苷酸包含第一可裂解接头和第二可裂解接头,其中所述第一可裂解接头位于所述多核苷酸中紧接核苷酸56的3’侧,并且所述第二可裂解接头位于所述多核苷酸中紧接核苷酸73的3’侧,其中
所述引导序列的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述引导序列的5’末端至所述多核苷酸的3’末端的顺序编号。
[0515]
74.根据实施方案26至70中任一项所述的crispr复合物,其中所述多核苷酸从5’至3’包含四环、连结、茎环1和茎环2,其中所述可裂解接头位于所述连结的环或所述茎环1的环中。
[0516]
75.根据实施方案26至70中任一项所述的crispr复合物,其中所述多核苷酸从5’至3’包含四环、连结、茎环1和茎环2,其中所述可裂解接头位于所述连结的环和所述茎环1的环中。
[0517]
76.根据实施方案26至72中任一项所述的crispr复合物,其中所述可裂解接头是光不稳定的。
[0518]
77.根据实施方案73所述的crispr复合物,其中所述可裂解接头被紫外(uv)光裂解。
[0519]
78.根据实施方案74所述的crispr复合物,其中所述可裂解接头被波长在100nm至400nm范围内的光裂解。
[0520]
79.根据实施方案73所述的crispr复合物,其中所述可裂解接头被可见光裂解。
[0521]
80.根据实施方案73所述的crispr复合物,其中所述可裂解接头被波长为400nm至700nm的光裂解。
[0522]
81.根据实施方案76所述的crispr复合物,其中所述可裂解接头被绿光裂解。
[0523]
82.根据实施方案76所述的crispr复合物,其中所述可裂解接头被紫光裂解。
[0524]
83.根据实施方案76所述的crispr复合物,其中所述可裂解接头被蓝光裂解。
[0525]
84.根据实施方案76所述的crispr复合物,其中所述可裂解接头被波长在490nm至570nm范围内的光裂解。
[0526]
85.根据实施方案76所述的crispr复合物,其中所述可裂解接头被波长在400nm至420nm范围内的光裂解。
[0527]
86.根据实施方案76所述的crispr复合物,其中所述可裂解接头被波长在420nm至430nm范围内的光裂解。
[0528]
87.根据实施方案76所述的crispr复合物,其中所述可裂解接头被波长在420nm至440nm范围内的光裂解。
[0529]
88.根据实施方案76所述的crispr复合物,其中所述crispr on多核苷酸的可裂解接头被绿光裂解。
[0530]
89.根据实施方案76所述的crispr复合物,其中所述crispr off多核苷酸的可裂解接头被紫光裂解。
[0531]
90.根据实施方案76所述的crispr复合物,其中所述crispr off多核苷酸的可裂解接头被蓝光裂解。
[0532]
91.根据实施方案76所述的crispr复合物,其中所述crispr on多核苷酸的可裂解接头被波长在490nm至570nm范围内的光裂解。
[0533]
92.根据实施方案76所述的crispr复合物,其中所述crispr on多核苷酸的可裂解接头被波长在420nm至430nm范围内的光裂解。
[0534]
93.根据实施方案76所述的crispr复合物,其中所述crispr off多核苷酸的可裂
解接头被波长在400nm至420nm范围内的光裂解。
[0535]
94.根据实施方案76所述的crispr复合物,其中所述crispr off多核苷酸的可裂解接头被波长在420nm至430nm范围内的光裂解。
[0536]
95.根据实施方案76所述的crispr复合物,其中所述crispr off多核苷酸的可裂解接头被波长在420nm至440nm范围内的光裂解。
[0537]
96.根据实施方案76所述的crispr复合物,其中所述crispr on/off多核苷酸的crispr on多核苷酸中的可裂解接头被比所述crispr on/off多核苷酸的crispr off多核苷酸中的可裂解接头更高波长的光裂解。
[0538]
97.根据实施方案93所述的crispr复合物,其中所述crispr on多核苷酸的可裂解接头被绿光裂解,并且所述crispr off多核苷酸的可裂解接头被紫光裂解。
[0539]
98.根据实施方案93所述的crispr复合物,其中所述crispr on多核苷酸的可裂解接头被绿光裂解,并且所述crispr off多核苷酸的可裂解接头被蓝光裂解。
[0540]
99.根据实施方案93所述的crispr复合物,其中所述crispr on多核苷酸的可裂解接头被波长在490nm至570nm范围内的光裂解,并且所述crispr off多核苷酸的可裂解接头被波长在400nm至420nm范围内的光裂解。
[0541]
100.根据实施方案93所述的crispr复合物,其中所述crispr on多核苷酸的可裂解接头被波长在490nm至570nm范围内的光裂解,并且所述crispr off多核苷酸的可裂解接头被波长在420nm至430nm范围内的光裂解。
[0542]
101.根据实施方案93所述的crispr复合物,其中所述crispr on多核苷酸的可裂解接头被波长在490nm至570nm范围内的光裂解,并且所述crispr off多核苷酸的可裂解接头被波长在420nm至440nm范围内的光裂解。
[0543]
102.根据实施方案26至98中任一项所述的crispr复合物,其中所述可裂解接头是亚磷酰胺衍生物。
[0544]
103.根据实施方案26至98中任一项所述的crispr复合物,其中所述可裂解接头是3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺衍生物。
[0545]
104.根据实施方案26至98中任一项所述的crispr复合物,其中所述可裂解接头是1
‑
(7
‑
(二乙基氨基)
‑2‑
氧代
‑
2h
‑
色烯
‑4‑
基)
‑3‑
(4,4'
‑
二甲氧基三苯甲基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]亚磷酰胺衍生物。
[0546]
105.根据实施方案26至98中任一项所述的crispr复合物,其中所述可裂解接头包括磷酸二酯。
[0547]
106.根据实施方案26至98中任一项所述的crispr复合物,其中所述可裂解接头包括磷酸单酯。
[0548]
107.根据实施方案26至98中任一项所述的crispr复合物,其中所述可裂解接头是香豆素衍生物。
[0549]
108.根据实施方案26至98中任一项所述的crispr复合物,其中所述可裂解接头包括由以下式表示的结构:
[0550][0551]
其中:
[0552]
x是o、s或cr
x
r
y
,其中r
x
和r
y
独立地选自任选地取代的烷基、任选地取代的烯基、任选地取代的炔基、任选地取代的杂烷基、卤素、卤代烷基、烷氧基、烷氧基烷基、卤代烷氧基、氨基、氨基烷基、烷基氨基、烷基氨基烷基、羟基、羟基烷基、c
‑
羧基、o
‑
羧基、酰基、硫醇、烷硫基、硫代烷基、c
‑
酰胺基、n
‑
酰胺基、脲基、硝基、氰基、磺酰基、磺基、磺酸酯、亚磺基、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;并且
[0553]
*表示与h或第一核苷酸的附接点;并且
[0554]
**表示与oh或第二核苷酸的附接点。
[0555]
109.根据实施方案105所述的crispr复合物,其中x是氧。
[0556]
110.根据实施方案105所述的crispr复合物,其中x是硫。
[0557]
111.根据实施方案105所述的crispr复合物,其中x是=c(cn)2。
[0558]
112.根据实施方案26至98中任一项所述的crispr复合物,其中所述可裂解接头包括由式(i)表示的结构:
[0559][0560]
其中:
[0561]
r1、r2、r3、r4和r5各自独立地选自h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0562]
可选地,r1、r2、r3和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0563]
可选地,r1、r2、r3、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任
选地取代的c5‑
10
碳环;并且
[0564]
m是选自1至10的整数;并且
[0565]
x是o、s或cr
x
r
y
,其中r
x
和r
y
独立地选自任选地取代的烷基、任选地取代的烯基、任选地取代的炔基、任选地取代的杂烷基、卤素、卤代烷基、烷氧基、烷氧基烷基、卤代烷氧基、氨基、氨基烷基、烷基氨基、烷基氨基烷基、羟基、羟基烷基、c
‑
羧基、o
‑
羧基、酰基、硫醇、烷硫基、硫代烷基、c
‑
酰胺基、n
‑
酰胺基、脲基、硝基、氰基、磺酰基、磺基、磺酸酯、亚磺基、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基,
[0566]
条件是,当x是o,并且r3是二烷基氨基时,则m是2至10的整数;并且
[0567]
*表示与h或第一核苷酸的附接点;并且
[0568]
**表示与oh或第二核苷酸的附接点。
[0569]
113.根据实施方案109所述的crispr复合物,其中x是氧。
[0570]
114.根据实施方案109所述的crispr复合物,其中x是硫。
[0571]
115.根据实施方案109所述的crispr复合物,其中x是=c(cn)2。
[0572]
116.根据实施方案109所述的crispr复合物,其中所述式(i)的结构由式(i’)表示:
[0573][0574]
其中:
[0575]
r1、r2、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0576]
r
3a
和r
3b
独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0577]
可选地,r1、r2、r
3a
、r
3b
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0578]
可选地,r1、r2、r
3a
、r
3b
、r4和r5中的两个或更多个连同它们所附接的原子一起形成
选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0579]
x是氧。
[0580]
117.根据实施方案109所述的crispr复合物,其中所述式(i)的结构由式(i’)表示:
[0581][0582]
其中:
[0583]
r1、r2、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0584]
r
3a
和r
3b
独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0585]
可选地,r1、r2、r
3a
、r
3b
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0586]
可选地,r1、r2、r
3a
、r
3b
、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0587]
x是硫。
[0588]
118.根据实施方案109所述的crispr复合物,其中所述式(i)的结构由式(i’)表示:
[0589][0590]
其中:
[0591]
r1、r2、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0592]
r
3a
和r
3b
独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0593]
可选地,r1、r2、r
3a
、r
3b
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0594]
可选地,r1、r2、r
3a
、r
3b
、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0595]
x是=c(cn)2。
[0596]
119.根据实施方案113至115中任一项所述的crispr复合物,其中:
[0597]
r1、r2、r4和r5各自独立地是h或c1‑6烷基;并且
[0598]
r
3a
和r
3b
是c1‑6烷基。
[0599]
120.根据实施方案113至115中任一项所述的crispr复合物,其中:
[0600]
r1、r2、r4和r5各自是h;并且
[0601]
r
3a
和r
3b
各自是乙基。
[0602]
121.根据实施方案1至117中任一项所述的crispr复合物,其中当所述grna与crispr效应蛋白复合时,形成第一交联的crispr复合物,与包含不与所述crispr效应蛋白交联的grna的第二crispr复合物相比,所述第一交联的crispr复合物对脱靶核酸分子具有更低的编辑活性。
[0603]
122.根据实施方案118所述的crispr复合物,其中所述编辑活性以被编辑的脱靶核酸分子的百分比来测量。
[0604]
123.根据实施方案118所述的crispr复合物,其中所述编辑活性以被编辑的靶核
酸分子的百分比来测量。
[0605]
124.根据实施方案118至120中任一项所述的crispr复合物,其中所述第一crispr复合物对所述脱靶核酸分子的编辑活性低于所述第二crispr复合物的编辑活性,其中p值≤0.0001。
[0606]
125.根据实施方案118至121中任一项所述的crispr复合物,其中所述第一crispr复合物对所述靶核酸分子的编辑活性和所述第二crispr复合物对所述靶核酸分子的编辑活性在5%以内。
[0607]
126.根据实施方案1至122中任一项所述的crispr复合物,其中所述crispr效应蛋白是ii型crispr效应蛋白。
[0608]
127.根据实施方案123所述的crispr复合物,其中所述ii型crispr效应蛋白是cas9多肽。
[0609]
128.根据实施方案1至122中任一项所述的crispr复合物,其中所述crispr效应蛋白是v型crispr效应蛋白。
[0610]
129.根据实施方案125所述的crispr复合物,其中所述v型crispr效应蛋白是cas12a、cas12b、cas12c、cas12d、cas12e、cas12f、cas12g、cas12h或cas12i多肽。
[0611]
130.根据实施方案1至122中任一项所述的crispr复合物,其中所述crispr效应蛋白是vi型crispr效应蛋白。
[0612]
131.根据实施方案127所述的crispr复合物,其中所述vi型crispr效应蛋白是cas13a、cas13b、cas13c或cas13d多肽。
[0613]
132.根据实施方案1至122中任一项所述的crispr复合物,其中所述crispr效应蛋白是cas14a、cas14b或cas14c多肽。
[0614]
133.一种单引导rna(sgrna),所述单引导rna(sgrna)包含crrna区和tracrrna区以及核苷酸位置49处的核苷酸,其中核苷酸位置1位于所述crrna区的靶结合区的5’末端处,并且所述sgrna的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。
[0615]
134.一种单引导rna(sgrna),所述单引导rna(sgrna)包含crrna区和tracrrna区以及核苷酸位置49处的尿嘧啶,其中核苷酸位置1位于所述crrna区的靶结合区的5’末端处,并且所述sgrna的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。
[0616]
135.一种多核苷酸,所述多核苷酸包含:(i)包含被配置为与靶核酸分子中的靶序列退火的靶结合区的引导序列,(ii)包含被配置为与crispr效应蛋白结合的crispr效应蛋白结合区的序列,以及(iii)被配置为与crispr效应蛋白交联的核苷酸,其中所述核苷酸位于所述引导序列的靶结合区之外。
[0617]
136.根据实施方案132所述的多核苷酸,其中所述核苷酸包含尿嘧啶。
[0618]
137.根据实施方案132或133所述的多核苷酸,其中所述核苷酸位于所述多核苷酸的核苷酸位置49处,其中核苷酸位置1位于所述引导序列的靶结合区的5’末端处,并且所述多核苷酸的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。
[0619]
138.根据实施方案132至134中任一项所述的多核苷酸,其中所述核苷酸位于所述多核苷酸的以下一个或更多个核苷酸位置处:22、23、24、25、31、37、44、49、45、50、56、59、63、64、66、71、72、77、78、80、84、90和94,其中核苷酸位置1位于所述引导序列的靶结合区的5’末端处,并且所述多核苷酸的核苷酸位置从核苷酸位置1开始从5’至3’连续编号。
[0620]
139.根据实施方案132至135中任一项所述的多核苷酸,其中所述核苷酸是非天然核苷酸。
[0621]
140.根据实施方案136所述的多核苷酸,其中所述非天然核苷酸包含糖的修饰。
[0622]
141.根据实施方案136或137所述的多核苷酸,其中所述非天然核苷酸包含碱基的修饰。
[0623]
142.根据实施方案136所述的多核苷酸,其中所述非天然核苷酸包含马来酰亚胺。
[0624]
143.根据实施方案139所述的多核苷酸,其中所述马来酰亚胺与所述crispr效应蛋白上的半胱氨酸共价连接。
[0625]
