1.本发明涉及一种林木四倍体的诱导方法,尤其涉及一种诱导合子染色体加倍创制桉树四倍体植株的诱导方法,属于植物遗传育种领域。
背景技术:
2.桉树(eucalyptus spp.)是世界性的重要用材林树种,每年为我国工业生产提供了超过40%的木材和木浆产量。目前全球以杂交育种为主的桉树遗传改良研究陷入瓶颈期,多年未见突破性品种诞生。为创制桉树高产优质新种质,促使生物产量进一步提高,急需创新桉树育种技术体系。近年来,人工诱导四倍体已成功应用于杨树遗传改良,这为桉树种质创新指明了方向。四倍体是指细胞内包含四套染色体的生物体。与普通二倍体相比,四倍体植物通常植株粗壮,花、叶和果实等器官显著增大,对生物和非生物逆境的抗性更强。同时,四倍体种质还可作为亲本,直接与二倍体杂交获得生长优势更明显的三倍体。因此,人工诱导桉树四倍体对选育桉树多倍体优良种质、大幅度提高桉树人工林生产力意义重大。
3.人工诱导获得植物四倍体通常有两种途径,其一是在实验室内采用组织培养的方法,使用秋水仙碱等细胞分裂抑制剂处理二倍体植物的分生组织等,诱发植物体细胞染色体加倍获得四倍性体细胞组织,从而获得四倍体再生植株。这种方法在桉树上已有报道(韩超等.秋水仙素诱导巨桉无性系eg5多倍体的研究.中国农学通报,2010,26(24):128-132;韩超等.秋水仙素诱导尾巨桉多倍体的研究.中国农学通报,2010,26(15):149-153;韩超.秋水仙素诱导桉树染色体加倍技术体系的研究.中国林业科学研究院,2010;谭德冠等.刚果12号桉离体组织的多倍体诱导.热带作物学报,2005(02):50-54;胡洲鹤等.秋水仙素诱导桉树多倍体的初步研究.广西林业科学,2004(04):195-196 203:谭德冠.刚果12号桉(eucalyptus 12abl)组织培养及多倍体诱导的研究.华南热带农业大学,2004;kapooi等.试验合成了桉树异源四倍体.湖南林业科技,1986(04):46-48),但也存在一些问题。比如针对分生组织多细胞施加诱变处理常会得到大量混倍体,再生植株倍性复杂且难以分离;依托组培再生体系的诱变处理方法往往仅针对个别基因型有效,一次有效的诱变处理仅能获得一份多倍体种质,诱导效率不高等。
4.人工诱导获得植物四倍体的另一途径是直接诱导植物合子染色体加倍,即采用一定的技术方法,在受精后合子细胞发育的特定时机施加诱变处理,诱发合子细胞染色体加倍获得四倍体植株。相较于已有报道的体细胞染色体加倍,诱导合子染色体加倍具有以下三个优势:一是诱导获得的四倍体是由单个合子细胞直接发育而来的整倍性的四倍体植株,不会产生混倍体;二是诱变过程不依赖特定实验室环境,可在野外非离体树上直接实施,适用范围广;三是一次诱变处理可作用于多个果序及子房内的多个受精合子,同时获得多个四倍体新种质,诱导效率较高。
5.目前,人工诱导植物合子染色体加倍主要采用秋水仙碱溶液、n2o气体等化学药剂,或极端温度处理等物理手段阻断合子细胞正常分裂过程,促使合子细胞染色体加倍获
得四倍体。其中又以高温处理诱导杨树合子染色体加倍效果最佳(王君等.理化处理诱导合子染色体加倍选育青杨派杂种四倍体.北京林业大学学报,2010,32(05):63-66;石乐等.高温处理诱导染色体加倍获得白杨杂种多倍体.核农学报,2012,26(08):1118-1123;鲁敏.响叶杨三倍体和四倍体诱导技术研究.北京林业大学,2013)。但是,现有高温诱导植物四倍体技术主要针对杨树等早春低温时开花、花期短且花芽发育状态易辨认区分、坐果期短可离体切枝室内水培收获种子的北方适生树种。