水稻osckx3基因在调控水稻叶夹角中的应用
技术领域
1.本发明属于植物基因工程领域,具体涉及水稻osckx3基因在控制水稻叶夹角大小中的应用。
背景技术:
2.到目前为止,我国年粮食总产量已超过6.6亿吨,其中水稻产量约占32%,但是随着我国人口增加和生活水平的提高,人们对粮食产量的需求日益增加。在耕地面积不变的现状下,粮食产量尤其是水稻产量的大幅提高迫切需要科学家培育出高产、优质作物新品种。叶夹角是指叶片和茎的倾角,其对于水稻的受光面积具有重要影响,是理想株型的重要因素,在作物培育中具有重要作用,直立的叶片适宜密植,可减少植株间的相互遮蔽,增加光合效率,提高作物产量(tong and chu,2018)。叶夹角的大小主要由叶枕控制,叶枕发育过程中的细胞分裂和伸长能导致叶片倾角的改变(feng et al.,2016)。
3.植物激素在调节叶夹角过程中发挥重要作用,已有研究发现油菜素内酯(brassinosteroid, br)、赤霉素(gibberellic acid,ga)和生长素(iaa)均参与调控叶夹角的形成(tong and chu, 2018;duan et al.,2019)。细胞分裂素(cytokinin,ck)是调控农作物形态生理和产量的重要激素之一,与干细胞分裂、穗发育、株型、氮肥利用、环境胁迫等均密切相关(hwang et al.,2012; wang et al.,2018;yang et al.,2021)。但是ck调控叶夹角的分子机制尚未见报道。作为调控 ck水平的重要酶细胞分裂素氧化酶(cytokinin dehydrogenase,ckx)可以催化ck氧化分解生成腺嘌呤和侧链,使ck彻底丧失活性,此反应不可逆(ashikari et al.,2005),因此ckx 对于ck活性水平的调节发挥关键作用。ckx基因表达量的下降可以显著提高ck含量,促进顶端分生组织的形成,拟南芥ckx3ckx5双突变体的花数目、果荚数及种子数均明显增加,种子数量提高50%以上(bartrina et al.,2011);水稻ckx2和dst(osckx2正向调节基因)突变体中ck含量显著增加,水稻穗粒数均增加20%以上(ashikari,2005;li et al.,2013)。水稻 osckx9通过d53参与独脚金内酯(strigolactone,sl)信号途径,调节水稻分蘖芽的发育(duanet al.,2019);水稻ckx11参与aba代谢途径调控叶片衰老(zhang et al.,2021)。水稻osckx4 还参与调控zn元素的吸收(gao et al.,2019)。ck和ckx基因已被证实参与水稻顶端分生组织发育、根部发育、叶片衰老以及分蘖芽发育等过程,但是ckx调控叶夹角形成的机制以及应用还尚未研究。
技术实现要素:
4.本发明要解决的技术问题是提供一种水稻细胞分裂素氧化酶osckx3基因在调控水稻叶夹角大小中的应用。
5.为解决上述技术问题,本发明提供一种水稻基因osckx3或其编码的蛋白在水稻作物改良中的应用:调控水稻叶夹角。
6.作为本发明应用的改进:调控水稻叶枕发育和叶夹角、改善水稻株型和提高水稻产量。
7.作为本发明应用的进一步改进:所述水稻基因osckx3的核苷酸序列如seq id no:1所示;还包括与其互补,同源,或该序列经过插入、突变几个核苷酸而形成的具有相同功能的核苷酸序列。
8.作为本发明应用的进一步改进:所述基因osckx3编码的氨基酸序列如seq id no:2所示。
9.作为本发明应用的进一步改进:敲除osckx3基因,使得叶夹角变小、籽粒增大。
10.作为本发明应用的进一步改进:过表达osckx3,使得水稻叶夹角增大。
11.作为本发明应用的进一步改进:osckx3突变体的核苷酸序列如seq id no:4~seq id no: 5的任一所示。
12.本发明还同时提供了一种用于编辑水稻osckx3基因的grna靶序列在调控植物叶夹角中的应用:所述grna靶序列为水稻osckx3基因的cds正义链的第332~351位,其核苷酸序列如seq id no.3所示。
