嵌合共刺激受体及其方法和用途
1.相关申请
2.本技术要求于2019年9月16日提交的美国临时专利申请第62/900,911号的优先权的权益,所述美国临时专利申请的内容通过引用以其整体并入本文。
3.领域
4.本技术涉及来源于肿瘤坏死因子受体超家族的嵌合受体。特别地,本技术涉及能够共刺激免疫细胞的嵌合肿瘤坏死因子受体超家族受体,及其用于治疗癌症的相关方法和用途。
5.背景
6.免疫监视是免疫系统不断监测身体的转化细胞的能力。癌症能够规避该系统,并且从免疫控制逃逸。t细胞被认为是免疫监视和癌症控制的主要贡献者。在过去的几十年里,已经开发了试图利用t细胞对肿瘤有响应并根除肿瘤的先天能力的疗法。第一种方法使用自体天然存在的肿瘤浸润淋巴细胞,将该肿瘤浸润淋巴细胞在体外扩增以获得大的数目并且再输注到患者中。通过从外周血单个核细胞(pbmc)中分离t细胞并将它们工程化以表达修饰的t细胞受体来对这些方法做出改进。其他方法已经集中在用靶向双特异性抗体体内刺激或重定向t细胞,但功效有限。
7.细胞毒性t淋巴细胞(ctl)通过t细胞受体(tcr)、细胞毒性效应物分子和分泌的细胞因子介导转化的和/或病毒感染的细胞的识别和清除。天然的tcr成熟经历了多轮选择以通过mhc依赖性tcr相互作用识别抗原。mhc-tcr相互作用启动t细胞活化,并且导致作用于肿瘤细胞以敏化和诱导凋亡的效应物分子的释放。癌细胞通常在mhc肽呈递机制方面具有损伤,导致抗原呈递下调。抗原损失以及因此对转化细胞的识别的损失减弱了适应性免疫反应。携带非mhc限制性合成抗原受体的工程化t细胞的使用提供了靶向其中适应性免疫系统尚未产生清除肿瘤负荷的足够稳健反应的肿瘤的策略。非mhc限制性合成抗原受体包括新类别的t细胞活化受体,所述新类别的t细胞活化受体包括:嵌合抗原受体(car)、三功能抗原受体(tri-functional antigen receptors,tac),以及其他能够在抗原结合后活化t细胞的受体。被工程化以表达这些新的合成抗原受体的t细胞除了其天然tcr外,还获得了靶向新的抗原的能力,并且在肿瘤抗原相互作用后介导识别和t细胞活化。有趣的是,这些受体允许靶向天然tcr库(repertoire)之外的表位的能力,将抗原选择扩展至诸如碳水化合物和糖脂等分子。工程化t细胞疗法因其靶向非mhc限制性肿瘤抗原、易于获得pbmc以及超过全身施用激动剂抗体的增加的安全性的能力,为肿瘤疗法提供了有吸引力的方法。car工程化t细胞的临床成功确定了这种新类别细胞药物的可行性和治疗效力。
8.共刺激/共抑制信号是与tcr连接同时发生并调节t细胞功能的那些信号。共刺激/共抑制信号的类型和强度决定了适应性免疫反应的进展。肿瘤坏死因子受体超家族(tnfrsf)及配体是关键的共刺激/共抑制分子。tnfrsf成员以多种方式介导t细胞中的反应。取决于炎性环境,tnfrsf传播的信号可以刺激或抑制效应物向记忆t细胞的分化,所述记忆t细胞由幼稚t细胞响应于抗原以及在记忆反应期间产生。t细胞在反应的每个阶段(包括幼稚阶段、效应物阶段和记忆阶段)都接收独特的活化或存活信号;共刺激信号是驱动这
些反应的主要组分。有趣的是,共刺激信号可以用来驱动t细胞反应,同时限制共抑制信号。然而,尚不完全了解这些反应是如何协调的。已经表明,数种tnfrsf的表达在t细胞活化后被诱导,并且这些分子的表达不是普遍存在的,表明这些分子可能在调节不同群体中的免疫反应中具有作用。此外,各种刺激物可以通过抗原呈递细胞(apc)促进tnfrsf配体(包括先天的(细菌配体)和适应性信号(促炎细胞因子))的表达。因此,在适应性免疫反应的各个阶段,tnfrsf通过受体在t细胞上的表达及其配体在其他免疫细胞上的表达二者来调节t细胞免疫。
9.概述
10.本发明人已经展示出,包括来自不同肿瘤坏死因子受体超家族(tnfrsf)受体的功能结构域的嵌合共刺激受体(ccr)能够将由tnfrsf配体引起的信号传导重编程至期望的反应,包括t细胞刺激反应。
11.因此,在某些实施方案中,本文公开了嵌合共刺激受体(ccr)核酸,所述嵌合共刺激受体(ccr)核酸包含(a)编码肿瘤坏死因子受体超家族(tnfrsf)成员的胞外结构域的第一多核苷酸;(b)编码跨膜结构域多肽的第二多核苷酸;和(c)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸来源于tnfrsf的不同成员。在一些实施方案中,第一多核苷酸编码具有死亡结构域的tnfrsf成员的胞外结构域。在一些实施方案中,第一多核苷酸编码以下的胞外结构域:肿瘤坏死因子受体1(tnfr1)、肿瘤坏死因子受体2(tnfr2)、fas受体、死亡结构域4(dr4)、死亡结构域5(dr5)、死亡结构域3(dr3)、死亡结构域6(dr6)、外胚层发育不良因子受体(ectodermal dysplasia receptor,edar)、外胚叶发育不良因子a2受体(ectodysplasin a2 receptor,xedar)、troy或神经生长因子受体(ngfr)。在一些实施方案中,第一多核苷酸编码tnfr1的胞外结构域。在一些实施方案中,第一多核苷酸编码tnfr2的胞外结构域。在一些实施方案中,第一多核苷酸编码fas的胞外结构域。在一些实施方案中,第一多核苷酸编码dr4的胞外结构域。在一些实施方案中,第一多核苷酸编码dr5的胞外结构域。在一些实施方案中,第一多核苷酸编码与seq id no:1至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,第一多核苷酸编码与seq id no:2至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,第三多核苷酸编码来自以下的胞质共刺激信号传导结构域:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第三多核苷酸编码来自baffr的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自4-1bb的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自cd27的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自hvem的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自ox40的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自gitr的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自taci的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码与seq id no:3至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,第三多核苷酸编码与seq id no:4至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,跨膜结构域多肽是来自肿瘤坏死因子受体超家族成员的跨膜结构域多肽。在一些实施方案