而以桉树为代表的南方适生树种,通常在夏季湿热时开花,适生环境要求使用更高的热激处理强度,繁茂的枝叶易受热损伤导致落果;花期长达数月且花蕾及果序形态变化不明显,难以通过经验或简单试验在长达数月的果序发育期内确定施加热激诱变处理的有效时机;坐果期超过半年无法采用离体水培收获种子,只能在野外复杂环境条件实施非离体诱变操作,更加放大了诱变处理时机的选择难度,并对诱变操作方法提出了更高的要求。因此,已有报道的方法对于以桉树为代表的南方适生树种并不适用,主要缺陷在于:无法解决野外非离体诱导合子染色体加倍处理时机的选择问题,没有有效方法判明植物合子发育阶段,使得诱变处理难以准确施加于合子第一次有丝分裂期附近造成加倍失败;无法解决野外非离体条件下对树体进行高温诱变处理的操作方法问题,导致树体枝叶易受到热激损伤;无法解决在夏季野外施加高温处理后保全花蕾及果序活性的问题,导致施加诱变后树体枝叶果序枯死掉落等。
技术实现要素:
6.本发明针对现有夏季湿热时开花、对高温不敏感、花期长且花蕾、果序形态变化不明确、坐果期长的林木在野外诱变四倍体过程中存在的诱导处理时期、诱导处理条件难以确定,导致四倍体得率差的技术问题,提供一种野外非离体诱导林木四倍体的方法,本发明方法对野外生长的林木在非离体条件下进行热激处理,诱导合子染色体加倍,获得四倍体植株。本发明方法克服了已有报道方法无法适用于桉树等南方树木四倍体诱导的难题。
7.本发明提出的技术方案解决了以下难题:明确了野外非离体热激处理诱导桉树合子染色体加倍获得四倍体的有效处理时机和处理条件;建立了一种能够在夏季有效保全桉树花蕾及果序活力的野外非离体热激处理诱导桉树四倍体的方法。
8.为实现本发明的目的,本发明一方面提供一种野外非离体诱导林木四倍体的方法,包括在非离体状态下对林木授粉后的果序进行热激处理。
9.其中,所述林木选择为桉树、相思树或柚木,优选为桉树。
10.特别是,所述桉树选择为尾叶桉。
11.其中,所述热激处理为对授粉后的果序着生花枝进行加热处理,即对授粉后的果序进行加热处理。
12.特别是,所述热激处理的温度为40-48℃;热激处理时间为3-6h。
13.尤其是,所述热激处理温度优选为44
±
1℃;热激处理时间优选为6h。
14.其中,林木授粉后在果序上花丝和花柱脱落阶段或果序开始膨大阶段,进行热激处理。
15.特别是,优选为在果序上花丝和花柱脱落阶段,进行热激处理。
16.本发明另一方面,提供一种野外非离体诱导林木四倍体的方法,包括如下顺序进行的步骤:
17.1)对林木花枝上花蕾的蒴盖由绿色变为黄色或白色,且花蕾蒴盖沿脱落环处出现松动时进行授粉;
18.2)对授粉后花枝上的果序进行观察,对处于果序上的花丝和花柱脱落阶段、果序开始膨大阶段的林木花枝进行加热,即对授粉后的处于花丝和花柱脱落阶段、果序开始膨大阶段的果序进行热激诱变处理;
19.3)收集热激诱变处理后成熟种子,并进行播种,育苗;
20.4)首先采用流式细胞仪对幼苗倍性水平进行初步鉴定;接着对经流式细胞仪初步鉴定出的四倍体植株,通过体细胞染色体计数进行二次鉴定,确定植株的倍性水平,获得四倍体植株。
21.其中,所述林木选择为桉树、相思树或柚木,优选为桉树。
22.特别是,所述桉树选择为尾叶桉。
23.其中,所述步骤1)中所述授粉为在将林木花枝上的花蕾蒴盖顶部连同柱头的顶端一起用刀片切除,接着在切口显露的花柱创口处点授花粉。
24.在切口显露的花柱创口处点授花粉,不经去雄而一次性完成授粉的方法。
25.特别是,在授粉后,对授粉花枝套装隔离纸袋,起隔绝花粉污染的作用。隔离纸袋于授粉两周后解除。