13.本发明提出了一个调控水稻叶夹角的正向调控因子及其应用,利用反向遗传学方法筛选到osckx3基因通过参与水稻叶枕组织细胞的分裂和生长来调控叶夹角的大小。本发明研究了该基因调控水稻叶夹角的分子生物学功能及应用,为选育密植型的高产水稻新品种提供了一个潜在的新基因资源。
14.本发明获得了一种调控水稻叶夹角的正向调控因子,正向调控因子为基因osckx3,碱基序列如seq id no.1所示,氨基酸序列如seq id no.2所示。水稻osckx3基因在调控植物叶夹角中的应用具体为:osckx3基因通过调节叶枕处细胞的不对称生长调控叶枕发育,以调控水稻叶夹角的形成。
15.本发明的技术方案是这样实现的:
16.本发明通过筛选11个osckx基因突变体发现osckx3突变体叶夹角显著变小,不同发育时期的水稻叶夹角表型均证明osckx3调控叶夹角的大小。
17.应用时,构建osckx3的crispr/cas9载体,并利用农杆菌介导法转入日本晴,敲除osckx3基因,所获得的敲除突变体叶夹角显著变小,因此可通过该方法增加叶枕组织中细胞分裂素的含量,增加叶枕处细胞层数,减小叶夹角,改善水稻株型和种植密度,提高产量。
18.与现有技术相比,本发明具有如下技术优势:
19.1.osckx3基因在改变作物叶夹角同时,不削弱其它产量性状,可以创建株型紧凑、叶夹角小的水稻育种材料,对水稻育种具有重要的实际应用意义。
20.2.osckx3基因是改变叶夹角大小的有效基因,可以改变叶枕处细胞层数和维管束数目,进而改变水稻的叶夹角。
21.3.目前,尚未有细胞分裂素调控叶夹角的基因的报道,本发明的基因揭示了其通过控制叶枕细胞分裂素的含量,参与叶夹角大小的调控。
22.本发明的osckx3调控叶夹角的发现和应用,填补了ck调控叶夹角的空白,提供了一个改善水稻密植和提高水稻产量的潜在应用价值的基因。
23.需要说明的是:促进细胞分裂素含量的增加与调控水稻叶夹角之间并没有必然的联系。例如目前已知的osckx2基因、osckx11基因,其均具有促进细胞分裂素含量的增加的功能,但是没有任何关于与水稻叶夹角关联性的文章被报道。
24.综上,本发明涉及植物基因工程领域,具体涉及水稻细胞分裂素氧化酶osckx3基
因在调控植物叶夹角中的应用。本发明利用crispr/cas9技术敲除日本晴水稻基因osckx3,利用转基因技术获得osckx3过表达材料。与野生型相比,突变体叶夹角显著减小并且株型更直立而过表达材料叶夹角显著增大。激素分析证明突变体细胞分裂素含量显著增加,而叶枕细胞学结构观察发现突变体的叶枕近轴端和远轴端的细胞层数和维管束数量显著增加,表明细胞分裂素通过增加细胞数量改变叶夹角的形成。农艺性状分析表明突变体的水稻籽粒大小显著增加,过表达材料籽粒显著变小。因此基因工程技术敲除osckx3基因能够改变水稻叶夹角的形成和籽粒大小,从而改善植物株型和种植密度,提高产量。
附图说明
25.下面结合附图对本发明的具体实施方式作进一步详细说明。
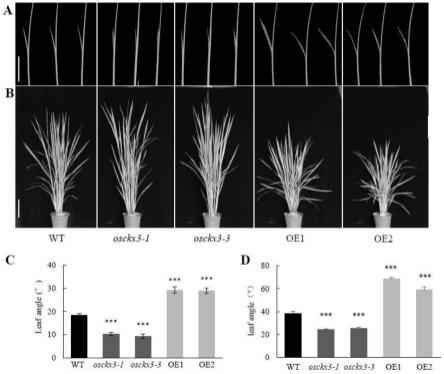
26.图1是本发明实施例提供的osckx3突变体的创制图和过表达材料osckx3基因表达量;
27.图1中:
28.a为osckx3基因结构和crispr靶点;
29.b为osckx3基因的突变位点;
30.c为osckx3过表达材料中osckx3基因表达水平。
31.图2是本发明提供的野生型(wt)、osckx3突变体和osckx3过表达材料生长10天(a) 和90天(b)的表型;c、d分别是a图和b图叶夹角统计数据。
32.图3是本发明实施例提供的野生型(wt)osckx3突变体和osckx3过表达材料生叶枕近轴端和远轴端表型;
33.图3中:
34.a~c:wt(a)、osckx3突变体(b)和osckx3过表达材料(c)叶枕切片结构;
35.d~f:wt(d)、osckx3突变体(e)和osckx3过表达材料(f)叶枕近轴端结构;
36.g~i:wt(g)、osckx3突变体(h)和osckx3过表达材料(i)叶枕远轴端结构;
37.j为野生型(wt,左)、osckx3突变体(中)和osckx3过表达材料(右)叶枕d1和d2 长度;
38.k为野生型(wt,左)、osckx3突变体(中)和osckx3过表达材料(右)叶枕d1和d2 薄壁细胞层数;