144.根据实施方案132至140中任一项所述的多核苷酸,其中所述多核苷酸通过交联剂与所述crispr效应蛋白交联,其中所述交联剂包括或衍生自吡啶基二硫化物、烷氧基胺、nhs酯、双吖丙啶、亚氨酸酯、卤代乙酰基基团、酰肼、芳基叠氮化物、异氰酸酯、二硫醇亚磷酰胺dtpa、4
‑
硫代
‑
utp、5
‑
叠氮基
‑
utp、5
‑
溴
‑
utp、8
‑
叠氮基
‑
atp、5
‑
apas
‑
utp或8
‑
n(3)amp。
[0626]
145.根据实施方案132至140中任一项所述的多核苷酸,其中所述多核苷酸通过交联剂与所述crispr效应蛋白交联,其中所述交联剂包括二硫化物、酰胺、亚胺、酰肼、o
‑
烷基肟、烷基、胺、醇、三唑、异噁唑啉、异噁唑烷、异噁唑或哒嗪。
[0627]
146.根据实施方案132至142中任一项所述的多核苷酸,其中(ii)的所述序列从5’至3’形成四环、连结、第一茎环和第二茎环。
[0628]
147.根据实施方案132或142所述的多核苷酸,其中所述多核苷酸不包含第三茎环。
[0629]
148.根据实施方案132至144中任一项所述的多核苷酸,其中所述多核苷酸在所述多核苷酸的5’末端处不包含茎环。
[0630]
149.根据实施方案142至145中任一项所述的多核苷酸,其中所述核苷酸位于茎环中。
[0631]
150.根据实施方案146所述的多核苷酸,其中所述茎环的结构相对于缺少所述核苷酸的多核苷酸的茎环的结构被维持。
[0632]
151.根据实施方案142至147中任一项所述的多核苷酸,其中所述核苷酸位于所述四环中。
[0633]
152.根据实施方案148所述的多核苷酸,其中所述四环的结构相对于缺少所述核苷酸的多核苷酸的四环的结构被维持。
[0634]
153.根据实施方案148或149所述的多核苷酸,其中所述四环包含凸起。
[0635]
154.根据实施方案150所述的多核苷酸,其中所述核苷酸位于所述凸起中。
[0636]
155.根据实施方案151所述的多核苷酸,其中所述凸起的结构相对于缺少所述核苷酸的多核苷酸的凸起的结构被维持。
[0637]
156.根据实施方案142至152中任一项所述的多核苷酸,其中所述核苷酸位于茎环之间。
[0638]
157.根据实施方案132至153中任一项所述的多核苷酸,其中所述核苷酸位于所述crispr效应蛋白的半胱氨酸的20埃以内。
[0639]
158.根据实施方案132至154中任一项所述的多核苷酸,其中所述核苷酸不是4
‑
硫
代尿苷或修饰的腺苷。
[0640]
159.根据实施方案132至155中任一项所述的多核苷酸,其中所述多核苷酸包含至少两个被配置为与crispr效应蛋白交联的核苷酸。
[0641]
160.根据实施方案132至156中任一项所述的多核苷酸,所述多核苷酸还包含被配置为调节所述crispr效应蛋白的活性的序列。
[0642]
161.根据实施方案157所述的多核苷酸,其中所述多核苷酸包括包含可裂解接头的crispr on多核苷酸、crispr off多核苷酸或crispr on/off多核苷酸。
[0643]
162.根据实施方案158所述的多核苷酸,其中所述crispr on/off多核苷酸包含crispr on多核苷酸和crispr off多核苷酸。
[0644]
163.根据实施方案158或159所述的多核苷酸,其中所述crispr on多核苷酸包含通过可裂解接头与所述引导序列的5’末端共价连接的序列元件。
[0645]
164.根据实施方案158或159所述的多核苷酸,其中所述crispr on多核苷酸包含与引导序列的5’末端共价连接的序列元件,其中所述引导序列包含被配置为与靶核酸分子中的靶序列退火的靶结合区。
[0646]
165.根据实施方案158或159所述的多核苷酸,其中所述序列元件通过可裂解接头与引导序列的5’末端共价连接。
[0647]
166.根据实施方案158或159所述的多核苷酸,其中所述crispr on多核苷酸包含含有一个或更多个可裂解接头的序列元件,其中所述序列元件不通过可裂解接头与引导序列的5’末端共价连接,并且其中所述引导序列包含被配置为与靶核酸分子中的靶序列退火的靶结合区。
[0648]
167.根据实施方案160至166中任一项所述的多核苷酸,其中所述序列元件包含至少15个核苷酸。
[0649]
168.根据实施方案160至166中任一项所述的多核苷酸,其中所述序列元件包含至少20个核苷酸。
[0650]
169.根据实施方案160至165中任一项所述的多核苷酸,其中所述序列元件包含24个核苷酸。
[0651]
170.根据实施方案160至166中任一项所述的多核苷酸,其中所述序列元件包括rna序列。
[0652]
171.根据实施方案167所述的多核苷酸,其中所述rna序列包含修饰的rna碱基。
[0653]
172.根据实施方案168所述的多核苷酸,其中所述修饰的rna碱基是2'
‑
o
‑
甲基rna碱基。
[0654]
173.根据实施方案160至169中任一项所述的多核苷酸,其中所述序列元件形成包含环的茎环。
[0655]
174.根据实施方案170所述的多核苷酸,其中所述环包含至少两个核苷酸。
[0656]
175.根据实施方案170或171所述的多核苷酸,其中所述环包含至少三个核苷酸。
[0657]
176.根据实施方案170至172中任一项所述的多核苷酸,其中所述环包含四个核苷酸。
[0658]
177.根据实施方案170至173中任一项所述的多核苷酸,其中所述序列元件包含与所述引导序列的碱基配对。
[0659]
178.根据实施方案174所述的多核苷酸,其中所述序列元件包含与所述引导序列的靶结合区的碱基配对。
[0660]
179.根据实施方案175所述的多核苷酸,其中所述序列元件与所述引导序列的靶结合区中的至少10个核苷酸碱基配对。
[0661]
180.根据实施方案175所述的多核苷酸,其中所述序列元件与所述引导序列的靶结合区中的至少15个核苷酸碱基配对。
[0662]
181.根据实施方案175所述的多核苷酸,其中所述序列元件与所述引导序列的靶结合区中的20个核苷酸碱基配对。
[0663]
182.根据实施方案170至173中任一项所述的多核苷酸,其中所述序列元件不包含与所述引导序列的碱基配对。
[0664]
183.根据实施方案179所述的多核苷酸,其中所述序列元件的最5’侧碱基与所述序列元件中紧接所述引导序列5’侧的碱基退火。
[0665]
184.根据实施方案160至180中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件中的一个或更多个可裂解接头。
[0666]
185.根据实施方案160至180中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件中的至少两个可裂解接头。
[0667]
186.根据实施方案160至180中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件中的至少三个可裂解接头。
[0668]
187.根据实施方案160至180中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件中的至少四个可裂解接头。
[0669]
188.根据实施方案160至180中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件中的至少五个或更多个、七个或更多个、十个或更多个、十五个或更多个或者二十个或更多个可裂解接头。
[0670]
189.根据实施方案170至180中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的环中的一个或更多个可裂解接头。
[0671]
190.根据实施方案170至180中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的环中的两个或更多个可裂解接头。
[0672]
191.根据实施方案170至180中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的环中的三个可裂解接头。
[0673]
192.根据实施方案170至180或186至188中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的中间位置的核苷酸处的一个或更多个可裂解接头。
[0674]
193.根据实施方案170至180或186至188中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的中间位置的核苷酸处的两个或更多个可裂解接头。
[0675]
194.根据实施方案170至180或186至188中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的中间位置的核苷酸处的三个或更多个可裂解接头。
[0676]
195.根据实施方案170至180或186至188中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的中间位置的核苷酸处的四个或更多个可裂解接头。
[0677]
196.根据实施方案181所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头和在所述序列元件中的位置11处的第二可裂解接头,其中
所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0678]
197.根据实施方案181所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头和在所述序列元件中的以下任一位置处的一个或更多个可裂解接头:5、6、10、11、14、15、16、21、22或23,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0679]
198.根据实施方案181所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头和在所述序列元件中的位置23处的第二可裂解接头,以及在所述序列元件中的以下任一位置处的一个或更多个可裂解接头:5、6、10、11、14、15或16,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0680]
199.根据实施方案181所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的可裂解接头和在所述序列元件中的以下任一位置处的第一一个或更多个可裂解接头:21、22或23,以及在所述序列元件中的以下任一位置处的第二一个或更多个可裂解接头:5、6、10、11、14、15或16,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0681]
200.根据实施方案181所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头、在所述序列元件中的位置23处的第二可裂解接头和在所述序列元件中的位置3处的第三可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0682]
201.根据实施方案181所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头、在所述序列元件中的位置23处的第二可裂解接头和在所述序列元件中的位置3处的第三可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0683]
202.根据实施方案181所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头、在所述序列元件中的位置23处的第二可裂解接头、在所述序列元件中的位置6处的第三可裂解接头、在所述序列元件中的位置16处的第四可裂解接头和在所述序列元件中的位置11处的第五可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0684]
203.根据实施方案181所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头、在所述序列元件中的位置23处的第二可裂解接头、在所述序列元件中的位置6处的第三可裂解接头和在所述序列元件中的位置14处的第四可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0685]
204.根据实施方案193至200中任一项所述的多核苷酸,其中所述序列元件的长度为24个核苷酸,所述序列元件的环包含从位置21至位置24的核苷酸,并且所述序列元件的从位置1至位置20的核苷酸与所述引导序列的靶结合区碱基配对。
[0686]
205.一种crispr on多核苷酸,所述crispr on多核苷酸包含通过可裂解接头与引
导序列的5’末端共价连接的序列元件。
[0687]
206.一种crispr on多核苷酸,所述crispr on多核苷酸包含与引导序列的5’末端共价连接的序列元件,其中所述引导序列包含被配置为与靶核酸分子中的靶序列退火的靶结合区。
[0688]
207.根据实施方案206所述的多核苷酸,其中所述序列元件通过可裂解接头与引导序列的5’末端共价连接。
[0689]
208.一种crispr on多核苷酸,所述crispr on多核苷酸包含含有一个或更多个可裂解接头的序列元件,其中所述序列元件不通过可裂解接头与引导序列的5’末端共价连接,并且其中所述引导序列包含被配置为与靶核酸分子中的靶序列退火的靶结合区。
[0690]
209.根据实施方案202至208中任一项所述的多核苷酸,其中所述序列元件包括rna序列。
[0691]
210.根据实施方案209所述的多核苷酸,其中所述rna序列包含修饰的rna碱基。
[0692]
211.一种crispr on多核苷酸,所述crispr on多核苷酸包含通过可裂解接头与引导序列的5’末端共价连接的序列元件,其中所述引导序列包含被配置为与靶核酸分子中的靶序列退火的靶结合区,并且所述序列元件包含修饰的rna碱基。
[0693]
212.根据实施方案210或211所述的多核苷酸,其中所述修饰的rna碱基是2'
‑
o
‑
甲基rna碱基。
[0694]
213.根据实施方案202至209中任一项所述的多核苷酸,其中所述序列元件包含至少15个核苷酸。
[0695]
214.根据实施方案202至210中任一项所述的多核苷酸,其中所述序列元件包含至少20个核苷酸。
[0696]
215.根据实施方案202至211中任一项所述的多核苷酸,其中所述序列元件包含24个核苷酸。
[0697]
216.根据实施方案202至212中任一项所述的多核苷酸,其中所述序列元件形成包含环的茎环。
[0698]
217.根据实施方案213所述的多核苷酸,其中所述环包含至少两个核苷酸。
[0699]
218.根据实施方案213或214所述的多核苷酸,其中所述环包含至少三个核苷酸。
[0700]
219.根据实施方案213至215中任一项所述的多核苷酸,其中所述环包含四个核苷酸。
[0701]
220.根据实施方案213至216中任一项所述的多核苷酸,其中所述序列元件包含与所述引导序列的碱基配对。
[0702]
221.根据实施方案217所述的多核苷酸,其中所述序列元件包含与所述引导序列的靶结合区的碱基配对。
[0703]
222.根据实施方案218所述的多核苷酸,其中所述序列元件与所述引导序列的靶结合区中的至少10个核苷酸碱基配对。
[0704]
223.根据实施方案218所述的多核苷酸,其中所述序列元件与所述引导序列的靶结合区中的至少15个核苷酸碱基配对。
[0705]
224.根据实施方案218所述的多核苷酸,其中所述序列元件与所述引导序列的靶结合区中的20个核苷酸碱基配对。
[0706]
225.根据实施方案213至216中任一项所述的多核苷酸,其中所述序列元件不包含与所述引导序列的碱基配对。
[0707]
226.根据实施方案222所述的多核苷酸,其中所述序列元件的最5’侧碱基与所述序列元件中紧接所述引导序列5’侧的碱基退火。
[0708]
227.根据实施方案202至223中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件中的一个或更多个可裂解接头。
[0709]
228.根据实施方案202至223中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件中的至少两个可裂解接头。
[0710]
229.根据实施方案202至223中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件中的至少三个可裂解接头。
[0711]
230.根据实施方案202至223中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件中的至少四个可裂解接头。
[0712]
231.根据实施方案202至223中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件中的至少五个或更多个、七个或更多个、十个或更多个、十五个或更多个或者二十个或更多个可裂解接头。
[0713]
232.根据实施方案202至223中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的环中的一个或更多个可裂解接头。
[0714]
233.根据实施方案202至223中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的环中的两个或更多个可裂解接头。
[0715]
234.根据实施方案202至223中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的环中的三个可裂解接头。
