核苷酸序列的应用
1.本发明为中国专利“自主异花授粉及自主单性结实番茄的培育方法”(申请日:2019.11.19,申请号:201911117060.0)的分案申请,
技术领域
2.本发明属于作物育种技术领域,具体涉及一种核苷酸序列的应用,尤其涉及一种核苷酸序列在培育番茄自主异花授粉及自主单性结实系中的应用。
背景技术:
3.番茄是全球消费最多的蔬菜之一,中国是世界上番茄栽培面积最大、生产总量最多的国家。作为未来市场占据主导地位的蔬菜,番茄在生产、栽培等方面仍存在两个重大难题:1、坐果率低,需人工点花;2、柱头无外露,需人工去雄,耗费大量人力成本。
4.为了提高番茄坐果率,一般采用以下传统单性结实方法进行:授粉刺激,即给番茄柱头授以不亲和花粉或无活力花粉,虽未受精但由于授粉本身的刺激可形成单性结实果;2)通过人工振动植株或振动花序刺激子房发育诱导形成单性结实果;3)化学诱导,即在花柄或花梗处涂抹赤霉素等激素诱导番茄单性结实。但上述传统单性结实方法都存在操作复杂、单性结实率低、激素浓度难掌握、耗费人力物力等缺点。为解决上述问题,现已有科研人员采用基因敲除技术对番茄基因组中的单性结实基因进行编辑,使其丧失功能得到番茄单性结实系,达到提高坐果率的目的,但由于其在靶位点选择及靶向序列设计等方面仍存在缺陷,导致利用该方法培育得到的番茄单性结实系在坐果率、尤其在高低温逆境条件下的坐果率仍不理想。
5.因此,如何开发出一种新的靶位点选择及靶向序列设计策略将其合理应用于培育番茄单性结实系当中是解决上述问题的关键。
技术实现要素:
6.本发明针对现有的番茄单性结实系的培育方法存在高低温等逆境条件下坐果率不理想等的技术问题,提出一种核苷酸序列的应用,将该核苷酸序列应用于番茄自主异花授粉及自主单性结实系的培育中,能够提高番茄在高低温等逆境条件下的坐果率,且无其他不良性状。
7.为了达到上述目的,本发明采用的技术方案为:
8.一组核苷酸序列在培育番茄自主异花授粉自主单性结实系中的应用,所述核苷酸序列包括:如seq id no.6、seq id no.2所示的核苷酸序列或如seq id no.7、seq id no.8所示的核苷酸序列;
9.其中,所述番茄自主异花授粉自主单性结实系通过以下方法培育得到,包括:
10.从番茄基因组上选取多个靶位点,并构建用于敲除靶位点的载体,所述靶位点的核苷酸序列为如seq id no.6、seq id no.2所示的核苷酸序列或如seq id no.7、seq id no.8所示的核苷酸序列;
11.将构建所得的载体导入番茄外植体进行组织培养获得转化苗;
12.从所述番茄转化苗中筛选出阳性株系,经鉴定后,得到所述番茄自主异花授粉自主单性结实系。
13.作为优选,所述多个靶位点选自番茄基因组中的agl6基因和ms10基因或者f3h基因和gst基因。
14.作为优选,如seq id no.6所示的靶位点纯合敲除的株系是自主异花授粉自主单性结实系,如seq id no.6所示的靶位点杂合敲除的株系是自主异花授粉自主单性结实系的恢复系。
15.作为优选,所述番茄自主异花授粉自主单性结实系的繁育方法,包括以下步骤:
16.将所述番茄自主异花授粉自主单性结实系与其恢复系进行杂交,获得子代植株,根据所述子代植株的茎秆颜色进行筛选。
17.作为优选,所述子代植株的茎秆颜色为绿色时,其为番茄自主异花授粉自主单性结实系,所述子代植株的茎秆颜色为紫色时,其为番茄自主异花授粉自主单性结实系的恢复系。
18.本发明还提供了一组核苷酸序列在培育番茄自主单性结实系中的应用,所述核苷酸序列包括:如seq id no.1、seq id no.2以及seq id no.3所示的核苷酸序列;
19.其中,所述番茄自主单性结实系通过以下方法培育得到,包括:
20.从番茄基因组上选取多个靶位点,并构建用于敲除靶位点的载体,其中,所述靶位点的核苷酸序列如seq id no.1、seq id no.2以及seq id no.3所示;
21.将构建所得的载体导入番茄外植体进行组织培养获得转化苗;
22.从所述番茄转化苗中筛选出阳性株系,经鉴定后,得到所述番茄自主单性结实系。
23.作为优选,所述多个靶位点选自番茄基因组上的agl6基因。
