调控水稻耐盐性的类受体蛋白激酶strk2蛋白、基因及应用
技术领域
1.本发明属于植物基因工程领域。具体涉及调控水稻耐盐性的类受体蛋白激酶strk2蛋白、基因及其在耐盐水稻培育中的应用。
背景技术:
2.水稻(oryza sativa l.)是超过世界一半人口的主粮,中国也是世界水稻产量第一的国家。近年来土壤的盐渍化是威胁和制约水稻产量的重大因素。我国现有近1 亿 hm2内陆盐碱面积和大约240万hm2沿海滩涂,若能将这些盐渍化的土地加以利用,它们将是潜力巨大的可利用土地后备资源。开发和创制耐盐水稻新品种是提高水稻产量和有效利用盐碱地等农业问题最有效的解决办法。土壤盐渍化最主要的判断依据为是否含有高浓度的盐,其中又以分布最广泛、最易溶解的nacl最为常见。盐碱地土壤中积累的盐主要来自于海水和灌溉水中nacl的积累。水稻属于中度盐敏感作物,在生产中定义能够在含0.3%以上盐度的土地中生长的品种属于能够耐受盐胁迫和碱胁迫水稻品种。因此,在利用改造盐碱地作为种植耕地的应用中,水稻是沿海滩涂和盐碱地改良的首选粮食作物。研究水稻的耐盐胁迫机制,筛选、培育和推广耐盐碱新品种,是保障耕地面积、维护粮食安全的重要举措。通过遗传改良的方法增强水稻耐受盐胁迫性能,就需要能发掘更多的潜力耐盐基因,并从基因功能和生理生化机制的层面,解析具体的作用机制,丰富水稻耐盐遗传改良的候选基因。
3.高盐浓度的生长环境会抑制植物的正常生长和发育,严重情况甚至引起植物的死亡,导致离子毒害(主要是na
)和高渗胁迫,同时伴随发生一些次级胁迫如氧化损伤等。同时,盐胁迫对植物的正常生理活动以及代谢的影响是复杂且多维度的,从生理层面理解的影响主要包括了体内离子平衡、渗透平衡、酶活性、氧化还原平衡以及相应的信号转导和基因表达的改变。此外,高浓度的na
对于细胞膜完整性和蛋白质功能也有一定程度的影响,植物也会通过调节自身机制抵消负面的影响,提高细胞的适应性,以增强对盐胁迫的耐受性。
4.高盐胁迫会引起严重的氧化胁迫。盐胁迫条件下,光合速率的降低能够激活梅勒反应,产生大量的活性氧(reactive oxygen species,ros)。其中,过氧化氢(h2o2)是一种非常重要的ros分子,其本身能够作为信号分子参与植物的生长发育调控,以及响应生物与非生物胁迫(周根友, 翟彩娇, 邓先亮, 张蛟, 张振良, 戴其根, 崔士友. 盐逆境对水稻产量、光合特性及品质的影响. 中国水稻科学, 2018, 32(2), 146
–
154)。然而ros的过量积累会氧化并破坏细胞组分,若不及时清除将会对植物产生严重的危害。因此,体内过量ros的及时清除能显著提高植物盐碱等胁迫的耐受性。为了消除ros 的毒害,植物已进化出有效清除ros的酶和非酶清除机制(xu j., duan x., yang j., beeching j.r., zhang p. enhanced reactive oxygen species scavenging by overproduction of superoxide dismutase and catalase delays postharvest physiological deterioration of cassava storage roots. plant physiology. 2013, 161, 1517
–
1528)。如过氧化氢酶(cat)和抗坏血酸(asa)就分别是细胞内重要的ros清除酶类和非酶清除抗氧化分子。这些
酶类和非酶类协同作用使ros保持稳态,维持ros在细胞信号传导中所需要的正常水平(刘聪, 董腊嫒, 林建中, 刘选明. 逆境胁迫下植物体内活性氧代谢及调控机理研究进展. 生命科学研究, 2019, 23, 253
–
258)。
