1.本发明属于生物农药技术领域,具体涉及抗真菌花生聚糖索马甜类蛋白 pg-tlp的制备和该抗菌花生聚糖索马甜类蛋白在制备抗真菌中的应用。
背景技术:
2.自从抗生素被发现以来,针对真菌感染性疾病的治疗有了根本的改善,抗生素的使用拯救了农作物经济,增加了农作物的产量。但随着抗生素的大规模使用或滥用,尤其是在发展中国家,导致了一些抗药性菌的产生和扩散,其中包括一些毒力很强的致病菌,像耐甲氧西林葡萄球菌与肺炎链球菌等。因此,寻找安全有效且不易产生耐药性质的抗菌药物成为全世界科学家竞争和努力的方向。
3.高等植物具有广泛的防御机制来应对物理、化学和生物胁迫,例如干旱、寒冷、重金属、污染物和来自真菌、细菌和病毒的病原体攻击。为应对多种病原体的感染,植物表现出上调一组与系统获得性抗性相关的基因 [stintzi a.,heitz t.,prasad v.,wiedemann-merdinoglu s.,kauffmann s., geoffroy p.,legrand m.,fritig b.plant“pathogenesis-related”proteins and their role in defense against pathogens.biochimie.1993;75:687
–
706.]。一般抗性是通过释放次级代谢物如植物抗毒素、单宁和多酚化合物以及产生发病机制相关(pr)蛋白来实现的。pr蛋白于1970年代初首次在烟草叶中发现,以应对烟草花叶病毒感染,后来被定义为在致病性发作期间释放的诱导蛋白[stintzi a.,heitz t.,prasad v.,wiedemann-merdinoglu s.,kauffmann s., geoffroy p.,legrand m.,fritig b.plant“pathogenesis-related”proteins and their role in defense against pathogens.biochimie.1993;75:687
–
706;sinha m., singh r.p.,kushwaha g.s.,iqbal n.,singh a.,kaushik s.,kaur p.,sharma s., singh t.p.current overview of allergens of plant pathogenesis related protein families.sci.world j.2014;2014:543195.]。根据最近的一项综述,至少有17 个家族已被检测和分离,它们具有广泛的防御相关特性,包括抗菌、抗真菌、抗病毒、抗氧化活性、几丁质酶和蛋白酶抑制活性[stintzi a.,heitz t., prasad v.,wiedemann-merdinoglu s.,kauffmann s.,geoffroy p.,legrand m., fritig b.plant“pathogenesis-related”proteins and their role in defense against pathogens.biochimie.1993;75:687
–
706;sinha m.,singh r.p.,kushwaha g.s., iqbal n.,singh a.,kaushik s.,kaur p.,sharma s.,singh t.p.current overview of allergens of plant pathogenesis related protein families.sci.world j. 2014;2014:543195;ebrahim s.,usha k.,singh b.pathogenesis related(pr) proteins in plant defense mechanism.sci.against microb.pathog. 2011;2:1043
–
1054;4.sels j.,mathys j.,de coninck b.m.,cammue b.p.,de bolle m.f.plant pathogenesis-related(pr)proteins:a focus on pr peptides.plant physiol.biochem.2008;46:941
–
950.],包括抗菌活性的肽,即硫素(pr-13 家族)、防御素(pr-12家族)、hevein样肽、knottin、α-发夹蛋白、脂质转
移蛋白(pr-14家族)和蛇形蛋白.
