鉴定或表征受试者中的免疫应答的方法
1.本发明提供了一种通过使用例如胰蛋白酶消化片段通过质谱来定量抗原特异性免疫球蛋白来鉴定和定量受试者中的免疫应答的方法。还提供了计算机实施的方法和在所述方法中使用的测定试剂盒。所述方法尤其可用于表征免疫应答,例如抗病毒,真菌和细菌感染,自身抗原和其它免疫应答,包括对候选疫苗的表征。
2.人免疫球蛋白含有通过二硫键结合在一起的两个相同的重链多肽和两个相同的轻链多肽。有两种不同的轻链同种型(κ和λ)和五种不同的重链同种型(igg、iga、igm、igd和ige)。
3.igg提供了大多数抗侵入病原体的基于抗体的免疫。
4.iga倾向于存在于粘膜区域,例如肠道,呼吸道和泌尿生殖道,并且经常防止病原体的定殖。它也在唾液,泪液和母乳中发现。通常发现它是通过所谓的j链肽连接的二聚体。
5.igm作为单体在b细胞的表面上表达并且作为五聚体以分泌形式表达,并在b细胞介导的(体液)免疫的早期阶段消除病原体。igm的五聚体通过j链肽连接在一起。
6.igd是这样的一种抗体,其构成约1%的未成熟b淋巴细胞的质膜中的蛋白质,并且构成约0.25%的血清免疫球蛋白。
7.ige在哺乳动物中被发现并且经常参与对寄生虫(例如蠕虫)的免疫,并且还在对过敏原的多种类型的超敏反应中起作用。
8.igg具有四个亚类,igg1、igg2、igg3和igg4。根据铰链区的大小,链间二硫键的位置,恒定区的氨基酸序列和分子量来区分这些亚类。亚类在激活补体的能力和它们与fc受体的结合方面也不同。
9.igg1占主要亚类igg的60-65%,并且主要负责针对蛋白质和多肽抗原的胸腺介导的免疫应答(也称为t细胞介导的免疫应答)。它也参与补体级联的调理和活化。
10.igg2占主要亚类的20-25%,并且是针对碳水化合物和多糖抗原的普遍的免疫应答。在所有igg同种型缺陷中,igg2的缺陷是最常见的,并且与婴儿中的反复气道和呼吸道感染有关。
11.igg3占总igg的约5-10%,并且在针对蛋白质或多肽抗原的免疫应答中起主要作用。
12.igg4通常占少于总igg的4%。igg4主要不与多糖结合。在igg4相关疾病(rd)中观察到升高的igg4水平,这是具有特征性组织病理学表象的免疫介导和慢性纤维炎性病况。血清igg4的定量包括在迄今可用的所有igg4-rd诊断指南中,包括1型自身免疫性胰腺炎(aip),它是igg4-rd的胰腺表现,尽管在所有igg4-rd患者中未观察到升高的igg4。最近的研究表明,在患有由浸润igg阳性浆细胞引起的硬化性胰腺炎,胆管炎和间质性肺炎的患者中发现了升高的igg4血清水平。igg4的精确作用仍然是未知的。
13.表1显示了人体igg抗体同种型或亚类的比较。这包括不同亚类对不同抗原的应答的概述。其还包括它们的补体激活和与参与抗原识别的fc受体结合的概述。后者位于例如b淋巴细胞,杀伤细胞,巨噬细胞,嗜中性粒细胞和肥大细胞的膜上。这种受体识别抗体的fc片段。
14.iga,其分泌形式(作为二聚体)也被称为siga,具有两个亚类,iga1和iga2。iga是补体系统的不良激活剂,而仅微弱地调理。尽管iga1在血清中占优势(约80%),但iga2在分泌物中的百分比高于血清中的百分比(约35%在分泌物中)。iga1是血清中发现的主要iga亚类,其中大多数淋巴组织具有大部分产生iga1的细胞。在一种同种异型iga2m(1)中,重链和轻链没有用二硫化物连接,而是用非共价键连接。在分泌性淋巴组织,例如肠相关淋巴组织中,iga2产生的份额大于非分泌性淋巴器官,例如脾和外周淋巴结中的份额。多糖抗原比蛋白质抗原更倾向于诱导更多的iga2。
15.在哺乳动物中还没有发现另外两种抗体同种型。igy在鸟和爬行动物中发现的,并且与哺乳动物igg有关。igd在鲨鱼和鳐鱼中发现的,并且与哺乳动物igd有关。
16.免疫球蛋白亚类激活补体和介导抗体细胞细胞毒性的能力的不同意味着被激活的免疫球蛋白类型和亚类的变化可以对免疫应答,例如对感染具有积极的或消极的影响。例如,igg3与对抗一系列的细胞内细菌,寄生虫和病毒的增强的控制或保护有关。与其它igg亚型相比,igg3抗体是效应或功能的有效介质,包括增强的adcc,调理吞噬作用,补体激活和中和。因此,认为未来基于抗体的治疗和疫苗应该基于增强的功能能力的特征考虑利用igg3。研究糖基化模式和同种异型对igg3功能的影响可以扩大我们对igg3应答及其治疗潜力的理解。
17.igg3仅包括小部分的igg,并且直到最近几年仍相对研究较少。最近的研究强调了igg3效应子功能对一系列病原体的重要性,并且已经提供了克服igg3相关限制的方法,所述igg3相关限制例如同种异型依赖性短抗体半衰期和过度的促炎性激活。因此,理解igg3和其它免疫球蛋白亚类的分子和功能特性可以促进改进的基于抗体的免疫疗法以及对抗感染性疾病的免疫疗法和疫苗的发展。
18.对免疫球蛋白及其亚类的检测是本领域已知的。然而,散射测浑法和浊度测定法将测量总免疫球蛋白的量,而不是抗原特异性的量。用于抗原特异性的量的elisa方法利用免疫球蛋白亚类特异性抗体进行检测。这些抗体对每一类或亚类具有不同的亲合力。因此,例如igg亚类或iga亚类测定之间的比较可能不准确。此外,它们将具有不同的校准曲线,并且除了相对较少的例外之外,还缺乏国际标准来测量对不同致病抗原的抗体应答,这意味着测定结果之间的比较是不可能的。由于观察到的明显的量的差异,这防止了在单一测定中不同类别、亚类或轻链类型之间的任何比率或比较的可能性。
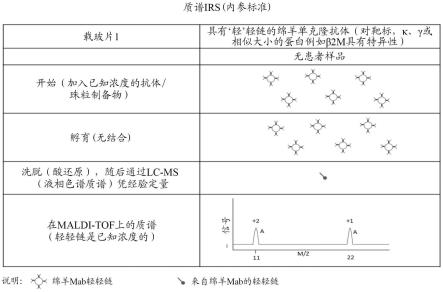
19.ladwig,p.m.等人,clinical chemistry(2014),第16卷,第1080-1088页讨论了使用质谱对igg亚类的绝对定量。他们描述了血清与稳定同位素标记的内肽标准以及完整纯化的马igg的组合。样品被变性,还原,烷基化并用肽酶如胰蛋白酶消化。他们然后通过lc-ms/ms分析消化的血清的igg亚类1-4和总igg。他们能够证明可以使用lc-ms/ms定量总igg和igg亚类1、2、3和4。重链和轻链同种型的同种型分析也描述在wo 2015/154052中,全文并入本文。这描述了通过质谱(ms)检测这种同种型。