26.尤其是,所述授粉时所用花粉为提前收集的其它优良桉树花粉。
27.特别是,自完成授粉时起,间隔24小时连续采集授粉后果序,观察果序外部形态变化并拍照记录,并根据授粉后果序发育时间和外部形态变化将授粉后果序发育过程分为5个阶段;
28.第1阶段为授粉后1至6天,果序外部形态为柱头顶部经切割后氧化变黑,黄色花丝挺立于花柱周围,此阶段合子未受精;
29.第2阶段为授粉后7至9天,果序外部形态为花丝枯萎卷曲但尚未脱落,此阶段为双受精时期;
30.第3阶段为授粉后10至22天,果序外部形态为花丝逐渐脱落,花柱逐渐变黄枯萎,此阶段为休眠合子时期;
31.第4阶段为授粉后23至26天,果序外部形态为花丝完全脱落,花柱完全脱落,此阶段为合子休眠结束至4细胞原胚时期;
32.第5阶段为授粉后27-28天,除果序开始明显膨大外,形态再无其它明显变化;此阶段为合子进一步发育至球形胚时期。
33.所述采集果序为采集授粉后套袋内的果序,每次采集时解除套袋并在采集后立刻套回。
34.其中,步骤2)中对授粉后的处于花丝和花柱脱落阶段的果序进行热激诱变处理。于授粉后合适时机(即果序处于花丝和花柱脱落阶段)对授粉后果序所在的整个花枝进行热激诱变处理(即加热处理)。
35.于授粉后果序发育的第4阶段和第5阶段,即授粉后第23-28天;优选为第4阶段,即授粉后第23-26天,花丝和花柱已完全脱落,合子发育处于合子休眠结束至4细胞原胚时期进行热激处理。
36.特别是,步骤2)中所述热激诱变处理的温度为40-48℃,优选为44
±
1℃;热激诱变
处理时间为3-6h,优选为6h。
37.尤其是,在热激诱变处理之后还包括:对花枝进行后处理,即对花枝上的叶片及果序喷洒预冷至4-20℃的清水或磷酸二氢钾水溶液,直至叶面温度降到10℃以下。
38.特别是,后处理优选使用喷洒预冷至4-20℃的磷酸二氢钾水溶液,直至叶面温度降到10℃以下。
39.尤其是,所述清水或磷酸二氢钾水溶液的温度优选为4℃;后处理喷洒的磷酸二氢钾水溶液的浓度为0.05-0.2%,优选为0.1%。
40.大量喷洒预冷的0.1%磷酸二氢钾水溶液的目的是为叶片和果序快速补水和降温、促进热激后植物组织修复损伤,从而提高果序生活力,提高四倍体得率。
41.尤其是,步骤2中所述热激诱变处理为采用专利号为zl200610113448.x的“树木非离体枝芽加热处理装置”对林木花枝进行加热。
42.特别是,所述热激诱变处理的温度为40-48℃,优选为44
±
1℃;热激诱变处理时间为3-6h,优选为6h。
43.尤其是,采用专利号为zl200610113448.x的“树木非离体枝芽加热处理装置”在非离体条件下,将整个待诱变花枝包裹,然后加热,提高处理装置内部温度,加热包裹在“树木非离体枝芽加热处理装置”内的花枝,对处理装置内部的花枝上着生的果序进行热激诱变处理。
44.本发明的热激诱变处理在树体上直接进行,花枝仍与树体连接,为非离体状态。在实施前处理的操作后,使用专用设备(即专利号为zl200610113448.x,专利名称为“树木非离体枝芽加热处理装置”)对着生有授粉果序的整个花枝进行控温控时加热处理。
45.特别是,采用“树木非离体枝芽加热处理装置”对林木花枝进行加热处理之前,还包括:先对待诱导的花枝上的叶片和果序喷水润湿;接着使用包裹材料包裹着生有果序的整个花枝。
46.前处理的目的是在后续热激处理中为果序和叶片保湿、避免果序与专利号为zl200610113448.x的“树木非离体枝芽加热处理装置”的极高温度电热片直接接触,从而提高果序生活力。
47.尤其是,喷水至有凝聚水珠自叶片流下后,立即使用二氧化硅气凝胶材料包裹着生有果序的整个花枝;;