24.作为优选,敲除所用方式为crispr/cas9系统。
25.作为优选,将构建所得的载体导入番茄受体时,所用的导入方法为农杆菌侵染法。
26.与现有技术相比,本发明的优点和积极效果在于:
27.1、本发明通过从番茄基因组上的agl6基因选择多个适宜的靶位点,得到相关的靶位点核苷酸序列,并将其合理利用于番茄自主单性结实系的培育之中进行串联敲除,从而得到一种新的番茄自主单性结实系,该结实系无需点花或授粉即可完成自主性单性结实、坐果率高(尤其是高低温环境)、柱头外露等性状,且无其他不良性状的产生;
28.2、本发明通过从番茄基因组上的agl6基因和ms10基因或者f3h基因和gst基因选择多个适宜的靶位点,得到相关的靶位点核苷酸序列,并将其合理利用于番茄自主异花授粉自主单性结实系的培育之中进行双敲除,从而得到一种新的番茄自主异花授粉自主单性结实系,不会产生花粉且可以自主进行单性结实,有利于进行其品种保护。
附图说明
29.图1为本发明实施例所提供的载体35s:cas9
‑
a3图谱;
30.图2为本发明实施例所提供的番茄自主单性结实系的酶切和测序鉴定图谱;
31.图3为本发明实施例所提供的番茄自主单性结实系的表型示意图;
32.图4为本发明实施例所提供的番茄自主单性结实系在正常条件下的表型示意图;
33.图5为本发明实施例所提供的不同品种番茄自主单性结实系花部位表型示意图;
34.图6为本发明实施例所提供的不同株系单性结实率结果图;
35.图7为本发明实施例所提供的番茄难结籽自主单性结实系或杂交种的繁育流程;
36.图8为本发明实施例所提供的载体ybk
‑
ms10
‑
agl6的图谱;
37.图9为本发明实施例所提供的番茄自主异花授粉自主单性结实系的表型结果;
38.图10为本发明实施例所提供的番茄自主异花授粉自主单性结实系的繁育流程;
39.图11为本发明实施例所提供的番茄自主单性结实系的indel标记的鉴定图谱及对其验证的测序图谱;
40.图12为本发明实施例所提供的番茄自主异花授粉自主单性结实系及其恢复系的区分表型;
41.图13为本发明实施例所提供的基因ms10和基因f3h之间遗传距离示意图。
具体实施方式
42.下面将对本发明实施例中的技术方案进行清楚、完整地描述,显然,所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动前提下所获得的所有其他实施例,都属于本发明保护的范围。
43.本发明实施例提供了一组核苷酸序列在培育番茄自主异花授粉自主单性结实系中的应用,所述核苷酸序列包括:如seq id no.6、seq id no.2所示的核苷酸序列或如seq id no.7、seq id no.8所示的核苷酸序列;
44.其中,所述番茄自主异花授粉自主单性结实系通过以下方法培育得到,包括:
45.(1)从番茄基因组上选取多个靶位点,并构建用于敲除靶位点的载体,所述靶位点的核苷酸序列为如seq id no.6、seq id no.2所示的核苷酸序列或如seq id no.7、seq id no.8所示的核苷酸序列;
46.(2)将构建所得的载体导入番茄外植体进行组织培养获得转化苗;
47.(3)从所述番茄转化苗中筛选出阳性株系,经鉴定后,得到所述番茄自主异花授粉自主单性结实系。
48.在上述实施例中,本发明提供了一组核苷酸序列在培育番茄自主异花授粉自主单性结实系中的应用,其中,选用靶位点如seq id no.6、seq id no.2所示的核苷酸序列进行agl6基因和ms10基因双敲除的原因在于:番茄是一种自花授粉作物,敲除ms10可以实现柱头外露异花授粉,敲除agl6可以实现自主单性结实,同时敲除二者,可以创造番茄自主异花授粉自主单性结实系,且两个基因之间不会产生干扰。同时,本发明所选择的靶点选取标准为:靶点位于基因编码区上游,潜在脱靶可能性小。
49.本发明选用靶位点如seq id no.7、seq id no.8所示的核苷酸序列进行f3h基因和gst基因双敲除的原因在于:为了便于后续筛选出番茄自主异花授粉自主单性结实系和其恢复系,本发明选择可ms10基因附近的可见连锁标记基因f3h和gst基因,f3h基因与ms10基因遗传距离较远,同时敲除f3h和gst基因可以降低可见表型与ms10基因的分离率。同时,本发明所选择的靶点选取标准:位于基因编码区上游,潜在脱靶可能性小。