20.发明人已经认识到,使单一方法同时和定量地剖析抗体应答,例如在感染之前,期间或之后,将允许更好的临床干预。他们认识到,通过固相捕获富集抗原或疾病特异性抗体,然后通过质谱分析,允许进行样品的自动分析,而没有例如基于elisa的测定的缺点。预期这将提高临床医生管理受试者对疾病的免疫应答的能力,并允许一般性研究对抗原的免疫应答,其可能提高疫苗和单克隆抗体的产生。
21.本发明提供了一种鉴定或表征受试者中的免疫应答的方法,其包括:
22.(a)使来自受试者的含有免疫球蛋白的样品与至少一种固定在支持物上的抗原接触;
23.(b)将未结合的非抗原特异性免疫球蛋白从所述支持物上洗去,以留下结合到所述支持物上的抗原的抗原特异性免疫球蛋白;
24.(c)任选地从所述支持物上的抗原洗脱所述抗原特异性免疫球蛋白;和
25.(d)对所述抗原特异性免疫球蛋白进行质谱分析以鉴定两种或更多种不同的抗原特异性免疫球蛋白类别、亚类和/或轻链同种型。所述免疫球蛋白本质上可以是单克隆的或多克隆的。
26.通常,提供两种或更多种不同的抗原。这些可以提供在不同的支持物上。因此,例如,可以将样品分成不同的等分试样,其中每个等分试样与不同支持物上的不同抗原接触。可选地,样品可以与支持物的不同部分上的抗原接触。
27.支持物可以是本领域通常已知的任何支持物。这些包括:琼脂糖,纤维素,玻璃,顺磁珠粒或磁性(例如铁)珠粒或聚苯乙烯珠粒。支持物也可以是例如微量滴定板上的孔,或者实际上用于例如maldi-tof ms的靶标。通常,使用顺磁珠粒或maldi-tof靶。顺磁珠粒是特别有用的,因为它们容易通过磁体与围绕珠粒的流体分离。通常使用本领域公知的化学方法将抗原共价连接到珠粒上。这些包括,例如,使用市售的链霉抗生物素蛋白包被的珠粒,并通过附着于抗原的生物素部分附着抗原。有许多可商购的用于珠粒的其它表面涂层,其促进抗原与这种珠粒的结合,包括各种氨基,羧基,烷基,硫醇,环氧基,肼和甲苯磺酰基。还使用例如cona来结合例如糖结合蛋白和rho1d4上的糖,它们在膜蛋白的纯化中是有效的。
28.未结合的非抗原特异性免疫球蛋白任选地用合适的缓冲液或其它洗涤液洗去。同样,这种洗涤在本领域中是通常是已知的。这种洗涤液包括例如含有例如0.1% tween的磷酸盐缓冲盐水。可以使用两次或更多次洗涤以确保去除非抗原特异性免疫球蛋白。
29.抗原特异性免疫球蛋白可以使用合适的洗脱缓冲液如酸缓冲液从抗原上洗脱下来。本发明还允许从抗原中洗脱不同特异性的抗体,例如通过在洗脱洗涤中使用不同浓度的盐或其它条件。因此,在去除非特异性结合标志物后,还可以用第一洗脱缓冲液去除具有较低抗原特异性的免疫球蛋白,然后用第二洗脱缓冲液去除具有较高抗原结合特异性的免疫球蛋白。例如,当目的抗原的不同部分对受试者的免疫系统具有不同水平的抗原性时,这可能是有用的。
30.可选地,如果例如抗原与质谱靶标如maldi-tof靶结合,则抗原结合的免疫球蛋白可以被电离并直接从靶标上分析。
31.可以对消化的抗原特异性免疫球蛋白进行质谱分析之前,对所述抗原特异性免疫球蛋白进行蛋白水解消化。通常,用于消化的酶是肽链内切酶,其在免疫球蛋白的链内而不是在免疫球蛋白的链的一端断裂免疫球蛋白。合适的肽链内切酶的实例包括胰蛋白酶。ludwig等人(同上)例如,在进行质谱分析之前,使用胰蛋白酶与水和碳酸氢铵的组合来进行免疫球蛋白的胰蛋白酶消化。
32.在gugten,clinical chemistry,第64卷,735-742(2018)中也已经证实了lc-ms/ms与胰蛋白酶消化的联合使用。这证明了基于免疫的诊断试剂盒偶尔在患者中产生可疑的
分析误差,例如总igg4增加。这导致总igg浓度的计算误差。使用lc-ms/ms,结合校准物,能够降低明显的差异。所述测定再次用于检测受试者中非抗原特异性免疫球蛋白和免疫球蛋白亚类的总量,例如对免疫缺陷和igg4相关疾病的评估。
33.remily-wood等人,proteomics clin appl.2014年10月;8(0):783
–
795也证实了lc-ms/ms与胰蛋白酶消化的联合使用。该作者描述了同种型和亚类特异性胰蛋白酶肽用于鉴定和监测血清中的单克隆免疫球蛋白的用途。本研究中使用的同种型和亚类特异性胰蛋白酶肽示于表2中。
34.该表显示了可以合成的同位素标记的肽的列表,标记的氨基酸加下划线,并且星号表示存在保守氨基酸置换的位置。该表显示了蛋白质、肽序列(以及它是同位素标记的还是通过保守的单个氨基酸置换产生的结构类似物)和转换。
35.可选地,抗原特异性免疫球蛋白的至少一部分在进行质谱分析之前不进行蛋白水解消化。也就是说,完整的重链和/或轻链可以通过质谱检测。
36.通常,抗原特异性免疫球蛋白的至少一部分可以用至少一种还原剂解离,以便在对分离的免疫球蛋白进行质谱分析之前将轻链从重链上分离下。这可以发生在例如蛋白水解消化(如果使用的话)之前。这在本领域中通常也是已知的,例如,如wo 2015/154052和ladwig等人所示,两者均通过引用全文并入本文中。
37.样品中的抗体与抗原结合,然后分成它们各自的类别、亚类或类型。这意味着可以将检测到的每一类别、亚类或类型的相对峰值彼此进行比较,以允许确定样品中的类别,亚类或轻链类型的相对量。这消除了对于elisa型测定经常看到的一些歧义。
38.可选地,可以使用内部消化对照。对照可以是,例如,来自另一种与受试者不同种类的动物的免疫球蛋白。例如,马igg由ladwig等人(同上)使用,因为马免疫球蛋白胰蛋白酶肽具有不同的氨基酸序列,因此具有与人免疫球蛋白胰蛋白酶肽等同物不同的质量。这允许对照区别于免疫球蛋白峰并用于确定胰蛋白酶消化的完成。预先确定量的合成的稳定同位素标记的内标肽可以例如在质谱分析之前加入。内标肽具有与同种型和亚类特异性胰蛋白酶肽完全相同的氨基酸序列,但是由于在用于合成肽的氨基酸中添加了稳定的同位素如c13和n15而更重。内标肽的化学性质与来自ig的天然胰蛋白酶肽相同,但其质量不同,因此可以在质谱仪中与天然肽一起监测。