5.植物对外界刺激的内部变化依赖于复杂的信号系统,细胞表面的类受体蛋白激酶(rlks)是通过感知外界信号变化和信号的传递来启动细胞信号通路机制。大量的研究结果表明,rlks和类受体胞质激酶(rlcks)在同一个信号通路中存在协同作用,且该协同模式在植物中普遍存在。蛋白的结构决定功能,由于rlck作为类受体蛋白激酶家族中缺失了胞外配体结合结构域的一类蛋白,无法单独完成对外界信号的接收及参与细胞跨膜信号转导的生物功能,因此需要与一个具有胞外结构域的rlk相互协同作用,通过蛋白磷酸化的级联反应传递信号,激活下游效应分子(li j, wen j, lease ka, doke jt, tax fe, walker jc. bak1, an arabidopsis lrr receptor-likeprotein kinase, interacts with bri1 and modulates brassinosteroid signaling. cell, 2002, 110, 213
–
22)。rlk和rlck蛋白相互作用,共同参与的信号转导通路广泛参与了植物的生长发育、免疫、繁殖、激素信号、胁迫响应等生命过程的调控
23.。例如,类受体胞质酪氨酸激酶strk1在盐胁迫条件下通过磷酸化过氧化氢酶catc,以提高catc活性从而提高水稻耐盐性性(zhou y, liu c, tang d, et al. the receptor-like cytoplasmic kinase strk1 phosphorylates and activates catc, thereby regulating h2o
2 homeostasis and improving salt tolerance in rice. the plant cell, 2018,30(5), 1100-1118)。因此,发掘参与盐胁迫响应的rlk和rlck,解析其参与的盐胁迫信号转导分子机制,有助于丰富水稻耐盐基因资源和解析盐胁迫响应的调控网络,同时为水稻耐盐品种的培育提供优质基因资源和理论指导。
技术实现要素:
6.本发明提供一种可调控水稻耐盐性的类受体蛋白激酶编码基因(salt tolerance receptor-like kinase 2, strk2),该基因编码蛋白通过磷酸化方式调控水稻的盐胁迫响应。在nacl处理条件下,strk2过表达株系在种子萌发期的苗长、根长均低于野生型,幼苗期的存活率也低于野生型,说明strk2过表达株系表现为对盐胁迫敏感。利用crispr/cas9基因编辑系统构建strk2敲除株系,经农杆菌介导转化,测序鉴定得到纯合的敲除株系strk2。在nacl处理条件下,敲除株系strk2在种子萌发期的苗长、根长均显著高于野生型,幼苗期的存活率也显著高于野生型,说明敲除strk2能显著提高水稻的耐盐性,在耐盐水稻品种培育中具有重要的应用价值。随后,通过质谱差异表达蛋白鉴定,筛选到一个strk2的相互作用蛋白lrrk1,发现strk2可磷酸化lrrk1,并通过在盐胁迫条件下对lrrk1的磷酸化调控,实现对盐胁迫的响应。本发明提供了一个负调控水稻耐盐性的新基因strk2,为培育耐盐水稻品种提供了优质候选基因,在农业领域具有广阔的应用前景。
7.本发明所提供的水稻盐胁迫响应调控基因strk2来源于水稻(oryza sativa l.),是具有下述氨基酸残基序列之一的蛋白质:1)序列表中的seq id no:3;2)将序列表中seq id no:3的氨基酸残基序列经过一至十个氨基酸残基的取代、缺失或添加且具有调控水稻负调控响应盐胁迫功能的蛋白质。序列表中的序列3由606个氨
基酸残基组成。
8.同时,本发明还提供strk2的编码基因,strk2的cdna基因,可具有下述核苷酸序列之一:1)序列表中seq id no:2的dna序列;2)编码序列表中seq id no:3蛋白质序列的多核苷酸;3)在高严谨条件下可与序列表中seq id no:2限定的dna序列杂交的核苷酸序列;4)与序列表中seq id no:2限定的dna序列具有70%以上同源性,且编码相同功能蛋白质的dna序列。
9.序列表中的序列2由121个碱基组成,其开放阅读框(orf)为自5’端第1位至1821位碱基。
10.strk2的基因组基因,可具有下述核苷酸序列之一:1)序列表中seq id no:1的dna序列;2)编码序列表中seq id no:3蛋白质序列的多核苷酸;3)在高严谨条件下可与序列表中seq id no:1限定的dna序列杂交的核苷酸序列;4)与序列表中seq id no:1限定的dna序列具有70%以上同源性,且编码相同功能蛋白质的dna序列。
