一种乙醛脱氢酶基因dkaldh10及其应用
技术领域
1.本发明属于植物基因工程技术领域,具体涉及一种乙醛脱氢酶基因 dkaldh10及其应用。
背景技术:
2.我国是柿(diospyros kaki thunb.)原产地,也是世界上柿树栽培历史最悠久、面积最大和产量最多的国家,但传统产区以涩柿为主。柿果中存在特化的单宁细胞,其液胞中积累的原花青素(又称“单宁”,proanthocyanidin,pa)是食用后产生涩感的主要原因。根据果实成熟时能否在树上自然脱涩及其性状遗传特点,现有柿品种分为完全甜柿(pollination-constant&non-astringent,pcna;简称“甜柿”)和非完全甜柿(non-pcna;简称“涩柿”)。涩柿果实成熟后需经脱涩处理才能食用,不仅消耗人力、物力和财力,且脱涩后的果实货架期短;而脱涩不完全的柿果不仅商品性降低,还引起消费者对其诱发胃柿石(gastricphytobezoar)的恐惧。因此,成熟果实可在树上自然脱涩的完全甜柿是目前世界范围内产业发展和遗传改良的重点目标。
3.已有研究表明,中国甜柿自然脱涩是乙醛介导的可溶性单宁的不溶化过程,亦即果实中的挥发性产物乙醛与可溶性单宁结合转变生成不导致涩感的不溶性凝胶类物质(通称“凝固效应”),该过程主要受乙醛代谢途径中的三个酶基因家族控制:丙酮酸脱羧酶(pyruvate decarboxylase,pdc),乙醇脱氢酶(alcoholdehydrogenase,adh)和醛脱氢酶(aldehyde dehydrogenases,aldh)。其中,乙醛脱氢酶(aldh)催化乙醛氧化形成乙酸、co2和h2o。已有研究结果表明,利用乙醇和co2等处理均显著增强adh和pdc的活性,诱导果实乙醛含量升高,促进可溶性单宁向不溶性单宁的转化,最终导致果实脱涩。
4.柿遗传背景复杂,染色体倍性高,尤其是基因组信息和甜涩性状遗传规律的研究相对滞后。乙醛脱氢酶及其相关基因在木本植物中研究较少。此外,柿单宁单体的种类、胞质运输与跨膜转运的选择性,以及在液泡中的聚合方式等导致中国甜柿和日本甜柿具有不同的自然脱涩机理。其中,乙醛代谢的下游关键基因dkaldh在中国甜柿自然脱涩中的效应尚无直接相关证据,还鲜有 dkaldh的基因功能被验证。
技术实现要素:
5.本发明针对现有技术的空白点,根据中国甜柿
‘
鄂柿1号’果实发育的关键时期的果肉转录组数据进行表达差异筛选,克隆鉴定到了一个差异表达基因即乙醛脱氢酶基因dkaldh10,并首次验证其与中国甜柿自然脱涩之间的关系,完善了中国甜柿自然脱涩分子调控网络,为进一步的遗传改良调节柿脱涩提供了新的科学依据和基因资源。
6.本发明的目的之一在于提供一种乙醛脱氢酶dkaldh10,所述乙醛脱氢酶 dkaldh10的氨基酸序列如seq id no:1所示。
7.进一步的,所述乙醛脱氢酶dkaldh10定位于细胞质膜和液泡膜中。
8.本发明的目的之二在于提供了一种编码上述乙醛脱氢酶dkaldh10的基因,所述基
因为dkaldh10基因,其核苷酸序列如seq id no:2所示。
9.本发明的目的之三在于提供了一种编码上述dkaldh10基因的扩增引物,所述扩增引物的核苷酸序列如seq id no:3-4所示。
10.本发明的目的之四在于提供一种用于检测dkaldh10基因的荧光定量pcr 特异引物,所述荧光定量pcr特异引物的核苷酸序列如seq id no:5-6所示。
11.本发明的目的之五在于提供一种包含上述dkaldh10基因的载体。
12.进一步的,所述载体为dkaldh10基因的超表达载体或干涉表达载体。
13.本发明的目的之六在于提供一种遗传工程菌,所述遗传工程菌中包含上述载体。
14.本发明的目的之七在于提供上述dkaldh10基因,或上述载体,或上述遗传工程菌在调节柿脱涩中的应用。
15.进一步的,所述dkaldh10基因,或载体,或遗传工程菌通过调节可溶性单宁和不溶性单宁的含量,调节柿脱涩。
16.与现有技术相比,本发明的有益效果是:
17.本发明通过对中国甜柿
‘