39.通过报道在质谱仪中观察到的未知样品中内标肽的丰度与天然肽的丰度的比率来实现绝对定量。然后将该比率与在不同浓度的已知校准物下得到的标准曲线进行比较,所述校准物以与未知样品相同的方式消化并含有相同量的内标肽。然后可以将结合了特异性抗原的免疫球蛋白的浓度报道为绝对浓度,例如g/l。
40.因此,样品中一种或多种不同抗原特异性免疫球蛋白类别,亚类和/或轻链类型的量可以是绝对或相对定量的。
41.免疫球蛋白类别可选自igg、iga、igm、igd和ige。亚类可以选自igg1、igg2、igg3、igg4、iga1和iga2。轻链可以选自λ轻链和κ轻链。根据所观察到的免疫应答,可以检测2、3、4、5、6、7、8、9、10、11、12或13种不同的类别、亚类和类型。如上所述,igg3具有特别的兴趣,因此通常检测到的至少两个类别、亚类或类型包括igg3。可选地,ige和igg4最有可能对于研究过敏症有意义。
42.可以测定λ:κ轻链的相对比率。
43.所述方法还可以包括鉴定以下的一种或多种:
44.(a)与iga和/或igm结合的j链;和/或
45.(b)与igm结合的cd5l。
46.对j-链的检测描述于例如wo2019/055632中,其全文并入本文。
47.在wo 2019/055634(全文并入本文)中描述了对cd5l(也称为cd5样抗原和aim-巨噬细胞的细胞凋亡抑制剂)的检测。对样品中j-链的检测提供了在洗脱和还原之后和样品消化之前对与特异性抗原结合的免疫球蛋白相关的iga和/或igm分子的量的快速定性评估。当要测定这样的j链或cdl5抑制剂时,可以在将iga或igm富集样品与基底上的抗原接触之前,纯化血清样品或富集iga或igm,例如使用抗iga或抗igm抗体或片段。
48.可以在与结合到支持物上的样品接触之前纯化或富集样品中的免疫球蛋白。例如,可以使用例如蛋白g或melon凝胶富集样品中的igg。免疫球蛋白和样品也可以用抗免疫球蛋白抗体富集。例如,抗igm,抗igg,抗iga,抗igd或抗ige特异性抗体,或其片段。抗亚类特异性抗体,抗轻链型特异性抗体,或实际上抗重链类别轻链类型特异性抗体也可用于从样品中特异性纯化免疫球蛋白。可以使用抗体片段。
49.此外,还可能需要在与结合于受试者的抗原接触之前从样品中去除一种或多种类型的抗体或抗体类别。例如,igg的存在量明显高于其它类别抗体。当研究例如iga或igm时,可能存在igg的高背景。因此,例如通过使用蛋白g,melon凝胶或抗igg特异性抗体或其片段去除igg可能是理想的。抗免疫球蛋白类别,亚类,轻链类型,重链类别-轻链类型特异性抗体都是可商购的,例如来自the binding site limited,birmingham,united kingdom。
50.用于从样品中选择性地去除或纯化免疫球蛋白的抗体或其片段可以是单结构域抗体片段,如例如wo2015/154052中所述,整体并入本文中。可选地,也可使用交联抗体,其中重链通过例如硫醚键或另一种交联化合物如双(马来酰亚胺基)乙烷与轻链交联,如wo 2017/144909(整体并入本文中)中所公开的。sdaf和交联抗体的使用减少了用重链或轻链纯化样品的抗体造成的污染,这可能影响通过本发明的方法产生的数据。
51.与基底结合的抗原可以是较大分子的抗原性部分。因此,例如,它可以是较大蛋白的亚基或片段,这将允许研究对该蛋白的不同部分的免疫应答。
52.抗原可以是来自以下的抗原:细胞,病毒,细菌,古细菌,真菌,原生动物,蠕虫,自身免疫抗原,癌症抗原的抗原,能够在受试者中诱导过敏反应的抗原。生物体例如可以是致病生物体,例如可以研究针对其的疫苗或一种治疗,或者追踪疫苗或治疗对由这种生物体引起的疾病进展的影响。此外,通过观察与中和免疫球蛋白结合后释放到样品缓冲液中的抗原的杀伤相关的分子,可以使用质谱来监测与抗原结合的免疫球蛋白的中和作用。
53.所述方法还可以研究和鉴定受体位点的激活和非激活抗体。
54.本发明的方法可用于在感染之前,期间或之后剖析抗体应答。
55.致病病毒的实例包括:甲型肝炎病毒、柯萨奇病毒和其他小核糖核酸病毒科;乙型肝炎病毒和其它嗜肝dna病毒科(hepadnaviridae);丙型肝炎病毒、登革病毒和其它黄病毒科;单纯疱疹病毒1&2、巨细胞病毒、eb病毒和其它疱疹病毒科;hiv和其它逆转录病毒科;流感病毒和其它正粘病毒科;乳头瘤病毒和其它乳头瘤病毒科;狂犬病病毒和其它弹状病毒科;呼吸道合胞病毒和其它副粘病毒科;sars cov 2和mers及其它冠状病毒科。
56.致病细菌的实例包括:葡萄球菌(例如金黄色葡萄球菌)、链球菌、大肠杆菌、奈瑟
氏球菌、假单胞菌、结核分枝杆菌、耶尔森氏菌、芽孢杆菌、梭菌(艰难梭菌和肉毒梭菌)、嗜血杆菌(流感嗜血杆菌)、李斯特氏菌、疏螺旋体和立克次氏体。
57.致病真菌包括:厌酷球孢子菌(coccidiodes immitis)、荚膜组织胞浆菌(histoplasma capsulatum)、巴斯特酵母属(bastomyces)和肺囊虫属(pneumocystis)。
58.抗原可以来自原生动物,例如疟疾或锥虫。由原生动物引起的常见感染性疾病包括疟疾、贾第鞭毛虫和弓形体病。另外,痢疾可能是由许多阿米巴引起的。这些包括溶组织内阿米巴(entamoeba histolytica)、布氏锥虫(trypanosoma brucei gambiense)、杜氏利什曼原虫(leishmania donovani)、间日疟原虫(plasmodium vivax)、疟原虫(plasmodium malariae)、恶性疟原虫(plasmodium falciparu)和弓形虫(toxoplasma gondil)。这种原生动物疾病通常难以生产疫苗,因为生物体通常具有避开免疫系统的系统。能够研究在患有这种疾病的受试者中的免疫应答可能有助于表征所述疾病并有希望鉴定可能的疫苗组分。