48.特别是,所述包裹材料选择二氧化硅气凝胶材料、防火隔热布或陶瓷纤维棉,优选为二氧化硅气凝胶材料。
49.尤其是,在花枝进行热激诱变处理后,撤去专利号为zl200610113448.x的“树木非离体枝芽加热处理装置”,以及包裹花枝的二氧化硅气凝胶材料,然后再喷洒所述的清水或磷酸二氢钾水溶液。
50.其中,步骤3)中种子成熟具体时间为授粉后8~10个月,即次年3~4月份前后。
51.其中,步骤4)中待幼苗长至约10cm高度时,进行子代幼苗植株倍性水平检测。
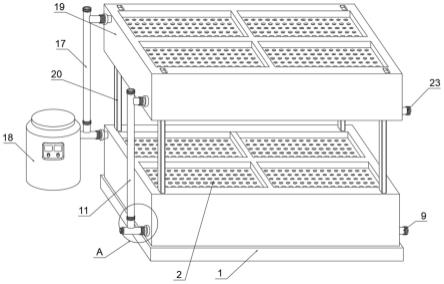
52.本发明的野外非离体热激处理诱导合子染色体加倍获得桉树四倍体的方法,包括:对野外定植的桉树进行控制授粉(一步授粉法对桉树进行人工授粉),经套袋隔离等形成果序;观察记录授粉后果序发育状态,待果序进一步发育,在特定时机对花枝上果序施加非离体热激处理;待果序内种子成熟后进行播种育苗,从中鉴定获得桉树四倍体植株;其
中,对花枝上果序施加非离体热激处理的特定时机为:授粉后果序发育至第4阶段,即授粉后第23-26天、花丝和花柱已完全脱落时;施加热激处理的方法为:处理前采用二氧化硅气凝胶材料包裹花枝上的果序,热激处理后及时对果序降温,其中经44
±
1℃热激处理6小时诱导桉树合子染色体加倍获得四倍体的效率高。
53.综合上述技术方案,本发明所具备的优点及积极效果为:
54.1、本发明方法首次明确了诱导夏季湿热开花、对高温不敏感、花期长且花蕾、果序形态变化不明确、坐果期长的林木(优选为桉树)合子染色体加倍获得四倍体的可行性,获得了桉树四倍体新种质,有效的推动了桉树多倍体育种技术进步。
55.2、本发明方法确定了非离体热激处理诱导桉树合子染色体加倍的最佳处理条件,即果序发育处于第4阶段(即授粉后第23至26天)、花丝和花柱完全脱落时,经44
±
1℃热激处理6小时诱导桉树合子染色体加倍获得四倍体的效率最高;提供了适宜非离体热激诱导桉树合子染色体加倍的完整操作方法,建立了桉树合子染色体加倍选育四倍体的技术体系,为林木育种增添了新途径,推进了南方林木种质创新发展。本发明诱变时机判定更加便捷,利用花丝、柱头的发育变化特征,结合授粉后的时长快速判定合子发育状态,从而确定实施诱变的时机,提高了诱导效率。
56.3、本发明方法旨在诱导桉树受精后合子染色体加倍获得四倍体,诱导过程没有混倍体、嵌合体产生,四倍体诱导效率高;可在特定亲本组合杂交授粉后进行热激诱变处理,一次性获得一批不同基因型的桉树四倍体新种质,有效的提高桉树多倍体诱导效率,诱变效率达到6%以上,加速了桉树良种选育进程。
57.4、本发明方法可推广性好,在热激处理前、后对待诱导花枝进行前处理、后处理,可有效保全花蕾及果序活力,提高四倍体得率,特别适用于夏季开花且坐果时间长的南方阔叶树种非离体诱导四倍体。
58.5、本发明提供的四倍体诱导方法适宜野外操作,破除了试验条件的制约,为不可离体培养的长坐果期林木诱导合子染色体加倍产生四倍体开辟了新途径。
59.6、本发明方法操作简单,诱变操作方法适用性强,可应用于各型立地条件下不同树种的野外非离体诱变育种;而且本发明方法处理成本低,可经一次处理获得多个四倍体新种质,避免嵌合体和混倍体产生等优点,对于推动南方林木种质创新发展具有良好前景和重大意义。
附图说明
60.图1为本发明实施例中人工授粉与花蕾、果序外部形态示意图,其中a为尾叶桉花蕾人工授粉示意图,b至e为不同发育阶段花蕾及果序外部形态示意图;
61.图2为本发明实施例中野外非离体热激处理示意图;