39.l为野生型(wt,左)、osckx3突变体(中)和osckx3过表达材料(右)叶枕维管束数目。
40.图4是osckx3基因的表达模式和亚细胞定位;
41.图4中:
42.a为osckx3基因在根、茎、叶枕和叶片中的表达量;
43.b~e:osckx3启动子驱动gus报告基因载体转基因水稻7天幼苗(b)、叶枕切片(c)、叶片(d)和节(e)的gus染色图;
44.f为osckx3的激素诱导表达模式;
45.g为osckx3的亚细胞定位;
46.iaa:3-indoleacetic acid,吲哚乙酸;bl,brassinolide,油菜素内酯;cz,cis-zeatin,顺式玉米素;tz,trans-zeatin,反式玉米素;ip,isopentenyladenine,异戊二烯腺
10min。获得osckx3 cds片段(扩增产物大小为1584bp,为seq id no:1的序列)。扩增产物经过琼脂糖凝胶回收试剂盒回收该片段,利用一步克隆法连接到pcr8载体,将构建好的载体利用lr反应构建到pmdc43载体,得osckx3-gfp载体。
88.2.osckx3过表达材料获得
89.(1)构建osckx3过表达材料
90.获得osckx3-gfp载体后由武汉伯远生物科技有限公司负责转化日本晴材料,获得 osckx3过表达材料。
91.(2)osckx3过表达材料osckx3表达量分析
92.提取osckx3过表达材料rna并反转,利用以下引物检测osckx3表达量,图1c结果显示, osckx3在过表达材料中高量表达。
93.用以下引物做osckx3基因的表达分析:
94.引物f:5
’‑
aggcccttgatggcattgtag-3’95.引物r:5
’‑
ccaccacccacatcagcataa-3’96.所用内参基因ubiquitin5引物如下:
97.引物f:5
’‑
gcacaagcacaagaaggtga-3’98.引物r:5
’‑
ccaaagaacaggagcctacg-3’。
99.图1c中:nip代表日本晴野生型,oe1~oe2代表osckx3过表达材料的2个株系。
100.实施例3、水稻突变体和过表达材料的表型鉴定
101.分别观察野生型日本晴、osckx3敲除突变体材料和osckx3过表达材料的叶夹角。播种后正常生长10天至三叶一心期,对第一叶叶枕部位拍照,使用软件image j对不完全叶的叶夹角进行测量,以叶枕为交点,叶片为一边线,叶鞘为一边线。其发育表型如图2所示:可以看到osckx3突变体材料对比野生型有显著的叶夹角减小的表型,过表达材料有显著的叶夹角增大的表型。表明osckx3可以改变水稻的叶夹角。
102.叶枕组织切片(图3a~i)观察发现突变体内叶枕近轴端边缘至维管束的距离(d1)略短,叶枕远轴端边缘至维管束的距离(d2)显著变长,而过表达材料d1和d2都变短(图3d~j)。突变体远轴端细胞层数和微管数量显著增加(图3k~l);结果表明osckx3通过调节叶枕长度和远轴端长度调控叶夹角的大小。
103.观察正常生长120d至成熟期的日本晴、osckx3突变体和osckx3过表达材料,农艺性状统计结果见图6。osckx3突变体籽粒变大且其他性状无显著性差异,表明osckx3突变体具有可以减小叶夹角和增大籽粒的有益性状,但不影响其它性状,这不同于许多叶夹角变小的突变体会影响产量性状。表明osckx3可以作为一个潜在的提高水稻产量的新基因资源。
104.实施例4、osckx3基因表达分析
105.1.pro
osckx3
::gus载体的构建
106.以日本晴水稻基因组dna为模板,采用引物对:
107.osckx3 pro-f(5
’‑
gcaggctccgaattcgtccattagcaccaccaaat-3’)和osckx3 pro-r(5
’‑
aagctgggtcgaattcggtgttggagtagtgattct-3’)进行pcr扩增。pcr扩增体系为总体积为50μl,模板为基因组dna1μl(约50ng)、2
×
kod酶反应缓冲液 25μl、2mm dntp 10μl、10um引物4μl(每条引物均为2μl)、kod酶1μl,加ddh2o(无菌去离子水)至50μl。