[0004]
索马甜类蛋白是过氧化物酶家族的成员,参与植物-微生物相互作用中的效应触发免疫。从香蕉中纯化的索马甜类蛋白通过诱导质膜紊乱和细胞壁解体对p.expansum发挥抗真菌活性[jiao,w.,li,x.,zhao,h.,cao,j.,& jiang,w.(2018).antifungal activity of an abundant thaumatin-like protein frombanana against penicillium expansum,and its possible mechanisms of action. molecules,23(6),1442.]。也有报道这种索马甜类蛋白抑制一些其它真菌(f. oxysporum f.sp.niveum race,fusarium solani f.sp.cucurbitae race,f. oxysporum f.sp.melonis,fusarium verticillioides and didymella bryoniae,l. piceae,fusarium proliferatum,botrytis cinerea,roussoella doimaesalongensi, rhizoctonia solani,和alternaria alternata)。在转基因马铃薯植物中过表达拟南芥索马甜类蛋白可增强对早疫病和晚疫病的抗性[.ali,g.s.,hu,x.,& reddy,a.s.n.(2019).overexpression of the arabidopsis thaumatin-likeprotein 1in transgenic potato plants enhances resistance against early and lateblights.biorxiv,621649.]。小麦马甜类蛋白抑制酿酒酵母生长[van der maelen,e.,rezaei,m.n.,struyf,n.,proost,p.,verstrepen,k.j.,&courtin,c. m.(2019).identification of a wheat thaumatin-like protein that inhibitssaccharomyces cerevisiae.journal of agricultural and food chemistry,67(37), 10423-10431.]。一些索马甜类蛋白被认为具有糖基化位点,蛋白的聚糖修饰对蛋白的活性和稳定性有很重要的作用[thimme gowda,c.,purama,s.n. s.,&kammara,r.(2020).tlpdb:a resource for thaumatin-like proteins. the protein journal,39(4),301-307.]。但是索马甜类蛋白的聚糖修饰很少有报道,我们在花生蛋白分离出有聚糖修饰的索马甜类蛋白,为真菌的控制提供了可靠的工具。
[0005]
黑曲霉和黄青霉属于活体营养型植物致病真菌,在营养上依赖于植物例如葡萄活细胞的新陈代谢。这类真菌属于活体营养型真菌类群。他们直接渗入到植物细胞表皮中,被渗入的细胞随后坏死。在渗入后,该真菌变换至专性活体营养型生活方式。主要遵照这样的感染策略的活体营养型真菌病原体亚群是半死体营养型。与半死体营养型病原体相比,半活体营养型病原体以活体营养型方式生活一段短时间,随后开始杀死寄主细胞和/或寄主生物。
[0006]
近年来,真菌病害例如黑曲霉在农业例如葡萄生产中作为病害越来越重要。比如锈菌病真菌中,由puccinia striiformis引起的小麦条锈病,是世界范围内最重要的小麦病害之一[chen,x.m.epidemiology and control of stripe rustpuccinia striiformis f.sp.tritici on wheat.canadian journal of plant pathology 2005;27.3:314-337.]。镰刀菌病真菌中,fusarium oxysporum在根际微生物区系中有很好的代表性。几乎所有菌株都以腐生方式存在,在植物上诱发枯萎或根腐病而闻名[fravel,d.,chantal olivain,and claude alabouvette. fusarium oxysporum and its biocontrol.new phytologist 2003;157.3: 493-502.]。诱导炭疽菌病的炭疽菌属物种存在于世界的热带和亚热带地区。是浆果病的病原体,在其发育的任何阶段感染叶子和浆果,多数是农作物褐枯病的病原体,经常感染成熟的浆果[chen z,franco cf,baptista rp, cabral jm,coelho av,rodrigues cj,melo ep.purification and identification of