50.在一优选实施例中,所述多个靶位点选自番茄基因组中的agl6基因和ms10基因或
者f3h基因和gst基因。
51.在一优选实施例中,如seq id no.6所示的靶位点纯合敲除的株系是自主异花授粉自主单性结实系,如seq id no.6所示的靶位点杂合敲除的株系是自主异花授粉自主单性结实系的恢复系。
52.在一优选实施例中,所述番茄自主异花授粉自主单性结实系的繁育方法,包括以下步骤:
53.将所述番茄自主异花授粉自主单性结实系与其恢复系进行杂交,获得子代植株,根据所述子代植株的茎秆颜色进行筛选。
54.在一优选实施例中,所述子代植株的茎秆颜色为绿色时,其为番茄自主异花授粉自主单性结实系,所述子代植株的茎秆颜色为紫色时,其为番茄自主异花授粉自主单性结实系的恢复系。
55.本发明实施例还提出了一组核苷酸序列在培育番茄自主单性结实系中的应用,所述核苷酸序列包括:如seq id no.1、seq id no.2以及seq id no.3所示的核苷酸序列;
56.其中,所述番茄自主单性结实系通过以下方法培育得到,包括:
57.(1)从番茄基因组上选取多个靶位点,并构建用于敲除靶位点的载体,其中,所述靶位点的核苷酸序列如seq id no.1、seq id no.2以及seq id no.3所示;
58.(2)将构建所得的载体导入番茄外植体进行组织培养获得转化苗;
59.(3)从所述番茄转化苗中筛选出阳性株系,经鉴定后,得到所述番茄自主单性结实系。
60.在上述技术方案中,本发明提出了一组核苷酸序列在培育番茄自主单性结实系中的应用,其中,选择靶位点如seq id no.1、seq id no.2以及seq id no.3所示的核苷酸序列进行agl6基因串联敲除的原因在于:作为多肽/蛋白质一级结构的氨基酸序列是多肽/蛋白质空间结构的基础,而空间结构又直接决定其功能,氨基酸序列的微小变化可能会导致空间结构的巨变,进而导致功能发生改变,难以预见单独的靶点突变后氨基酸序列组成会产生座果率表现好的单性结实系,因此在agl6基因上选择三个靶点,为了获得不同突变的核苷选组成,获得座果率表现好的单性结实系。
61.还需进一步说明的是,靶位点选择标准为:因现有技术中两个靶点所培育得到的番茄单性结实系座果率不理想,本发明选择三个靶位点可以产生更多丰富的突变类型,能够从中筛选出座果率表现好的单性结实系。
62.在一优选实施例中,所述多个靶位点选自番茄基因组上的agl6基因。
63.在一优选实施例中,敲除所用方式为crispr/cas9系统。
64.在一优选实施例中,将构建所得的载体导入番茄受体时,所用的导入方法为农杆菌侵染法。
65.为了更清楚详细地介绍本发明实施例所提供的核苷酸序列的应用,下面将结合具体实施例进行描述。
66.实施例1
67.本实施例提出了一种番茄自主单性结实系的培育方法,该方法从番茄基因组上的agl6基因上选取了3个靶位点,分别是如seq id no.1所示的agl6
‑
target1核苷酸序列、如seq id no.2所示的agl6
‑
target2核苷酸序列以及如seq id no.3所示的agl6
‑
target3核
苷酸序列,将上述3个靶位点构建成串联敲除载体,经农杆菌侵染番茄外植体,经筛选获得番茄自主单性结实系,具体步骤如下:
68.(1)引物设计及载体构建:
69.采用已经商业化的bgk01载体试剂盒进行构建,针对agl6
‑
target1设计的连接引物如下表所示:
70.表1 agl6
‑
target1连接引物序列
71.序列名称序列agl6
‑
sg
‑
f15
’‑
tgattgaggattaaggcaacaacgt
‑3’
agl6
‑
sg
‑
r15
’‑
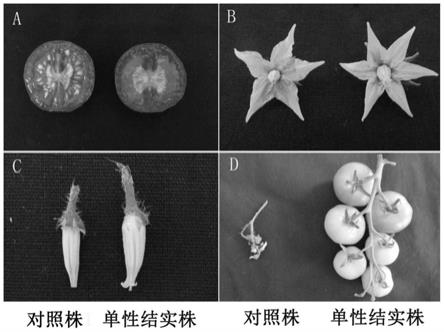
aaacacgttgttgccttaatcctca
‑3’
72.将上述引物agl6
‑
sg
‑
f1和agl6