62.图3为本发明实施例中播种育苗示意图;
63.图4为本发明实施例中植株流式细胞仪倍性检测和体细胞染色体计数结果图,其中a和b为流式细胞仪倍性检测结果,a图峰值在坐标轴50左右为二倍体,b图峰值在坐标轴100左右为四倍体;c和d为体细胞染色体计数结果,c图中细胞具22条染色体为二倍体,d图中细胞具44条染色体为四倍体;
64.图5为本发明实施例中尾叶桉二倍体、四倍体植株对比图,其中a为二倍体,b为四
倍体。
具体实施方式
65.以下结合具体实施例子来进一步阐述本发明,本发明的优点和特点将会随着描述而更为清楚。但这些实施例仅是范例性的,并不对本发明的范围构成任何限制。本领域普通技术人员在没有做出创造性劳动前提下所获得的所有其他实施例均属于本发明保护范围。
66.实施例1
67.1、人工授粉:
68.在尾叶桉散粉前选择花量大的花枝作为诱变处理候选花枝;待观察到多数花蕾蒴盖由绿色变为黄色或白色、并且花蕾蒴盖沿脱落环处出现松动、即将脱落时进行人工授粉;
69.授粉方法采用一步授粉法,将花蕾蒴盖顶部连同柱头的顶端一起用刀片切除,在切口显露的花柱创口处用棉签点授花粉;其中:人工授粉所用花粉来自其它优良桉树花粉。授粉完成后对花枝套装隔离袋(醋酸纸袋,如图1a)。
70.图1a中对授粉后花枝套装隔离醋酸纸袋,起隔绝花粉污染的作用,于授粉两周后解除套装的隔离袋。
71.2、果序发育阶段观察:
72.以尾叶桉完成授粉时起,以间隔24小时的采样时间间距,连续采集授粉后果序,观察果序外部形态变化并拍照记录。
73.以尾叶桉完成人工授粉时间为起始点,授粉后花蕾、果序外部形态发育变化示意图(如图1a),其中图1a的a
′
为完成一次性人工授粉后的花蕾。
74.授粉后的花蕾、果序的外部形态观察结果如表1。根据授粉后果序发育时长和外部形态变化将果序发育分为5个阶段(表1)。
75.第1阶段:授粉后1至6天,柱头顶部经切割后氧化变黑,黄色花丝挺立于花柱周围(图1b);此阶段为合子尚未受精时期。
76.第2阶段:授粉后第7至9天,可观察到果序上花丝枯萎卷曲但尚未脱落(图1c);此阶段为双受精时期。
77.第3阶段:授粉后第10至22天,此期间果序上花丝逐渐脱落,花柱逐渐变黄枯萎(图1d);此阶段合子处于休眠时期。
78.第4阶段,授粉后第23至26天,可观察到果序上花丝和花柱已完全脱落(图1e);此阶段合子陆续结束休眠,开始第一次有丝分裂。
79.第5阶段,授粉后第27至28天,可观察到除果序开始明显膨大外,外形再无其他明显变化;此阶段合子继续分裂,发育至球形胚。
80.表1授粉后花蕾及果序发育阶段特点
81.[0082][0083]
实施例2野外非离体热激诱变处理:
[0084]
根据授粉后果序发育时长和外部形态特征,对处于授粉后发育至第4和第5阶段的果序进行野外非离体热激诱变处理(图2)。
[0085]
1、前处理
[0086]
对发育至第4和第5阶段的果序所在的花枝,每个阶段依据授粉后天数各自选定2组目标花枝进行前处理,即对发育至第4、5阶段的每个发育阶段的不同授粉天数的果序着生花枝各自选择2组进行前处理,也就是对授粉后第23天至第28天的果序着生花枝各自选择2组进行前处理,其中前处理为对目标花枝上的叶片和果序进行充分的喷水润湿,至有凝聚水珠自叶片流下后立即使用二氧化硅气凝胶材料包裹着生有果序的整个花枝;
[0087]
二氧化硅气凝胶材料的作用是在后续热激处理过程中为果序和叶片保湿、避免果序与热激处理装置的极高温度电热片直接接触,从而提高果序生活力。
[0088]