59.有许多蠕虫疾病或生物体,例如绦虫病、吸虫病、蛔虫病、鞭虫病(trichuriasis)、钩虫病、蛲虫病、类圆线虫病、丝虫病、旋毛虫病、恶丝虫病和血管圆线虫病(大鼠肺线虫病)。
60.自身免疫性疾病通常是使人非常衰弱的。这些疾病包括,例如,类风湿性关节炎、系统性红斑狼疮(sle)、混合性结缔组织病(mctd)、炎性肠病、多发性硬化、1型糖尿病、吉兰-巴雷综合征、慢性炎性脱髓鞘性多发性神经病和银屑病。表征疾病和鉴定诱导疾病的抗原,或抗体或抗体亚类结合的抗原的能力将有助于鉴定治疗策略和实际上表征受试者中的疾病和疾病进展。抗原也可以是癌抗原。
61.许多癌症在肿瘤表面上表达癌症抗原,例如mhc i类或ii类分子,或者将它们分泌到体内。新抗原也被鉴定用于癌症。同样,本发明的方法将允许表征和检测这样的癌症,并有助于鉴定新的治疗。
62.受试者可能变得对抗原,例如对花粉,蜂蜇伤或例如镍过敏。当具有这种过敏症时,观察到不同的症状,它们是哮喘,皮肤瘙痒,或更极端的反应,例如过敏性休克,通常依赖于受试者系统内不同类型的抗体或这种不同类型的抗体的量。ige和igg4对这种病况最有可能让人感兴趣的。
63.抗原可以是病毒抗原,例如病毒包膜蛋白,衣壳蛋白,酶或血凝素。也可以使用完整的病毒裂解物或复合提取物。酶包括,例如,神经氨酸酶或甲基转移酶。后者常见于冠状病毒,例如sars-cov-2。基质蛋白通常被称为“m1”蛋白。在一些病毒中发现了离子通道蛋白,也称为“m2”蛋白。
64.类似地,细菌抗原可以是细胞抗原,鞭毛抗原,体细胞抗原,毒性抗原,菌毛抗原或类毒素的至少一个抗原部分。
65.受试者可以是任何产生抗体的生物体,例如鱼,哺乳动物,鸟或爬行动物。更通常地,受试者是哺乳动物,例如人,非人猿,猴,马,绵羊,山羊,牛,狗,猫或啮齿动物,例如小鼠,仓鼠或大鼠。哺乳动物也可以是骆驼科动物。后者是特别令人感兴趣的,因为它们通常产生缺乏轻链的抗体,并且正变得越来越多地用于产生例如sdaf(单结构域抗体片段)。
66.样品通常是任何生物流体,例如血液,血清,血浆,脑脊液,尿液,泪液,痰液,灌洗液或唾液。灌洗液可以是例如支气管肺泡灌洗液,其可以是例如经支气管镜检查获得的。还
可以使用鼻咽或口咽拭子样品,作为另外的鼻分泌物。
67.在进行质谱分析之前,可以向样品中加入电离对照。这种电离对照通常是与要通过质谱鉴定的化合物具有不同分子量的蛋白质。这确保质谱技术在样品之间一致地操作。
68.本发明的方法允许产生例如针对不同免疫应答的不同抗原的矩阵,如通过本发明的方法鉴定的不同抗原特异性免疫球蛋白类别,亚类或轻链类型所测量的。这允许容易地鉴定针对不同抗原的免疫应答的变化。因此,本发明的另一方面提供了一种产生矩阵或布置(profile)的方法。
69.本发明的方法可以与受试者中的免疫应答或免疫功能的一个或多个另外的指标组合。这些包括免疫调节和促炎细胞因子,例如干扰素,白细胞介素,白细胞介素-1,白细胞介素-2,tnf-α,血液或其它流体如灌洗液内的循环巨噬细胞或其它白细胞的数量,样品中补体蛋白的量和其它这样的因子。
70.用于分析的仪器可以例如由与质谱仪耦合的液相色谱仪(lc-ms,lc-ms/ms)组成。其它仪器配置包括但不限于与质谱仪耦合的cze或与质谱仪耦合的离子迁移装置。所用的典型电离技术包括但不限于电喷雾电离和maldi电离。用于分析的质谱仪可以包括但不限于四极杆飞行时间质谱仪,轨道阱质谱仪,三重四极质谱仪,离子阱质谱仪或飞行时间质谱仪。
71.提供了选择一种或多种疫苗靶标的方法(包括使用本发明的方法),以及鉴定受试者目标的免疫状态的方法(包括使用本发明的方法)。还进一步提供了表征受试者对病原体,过敏原或其它抗原的免疫应答的方法,包括使用本发明的方法。可以确定由病原体引起的病况的严重性或进展或治疗。本发明的另一方面提供了表征受试者的自身免疫应答的方法(包括使用本发明的方法),或表征受试者的过敏反应的方法。所述方法也可用于监测受试者中疾病的进展。
72.本发明的方法还可以提供使用或刺激对抗原的最大应答的最佳类型的抗体或抗体亚类的证据。例如,这可以允许优化单克隆抗体类别或其它单克隆抗体特性。
73.本发明的另一方面提供了用于鉴定或表征受试者中的免疫应答的计算机实施的方法,包括使用本发明的方法。所述方法可以包括将第一抗原特异性免疫球蛋白类别,亚类和/或轻链类型获得的质谱与第二抗原特异性免疫球蛋白类别,亚类和/或轻链类型获得的质谱进行比较,其中所述质谱通过本发明的方法获得。质谱可由计算机接收并进行比较,例如以提供与所述类别、亚类或轻链类型相关的一个或多个峰的量或比率。
74.计算机可以包括计算机处理器和计算机存储器。
75.还提供了用于通过根据本发明的方法鉴定或表征受试者中的免疫应答,并且包括使用根据本发明的计算机实施的方法的设备。所述设备可以包括质谱仪。
76.还提供了用于本发明方法的测定试剂盒,其包含附着于一种或多种基底的多种抗原以及一种或多种免疫球蛋白校准物。
77.现在将仅通过实例并参考以下附图来描述本发明:
78.图1显示了用抗igg,抗iga,抗igm,抗κ和抗λ抗体免疫纯化的免疫球蛋白的5个质谱。免疫纯化后,使用还原剂二硫苏糖醇解离重链和轻链,然后使用maldi-tof进行质谱分析。
79.图2来自ladwig,p.m.等人,clinical chemistry(2014),第16卷,第1080-1088页。
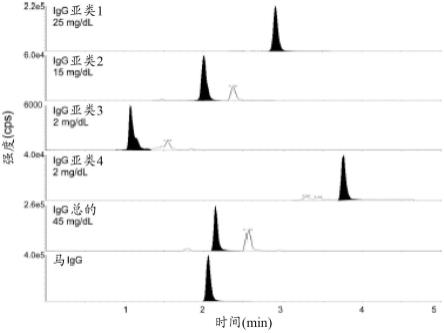
它显示了稀释在1:16牛血清中的igg亚低对照(the binding site limited,birmingham,united kingdom)的lc-ms/ms离子色谱图,显示了用于定量每种单独的亚类肽的峰(阴影),用于igg总量的共有肽,和用于定量的马igg肽。所见的许多非特异性背景峰经色谱分离且不干扰。