[0716]
235.根据实施方案202至223或229至232中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的中间位置的核苷酸处的一个或更多个可裂解接头。
[0717]
236.根据实施方案202至223或229至232中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的中间位置的核苷酸处的两个或更多个可裂解接头。
[0718]
237.根据实施方案202至223或229至232中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的中间位置的核苷酸处的三个或更多个可裂解接头。
[0719]
238.根据实施方案202至223或229至232中任一项所述的多核苷酸,所述多核苷酸还包含位于所述序列元件的中间位置的核苷酸处的四个或更多个可裂解接头。
[0720]
239.根据实施方案224所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头和在所述序列元件中的位置11处的第二可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0721]
240.根据实施方案224所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头和在所述序列元件中的以下任一位置处的一个或更多个可裂解接头:5、6、10、11、14、15、16、21、22或23,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0722]
241.根据实施方案224所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头和在所述序列元件中的位置23处的第二可裂解接头,以及
在所述序列元件中的以下任一位置处的一个或更多个可裂解接头:5、6、10、11、14、15或16,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0723]
242.根据实施方案224所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的可裂解接头和在所述序列元件中的以下任一位置处的第一一个或更多个可裂解接头:21、22或23,以及在所述序列元件中的以下任一位置处的第二一个或更多个可裂解接头:5、6、10、11、14、15或16,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0724]
243.根据实施方案224所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头、在所述序列元件中的位置23处的第二可裂解接头和在所述序列元件中的位置3处的第三可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0725]
244.根据实施方案224所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头、在所述序列元件中的位置23处的第二可裂解接头和在所述序列元件中的位置3处的第三可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0726]
245.根据实施方案224所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头、在所述序列元件中的位置23处的第二可裂解接头、在所述序列元件中的位置6处的第三可裂解接头、在所述序列元件中的位置16处的第四可裂解接头和在所述序列元件中的位置11处的第五可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0727]
246.根据实施方案224所述的多核苷酸,其中所述多核苷酸包含在所述序列元件中的位置24处的第一可裂解接头、在所述序列元件中的位置23处的第二可裂解接头、在所述序列元件中的位置6处的第三可裂解接头和在所述序列元件中的位置14处的第四可裂解接头,其中所述序列元件的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述序列元件的5’末端至所述序列元件的3’末端的顺序编号。
[0728]
247.根据实施方案236至243中任一项所述的多核苷酸,其中所述序列元件的长度为24个核苷酸,所述序列元件的环包含从位置21至位置24的核苷酸,并且所述序列元件的从位置1至位置20的核苷酸与所述引导序列的靶结合区碱基配对。
[0729]
248.一种crispr off多核苷酸,所述crispr off多核苷酸包含引导序列的最5’侧核苷酸的3’侧的可裂解接头。
[0730]
249.根据实施方案245所述的多核苷酸,其中所述可裂解接头不位于所述多核苷酸的3’末端处。
[0731]
250.根据实施方案158至246中任一项所述的多核苷酸,其中所述可裂解接头位于crispr效应蛋白结合区中。
[0732]
251.根据实施方案158至247中任一项所述的多核苷酸,其中所述可裂解接头位于所述多核苷酸中紧接核苷酸56或73的3’侧,其中所述引导序列的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述引导序列的5’末端至所述多核苷酸的3’末端的顺序编号。
[0733]
252.根据实施方案158至248中任一项所述的多核苷酸,其中所述多核苷酸包含第一可裂解接头和第二可裂解接头,其中所述第一可裂解接头位于所述多核苷酸中紧接核苷酸56的3’侧,并且所述第二可裂解接头位于所述多核苷酸中紧接核苷酸73的3’侧,其中所述引导序列的5’末端处的核苷酸是核苷酸1,并且核苷酸以从所述引导序列的5’末端至所述多核苷酸的3’末端的顺序编号。
[0734]
253.根据实施方案158至249中任一项所述的多核苷酸,其中所述多核苷酸从5’至3’包含四环、连结、茎环1和茎环2,其中所述可裂解接头位于所述连结的环或所述茎环1的环中。
[0735]
254.根据实施方案158至249中任一项所述的多核苷酸,其中所述多核苷酸从5’至3’包含四环、连结、茎环1和茎环2,其中所述可裂解接头位于所述连结的环和所述茎环1的环中。
[0736]
255.一种crispr on/off多核苷酸,所述crispr on/off多核苷酸包含实施方案202至244中任一项所述的crispr on多核苷酸和实施方案245至251中任一项所述的crispr off多核苷酸。
[0737]
256.根据实施方案158至252中任一项所述的多核苷酸,其中所述可裂解接头是光不稳定的。
[0738]
257.根据实施方案253所述的多核苷酸,其中所述可裂解接头被紫外(uv)光裂解。
[0739]
258.根据实施方案254所述的多核苷酸,其中所述可裂解接头被波长在100nm至400nm范围内的光裂解。
[0740]
259.根据实施方案253所述的多核苷酸,其中所述可裂解接头被可见光裂解。
[0741]
260.根据实施方案253所述的多核苷酸,其中所述可裂解接头被波长为400nm至700nm的光裂解。
[0742]
261.根据实施方案256所述的多核苷酸,其中所述可裂解接头被绿光裂解。
[0743]
262.根据实施方案256所述的多核苷酸,其中所述可裂解接头被紫光裂解。
[0744]
263.根据实施方案256所述的多核苷酸,其中所述可裂解接头被蓝光裂解。
[0745]
264.根据实施方案256所述的多核苷酸,其中所述可裂解接头被波长在490nm至570nm范围内的光裂解。
[0746]
265.根据实施方案256所述的多核苷酸,其中所述可裂解接头被波长在400nm至420nm范围内的光裂解。
[0747]
266.根据实施方案256所述的多核苷酸,其中所述可裂解接头被波长在420nm至430nm范围内的光裂解。
[0748]
267.根据实施方案256所述的多核苷酸,其中所述可裂解接头被波长在420nm至440nm范围内的光裂解。
[0749]
268.根据实施方案256所述的多核苷酸,其中所述crispr on多核苷酸的可裂解接头被绿光裂解。
[0750]
269.根据实施方案256所述的多核苷酸,其中所述crispr off多核苷酸的可裂解接头被紫光裂解。
[0751]
270.根据实施方案256所述的多核苷酸,其中所述crispr off多核苷酸的可裂解接头被蓝光裂解。
[0752]
271.根据实施方案256所述的多核苷酸,其中所述crispr on多核苷酸的可裂解接头被波长在490nm至570nm范围内的光裂解。
[0753]
272.根据实施方案256所述的多核苷酸,其中所述crispr on多核苷酸的可裂解接头被波长在420nm至430nm范围内的光裂解。
[0754]
273.根据实施方案256所述的多核苷酸,其中所述crispr off多核苷酸的可裂解接头被波长在400nm至420nm范围内的光裂解。
[0755]
274.根据实施方案256所述的多核苷酸,其中所述crispr off多核苷酸的可裂解接头被波长在420nm至430nm范围内的光裂解。
[0756]
275.根据实施方案256所述的多核苷酸,其中所述crispr off多核苷酸的可裂解接头被波长在420nm至440nm范围内的光裂解。
[0757]
276.根据实施方案256所述的多核苷酸,其中所述crispr on/off多核苷酸的crispr on多核苷酸中的可裂解接头被比所述crispr on/off多核苷酸的crispr off多核苷酸中的可裂解接头更高波长的光裂解。
[0758]
277.根据实施方案268所述的多核苷酸,其中所述crispr on多核苷酸的可裂解接头被波长在490nm至570nm范围内的光裂解,并且所述crispr off多核苷酸的可裂解接头被波长在400nm至420nm范围内的光裂解。
[0759]
278.根据实施方案268所述的多核苷酸,其中所述crispr on多核苷酸的可裂解接头被波长在490nm至570nm范围内的光裂解,并且所述crispr off多核苷酸的可裂解接头被波长在420nm至430nm范围内的光裂解。
[0760]
279.根据实施方案268所述的多核苷酸,其中所述crispr on多核苷酸的可裂解接头被波长在490nm至570nm范围内的光裂解,并且所述crispr off多核苷酸的可裂解接头被波长在420nm至440nm范围内的光裂解。
[0761]
280.根据实施方案268所述的多核苷酸,其中所述crispr on多核苷酸的可裂解接头被绿光裂解,并且所述crispr off多核苷酸的可裂解接头被紫光裂解。
[0762]
281.根据实施方案268所述的多核苷酸,其中所述crispr on多核苷酸的可裂解接头被绿光裂解,并且所述crispr off多核苷酸的可裂解接头被蓝光裂解。
[0763]
282.根据实施方案158至278中任一项所述的多核苷酸,其中所述可裂解接头是亚磷酰胺衍生物。
[0764]
283.根据实施方案158至278中任一项所述的多核苷酸,其中所述可裂解接头是3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺衍生物。
[0765]
284.根据实施方案158至278中任一项所述的多核苷酸,其中所述可裂解接头是1
‑
(7
‑
(二乙基氨基)
‑2‑
氧代
‑
2h
‑
色烯
‑4‑
基)
‑3‑
(4,4'
‑
二甲氧基三苯甲基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]亚磷酰胺衍生物。
[0766]
285.根据实施方案158至278中任一项所述的多核苷酸,其中所述可裂解接头包括磷酸二酯。
[0767]
286.根据实施方案158至278中任一项所述的多核苷酸,其中所述可裂解接头包括磷酸单酯。
[0768]
287.根据实施方案158至278中任一项所述的多核苷酸,其中所述可裂解接头是香
豆素衍生物。
[0769]
288.根据实施方案158至278中任一项所述的多核苷酸,其中所述可裂解接头包括由以下式表示的结构:
[0770][0771]
其中:
[0772]
x是o、s或cr
x
r
y
,其中r
x
和r
y
独立地选自任选地取代的烷基、任选地取代的烯基、任选地取代的炔基、任选地取代的杂烷基、卤素、卤代烷基、烷氧基、烷氧基烷基、卤代烷氧基、氨基、氨基烷基、烷基氨基、烷基氨基烷基、羟基、羟基烷基、c
‑
羧基、o
‑
羧基、酰基、硫醇、烷硫基、硫代烷基、c
‑
酰胺基、n
‑
酰胺基、脲基、硝基、氰基、磺酰基、磺基、磺酸酯、亚磺基、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;并且
[0773]
*表示与h或第一核苷酸的附接点;并且
[0774]
**表示与oh或第二核苷酸的附接点。
[0775]
289.根据实施方案285所述的多核苷酸,其中x是氧。
[0776]
290.根据实施方案285所述的多核苷酸,其中x是硫。
[0777]
291.根据实施方案285所述的多核苷酸,其中x是=c(cn)2。
[0778]
292.根据实施方案158至278中任一项所述的多核苷酸,其中所述可裂解接头包括由式(i)表示的结构:
[0779][0780]
其中:
[0781]
r1、r2、r3、r4和r5各自独立地选自h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0782]
可选地,r1、r2、r3和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选
地取代的c5‑
10
碳环;
[0783]
可选地,r1、r2、r3、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0784]
m是选自1至10的整数;并且
[0785]
x是o、s或cr
x
r
y
,其中r
x
和r
y
独立地选自任选地取代的烷基、任选地取代的烯基、任选地取代的炔基、任选地取代的杂烷基、卤素、卤代烷基、烷氧基、烷氧基烷基、卤代烷氧基、氨基、氨基烷基、烷基氨基、烷基氨基烷基、羟基、羟基烷基、c
‑
羧基、o
‑
羧基、酰基、硫醇、烷硫基、硫代烷基、c
‑
酰胺基、n
‑
酰胺基、脲基、硝基、氰基、磺酰基、磺基、磺酸酯、亚磺基、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基,
[0786]
条件是,当x是o,并且r3是二烷基氨基时,则m是2至10的整数;并且
[0787]
*表示与h或第一核苷酸的附接点;并且
[0788]
**表示与oh或第二核苷酸的附接点。
[0789]
293.根据实施方案289所述的多核苷酸,其中x是氧。
[0790]
294.根据实施方案289所述的多核苷酸,其中x是硫。
[0791]
295.根据实施方案289所述的多核苷酸,其中x是=c(cn)2。
[0792]
296.根据实施方案289所述的多核苷酸,其中所述式(i)的结构由式(i’)表示:
[0793][0794]
其中:
[0795]
r1、r2、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0796]
r
3a
和r
3b
独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0797]
可选地,r1、r2、r
3a
、r
3b
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和
任选地取代的c5‑
10
碳环;
[0798]
可选地,r1、r2、r
3a
、r
3b
、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0799]
x是氧。
[0800]
297.根据实施方案289所述的多核苷酸,其中所述式(i)的结构由式(i’)表示:
[0801][0802]
其中:
[0803]
r1、r2、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0804]
r
3a
和r
3b
独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0805]
可选地,r1、r2、r
3a
、r
3b
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0806]
可选地,r1、r2、r
3a
、r
3b
、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0807]
x是硫。
[0808]
298.根据实施方案289所述的多核苷酸,其中所述式(i)的结构由式(i’)表示:
[0809][0810]
其中:
[0811]
r1、r2、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0812]
r
3a
和r
3b
独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0813]
可选地,r1、r2、r
3a
、r
3b
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0814]
可选地,r1、r2、r
3a
、r
3b
、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0815]
x是=c(cn)2。
[0816]
299.根据实施方案293至296中任一项所述的多核苷酸,其中:
[0817]
r1、r2、r4和r5各自独立地是h或c1‑6烷基;并且
[0818]
r
3a
和r
3b
是c1‑6烷基。
[0819]
300.根据实施方案293至296中任一项所述的多核苷酸,其中:
[0820]
r1、r2、r4和r5各自是h;并且
[0821]
r
3a
和r
3b
各自是乙基。
[0822]
301.根据实施方案132至297中任一项所述的多核苷酸,其中当所述多核苷酸与crispr效应蛋白复合时,形成第一交联的crispr复合物,与包含不与所述crispr效应蛋白交联的多核苷酸的第二crispr复合物相比,所述第一交联的crispr复合物对脱靶核酸分子具有更低的编辑活性。
[0823]
302.根据实施方案298所述的多核苷酸,其中所述编辑活性以被编辑的脱靶核酸分子的百分比来测量。
[0824]
303.根据实施方案298所述的多核苷酸,其中所述编辑活性以被编辑的靶核酸分
子的百分比来测量。
[0825]
304.根据实施方案298至300中任一项所述的多核苷酸,其中所述第一crispr复合物对所述脱靶核酸分子的编辑活性低于所述第二crispr复合物的编辑活性,其中p值≤0.0001。
[0826]
305.根据实施方案298至301中任一项所述的多核苷酸,其中所述第一crispr复合物对所述靶核酸分子的编辑活性和所述第二crispr复合物对所述靶核酸分子的编辑活性在5%以内。
[0827]
306.一种crispr复合物,所述crispr复合物包含实施方案132至302中任一项所述的多核苷酸和crispr效应蛋白。
[0828]
307.根据实施方案303所述的crispr复合物,其中所述crispr复合物包括核酸酶活性。
[0829]
308.一种核苷酸或寡核苷酸或多核苷酸,所述核苷酸或寡核苷酸或多核苷酸包含可裂解接头,所述可裂解接头包括式(i)的结构:
[0830][0831]
其中:
[0832]
r1、r2、r3、r4和r5各自独立地选自h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0833]
可选地,r1、r2、r3和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0834]
可选地,r1、r2、r3、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0835]
m是选自1至10的整数;并且
[0836]
x是o、s或cr
x
r
y
,其中r
x
和r
y
独立地选自任选地取代的烷基、任选地取代的烯基、任选地取代的炔基、任选地取代的杂烷基、卤素、卤代烷基、烷氧基、烷氧基烷基、卤代烷氧基、氨基、氨基烷基、烷基氨基、烷基氨基烷基、羟基、羟基烷基、c
‑
羧基、o
‑
羧基、酰基、硫醇、烷硫基、硫代烷基、c
‑
酰胺基、n
‑
酰胺基、脲基、硝基、氰基、磺酰基、磺基、磺酸酯、亚磺基、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基,
[0837]
条件是,当x是o,并且r3是二烷基氨基时,则m是2至10的整数;并且
[0838]
*表示与h或第一核苷酸的附接点;并且
[0839]
**表示与oh或第二核苷酸的附接点。
[0840]
309.根据实施方案305所述的核苷酸或寡核苷酸或多核苷酸,其中x是氧。
[0841]
310.根据实施方案305所述的核苷酸或寡核苷酸或多核苷酸,其中x是硫。
[0842]
311.根据实施方案305所述的核苷酸或寡核苷酸或多核苷酸,其中x是=c(cn)2。
[0843]
312.根据实施方案305所述的核苷酸或寡核苷酸或多核苷酸,其中所述式(i)的结构由式(i’)表示:
[0844][0845]
其中:
[0846]
r1、r2、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0847]
r
3a
和r
3b
独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0848]
可选地,r1、r2、r
3a
、r
3b
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0849]
可选地,r1、r2、r
3a
、r
3b
、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0850]
x是氧。
[0851]
313.根据实施方案305所述的核苷酸或寡核苷酸或多核苷酸,其中所述式(i)的结构由式(i’)表示:
[0852][0853]
其中:
[0854]
r1、r2、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0855]
r
3a
和r
3b
独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0856]
可选地,r1、r2、r
3a
、r
3b
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0857]
可选地,r1、r2、r
3a
、r
3b
、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0858]
x是硫。
[0859]
314.根据实施方案305所述的核苷酸或寡核苷酸或多核苷酸,其中所述式(i)的结构由式(i’)表示:
[0860][0861]
其中:
[0862]
r1、r2、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤
素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0863]
r
3a
和r
3b
独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0864]
可选地,r1、r2、r
3a
、r
3b
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0865]
可选地,r1、r2、r
3a
、r
3b
、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0866]
x是=c(cn)2。
[0867]
315.根据实施方案309至311中任一项所述的核苷酸或寡核苷酸或多核苷酸,其中:
[0868]
r1、r2、r4和r5各自独立地是h或c1‑6烷基;并且
[0869]
r
3a
和r
3b
是c1‑6烷基。
[0870]
316.根据实施方案309至311中任一项所述的核苷酸或寡核苷酸或多核苷酸,其中:
[0871]
r1、r2、r4和r5各自是h;并且
[0872]
r
3a
和r
3b
各自是乙基。
[0873]
317.一种核苷酸或寡核苷酸或多核苷酸,所述核苷酸或寡核苷酸或多核苷酸包含可裂解接头,所述可裂解接头包含由以下式表示的结构:
[0874][0875]
其中:
[0876]
x是o、s或cr
x
r
y
,其中r
x
和r
y
独立地选自任选地取代的烷基、任选地取代的烯基、任选地取代的炔基、任选地取代的杂烷基、卤素、卤代烷基、烷氧基、烷氧基烷基、卤代烷氧基、氨基、氨基烷基、烷基氨基、烷基氨基烷基、羟基、羟基烷基、c
‑
羧基、o
‑
羧基、酰基、硫醇、烷硫基、硫代烷基、c
‑
酰胺基、n
‑
酰胺基、脲基、硝基、氰基、磺酰基、磺基、磺酸酯、亚磺基、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;并且
[0877]
*表示与h或第一核苷酸的附接点;并且
[0878]
**表示与oh或第二核苷酸的附接点。
[0879]
318.根据实施方案314所述的核苷酸或寡核苷酸或多核苷酸,其中x是氧。
[0880]
319.根据实施方案314所述的核苷酸或寡核苷酸或多核苷酸,其中x是硫。
[0881]
320.根据实施方案314所述的核苷酸或寡核苷酸或多核苷酸,其中x是=c(cn)2。
[0882]
321.根据实施方案305至317中任一项所述的核苷酸或寡核苷酸或多核苷酸,其中所述核苷酸或寡核苷酸或多核苷酸还包含交联剂。
[0883]
322.根据实施方案318所述的核苷酸或寡核苷酸或多核苷酸,其中所述交联剂包括或衍生自吡啶基二硫化物、烷氧基胺、nhs酯、双吖丙啶、亚氨酸酯、卤代乙酰基基团、酰肼、芳基叠氮化物、异氰酸酯、二硫醇亚磷酰胺dtpa、4
‑
硫代
‑
utp、5
‑
叠氮基
‑
utp、5
‑
溴
‑
utp、8
‑
叠氮基
‑
atp、5
‑
apas
‑
utp或8
‑
n(3)amp。
[0884]
323.根据实施方案318所述的核苷酸或寡核苷酸或多核苷酸,其中所述交联剂包括二硫化物、酰胺、亚胺、酰肼、o
‑
烷基肟、烷基、胺、醇、三唑、异噁唑啉、异噁唑烷、异噁唑或哒嗪。
[0885]
324.根据实施方案318所述的核苷酸或寡核苷酸或多核苷酸,其中所述交联剂包括或衍生自吡啶基二硫化物、烷氧基胺、nhs酯、双吖丙啶、亚氨酸酯、卤代乙酰基基团、酰肼、芳基叠氮化物、异氰酸酯、二硫醇亚磷酰胺dtpa、4
‑
硫代
‑
utp、5
‑
叠氮基
‑
utp、5
‑
溴
‑
utp、8
‑
叠氮基
‑
atp、5
‑
apas
‑
utp或8
‑
n(3)amp。
[0886]
325.一种核苷酸或寡核苷酸或多核苷酸,所述核苷酸或寡核苷酸或多核苷酸包含交联剂,其中所述交联剂包括二硫化物、酰胺、亚胺、酰肼、o
‑
烷基肟、烷基、胺、醇、三唑、异噁唑啉、异噁唑烷、异噁唑或哒嗪。
[0887]
326.一种核苷酸或寡核苷酸或多核苷酸,所述核苷酸或寡核苷酸或多核苷酸包含交联剂,其中所述交联剂包括或衍生自吡啶基二硫化物、烷氧基胺、nhs酯、双吖丙啶、亚氨酸酯、卤代乙酰基基团、酰肼、芳基叠氮化物、异氰酸酯、二硫醇亚磷酰胺dtpa、4
‑
硫代
‑
utp、5
‑
叠氮基
‑
utp、5
‑
溴
‑
utp、8
‑
叠氮基
‑
atp、5
‑
apas
‑
utp或8
‑
n(3)amp。
[0888]
327.一种细胞,所述细胞包含以下中的一种或更多种:实施方案1至122、303或304中任一项所述的crispr复合物、实施方案130或131所述的sgrna或实施方案132至302中任一项所述的多核苷酸。
[0889]
328.根据实施方案320所述的细胞,其中所述细胞是干细胞。
[0890]
329.根据实施方案320所述的细胞,其中所述细胞是免疫细胞。
[0891]
330.根据实施方案325所述的细胞,其中所述干细胞是诱导性多能干细胞。
[0892]
331.根据实施方案326所述的细胞,其中所述免疫细胞是t细胞。
[0893]
332.根据实施方案326所述的细胞,其中所述免疫细胞是自然杀伤细胞(nk细胞)。
[0894]
333.一种化合物,所述化合物包含由以下式表示的结构:
[0895][0896]
其中:
[0897]
x是o、s或cr
x
r
y
,其中r
x
和r
y
独立地选自任选地取代的烷基、任选地取代的烯基、任选地取代的炔基、任选地取代的杂烷基、卤素、卤代烷基、烷氧基、烷氧基烷基、卤代烷氧基、氨基、氨基烷基、烷基氨基、烷基氨基烷基、羟基、羟基烷基、c
‑
羧基、o
‑
羧基、酰基、硫醇、烷硫基、硫代烷基、c
‑
酰胺基、n
‑
酰胺基、脲基、硝基、氰基、磺酰基、磺基、磺酸酯、亚磺基、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;并且
[0898]
*表示与h或第一核苷酸的附接点;并且
[0899]
**表示与oh或第二核苷酸的附接点。
[0900]
334.根据实施方案330所述的化合物,其中x是氧。
[0901]
335.根据实施方案330所述的化合物,其中x是硫。
[0902]
336.根据实施方案330所述的化合物,其中x是=c(cn)2。
[0903]
337.一种化合物,所述化合物包含由式(i)表示的结构:
[0904][0905]
其中:
[0906]
r1、r2、r3、r4和r5各自独立地选自h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0907]
可选地,r1、r2、r3和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0908]
可选地,r1、r2、r3、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0909]
m是选自1至10的整数;并且
[0910]
x是o、s或cr
x
r
y
,其中r
x
和r
y
独立地选自任选地取代的烷基、任选地取代的烯基、任选地取代的炔基、任选地取代的杂烷基、卤素、卤代烷基、烷氧基、烷氧基烷基、卤代烷氧基、氨基、氨基烷基、烷基氨基、烷基氨基烷基、羟基、羟基烷基、c
‑
羧基、o
‑
羧基、酰基、硫醇、烷硫基、硫代烷基、c
‑
酰胺基、n
‑
酰胺基、脲基、硝基、氰基、磺酰基、磺基、磺酸酯、亚磺基、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基,
[0911]
条件是,当x是o,并且r3是二烷基氨基时,则m是2至10的整数;并且
[0912]
*表示与h或第一核苷酸的附接点;并且
[0913]
**表示与oh或第二核苷酸的附接点。
[0914]
338.根据实施方案334所述的化合物,其中x是氧。
[0915]
339.根据实施方案334所述的化合物,其中x是硫。
[0916]
340.根据实施方案334所述的化合物,其中x是=c(cn)2。
[0917]
341.根据实施方案334所述的化合物,其中所述式(i)的结构由式(i’)表示:
[0918][0919]
其中:
[0920]
r1、r2、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0921]
r
3a
和r
3b
独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0922]
可选地,r1、r2、r
3a
、r
3b
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0923]
可选地,r1、r2、r
3a
、r
3b
、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0924]
x是氧。
[0925]
342.根据实施方案334所述的化合物,其中所述式(i)的结构由式(i’)表示:
[0926][0927]
其中:
[0928]
r1、r2、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0929]
r
3a
和r
3b
独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0930]
可选地,r1、r2、r
3a
、r
3b
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0931]
可选地,r1、r2、r
3a
、r
3b
、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0932]
x是硫。
[0933]
343.根据实施方案334所述的化合物,其中所述式(i)的结构由式(i’)表示:
[0934][0935]
其中:
[0936]
r1、r2、r4和r5各自独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯
基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、单烷基氨基、二烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0937]
r
3a
和r
3b
独立地选自由以下组成的组:h、烷基、取代的烷基、烷氧基、烯基、炔基、卤代烷基、卤代烷氧基、烷氧基烷基、氨基、烷基氨基、氨基烷基、卤素、氰基、羟基、羟基烷基、杂烷基、c
‑
羧基、o
‑
羧基、c
‑
酰胺基、n
‑
酰胺基、硝基、磺酰基、磺基、亚磺基、磺酸酯、s
‑
磺酰胺基、n
‑
磺酰胺基、任选地取代的碳环基、任选地取代的芳基、任选地取代的杂芳基和任选地取代的杂环基;
[0938]
可选地,r1、r2、r
3a
、r
3b
和r4中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;
[0939]
可选地,r1、r2、r
3a
、r
3b
、r4和r5中的两个或更多个连同它们所附接的原子一起形成选自以下的环或环体系:任选地取代的5元至10元杂芳基、任选地取代的5元至10元杂环基和任选地取代的c5‑
10
碳环;并且
[0940]
x是=c(cn)2。
[0941]
344.一种药物制剂,所述药物制剂包含以下中的一种或更多种:实施方案1至129、303或304中任一项所述的crispr复合物、实施方案130或131所述的sgrna、实施方案132至302中任一项所述的多核苷酸、实施方案324至329中任一项所述的细胞或实施方案330至340中任一项所述的化合物。
[0942]
345.根据实施方案341所述的药物制剂,所述药物制剂还包含药学上可接受的赋形剂。
[0943]
346.一种方法,所述方法包括向受试者施用实施方案341或342所述的药物制剂。
[0944]
347.根据实施方案343所述的方法,其中所述受试者是哺乳动物。
[0945]
348.根据实施方案344所述的方法,其中所述哺乳动物是人类。
[0946]