鄂柿1号’果实发育期(花后10周和20周)的转录组数据进行表达差异筛选,首次获得并鉴定一个中国甜柿乙醛脱氢酶基因家族重要成员dkaldh10基因,并获得了dkaldh10基因的全长序列。本发明通过构建dkaldh10基因干涉载体和超表达载体,并测定不同载体转化后果实中可溶性单宁和不溶性单宁的含量,验证了dkaldh10基因是通过分解乙醛,抑制可溶性单宁向不溶性单宁的转化,进而影响了中国甜柿自然脱涩中的“凝固效应”,即dkaldh10基因通过调控乙醛代谢,调节柿果实中可溶性单宁和不溶性单宁的含量,进而调节柿脱涩过程。本发明首次阐明了dkaldh10基因与中国甜柿自然脱涩之间的关系,完善了甜柿自然脱涩分子调控网络,为调节柿脱涩提供了新的科学依据和基因资源。
附图说明
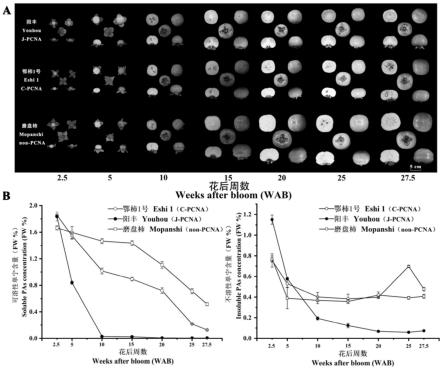
18.图1为本发明实施例1中三个柿品种的果实形态图和单宁含量变化图,其中图1-a为中国甜柿
‘
鄂柿1号’,日本甜柿
‘
阳丰’,非完全甜柿
‘
磨盘柿’花后2.5-27.5周的果实形态图,图1-b为三种柿的可溶性单宁和不溶性单宁含量动态变化;
19.图2为本发明实施例2中差异表达基因dkaldh10基因在
‘
鄂柿1号’花后2.5-27.5周的表达量变化趋势;
20.图3为本发明实施例2中不同物种的aldh家族中的聚类分析结果;
21.图4为本发明实施例2中dkaldh10在烟草叶片中的亚细胞定位;
22.图5为本发明实施例3中柿果实圆片瞬时干涉表达dkaldh10后的结果,其中图5-a为果实圆片的dmaca染色结果,图5-b为可溶性单宁含量变化,图5-c为不溶性单宁含量变化;
23.图6为本发明实施例3中利用
‘
恭城水柿’组培苗构建dkaldh10超表达株系或干涉表达株系后,不同株系中的可溶性单宁和不溶性单宁含量分析结果,其中图6-a为dkaldh10超表达株系中可溶性单宁和不溶性单宁含量,图6-b 为dkaldh10干涉表达株系中可溶性单宁和不溶性单宁含量。
具体实施方式
24.下面将结合本发明中的实施例,对本发明的技术方案进行清楚、完整地描述,显然,所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动条件下所获得的所有其它实施例,都属于本发明保护的范围。
25.实施例1不同柿品种中单宁含量测定
26.选取三个柿品种:中国甜柿
‘
鄂柿1号’(c-pcna),日本甜柿
‘
阳丰
’ꢀ
(j-pcna)和非完全甜柿
‘
磨盘柿’(non-pcna)。于花后2.5周、5周、10 周、15周、20周、25周和27.5周,分别取样观察三个柿品种的果实形态图,并采用folin-ciocalteu法定量测定不同柿品种中的可溶性单宁和不溶性单宁含量,分析单宁在果实发育过程中的积累模式。结果如图1所示。
27.folin-ciocalteu法具体为:
28.(1)单宁酸标准曲线制作:分别取0ml、0.2ml、0.4ml、0.6ml、0.8ml、 1.0ml的标准单宁酸溶液于刻度试管中,向每管中分别加入7.5ml蒸馏水,0.5 ml酚类试剂,3min后,加入1.0ml饱和碳酸钠,最后用水稀释到10.0ml,1 h后在725nm下比色测出样品的吸光度。
29.(2)单宁含量测定:从果实赤道取0.1g果肉,切碎放入研钵,加入0.6ml 80%甲醇溶液充分研磨后转入2ml离心管中,5000g离心10min,取上清液至 2ml离心管中,再用0.4ml 80%甲醇溶液清洗残渣,重复以上步骤,最后定容于1ml离心管,然后用蒸馏水将其稀释10倍,测定时取1.0ml样品,加入7.5 ml蒸馏水,0.5ml酚类试剂,3min以后,加入1.0ml饱和碳酸钠,反应1h 后用分光光度计在725nm下,测定吸光度,由单宁酸标准曲线上查出对应的可溶性单宁含量。
30.将测定完可溶性单宁含量后的柿果残渣用于不溶性单宁含量的测定,具体为:用0.6ml 1%盐酸-甲醇溶液溶解,于室温放置30min,5000g离心10min,取上清液,再用0.4ml 1%盐酸-甲醇溶液清洗残渣,重复以上步骤,最后取上清液定容至1ml离心管,后续测定步骤同可溶性单宁。