80.图3显示了使用固定在顺磁珠粒中的冠状病毒病毒突刺蛋白的maldi-tof光谱。光谱在扩展的7000-30000m/z范围内(显示所有3种电荷状态,a)和降低的11000至14000m/z范围(仅摆动 2-电荷状态,b)内呈现。单克隆抗体(对突刺蛋白特异)与珠粒结合,洗脱,并通过maldi-tof质谱分离峰。另外,健康人血浆和血清也分别与病毒突刺蛋白珠粒孵育。珠粒标记的病毒突刺蛋白(预先清除的)是对照以证明缀合后的珠粒洗涤不损害固定的蛋白。珠粒标记的α-人igg是其中病毒蛋白已经被α-igg特异性抗体替代的珠粒,以证明该抗体不仅结合单克隆抗体(其是igg单克隆抗体),而且当与正常血浆和正常血清一起使用时,还结合正常血浆和正常血清中的免疫球蛋白。
81.图4测试抗病毒sars-cov-2突刺蛋白和细菌肺炎球菌细胞壁多糖(cwps)的covid-19阴性(健康)和pcr-阳性(患病)个体的maldi质谱。用抗原缀合的珠粒免疫捕获4个个体的血清或血浆样品。个体2和4是“患病的”(对covid-19测试阳性)。个体1和3是“健康的”类别(对covid-19测试阴性)。
82.图5使用各种感染特异性抗原,测试抗目标感染的健康和患病个体的maldi质谱。sars-cov-2病毒特异性抗原用病毒突刺蛋白和核衣壳蛋白缀合到珠粒上。用抗原缀合的珠粒免疫捕获4个个体的样品。两个个体是“患病的”(对covid-19测试阳性,个体2和4)和两个是健康的(个体1和3)。
83.图6在erm-da470k参考血清的消化物和使用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的血清和血浆样品的洗脱液消化物中检测到的igg1肽tpevtc(cam)vvvdvshedpevk的提取离子色谱图(xic)。(a)erm-da470k中igg1肽的xic。(b-e)免疫沉淀的(b)covid-19阴性血清,(c)covid-19阴性血浆,(d)covid-19阳性血清,和(e)covid-19阳性血浆的洗脱液消化物中igg1肽的xic。*(cam)=氨基甲酰化半胱氨酸。
84.图7在erm-da470k参考血清的消化物和使用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的血清和血浆样品的洗脱液消化物中检测到的igg2肽glpapiek的提取离子色谱图(xic)。(a)erm-da470k中igg2肽的xic。(b-e)免疫沉淀的(b)covid-19阴性血清,(c)covid-19阴性血浆,(d)covid-19阳性血清,和(e)covid-19阳性血浆的洗脱液消化物中igg2肽的xic。
85.图8在erm-da470k参考血清的消化物和使用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的血清和血浆样品的洗脱液消化物中检测到的igg3肽tpevtc(cam)vvvdvshedpevqfk的提取离子色谱图(xic)。(a)erm-da470k中igg3肽的xic。(b-e)免疫沉淀的(b)covid-19阴性血清,(c)covid-19阴性血浆,(d)covid-19阳性血清,和(e)covid-19阳性血浆的洗脱液消化物中igg3肽的xic。*(cam)=氨基甲酰化半胱氨酸。
86.图9在erm-da470k参考血清的消化物和使用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的血清和血浆样品的洗脱液消化物中检测到的igg4肽glpssiek的提取离子色谱图(xic)。(a)erm-da470k中igg4肽的xic。(b-e)免疫沉淀的(b)covid-19阴性血清,(c)covid-19阴性血浆,(d)covid-19阳性血清,和(e)covid-19阳性血浆的洗脱液消化物中igg4肽的
xic。
87.图10在erm-da470k参考血清的消化物和使用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的血清和血浆样品的洗脱液消化物中检测到的iga1肽dasgvtftwtpssgk的提取离子色谱图(xic)。(a)erm-da470k中iga1肽的xic。(b-e)免疫沉淀的(b)covid-19阴性血清,(c)covid-19阴性血浆,(d)covid-19阳性血清,和(e)covid-19阳性血浆的洗脱液消化物中iga1肽的xic。
88.图11在erm-da470k参考血清的消化物和使用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的血清和血浆样品的洗脱液消化物中检测到的iga2肽dasgatftwtpssgk的提取离子色谱图(xic)。(a)erm-da470k中iga2肽的xic。(b-e)免疫沉淀的(b)covid-19阴性血清,(c)covid-19阴性血浆,(d)covid-19阳性血清,和(e)covid-19阳性血浆的洗脱液消化物中iga2肽的xic。
89.图12在erm-da470k参考血清的消化物和使用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的血清和血浆样品的洗脱液消化物中检测到的κlc肽sgtasvvc(cam)llnnfypr的提取离子色谱图(xic)。(a)erm-da470k中κlc肽的xic。(b-e)免疫沉淀的(b)covid-19阴性血清,(c)covid-19阴性血浆,(d)covid-19阳性血清,和(e)covid-19阳性血浆的洗脱液消化物中κlc肽的xic。*(cam)=氨基甲酰化半胱氨酸。