cutinases from colletotrichum kahawae and colletotrichum gloeosporioides. applied microbiology and biotechnology.2007;73:1306-13.]。丝核菌真菌中,立枯丝核菌复合体代表了经济上重要的一组土传担子菌病原体,它们发生在世界各地的许多植物物种上[cubeta,m.a.,and r.vilgalys."populationbiology of the rhizoctonia solani complex."phytopathology 1997;87.4: 480-484.]。长期以来,我国农作物的病虫害防治主要依赖化学农药,农药污染成为食品安全生产中急需解决的难题。相比化学农药,生物农药具有安全、环保、长效、无残留等优点,注重低毒、高效、选择性强,成为当前的研究热点之一。
[0007]
因此,在本领域需要开发控制真菌生物制剂和提供抗真菌植物的方法。花生粕是从花生仁经压榨提炼油料后的产品,通常花生粕分一次粕,二次粕。一次粕的意思是经过初次压榨剩余的花生渣,二次粕即压榨过两次的花生渣。通常花生粕的产量可以达到44%以上,是一种丰富的副产品资源。花生粕中含有丰富的蛋白质和碳水化合物。我们从花生粕的浸泡液获得了一系列蛋白。我们对这些蛋白分离,发现有些蛋白有抗真菌活性,对其中的一条蛋白去聚糖修饰测序为花生索马甜类蛋白。该蛋白由于聚糖修饰比理论分子量多了4-14kda,对多种植物病原真菌有抗性,包括葡萄霉菌、黑曲霉和青霉属。
技术实现要素:
[0008]
本发明的目的在于提供一种新的用于农作物的抗真菌花生聚糖索马甜类蛋白pg-tlp及制备。
[0009]
一种抗真菌花生聚糖索马甜类蛋白pg-tlp,所述的抗真菌花生聚糖索马甜类蛋白pg-tlp的氨基酸序列,没有聚糖的序列包含246个氨基酸残基,分子量25.9kda,等电点8.34。花生聚糖索马甜类蛋白有两个糖基化位点(n208 和n239位置),每个糖基化位点聚糖的聚合度在15-40之间,主要单糖成分是甘露糖(mannose),n-乙酰葡糖胺(glcnac),半乳糖(galactose),木糖(xylose)和果糖(fucose)。聚糖结合以后的分子量在30kda-40kda 之间。
[0010]
一种如上所述的抗真菌花生聚糖索马甜类蛋白的制备方法,利用离子交换的方法,花生聚糖索马甜类蛋白通过离子交换层析和凝胶过滤层析逐级分离出来,通过反相柱层析脱盐纯化。
[0011]
所述的抗真菌花生聚糖索马甜类蛋白的应用,其特征是抗葡萄霉菌、黑曲霉和青霉属真菌。
[0012]
本发明的有益效果在于:本发明中的纯化的抗真菌花生聚糖索马甜类蛋白,具有分子量适中、储存稳定,杀真菌能力强等优点。抗真菌花生聚糖索马甜类蛋白对葡萄霉菌、黑曲霉和青霉属真菌均具有较强的抑菌作用,但对益生菌,比如枯草杆菌抑菌作用弱,毒性低,说明抗真菌花生聚糖索马甜类蛋白抗菌选择性好,毒副作用小。
附图说明
[0013]
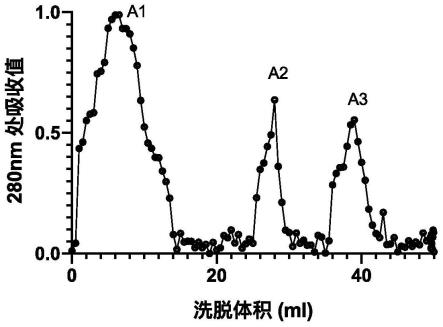
图1.sephacryl s-200色谱柱花生蛋白的洗脱曲线,形成a1(洗脱的第2 管),a2(洗脱的第6管),和a3峰(洗脱的第8管)。
[0014]
图2.分离的抗真菌花生聚糖索马甜类蛋白pg-tlp(a1,a2,和a3)对 a.niger和
p.chrysogenum抑菌透明区面积。
[0015]
图3.高压液相色谱分析抗真菌花生聚糖索马甜类蛋白的纯度。
[0016]
图4.电喷雾质谱鉴定抗真菌花生聚糖索马甜类蛋白pg-tlp。
[0017]
图5.抗真菌花生聚糖索马甜类蛋白pg-tlp分析。
[0018]
图6.平板培养法分析葡萄果品的真菌感染数。
[0019]
图7.葡萄果品呼吸频率、体重减轻、硬度比较。
[0020]
图8.ampd2抑菌效果比较。
[0021]
图9.糖苷合成酶活性(gs)比较。
具体实施方式
[0022]
根据下述实施例可以更好的理解本发明。然而,本领域的技术人员容易理解,实施例所描述的内容仅用于说明本发明,而不应当也不会限制权利要求书所详细描述的本发明。
[0023]
实施例1:抗真菌花生聚糖索马甜类蛋白pg-tlp的制备
[0024]
称取5.0g花生粕,于液氮中反复研磨成细粉状,于200ml烧杯中,加入 100ml蛋白质提取缓冲液(组成为:20mm三羟甲基氨基甲烷盐酸盐(tris-hcl),ph8.0,10mm乙二胺四乙酸(edta)、150mm nacl、体积浓度1%二甲基亚砜(dmso)和0.1%β-巯基乙醇,于水中),搅拌混匀。在10000g的转速下离心20min,取60ml上清。
[0025]
将767g(nh4)2so4边搅拌边慢慢加到1升蒸馏水中。用质量浓度25%- 28%氨水或95%硫酸调硫到ph7.0。此即饱和度为100%的硫酸铵溶液(4.1 mol/l,25℃).
