一种番茄肌醇半乳糖苷合成酶基因slgols2在调控果实着色与成熟中的应用
技术领域
1.本发明涉及植物基因工程技术领域,具体涉及一种番茄肌醇半乳糖苷合成酶基因slgols2在调控果实着色与成熟中的应用。
背景技术:
2.番茄果实的风味独特,能够为人类提供多种营养元素,如多种维生素、碳水化合物、类胡萝卜素、酚类化合物等,可以起到抗氧化、抗血栓、抗过敏和抗炎症等作用(raiola et al., 2014)。可鲜食、烹饪、制成番茄酱等多种食用方式(li et al., 2018),是人们每日饮食必不可少的果蔬之一。番茄的产量和种植面积在全世界范围内始终处于增长水平(quinet et al., 2019)。
3.番茄果实的发育起始于花分生组织,此时的组织和结构决定了果实生长的状态。果实生长是果实发育过程中最长的阶段,根据不同的基因型生长时长可达5至8周。花后1至2周是果实生长的初始阶段,此时细胞进行强烈的有丝分裂,果皮细胞层数增多,并伴随着细胞膨大(cheniclet et al., 2005)。随后,细胞分裂逐渐减缓,进入细胞膨大期,此时果实的体积和重量显著增加并达到最终大小,进入完全绿熟(mature green,mg)时期(pesaresi et al., 2014)。进入mg时期几天后,果实的代谢物质开始了大量重组,标志着果实成熟过程的开始。主要包括破色(breaker,br)阶段和成熟(ripening,rr)阶段。果实进入破色期后,叶绿素降解,类胡萝卜素大量积累,果实逐渐软化,并伴随着呼吸和乙烯释放量的激增形成高峰,这是呼吸跃变型果实的显著特征(alexander and grierson, 2002)。营养物质积累,风味、质地、颜色和香气发生变化,最终赋予番茄特有的品质和营养价值。
4.番茄果实的风味、色泽的形成和营养物质的积累与成熟阶段密切相关,明确果实成熟的分子机制才能为改善果实品质提供理论依据,与番茄果实成熟相关的研究始终吸引着众多学者的目光(yuan et al., 2018; ji et al., 2020; wu et al., 2020)。在番茄果实逐渐成熟的过程中,色泽的变化是最明显的,色泽发生变化的内在原因是果实中叶绿素降解同时类胡萝卜素积累。
5.类胡萝卜素的合成和积累在质体中进行完成(sun et al., 2018)。植物中存在着多种质体,包括:叶绿体、有色体、原质体、淀粉体等(lopez
‑
juez and pyke, 2005; jarvis and lopez
‑
juez, 2013)。其中原质体是还未分化的质体,是其他质体的前体结构。类胡萝卜素大量存在于叶绿体和有色体中(egea et al., 2010)。但类胡萝卜素的颜色在叶绿体中通常被叶绿素遮盖住,在有色体中才充分显露出来。番茄果实中的有色体通常由叶绿体转化而来,在番茄果实成熟过程中,果皮细胞中叶绿素降解造成叶绿体的类囊体结构瓦解,从而逐步转变为有色体同时积累大量类胡萝卜素(min and chieri, 2008; egea et al., 2011)。大量研究表明番茄果实中叶绿素的含量和叶绿体结构往往影响着番茄果实成熟后的类胡萝卜素含量。de
‑
etiolated1(det1)基因是光信号转导负调节因子,番茄high pigment 2(hp2)突变体在mg时期表现出深绿色,这是det1基因发生突变的结果。mg时期hp2
果实中叶绿体体积增大,数量增多,叶绿素含量达到野生型番茄的2.9倍,待果实完全成熟后,类胡萝卜素水平显著提高(mustilli et al., 1999; bino et al., 2005)。high pigment 1(hp1)突变体与hp2表型类似,是uv