90.图13在erm-da470k参考血清的消化物和使用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的血清和血浆样品的洗脱液消化物中检测到的λlc肽yaassylsltpeqwk的提取离子色谱图(xic)。(a)erm-da470k中λlc肽的xic。(b-e)免疫沉淀的(b)covid-19阴性血清,(c)covid-19阴性血浆,(d)covid-19阳性血清,和(e)covid-19阳性血浆的洗脱液消化物中λlc肽的xic。
91.图14在erm-da470k参考血清的消化物和使用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的血清和血浆样品的洗脱液消化物中检测到的igg肽dtlmisr的提取离子色谱图(xic)。(a)erm-da470k中igg肽的xic。(b-e)免疫沉淀的(b)covid-19阴性血清,(c)covid-19阴性血浆,(d)covid-19阳性血清,和(e)covid-19阳性血浆的洗脱液消化物中igg肽的xic。
92.图15在erm-da470k参考血清的消化物和使用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的血清和血浆样品的洗脱液消化物中检测到的iga肽sgntfrpevhllpppseelalnelvtltc(cam)lar的提取离子色谱图(xic)。(a)erm-da470k中iga肽的xic。(b-e)免疫沉淀的(b)covid-19阴性血清,(c)covid-19阴性血浆,(d)covid-19阳性血清,和(e)covid-19阳性血浆的洗脱液消化物中iga肽的xic。*(cam)=氨基甲酰化半胱氨酸。
93.图16在erm-da470k参考血清的消化物和使用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的血清和血浆样品的洗脱液消化物中检测到的igm肽gvalhrpdvyllppar的提取离子色谱图(xic)。(a)erm-da470k中igm肽的xic。(b-e)免疫沉淀的(b)covid-19阴性血清,(c)covid-19阴性血浆,(d)covid-19阳性血清,和(e)covid-19阳性血浆的洗脱液消化物中igm肽的xic。
94.图17在用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的covid-19阳性血清的消化洗脱液中对应于igg1肽tpevtc(cam)vvvdvshedpevk的片段化光谱。(a)突出c-末端片段
离子(y-离子系列)的片段化光谱。(b)预期的和观察到的y离子系列片段的表。*(cam)=氨基甲酰化半胱氨酸。
95.图18在用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的covid-19阳性血清的消化洗脱液中对应于igg2肽glpapiek的片段化光谱。(a)突出c-末端片段离子(y-离子系列)的片段化光谱。(b)预期的和观察到的y离子系列片段的表。
96.图19在用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的covid-19阳性血清的消化洗脱液中对应于igg3肽tpevtc(cam)vvvdvshedpevqfk的片段化光谱。(a)突出c-末端片段离子(y-离子系列)的片段化光谱。(b)预期的和观察到的y离子系列片段的表。*(cam)=氨基甲酰化半胱氨酸。
97.图20在消化的erm-da470k参考血清中对应于igg4肽glpssiek的片段化光谱。(a)突出c-末端碎片离子(y-离子系列)的片段化光谱。(b)预期的和观察到的y离子系列片段的表。
98.图21在用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的covid-19阳性血清的消化洗脱液中对应于iga1肽dasgvtftwtpssgk的片段化光谱。(a)突出c-末端片段离子(y-离子系列)的片段化光谱。(b)预期的和观察到的y离子系列片段的表。
99.图22在用与sars-cov-2核衣壳蛋白缀合的珠粒免疫沉淀的covid-19阴性血清的消化洗脱液中对应于iga2肽dasgatftwtpssgk的片段化光谱。(a)突出c-末端片段离子(y-离子系列)的片段化光谱。(b)预期的和观察到的y离子系列片段的表。
100.图23在用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的covid-19阳性血清的消化洗脱液中对应于κlc肽sgtasvvc(cam)llnnfypr的片段化光谱。(a)突出c-末端片段离子(y-离子系列)的片段化光谱。(b)预期的和观察到的y离子系列片段的表。*(cam)=氨基甲酰化半胱氨酸。
101.图24在用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的covid-19阳性血清的消化洗脱液中对应于λlc肽yaassylsltpeqwk的片段化光谱。(a)突出c-末端片段离子(y-离子系列)的片段化光谱。(b)预期的和观察到的y离子系列片段的表。
102.图25在用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的covid-19阳性血清的消化洗脱液中对应于igg肽dtlmisr的片段化光谱。(a)突出c-末端片段离子(y-离子系列)的片段化光谱。(b)预期的和观察到的y离子系列片段的表。
103.图26在用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的covid-19阳性血清的消化洗脱液中对应于iga肽sgntfrpevhllpppseelalnelvtltc(cam)lar的片段化光谱。(a)突出c-末端片段离子(y-离子系列)的片段化光谱。(b)预期的和观察到的y离子系列片段的表。*(cam)=氨基甲酰化半胱氨酸。