[0026]
边搅拌边慢慢加入等体积的饱和度为100%的硫酸铵溶液到上清液中。将溶液放在磁力搅拌器上搅拌过夜(4℃),使蛋白质充分沉淀。蛋白质溶液 10000g离心30min(4℃)。弃上清保留沉淀。将沉淀溶于20ml蛋白质提取缓冲液(组成为:20mm tris-hcl,ph8.0,10mm edta、150mm nacl、 1%dmso,0.1%β-巯基乙醇和0.2g/l叠氮钠,于水中)。放入40ml透析袋 (截留分子量10kda),用1升蛋白质提取缓冲液透析24小时(4℃),每隔6小时换透析缓冲液一次,以彻底除去硫酸氨。用三柱体积的蛋白质提取缓冲液(组成为:20mm tris-hcl,ph8.0,10mm edta、150mm nacl、体积浓度1%dmso和0.1%β-巯基乙醇,于水中)平衡deae-sepharose fastflow柱(内径与长度2.6
×
100cm,默克,型号17-0709-10),流速0.5 ml/min。将25ml粗提溶液加载到deae-sepharose fast flow柱(2.6
×
100 cm)上,再次用三柱体积的蛋白质提取缓冲液(组成为:20mm tris-hcl, ph8.0,10mm edta、150mm nacl、体积浓度1%dmso和0.1%β-巯基乙醇,于水中)平衡deae-sepharose fast flow柱(2.6
×
100cm),流速0.5 ml/min。最后用蛋白质提取缓冲液(组成为:20mm tris-hcl,ph8.0,10mmedta、150mm nacl、1%dmso和0.1%β-巯基乙醇,于水中)和0.3m nacl 梯度以1.0ml/min的流速洗脱柱子。从检测器可以检测到蛋白开始收集蛋白洗脱液,到检测器不能检测到蛋白结素,共收集蛋白45ml。将收集物加到用3柱体积20mm tris-hcl缓冲液(ph 8.0)(内含0.15m nacl)平衡的凝胶过滤柱(sephacryl s-200,内径与长度5x90 cm,美国ge公司,型号17-0584-01)上,收集3-ml级分。以0.5ml/min的20mm tris-hcl缓冲液(ph 8.0)(内含0.3m nacl)进行洗脱。从检测器可以检测到蛋白开始,每5ml收集一管,共收集9管。通过280测量nm吸光度,集中显示较高蛋白浓度的峰是图1.a1(第2管)、a2(第6管),和a3(第8管)。对3部
分蛋白(a1(第2管)、a2(第6管),和a3(第8管)),分别分析每一部分的抗真菌活性。使用马铃薯葡萄糖琼脂平板(直径9cm,厚度0.3 cm)(配方如下:马铃薯20g制成浸出液,葡萄糖2g,琼脂1.5g,水100 ml,不调节ph(ph自然)。制备方法:取新鲜马铃薯20g,洗净去皮切成小块,加水80毫升煮沸30min,用四层纱布过滤,加入葡萄糖2g和琼脂 1.5g,继续加热搅匀,稍冷却后补足水分至100ml,121℃灭菌20分钟)。平板中央滴加10μl冻存的菌液(分别为a.niger和p.chrysogenum,菌体密度为1
×
105~2
×
105cfu/g冻干粉,每一克样品溶于10毫升的质量浓度 0.85%生理盐水)。分别评估洗脱的花生蛋白的第2,6,8管花生蛋白的抗真菌活性。将直径为1cm的圆形无菌纸盘,以平板的中央(为中心)画“十”字形,中心交点放一纸盘,沿二条线向远离中心方向分别距离中心每1.5cm 处放一纸盘,直径9cm的平板共放5个纸盘。将在20mm pbs,ph7.5缓冲液中的蛋白10μl滴加到纸盘上,对照只滴加10μl的20mm pbs, ph7.5。在25℃下培养72小时,菌丝体生长包围含有对照的外周圆盘,并在带有抗真菌样品的圆盘周围产生透明圈抑制。透明圈的面积越大代表蛋白的抑制率越高。抑菌实验显示洗脱的花生聚糖索马甜类蛋白pg-tlp 中a2(洗脱的第6管)对a.niger和p.chrysogenum有明显的抑菌活性, a1(第2管),和a3(第8管)对a.niger和p.chrysogenum抑菌活性很弱(图2)。因此,a2(洗脱的第6管)可能是理想的抑菌花生蛋白,命名为花生聚糖索马甜类蛋白pg-tlp,用于后续实验。
[0027]