349.一种方法,所述方法包括将以下中的一种或更多种引入细胞中:实施方案1至129、303或304中任一项所述的crispr复合物、实施方案130或131所述的sgrna或实施方案132至302中任一项所述的多核苷酸。
[0947]
350.根据实施方案346所述的方法,其中所述细胞是干细胞。
[0948]
351.根据实施方案346所述的方法,其中所述细胞是免疫细胞。
[0949]
352.根据实施方案347所述的方法,其中所述干细胞是诱导性多能干细胞。
[0950]
353.根据实施方案348所述的方法,其中所述免疫细胞是t细胞。
[0951]
354.根据实施方案348所述的方法,其中所述免疫细胞是自然杀伤细胞(nk细胞)。
[0952]
355.一种编辑核酸分子的方法,所述方法包括使实施方案1至129、303或304中任一项所述的crispr复合物与核酸分子接触。
[0953]
356.根据实施方案352所述的方法,其中所述crispr复合物包括小于2%裂解事件的脱靶裂解活性。
[0954]
357.一种编辑核酸分子的方法,所述方法包括使所述核酸分子与crispr效应蛋白和以下中的一种或更多种接触:实施方案130或131所述的sgrna或实施方案132至302中任
一项所述的多核苷酸。
[0955]
358.一种在一个或更多个细胞中编辑靶基因的方法,所述方法包括将实施方案1至129、303或304中任一项所述的crispr复合物施用至包含靶基因的所述一个或更多个细胞,从而产生一个或更多个包含编辑的靶基因的细胞。
[0956]
359.一种在一个或更多个细胞中编辑靶基因的方法,所述方法包括向包含靶基因的所述一个或更多个细胞施用crispr效应蛋白或编码crispr效应蛋白的多核苷酸以及以下中的一种或更多种:实施方案130或131所述的sgrna或实施方案132至302中任一项所述的多核苷酸,从而产生一个或更多个包含编辑的靶基因的细胞,其中99%的所述包含编辑的靶基因的细胞在施用之后保持存活。
[0957]
360.根据实施方案355或356所述的方法,其中99%的所述包含编辑的靶基因的细胞在施用之后保持存活。
[0958]
361.根据实施方案357所述的方法,所述方法还包括通过刃天青测定来测量细胞存活力。
[0959]
362.一种产生crispr复合物的方法,所述方法包括将引导rna(grna)交联,其中所述交联发生在所述grna的靶结合区之外的核苷酸处,并且其中crispr效应蛋白的核酸酶活性在所述交联之后被维持。
[0960]
363.根据实施方案359所述的方法,其中所述交联发生在溶液中,并且其中所述溶液中的所述grna与与所述crispr效应蛋白的比例是至少9:1。
[0961]
364.根据实施方案360所述的方法,其中所述交联包括将所述溶液暴露于紫外(uv)光。
[0962]
365.根据实施方案359至361中任一项所述的方法,其中所述交联在所述grna与所述crispr效应蛋白混合时发生。
[0963]
366.一种裂解实施方案202至302中任一项所述的多核苷酸的方法,所述方法包括将多所述核苷酸暴露于裂解媒介,从而使所述可裂解接头裂解。
[0964]
367.一种方法,所述方法包括:
[0965]
引入crispr效应蛋白和实施方案202至302中任一项所述的多核苷酸;并且
[0966]
将所述多核苷酸暴露于裂解媒介,从而使所述可裂解接头裂解。
[0967]
368.根据实施方案363或364所述的方法,其中所述多核苷酸与crispr效应蛋白复合。
[0968]
369.根据实施方案363至365中任一项所述的方法,其中所述裂解媒介是紫外(uv)光。
[0969]
370.根据实施方案366所述的方法,其中所述裂解媒介是波长在100nm至400nm范围内的光。
[0970]
371.根据实施方案363至365中任一项所述的方法,其中所述裂解媒介是可见光。
[0971]
372.根据实施方案363至365中任一项所述的方法,其中所述裂解媒介是波长为400nm至700nm的光。
[0972]
373.根据实施方案363至365中任一项所述的方法,其中所述裂解媒介是绿光。
[0973]
374.根据实施方案365至365中任一项所述的方法,其中所述裂解媒介是紫光。
[0974]
375.根据实施方案363至365中任一项所述的方法,其中所述裂解媒介是蓝光。
[0975]
376.根据实施方案363至365中任一项所述的方法,其中所述裂解媒介是波长在490nm至570nm范围内的光。
[0976]
377.根据实施方案363至365中任一项所述的方法,其中所述裂解媒介是波长在400nm至420nm范围内的光。
[0977]
378.根据实施方案363至365中任一项所述的方法,其中所述裂解媒介是波长在420nm至430nm范围内的光。
[0978]
379.根据实施方案363至365中任一项所述的方法,其中所述裂解媒介是波长在420nm至440nm范围内的光。
[0979]
380.根据实施方案363至371中任一项所述的方法,其中所述暴露增加与所述多核苷酸复合的crispr效应蛋白的靶特异性裂解活性。
[0980]
381.根据实施方案363至373中任一项所述的方法,其中所述多核苷酸不位于细胞中。
[0981]
382.根据实施方案363至373中任一项所述的方法,其中所述多核苷酸位于细胞中。
[0982]
383.根据实施方案379所述的方法,其中所述细胞是干细胞。
[0983]
384.根据实施方案379所述的方法,其中所述细胞是免疫细胞。
[0984]
385.根据实施方案380所述的方法,其中所述干细胞是诱导性多能干细胞。
[0985]
386.根据实施方案381所述的方法,其中所述免疫细胞是t细胞。
[0986]
387.根据实施方案381所述的方法,其中所述免疫细胞是自然杀伤细胞(nk细胞)。
[0987]
388.一种系统,所述系统包括以下中的一种或更多种:实施方案1至129、303或304中任一项所述的crispr复合物、实施方案130或131所述的sgrna、实施方案132至302中任一项所述的多核苷酸、实施方案324至329中任一项所述的细胞、实施方案330至340中任一项所述的化合物或实施方案341或342所述的药物制剂。
[0988]
389.一种试剂盒,所述试剂盒包括以下中的一种或更多种:实施方案1至129、303或304中任一项所述的crispr复合物、实施方案130或131所述的sgrna、实施方案132至302中任一项所述的多核苷酸、实施方案324至329中任一项所述的细胞、实施方案330至340中任一项所述的化合物、实施方案341或342所述的药物制剂或实施方案385所述的系统。
[0989]
390.根据实施方案386所述的试剂盒,所述试剂盒还包括用于进行实施方案343至384中任一项所述的方法的说明书。
实施例
[0990]
实施例1:具有位于四环中的交联剂的修饰的多核苷酸。
[0991]
最初的研究创建了一种修饰的多核苷酸,其中在位置22和位置49处的特定核苷酸被交换。如图1中可见的,所述核苷酸位于四环的碱基处。此外,该位置处的核苷酸被转化为脱氧核糖核酸。这种转化对多核苷酸或核糖核蛋白活性具有极小的影响,图2。可选地,位置50处的尿嘧啶分子可以被转化为脱氧胸苷。
[0992]
接下来,多核苷酸通过用包含反应性接头化合物的类似物取代位置49或50处的脱氧核苷酸被进一步修饰。通过已知化学将马来酰亚胺共价附接至该反应性接头。此外,这种马来酰亚胺化合物可以包含可变长度的间隔物。然后将修饰的多核苷酸与cas9混合以形成
核糖核蛋白。核糖核蛋白在生理条件下将经历交联反应。
[0993]
接下来,将交联的核糖核蛋白(锁定的rnp)从单独游离组分和非特异性交联物质中纯化。首先将混合溶液使用尺寸排阻层析来纯化,以分离游离的多核苷酸。接下来,将收集的级分通过具有固定的sgrna的亲和层析柱,以使游离的多肽与形成的rnp分开。最后,将纯化的rnp在高盐条件(例如,300mm nacl)下通过阳离子交换柱以分离锁定的rnp(如图16中可见的)。高盐条件将引起rnp解离为组分物质cas9和sgrna。由于锁定的rnp的共价键性质,这些物质将保持在一起,并且可以相应地纯化。
[0994]
将纯化的锁定的rnp转染到永生化细胞系中,以测定它们在人类细胞内形成双链断裂的能力。合成靶向基因组特定区域的多核苷酸并形成锁定的rnp。转染后,对转染的池(pool)进行sanger测序,并且所得基因组数据通过crispr编辑的推理(ice)软件进行分析,以检测插入/缺失突变的存在。
[0995]
为了测试锁定的rnp复合物的特异性,包含特定多核苷酸序列的锁定的rnp将被形成并纯化。将向这种纯化物添加10倍过量的靶向单独基因组区域的多核苷酸并且允许混合。然后将这种混合物转染到细胞中。对两个基因座处的编辑进行分析,并且显示crispr诱导的编辑仅发生在被锁定的rnp靶向的基因座处。
[0996]
实施例2:具有被修饰以允许交联的sgrna的crispr复合物的靶dna切割效率的表征
[0997]
将cas9核酸酶(来自aldevron的nls
‑
cas9
‑
nls;pdb:4008)与被设计为在锁定的crispr复合物中使用的包含不同修饰的3种不同的sgrna引入hek 293细胞中。对于每种修饰,产生靶向五个dna靶之一的sgrna。将15种不同的sgrna与cas9核酸酶进行测试,将每一种引入相应的hek293细胞培养物中。图15示出了靶向emx1、fancf、grk1、prgn和vegfa的15种不同sgrna的基因组编辑效率示意图。sgrna的第一种修饰(“u50t”)用胸苷dna核苷酸替代位置50处的尿嘧啶核苷酸。sgrna的第二种修饰(“u49a”)使核苷酸位置u22和a49翻转,保存茎环结构,同时允许进一步的功能修饰。sgrna的第三种修饰(“u49t”)基于u49a进一步构建,但用脱氧胸苷替代新的u49。每种修饰与五个靶结合区中的每一个组合,以靶向五个靶中的每一个。五个dna靶区域是:emx1(gagtccgagcagaagaagaa);fancf(gctgcagaagggattccatg);grk1(gccgtcaaagctgcctcggg);prgn(cagatgcctgctcagtgttg);和vegfa(ggtgagtgagtgtgtgcgtg)。
[0998]
从所有样品收获基因组dna,并且使用ice(crispr编辑的推理)分析插入和缺失的存在。ice通过分析sanger测序示踪和将序列断裂水平映射至插入/缺失形成频率来测量基因编辑的量,如hsiau等人,“inference of crispr edits from sanger trace data”,2019biorxiv中描述的。图15中的图表示编辑效率。将样品使用pcr扩增并提交测序。在测序之后,通过ice来分析扩增后野生型或被编辑的序列的数目。编辑表示为非野生型序列的百分比。除了在pgrn基因的基因座处使用u50t引导观察到的编辑丧失以外,大多数修饰的引导sgrna能够诱导遍及五个靶的基因编辑。
[0999]
实施例3:具有可光裂解接头的修饰的多核苷酸的产生及功能表征.
[1000]
合成了四种sgrna。下文提供了示出sgrna的一部分的序列。对sgrna的修饰包括在前三个5’末端rna核苷酸和前三个3’末端rna核苷酸处的2
’‑
o
‑
甲基类似物和3’硫代磷酸酯核苷酸间连接。
[1001]“对照”:
[1002]
nnnnnnnnnnnnnnnnnnnnguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu
[1003]“无二级”:
[1004]
gaaannnnnnnnnnnnnnnnnnnnguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu
[1005]“3bp茎”:
[1006]
ugagaaaucannnnnnnnnnnnnnnnnnnnguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu
[1007]“6bp茎”:
[1008]
cacugagaaaucagugnnnnnnnnnnnnnnnnnnnnguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu
[1009]
第一sgrna(“对照”或“mod”)是缺少引导序列5’侧的多核苷酸前导序列的sgrna。第二sgrna(“无二级”或“无二级结构(no secondary)”)具有被设计为不形成茎环的引导序列5’测的多核苷酸前导序列,随后是插入多核苷酸前导序列3’末端和引导序列5’侧之间的可光裂解接头3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺(www.glenresearch.com/data/productinfo.php?item=10
‑
4920)。两种另外的sgrna被合成为具有设计成在引导序列的5’碱基之前形成茎环的添加的多核苷酸前导序列,随后是插入添加的多核苷酸前导序列的3’末端和引导序列的5’碱基之间的3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺(http://www.glenresearch.com/data/productinfo.php?item=10
‑
4920)可光裂解接头。第三sgrna(“3bp茎”)具有被设计为形成3bp茎环的多核苷酸前导序列,并且第四sgrna(“6bp茎”)具有被设计为形成6bp茎环的多核苷酸前导序列。然后,使用已知足以在体外光裂解sgrna的条件,将这四种类型的sgrna暴露于uva光(320
‑
390nm)。
[1010]
图19是使用来自advanced analytical的小rna分析试剂盒在片段分析仪上运行的凝胶图像,描绘了在暴露于uv光0分钟、5分钟、10分钟或15分钟之后的四种sgrna。来自设定时间点的所有图像是在同一凝胶上的相邻泳道中运行的,即,所有10分钟的样品彼此相邻运行。来自不同时间点的样品在不同的凝胶上运行,以允许测试多个sgrna靶位点。条件之间的比较主要基于定性观察。暴露于uv光5分钟之后,第二sgrna、第三sgrna和第四sgrna显示出与多核苷酸前导序列的裂解一致的条带模式。暴露10分钟和15分钟之后的条带模式也与来自第二sgrna、第三sgrna和第四sgrna的多核苷酸前导序列的裂解一致。
[1011]
实施例4:sgrna激活后靶dna切割效率的表征
[1012]
如以上描述的,在体外将四种sgrna与spcas9复合,并与靶dna一起孵育适当的持续时间。然后,如图20中示出的,将四种sgrna各自暴露于175mw/cm2的uv光(320
‑
390nm),持续所示的周期间隔。uv介导的具有设计的茎(“3bp茎”和“6bp茎”)的sgrna的裂解用于激活crispr复合物,导致靶特异性dna的切割。然后,将靶dna在片段分析仪上运行,以展示crispr介导的切割。图20示出了与靶dna(fancf)一起孵育并以规律间隔暴露于裂解媒介的sgrna
‑
cas9 crispr复合物的实例。在暴露后0分钟,与第一sgrna和第二sgrna相比,第三sgrna和第四sgrna(“3bp茎”和“6bp茎”)显示出降低的切割效率。在暴露于裂解媒介15min
之后,激活的sgrna记录到靶dna切割效率的增加。对未切割dna与切割dna的比例的测量显示,“6bp茎”从暴露15分钟时的~45%降低到暴露30分钟时的~20%。相比之下,缺少5’多核苷酸前导序列(“mod”)或5’二级结构(“无二级结构”)的第一sgrna没有表现出被裂解媒介激活,如通过未切割dna与切割dna的比例测量的。“mod”是在前三个5’末端rna核苷酸和前三个3’末端rna核苷酸处具有2
’‑
o
‑
甲基类似物和3’硫代磷酸酯核苷酸间连接、缺少向引导序列的任何5’添加碱基的修饰的合成sgrna,并且“无二级结构”条件使用具有向引导序列的非茎形成性5’侧添加的sgrna。“3bp茎”和“6bp茎”条件使用具有被设计为分别在sgrna的5’末端处形成3bp和6bp长度的茎的区域的sgrna。
[1013]
实施例5:可失活sgrna的产生和表征
[1014]
合成了6种sgrna。下文提供了示出sgrna的一部分的序列。对sgrna的修饰包括在前三个5’末端rna核苷酸和前三个3’末端rna核苷酸处的2
’‑
o
‑
甲基类似物和3’硫代磷酸酯核苷酸间连接。
[1015]
对照:
[1016]
nnnnnnnnnnnnnnnnnnnnguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu
[1017]
21
[1018]
nnnnnnnnnnnnnnnnnnnn*uuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu
[1019]
24
[1020]
nnnnnnnnnnnnnnnnnnnnguu*uagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu
[1021]
50
[1022]
nnnnnnnnnnnnnnnnnnnnguuuuagagcuagaaauagcaaguuaaaa*aaggcuaguccguuaucaacuugaaaaaguggcaccgagucggugcuuuu
[1023]
57
[1024]
nnnnnnnnnnnnnnnnnnnnguuuuagagcuagaaauagcaaguuaaaauaaggcu*guccguuaucaacuugaaaaaguggcaccgagucggugcuuuu
[1025]
74
[1026]
nnnnnnnnnnnnnnnnnnnnguuuuagagcuagaaauagcaaguuaaaauaaggcuaguccguuaucaacuug*aaaaguggcaccgagucggugcuuuu
[1027]