前处理过程中以二氧化硅气凝胶材料包裹花枝为例,其他材料如防火隔热布、陶瓷纤维棉等均适用于本发明,由于在野外使用,为防止花枝处理过程中防风、防雨、轻便考虑,选择二氧化硅气凝胶材料适于野外非离体使用。
[0089]
2、热激处理
[0090]
采用申请人已获得授权的发明专利“树木非离体枝芽加热处理装置”(专利号:zl200610113448.x)对每个目标花枝分别进行热激处理。该“树木非离体枝芽加热处理装置”的高温加热袋和温度控制器通过电缆线相连,且与电源相连通。
[0091]
将“树木非离体枝芽加热处理装置”的加热袋(如图2)袋体通过开口端套装在待诱变处理花枝上,并将袋体的开口端通过挠性绳紧固在待热激诱变处理的花枝上,并密封包裹于花枝上。
[0092]
将“树木非离体枝芽加热处理装置”的温度控制器放于地面安全处,将加热袋的袋体与温度控制器两者通过电线相连(图2)。
[0093]
接通电源后加热袋按照温度控制器的设定将温度升至预设温度(44
±
1℃)后保持恒温,保证加热袋包裹的花枝处于44
±
1℃的预设温度。处理温度恒定后开始计时,于开始处理后3小时、6小时分别解除加热处理。
[0094]
3、后处理
[0095]
热激处理结束后,立即对目标花枝进行后处理,及时撤去加热处理装置及二氧化硅气凝胶材料,并立即对花枝上的叶片及果序大量喷洒预冷至4℃的0.1%磷酸二氢钾水溶液,直至叶面温度降到10℃以下。
[0096]
大量喷洒预冷的0.1%磷酸二氢钾水溶液的目的是为叶片和果序快速补水和降温、促进热激后植物组织修复损伤,从而提高果序生活力,提高四倍体得率。
[0097]
本发明中后处理以喷洒0.1%磷酸二氢钾水溶液为例进行说明,其他浓度0.05-0.2%的磷酸二氢钾水溶液也适用于本发明;而且还可以喷洒大量的清水;
[0098]
后处理喷洒清水或磷酸二氢钾水溶液的温度以4℃为例进行说明,其他温度如4-20℃也适用于本发明。
[0099]
根据热激处理温度和时长为目标花枝分组编号并标记。热激处理温度和时长如表2。
[0100]
实施例2a
[0101]
除了步骤2)热激处理过程中,控制加热袋体内温度保持为40℃,其余与实施例2相同。
[0102]
实施例2b
[0103]
除了步骤2)热激处理过程中,控制加热袋体内温度保持为48℃,其余与实施例2相同。
[0104]
温度设置为48℃的处理条件下,不论处理时长为3小时或6小时,枝条受热激影响干枯,无法收到种子。
[0105]
而本发明方法热激处理温度为40℃与44℃处理条件下的枝条未显出异常,果序能够正常生长结实。次年种子成熟时,按处理组合采集种子,共收获经处理的尾叶桉种子2197粒。
[0106]
实施例3倍性检测:
[0107]
1、育苗
[0108]
次年3-4月份待种子成熟后,按照标记收集实施例2、2a桉树种子,共收获经处理的尾叶桉种子2197粒。
[0109]
将收获的种子按照标记组别分别进行播种,育苗(图3)。热激温度为40℃和44℃的处理条件下,3小时和6小时的处理时长下收获的种子均可出苗(表2)。苗木长至三对叶时进行移栽,移栽后有1074株尾叶桉种子苗成活(如表2)。
[0110]
2、流式细胞仪对尾叶桉子代倍性水平初步鉴定
[0111]
待幼苗长至10cm高后,从植株上摘取部分叶片,叶片切碎后经50μm尼龙网过滤,过滤后细胞悬浮液加入50μg ml-1
的4’,6-diamidino-2-phenylindole(dapi)中,并使用流式细胞仪分析尾叶桉子代的倍性水平,分析时使用尾叶桉二倍体叶片作为对照。
[0112]