图1.sephacryl s-200色谱柱花生蛋白的洗脱曲线,形成a1(洗脱的第2 管),a2(洗脱的第6管),和a3峰(洗脱的第8管)。以5ml/min的20 mm tris-hcl缓冲液(ph 8.0)(内含0.35m nacl)进行洗脱。从检测器可以检测到蛋白开始,每5ml收集一管,共收集9管。通过280测量nm吸光度,集中显示较高蛋白浓度的峰是a1(洗脱的第2管),a2(洗脱的第6 管),和a3峰(洗脱的第8管)。
[0028]
图2:分离的抗真菌花生聚糖索马甜类蛋白pg-tlp(a1,a2,和a3)对 a.niger和p.chrysogenum抑菌透明区面积。每个抑菌圈的直径测量3次,计算抑菌圈直径的平均值,根据面积公式(s=π*(d/2)2,π表示圆周率,r 表示半径,d表示直径,s代表面积)求透明圈的面积。
[0029]
实施例2:实施例1获得的抗真菌花生聚糖索马甜类蛋白pg-tlp的分子量和序列鉴定为了测定原序列进行了蛋白去聚糖化。去糖基化步骤如下:用100mm甘氨酸-hcl ph 2.5配制1ml 100pmol pg-tlp。2分钟后,将100μl样品转到含有5μl n-糖苷酶f(一种酰胺水解酶,可以裂解由天冬酰胺连接的高甘露糖、杂合和复杂的寡糖糖蛋白中聚糖和蛋白之间的糖苷键)的试管中。 n-糖苷酶f消化去糖基化在4℃下进行30分钟。样品注入液相质谱 (lc-ms)系统检测n-糖苷酶f作用后的抗真菌花生聚糖索马甜类蛋白去糖基化程度,达到理论分子量25.9kda,即246个氨基酸残基的分子量表明所有的糖基化基团被n-糖苷酶f全部消化掉。
[0030]
1ml抗真菌花生聚糖索马甜类蛋白pg-tlp或去糖基的pg-tlp通过液相色谱(hplc)反相柱(welchxb,内径与长度c18 4.6
×
150mm)层析脱盐纯化,流动相为40%v/v纯水(含体积浓度0.1%三氟乙酸(tfa))-60%v/v乙腈 (acn)(含体积浓度0.1%三氟乙酸),流速为1ml/min,220nm波长下检测,收集流出峰,冻干,然后重新悬浮在体积浓度0.1%甲酸中用于液相色谱-质谱(hplc-ms)联用分析。使用电喷雾质谱对合成的抗真菌花生聚糖索马甜类蛋白pg-tlp的分子量进行鉴定。样品使用液相系统进样,流动相为体积浓度50%h2o/50%
can,流速为0.2ml/min,保护气体氮气流速1.5 l/min,碰撞能量4.5kv,使用阴离子模式。
[0031]
纯化的抗真菌花生聚糖索马甜类蛋白pg-tlp用高效液相色谱hplc方法 (welch xbc,内径与长度184.6x250mm)鉴定其纯度,分子量测定采用电喷雾质谱结构。
[0032]
高效液相色谱hplc纯度鉴定结果如图3所示:抗真菌花生聚糖索马甜类蛋白pg-tlp去糖基化后在25.6min处显示为单峰(图3a)。抗真菌花生聚糖索马甜类蛋白pg-tlp在28-42min处显示为宽峰(图3b)。
[0033]
图3:高压液相色谱分析抗真菌花生聚糖索马甜类蛋白的纯度。a,去聚糖后。 b,未去聚糖修饰。
[0034]
电喷雾质谱鉴定结果如图4所示:去掉聚糖修饰后,蛋白的分子量接近理论数值25.9kda(图4a)。未去糖基化前抗真菌花生聚糖索马甜类蛋白 pg-tlp分子量为30-40kda,比理论分子量多了4-14kda(图4b)。
[0035]
图4:电喷雾质谱鉴定抗真菌花生聚糖索马甜类蛋白pg-tlp。a,去聚糖后。b,未去聚糖修饰。标记分子量的三个峰(多糖的聚合度分别为32,39和 87)分子量差是162的整倍数,代表相应的葡萄糖和/或半乳糖单元。
[0036]
实施例3:实施例1获得的抗真菌花生聚糖索马甜类蛋白pg-tlp测序参考以前的文献使用异硫氰酸苯酯法(edman)降解法对抗真菌花生聚糖索马甜类蛋白pg-tlp进行测序[sels j.,mathys j.,de coninck b.m.,cammueb.p.,de bolle m.f.plant pathogenesis-related(pr)proteins:a focus on prpeptides.plant physiol.biochem.2008;46:941
–