31.其中图1-a为三个柿品种的果实形态观察图,图1-b为三个柿品种的单宁含量变化。根据图1-b结果,三个品种的可溶性单宁含量随着果实的发育均呈下降趋势,其中
‘
阳丰’(j-pcna)在果实花后2.5周可溶性单宁浓度为1.85% fw,在花后5周时其可溶性单宁浓度迅速减少到0.83%fw,但在花后15周其下降趋势有所减缓,此时可溶性单宁浓度已经降到0.2%fw以下并一直保持极低水平,结合品尝实验,
‘
阳丰’已经没有涩味,与先前报道结果相符。
‘
鄂柿1号’(c-pcna)在花后25周时,果肉可溶性单宁含量下降到极低水平(0.21% fw),可以直接食用,而磨盘柿(non-pcna)果实的可溶性单宁含量始终高于两种完全甜柿,直到花后25周,果实仍有明显涩感,不能直接食用。而与 j-pcna和non-pcna相比较,c-pcna的不溶性单宁含量在果实自然发育后期 (花后20-25周)有呈上升趋势,上述结果表明c-pcna的自然脱涩过程与可溶性单宁向不溶性单宁转化有关。
32.实施例2 dkaldh10表达分析、基因克隆和序列分析
33.1、表达分析
34.根据实施例1的分析结果,中国甜柿
‘
鄂柿1号’在果实发育期存在可溶性单宁向不溶性单宁转化的现象,因此对
‘
鄂柿1号’花后10周和20周的转录组数据进行表达差异筛选,鉴定到了一个差异表达基因dkaldh10,对该差异表达基因进行qrt-pcr表达分析。
35.(1)rna提取和cdna合成
36.于花后不同时期,分别取
‘
鄂柿1号’果实,采用rna plant plus(tiangen) 试剂盒,按说明书操作提取总rna。利用1%凝胶电泳来检测rna的纯度和完整性,通过超微量分光光度计测定其在260nm处吸光值进行定量。然后采用 primescript
tm rt reagent kit with gdna eraser(takara)试剂盒,按试剂盒说明书合成cdna。
37.(2)qrt-pcr表达分析
38.根据该差异表达基因,设计其相应的荧光定量pcr特异性引物,具体为:
39.f:ccgattccttctcgtatgct(如seq id no:5所示);
40.r:actttgctcgatgtgctcca(如seq id no:6所示).
41.并以柿(d.kaki thunb.)actin(引物f:catggagaaaatctggcatcatac; r:gaagcactgggtgctcttctg)作为qrt-pcr的内参基因,采用 green realtime pcr master mix荧光染料(toyobo,japan),按说明书操作并在quantstudio 7flex实时定量pcr仪(applied biosystems)上进行荧光定量 pcr分析,反应程序为95℃30s,接着95℃5s,58℃10s,72℃15s,运行 45个循环,然后95℃60s,40℃30s,每个样品进行4次机械重复。
42.该差异表达基因的表达量变化如图2所示。dkaldh10基因在果实花后5-15 周呈现上调表达的趋势,而在花后25-27.5周的表达呈现下调表达的趋势,这与实施例1中测定的
‘
鄂柿1号’不溶性单宁含量的变化趋势一致。说明该基因与
‘
鄂柿1号’中可溶性单宁向不溶性单宁的转化,即与中国甜柿
‘
鄂柿1号’的自然脱涩相关。
43.2、基因克隆和序列分析
44.栽培柿品种遗传背景复杂且染色体倍性高,童期长,且目前没有栽培柿的基因组序列信息,因此获得dkaldh10基因的全长序列较为困难。本发明通过常规的3
’5’‑
race方法获得该差异表达基因的unigene序列全长后,设计特异性引物:
45.f:atggggaaagggcgagagagg(如seq id no:3所示);
46.r;aagctttggcttttcaggaga(如seq id no:4所示),
47.以
‘