‑
sg
‑
r1以表2所示的反应体系混合,95℃加热3min,然后以约0.2℃/s缓慢降至20℃,形成oligo二聚体;
73.表2反应体系
[0074][0075][0076]
以表3所示的反应体系在冰上混合,于室温反应1h,然后按照大肠杆菌转化标准步骤转化trans1
‑
t1 phage resistant chemically competent cell;
[0077]
表3反应体系
[0078]
组分体积(μl)h2o6crispr/cas9 vector2oligo二聚体1enzyme mix1总计10
[0079]
抽提包含agl6
‑
target1靶点的编辑载体质粒(命名为:35s:cas9
‑
a1),用hindⅲ酶切待用;
[0080]
本实施例针对agl6
‑
target2和agl6
‑
target3设计的连接引物如下表所示:
[0081]
表4 agl6
‑
target2和agl6
‑
target3连接引物序列
[0082][0083][0084]
以seq id no.4所示的核苷酸序列为模板,分别以agl6
‑
u6
‑
f和agl6
‑
u6
‑
r2、agl6
‑
u6
‑
f和agl6
‑
u6
‑
r3为引物进行pcr扩增,得到两个扩增片段,分别命名为:u6
‑
2和u6
‑
3;
[0085]
以seq id no.5所示的核苷酸序列为模板,分别以agl6
‑
sgrna
‑
f2和agl6
‑
sgrna
‑
r、agl6
‑
sgrna
‑
f3和agl6
‑
sgrna
‑
r为引物进行pcr扩增,得到两个扩增片段,分别命名为:sgrna
‑
2和sgrna
‑
3;
[0086]
分别以u6
‑
2和sgrna
‑
2、u6
‑
3和grna
‑
3为模板,以agl6
‑
u6
‑
f和agl6
‑
sgrna
‑
r为引物进行重叠pcr,分别得到两个扩增产物,分别命名为:agl6
‑
u6
‑
sgrna
‑
2和agl6
‑
u6
‑
sgrna
‑
3;
[0087]
上述pcr扩增的反应体系如表5所示,pcr扩增程序如下:94℃2min;94℃15s,55℃30s,68℃30s,35个循环;68℃10min。
[0088]
表5反应体系
[0089][0090][0091]
将重叠pcr产物agl6
‑
u6
‑
sgrna
‑
2与35s:cas9
‑
a1线性化大片段用重组酶试剂盒(peasy
‑
uni seamless cloning and assembly kit)进行重组,50℃重组15min,然后转化测序,将正确的测序载体用hindⅲ限制性内切酶酶切,以同样的方法连接agl6
‑
u6
‑
sgrna
‑
3,并测序,将正确的测序载体命名为35s:cas9
‑
a3(其图谱如图1所示),除了靶位点之外,还包括rna聚合酶ⅲ识别的启动子atu6驱动的向导rna、35s启动子驱动的cas9核酸酶编码基因、35s启动子驱动的潮霉素筛选基因。
[0092]
(2)农杆菌侵染:
[0093]
将上述质粒35s:cas9
‑
a3转化gv3101农杆菌感受态,挑取阳性菌落培养至对数中期,离心,用ms培养基悬浮至od
600
值为0.2,将外植体浸泡悬浮液5min,25℃暗处理共培养2天。
[0094]
(3)再生及筛选:
[0095]
将共培养后的外植体移至含15mg/l hyg的再生培养基(ms培养基4.3g/l,蔗糖30g/l,玉米素2mg/l,100mg/l肌醇,1x尼许维生素,0.8%琼脂)中,每两周继代一次直至长出抗性芽。
[0096]
(4)生根:
[0097]
将抗性芽移至生根培养基(ms培养基4.3g/l,蔗糖30g/l,1x尼许维生素,0.8%琼脂)中,约20天左右,将生根苗移至育苗钵中。
[0098]
(5)阳性株及靶点基因型的鉴定:
[0099]
以上述番茄抗性芽发育成的抗性株的dna为模板,利用阳性鉴定引物和进行pcr鉴定,阳性鉴定引物序列如表6所示,将pcr阳性苗用bar试纸条进行复筛,获得番茄阳性株。
[0100]
表6阳性鉴定引物
[0101][0102][0103]