11.序列表中的序列1为strk2的基因组序列,包含了3729个碱基,该基因含有2个外显子(序列1的 5’端起:872-1695,2222-3219)和3个内含子(序列 1的 5’端起:1-871,1695-2221,3220-3729)。
12.所述高严谨条件可为在 0.1
×
sspe (或0.1
×
ss),0.1
×
sds的溶液中,在65 ℃下杂交并洗膜。
13.含有strk2的表达载体、转基因细胞系和宿主菌均属于本发明的保护范围。具体地,strk2的基因表达为组成型表达。
14.本发明还提供一种提高水稻耐盐性的方法:利用基因编辑例如crispr/cas9基因编辑系统敲除strk2,获得显著提高耐盐性的水稻敲除株系strk1。具体地,将所述的crispr/cas9-strk2载体通过农杆菌介导的方式导入水稻组织或细胞,获得耐盐性水稻。
15.所述水稻盐胁迫负调控控制基因strk2可通过含有strk2的植物表达载体导入外植体;用于构建所述植物表达载体的出发载体可为任意一种双元农杆菌载体或可用于植物基因枪轰击的载体等,如pcambia1301-ubin (genbank号:af234296)、pcambia2301、pcambia1300或其他衍生植物表达载体。
16.使用strk2构建植物表达载体时,在其转录起始核苷酸前可加上任何一种增强型、组成型、组织特异型或诱导型启动子,如花椰菜花叶病毒(camv)35s启动子、泛素化基因ubiquitin启动子(pubi)等,它们可单独使用或与其它的植物启动子结合使用。此外,使用本发明的基因构建植物表达载体时,还可使用增强子,包括翻译增强子或转录增强子。这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必须与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。
17.为了便于对转基因植物或转基因植物细胞进行鉴定及筛选,可对所使用的载体进行加工,如加入选择性标记(gus基因、gfp、yfp、as-red和荧光素酶基因等)或具有抗性的抗
生素标记基因(潮霉素、卡那霉素、氨苄青霉素、博莱霉素等)。为了转基因植物释放的安全性,在构建植物表达载体时也可不携带任何标记基因,在苗期进行特定pcr分子标记筛选。
18.含有本发明的strk2的植物表达载体可通过使用ti质粒、ri质粒、植物病毒载体、直接dna转化、显微注射、电导、农杆菌介导或基因枪等常规生物学方法转化水稻细胞或组织,并将转化的水稻组织培育成植株。被转化的植物宿主既可以是单子叶植物,也可以是双子叶植物,如:水稻、小麦、玉米、黄瓜、番茄、杨树、草坪草、苜蓿等。
19.本发明具有以下有益效果:在水稻中过量表达strk2基因,幼苗期、苗期均出现盐敏感表型,存活率均低于野生型。相反,当敲除strk1后,其敲除突变株系strk1的耐盐性显著提高,存活率和生长量均显著高于野生型。进一步研究发现,在盐胁迫条件下,strk2感知信号后,通过磷酸化其互作蛋白lrrk1,参与盐胁迫信号的转导,最终负调控水稻的耐盐性。总之,strk2负调控水稻的耐盐性,其敲除突变体在耐盐水稻培育中具有很好的应用前景。
20.本发明strk2的发掘及其作用机理的解析为水稻耐盐品种选育提供分子标记和理论指导,尤其strk2敲除株系的创制是提高盐碱地水稻产量的有效途径。
附图说明
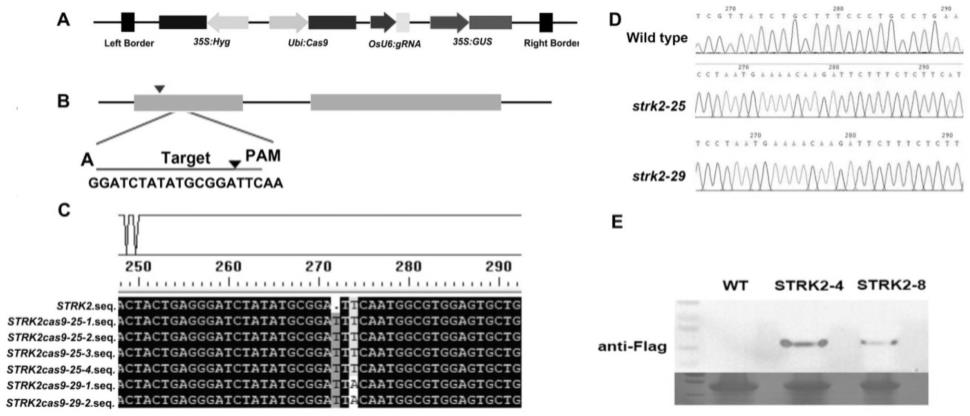
21.图1 strk2敲除株系及其过表达株系的构建。其中,a. pylcrispr/cas9双元载体的结构。b. strk2基因的crispr/cas9敲除靶点的引物设计。c. strk2基因的crispr/cas9敲除株系的编辑区域测序。d. strk2基因敲除株系与野生型序列比对。e. strk2过表达株系的western blot检测。用抗flag抗体免疫检测融合蛋白flag-strk2。