使用流式细胞仪对1074棵尾叶桉种子苗进行倍性检测,初步鉴定出4株四倍体尾叶桉(见表2)。流式细胞仪鉴定种子苗倍性检测结果如图4,图4a为二倍体植株倍性检测,a图峰值在坐标轴50左右为二倍体;图4b为四倍体植株倍性检测,b图峰值在坐标轴100左右为四倍体。
[0113]
3、体细胞染色体计数
[0114]
经流式细胞仪初步鉴定出的四倍体植株,最终通过体细胞染色体计数进行二次鉴定,用以确定植株的倍性水平。
[0115]
流式细胞仪鉴定出的四倍体植株的嫩叶,置于卡诺固定液中固定处理24h;接着将固定样品水洗至完全去除卡诺溶液后,置于1n盐酸中,于室温下解离处理15分钟;然后将解离处理后的样品使用卡宝品红染色剂进行染色并压碎制片;使用光学显微镜观察样品制片,并对样品的染色体数目进行计数,以确定倍性水平。
[0116]
表2非离体热激诱导尾叶桉合子染色体加倍
[0117][0118][0119]
桉树二倍体、四倍体染色体计数结果如图4,其中光学显微镜观察并计数二倍体尾叶桉的染色体数目为2n=2x=22,如图4c,2n表示体细胞,x表示染色体基数,本实施例中桉树的x=11,2x表示两套染色体组,二倍体,共计2
×
11=22条染色体;光学显微镜观察本发明流式细胞仪初步鉴定的四倍体尾叶桉植株染色体数目均为2n=4x=44,如图4d,四倍体2n=4x=44,表示:体细胞具有4套染色体,是四倍体,共计4
×
11=44条染色体。通过体细胞染色体计数,最终鉴定确认4株四倍体尾叶桉
[0120]
二倍体、四倍体植株盆栽苗如图5,其中5a为二倍体;5b为四倍体。
[0121]
检测结果表明:当果序发育至第4阶段时,采用44
±
1℃热激处理3-6小时,此阶段中授粉后第24-26天的组别均获得了四倍体。尤其是以第4阶段,即授粉后25天、果序上花丝和花柱完全脱落时,采用44
±
1℃热激处理6小时获得四倍体的效率最高,四倍体得率达到6.45%。
[0122]
本发明的多次试验中,在授粉后果序花丝、花柱完全脱落阶段(通常在授粉后23-26天),均可获得四倍体。
[0123]
对照例1
[0124]
将按照实施例1方法步骤获得的授粉后发育至第4和第5阶段的果序不经过热激诱导处理,直接于次年3-4月份种子成熟后,进行播种,育苗;育苗获得的子代植株按照实施例3的方式进行倍性检测,检测结果为:流式细胞仪鉴定、体细胞染色体计数均未发现四倍体。
[0125]
对照例2
[0126]
将按照实施例1方法步骤获得的授粉后发育至第4和第5阶段的果序在进行热激诱导处理后不对目标花枝及其着生的花蕾、果序进行后处理,即未对花枝上的叶片及果序大量喷洒预冷至4℃的0.1%磷酸二氢钾水溶液直至叶片和果序表面温度达到10℃以下,其余与实施例2相同。各处理组花枝及果序均因热激处理后未能及时降温、补水而失水、干枯、失活。未能收集到成熟种子,未能获得四倍体桉树植株。
[0127]
对照例3
[0128]
除了使用较大空间体积的加热处理装置,在加热袋内使用支架将待诱变处理的花枝远离袋体,不与加热袋的袋体靠近,且不使用二氧化硅气凝胶材料包裹着生有果序的整个花枝之外,其余与实施例2相同。
[0129]
热激诱导处理后于次年3-4月份种子成熟后,进行播种,育苗;育苗获得的子代植株按照实施例3的方式进行倍性检测,检测结果为:果序发育至第4阶段时,采用44
±
1℃热激处理3-6小时,可获得四倍体。其中以第4阶段,即授粉后25天、果序上花丝和花柱完全脱落时,采用44
±
1℃热激处理6小时获得四倍体的效率最高。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。