中,第二多核苷酸编码与seq id no:5至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,第二多核苷酸编码与seq id no:6至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,跨膜结构域和胞质共刺激信号传导结构域来自同一共刺激信号传导蛋白。在一些实施方案中,跨膜结构域和共刺激信号传导结构域来源于4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第一多核苷酸和第二多核苷酸被直接和/或间接(例如,通过接头)连接至第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸被直接和/或间接(例如,通过接头)连接至第二多核苷酸。
12.在某些实施方案中,本文还公开了嵌合共刺激受体(ccr)核酸,所述嵌合共刺激受体(ccr)核酸包含:(a)编码肿瘤坏死因子受体1的胞外结构域的第一多核苷酸;(b)编码跨膜结构域多肽的第二多核苷酸;和(c)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸。在一些实施方案中,第一多核苷酸编码与seq id no:1至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,胞质共刺激信号传导结构域是来自以下的胞质共刺激信号传导结构域:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,胞质共刺激信号传导结构域是来自baffr的胞质共刺激信号传导结构域。在一些实施方案中,胞质共刺激信号传导结构域是来自4-1bb的胞质共刺激信号传导结构域。在一些实施方案中,胞质共刺激信号传导结构域是来自cd27的胞质共刺激信号传导结构域。在一些实施方案中,胞质共刺激信号传导结构域是来自hvem的胞质共刺激信号传导结构域。在一些实施方案中,胞质共刺激信号传导结构域是来自ox40的胞质共刺激信号传导结构域。在一些实施方案中,胞质共刺激信号传导结构域是来自gitr的胞质共刺激信号传导结构域。在一些实施方案中,胞质共刺激信号传导结构域是来自taci的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码与seq id no:3至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,第三多核苷酸编码与seq id no:4至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,跨膜结构域多肽是来自肿瘤坏死因子受体超家族成员的跨膜结构域多肽。在一些实施方案中,第二多核苷酸编码与seq id no:5至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,第二多核苷酸编码与seq id no:6至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,跨膜结构域和胞质共刺激信号传导结构域来自同一共刺激信号传导蛋白。在一些实施方案中,跨膜结构域和共刺激信号传导结构域来源于4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第一多核苷酸和第二多核苷酸被直接和/或间接(例如,通过接头)连接至第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸被直接或间接(例如,通过接头)连接至第二多核苷酸。
13.在某些实施方案中,本文还公开了嵌合共刺激受体(ccr)核酸,所述嵌合共刺激受体(ccr)核酸包含:(a)编码肿瘤坏死因子受体2的胞外结构域的第一多核苷酸;(b)编码跨膜结构域多肽的第二多核苷酸;和(c)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸。在一些实施方案中,胞质共刺激信号传导结构域是来自以下的胞质共刺激信号传导结构域:4-1bb、baffr、ox40、cd27、cd40、
gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,胞质共刺激信号传导结构域是来自baffr的胞质共刺激信号传导结构域。在一些实施方案中,胞质共刺激信号传导结构域是来自4-1bb的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自cd27的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自hvem的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自ox40的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自gitr的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自taci的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码与seq id no:3至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,第三多核苷酸编码与seq id no:4至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,跨膜结构域多肽是来自肿瘤坏死因子受体超家族成员的跨膜结构域多肽。在一些实施方案中,第二多核苷酸编码与seq id no:5至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,第二多核苷酸编码与seq id no:6至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,跨膜结构域和胞质共刺激信号传导结构域来自同一共刺激信号传导蛋白。在一些实施方案中,跨膜结构域和共刺激信号传导结构域来源于4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第一多核苷酸和第二多核苷酸被直接和/或间接(例如,通过接头)连接至第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸被直接和/或间接(例如,通过接头)连接至第二多核苷酸。
14.在某些实施方案中,本文还公开了嵌合共刺激受体(ccr)核酸,所述嵌合共刺激受体(ccr)核酸包含:(a)编码肿瘤坏死因子受体超家族成员6(tnfrsf6;fas)的胞外结构域的第一多核苷酸;(b)编码跨膜结构域多肽的第二多核苷酸;和(c)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸。在一些实施方案中,第一多核苷酸编码与seq id no:2至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,胞质共刺激信号传导结构域是来自以下的胞质共刺激信号传导结构域:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,胞质共刺激信号传导结构域是来自baffr的胞质共刺激信号传导结构域。在一些实施方案中,胞质共刺激信号传导结构域是来自4-1bb的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自cd27的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自hvem的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自ox40的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自gitr的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自taci的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码与seq id no:3至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,第三多核苷酸编码与seq id no:4至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,跨膜结构域多肽是来自肿瘤坏死因子受体超家族成员的跨膜结构域多肽。在一些实施方案中,第二多核苷酸编码与seq id no:5至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的寡肽。在一些实施方案中,第二多核苷酸编码与seq id no:6至少80%、至少85%、至少90%、至少