950.]。将纯化去糖基化的花生索马甜类蛋白(20μg)溶于50μl 0.2m碳酸氢铵(ph 8.0,其中含有4m盐酸胍)与5ml 45mm二硫苏糖醇混合。在自动蛋白质测序仪(appliedbiosystems,476a型)上进行s-羧酰胺甲基化肽的自动edman降解和苯硫基乙缩醛衍生物的检测。在6mol/l hcl存在的情况下,裂解发生在第一个肽键上,从而得到减去第一个碱基的肽段和释放的第一个 anilinothiazolinone(atz)形式的残基。用20mm trishcl(ph8.0)缓冲洗掉其他反应物和释出的残基,缩短的肽段就可以通过另一轮偶联和裂解而释放出第二个残基,依此类推,直到最后一个氨基酸残基被释出。测序结果,经确定为花生抗真菌花生聚糖索马甜类蛋白pg-tlp。
[0037]
序列表seq id no.1为:
[0038]
maiiitrvialfflgfaflacvaqgarvtfknkcqytvwpgtltgsqkaq lsqtgftlapgatnslnlpsswsgrfwgrtgcsnnggrftcatgdcgtg qvacngnggatpatlveitvasnggqdfydvsnvdgfnlplsvstqggr gtcktsscptninracpselqvkgsngnvvgcksacvafgkpeycctgs hntektcpptnyskffkqqcpdaysyayddlrstftcsgkpnytitfcp。
[0039]
(246aa)
[0040]
(a)序列特征:
[0041]
·
长度:246
[0042]
·
类型:氨基酸序列
[0043]
·
链型:单链
[0044]
·
拓扑结构:线性
[0045]
(b)分子类型:蛋白质
[0046]
(c)假设:否
[0047]
(d)反义:否
[0048]
(e)最初来源:花生
[0049]
实施例4:实施例1获得的抗真菌花生聚糖索马甜类蛋白pg-tlp测序糖基化成分分析
[0050]
胰蛋白酶消化
[0051]
1ml实施例1获得的抗真菌花生聚糖索马甜类蛋白溶液(1mg/ml)中加终浓度为10mm二硫苏糖醇(dtt),在55℃下还原二硫键20分钟。冷却至室温后,加入终浓度为20mm碘乙酰胺(iaa),在室温下在黑暗中诱导蛋白质烷基化30分钟。用1mg/ml胰蛋白酶(酶:底物溶液=1:50(w/v、mg/ml))在37℃下消化16小时,同时轻轻摇动。消化的肽用c18 (phenomenex jupiter,填料粒径15μm,飞诺美公司,型号00g-4053-p0) 固相萃取柱辛基非极性柱脱盐,并在真空干燥器中干燥2小时。
[0052]
完整糖基化肽段的富集与质谱解析
[0053]
用100μl结合缓冲液at(体积浓度80%乙腈(can)、体积浓度1%三氟乙酸(tfa))使用脱盐柱对实施例4获得的胰蛋白酶消化的聚糖索马甜类蛋白进行脱盐。具体步骤如下,用100μl acn激活柱子亲水色谱柱(默克色谱柱zic-hilic,型号:1.50478.0001),并用300μl结合缓冲液at平衡,将上面胰蛋白酶消化的实施例4获得的胰蛋白酶消化的聚糖索马甜类蛋白500μl加载到亲水色谱柱zic-hilic上样,5分钟。随后,用800μl at结合缓冲液洗涤柱子,5分钟。最后,n-糖肽用300μl体积浓度0.1% tfa 洗脱,最后用100μl 50mm nh4hco3洗脱1次。洗脱液在真空干燥器中干燥并重新悬浮在100μl体积浓度0.1%tfa中。
[0054]
反相液体色谱法电喷雾质谱(rplc esi-ms/ms)分析在反相液体色谱法电喷雾质谱系统上分析花生聚糖银黄素pg-ahgp和未糖基化的花生银黄素质量(100μl 1mg/ml蛋白),在正离子模式下运行的光谱仪。在c18色谱柱(waters,5μm,4.6mm x 250mm,型号 186001265)上以300nl/min的流速进行色谱分离,流动相为体积浓度 99.8%h2o、0.2%tfa(a)和体积浓度95%acn、4.8%h2o和0.2%tfa (b)。lc梯度如下:2%b,用时10min;线性2%-40%b,用时190分钟;线性40%-95%b,用时10分钟;95%b,用时5分钟;线性95%-2% b,用时5分钟;2%b,用时20分钟。esi条件如下:喷雾电压2.8kv,毛细管温度320℃,高压环形离子导入装置(s-lens rf)电平75v。全扫描质谱在700-2000m/z范围内获得,主要esi源设置:显微扫描1、质量分辨率70k,自动增益控制(agc)目标2e5,最大离子注入时间50ms;msms 光谱是在top20数据相关采集模式下采集的,设置如下:微扫描1,质量分辨率17.5k,agc目标5e5,最大离子注入时间250ms,隔离窗口3 m/z,更高能量的碰撞解离(hcd),阶梯式归一化碰撞能量20%、30%、 40%;动态排除20秒。