104.图27在用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的covid-19阳性血清的消化洗脱液中对应于igm肽gvalhrpdvyllppar的片段化光谱。(a)突出c-末端片段离子(y-离子系列)的片段化光谱。(b)预期的和观察到的y离子系列片段的表。
105.图28比较了在用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的阴性和阳性covid-19血清样品的消化洗脱液中,代表免疫球蛋白同种型igg/iga/igm,igg亚类,iga亚类和轻链的标志物的峰面积的柱形图。(a)igg亚类,(b)iga亚类,(c)轻链和(d)igg/iga/igm免疫球蛋白的峰面积。
106.图29比较了在用与sars-cov-2突刺蛋白缀合的珠粒免疫沉淀的阴性和阳性covid-19血浆样品的消化洗脱液中,代表免疫球蛋白同种型igg/iga/igm,igg亚类,iga亚类和轻链的标志物的峰面积的柱形图。(a)igg亚类,(b)iga亚类,(c)轻链和(d)igg/iga/igm免疫球蛋白的峰面积。
107.igg类别和轻链类型可以使用本领域通常已知的技术进行鉴定。
108.使用maldi-tof光谱获得的不同ig类别和轻链类型的光谱显示在例如图1中。这利用了已经用二硫苏糖醇解离的免疫纯化的免疫球蛋白,如本领域通常已知的。这表明可以鉴定不同的重链类别和轻链类型。在例如wo2015/154052中显示了进行这种测定的方法。
109.图2是来自ladwig等人(同上)的一个实例,其证明也可以使用质谱确定不同免疫球蛋白的亚类并对其进行定量。用于进行该测定的方法描述于例如该篇论文中。
110.本发明中的这些技术被应用于抗原特异性免疫球蛋白。将含有免疫球蛋白的样品与附着于基底(如顺磁珠粒)上的抗原接触。然后在质谱分析之前洗涤抗原特异性抗体以去除非特异性结合,并将其从珠粒中洗脱。
111.图3(a和b)显示了附着于顺磁珠粒的冠状病毒病毒突刺蛋白的实例。上图显示了用于受体位点单克隆抗体的激活和非激活抗体特异性结合病毒突刺蛋白珠粒,然后洗脱并通过质谱检测为免疫球蛋白轻链和重链的不同的峰。正常血清和正常血浆免疫球蛋白不与病毒突刺蛋白结合,因此从珠粒上洗掉,因此在洗脱液中不能检测到。中间图显示了病毒突刺蛋白珠粒的预洗涤基本上不影响固定的蛋白,并且仍然允许单克隆抗体与其结合。下图显示了如果病毒突刺蛋白取代α-igg特异性抗体,则该单克隆抗体结合珠粒。当这种珠粒与正常血浆或正常血清孵育时,检测血浆或血清中的igg抗体,导致仅在下图中观察到的重叠的宽的较低的峰。
112.表3显示了典型量的不同igg亚类和igg的矩阵的实例,在该实例中,所述不同igg亚类和igg可以针对不同的病毒蛋白生成。这种矩阵可以被扩展以包括其它类别,亚类型或轻链类型。例如,ige可集中于例如ige相关的慢性疾病,如可能正在研究中的过敏性哮喘或慢性荨麻疹。
113.这种矩阵也可以被转化成图形格式或其它格式,以允许比较不同免疫球蛋白类别,亚类或轻链类型与不同抗原之间的数据。它可以使用适当的计算实施的方法自动地输入数据。
114.为了说明我们的方法,我们分析了四个样品中对3种不同的感染相关抗原,sars cov-2突刺和核衣壳蛋白以及肺炎球菌细胞壁多糖(cwps)的免疫应答;2个“患病的”个体(covid-19 pcr阳性)和2个“健康的”个体(covid-19的测试阴性)。这4个个体的血清(样品1和2)或血浆样品(3和4)用这3种抗原缀合的珠粒免疫捕获。洗脱、还原由珠粒捕获的抗体和抗原特异性蛋白并通过maldi-tof ms分析。此外,在胰蛋白酶消化后,洗脱由珠粒捕获的抗体和抗原特异性蛋白,并通过lc-ms进行分析。igg1、igg2、igg3、igg4、iga1、iga2、igg、iga、igm重链以及κ和λ轻链的胰蛋白酶肽被用于“剖析”免疫应答。对于lc-ms分析,国际蛋白质参考材料da470k也用作血清对照。
115.方法
116.免疫沉淀和maldi-tof-ms
117.抗体和抗原特异性蛋白被抗原缀合的顺磁珠粒捕获,并在还原条件下洗脱以将轻
链与重链解离。简言之,用磷酸盐缓冲盐水,0.1% tween-20(pbst)洗涤50-150μl珠粒三次。将稀释的样品加入到珠粒中,并在室温下振荡孵育30分钟。用pbst洗涤珠粒3次,然后用标准去离子水再洗涤3次。使用50μl的0.1%甲酸(lc-ms)或5%乙酸(包括还原剂)(maldi-tof)通过在室温下振荡孵育15分钟来洗脱珠粒。随后用maldi矩阵(α-氰基-4-羟基肉桂酸)将洗脱液夹点在maldi-tof目标板上并干燥。在覆盖5000至80,000的m/z范围的bruker microflex maldi-tof-ms上以正离子模式获得质谱,其包括分析物(人κ或λ轻链)的双电荷([m 2h]2 ,m/z 10900-12300)离子。在2 电荷状态下观察到的轻链可被分割成对每条轻链具有特异性的3个区域;λ(11200-11560m/z),κ(11570-11850m/z)和重κ(11900-12400m/z)。
[0118]
lc-ms/ms和消化条件
[0119]
将洗脱液转移到新鲜的微量离心管中并通过加入1m的三乙基碳酸氢铵(teab)来中和。然后将样品用200mm三(2-羧乙基)膦(tcep)(中性ph)在60℃下以1000rpm还原30分钟,然后冷却至室温。通过添加375mm碘乙酰胺进行烷基化并在室温下在黑暗中孵育30分钟。用2.5μl的1μg/μl胰蛋白酶进行酶促消化,并将样品在37℃以1000rpm孵育2小时。用1μl的100%甲酸终止消化反应。在通过液相色谱耦合的电喷雾离子化质谱(lc-esi-ms)进行分析之前,使用真空浓缩器在60℃下在水性模式下减小样品体积。
[0120]