95%、至少99%或100%相同的寡肽。在一些实施方案中,跨膜结构域和胞质共刺激信号传导结构域来自同一共刺激信号传导蛋白。在一些实施方案中,跨膜结构域和共刺激信号传导结构域来源于4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第一多核苷酸和第二多核苷酸被直接和/或间接(例如,通过接头)连接至第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸被直接和/或间接(例如,通过接头)连接至第二多核苷酸。
15.在某些实施方案中,本文还公开了由本文描述的嵌合共刺激受体(ccr)核酸编码的多肽。
16.在某些实施方案中,本文还公开了与seq id no:7至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的嵌合共刺激受体(ccr)多肽。
17.在某些实施方案中,本文还公开了与seq id no:8至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的嵌合共刺激受体(ccr)多肽。
18.在某些实施方案中,本文还公开了与seq id no:9至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的嵌合共刺激受体(ccr)多肽。
19.在某些实施方案中,本文还公开了与seq id no:10至少80%、至少85%、至少90%、至少95%、至少99%或100%相同的嵌合共刺激受体(ccr)多肽。
20.另外地,在某些实施方案中,本文公开了t细胞,所述t细胞表达本文描述的嵌合共刺激受体(ccr)核酸或包含本文公开的多肽。在一些实施方案中,t细胞是细胞毒性t细胞、辅助性t细胞、调节性t细胞或γ-δt细胞。
21.在某些实施方案中,本文还公开了t细胞(例如,用于治疗有相应需要的受试者中的癌症),所述t细胞包含(a)本文公开的嵌合共刺激受体(ccr)核酸;和(b)编码可以识别靶特异性配体的工程化t细胞受体(tcr)或合成抗原受体多肽的第二核酸。在一些实施方案中,靶特异性配体结合癌细胞上的抗原。在一些实施方案中,合成抗原受体多核苷酸是嵌合抗原受体(car)、t细胞抗原偶联剂(tac)或bite(双特异性t细胞衔接物(bispecific t-cell engager))。在一些实施方案中,合成抗原受体多核苷酸是t细胞抗原偶联剂(tac)。在一些实施方案中,t细胞是细胞毒性t细胞、辅助性t细胞、调节性t细胞或γ-δt细胞。
22.在某些实施方案中,本文还公开了t细胞(例如,用于治疗有相应需要的受试者中的癌症),所述t细胞包含(a)嵌合共刺激受体(ccr)核酸,所述嵌合共刺激受体(ccr)核酸包含:(i)编码来自肿瘤坏死因子受体1的胞外结构域的第一多核苷酸;(ii)编码跨膜结构域多肽的第二多核苷酸;和(iii)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸;以及(b)编码可以识别靶特异性配体的工程化t细胞受体(tcr)或合成抗原受体多肽的第二核酸。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自以下的胞质共刺激信号传导结构域:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自baffr的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自4-1bb的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自cd27的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自hvem的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自ox40的胞质共刺激信号传导结构域。在一些实施方案中,
第三多核苷酸编码来自gitr的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自taci的胞质共刺激信号传导结构域。在一些实施方案中,跨膜结构域和胞质共刺激信号传导结构域来自同一共刺激信号传导蛋白。在一些实施方案中,跨膜结构域和共刺激信号传导结构域来源于4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第一多核苷酸和第二多核苷酸被直接或间接(例如,通过接头)连接至第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸被直接或间接(例如,通过接头)连接至第二多核苷酸。在一些实施方案中,靶特异性配体结合癌细胞上的抗原。在一些实施方案中,合成抗原受体多核苷酸是嵌合抗原受体(car)、t细胞抗原偶联剂(tac)或bite(双特异性t细胞衔接物)。在一些实施方案中,合成抗原受体多核苷酸是t细胞抗原偶联剂(tac)。在一些实施方案中,t细胞是细胞毒性t细胞、辅助性t细胞、调节性t细胞或γ-δt细胞。
23.在某些实施方案中,本文还公开了t细胞(例如,用于治疗有相应需要的受试者中的癌症),所述t细胞包含(a)嵌合共刺激受体(ccr)核酸,所述嵌合共刺激受体(ccr)核酸包含:(i)编码来自肿瘤坏死因子受体2的胞外结构域的第一多核苷酸;(ii)编码跨膜结构域多肽的第二多核苷酸;和(iii)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸;以及(b)编码可以识别靶特异性配体的工程化t细胞受体(tcr)或合成抗原受体多肽的第二核酸。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自以下的胞质共刺激信号传导结构域:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自baffr的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自4-1bb的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自cd27的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自hvem的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自ox40的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自gitr的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自taci的胞质共刺激信号传导结构域。