22.图2 strk2蛋白结构及亚细胞定位分析。其中,a. strk2的结构域预测分析。b. strk2的三维结构预测。c. strk2的组织特异性表达分析。d. strk2定位于细胞膜。
23.图3 strk2过表达及敲除株系的盐胁迫表型鉴定。其中,a. 盐胁迫条件下strk2过表达及敲除株系苗期的表型鉴定。b. 盐胁迫处理后恢复5d后存活率的统计。以15 d幼苗经140 mm nacl处理15 d,再正常水培恢复5 d。c. 盐胁迫条件下strk2过表达及敲除株系早期幼苗的表型鉴定。d. 盐胁迫处理后早期幼苗的苗长统计。以刚萌发露白的种子分别在正常条件及250 mm nacl条件下,测定盐胁迫对其苗长的影响。数值代表的是平均值
±
标准方差(t 检验,* p ≤ 0.05; ** p ≤ 0.01)。
24.图4 lrrk1与strk2相互作用验证。其中,a. 酵母双杂分析lrrk1与strk2的相互作用。b. 采用酵母双杂分段分析lrrk1与strk2的相互作用。 lrrk1分为5段,lrrk1a为n端,lrrk1e为c端, lrrk1b-d为激酶结构域,分为3段。c. lrrk1与strk2双分子荧光互补互作分析。
25.图5 strk2磷酸化lrrk1。其中,a. lrrk1激酶失活蛋白体外磷酸化检测。b. strk2激酶失活蛋白体外磷酸化检测。c.磷酸化抗体检测lrrk1与strk2体外磷酸化。
26.图6 nacl处理后strk2磷酸化lrrk1水平升高。其中,nacl处理增强了体内lrrk1的磷酸化水平。flag-lrrk1为经100 mm nacl处理不同时间的lrrk1过表达株系中ip得到的融合蛋白。
27.图7 盐胁迫对lrrk1过表达和敲除株系中ros和asa含量的影响。其中,a. 活性氧荧光探针检测根中ros的积累。b-d. 盐胁迫与正常条件下野生型、lrrk1过表达及敲除株系
中h2o
2 (b)、超氧阴离子(c)和asa (d)的含量测定。
具体实施方式
28.下面结合具体实施例对本发明做进一步说明,下述实施例中提到的实验方法如无特别说明均为常规方法。
29.1. strk2结构分析及其过表达和敲除株系的构建(1)strk2基因的结构分析从水稻数据库(http://rice.plantbiology.msu.edu)中下载strk2(loc_os11g14420)的编码序列,通过比较粳稻品种日本晴(oryza sativa l. japonica cv. nipponbare)的strk2的cdna及其基因组dna序列,发现strk2共有2个外显子(seq id no:1的 5’端起:872-1694,2222-3219),3个内含子(seq id no:1的 5’端起:1-871,1695-2221,3220-3729);其基因组全长为3729 bp(seq id no:1),cdna全长为1821 bp(seq id no:2),开放阅读框架为seq id no:2自5’端第1至1821位点,共有1821个碱基;其编码蛋白长度为607个氨基酸。用引物设计软件primer premier 5设计扩增strk2全长的特异性正向引物strk2-f: 5'-atgtctgtcaagtgttcc-3' 和反向引物strk2-r: 5'-ctagtttgctacaataa-3',以日本晴的cdna为模板,pcr方法克隆得到全长strk2基因。
30.(2)strk2过表达株系的构建通过同源重组的方法将strk2基因克隆到gateway入门载体pgwc中并测序。测序正确后将其通过lr反应重组到改造后的pcambia1301gw载体中。改造后的pcambia1301gw载体含有潮霉素抗性筛选基因hpt和红色荧光蛋白筛选标记基因asred。通过农杆菌介导转化,同时由玉米的泛素基因ubiquitin启动子(pubi)驱动目标基因strk2过表达或组成型表达,且strk2的n端融合有标签蛋白flag。然后通过农杆菌介导的方式侵染粳稻品种kitaake的愈伤组织,经筛选、分化、生根、炼苗后得到再生植株,并经鉴定后得到阳性过表达株系strk2-4和strk2-8(图1中e)。