[0055]
结合实施例3和4,一种抗真菌花生聚糖索马甜类蛋白pg-tlp,所述的抗真菌花生聚糖索马甜类蛋白pg-tlp的氨基酸序列为如图5a所示。没有聚糖的序列包含246个氨基酸残基,分子量25.9kda,等电点8.34。聚糖的聚合度在15-40之间,真菌花生聚糖索马甜类蛋白pg-tlp氨基酸序列中 n208和n239位置是索马甜类蛋白聚糖结合的位置,n-乙酰葡糖胺与天冬酰胺的γ-酰胺n原子形成糖苷键(图5a)。花生聚糖索马甜类蛋白糖型主要有4种(图5b),天冬酰胺一糖苷键的形式线性连接2个n-乙酰葡糖胺,再结合结合多个分支状线性连接甘露糖单元聚糖(75%-90%,按聚糖总质量计),后面以同样的方式连接,n-乙酰葡糖胺结合多个甘露糖单元,外加半乳糖修饰聚糖(2%-15%,按聚糖总质量计),n-乙酰葡糖胺结
合多个甘露糖单元,外加半乳糖和木糖修饰聚糖(0%-10%,按聚糖总质量计), n-乙酰葡糖胺结合多个甘露糖单元,外加半乳糖,木糖,果糖修饰聚糖 (0%-5%,按聚糖总质量计)。
[0056]a[0057][0058]
图5:抗真菌花生聚糖索马甜类蛋白pg-tlp分析。a,氨基酸序列。b,聚糖种类和结合位置。真菌花生聚糖索马甜类蛋白pg-tlp氨基酸序列中 n208和n239(双下划线)位置是索马甜类蛋白聚糖结合的位置。
[0059]
实施例5:实施例1获得的pg-tlp对葡萄感染真菌的防治
[0060]
葡萄到达实验室后,在30分钟内根据统一的颜色、大小、无损伤以及健康或绿色的葡萄串选择葡萄串。然后将选定的串葡萄浸入实施例1获得的抗真菌花生聚糖索马甜类蛋白pg-tlp(0.1mg/ml,200ml生理盐水为溶剂) 溶液1分钟,在无菌空气(20℃)下表面干燥,放入纸板箱中,然后在最佳冷藏温度(-1至0℃)下储存在冰箱里,作为处理组。同等(或同样) 的3串葡萄只有200ml生理盐水处理,作为对照(过程和条件同前处理组)。葡萄表面的真菌污染和生物指数每5天测量一次,测量过程为:从每串葡萄中随机选择五个浆果,称重并浸渍在50ml无菌0.1%蛋白胨(w/v, g/ml)。剧烈摇晃5分钟以去除葡萄表面的真菌,获得洗涤液。洗涤液用水连续稀释3次至稀释前体积的10倍,以分别获得1/10、1/100和1/1000的稀释度的溶液。将每种稀释溶液(0.1ml)涂在马铃薯琼脂培养基ypd固体平板上(直径9cm,培养基厚度3-4mm)。平板在25℃下孵育2天,cfu 数以每克表示。对观察到的菌落,利用显微镜确定是霉菌菌丝(无隔膜菌丝(无隔菌丝)和有隔膜菌丝(有隔菌丝),呈长管状,宽度2~10微米,可不断自前端生长并分枝,无隔或有隔,具1至多个细胞核),然后根据平板计数法确定感染的真菌数量。结果如下:储藏30天后,对照组真菌感染数量明显高于对照组(图6)。
[0061]
图6:平板培养法分析葡萄果品的真菌感染数。对照,生理盐水。处理组,生理盐水溶解的pg-tlp溶液。
[0062]
实施例6:实施例1获得的pg-tlp的使用对果品质量的影响
[0063]
使用果蔬呼吸器(gxh-3051,北京,中国)对实施例5中储存的葡萄的呼吸频率进行检测,结果以co
2 mg/kg
·
h表示。通过记录从各组鲜食葡萄在储存期间的重量差异(储存前的重量为初始重量-储存后5,10,15,20,25和 30天后重量),重量损失率测量为重量差异与初始重量的百分比。使用 tvt-6700质地分析仪(perten,北京,中国)以25%的变形和5毫米直径的圆柱形探头以1毫米/秒的前速和后速测量葡萄硬度。在储存期间对每个处理的10个葡萄浆果进行测量,硬度以牛顿(n)表示。储藏30天后,对照组的呼吸率和重量丢失率明显高于对照组(图7a和7b)。相反,储藏30 天后,对照组的葡萄硬度明显低于对照组(图7c)。