在一些实施方案中,跨膜结构域和胞质共刺激信号传导结构域来自同一共刺激信号传导蛋白。在一些实施方案中,跨膜结构域和共刺激信号传导结构域来源于4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第一多核苷酸和第二多核苷酸被直接或间接(例如,通过接头)连接至第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸被直接或间接(例如,通过接头)连接至第二多核苷酸。在一些实施方案中,靶特异性配体结合癌细胞上的抗原。在一些实施方案中,合成抗原受体多核苷酸是嵌合抗原受体(car)、t细胞抗原偶联剂(tac)或bite(双特异性t细胞衔接物)。在一些实施方案中,合成抗原受体多核苷酸是t细胞抗原偶联剂(tac)。在一些实施方案中,t细胞是细胞毒性t细胞、辅助性t细胞、调节性t细胞或γ-δt细胞。
24.在某些实施方案中,本文还公开了t细胞(例如,用于治疗有相应需要的受试者中的癌症),所述t细胞包含(a)嵌合共刺激受体(ccr)核酸,所述嵌合共刺激受体(ccr)核酸包含:(i)编码来自fas受体的胞外结构域的第一多核苷酸;(ii)编码跨膜结构域多肽的第二
多核苷酸;和(iii)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸;以及(b)编码可以识别靶特异性配体的工程化t细胞受体(tcr)或合成抗原受体多肽的第二核酸。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自以下的胞质共刺激信号传导结构域:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自baffr的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自4-1bb的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自cd27的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自hvem的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自ox40的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自gitr的胞质共刺激信号传导结构域。在一些实施方案中,嵌合共刺激受体(ccr)核酸的第三多核苷酸编码来自taci的胞质共刺激信号传导结构域。在一些实施方案中,跨膜结构域和胞质共刺激信号传导结构域来自同一共刺激信号传导蛋白。在一些实施方案中,跨膜结构域和共刺激信号传导结构域来源于4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第一多核苷酸和第二多核苷酸被直接或间接(例如,通过接头)连接至第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸被直接或间接(例如,通过接头)连接至第二多核苷酸。在一些实施方案中,靶特异性配体结合癌细胞上的抗原。在一些实施方案中,合成抗原受体多核苷酸是嵌合抗原受体(car)、t细胞抗原偶联剂(tac)或bite(双特异性t细胞衔接物)。在一些实施方案中,合成抗原受体多核苷酸是t细胞抗原偶联剂(tac)。在一些实施方案中,t细胞是细胞毒性t细胞、辅助性t细胞、调节性t细胞或γ-δt细胞。
25.在某些实施方案中,本文公开了治疗有相应需要的个体中的癌症的方法,所述方法包括向个体施用包含嵌合共刺激受体(ccr)核酸的免疫细胞,所述嵌合共刺激受体(ccr)核酸包含(a)编码肿瘤坏死因子受体超家族成员的胞外结构域的第一多核苷酸;(b)编码跨膜结构域多肽的第二多核苷酸;和(c)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸来源于tnfrsf的不同成员。在一些实施方案中,第一多核苷酸编码具有死亡结构域的tnfrsf成员的胞外结构域。在一些实施方案中,第一多核苷酸编码以下的胞外结构域:tnfr1、tnfr2、fas、dr4、dr5、dr3、dr6、edar、xedar、troy或ngfr。在一些实施方案中,第三多核苷酸编码来自以下的胞质共刺激信号传导结构域多肽:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,免疫细胞是t细胞。在一些实施方案中,t细胞是工程化t细胞。在一些实施方案中,免疫细胞是自然杀伤细胞(nk细胞)。在一些实施方案中,免疫细胞是巨噬细胞。在一些实施方案中,免疫细胞是肿瘤浸润淋巴细胞(til)。在一些实施方案中,免疫细胞是单核细胞。在一些实施方案中,免疫细胞是b细胞。在一些实施方案中,免疫细胞是本文公开的免疫细胞。在一些实施方案中,癌症是白血病或淋巴瘤。在一些实施方案中,癌症是混合谱系白血病(mll)、慢性淋巴细胞白血病(cll)、急性淋巴细胞母白血病(all)、大b细胞淋巴瘤、弥漫性大b细胞淋巴瘤、原发性纵隔b细胞淋巴瘤、高级别b细胞淋巴瘤或源于滤泡性淋巴瘤的大b细胞淋巴瘤。在一些实施
方案中,癌症是肺癌、乳腺癌、结肠癌、多发性骨髓瘤、胶质母细胞瘤、胃癌、卵巢癌、胃癌、结肠直肠癌、尿路上皮癌、子宫内膜癌或黑素瘤。在一些实施方案中,癌症是肺癌。在一些实施方案中,癌症是乳腺癌。在一些实施方案中,癌症是结肠癌。在一些实施方案中,癌症是多发性骨髓瘤。在一些实施方案中,癌症是胶质母细胞瘤。在一些实施方案中,癌症是胃癌(gastric cancer)。在一些实施方案中,癌症是卵巢癌。在一些实施方案中,癌症是胃癌(stomach cancer)。在一些实施方案中,癌症是尿路上皮癌。在一些实施方案中,癌症是子宫内膜癌。在一些实施方案中,癌症是黑素瘤。
26.在某些实施方案中,本文公开了药物组合物,所述药物组合物包含(a)免疫细胞,所述免疫细胞包含(i)编码肿瘤坏死因子受体超家族成员的胞外结构域的第一多核苷酸;(ii)编码跨膜结构域多肽的第二多核苷酸;和(iii)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸;以及(b)药学上可接受的载体。在一些实施方案中,第一多核苷酸和第三多核苷酸来源于tnfrsf的不同成员。在一些实施方案中,第一多核苷酸编码具有死亡结构域的tnfrsf成员的胞外结构域。在一些实施方案中,第一多核苷酸编码以下的胞外结构域:tnfr1、tnfr2、fas、dr4、dr5、dr3、dr6、edar、xedar、troy或ngfr。在一些实施方案中,第一多核苷酸编码tnfr1的胞外结构域。在一些实施方案中,第一多核苷酸编码tnfr2的胞外结构域。在一些实施方案中,第一多核苷酸编码fas的胞外结构域。