31.(3)通过crispr/cas9技术构建strk2敲除株系构建strk2敲除株系crispr/cas9载体分为两步,即sgrna表达盒的构建及通过边切边连,将sgrna表达盒克隆到pylcrispr/cas9载体上。经靶点双链接头的制备,靶点接头连接扩增,两轮pcr反应后构建sgrna表达盒,再通过边切边连,将sgrna表达盒克隆到pylcrispr/cas9载体上。crispr/cas9载体骨架及编辑位点设计引物见图1b。利用pylcrispr/cas9双元载体构建strk2的crispr/cas9敲除株系。对strk2在可读框(open reading frame, orf)5
′
区设计crispr/cas9编辑靶点,利用pylcrispr/cas9双元载体(图1中a)构建strk2的crispr/cas9敲除株系。对strk2在orf 5
′
区设计crispr-cas9编辑靶点(图1中b),crispr/cas9系统靶点引物在crispr-ge网页进行靶点设计,根据靶点位置、gc含量、潜在脱靶位置以及估值等选择gc碱基含量较低(45%-70%之间脱靶效率较低)且符合要求的靶点引物。将构建成功的表达盒子连入pylcrispr/cas9双元载体,并转入农杆菌eha105中,经农杆菌介导侵染转化水稻kitaake愈伤组织,获得敲除株系的t0代幼苗。提取各敲除株系叶片的dna,对包含编辑位点的一段区域进行pcr扩增,通过与野生型的序列比对鉴定strk2编辑靶位点的编辑情况。在t1代鉴定到2种编辑方式的纯合strk2敲除株系,并命名为strk2-25和 strk2-29(图1中c-d)。
32.2. strk2蛋白结构及亚细胞定位分析由结构分析可知,strk2为典型亮氨酸重复序列类受体蛋白激酶(lrr-rlk),具有胞外配体结合结构域、跨膜结构域和胞内激酶结构域(图2中a)。根据swiss-model数据库分析,strk2的蛋白三维结构如图2b所示。strk2在水稻各个阶段的器官中均有表达,其在幼嫩的叶、根及种子中的表达量比较高(图2中c)。将strk2全长cds序列构建到1300-yfp的亚细胞定位载体,于水稻原生质体中表达绿色荧光蛋白(gfp)-strk2的融合蛋白。通过激光共聚焦显微镜检测发现,gfp-strk2定位于原生质体的细胞膜,与专一性的细胞膜定位染料fm4-64的荧光信号高度重合(图2中d),说明strk2定位于细胞膜,与其具有胞外结构域、跨膜结构域及胞内结构域的蛋白结构是一致的。
33.3. strk2过表达植株及crispr/cas9敲除株系的盐胁迫表型鉴定将野生型、strk2过表达及敲除株系的15 d幼苗经140 mm nacl处理15 d,发现与野生型(wt)相比,strk2过表达株系strk2-4和strk2-8表现出对盐胁迫敏感的表型,而敲除株系strk2-25和strk2-29表现出明显的对盐胁迫的耐受性(图3中a)。再经正常水培恢复5 d后统计存活率可知,过表达株系strk2-4和strk2-8的存活率显著低于野生型,而敲除株系strk2-25和strk2-29的存活率显著高于野生型,表现表现出较高的耐盐性(图3中a和b)。以250 mm nacl处理刚萌发露白的水稻种子5 d,然后观察幼苗材料在盐胁迫条件下的表型。在正常生长条件下,野生型与所有转基因材料的幼苗生长没有明显差异;250 mm nacl处理条件下,与野生型相比,过表达株系strk2-4和strk2-8的生长被显著抑制,苗长显著降低,而敲除株系strk2-25和strk2-29的苗长显著升高,表现出对盐胁迫较高的耐受性(图3中c)。因此,在苗期的盐胁迫条件下,strk2过表达株系表现为盐敏感的表型,而敲除株系则表现为较高的耐盐性。
34.4. strk2与下游蛋白lrrk1相互作用植物中rlk-rlck相互作用,且通过磷酸化方式转导胁迫信号是普遍存在的。我们推测strk2作为rlk能够在盐胁迫条件下感知na