第一sgrna(“对照”)不包含可光裂解元件。第二sgrna、第三sgrna、第四sgrna和第五sgrna在距sgrna的5’末端的位置21、24、50、57和74处具有可光裂解键。然后,将五种sgrna暴露于uv光5分钟。图21是描绘暴露于uv光之后的五种sgrna使用来自advanced analytical的片段分析仪运行的凝胶图像。根据制造商的方案,将所有样品用小rna试剂盒来运行。在暴露于uv光5分钟之后,所有五种sgrna均显示出与在各自的可光裂解键位置处的裂解一致的条带模式。
[1028]
实施例6:基因组编辑的细胞系的快速产生
[1029]
将表达cas9的hek 293t细胞用包含可光裂解接头的sgrna转染,并且经历裂解媒介。图22示出了靶向dnmt1的六种不同的sgrna的可编程基因组编辑效率的示意图。第一
sgrna(“mod”)缺少可光裂解位点。第二sgrna、第三sgrna、第四sgrna和第五sgrna在距sgrna的5’末端的位置21、24、50、57和74(分别为b21、b24、b50、b57和b74)处具有可光裂解键。将sgrna:cas9混合物[9:1的比]引入细胞中。将细胞每两小时一次暴露于裂解媒介,持续48小时。将每个样品保持在黑暗中直到指定的时间点,然后暴露于uv光一次以诱导裂解。然后将细胞留置于黑暗中,直到转染后48小时。所有样品在转染后48小时收集。在转染后48小时之后,从所有样品收获基因组dna,并且使用ice(crispr编辑的推理)分析插入和缺失的存在。ice通过分析sanger测序示踪和将序列断裂水平映射至插入/缺失形成频率来测量基因编辑的量,如hsiau等人,“inference of crispr edits from sanger trace data”,2019biorxiv中描述的。图22中的图表示编辑效率。将样品使用pcr扩增并提交测序。在测序之后,扩增后野生型或被编辑的序列的数目通过ice来分析。编辑表示为非野生型序列的百分比。在位置57和位置74处具有可光裂解键的sgrna展示出基因组编辑效率的时间依赖性失活。
[1030]
实施例7:编辑的hek 293细胞系的产生
[1031]
将hek 293细胞用cas9和包含可光裂解(pc)接头的sgrna转染,并且经历uv光以使接头裂解。将cas9与12种不同的sgrna(crispr off)复合以产生12种实验群体,所述12种不同的sgrna(crispr off)在位置57和位置74处包含掺入的亚磷酰胺(3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺),且具有靶向以下的靶结合区域:bub1b(agtgaagccatgtccctgga)、camk1(sg1:tgccaggatcacctccgaga)、prkag3(sg1
‑
agcaagaaaacagcagctca;sg2
‑
agcaagaaaacagcagcuca)、stk3(sg1
‑
tcctgaagatctgattcaac;sg2
‑
aaagcaatacacaaggaatc;sg3
‑
ccataatgcagcaatgtgac;sg4
‑
uuuaauugcgacaacuugac)、irak4(gtcctgtctttgtcacagaa)、和chr8q23(sg1
‑
agtctactatgagttttctg;sg2
‑
ttatagttacgatgtttgat;sg3
‑
aagcctcaaattaggagaaa)。还将cas9与不含可光裂解接头且具有以上描述的靶结合区的12种不同的sgrna(标准)复合。为了形成24种复合物溶液中的每一种,将10pmol的cas9蛋白与30pmol的sgrna混合。将每种溶液使用转染缓冲液稀释至20μl,并允许在室温混合15分钟。在室温使用tryple 5分钟以使细胞单个化来收获hek293细胞。对群体进行计数以确定适当的细胞数目,随后以100
×
g离心3分钟。然后将所得沉淀物以每200,000个细胞/5μl的浓度重悬于核转染缓冲液中。然后将细胞悬浮液添加至预先复合的cas9 sgrna溶液并转染。将每种实验群体分到两个孔中,以形成对照和处理细胞的成对重复。在转染之后4小时,将处理细胞暴露于uv光1分15秒(用带通滤光器将波长限制在大于345nm的波长)。随后将细胞放回培养箱。转染后48小时,收获对照和处理样品并提取基因组dna。使用amplitaq以及对中靶位点和脱靶位点特异性的引物对基因组dna进行pcr。使用ice来分析测序数据的编辑的存在。ice(crispr编辑的推理)通过分析sanger测序示踪和提供序列断裂水平的信息以确定插入/缺失形成频率来测量基因编辑的量,如hsiau等人“inference of crispr edits from sanger trace data”,january 14,2019biorxiv中描述的。编辑表示为非野生型序列的百分比。
[1032]
图23示出了带有12种不同的crispr off sgrna的cas9的编辑效率的图。灰色条表示在没有uv光暴露的情况下与crispr off sgrna复合的cas9的编辑效率。黑色条表示在uv光暴露之后与crispr off sgrna复合的cas9的编辑效率。
[1033]
图24示出了带有12种不同的标准sgrna(不含可光裂解接头,具有与图23中的sgrna相同的靶结合区域)的cas9的编辑效率。灰色条表示在没有uv光暴露的情况下与标准sgrna复合的cas9的编辑效率。黑色条表示在uv光暴露之后与标准sgrna复合的cas9的编辑效率。
[1034]
实施例8:编辑的u2os细胞系的产生
[1035]
将u2os细胞用cas9和包含可光裂解接头的sgrna转染,并且经历uv光以使接头裂解。将cas9与6种不同的sgrna(crispr off)复合以产生6种实验群体,所述6种不同的sgrna(crispr off)在位置57和位置74处包含掺入的亚磷酰胺(3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺),且具有靶向以下的靶结合区域:dnmt1(ggagtgagggaaacggcccc)、emx1(gagtccgagcagaagaagaa)、fancf(gctgcagaagggattccatg)、grk1(gccgtcaaagctgcctcggg)、prgn(cagatgcctgctcagtgttg)和vegfa(ggtgagtgagtgtgtgcgtg)。还将cas9与不含可光裂解接头且具有以上描述的靶结合区的6种不同的sgrna(标准)复合。为了形成12种复合物溶液中的每一种,将10pmol的cas9蛋白与30pmol的sgrna混合。将每种溶液使用转染缓冲液稀释至20μl,并允许在室温混合15分钟。在室温使用tryple 5分钟以使细胞单个化来收获u20s细胞。对群体进行计数以确定适当的细胞数目,随后以100
×
g离心3分钟。然后将所得沉淀物以每200,000个细胞/5μl的浓度重悬于核转染缓冲液中。然后将细胞悬浮液添加至预先复合的cas9 sgrna溶液并转染。将每种实验群体分到两个孔中,以形成对照和处理细胞的成对重复。在转染之后4小时,将处理细胞暴露于uv光1分15秒(用带通滤光器将波长限制在大于345nm的波长)。随后将细胞放回培养箱。转染后48小时,收获对照和处理样品并提取基因组dna。使用amplitaq以及对中靶位点和脱靶位点特异性的引物对基因组dna进行pcr。使用ice来分析测序数据的编辑的存在。ice(crispr编辑的推理)通过分析sanger测序示踪和提供序列断裂水平的信息以确定插入/缺失形成频率来测量基因编辑的量,如hsiau等人“inference of crispr edits from sanger trace data”,january 14,2019biorxiv中描述的。编辑表示为非野生型序列的百分比。
[1036]
图25示出了带有6种不同的crispr off sgrna的cas9的编辑效率的图。灰色条表示在没有uv光暴露的情况下与crispr off sgrna复合的cas9的编辑效率。黑色条表示在uv光暴露之后与crispr off sgrna复合的cas9的编辑效率。
[1037]
图26示出了带有6种不同的标准sgrna(不含可光裂解接头,具有与图25中的sgrna相同的靶结合区域)的cas9的编辑效率。灰色条表示在没有uv光暴露的情况下与标准sgrna复合的cas9的编辑效率。黑色条表示在uv光暴露之后与标准sgrna复合的cas9的编辑效率。
[1038]
实施例9:在未暴露于uv光的情况下,crispr
‑
off cas9复合物在hek293t细胞中的脱靶编辑的分析
[1039]
图29包括描绘上述靶向dnmt1、fancf和vegfa的sgrna的已知具有高度脱靶编辑的脱靶位点处的编辑百分比的图。
[1040]
使用的序列如下,其中*表示接头(3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺)的位置:
[1041]
dnmt1
[1042]
中靶sgrna:
sgrna复合的crispr酶,其中p值≤0.0001。如通过上述ice测量的,每个以上提及的crispr
‑
off sgrna在靶位点处的编辑效率与每个以上提及的标准sgrna在靶位点处的编辑效率相同或比每个以上提及的标准sgrna在靶位点处的编辑效率低1%
‑
3%。结果表明,相对于使用连结和茎环1中缺少3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺的sgrna,在编辑测定中使用连结和茎环1中具有3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺的sgrna导致更低的脱靶编辑活性。
[1060]
实施例10:与crispr off复合的cas9在u2os细胞中的时间依赖性活性的分析
[1061]
图29
‑
图32是描绘与靶向dnmt1、grk1和vegfa的“crispr off”复合的cas9的时间依赖性活性对比与“标准sgrna”复合的cas9的时间依赖性活性的缺乏的图。将细胞每两小时一次暴露于uv光,持续48小时。ice(crispr编辑的推理)通过分析sanger测序示踪和提供序列断裂水平的信息以确定插入/缺失形成频率来测量基因编辑的量,如hsiau等人“inference of crispr edits from sanger trace data”,january 14,2019biorxiv中描述的。
[1062]
实施例11:编辑的k562细胞系的产生
[1063]
将k562细胞用cas9和包含可光裂解接头的sgrna转染,并且经历uv光以使接头裂解。将cas9与两种不同的sgrna(crispr off)复合以产生两种实验群体,所述两种不同的sgrna(crispr off)在位置57和位置74处包含掺入的亚磷酰胺(3
‑
(4,4'
‑
二甲氧基三苯甲基)
‑1‑
(2
‑
硝基苯基)
‑
丙
‑1‑
基
‑
[(2
‑
氰乙基)
‑
(n,n
‑
二异丙基)]
‑
亚磷酰胺)(crispr off),且具有靶向emx1(gagtccgagcagaagaagaa)和grk1(gccgtcaaagctgcctcggg)的靶结合区域。还将cas9与不含可光裂解接头且具有以上描述的靶结合区的2种不同的sgrna(标准)复合。为了形成4种复合物溶液中的每一种,将10pmol的cas9蛋白与30pmol的sgrna混合。将每种溶液使用转染缓冲液稀释至20μl,并允许在室温混合15分钟。在室温使用tryple 5分钟以使细胞单个化来收获k562细胞。对群体进行计数以确定适当的细胞数目,随后以100
×
g离心3分钟。然后将所得沉淀物以每200,000个细胞/5μl的浓度重悬于核转染缓冲液中。然后将细胞悬浮液添加预先复合的cas9 sgrna溶液并转染。将每种实验群体分到两个孔中,以形成对照和处理细胞的成对重复。在转染之后4小时,将处理细胞暴露于uv光~1分15秒(用带通滤光器将波长限制在大于345nm的波长)。随后将细胞放回培养箱。转染后48小时,收获对照和处理样品并提取基因组dna。使用amplitaq以及对中靶位点和脱靶位点特异性的引物对基因组dna进行pcr。使用ice来分析测序数据的编辑的存在。ice(crispr编辑的推理)通过分析sanger测序示踪和提供序列断裂水平的信息以确定插入/缺失形成频率来测量基因编辑的量,如hsiau等人“inference of crispr edits from sanger trace data”,january 14,2019biorxiv中描述的。编辑表示为非野生型序列的百分比。
[1064]
图27示出了带有两种不同的crispr off sgrna的cas9的编辑效率的图。灰色条表示在没有uv光暴露的情况下与crispr off sgrna复合的cas9的编辑效率。黑色条表示在uv光暴露之后与crispr off sgrna复合的cas9的编辑效率。
[1065]
图28示出了带有2种不同的标准sgrna(不含可光裂解接头,具有与图27中的sgrna相同的靶结合区域)的cas9的编辑效率。灰色条表示在没有uv光暴露的情况下与标准sgrna复合的cas9的编辑效率。黑色条表示在uv光暴露之后与标准sgrna复合的cas9的编辑效率。
[1066]
实施例12:转录调节
[1067]
包含形成10bp茎环的5’多核苷酸前导序列和将多核苷酸与crispr效应蛋白交联的非天然核苷酸的修饰的可激活(crispr on)sgrna多核苷酸与跟转录激活因子结构域vp64融合的无活性的cas9核酸酶(dcas9)交联。可光裂解元件插入多核苷酸前导序列的3’侧和紧接引导序列的5’侧。将包含与dcas9融合酶交联的sgrna的crispr复合物转染到hek293t细胞中。5’多核苷酸前导序列使得crispr复合物不能够有效地与跟引导序列互补的靶序列的启动子退火。靶基因具有相对较低的转录活性。在期望的时间,将转染的细胞暴露于uv光,导致可光裂解键的裂解和多核苷酸前导序列的释放。crispr复合物现在更有效地与靶序列的启动子结合,并且产生更有效的靶序列转录。
[1068]
实施例13:在暴露于uv光和未暴露于uv光的情况下,crispr
‑
off cas9复合物在hek293细胞中的中靶编辑的分析
[1069]
将人类胚胎肾细胞(hek293)在高级改良eagles培养基(advanced modified eagles medium,life technologies)和10%v/v fbs中维持在第5
‑
20代之间。用tryple(life technologies)将细胞以1:8的比例每周两次进行传代。
[1070]
将hek 293细胞用cas9和包含可光裂解(pc)接头的sgrna转染,并且经历用345nm带通滤光器过滤的光,以使接头裂解。将cas9与23种不同的sgrna(crispr off)复合以产生23种实验群体,所述sgrna(crispr off)在位置57和位置74处包含掺入的可光裂解接头(1
‑
(7
‑
(二乙基氨基)
‑2‑
氧代
‑
2h
‑
色烯
‑4‑
基)丙基),且具有靶向以下的靶结合区域:aavs1(ggggccacuagggacaggau)、bub1b(agugaagccaugucccugga)、camk1_sg1(ugccaggaucaccuccgaga)、camk1_sg2(gcguccucuuaucuucugcc)、cel(aaccaguugcaggcgcccca)、chr8q23_sg1(uuauaguuacgauguuugau)、cxcr4(gauaacuacaccgaggaaau)、dnmt1(ggagugagggaaacggcccc)、emx1(gaguccgagcagaagaagaa)、fam163a(cugcagggcucgcuggugag)、fancf(gcugcagaagggauuccaug)、gaa(aggagccggugggagcaggg)、grk1(gccgucaaagcugccucggg)、itga7(ggugcuggagggcgaggcug)、irak4(guccugucuuugucacagaa)、mapre1(uucucugcagauaauuccug)、mip(gcugggguccucacugcgcu)、omp(gaacuguagccgcugcugcu)、opn1sw(acaggggcaaugugguacug)、prgn(cagaugccugcucaguguug)、prkag3(agcaagaaaacagcagcuca)、stk3_sg1(aaagcaauacacaaggaauc)、stk3_sg2(ccauaaugcagcaaugugac)和vegfa(ggugagugagugugugcgug)。然后,将每种实验群体分成三组,一组保持在黑暗中,一组暴露于环境光,并且一组暴露于用345nm带通滤光器过滤以将波长限制在大于345nm的波长的光。为了形成23种复合物溶液中的每一种,将10pmol的cas9蛋白与30pmol的sgrna混合。将每种溶液使用转染缓冲液稀释至20μl,并允许混合10分钟,然后转染。在转染之后4小时,将处理的细胞暴露于环境光20分钟,或者暴露于用345nm带通滤光器过滤以将波长限制在大于345nm的波长的光60秒。转染后48小时,收获样品并提取基因组dna。
[1071]
基因组分析
[1072]
遵循制造商的方案,使用dna quickextract(lucigen)来分离基因组dna。在收获之后,将提取溶液在65℃孵育15分钟,在68℃孵育15分钟,随后在98℃孵育10分钟。基因组pcr使用可见于表1中的引物序列使用amplitaq gold 360主混合物(thermo fisher)来进
行。在sanger测序后,插入/缺失的存在通过ice(synthego)来分析。
[1073]