在一些实施方案中,第三多核苷酸编码来自以下的胞质共刺激信号传导结构域:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第三多核苷酸编码来自baffr的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自4-1bb的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自cd27的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自hvem的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自ox40的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自gitr的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自taci的胞质共刺激信号传导结构域。在一些实施方案中,跨膜结构域和胞质共刺激信号传导结构域来自同一共刺激信号传导蛋白。在一些实施方案中,跨膜结构域和共刺激信号传导结构域来源于4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第一多核苷酸和第二多核苷酸被直接或间接(例如,通过接头)连接至第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸被直接或间接(例如,通过接头)连接至第二多核苷酸。在一些实施方案中,免疫细胞是t细胞、自然杀伤细胞(nk细胞)、巨噬细胞、肿瘤浸润淋巴细胞(til)、单核细胞或b细胞。在一些实施方案中,免疫细胞是本文公开的免疫细胞。
27.在某些实施方案中,本文公开了载体构建体,所述载体构建体包含:(a)本文公开的嵌合共刺激受体(ccr);和(b)在哺乳动物细胞中有功能的启动子。
28.在某些实施方案中,本文公开了用本文公开的载体构建体转染的分离的或工程化的t淋巴细胞、自然杀伤细胞、巨噬细胞、肿瘤浸润淋巴细胞或单核细胞。
29.本技术的其他特征和优势将由以下详细描述变得清晰。然而,应该理解的是,详细描述和特定实施例,虽然指示本技术的实施方案,但仅通过说明性方式给出,并且权利要求书的范围不应受这些实施方案的限制,而应给出与作为整体的描述一致的最宽泛解释。
withdrawal)后的存活
48.图8a示出了在没有补充生长因子的情况下用单独的抗cd3刺激的t细胞。图8b示出了在没有补充细胞因子的情况下用单独的tnfα刺激的t细胞。
49.图9:tnfr1-4-1bb ccr刺激改变细胞因子谱(cytokine profile)
50.在培养第14天测试用tnfr1-4-1bb ccr工程化的t细胞在抗cd3刺激后的细胞因子产生。将细胞在存在高尔基体转运抑制剂的情况下在抗cd3涂覆的板上孵育4小时。通过流式细胞术将细胞针对胞内细胞因子产生染色并评价。
51.图10:用2a表达系统工程化的tac ccr t细胞的表型。
52.将t细胞在第0天用抗cd3/cd28珠活化。一天后,将t细胞用携带针对骨髓瘤蛋白bcma的tac受体的慢病毒或编码相同tac受体连同由小rna病毒(picornavirus)2a序列分开的tnfr1-4-1bb ccr(tac ccr)的慢病毒进行转导。将细胞保持在具有il2和il7的培养物中,每2天添加新鲜培养基。表面tac和ccr的染色以及检测在培养的第14天通过流式细胞术来进行。2a系统中tac的表面蛋白水平低于单一表达系统。在2a工程化t细胞的表面上检测到tnfr1-4-1bb ccr蛋白。
53.图11:tac ccr t细胞显示出bcma 肿瘤细胞靶的裂解
54.t细胞介导体外肿瘤靶的裂解。将t细胞与表达萤光素酶的kms11肿瘤靶共孵育24小时。使用萤光素酶活性作为肿瘤细胞裂解的度量。ut(非工程化)t细胞显示出kms11肿瘤靶的检测不到的肿瘤裂解。用tac或tac ccr工程化的t细胞如图10中描述的。经24小时的测定,两个工程化t细胞群体显示出相似的肿瘤靶裂解。
55.图12:ccr工程化tac t细胞在刺激后产生较少的炎性细胞因子
56.将工程化t细胞在存在高尔基体转运抑制剂(golgi plug)的情况下与kms11肿瘤靶共孵育。共孵育4小时后,将细胞针对胞内细胞因子产生进行染色。对细胞因子il2、tnfα和ifnγ进行评价。tac t细胞在识别其同源配体后容易地产生tnfα和ifnγ。共表达tnfr1-4-1bb ccr的tac t细胞看起来产生较少的细胞因子分泌细胞。
57.图13a-图13b:tac ccr工程化t细胞的增殖与富集
58.图13a图示出描绘增殖的工程化t细胞(proliferating engineered t cells)的稀释峰的celltrace
tm violet直方图。图13b示出了cd4和cd8 t细胞的分裂指数和增殖指数的定量。数据点指示成对的增殖测定。分裂指数表示分裂的细胞变成的t细胞的平均数。增殖测定表示初始细胞变成的t细胞的平均数。相对于表达单独的bcma特异性tac(tac)的t细胞,被工程化为表达bcma特异性tac和tnfr1-4-1bb ccr(tac costim)的cd8 和cd4 t细胞表现出更高的分裂指数。被工程化为表达bcma特异性tac和tnfr1-4-1bb ccr(tac costim)的cd4 t细胞表现出比表达单独的bcma特异性tac(tac)的cd4 t细胞更高的增殖指数。(*p《0.05)。
59.图14:抗cd3刺激的tnfrsf增殖的筛选
60.将用抗cd3/抗cd28珠刺激并在il2和il7中生长的pbmc用tnfr1-4-1bb ccr、tnf-阻断剂和ngfr(转导对照)工程化。在培养第14天,工程化细胞在板结合的抗cd3刺激5天后评价其增殖。将细胞用celltrace
tm violet标记,并且世代峰(generation peaks)用流式细胞术来评价。板结合的抗cd3刺激所有组中的cd4和cd8 t细胞二者的增殖。用tnfr1-4-1bb ccr工程化的cd4 和cd8 t细胞比ngfr或tnf-阻断剂工程化的细胞更多地增殖。
61.图15:cd4 和cd8 t细胞的tnfrsf ccr工程化效率
62.将用抗cd3/抗cd28珠刺激并在il2和il7中生长的pbmc用y轴上列出的tnfrsf ccr构建体工程化。在培养第14天,评价主体(bulk)细胞群体的工程化效率。将细胞针对基因盒中包含的转导标志物ngfr染色。将细胞针对淋巴细胞/单细胞/cd4 或cd8 /ngfr 进行门控。(n=1-5)。
63.图16:ccr的平均萦光强度
64.将用抗cd3/抗cd28珠刺激并在il2和il7中生长的pbmc用y轴上列出的tnfrsf ccr筛选构建体工程化。在培养第14天,评价工程化细胞的ccr表面表达。将细胞针对ccr的胞外部分(tnfr1)染色。ccr mfi根据针对淋巴细胞/单细胞/cd4 或cd8 /ngfr 门控的群体进行计算。报告的ccr mfi归一化至原始tnfr1-4-1bb ccr(og4-1bb)mfi以校正实验间批次效应。(n=1-5)。
65.图17:ccr t细胞的生长
66.将用抗cd3/抗cd28珠刺激并在il2和il7中生长的pbmc用y轴上列出的tnfrsf ccr筛选构建体工程化。每两天向培养物补充新鲜培养基和细胞因子。在培养第14天,记录主体细胞群体的活细胞计数。培养物以105个pbmc开始。ngfr转导的对照以红色标记。(n=1-5)。
67.图18:cd4 ccr t细胞增殖
68.将用抗cd3/抗cd28珠刺激并在il2和il7中生长的pbmc用y轴上列出的tnfrsf ccr筛选构建体工程化。在培养第14天,评价工程化细胞用板结合的抗cd3刺激后的增殖持续5天。将细胞用celltrace
tm violet标记,并且世代峰通过流式细胞术来评价。增殖通过fcs增殖模型来定量,并且通过增殖指数来表示。用单独的ngfr转导标志物工程化的对照细胞被标识(红线)。将细胞群体针对活细胞/淋巴细胞/单细胞/cd4 /ngfr 门控。ngfr转导的对照以红色标记。