108.pcr程序为预变性95℃,5min,进入pcr循环,参数为变性95℃,30s,退火55℃, 30s,延伸72℃,30s,共35个循环。获得osckx3启动子序列(序列为序列表中的seqidno.6),扩增产物经过琼脂糖凝胶回收试剂盒回收该片段,利用一步克隆法连接到 pcr8载体,将构建好的载体利用lr反应构建到pmdc163载体,得pro
osckx3
::gus载体。然后按照常规方法进行遗传转化(委托武汉伯远生物科技有限公司),获得pro
osckx3
::gus 转化株植物。
109.pro
osckx3
的序列如seq id no:6。
110.2.osckx3基因表达分析
111.将步骤1获得的pro
osckx3
::gus转化株植物的幼苗、成熟叶片、节、叶枕等组织进行取样通过gus染色法对目的基因的表达情况进行分析。将转基因植株的样品放入盛有90%丙酮的预冷的玻璃瓶中,室温放置30min。用冰水清洗2次后,将样品转移到gus染色液中,37℃染色4~20h。染色完成的样品放入乙醇:乙酸=3:1的脱色液中进行脱色,时间2~12h。脱色完成后使用数码相机或者显微镜对染色结果进行拍照记录。叶枕切片观察的样品需要使用faa固定液进行固定,具体成分为1ml 38%甲醛(终浓度2%),1ml冰醋酸,18ml 50%的酒精,之后通过树脂包埋。利用半薄切片机获得7μm厚的切片用于观察叶枕组织。osckx3表达分析按照takara公司sybr green的操作说明以不同组织为模板、以osckx3和内参基因ubiquitin5 的定量引物扩增目的基因,检测osckx3在不同组织的表达量,图4a~e结果显示osckx3在叶枕中高量表达。图4f结果显示osckx3基因受bl、tz和ip激素的诱导。
112.用以下引物做osckx3基因的表达分析:
113.引物f:5
’‑
aggcccttgatggcattgtag-3’114.引物r:5
’‑
ccaccacccacatcagcataa-3’115.所用内参基因ubiquitin5引物如下:
116.引物f:5
’‑
gcacaagcacaagaaggtga-3’117.引物r:5
’‑
ccaaagaacaggagcctacg-3’。
118.实施例5、osckx3亚细胞定位
119.实施例2所得的osckx3-gfp载体转化至农杆菌gv3101中,在lb固体培养基上进行划线,培养1~2天后选取单菌落进行实验;挑取单克隆于5ml含有相应抗生素的lb液体培养中,28℃,200~220rpm震荡培养1天;将农杆菌转接到20ml含有卡那抗生素的lb液体培养基中,使od
600
达到0.3左右(20ml培养基中加入1.5ml菌液),28℃震荡培养;检测菌液od
600
的值,数值达到1.5时即可;提前配置好重悬液(灭菌后的ddh2o,0.01m mgcl2, 0.01m mes及100μm as),6000rpm,5min集菌;先用2ml重悬液重悬菌液,随后加入 6~8ml重悬液,6000rpm,离心5min;倒出上清液,2ml重悬液重悬菌液,最终加入15ml 重悬液;28℃暗培养2~3h后注射烟草;将含有不同菌液的侵染液按照1:1:1的比例在5ml 离心管中进行混合,摇匀,随后装入1ml注射器内,用拇指按压注射器将液体从叶片下表皮注射到烟草叶片内(勿使用子叶)。注射后,烟草叶片会出现湿润的现象,尽量减少注射孔的数目;烟草培养3天后观察,结果如图4g所示,osckx3-gfp(左一)和内质网maker 荧光(左二)相融合(左四),明场为左三,表明osckx3定位于内质网中。
120.实施例6、osckx3生化活性检测
121.1.osckx3体外活性检测
122.(1)osckx3-mbp载体构建
123.利用以下引物扩增osckx3并连接到pmal载体上,转化到大肠杆菌bl21(de3 invitrogen)中,加入0.5mm iptg诱导表达,并利用amylose resin(new england biolabs inc.) 纯化蛋白目的蛋白,从而获得mbp-osckx3融合蛋白。
124.构建pmal-c2x载体的osckx3扩增引物:
125.osckx3-pmal f:gaaggatttcagaattc atggaggttgccatggtctg