鄂柿1号’cdna为模板进行pcr扩增获得该基因的全长序列,其中pcr程序为94℃3min,接着94℃30s,58℃30s,72℃80s,运行35个循环,最后进行72℃5min的延伸。扩增产物通过凝胶电泳分离,切胶回收。将该基因克隆到peasytm-blunt simple cloning vector(全式金,中国北京)克隆载体中,获得peasy-dkaldh10重组质粒并进行测序。
48.根据测序结果,采用orffinder在线软件对其全长序列以及对应的氨基酸序列进行分析,并利用mega7.0(邻接法,bootstrap value=2000)对dkaldh10 和相关蛋白进行聚类分析。聚类分析结果如图3所示,该基因聚类于乙醛脱氢酶aldh第10亚家族。将该基因命名为dkaldh10基团,该基因的核苷酸序列如seq id no:2所示,该基因全长1566bp,其对应的蛋白质dkaldh10的氨基酸序列如seq id no:1所示,预测编码521个氨基酸。
49.3、亚细胞定位
50.设计包含限制性酶切位点bamh i和kpn i的引物,引物序列具体为:
51.f:cgcggatccatggggaaagggcgagagagg(如seq id no:7所示);
52.r:cggggtacc aagctttggcttttcaggaga(如seq id no:8所示)
53.利用pcr扩增获得的dkaldh10的全长序列,pcr程序为94℃3min,94℃ 30s,58℃30s,72℃80s,35个循环,72℃额外延伸5min。pcr产物通过凝胶电泳回收,通过双酶切法连
接到yfp载体,构建35s:dkaldh10:yfp融合质粒,通过测序确认序列准确性后,利用热激法将重组质粒及对照转入农杆菌 gv3101。
54.将含有目的基因的农杆菌gv3101单克隆加入1ml含相应抗生素的lb液体培养基中,28℃培养1d;然后加入到50ml含相应抗生素的lb液体培养基中,28℃,250r/min过夜培养至od
600
为1.6-1.8或菌液呈现金黄色即可;25℃ 4000
×
g离心10min收集菌体;用渗透液重悬浮至菌液浓度od
600
为0.75,渗透液为无菌双蒸水含10mm mgcl2,10mm mes,ph 5.6和150μm乙酰丁香酮;注射侵染前,将重悬浮的菌液于28℃,100r/min条件与培养3h,使参与 t-dna转移的基因进行预表达,以提高农杆菌的侵染力。
55.利用注射渗透法注入6周苗龄烟草叶片,转化后的烟草培养2-3天后,共聚焦显微镜观察yfp的瞬时表达,观察结果如图4所示,表明dkaldh10定位于细胞质膜和液泡膜上。
56.实施例3柿果圆片瞬时转化和叶盘法稳定遗传转化
57.1、柿果圆片瞬时转化
58.柿属于较难进行遗传转化的木本植物之一,存在基因型差异大、转化率低、再生困难等难题,使得克隆并验证dkaldh10基因的功能十分困难。本发明则利用柿果实圆片作为转化对象,规避了温度、环境以及组织差异性因素的干扰,构建得到了稳定遗传转化株系。
59.柿果圆片瞬时转化的实验过程具体如下:
60.采用gateway技术,分别通过bp重组反应将dkaldh10全长序列和150-300 bp的5’端序列导入pdonrtm222中间载体和pmdc32表达载体或 ph7gwiwg1干涉载体,重组质粒通过测序验证准确性后利用热激法导入 gv3101农杆菌。
61.其中,扩增dkaldh10全长序列的引物为:
62.f:
[0063]5’‑
ggggacaactttgtacaaaaaagttggaatggggaaagggcgagaga gg-3’;
[0064]
r:
[0065]5’‑
ggcggccgcacaactttgtacaagaaagttgggtaaagctttggctttt caggaga-3’;
[0066]
扩增150-300bp的5’端序列所采用的引物为:
[0067]
f:
[0068]5’‑
ggggacaactttgtacaaaaaagttggatgctgttcatcgacggcgagt
ꢀ‑3’
;
[0069]
r:
[0070]5’‑
ggcggccgcacaactttgtacaagaaagttgggtacagcaaggtctgc ataatact-3’。