‑
damaged dna
‑
binding protein 1(ddb1)基因发生突变的结果(liu et al., 2004; bernhardt et al., 2006)。番茄中存在两个golden2
‑
like(glk)基因,其中glk2基因已被证实与果实叶绿素积累和叶绿体发育相关,超表达slglk2基因会使果实中叶绿素含量增加,叶绿体体积增大,类囊体数目增多,从而提高成熟时期果实中的类胡萝卜素含量(powell et al., 2012; nguyen et al., 2014)。arabidopsis pseudo response regulator2
‑
like(aprr2
‑
like)与glk基因结构类似,超表达slaprr2
‑
like基因能够增加番茄果实的质体数量、面积和类胡萝卜素含量(pan et al., 2013)。tkn2和tkn4是knox家族中的两个转录因子,研究表明这作用于slglk和slaprr2
‑
like上游,间接影响质体发育,类胡萝卜素积累(nadakuduti et al., 2014)。
6.肌醇半乳糖苷合成酶(galactinol synthase,gols)属于糖基转移酶家族8(glycosyltransferase family 8,gt8),该家族包含多种参与寡糖、多糖和糖结合物生物合成的酶,在植物生长发育的过程中起着重要作用。rfos是植物中重要的可溶性碳水化合物,包括三糖棉子糖(raffinose)、四糖水苏糖(stachyose)和五糖毛蕊花糖(verbascose)(sengupta et al., 2015; jorge et al., 2017)。具有储备碳水化合物、稳定膜结构、抵御逆境胁迫、参与渗透调节的功能(li et al., 2017a; 韦玉霞, 2017)。gols能够催化半乳糖苷的转移,以udp
‑
半乳糖(udp
‑
galactose,udp
‑
gal)为糖基供体,将半乳糖苷转移至肌醇(myo
‑
inositol)合成肌醇半乳糖苷(galactinol,gol)(sengupta et al., 2012),为rfos的生物合成提供重要前体物质。这一反应是rfos合成过程中的关键步骤,也是限速步骤(sengupta et al., 2015)。gols作为rfos生物合成的关键酶,通过影响rfos的含量,参与调节植物生长发育以及抵御胁迫。在番茄中存在4个gols基因,目前对这4个基因的功能尚未有报道。
技术实现要素:
7.发明目的:本发明提供一种番茄肌醇半乳糖苷合成酶基因slgols2在调控果实着色与成熟中的应用,其目的在于调控番茄果实绿果肩形成及果实成熟进程。
8.技术方案:番茄肌醇半乳糖苷合成酶基因slgols2在调控果实着色与成熟中的应用。
9.所述slgols2基因核苷酸序列如seq id no.1所示。
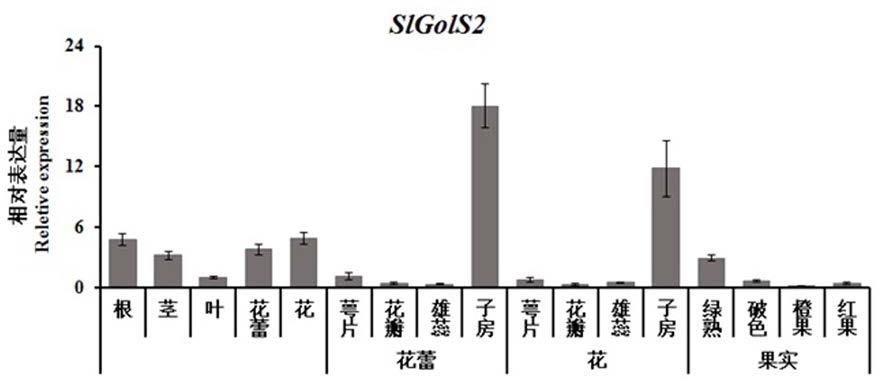
10.slgols2基因表达水平在花蕾的子房和花的子房中高,在子房发育成的果实中的表达量表现出低水平。
11.所述番茄为野生型番茄ailsa craig。
12.利用crispr
‑
cas9技术构建基因slgols2敲除突变体,获得两个纯合突变株系slgols2#23和slgols2#24;slgols2#23株系中,靶点位置前插入4个碱基ttta,替换1个碱基t
→
g,靶点位置缺失5个碱基atata;slgols2#24株系中,靶点位置前插入4个碱基ttta,靶点位置缺失12个碱基gatgatata。