69.图19:cd8 ccr t细胞增殖筛选
70.将用抗cd3/抗cd28珠刺激并在il2和il7中生长的pbmc用y轴上列出的tnfrsf ccr筛选构建体工程化。在培养第14天,评价工程化细胞用板结合的抗cd3刺激后的增殖持续5天。将细胞用celltrace
tm violet标记,并且世代峰通过流式细胞术来评价。增殖通过fcs增殖模型来定量,并且通过增殖指数来表示。用单独的ngfr转导标志物工程化的对照细胞被标识(红线)。将细胞群体针对活细胞/淋巴细胞/单细胞/cd8 /ngfr 门控。ngfr转导的对照以红色标记。
71.图20:ccr筛选构建体聚类中心(clustering centroid)的相关热图
72.tnfrsf ccr工程化t细胞的表型和功能数据通过主成分分析(pca)和k-均值聚类进行分析。聚类中心的相关热图将具有相似属性的ccr聚类在一起。在树状图中,不同的跨膜结构域和信号传导结构域显示为不同的颜色以显示分组。分析中鉴定的组用于突出ccr对进一步研究的重要性。
73.图21a-图21b:ccr t细胞的增殖
74.将抗cd3/抗cd28珠刺激并在il2和il7中生长的pbmc用tnfr1-4-1bb、tnfr1-baffr、tnf-阻断剂和ngfr(转导对照)工程化。在培养第14天,评价工程化细胞用板结合的抗cd3刺激后的增殖持续5天。将细胞用celltrace
tm violet标记,并且世代峰用流式细胞术
来评价。增殖通过fcs增殖模型来定量,并且通过增殖指数来表示。所示值在供体内归一化至ngfr(转导对照)。在所有条件下,板结合的抗cd3刺激都导致cd4和cd8 t细胞子集二者的增殖。图21a示出了经5天,用tnfr-baffr和tnfr1-4-1bb ccr工程化的cd4 t细胞比ngfr或tnf-阻断剂工程化的细胞更多地增殖。图21b,经5天,用tnfr-baffr和tnfr1-4-1bb ccr工程化的cd8 t细胞比ngfr或tnf-阻断剂工程化的细胞更多地增殖。(*=p《0.05)。
75.图22a-图22b:ccr工程化t细胞的胞内细胞因子产生
76.在培养第14天测试用ccr受体工程化的t细胞在抗cd3刺激后的细胞因子产生。将细胞在存在高尔基体转运抑制剂的情况下在抗cd3涂覆的板上孵育4小时。通过流式细胞术将细胞针对胞内细胞因子产生染色并评价。图22a示出了在刺激后高百分比的t细胞分泌ifnγ。与转导ngfr的对照相比,用4-1bb/baffr ccr或tnf-阻断剂工程化的t细胞均未改变ifnγ产生。图22b示出了在抗cd3刺激后,高百分比的531(ngfr转导的对照)工程化t细胞产生tnfα。用tnf-阻断剂、baffr或4-1bb ccr工程化的细胞在刺激后,在cd4 和cd8 t细胞子集二者中具有降低量的表达tnfα的t细胞。
77.图23a-图23c:tnfr融合构建体
78.用于pcr并连接到pccl载体的tnfr融合构建体的代表性图。图23a图示了全长tnfr1天然受体被克隆用于在tnfr-融合的表征中作为对照受体使用。图23b图示了ccr受体通过将tnfr1受体的胞外结构域和跨膜结构域二者与4-1bb的胞内信号传导结构域融合,产生共刺激tnfr1-4-1bb ccr受体而克隆。图23c图示了作为显性负性受体(dominant negative receptor)使用,天然tnfr1受体被截短以去除胞质信号传导结构域并且被称为tnf-阻断剂受体。
79.图24:fas-嵌合体t细胞的工程化效率
80.将用抗cd3/抗cd28珠活化的人类pbmc用编码截短的fas(fasr-trunc)或者包含fas的胞外结构域和跨膜结构域以及4-1bb或baff-r的胞质结构域的fas-嵌合体(分别地,fas-41bb和fas-baffr)的慢病毒进行转导,如图2b中描述的。作为对照,pbmc用编码ngfr的慢病毒进行转导。细胞在il2/il7中生长,每两天添加新鲜培养基。培养第14天,收集细胞,并且通过流式细胞术针对转导效率染色。图的x轴描绘了被工程化以表达所命名的受体的t细胞。(n=3,独立实验)。
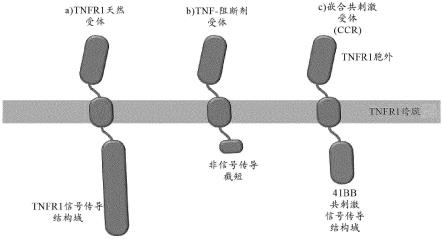
81.图25a-图25b:fas嵌合体工程化t细胞的表面表达
82.将用抗cd3/抗cd28珠活化的人类pbmc用编码截短的fas(fasr-trunc)或者包含fas的胞外结构域和跨膜结构域以及4-1bb或baff-r的胞质结构域的fas嵌合体(分别地,fas-41bb和fas-baffr)的慢病毒进行转导,如图2b中描述的。作为对照,pbmc用编码ngfr的慢病毒进行转导。细胞在il2/il7中生长,每两天添加新鲜培养基。培养第14天,收集细胞,并且通过流式细胞术针对fas染色。图25a.描绘为展示高于基线表达的fas的%ngfr阳性细胞的fas-嵌合体工程化t细胞中fas和ngfr表达的点图。图25b.工程化t细胞中fas表达(mfi)的条形图。高于ngfr的fas表达是对修饰的fas受体的间接测量。x轴描绘了被工程化以表达所命名的受体的t细胞。((n=3),独立实验)。
83.图26:fas-嵌合体t细胞在初始扩增期间的生长
84.将用抗cd3/抗cd28珠活化的人类pbmc用编码截短的fas(fasr-trunc)或者包含
fas的胞外结构域和跨膜结构域以及4-1bb或baff-r的胞质结构域的fas-嵌合体(分别地,fas-41bb和fas-baffr)的慢病毒进行转导,如图2b中描述的。作为对照,pbmc用编码ngfr的慢病毒进行转导。细胞在il2/il7中生长,每两天补充新鲜培养基。在培养第14天,细胞扩增被测量为总活细胞计数。x轴描绘了被工程化以表达所命名的受体的t细胞。(n=3,独立实验)。
85.图27:fas工程化t细胞的增殖。
86.用抗cd3/抗cd28珠活化的人类pbmc被工程化以表达修饰的fas受体(fas-trunc、fas-4-1bb、fas-baffr)或不表达受体(ngfr)。将工程化t细胞用cell trace violet标记,用抗cd3刺激并培养4天。增殖通过fcs 增殖模型来定量,并且通过归一化至未工程化的增殖指数的增殖指数来表示。x轴描绘了被工程化以表达cd8(图27a)或cd4(图27b)的经刺激的t细胞。(n=3,独立实验)。
87.图28:fas-嵌合体t细胞在存在fasl的情况下的存活力。
88.用抗cd3/抗cd28珠活化的人类pbmc被工程化以表达修饰的fas受体(fas-trunc、fas-4-1bb、fas-baffr)或不表达受体(ngfr)。工程化t细胞在存在递增浓度的fasl的情况下培养。48小时之后通过alamarblue测量存活力。(n=3个重复,图表示3个独立实验)。
89.图29:正增殖的fas工程化t细胞的alamarblue测定。
90.用抗cd3/抗cd28珠活化的人类pbmc被工程化以表达修饰的fas受体(fas-trunc、fas-4-1bb、fas-baffr)或不表达受体(ngfr)。工程化t细胞用cell trace violet标记,用抗cd3刺激,并且在存在或不存在递增浓度的fasl(x轴)的情况下培养4天。增殖通过alamarblue测量。图例:用所命名的受体工程化的t细胞。(n=3个重复,表示3个独立实验)。
91.详细描述
92.定义
93.如本文使用的术语“细胞”包括单细胞以及多于一个细胞。
94.如本文使用的术语“t细胞”是指在细胞介导的免疫中发挥核心作用的一种类型的淋巴细胞。t细胞,也称为t淋巴细胞,可以通过细胞表面上t细胞受体(tcr)的存在与其他淋巴细胞,诸如b细胞和自然杀伤细胞区分开。存在数种具有不同功能的t细胞子集,包括但不限于,辅助性t细胞、细胞毒性t细胞、记忆t细胞、调节性t细胞和自然杀伤t细胞。在一些实施方案中,t细胞是工程化t细胞。