[0071]
农杆菌侵染液制备方法同上。
[0072]
将采好的
‘
鄂柿1号’柿果实用流水清洗,放入灭菌烧杯中,加入75%无水乙醇消毒,然后用1%的次氯酸钠消毒,最后用无菌双蒸水清洗;将消毒完成的柿果实置于灭菌干燥滤纸上,用已灭菌的水果刀将其切成厚度约0.2cm的薄片;用已灭菌的直径为1.0cm的打孔器在切好的薄片上均匀打成圆片,置于干净的灭菌干燥的滤纸上;将打好的柿果实圆片放入已制备好的渗透悬浮液中,在振荡器上轻微震荡一段时间;将侵染完成的柿果实圆片取出置于干燥无菌滤纸上,以吸干圆片表面菌液,均匀置于铺有滤纸的ms固体培养基的培养皿上;将培养皿置于25℃暗培养3d。
[0073]
分别取空载体和连接有dkaldh10基因的干涉载体(phg-dkaldh10),转入
‘
鄂柿1号’果实圆片中,每一天取15片圆片进行分析。通过dmaca染色法对
‘
鄂柿1号’的圆片进行染
色,其中dmaca染色法具体步骤如下:将果实圆片置于30%的冰醋酸乙醇溶液中浸泡12-20h以确保完全去除花青苷和叶绿素等色素,然后用75%的乙醇漂洗12h,之后用蒸馏水冲洗10s,最后在0.6% dmaca溶液(6n hcl:甲醇=1:1)中染色2min,观察和拍照记录,染色颜色越深表示单宁含量越高。采用folin-ciocalteu法定量测定果实圆片中可溶性单宁和不溶性单宁的含量。测定结果如图5所示。
[0074]
其中图5-a为dmaca染色结果,图5-b为可溶性单宁含量,图5-c为不溶性单宁含量。结果显示,与空载体的对照组相比,dkaldh10基因的干涉载体(phg-dkaldh10)染色后颜色明显较浅,即干涉载体会影响果实圆片中的总单宁含量。根据定量检测结果可知,干涉载体phg-dkaldh10表达后果实圆片中可溶性单宁含量降低,不溶性单宁含量升高,即表明dkaldh10可通过分解乙醛,抑制可溶性单宁向不溶性单宁转化,进而影响中国甜柿自然脱涩中的凝固效应。
[0075]
2、叶盘法稳定遗传转化
[0076]
农杆菌侵染液制备方法同上。
[0077]
从继代培养一个月,长势良好的
‘
恭城水柿’组培苗中切下叶片,切成边长为1.0cm的方形叶块,将切割好的叶片叶背朝下,放置于垫有滤纸的愈伤组织预培养基上,暗培养3d;将预培养后的叶片放入活化完成的菌液,缓慢震荡数分钟后,取出叶片于无菌滤纸上,除去叶片表面过量的菌液;将叶片放置到置有滤纸的共培养培养基上,在25℃的黑暗条件下共培养3天;用含有cef(400 mg/l)的无菌水清洗叶片,再用无菌水清洗,将叶片置于无菌滤纸上吸干表面的水份;将叶片转移至愈伤诱导培养基中,25℃黑暗条件下培养1个月;切下叶片产生的愈伤组织,继代到不定芽形成培养基中,25℃光照条件下以诱导不定芽。通过对愈伤组织使用相同培养基的继代培养,可以保持它多年形成不定芽的能力。
[0078]
分别构建获得dkaldh10超表达以及干涉稳定表达株系,并分别定量测定不同株系中可溶性单宁和不溶性单宁的含量,检测结果如图6所示。其中图6-a 为dkaldh10超表达株系中可溶性和不溶性单宁含量,图6-b为dkaldh10干涉表达株系中可溶性和不溶性单宁含量。结果显示,dkaldh10超表达后,植株#20、#21、#23中可溶性单宁含量升高,不溶性单宁含量降低,dkaldh10干涉表达后,植株#1、#3、#11中可溶性单宁含量显著降低,不溶性单宁含量显著升高,该结果也进一步证实了,dkaldh10通过调控乙醛代谢,调节了植株的可溶性单宁和不溶性单宁含量,进而参与脱涩过程的调控。本发明阐明了 dkaldh10基因与中国甜柿果实自然脱涩之间的关系,完善了中国甜柿自然脱涩分子调控网络,为进一步的遗传改良提供了新的科学依据和基因资源。
[0079]
以上所述,仅为本发明较佳的具体实施方式,但本发明的保护范围并不局限于此,任何熟悉本技术领域的技术人员在本发明揭露的技术范围内,可轻易想到的变化或替换,都应涵盖在本发明的保护范围之内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。