13.有益效果:本发明中基因表达量检测结果显示,slgols2基因在番茄子房中特异性表达,而番
茄果实发育起始于子房,这说明该基因在番茄果实发育过程中潜在重要作用。
14.利用crispr
‑
cas9技术构建slgols2敲除突变体,突变体果实表现为绿熟期无绿果肩、转色提前并提早成熟。即番茄slgols2基因缺失后消除番茄果实绿果肩形成,促进果实成熟。
15.基于上述研究成果,可将slgols2基因通过crispr
‑
cas9基因编辑技术敲除后,用于培养番茄绿熟期无绿果肩的早熟品种。
附图说明
16.图1为本发明slgols2基因在野生型番茄ailsa craig不同组织部位的表达模式分析;图2为本发明slgols2基因突变方式;图3为本发明slgols2与野生型番茄表型;图4为本发明slgols2与野生型番茄叶绿素含量,其中,图4a是番茄各部分果皮;图4b是番茄果实绿熟期果皮叶绿素含量;图5为本发明slgols2与野生型番茄叶绿素合成前体含量,其中,图5a是δ
‑
氨基酮戊酸含量,图5b是胆色素原含量,图5c是尿卟啉原iii含量,图5d是尿卟啉原iii含量,图5e是镁原卟啉ix含量,图5f是原叶绿素酸酯;图6为本发明slgols2与野生型番茄显微镜观察,其中图6a
‑
c为叶绿素荧光观察;图6d
‑
f为质体观察;图6g
‑
i为叶绿体结构观察;图6j
‑
l为类囊体结构观察;图7为本发明slgols2与野生型番茄果实的乙烯释放量;图8为本发明slgols2与野生型番茄果实的硬度;图9为本发明slgols2与野生型番茄果实的色度;图10为本发明slgols2与野生型番茄果实总类胡萝卜素的含量;图11为本发明slgols2与野生型番茄果实番茄红素的含量。
具体实施方式
17.以下结合说明书附图更详细的说明本发明。
18.本发明中slgols2基因属于糖基转移酶家族8,通过限速调节棉子糖系列寡糖的生物合成,参与调节植物生长发育,抵御逆境胁迫。在番茄子房中特异性表达,本发明利用crispr
‑
cas9基因编辑技术获得的slgols2突变番茄不具有绿果肩,并且果实提前转色成熟。基于此研究结果,可为番茄果实绿果肩形成和果实成熟的分子机制研究,加快番茄早熟品种的选育提供新的应用基础。
19.本发明提供了一种番茄肌醇半乳糖苷合成酶基因slgols2,所述slgols2基因具有seq id no.1所示的核苷酸序列。所述番茄肌醇半乳糖苷合成酶基因slgols2在调控果实着色与成熟中的应用。
20.番茄材料为野生型番茄ailsa craig,slgols2突变番茄株系(slgols2#23和slgols2#24);实验试剂:超纯rna提取试剂盒(ultrapure rna kit),购自康为世纪;
实时荧光定量试剂盒(tbgreenpremixextaq),购自takara;dnamarker2000,2xtaqpcrmastermix,购自中科瑞泰;其他分子生物学试剂,购自sigma;其他化学试剂,购自科隆。
21.实施例1基因表达的特殊部位或时期,往往决定了该基因的潜在生物学功能。因此,本发明测定了slgols2基因在野生型番茄的各个组织部位中的表达含量,实验步骤如下:取野生型番茄根、茎、叶片、花蕾、花、花蕾的花器官(萼片、花瓣、雄蕊和子房)和花的花器官(萼片、花瓣、雄蕊和子房)组织,提取rna,反转录为cdna后,稀释至3倍用slgols2基因的qrt
‑
pcr引物进行扩增;qrt
‑
pcr引物序列如下:slgols2f:5
’‑
ttgtcaagccgtgcctatgt
‑3’
(seqidno.2);slgols2r:5
’‑