以上述阳性株dna为模板,利用扩增引物进行pcr扩增,其中,扩增引物的序列如表7所示,将pcr产物用限制性内切酶aclⅰ进行酶切,条带类型为503bp和173bp的为对照株,条带类型为676bp、503bp和173bp的为杂合编辑株,条带类型为676bp的为纯合编辑株;
[0104]
阳性株基因型鉴定结果如图2所示,图2a为t0代阳性株基因型鉴定结果,图2b为筛选所得的t0
‑
5测序鉴定结果(即番茄自主单性结实系),t0
‑
5缺失2个碱基,造成密码子移码,丧失原有的蛋白功能,导致雌性器官激素失调,无需花粉受精子房提前膨大。同时根据多靶点突变,还获得了大片段删除的编辑类型如图11所示。
[0105]
表7阳性株扩增引物
[0106][0107]
(6)番茄自主单性结实系表型的鉴定:
[0108]
本实施例培育得到的番茄自主单性结实系呈现兼性单性结实株系的表型,如图3所示,其中,t0
‑5‑
t1兼性单性结实株系表现为子房自主膨大,因此无种子产生(如图3a所示,图3中,对照株系是h18 wt株系),同时柱头外露有利于异株授粉(如图3c所示),花瓣相对于对照株系表现为淡黄色(如图3b所示)。
[0109]
(7)自主单性结实系种子的生产方法:
[0110]
本实施例培育出的自主单性结实系结籽困难,但是可以通过与相应敲除杂合的株系杂交进行扩繁,其流程如图7所示。
[0111]
还需进一步说明的是,本发明参照实施例所述的方法同时对不同的番茄品种中培育了番茄单性结实系,得出了以下结论:1、不同品种的单性结实系果实膨大的时间不同,仅有一个品种(h18)表现为子房提前膨大且具有很高的单性结实率,另外两个品种(h17/ailsa craig)表现为子房膨大较晚(如图5所示),这一点是非常关键的,果实膨大的早晚直接影响了何时提前进行授粉留种的关键问题;2、不同品种柱头外露情况不同,本发明发现仅h18表现为全部柱头外露,h17和ailsa craig表现为部分柱头外露且不明显(如图5所示),柱头外露对制种非常重要,可以节约大量的人工去雄或剥开花瓣等工作量,能很好地保证雌蕊不受损,使种子的质量、纯度、产量都大幅度提高。在目前人工成本越来越高的情况下,该特性具有很高的实用价值。
[0112]
本发明实施例选择了agl6基因的三个靶位点目的在于:验证是否不同编辑类型会产生不同的表型效果,本发明培育出了h17不同编辑基因型的株系,同时统计了其单性结实系率,结果表明不同株系单性结实率差别很大(如图6所示),在实际应用中需要进行筛选强
单性结实系后进行下一步育种操作。
[0113]
实施例2
[0114]
本实施例对实施例1培育得到的番茄自主单性结实系t0
‑5‑
t1进行不同生长条件下的坐果率测定,具体为:
[0115]
(1)实验分组:
[0116]
实验组1:t0
‑5‑
t1兼性单性结实株系在非正常环境下生长,生长条件为:植物生长室,高温32
‑
35℃每天持续6
‑
8h、弱光;
[0117]
实验组2:t0
‑5‑
t1兼性单性结实株系在非正常环境下生长,生长条件为:植物生长室,低温10
‑
20℃,每天持续24h、弱光;
[0118]
对照组:t0
‑5‑
t1兼性单性结实株系在正常环境下(山东潍坊温室大棚10月份定植)生长;
[0119]
(2)坐果率测定方法:坐果率=果实膨大数/总花数;
[0120]
(3)坐果率测定结果,如下表所示:
[0121]
表8实验组与对照组的坐果率
[0122][0123][0124]
由上表数据结合附图3
‑
4可知,当t0
‑5‑
t1兼性单性结实株系在非正常环境下生长(高温弱光)时,表现为高的坐果率(如图3d所示),在正常环境下(山东潍坊温室大棚10月份定植)同样表现为高座果率(如图4所示,其中,对照株系是h18 wt株系)。
[0125]
实施例3
[0126]
本实施例提出了一种番茄自主异花授粉自主单性结实系的培育方法,从番茄基因组上选取了2个靶位点,分别是核苷酸序列如seq id no.6所示的ms10
‑
target1和核苷酸序列如seq id no.2所示的agl6
‑
target2,将上述2个靶位点构建成双敲除载体,经农杆菌侵染番茄外植体,经筛选获得番茄自主异花授粉自主单性结实系,具体为:
[0127]
(1)引物设计及载体构建:
[0128]
首先,以cpb载体为基础载体,参照yan et al.,2015.high
‑
efficiency genome editing in arabidopsis using yao promoter
‑
driven crispr/cas9 system.molecular plant,8(12),1820
‑
1823,将cas9的启动子pubi更换成pyao启动子,改造后的载体命名为ybk载体;
[0129]
参照实施例1所述的方法,以ybk载体为基础构建载体,其中针对ms10
‑
target1、agl6
‑
target2和sgrna设计的扩增引物如下:
[0130]
表9 ms10
‑
target1、agl6
‑
target2和sgrna的扩增引物序列
[0131][0132][0133]
最终成功构建的载体被命名为ybk
‑
ms10
‑
agl6,其图谱如图8所示,除了靶位点之外,还包括rna聚合酶ⅲ识别的启动子atu6驱动的向导rna、pyao启动子驱动的cas9核酸酶编码基因、35s启动子驱动的卡那霉素筛选基因。
[0134]
(2)番茄遗传转化:
[0135]
利用ybk
‑
ms10
‑
agl6载体,参照实施例1所述的番茄遗传转化步骤进行。
[0136]
(3)阳性株及靶点基因型的鉴定:
[0137]
以上述番茄抗性芽发育成的抗性株的dna为模板,利用阳性鉴定引物进行pcr鉴定,将pcr阳性苗用bar试纸条进行复筛,获得番茄阳性株,其中,阳性鉴定引物序列如下表所示:
[0138]
表10阳性鉴定引物序列
[0139][0140][0141]
以上述阳性株dna为模板,分别利用扩增引物进行扩增,测序,鉴定出双基因敲除纯合株系,其中,扩增引物序列如下表所示:
[0142]
表11扩增引物序列
[0143]
引物名称序列信息ms10
‑
鉴定
‑
f5
’‑
acaagaagattcccacaaccaag
‑3’
ms10
‑
鉴定
‑
r5
’‑
cacttgaacttcaggctctatgc
‑3’
agl6
‑
鉴定
‑
f25
’‑
ggaggaatggtttgttgaag
‑3’
agl6
‑
鉴定
‑
r25
’‑
agttactatgaaccccgtatga
‑3’
[0144]
(4)番茄自主异花授粉自主单性结实系表型的鉴定:
[0145]
培育获得的双基因敲除株系为自主异花授粉自主单性结实系,表现为自主性单性结实且花粉无活性、柱头外露(如图9所示)具体地,其花柱外露(如图9b所示),可以作为母本利于生产杂交种;而且,它单性结实子房自主膨大(如图9a所示),所以可以不接受花粉,实现常规种保护;另外,它无花粉(如图9c所示),所以不能作为父本进行授粉,实现对目标品系的严格保护。
[0146]
(5)番茄自主异花授粉自主单性结实系的繁育:
[0147]
如图10所示,番茄自主异花授粉自主单性结实系不会产生种子且花粉无活性,生产上需要杂合有野生型ms10基因(即未经ms10
‑
target1突变)的恢复系(aabb)进行扩繁,f1代50%为aabb,50%为aabb,该过程需要通过分子标记或可见标记对番茄自主异花授粉自主单性结实系及其恢复系进行分辨,所以开发相应标记是番茄自主异花授粉自主单性结实系的繁育的关键。
[0148]
实施例4
[0149]
本实施例提供了一种番茄基因组上ag6基因和ms10基因的双敲除策略的筛选方法,具体为:
[0150]
(1)实验分组及实验方法:
[0151]
实验组1:以实施例1所述的方法,在番茄基因组上的agl6基因上选取了3个靶位点,分别是如seq id no.1所示的agl6
‑
target1核苷酸序列、如seq id no.2所示的agl6
‑
target2核苷酸序列以及如seq id no.3所示的agl6
‑
target3核苷酸序列,将上述3个靶位点构建成串联敲除载体,再参照实施例2所述的方法,在ms10基因上选择1个靶位点,如seq id no.6所示的ms10
‑