在与acquityi-class uplc系统耦合的xevo g2-xs qtof质谱仪(waters ltd.,wilmslow,uk)上分析样品。将10μl经消化的样品注射到维持在40℃下的acquity uplc peptide beh c18,1.7μm,2.1
×
150mm柱(waters ltd.,wilmslow,uk)上,流速为0.2ml/min。使用从0.1%(v/v)甲酸水溶液(a)至0.1%甲酸(v/v)/乙腈(b)的梯度。(梯度:0-1min,1% b;1-60min,40% b;60-70min,60% b;70min,95% b;70-80min,95% b;80min 1% b;80-90min 1% b)。用40v锥电压将毛细管电压设定为1.5kv。将源温度设定为120℃并且将去溶剂化温度设定为250℃。将锥形气流保持在50l/h并且去溶剂化气体流保持在600l/h。在100-2000m/z之间扫描90分钟后获得mse。扫描在6ev的低碰撞能量持续0.5秒和25ev至45ev的高碰撞能量斜坡持续另外0.5秒之间交替。启用lockspray
tm
并且在3kv毛细管电压和30v锥电压下每分钟测量亮氨酸脑啡肽0.25秒。在表5中概述了监测用于mse相对定量的离子。每个单独的免疫标记肽的片段化谱示于图17至27中。
[0121]
结果
[0122]
maldi-tof
[0123]
以峰强度(a.u)测量针对抗原缀合的珠粒的总抗体应答,并通过来自maldi-ms数据的峰分布来表征。大多数测试抗细菌多糖的个体呈现低水平的天然抗体应答(图4)。这排除了个体3(covid-19阳性),其对细菌抗原具有显著高的应答,提示最近的细菌感染。总的来说,针对细菌抗原观察到的轻链分布主要是多克隆的和寡克隆的,对κ轻链的使用,特别是重κ有偏向。针对病毒sars-cov-2突刺蛋白测试相同的个体(图4)。covid-19阳性个体(2和4)表现出针对突刺蛋白的高抗体应答,由下面的多克隆轻链分布(平滑峰)和寡克隆轻链(尖峰)组成。“健康的”covid-19阴性个体(2和4)存在基线抗体应答。随后,我们比较对sars-cov-2核衣壳蛋白的免疫应答与对突刺蛋白的免疫应答,以表征对大小,结构,功能和定位不同的抗原的应答(图5)。如用突刺蛋白所观察到的,来自covid-19阳性个体(2和4)的抗体应答也引起针对核衣壳蛋白的高应答。核衣壳应答主要由具有很少寡克隆峰的λ轻链组成(图5)。“健康”covid-19阴性个体(1和3)对核衣壳蛋白的相对应答接近基线(图5)。所
有4个样品对细菌肺炎球菌细胞壁多糖的κ∶λ比具有高κ∶λ比,表明κ轻链偏向(表4)。针对珠粒上的病毒蛋白抗原(sar-cov-2突刺蛋白和核衣壳蛋白)的应答的κ∶λ比是可变的,没有偏向的证据。两个covid-19阳性个体(2和4)显示出针对核衣壳蛋白的应答,其中κ∶λ比《1,表明该应答由λ轻链使用控制(表4)。
[0124]
总之,maldi-tof分析提供了对细菌和病毒抗原的抗体应答的概述,并指出了这4个个体之间免疫应答的数量和品质的差异。
[0125]
lc-ms/ms
[0126]
为每种人免疫球蛋白(igg1、igg2、igg3、igg4、iga1、iga2、igg、iga、igm重链以及κ和λ轻链)选择对所检测的免疫球蛋白具有特异性(诊断)的免疫标记胰蛋白酶-肽(表5)。这些肽的氨基酸序列和同一性使用如图17-27所示的相应片段化离子光谱来确认。在每种情况下,ms光谱(a)伴随有片段化离子表(b)。这些肽是根据它们的长度,相对丰度选择的,并且它们只在蛋白质的多肽序列中出现一次。
[0127]
这些肽用于剖析四个人样品中对3种不同的感染相关抗原,sars cov-2突刺和核衣壳蛋白以及肺炎球菌cwps的免疫应答;2个“疾病状态”个体(covid-19 pcr阳性)和2个“健康的”个体(covid-19的测试阴性)。在每种情况下,获得提取的离子色谱图以鉴定该肽的存在,并且计算峰下面积并用作替代标志物,用于对其来源的完整免疫球蛋白进行比较测量。例如,针对sars-cov-2突刺蛋白获得的这4个人临床样品的提取的离子色谱图显示了igg1(图6),igg2(图7),igg3(图8),igg4(图9),iga1(图10),iga2(图11)以及κ(图12)和λ轻链(图13)。此外,还获得了总igg(图14),总iga(图15)和总igm(图16)的肽标志物。为了比较,独立地消化国际蛋白质参考材料erm-da470k并作为血清免疫球蛋白对照运行。可以比较图6-16中所示的相对丰度,并将其用于剖析健康和疾病状态样品之间的抗体应答。这图示了血清(图28)和血浆(图29)矩阵。为了容易比较,已经将这些分为igg(图28a和29a)和iga亚类(图28b和29b),免疫球蛋白轻链(图28c和29c)和总免疫球蛋白igg、iga和igm(图28d和29d)。以igg应答来看,covid阳性样品的免疫应答远大于covid阴性样品的免疫应答。就亚类应答而言,igg1和iga1都是存在的主要亚类。轻链的使用在κ和λ之间大致相同。这类似于使用maldi-tof平台观察到的(表4)。
[0128]
还产生了由核衣壳蛋白和肺炎球菌cwps珠粒免疫捕获的免疫球蛋白的等同分析。该数据与表6中的突刺蛋白的数据相结合,并表示为相对丰度谱。如预期的,在covid阴性(健康)患者中对核衣壳蛋白抗原的抗体免疫应答比阳性(疾病状态)患者低。与两种sars-cov-2抗原相比,对肺炎球菌cwps的免疫应答在covid阳性和阴性患者之间更均衡,但强烈偏向于igg2和κ轻链应答。这也支持使用maldi-tof-ms观察到的结果。
[0129]
总之,胰蛋白酶-肽lc-ms/ms分析提供了对细菌和病毒抗原的抗体应答的详细概述,并允许对这4个个体之间的抗体免疫应答进行剖析。
[0130][0131]
表1
[0132][0133]
表2
[0134][0135]
表3
[0136]
[0137]
[0138]
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。