caggcaaacaagccacaaca
‑3’
(seqidno.3);qrt
‑
pcr反应体系:2
×
realmastermix10
µ
l,primerf(10mm)0.4
µ
l,primerr(10mm)0.4
µ
l,cdna3
µ
l,ddh2o6.2
µ
l;qrt
‑
pcr反应程序:95℃30s,95℃5s,60℃30s,45个循环,melt15s;slgols2基因在野生型番茄各组织中的表达情况如图1所示,slgols2基因在番茄的各个组织部位均有表达,特别的是,slgols2基因表达水平在花蕾的子房和花的子房中均较高,而在子房发育成的果实中的表达量表现出低水平。
22.实施例2由百格基因公司构建slgols2基因的crispr
‑
cas9敲除载体,通过农杆菌介导法转入野生型番茄中。发明人对slgols2t0代突变体进行纯化,筛选出稳定遗传的纯合株系,实验步骤如下:采用ctab法提取slgols2突变番茄幼嫩叶片基因组dna,具体步骤包括:将番茄叶片置于ctab提取液中研磨,65℃水浴后加入饱和酚,振荡混匀离心后,转移上清液至新的离心管,加入异丙醇并置于
‑
20℃,5min后离心,转移上清至新的离心管中,用75%乙醇漂洗两次,晾干乙醇后,加入ddh2o溶解dna,
‑
20℃保存;ctab提取液配制方法:tris1.21g,edta
‑
na20.74g,dtt1g,ctab2g,nacl4.1g,蒸馏水定容至100ml。水系滤膜过滤后室温保存;将提取的dna进行扩增用于后续测序分析,用slgols2基因的pcr引物进行扩增;pcr引物序列如下:slgols2f:5
’‑
atgtttttggtcttgcaactaaggc
‑3’
(seqidno.4);slgols2r:5
’‑
acaattagagtccagaaaaagggct
‑3’
(seqidno.5);pcr反应体系:2
×
taqpcrmaster12.5
µ
l,primerf(10mm)1
µ
l,primerr(10mm)1
µ
l,dna1
µ
l,ddh2o9.5
µ
l;pcr反应程序:95℃5min,95℃30s,57℃30s,72℃1min,共35个循环,72℃5min,16℃10min;得到的pcr产物送至北京诺赛基因组研究中心有限公司进行dna测序。利用dnaman软件分析测序结果,比对靶点位置碱基序列,如图2。获得两个纯合突变株系slgols2#23和
slgols2#24。slgols2#23株系中,靶点位置前插入4个碱基(ttta),替换1个碱基(t
→
g),靶点位置缺失5个碱基(atata);slgols2#24株系中,靶点位置前插入4个碱基(ttta),靶点位置缺失12个碱基(gatgatata)。
23.实施例3对slgols2突变体和野生型番茄表型进行观察比较,结果表明,slgols2基因能够影响番茄果实叶绿素积累、叶绿体发育及果实成熟,相关实验步骤如下:番茄果实表型观察和成熟时间统计:为了对番茄果实成熟过程进行统一标定,在番茄花瓣完全展开当天对花朵进行标记,之后的时间记录为开花后天数(days post anthesis,dpa)。选取相同dpa的果实进行表型观察、生理指标测定和分子生物实验。当番茄果实表现出肉眼可见的一抹黄时标记为破色期并记录此时的生长天数,作为统计番茄果实成熟时间的依据;叶绿素含量测定:取番茄各部位果皮0.1 g,加入提取液(v丙酮:v无水乙醇=2:1)5 ml,放置在黑暗处24 h,待样品全部脱色变白后,测定663 nm和645 nm波长处的吸光度。每次测定3次重复;叶绿素合成前体含量测定:δ
‑