95.术语“工程化tcr”或“工程化t细胞受体”意指已从其天然存在的形式被修饰的任何tcr。工程化tcr可以具有对α链和/或β链或者γ链和/或δ链进行的使tcr能够识别特定抗原(例如,新抗原)的修饰(包括替代任何前述链)。工程化tcr可以具有对任何cd3亚基(例如cd3ε,如在truc受体的情况下)的修饰,包括添加抗原识别结构域(例如,抗体、scfv、darpin)。工程化tcr可以具有与α链和/或β链或者γ链和/或δ链的跨膜结构域连接的抗原识别结构域(例如,抗体、scfv、darpin)。
96.如本文使用的术语“多核苷酸”和/或“核酸序列”和/或“核酸”是指由碱基、糖和糖间(骨架)连接组成的核苷或核苷酸单体的序列。该术语还包括包含非天然存在的单体或其部分的修饰或取代的序列。本技术的核酸序列可以是脱氧核糖核酸序列(dna)或核糖核酸序列(rna),并且可以包括天然存在的碱基,包括腺嘌呤、鸟嘌呤、胞嘧啶、胸苷和尿嘧啶。序列也可以包含修饰的碱基。这样的修饰的碱基的实例包括氮杂和脱氮杂的腺嘌呤、鸟嘌呤、
胞嘧啶、胸苷和尿嘧啶;黄嘌呤和次黄嘌呤。本公开内容的核酸可以从生物有机体中分离、通过实验室遗传重组方法形成或者通过化学合成或其他已知的产生核酸的方案获得。
97.如本文使用的术语“分离的多核苷酸”或“分离的核酸序列”是指在通过重组dna技术产生时大体上不含细胞物质或培养基的核酸,或者在化学合成时大体上不含化学前体或其他化学物质的核酸。分离的核酸也大体上不含天然位于该核酸来源于其中的核酸侧翼的序列(即,位于核酸5'末端和3'末端的序列)。术语“核酸”意图包括dna和rna,并且可以是双链或单链的,并且表示有义链或反义链。此外,术语“核酸”包括互补核酸序列。
98.如本文使用的术语“重组核酸”或“工程化核酸”是指未在生物有机体中发现的核酸或多核苷酸。例如,重组核酸可以通过实验室遗传重组方法(诸如分子克隆)产生本来不会在自然界中发现的序列来形成。重组核酸也可以通过化学合成或其他已知的产生核酸的方案来产生。除非另外指示,否则本部分和其他部分中描述的定义和实施方案意图适用于本文关于它们如本领域技术人员将理解的所适合的实施方案和方面描述的本技术的所有实施方案和方面。
99.如本文使用的术语“多肽”或“蛋白”描述了与核酸编码的氨基酸对应的氨基酸链。本公开内容的多肽或蛋白可以是通常描述2个至约30个氨基酸的氨基酸链的肽。如本文使用的术语蛋白还描述具有多于30个氨基酸的氨基酸链,并且可以是蛋白或全长蛋白的片段或结构域。此外,如本文使用的,术语蛋白可以指氨基酸的线性链,或者它可以指已经加工和折叠为功能蛋白的氨基酸链。然而,应当理解,30是关于区分肽和蛋白的任意数,并且这些术语可以可互换地用于氨基酸链。本公开内容的蛋白可以通过从天然产生蛋白的细胞中分离和纯化蛋白、通过酶促(例如,蛋白水解)裂解和/或通过表达编码本公开内容的蛋白或片段的核酸通过重组来获得。本公开内容的蛋白和/或片段也可以通过化学合成或其他已知的产生蛋白和片段的方案获得。
100.术语“分离的多肽”是指在通过重组dna技术产生时大体上不含细胞物质或培养基的多肽,或者在化学合成时大体上不含化学前体或其他化学物质的多肽。
101.如本文使用的术语“载体”是指可以用于将核酸递送至细胞内部的多核苷酸。在一种实施方案中,载体是包含与待在细胞中表达的核酸可操作地连接的表达控制序列(例如,启动子)的表达载体。本领域已知的载体包括但不限于质粒、噬菌体、黏粒和病毒。
102.术语“受者”、“个体”、“受试者”、“宿主”和“患者”在本文可互换地使用,并且指期望诊断、治疗或疗法的任何哺乳动物受试者,特别地人类。为了治疗的目的,“哺乳动物”是指被归类为哺乳动物的任何动物,包括人类、家畜和农场动物,以及实验室动物、动物园动物、运动动物或宠物动物,诸如犬、马、猫、奶牛、绵羊、山羊、猪、小鼠、大鼠、兔、豚鼠、猴子等。在一些实施方案中,哺乳动物是人类。这些术语都不需要医务人员的监督。
103.如本文使用的,在一些实施方案中,术语“治疗(treatment)”、“治疗(treating)”等是指为了获得某种作用的目的而施用剂或进行程序。所述作用在完全或部分预防疾病或其症状方面可以是预防性的,和/或在影响部分或完全治愈疾病和/或疾病的症状方面可以是治疗性的。如本文使用的“治疗”可以包括哺乳动物中,特别地人类中疾病或病症的治疗(例如癌症),并且包括:(a)预防疾病或疾病的症状在可能易患该疾病但尚未被诊断患有该疾病(例如,包括可能与原发病相关或由原发病引起的疾病)的受试者中发生;(b)抑制疾病,即,阻止其发展;以及(c)缓解疾病,即,引起疾病消退。治疗可以指在治疗或改善或预防
癌症方面的任何成功征象,包括任何客观或主观的参数,诸如症状的减轻、缓解、减弱;或者使疾病状况对患者更耐受;减慢退化或衰退的速率;或者使退化的终点不那么衰弱。症状的治疗或改善基于一个或更多个客观或主观的参数;包括医师检查的结果。因此,术语“治疗”包括施用本发明的化合物或剂,以预防、延迟、缓解或阻滞或抑制与疾病(例如癌症)相关的症状或状况的发展。术语“治疗作用”是指降低、消除或预防受试者中的疾病、疾病的症状或疾病的副作用。
104.除非上下文另外清楚指明,如本文使用的单数形式"一(a)"、"一(an)"和"该(the)"包括复数指代物。因此,例如,提及“抗体”包括多于一种抗体,并且在一些实施方案中提及“抗体”包括多种抗体,等等。
105.除非上下文另外清楚指明,如本文使用的,所有数值或数值范围包括这样的范围内或涵盖这样的范围的全部整数以及范围内或涵盖范围的值或整数的分数。因此,例如,提及90%-100%的范围包括91%、92%、93%、94%、95%、95%、96%、97%等,以及91.1%、91.2%、91.3%、91.4%、91.5%等,92.1%、92.2%、92.3%、92.4%、92.5%等,等等。在另一个实例中,提及1倍-5,000倍的范围包括1倍、2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍、10倍、11倍、12倍、13倍、14倍、15倍、16倍、17倍、18倍、19倍、20倍等,以及1.1倍、1.2倍、1.3倍、1.4倍、1.5倍等,2.1倍、2.2倍、2.3倍、2.4倍、2.5倍等,等等。
106.如本文使用的,“约”数字,是指包括该数字并且范围从低于该数字10%至高于该数字10%的范围。“约”范围是指低于该范围下限10%,跨越至高于该范围上限10%。
[0107]“同一性百分比(%)”是指两个序列(核苷酸或氨基酸)在比对中的相同位置处具有相同残基的程度。例如,“氨基酸序列与seq id no:y x%相同”是指氨基酸序列与seq id no:y的同一性%并且详细说明为氨基酸序列中x%的残基与seq id no:y中公开的序列的残基相同。通常,应用计算机程序进行这样的计算。比较和比对序列对的示例性程序包括align(myers和miller,1988)、fasta(pearson和lipman,1988;pearson,1990)和带空位的blast(altschul等人,1997)、blastp、blastn或gcg(devereux等人,1984)。
[0108]