126.osckx3-pmal r:ctagaggatccgaattc tcagctatagcttgcaaatg
127.(2)活性检测
128.每个反应(100μl)中加入14μg mbp-osckx3融合蛋白、0.1mm细胞分裂素底物(ip 和tz)、0.5mm 2,6-二氯酚吲哚酚(dcip)、75mm tris/hcl buffer(ph 8.5)37℃水浴30min。后立即加入三氯乙酸终止反应。4℃,12000g离心10min;上清液过滤后利用hplc定量检测底物减少量。图5a和5b结果表明osckx3可以催化tz和ip。
129.2.体内活性检测
130.(1)ck的提取
131.取50mg样品放入2ml离心管中(eppendorf)并加入钢珠,迅速放入液氮中,然后使用碾磨机(tissuelyser-48,上海精鑫实验技术有限公司)粉碎样品,研磨至粉末后加入1.6ml 80%的甲醇以及相应的内标,内标具体添加量为50pg[2h5]tz,[2h5]tzr,[2h6]ip, [2h6]ipr。将装有混合液的离心管在4℃条件下置于旋转仪上旋转混匀2小时,4℃,15000g,离心10分钟。小心吸取上清液移至新离心管中,氮气吹干。剩余的沉淀物加入0.6ml 80%甲醇溶液重新混匀,重复上述操作,最后将上清液补加至2ml离心管中,氮气吹干,加入300μl 30%甲醇溶液进行重溶,使用0.22μm水相过滤膜过滤溶液备用。
[0132]
(2)使用gc-ms系统测定ck含量
[0133]
ck提取液通过装备acquity uplc beh c18型分离柱组件的exionlc(ab sciex) 液相色谱系统进行分离。色谱柱首先在40℃条件下进行平衡,之后添加30μl的样品用于后续分析。在针对ck的相关分析中,流动相成分a为水,b为甲醇,采用了多步线性梯度洗脱法对其进行洗脱,具体步骤为:0~2.5min:5%a;2.5~3min:5~20%b;3~12.5min: 20~50%b;12.5~13min:50~100%b;13~15min:100%b;15~15.2min:100~5%b; 15.2~18min:5%b。流速设置为0.3ml min-1
。
[0134]
ck分析通过qtrap 5500型三重四极杆质谱系统(ab sciex)的多反应检测扫描模式(mrm mode)进行。根据相关资料的参考选择正确的离子对激素含量进行定量分析。 mrm的最优条件如下:气帘气气压:40psi,离子喷雾电压:正离子模式5000v,负离子模式-4500v;涡轮加热器温度:600℃;雾化气气压(gas 1):60psi;加热气气压(gas 2):60psi。analyst(版本1.6.3ab scoex)软件用于控制相关仪器以及获取结果数据。得到的原始数据之后被导入multiquant软件(版本3.0.2ab sciex)中进行进一步的分析和处理,ck 的结果通过内标参照进行精确定量。图5c表明osckx3在体内可以催化tz和ip,突变后水稻体内tz和ip含量增加,造成水稻叶夹角变小;而过表达后水稻体内tz和ip含量减少,造成水稻叶夹角变大。
[0135]
实施例7、农艺性状分析
[0136]
将日本晴野生型、osckx3基因突变体和过表达材料种在田中,生长120天后对材料进行拍照和农艺性状统计分析,所有农艺性状包括株高、穗粒数、单株分蘖数、千粒重、粒长
和粒宽均以单株数据为基础,利用spss软件进行统计分析。所得结果如图6所示。
[0137]
根据图6,可获得以下总结性结论:osckx3基因突变后水稻叶夹角变小、千粒重增加、结实率略微减低,而其他农艺性状无显著性差异;osckx3基因过表后叶夹角变大、千粒重减少、结实率略微减低、株高变矮、穗粒数减少,其他农艺性状无显著性差异,表明osckx3 基因可以作为育种改良潜在的基因资源。
[0138]
最后,还需要注意的是,以上列举的仅是本发明的若干个具体实施例。显然,本发明不限于以上实施例,还可以有许多变形。本领域的普通技术人员能从本发明公开的内容直接导出或联想到的所有变形,均应认为是本发明的保护范围。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。