氨基酮戊酸(ala)含量测定:将番茄果皮称重后,加入4%三氯乙酸研磨,将研磨后的溶液离心,转移上清至新的离心管,并加入醋酸钠和乙酰丙酮,之后置于在沸水浴下反应10 min,待冷却后加入ehrhch
‑
hg试剂,暗处显色15 min,测定553 nm波长下的吸光值;胆色素原(pbg)含量测定:将番茄果皮称重后加入提取液(0.6 mol
·
l
‑
1 tris,0.1 mol
·
l
‑
1 edta,ph 8.2)充分研磨,将研磨后的溶液离心,转移上清至新的离心管,加入ehrhch
‑
hg试剂,暗处显色15 min,测定553 nm波长下的吸光值;尿卟啉原iii(urogeniii)含量测定:将番茄果皮称重后加入提取液(0.067 mol
·
l
‑
1 磷酸缓冲液,ph 6.8)研磨提取。将研磨后的溶液离心,转移上清至新的离心管,加入1% na2s2o3溶液,置于强光下照射20 min后,用冰醋酸(1 mol
·
l
‑1)将ph调至3.5后,用乙醚萃取3次,合并后测定水相在405.5 nm波长处的吸光值;原卟啉ix(proto ix)、镁原卟啉ix(mg
‑
proto ix)和原叶绿素酸酯(pchlide a)含量测定:将番茄果皮称重后,加入80%碱性丙酮后在冰上充分研磨,将研磨后的溶液离心,转移上清至新的离心管,用80%碱性丙酮定容,测定od575、od590和od628下吸光值;激光共聚焦显微镜观察:取开花后30天的番茄果实,用干净的手术刀快速切割下一层薄薄的番茄果皮立即放置在载玻片上。滴加pbs缓冲液,轻轻盖上盖玻片。在激光共聚焦显微镜下观察;质体观察:取开花后30天的番茄果实,用干净的手术刀将果皮部分切成2 mm
×
1 mm
×
1 mm的立方小块,迅速置于装3.5%的戊二醛溶液中,使样品完全浸没于溶液中,置于黑暗中反应1h。将样品转移至0.1 mol
·
l
‑
1 edta
‑
na2(ph 9.0)溶液中,置于65 ℃水浴锅中,20
‑
30 min。(样品置于在0.1 mol
·
l
‑
1 edta
‑
na2(ph 9.0)溶液中后,可在4 ℃保存半年。)用移液器将未完全分散的细胞分离开,吸取少量组织于载玻片上,轻轻盖上盖玻片,使细胞完全分散为单细胞。在显微镜下观察并拍照;透射电子显微镜观察:取开花后30天的番茄果实,用干净的手术刀将果皮部分切成2 mm
×
1 mm
×
1 mm的立方小块,迅速置于装有戊二醛溶液的青霉素瓶中。用50 ml注射器
抽出青霉素瓶中的空气,使样品完全浸没于溶液中,避光保存于4 ℃。后续漂洗、固定、脱水、包埋、切片和染色步骤由沈阳农业大学分析测试中心完成。在日立ht7700透射电子显微镜下观察叶绿体结构并拍照;总类胡萝卜素含量测定:取番茄果实赤道处果皮1 g,加入提取液(v正己烷:v丙酮=6:4)充分研磨后,涡旋振荡2 min,4000 g离心5 min。将上清液转移至新的离心管中,向沉淀中再加入提取液研磨,涡旋振荡,离心,合并上清液。直至色素完全洗脱下来,沉淀为白色。测定450 nm处的吸光值,总类胡萝卜素浓度计算公式为:总类胡萝卜素浓度(mg
·
ml
‑1)=od450/0.25;番茄红素含量测定:取番茄果实赤道处果皮1 g,加入10ml提取液(v正己烷:v无水乙醇:v丙酮=2:1:1,在丙酮中提前加入0.05%的bht抗氧化剂)充分研磨后涡旋振荡15 min。加入3 ml去离子水后再振荡2 min,1000 g离心1 min至溶剂分层。吸取上层溶液(正己烷),用正己烷调零,测定503 nm波长下的吸光值。番茄红素计算公式为:番茄红素浓度(mg
·
ml
‑1)=a503
×
3.12;利用tms