target1核苷酸序列,将上述1个靶位点构建敲除载体,共同敲除番茄基因组上的agl6基因和ms10基因,经农杆菌侵染番茄外植体,经筛选获得番茄自主单性结实系,具体操作步骤同实施例1;
[0152]
实验组2:在番茄基因组上的agl6基因上选取了1个靶位点,如seq id no.1所示的agl6
‑
target1核苷酸序列,构建敲除载体,在在番茄基因组上的ms10基因选择1个靶位点,
如seq id no.6所示的ms10
‑
target1核苷酸序列,敲除载体,共同共同敲除番茄基因组上的agl6基因和ms10基因,经农杆菌侵染番茄外植体,经筛选获得番茄自主单性结实系,具体操作步骤同实施例1
‑
2;
[0153]
实验组3:在番茄基因组上的agl6基因上选取了1个靶位点,如seq id no.2所示的agl6
‑
target2核苷酸序列,构建敲除载体,在在番茄基因组上的ms10基因选择1个靶位点,如seq id no.6所示的ms10
‑
target1核苷酸序列,敲除载体,共同共同敲除番茄基因组上的agl6基因和ms10基因,经农杆菌侵染番茄外植体,经筛选获得番茄自主单性结实系,具体操作步骤同实施例1
‑
2;
[0154]
实验组4:在番茄基因组上的agl6基因上选取了1个靶位点,如seq id no.3所示的agl6
‑
target3核苷酸序列,构建敲除载体,在在番茄基因组上的ms10基因选择1个靶位点,如seq id no.6所示的ms10
‑
target1核苷酸序列,敲除载体,共同共同敲除番茄基因组上的agl6基因和ms10基因,经农杆菌侵染番茄外植体,经筛选获得番茄自主单性结实系,具体操作步骤同实施例1
‑
2;
[0155]
(2)坐果率测定方法及测定结果:
[0156]
坐果率测定方法:坐果率=果实膨大数/总花数;
[0157]
表12实验组及对照组的坐果率测定结果
[0158]
实验分组坐果率/%实验组190实验组292实验组370实验组475
[0159]
实施例5
[0160]
本实施例提出了一种番茄自主单性结实系的标记开发,具体为:
[0161]
(1)番茄自主单性结实系的indel标记:
[0162]
本实施例设计了番茄自主单性结实系的插入
‑
缺失鉴定引物,上下游引物序列如下所示:
[0163]
表13插入
‑
缺失鉴定引物
[0164]
引物名称引物序列agl6
‑
鉴定
‑
f35
’‑
agcttgactcccatacacct
‑3’
agl6
‑
鉴定
‑
r35
’‑
tgcttcaaacttggccttta
‑3’
[0165]
利用agl6
‑
鉴定
‑
f3和agl6
‑
鉴定
‑
r3进行pcr扩增,得到676bp条带的为未编辑株系,得到676bp条带和452bp条带的为杂合敲除突变体,只有452bp条带的为纯合删除突变体(如图11a所示),其中t0
‑
3株系的测序验证结果如图11b所示,利用该indel标记可以有效的区分番茄自主单性结实系的突变类型。
[0166]
(2)番茄自主异花授粉自主单性结实系的绿色茎秆连锁标记:
[0167]
参照实施例2的方法,对核苷酸序列如seq id no.7所示的f3h
‑
target1和核苷酸序列如seq id no.8所示的gst
‑
target1这两个靶位点进行进一步双敲除,如图12所示,纯合突变的番茄自主异花授粉自主单性结实系的茎秆为绿色,其杂合突变的恢复系的茎秆为紫色,十分容易区分自主异花授粉自主单性结实系(aabb)和恢复系(aabb);ms10和f3h之间
遗传距离为5.8cm(如图13所示),理论上两者发生遗传交换的概率为5.8%,也就是说通过f3h表型筛选不育系理论上会有5.8%的出错率,本发明找到一个与ms10更接近的可见表型基因gst(gst与ms10的遗传距离仅为2.2cm),大大减少了可见标记与目标性状分离的几率,更具实用性。进一步,为了加强可见表型的可辨识率,同时达到双保险的目前的,本发明将ms10、gst、f3h同时敲除,获得了理想的改良效果。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。