信号并实现信号的跨膜转导,然后通过下游的rlck将信号转导至相关的效应蛋白分子。为了进一步揭示strk2调控盐胁迫响应的机制,我们通过免疫共沉淀(co-ip)从水稻中分离strk2的蛋白复合体,并进行质谱鉴定。结果从复合体中筛选和鉴定到一个rlck蛋白lrrk1,并对lrrk1与strk2的相互作用进行了验证。酵母双杂实验证明,lrrk1能与strk2互作(图4中a)。进一步的分段互作分析发现,lrrk1与strk2的直接相互作用是发生在lrrk1a段,即lrrk1的n端(图4中b)。随后的双分子荧光互补(bimolecular fluorescence complementation, bifc)实验也再次验证lrrk1与strk2在水稻原生质体的细胞膜上相互作用(图4中c)。
35.5. 类受体激酶strk2能够磷酸化下游lrrk1首先,在大肠杆菌(e. coli)bl21中原核表达并纯化lrrk1蛋白(his-lrrk1和gst-lrrk1)及strk2蛋白(his-strk2和gst-strk2),然后在体外磷酸化反应体系中孵育。经磷酸化反应后的体系进行10%分离胶sds-page,然后以抗his、gst及磷酸化丝氨酸(pser)的抗体进行免疫杂交分析。研究结果发现,当strk2与lrrk1共孵育时,lrrk1的激酶失活突变体(lrrk1ke)失去自我磷酸化活性和激酶活性,但strk2仍能磷酸化lrrk1,且lrrk1ke磷酸化水平仍然显著升高,说明strk2可以磷酸化lrrk1(图5中 a)。将strk2与lrrk1共同孵育,可检测到lrrk1磷酸化水平显著升高;当激酶失活蛋白strk2(gst-strk2ke)与lrrk1进行磷酸
化反应,gst-strk2ke并未出现磷酸化平升高,说明lrrk1不能磷酸化strk2(图5中b)。当单独孵育strk2与lrrk1,以磷酸化丝氨酸抗体检测发现,lrrk1的磷酸化水平显著上升(图5中c)。由此可知,strk2可磷酸化lrrk1,通过磷酸化方式将信号传递到下游效应分子。
36.6. lrrk1的磷酸化水平受盐胁迫诱导构建了flag-lrrk1的过表达转基因水稻,用于研究盐胁迫对lrrk1磷酸化水平的影响。在盐胁迫条件下将lrrk1过表达株系幼苗用100 mm nacl分别处理0、20、40和60 min,液氮研磨后ip buffer提取蛋白,然后用flag beads免疫沉淀融合蛋白flag-lrrk1。flag-lrrk1蛋白进行sds-page后,以抗pser抗体检测磷酸化水平。结果显示,随着nacl处理时间的延长,lrrk1的磷酸化水平逐渐升高(图6),说明strk2对lrrk1的磷酸化程度与nacl处理时间相关。我们推测,strk2通过磷酸化lrrk1而参与盐胁迫响应调控。
37.7. 盐胁迫下lrrk1促进水稻体内活性氧(ros)积累和抗坏血酸(asa)含量下降植物受到非生物胁迫后,ros的爆发是一个标志事件。采用ros荧光探针dcfh-da检测发现,正常条件下,野生型及lrrk1转基因株系中的ros荧光信号强度没有显著差别;盐胁迫处理后,lrrk1过量表达株系中ros荧光信号强度显著高于野生型,而其敲除株系lrrk1-1的ros荧光信号强度则显著低于野生型(图7中a)。对ros中最稳定的h2o2进行测定发现,正常条件下,野生型及lrrk1转基因株系中的h2o2含量没有显著差别;盐胁迫处理后,lrrk1过量表达株系中h2o2的含量高于野生型,而其敲除株系lrrk1-1则低于野生型(图7中b)。另外,超氧阴离子(o2·-)含量的测定结果与h2o2含量测定结果的趋势一致(图7中c)。asa是植物体内清除ros的重要抗氧化剂之一,在ros 的清除机制中发挥着重要作用。为了探究lrrk1对水稻体内asa的影响,我们对lrrk1转基因株系在盐胁迫下的asa含量进行测定。结果发现,在正常生长条件下,lrrk1过表达株系和敲除株系lrrk1-1的asa含量与野生型的无显著差异;经过盐胁迫处理之后,lrrk1-1的asa含量显著高于野生型,而过表达株系显著低于野生型(图7中d)。该结果说明,lrrk1通过负调控asa的生物合成而参与水稻的盐胁迫响应。
38.由此可以推知,strk2通过磷酸化lrrk1来负调控水稻体内asa的合成,从而最终参与水稻盐胁迫响应的调控。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。