‑
pilot质构仪对番茄果实的硬度进行测定。设置参数:起始力0.75 n,输入速度30 mm
·
min
‑1,穿刺距离8 mm,输出速度30 mm
·
min
‑1,回程距离50 mm。将番茄果实赤道处置于探头下,记录穿刺过程中最大压力值,每个果实沿赤道至少测定3个点;乙烯释放量测定:将采摘下来的番茄置于室温放置2 h,以充分释放伤乙烯。测定果实的体积和质量。将果实密封于300 ml的保鲜盒内,置于25 ℃培养箱中4 h。用注射器抽取1 ml气体,经高效气相色谱仪检测每个样品中的乙烯峰面积,再根据乙烯标准曲线计算乙烯浓度,根据果实的体积和质量确定果实的乙烯释放量;色度测定:利用色差仪对番茄果实的色度进行测定。沿赤道处测定番茄果实的色度,记录l、a和b数值,每个果实沿赤道至少测定3个点。根据(pan et al., 2013)公式计算番茄色度(tomato color index,tci):tci=);通过slgols2突变株系的表型观察发现(图3),在果实绿熟期,slgols2突变果实表现为比wt果实更浅的绿色,同时不存在野生型番茄深绿色的果肩。slgols2突变番茄果实提前4
‑
6天破色。
24.slgols2突变株系中叶绿素含量减少,特别是果肩处。原本野生型特有的深绿色果肩,在slgols2番茄果实中不复存在(图4);slgols2突变株系中叶绿素合成过程中的6种前体物质含量也减少(图5);通过多种显微镜对slgols2突变株系和野生型番茄果皮细胞的内部结构观察,结果显示(图6):slgols2突变株系中叶绿素荧光水平降低,细胞内质体呈现为浅绿色至白色,同时叶绿体减小,类囊体垛堞层数减少;乙烯释放量结果显示(图7):野生型番茄在42 dpa时乙烯释放量激增,slgols2#24和slgols2#23番茄分别在38 dpa和36 dpa时乙烯释放量激增,均早于野生型番茄。野生型番茄在高峰期的乙烯释放量约为12.32 μl
∙
kg
‑1∙
h
‑1,slgols2#24和slgols2#23的释放量分别约为7.65 μl
∙
kg
‑1∙
h
‑1和8.50 μl
∙
kg
‑1∙
h
‑1,均低于野生型番茄。这些结果显示slgols2基因敲除后使乙烯释放量提早激增,减少乙烯的释放量,表明slgols2基因能够影响乙烯激增的发生时间和乙烯释放量;
硬度测定结果显示(图8):wt、slgols2#24和slgols2#23番茄硬度分别在40 dpa、36 dpa和34 dpa后开始大幅度下降,在44 dpa、42 dpa和40 dpa后趋向平稳,在48 dpa后,三者的硬度无明显差异。这些结果显示slgols2基因敲除后使果实硬度提早降低,但不影响果实完全成熟后的硬度;色度测定结果显示(图9):wt、slgols2#24和slgols2#23番茄果实色度值分别40dpa、36 dpa和34 dpa后开始大幅度下降,在46 dpa、42 dpa和40 dpa后趋向平稳,在48 dpa后,三者的tci值无明显差异。这些结果显示slgols2基因敲除后使果实提前转色,但不影响果实完全成熟后的色度;类胡萝卜素含量测定结果显示(图10和图11),42 dpa时slgols2#24和slgols2#23中总类胡萝卜素和番茄红素含量均显著高于野生型番茄,但48 dpa时slgols2#24和slgols2#23与野生型番茄果皮中的总类胡萝卜素含量和番茄红素含量并没有显著差异;综上所述,slgols2基因缺失后可以消除番茄果实绿果肩,促进果实提早成熟。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。