[0064]
图7.葡萄果品呼吸频率、体重减轻、硬度比较。对照,生理盐水。处理组,生理盐水溶解的实施例1获得的抗真菌花生聚糖索马甜类蛋白pg-tlp 溶液。
[0065]
实施例7:实施例1获得的pg-tlp体外抑制a.niger和p.chrysogenum 效果
[0066]
取a.niger和p.chrysogenum冻存菌液各0.1毫升分别涂布在20毫升pda 培养基平板上(直径9cm,培养基厚度3-4mm),25℃下培养5天。然后用 20毫升无菌水分别冲洗霉菌和曲霉的表面的孢子。用无菌水调整孢子浓度为1
×
105个/毫升。将a.niger和p.chrysogenum孢子悬浮液(100μl) 分别涂在20毫升pda培养基平板上(直径9cm,培养基厚度3-4mm),上面均匀放置10个牛津杯中心(不锈钢小管,内径6mm,外径8mm,高10mm)。然后,将20μl pg-tlp(0.1mg/ml)或生理盐水(对照)添加到牛津杯中心(直径8毫米)在25℃下培养。3天后测量抑菌圈的直径。结果显示,短肽有很好的抑菌效果,0.1mg/ml实施例1获得的抗真菌花生聚糖索马甜类蛋白pg-tlp溶液可以完全抑制两种霉菌糖苷酶的活性(图8,p《0.0001)
[0067]
图8.ampd2抑菌效果比较。对照,生理盐水。处理组,生理盐水溶解的实施例1获得的抗真菌花生聚糖索马甜类蛋白pg-tlp溶液。
[0068]
实施例8:实施例1获得的pg-tlp对a.niger和p.chrysogenum的糖苷合成酶活性影响
[0069]
取a.niger和p.chrysogenum冻存菌液0.1毫升分别涂布在20毫升pda 培养基平板上(直径9cm,培养基厚度3-4mm),25℃下培养5天。两种霉菌的菌丝分别都用20毫升蒸馏水冲洗下来,并将冲洗下来的菌丝冻干。将干燥的菌丝(1.0g)添加到4ml醋酸钠缓冲液(50mmol/l,ph=5)中并在预冷(0-4℃)的研钵中研磨以获得匀浆。然后将匀浆以10,000g离心15 分钟以获得含有粗糖苷合成酶(gs)的上清液。使用bca检测试剂盒测定含有粗酶的上清液的蛋白质含量。粗酶上清液(1.0ml)根据文献测定gs 活性[kawaguchi,k.,fox,j.,holmes,e.,boyer,c.,&preiss,j.de novo synthesis of escherichia coli glycogen is due to primer associated with glycogen synthase and activation by branching enzyme.archives of biochemistry and biophysics.1978;190:385-397]。酶活性表示为酶单位/mg蛋白质。结果显示,实施例1获得的抗真菌花生聚糖索马甜类蛋白pg-tlp有很好的抑制霉菌糖苷合成酶的活性(图9,p《0.0001)
[0070]
图9.糖苷合成酶活性(gs)比较。对照,生理盐水。处理组,生理盐水溶解的实施例1获得的抗真菌花生聚糖索马甜类蛋白pg-tlp(0.1mg/ml)溶液。
[0071]
附录
[0072]
seq id no.1
[0073]
抗真菌花生聚糖索马甜类蛋白pg-tlp氨基酸序列
[0074]
maiiitrvialfflgfaflacvaqgarvtfknkcqytvwpgtltgsqkaq lsqtgftlapgatnslnlpsswsgrfwgrtgcsnnggrftcatgdcgtg qvacngnggatpatlveitvasnggqdfydvsnvdgfnlplsvstqggr gtcktsscptninracpselqvkgsngnvvgcksacvafgkpeycctgs hntektcpptnyskffkqqcpdaysyayddlrstftcsgkpnytitfcp。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。