图38示出了带有23种不同的crispr off sgrna的cas9的编辑效率的图。从左至右,对于每种sgrna:黑色条(圆圈)表示在没有光暴露的情况下,与crispr off sgrna复合的cas9的编辑效率;灰色条(方块)表示在环境光暴露之后,与crispr off sgrna复合的cas9的编辑效率;浅灰色条(三角形)表示在暴露于波长大于345nm的光之后,与crispr off sgrna复合的cas9的编辑效率。如箭头所指出的,暴露之后,fancf和fam163位点未显示出编辑减少。使用的灯为600w,强度为90
‑
120mw/cm2。在所有实验中使用来自aldevron的具有核定位信号的cas9(nls
‑
sp.cas9
‑
nls)。
[1074]
图39示出了带有26种不同crispr off sgrna的cas9与带有含相同靶结合序列的未修饰的sgrna的cas9相比的编辑效率的图。
[1075]
图46示出了展示以下的图:与未暴露于光的表达与cas9复合的crispr off的细胞以及暴露于光和未暴露于光的表达与标准sgrna复合的cas9的细胞相比,在暴露于光之后的表达与cas9复合的crispr off的细胞中观察到的编辑百分比减少。
[1076]
图47示出了展示以下的图:在用光使测试细胞表达的cas9
‑
crispr off复合物使失活之前,随时间段的增加观察到的编辑百分比的增加。
[1077]
图53是靶向camk1的与cas9核酸酶复合的以上提及的多核苷酸相比于与cas9核酸酶复合的标准sgrna的插入/缺失谱。
[1078]
图51示出了光暴露持续时间对编辑消除的影响的图,其中在45
‑
60秒之间实现了完全消除。
[1079]
图52是示出增加细胞暴露于广谱光的时间对细胞存活力的影响的图。
[1080]
表1:靶序列引物
[1081]
[1082][1083]
实施例14:在暴露于uv光和未暴露于uv光的情况下,crispr
‑
off cas9复合物在u2os细胞中的中靶编辑的分析
[1084]
将u2os细胞在补充有10%v/v fbs的rpmi 1640中维持在第5
‑
15代之间。用tryple将细胞以1:4的比例每周一次进行传代。所有细胞都维持在37℃和5%co2。
[1085]
将u2os细胞用cas9和包含可光裂解(pc)接头的sgrna转染,并且经历用345nm带通滤光器过滤的光,以使接头裂解。将cas9与18种不同的sgrna(crispr off)复合以产生18种实验群体,所述sgrna(crispr off)在位置57和位置74处包含掺入的可光裂解接头(1
‑
(7
‑
(二乙基氨基)
‑2‑
氧代
‑
2h
‑
色烯
‑4‑
基)丙基),且具有靶向以下的靶结合区域:aavs1(ggggccacuagggacaggau)、bub1b(agugaagccaugucccugga)、camk1_sg1(ugccaggaucaccuccgaga)、camk1_sg2(gcguccucuuaucuucugcc)、chr8q23_sg1(uuauaguuacgauguuugau)、chr8q23_sg2(agucuacuaugaguuuucug)、dnmt1(ggagugagggaaacggcccc)、emx1(gaguccgagcagaagaagaa)、fam163a(cugcagggcucgcuggugag)、fancf(gcugcagaagggauuccaug)、grk1(gccgucaaagcugccucggg)、itga7(ggugcuggagggcgaggcug)、irak4(guccugucuuugucacagaa)、prgn(cagaugccugcucaguguug)、prkag3(agcaagaaaacagcagcuca)、stk3_sg1(aaagcaauacacaaggaauc)、stk3_sg2(ccauaaugcagcaaugugac)和vegfa(ggugagugagugugugcgug)。然后,将每种实验群体分成三组,一组保持在黑暗中,一组暴露于环境光,并且一组暴露于用345nm带通滤光器过滤以将波长限制在大于345nm的波长的光。为了形成18种复合物溶液中的每一种,将10pmol的cas9
蛋白与30pmol的sgrna混合。将每种溶液使用转染缓冲液稀释至20μl,并允许混合10分钟,然后转染。在转染之后4小时,将处理的细胞暴露于环境光20分钟,或者暴露于用345nm带通滤光器过滤以将波长限制在大于345nm的波长的光60秒。转染后48小时,收获样品并提取基因组dna。
[1086]
基因组分析
[1087]
遵循制造商的方案,使用dna quickextract(lucigen)来分离基因组dna。在收获之后,将提取溶液在65℃孵育15分钟,在68℃孵育15分钟,随后在98℃孵育10分钟。基因组pcr使用可见于表1中的引物序列使用amplitaq gold 360主混合物(thermo fisher)来进行。在sanger测序后,插入/缺失的存在通过ice(synthego)来分析。
[1088]
图40示出了带有18种不同crispr off sgrna的cas9与带有含相同靶结合序列的未修饰的sgrna的cas9相比的编辑效率的图。
[1089]
实施例15:在暴露于uv光和未暴露于uv光的情况下,crispr
‑
off cas9复合物在hep3b细胞中的中靶编辑的分析
[1090]
将hep3b细胞在高级改良eagles培养基(life technologies)和10%v/v fbs中维持在第5
‑
20代之间。用tryple(life technologies)将细胞以1:8的比例每周两次进行传代。
[1091]
将hep3b细胞用cas9和包含可光裂解(pc)接头的sgrna转染,并且经历用345nm带通滤光器过滤的光,以使接头裂解。将cas9与23种不同的sgrna(crispr off)复合以产生23种实验群体,所述sgrna(crispr off)在位置57和位置74处包含掺入的可光裂解接头(1
‑
(7
‑
(二乙基氨基)
‑2‑
氧代
‑
2h
‑
色烯
‑4‑
基)丙基),且具有靶向以下的靶结合区域:aavs1(ggggccacuagggacaggau)、bub1b(agugaagccaugucccugga)、camk1_sg1(ugccaggaucaccuccgaga)、camk1_sg2(gcguccucuuaucuucugcc)、cel(aaccaguugcaggcgcccca)、chr8q23_sg1(uuauaguuacgauguuugau)、cxcr4(gauaacuacaccgaggaaau)、emx1(gaguccgagcagaagaagaa)、fam163a(cugcagggcucgcuggugag)、fancf(gcugcagaagggauuccaug)、gaa(aggagccggugggagcaggg)、grk1(gccgucaaagcugccucggg)、itga7(ggugcuggagggcgaggcug)、irak4(guccugucuuugucacagaa)、mapre1(uucucugcagauaauuccug)、mip(gcugggguccucacugcgcu)、omp(gaacuguagccgcugcugcu)、opn1sw(acaggggcaaugugguacug)、prgn(cagaugccugcucaguguug)、prkag3(agcaagaaaacagcagcuca)、stk3_sg1(aaagcaauacacaaggaauc)、stk3_sg2(ccauaaugcagcaaugugac)和vegfa(ggugagugagugugugcgug)。然后,将每种实验群体分成三组,一组保持在黑暗中,一组暴露于环境光,并且一组暴露于用345nm带通滤光器过滤以将波长限制在大于345nm的波长的光。为了形成23种复合物溶液中的每一种,将10pmol的cas9蛋白与30pmol的sgrna混合。将每种溶液使用转染缓冲液稀释至20μl,并允许混合10分钟,然后转染。在转染之后4小时,将处理的细胞暴露于环境光20分钟,或者暴露于用345nm带通滤光器过滤以将波长限制在大于345nm的波长的光60秒。转染后48小时,收获样品并提取基因组dna。
[1092]
基因组分析
[1093]
遵循制造商的方案,使用dna quickextract(lucigen)来分离基因组dna。在收获
之后,将提取溶液在65℃孵育15分钟,在68℃孵育15分钟,随后在98℃孵育10分钟。基因组pcr使用可见于表1中的引物序列使用amplitaq gold 360主混合物(thermo fisher)来进行。在sanger测序后,插入/缺失的存在通过ice(synthego)来分析。
[1094]
图41示出了带有23种不同crispr off sgrna的cas9与带有含相同靶结合序列的未修饰的sgrna的cas9相比的编辑效率的图。
[1095]
实施例16:具有香豆素接头的crispr off sgrna暴露于可见光
[1096]
图34是在sgrna的位置57和位置74处包含香豆素接头(二乙基氨基香豆素(1
‑
(7
‑
(二乙基氨基)
‑2‑
氧代
‑
2h
‑
色烯
‑4‑
基)丙基))的crispr多核苷酸的图。香豆素接头明显红移,并且可以用于使用可见光裂解寡核苷酸。香豆素接头的释放通过形成紧密的离子对、随后香豆素基甲基阳离子与水和其他可用的亲核试剂反应来发生。
[1097]
电喷雾电离
[1098]
te缓冲液中的rna样品(3um)通过质谱(与agilent 6530b q
‑
tof质谱仪(ms)耦合的agilent 1290infinity ii液相色谱系统(lc))以负离子极性模式进行分析。lc在acquity uplc beh c18 vanguard pre
‑
column(1.7um,2.1
×
5mm)上用梯度洗脱(缓冲液a:50mm hfip;15mm己胺2%meoh;缓冲液b:meoh,0.75ml/min,2%
‑
95%b,在1.05min中)进行。电喷雾电离用双esi源(气体温度325℃,干燥气体12l/min,雾化器40psi,vcap 4kv,碎裂电压(fragmentor)250,锥孔(skimmer)65)进行。数据在100
‑
3200m/z范围内获得,并且在4000
‑
35000m/z范围内解卷积。
[1099]
图36a是以上描述的靶向vegfa(ggugagugagugugugcgug)的crispr off sgrna在暴露于光之前的esi示踪。
[1100]
图36b是以上描述的crispr off sgrna在暴露于通过420nm长通滤光器过滤的光之后的esi示踪。未经历光的crispr off sgrna保持了与未修饰的sgrna相同的分子量。对于未经历光的crispr off sgrna未观察到片段化。图20b展示了crispr off sgrna在暴露于420nm光时在两个可光裂解位点处被裂解。
[1101]
实施例17:具有uv可裂解接头的crispr off sgrna暴露于uv光
[1102]
电喷雾电离
[1103]
te缓冲液中的rna样品(3um)通过质谱(与agilent 6530b q
‑
tof质谱仪(ms)耦合的agilent 1290infinity ii液相色谱系统(lc))以负离子极性模式进行分析。lc在acquity uplc beh c18 vanguard pre
‑
column(1.7um,2.1
×
5mm)上用梯度洗脱(缓冲液a:50mm hfip;15mm己胺2%meoh;缓冲液b:meoh,0.75ml/min,2%
‑
95%b,在1.05min中)进行。电喷雾电离用双esi源(气体温度325℃,干燥气体12l/min,雾化器40psi,vcap 4kv,碎裂电压250,锥孔65)进行。数据在100
‑
3200m/z范围内获得,并且在4000
‑
35000m/z范围内解卷积。
[1104]
图57a是在位置57和74处具有可光裂解接头的靶向vegfa(ggugagugagugugugcgug)的crispr off sgrna在暴露于uv光之前的esi示踪。
[1105]
图57b是图41a的crispr off sgrna在暴露于通过345nm带通滤光器过滤的光之后的esi示踪。未经历光的crispr off sgrna保持了与未修饰的sgrna相同的分子量。图57b展示了crispr off sgrna在暴露于345nm光时在两个可光裂解位点处被裂解。
[1106]
实施例18:用uv光使与crispr off sgrna复合的cas9失活
[1107]
将10pmol nls
‑
cas9
‑
nls蛋白(aldevron)与30pmol的合成sgrna以20ul的总体积混合,并且允许复合10分钟。在该孵育期间,收获细胞并计数。向rnp溶液中添加5μl浓度为4
×
104个细胞/μl的细胞溶液,并且温和混合。
[1108]
细胞 rnp溶液使用20μl形式的4d
‑
nucleofector系统(lonza)进行转染。转染根据制造商的方案进行。转染后,使细胞在培养基中恢复,并且铺于96孔板中。
[1109]
crispr off失活使用sunray 600uv flood lamp(uvitron international)进行。345nm和355nm 6.5
”×
6.5”有色玻璃可选长通滤光器从newport.com获得,并且使用定制的3d打印容器安装。
[1110]
使用正立显微镜进行的失活使用配有colibri 7柔性光源和385nm led的zeiss axios observer来进行。
[1111]
图54是细胞培养物的图片,其中与cas9核酸酶复合的crispr off用于靶向必需基因。暴露于光( hv)的细胞培养物展示出比未暴露于光的细胞培养物更高的汇合,表明缺乏失活引起高度的细胞死亡。
[1112]
图42示出了与用标准sgrna观察到的比率相比,通过在脱靶编辑发生之前使crispr off sgrna失活来调节中靶编辑与脱靶编辑的比率。crispr off sgrna的失活通过在转染后的不连续时间照射细胞来实现。选择在如表2中可见的基因组中的一个或两个位点处具有显著水平的脱靶编辑的靶位点。
[1113]
图55是示出转染后在hek293细胞中在不同时间点的中靶编辑:脱靶编辑的比率的图。
[1114]
表2:脱靶位点
[1115]
靶靶序列mip_ot1agtggggtcctcactgcactmip_ot2tgtggggcactcactgcgctfam163_ot1ctgcagggcccgctggagagfam163_ot2ctgcaggggacactggtgagomp_ot1aggctgtagcccctgctgctomp_ot2gaactacagccactgctgctfancf_ot1gctgcagaagggattccaagmapre_ot1atctctgcagataatccctgopn1sw_ot1ttagaggcaatgtggtactgvegfa_ot1tgtgggtgagtgtgtgcgtg
[1116]
表3:脱靶测序引物
[1117][1118]
基因组分析
[1119]
遵循制造商的方案,使用dnaquickextract(lucigen)来分离基因组dna。在收获之后,将提取溶液在65℃孵育15分钟,在68℃孵育15分钟,随后在98℃孵育10分钟。基因组pcr使用可见于表1中的引物序列使用amplitaq gold 360主混合物(thermo fisher)来进行。在sanger测序后,插入/缺失的存在通过ice(synthego)来分析。
[1120]
实施例19:使用cas9
‑
crispr off进行gfp敲除
[1121]
将10pmol nls
‑
cas9
‑
nls蛋白(aldevron)与30pmol的合成sgrna以20ul的总体积混合,并且允许复合10分钟。在该孵育期间,收获细胞并计数。向rnp溶液中添加5μl浓度为4
×
104个细胞/μl的细胞溶液,并且温和混合。
[1122]
细胞 rnp溶液使用20μl形式的4d
‑
nucleofector系统(lonza)进行转染。转染根据制造商的方案进行。转染后,使细胞在培养基中恢复,并且铺于96孔板中。
[1123]
crispr off失活使用sunray 600uv flood lamp(uvitron international)进行。345nm和355nm 6.5
”×
6.5”有色玻璃可选长通滤光器从newport.com获得,并且使用定制的3d打印容器安装。
[1124]
使用正立显微镜进行的失活使用配有colibri 7柔性光源和385nm led的zeiss axios observer来进行。
[1125]
图43是示出在细胞中观察到的编辑百分比随着暴露于385nm的光的增加而降低的图。
[1126]
图44示出了细胞培养物,其中使用掩蔽物选择性地将表达crispr off sgrna的细胞暴露于光,以使暴露于光的细胞中编码gfp的基因失活,同时允许未暴露于光的细胞继续表达gfp。
[1127]
图24是施加至图44的细胞培养物的薄膜掩蔽物的图片,使得透明区域允许光通
过,使与crispr off复合的cas9核酸酶的编辑活性失活,而黑暗区域是不透明的,以允许编辑不受阻碍地进行。
[1128]
实施例20:用可见光使与crispr off sgrna复合的cas9失活
[1129]
图48示出了在位置57和位置74处具有香豆素接头的靶向mip的crispr off sgrna被led光源失活有多快速。用具有香豆素接头的crispr off sgrna转染的hek293细胞被分到五个独立的孔中。在四小时之后,成对的重复被覆盖以去除环境光或被暴露于430
±
23nm led 1min、2min、3min或4min。1分钟足以使基因编辑失活。colibri 7光源以430
±
23nm led的100%强度与能够一次照射单个孔的标准倒置荧光显微镜一起使用。
[1130]
实施例21:sgrna上多个接头位置的测试.
[1131]
rnp形成和递送
[1132]
将10pmol nls
‑
cas9
‑
nls蛋白(aldevron)与30pmol的合成sgrna以20ul的总体积混合,并且允许复合10分钟。在该孵育期间,收获细胞并计数。向rnp溶液中添加5μl浓度为4
×
104个细胞/μl的细胞溶液,并且温和混合。
[1133]
细胞 rnp溶液使用20μl形式的4d
‑
nucleofector系统(lonza)进行转染。转染根据制造商的方案进行。转染后,使细胞在培养基中恢复,并且铺于96孔板中。
[1134]
crispr off失活
[1135]
crispr off失活使用sunray 600uv flood lamp(uvitron international)进行。345nm和355nm 6.5
”×
6.5”有色玻璃可选长通滤光器从newport.com获得,并且使用定制的3d打印容器安装。
[1136]
使用正立显微镜进行的失活使用配有colibri 7柔性光源和385nm led的zeiss axios observer来进行。
[1137]
基因组分析
[1138]
遵循制造商的方案,使用dna quickextract(lucigen)来分离基因组dna。在收获之后,将提取溶液在65℃孵育15分钟,在68℃孵育15分钟,随后在98℃孵育10分钟。基因组pcr使用可见于表1中的引物序列使用amplitaq gold 360主混合物(thermo fisher)来进行。在sanger测序后,插入/缺失的存在通过ice(synthego)来分析。
[1139]
图50a
‑
图50c示出了观察到的如以上描述的与cas9核酸酶复合的18种不同sgrna的编辑百分比。图50a示出了在与cas9核酸酶复合的各自靶向dnmt1的6种不同的sgrna中观察到的编辑百分比。sgrna是标准的(mod)或在位置21、24、50、57或74处具有单个可裂解接头。图50b示出了在与cas9核酸酶复合的各自靶向fancf的6种不同的sgrna中观察到的编辑百分比。sgrna是标准的(mod)或在位置21、24、50、57或74处具有单个可裂解接头。图50c示出了在与cas9核酸酶复合的各自靶向vegfa的6种不同的sgrna中观察到的编辑百分比。sgrna是标准的(mod)或在位置21、24、50、57或74处具有单个可裂解接头。
[1140]
实施例22:液滴pcr检测暴露于光之后crispr off的片段化
[1141]
数字液滴pcr
[1142]
在没有dna酶的情况下使用rna quickextract(lucigen)来提取细胞rna。rna使用ribogreen(thermo fisher)来定量和归一化。
[1143]
crispr off失活使用sunray 600uv flood lamp(uvitron international)进行。345nm和355nm 6.5
”×
6.5”有色玻璃可选长通滤光器从newport.com获得,并且使用定制的
3d打印容器安装。
[1144]
使用正立显微镜进行的失活使用配有colibri 7柔性光源和385nm led的zeiss axios observer来进行。
[1145]
总rna使用iscript advanced cdna合成试剂盒(biorad)以0.4um逆转录引物进行逆转录。逆转录产物使用2
×
evagreen ddpcr主混合物并以以下热循环进行扩增:在95℃持续3分钟,随后为95℃持续30秒和52.4℃持续1分钟的40个循环。然后信号在4℃稳定5分钟,随后在90℃失活5分钟。然后,将液滴通过qx200液滴数字pcr系统(biorad)进行读取。
[1146]
表4:ddpcr试剂:
[1147]
引物名称序列sgrna_fagagctagaaatagcaagttaaasgrna_rgactcggtgccacttt
[1148]
图45示出了展示与标准sgrna相比暴露于光之后crispr off丰度降低的图。
[1149]
实施例23:1
‑
(7
‑
(二乙基氨基)
‑2‑
氧代
‑
2h
‑
色烯
‑4‑
基)丙基的制备
[1150]
亚磷酰胺化合物3(3
‑
(双(4
‑
甲氧基苯基)(苯基)甲氧基)
‑1‑
(7
‑
(二乙基氨基)
‑2‑
氧代
‑
2h
‑
色烯
‑4‑
基)丙基(2
‑
氰乙基)二异丙基亚磷酰胺)遵循wenzel等人(2003)(nucleosides,nucleotides&nucleic acids,vol.22,nos.5
‑
8,第1579
‑
1581页)中公开的方法,通过使醛化合物1(7
‑
(二乙基氨基)
‑2‑
氧代
‑
2h
‑
色烯
‑4‑
甲醛)与烯丙基三甲基硅烷在ticl4存在的情况下反应来合成。接下来,二醇化合物2(7
‑
(二乙基氨基)
‑4‑
(1,3
‑
二羟基丙基)
‑
2h
‑
色烯
‑2‑
酮)通过前一化合物的臭氧分解和用nabh4进行还原处理产生。2的二甲氧基三苯甲基化,随后是亚磷酰化(phosphitylation),以优异的产率得到亚磷酰胺化合物3。
[1151][1152]
实施例24:将1
‑
(7
‑
(二乙基氨基)
‑2‑
氧代
‑
2h
‑
色烯
‑4‑
基)丙基连接至核苷酸
[1153]
在添加化合物3之后形成的带有接头的rna的dmt(dmt=4,4
’‑
二甲氧基三苯甲基)保护基团在酸催化的脱三苯甲基反应中被去除。脱三苯甲基的rna已准备好与以核苷亚磷酰胺单体的形式添加的核苷酸反应。将适当的核苷亚磷酰胺与激活物(四唑或衍生物)混合,两者都溶解在乙腈中。核苷亚磷酰胺的二异丙基氨基基团被激活物质子化,并且从而转化为良好的离去基团。该离去基团被脱三苯甲基的rna的脱保护的羟基基团对离去基团相邻磷原子的攻击快速替代,并且形成新的磷氧键,产生亚磷酸三酯键(如紧接的下图中示出的)。核苷亚磷酰胺在惰性气氛中相当稳定,并且可以以大量制备。
[1154][1155]
x可以是h、otbdms(o
‑
叔丁基二甲基硅烷基醚)或ome。
[1156]
在一些实施方案中,亚磷酰胺接头化合物3的二异丙基氨基基团被激活物质子化,并且从而转化为良好的离去基团。该离去基团被核苷碱基的3’或5’羟基基团快速替代,并且形成新的磷氧键(如紧接的下图中示出的)
[1157][1158]
x可以是h、otbdms(o
‑
叔丁基二甲基硅烷基醚)或ome。
[1159]
本领域技术人员应当理解,前述实施例中描述的亚磷酰胺方法通常包括四个步骤:步骤1(脱三苯甲基)、步骤2(偶联)、步骤3(加帽)和步骤4(氧化)。
[1160]
实施例25:用uv光使与crispr off sgrna连接的cas9失活
[1161]
将nls
‑
cas9
‑
nls蛋白(aldevron)与合成的crispr off sgrna组合,该合成的crispr off sgrna包含被配置为与cas9形成共价键的接头和在位置54和位置74处的可光裂解接头。sgrna通过接头与cas9核酸酶共价连接以形成连接的rnp复合物。
[1162]
用连接的rnp复合物转染细胞。转染后,使细胞在培养基中恢复,并且铺于96孔板中。将细胞培养48小时,以允许rnp复合物编辑靶序列。
[1163]
使用sunray 600uv flood lamp(uvitron international)用345nm和355nm 6.5
”×
6.5”有色玻璃可选长通滤光器使crispr复合物失活。在使用该泛光灯(flood lamp)之前和之后以一定的时间间隔收集细胞。
[1164]
可选地,使用正立显微镜进行的失活使用配有colibri 7柔性光源和385nm led的zeiss axios observer来进行。从收获的细胞中提取核酸,并且用于测量在该时间间隔的编辑效率。
[1165]
实施例26:使用crispr on v1 sgrna变体进行靶编辑
[1166]
图58示出了crispr on v1 sgrna的结构,所述crispr on v1 sgrna的结构与酿脓链球菌cas9(spcas9)sgrna相同,但crispr on v1 sgrna包含与前间区互补的20个核苷酸(nt)序列(折回序列),随后是紧接前间区5’侧的4nt环结构。图59示出了使用靶向四个独特基因座的crispr on v1 sgrna的三种变体进行的靶编辑。三种变体包括:一种预期对光刺激没有响应(非可裂解对照),一种在距sgrna的5’末端的位置24处包含单个可光裂解接头(1个可光裂解),以及一种在位置11和位置24处包含两个可光裂解接头(2个可光裂解)。将crispr on sgrnagrna与spcas9混合以形成rnp并转染到hek293中。4小时后,将细胞分为两
个实验组,一组经历光照射,并且一组置于黑暗中。转染后48小时,从两个组中分离基因组dna并分析插入/缺失的存在。前间区序列的互补序列的掺入降低了所分析的所有靶处的编辑效率。将可光裂解接头掺入到发夹中,并且任选地掺入到前间区的互补序列中未导致编辑的完全中断,但也没有允许在暴露于光之后编辑的完全恢复。
[1167]
实施例27:使用crispr on v2 sgrna进行靶编辑
[1168]
图60示出了crispr on v2 sgrna的结构,所述crispr on v2 sgrna的结构使用与crispr on v1相同的结构,但crispr on v2 sgrna用2
’‑
o
‑
甲基(2`o
‑
me)rna替代折回序列(与前间区互补的前20nt的rna)。2`o
‑
me rna与rna更紧密结合,并且在r
‑
环形成期间较不可能被替代。图61示出了标准sgrna(mod)、crispr on v1(rna)和crispr on v2(o
‑
me)之间编辑活性的比较。将crispr on rnp转染到hek293中并且允许其恢复。转染后48小时,收获基因组dna并分析插入/缺失的存在。crispr on v2显著降低了测试的所有5个基因座处的插入/缺失的诱导。将crispr on rnp转染到hek293中并且允许其恢复。转染后48小时,收获基因组dna并分析插入/缺失的存在。crispr on v2显著降低了测试的所有5个基因座处的插入/缺失的诱导。
[1169]
实施例28:使用crispr on v3 sgrna进行靶编辑
[1170]
图62示出了crispr on v3 sgrna的结构,所述crispr on v3 sgrna的结构基于crispr on v2构建,但crispr on v3 sgrna在前间区折回序列的中间(位置11)和紧接前间区序列的5’侧(位置24)掺入了可光裂解接头。图63示出了与在位置24(k24)处包含单个可光裂解接头的crispr on v2变体、crispr on v2(o
‑
me)和标准sgrna(mod)相比,使用靶向5个独特基因座的crispr on v3 sgrna(k11,24)进行的编辑。将可光裂解接头掺入到crispr on v3中允许部分恢复编辑活性。将包含crispr on v3 sgrna的rnp转染到hek293细胞中。转染后4小时,将池分为两组,其中一组暴露于光,而另一组保持在黑暗中。转染后48小时,从所有组中提取dna并分析插入/缺失的存在。置于黑暗中的crispr on v3 sgrna一致地展示出低至检测不到的编辑水平。当相同的样品暴露于光时,在基因组dna中检测到增加的插入/缺失水平。这种插入/缺失存在的增加在缺少可光裂解接头的引导中未被观察到。
[1171]
实施例29:使用crispr on v4 sgrna变体进行靶编辑
[1172]
图64示出了crispr onv4 sgrna的结构,所述crispr on v4 sgrna的结构基于crispr on v3构建,但引入了另外的可光裂解接头,以确保折回区域被dna靶有效替代。可光裂解残基位于位置23和位置24处,以增加折回序列从sgrna释放的可能性。另外的可光裂解残基位于位置6和位置14处,以辅助解离。图65示出了与5rp(在sgrna的5’侧包含另外的序列5
’‑
ucucccugagcuucagggag
‑3’
)、crispr on v2(me)和标准sgrna(mod)相比,使用crispr on v4 sgrna变体在两个基因座处进行的编辑。crispr on v4 sgrna变体在以下核苷酸处包含可光裂解接头:3、23和24(k3,23,24);6、11、16、23和24(k6,11,16,23,24);6、14、23和24(k6,14,23,24)。将与crispr on v4 sgrna形成的rnp转染到hek293细胞中。转染后1小时,将细胞分为两组。一组暴露于靶向的光,而另一组保持在黑暗中。转染后48小时,从两个组中分离dna并分析插入/缺失的存在。在位置23和位置24处添加可光裂解接头显示出增加了在所有靶处的编辑效率的恢复。通过在位置6和位置14处包含可光裂解接头的进一步优化允许编辑活性有效恢复至与标准(mod)sgrna相似的编辑活性。
[1173]
材料和方法:
[1174]
crispr on sgrna合成
[1175]
所有rna均使用synthego的crisprevolution平台通过固相亚磷酰胺化学合成,并且rna的身份通过电喷雾电离质谱(esi
‑
ms)证实。
[1176]
细胞培养
[1177]
将人类胚胎肾细胞(hek293)在高级改良eagles培养基(advanced modified eagles medium,life technologies)和10%v/v fbs中维持在第5
‑
20代之间。用tryple(life technologies)将细胞以1:8的比例每周两次进行传代。所有细胞都维持在37℃和5%co2。
[1178]
rnp形成和递送
[1179]
将10pmol酿脓链球菌nls
‑
spcas9
‑
nls蛋白(aldevron cat#9212)与30pmol的合成sgrna(synthego)以20ul的总体积混合,并且允许复合10分钟。在该孵育期间,收获细胞并计数。向rnp溶液中添加5μl浓度为4
×
104个细胞/μl的细胞溶液,并且温和混合。
[1180]
细胞 rnp溶液使用20μl形式的4d
‑
nucleofector系统(lonza)进行转染。hek293转染使用方案cm
‑
130在sf缓冲液中进行。转染后,使细胞在培养基中恢复,并且铺于96孔板中。为了创建成对的重复,将转染产物贴印到(stamp into)第二个96孔板中,并且允许独立恢复。
[1181]
sgrna激活
[1182]
crispr on激活通过定制的arduino控制的光板进行。光板包含24个独特的单独控制的具有420
‑
430nm的led。将96孔板照射4
‑
7分钟以激活sgrna。
[1183]
基因组分析
[1184]
遵循制造商的方案,使用dna quickextract(lucigen)来分离基因组dna。在收获之后,将提取溶液在65℃孵育15分钟,在68℃孵育15分钟,随后在98℃孵育10分钟。基因组pcr使用可见于补充表2中的引物序列使用amplitaq gold 360主混合物(thermo fisher)来进行。在sanger测序后,插入/缺失的存在通过ice(synthego)来分析。
[1185]
虽然本文已经示出和描述了本发明的优选实施方案,但对于本领域技术人员将明显的是,这样的实施方案仅通过实例的方式提供。在不偏离本发明的情况下,本领域技术人员现在将想到许多变化、改变和替代。应当理解,在实践本发明时可以以任何组合采用本文描述的本发明的实施方案的各种替代。所附权利要求意图限定本发明的范围,并且从而涵盖在这些权利要求的范围内的方法和结构及其等同物。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。