在理解本技术的范围时,如本文使用的术语“包含(comprising)”及其衍生词意图是开放式术语,其指定所陈述的特征、要素、组件、组、整数和/或步骤的存在,但不排除其他未陈述的特征、要素、组件、组、整数和/或步骤的存在。前述情况也适用于具有相似含义的词,诸如术语“包括(including)”、“具有(having)”及其衍生词。如本文使用的,术语“由...组成(consisting)”及其衍生词意图是封闭式术语,其指定所陈述的特征、要素、组件、组、整数和/或步骤的存在,但排除其他未陈述的特征、要素、组件、组、整数和/或步骤的存在。如本文使用的,术语“基本上由...组成(consisting essentially of)”意图指定所陈述的特征、要素、组件、组、整数和/或步骤以及那些不实质性影响特征、要素、组件、组、整数和/或步骤的一个或更多个基本和新颖特性的存在。
[0109]
如本文使用的,诸如“大体上”、“约”和“大约”的程度术语意指修饰的术语的合理偏差量,使得最终结果不被显著改变。这些程度术语应解释为如果这种偏差不会否定它所修饰的词的含义,则包括修饰的术语的至少
±
5%的偏差。
[0110]
如本文使用的术语“和/或”意味着所列出的项目单独地或组合地存在或使用。实际上,该术语意味着“至少一个”或“一个或更多个”所列项目被使用或存在。
[0111]
嵌合共刺激受体(ccr)
[0112]
在某些实施方案中,本文公开了嵌合共刺激受体(ccr)核酸,所述嵌合共刺激受体(ccr)核酸包含(a)编码肿瘤坏死因子受体超家族(tnfrsf)成员的胞外结构域的第一多核苷酸;(b)编码跨膜结构域多肽的第二多核苷酸;和(c)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸来源于tnfrsf的不同成员。
[0113]
在一些实施方案中,第一多核苷酸编码tnfrsf成员的胞外结构域。在一些实施方案中,第一多核苷酸和第三多核苷酸来源于tnfrsf的不同成员。在一些实施方案中,第一多核苷酸编码具有死亡结构域的tnfrsf成员的胞外结构域。在一些实施方案中,第一多核苷酸编码以下的胞外结构域:tnfr1、tnfr2、fas、dr4、dr5、dr3、dr6、edar、xedar、troy或ngfr。在一些实施方案中,第一多核苷酸编码tnfr1的胞外结构域。在一些实施方案中,第一多核苷酸编码tnfr2的胞外结构域。在一些实施方案中,第一多核苷酸编码fas的胞外结构域。在一些实施方案中,第一多核苷酸编码dr4的胞外结构域。在一些实施方案中,第一多核苷酸编码dr5的胞外结构域。在一些实施方案中,第一多核苷酸编码dr3的胞外结构域。在一些实施方案中,第一多核苷酸编码dr6的胞外结构域。在一些实施方案中,第一多核苷酸编码edar的胞外结构域。在一些实施方案中,第一多核苷酸编码xedar的胞外结构域。在一些实施方案中,第一多核苷酸编码troy的胞外结构域。在一些实施方案中,第一多核苷酸编码ngfr的胞外结构域。
[0114]
肿瘤坏死因子受体超家族(tnfrsf)是细胞因子受体的蛋白超家族,其特征在于通过胞外富含半胱氨酸的结构域结合肿瘤坏死因子(tnf)的能力。tnfr超家族存在27个成员,包括tnfr1、tnfr2、fas、dr4、dr5、dr3、dr6、edar、xedar、troy或ngfr。
[0115]
肿瘤坏死因子受体1(tnfr1),又称为肿瘤坏死因子受体超家族成员1a(tnfrsf1a)和cd120a,是结合肿瘤坏死因子α(tnfα)的膜结合受体。tnfr1活化转录因子nf-κb,介导凋亡,并且作为炎症的调节物发挥功能。
[0116]
肿瘤坏死因子受体2(tnfr2),又称为肿瘤坏死因子受体超家族成员1b(tnfrsf1b)和cd120b,是结合肿瘤坏死因子α(tnfα)的膜结合受体。
[0117]
fas受体,又称为fas、fasr、凋亡抗原1(apo-1或apt)、分化簇95(cd95)或肿瘤坏死因子受体超家族成员6(tnfrsf6),是人类中由fas基因编码的蛋白。fas的多个剪接变体已经被鉴定出来,它们被翻译为该蛋白的七种同种型。诱导凋亡的fas受体被称为同种型1,并且是一种1型跨膜蛋白。许多其他同种型是通常与疾病状态相关的罕见单体型。预期任何合适的fas同种型用于与本文公开的实施方案一起使用。
[0118]
死亡结构域4(dr4),又称为trail受体1(trailr1)和肿瘤坏死因子受体超家族成员10a(tnfrsf10a),是tnf受体超家族中结合trail并介导凋亡的细胞表面受体。
[0119]
死亡结构域5(dr5),又称为trail受体2(trailr2)和肿瘤坏死因子受体超家族成员10b(tnfrsf10b),是tnf受体超家族中结合trail并介导凋亡的细胞表面受体。
[0120]
死亡结构域3(dr3),又称为肿瘤坏死因子受体超家族成员25(tnfrsf25),是肿瘤坏死因子受体超家族中介导凋亡信号传导和分化的细胞表面受体。其唯一已知的tnfsf配体是tnf样蛋白1a(tl1a)。
[0121]
死亡结构域6(dr6),又称为肿瘤坏死因子受体超家族成员21(tnfrsf21),是肿瘤坏死因子受体超家族中活化jnk和nf-κb途径的细胞表面受体。
[0122]
外胚层发育不良因子受体(edar)是tnf受体超家族的成员。它在外胚层分化过程中发挥关键作用。
[0123]
外胚叶发育不良因子a2受体(xedar;肿瘤坏死因子受体超家族成员27)是人类中由eda2r基因编码的蛋白。eda-a1和eda-a2是由无汗性外胚层发育不良因子(eda)基因编码的外胚叶发育不良素(ectodysplasin)的两种同种型。
[0124]
troy(肿瘤坏死因子受体超家族成员19,tnfrsf19)是tnf受体超家族的成员。该受体在胚胎发育期间高度表达。已知,它与tnf受体相关因子(traf)家族成员相互作用,并且在细胞中过表达时活化c-jun n末端激酶(jnk)信号传导途径。该受体能够通过不依赖于胱天蛋白酶的机制诱导凋亡,并且被认为在胚胎发育中发挥重要作用。
[0125]
ngfr(低亲和力神经生长因子受体;神经生长因子受体;tnfr超家族成员16;lngfr;p75神经营养因子受体)是肿瘤坏死因子受体(tnf受体)超家族的成员。它是神经营养因子的两种受体类型之一,是刺激神经元细胞存活和分化的蛋白生长因子家族。
[0126]
在一些实施方案中,第一多核苷酸编码具有根据seq id no:1的序列的寡肽。在一些实施方案中,多核苷酸编码具有与根据seq id no:1的序列有至少80%、85%、90%、95%或99%同一性的序列的寡肽。在一些实施方案中,第一多核苷酸编码具有根据seq id no:2的序列的寡肽。在一些实施方案中,多核苷酸编码具有与根据seq id no:2的序列有至少80%、85%、90%、95%或99%同一性的序列的寡肽。
[0127][0128]
在一些实施方案中,第三多核苷酸编码来自以下的胞质共刺激信号传导结构域多肽:cd28、cd28t,ox-40、4-1bb/cd137、cd2、cd7、cd27、cd30、cd40、程序性死亡-1(pd-l)、诱导型t细胞共刺激物(icos)、淋巴细胞功能相关抗原-l(lfa-l、cd1-cd18)、cd247、cd276(b7-h3)、light(tnfsf14)、nkg2c、igα(cd79a)、dap-10、icam-l、b7-h3、cds、icam-l、gitr、baffr、light、hvem(lightr)、kirds2、slamf7、nkp80(klrf1)、nkp44、nkp30、nkp46、cd19、cd4、cd8α、cd8β、il-2rβ、il-2rγ、il-7rα、itga4、vla1、cd49a、itga4、ia4、cd49d、itga6、vla-6、cd49f、itgad、cd1 ld、itgae、cd 103、itgal、cd1 la、lfa-l、itgam、cd1 lb、itgax、
cd1 lc、itgb1、cd29、itgb2、cd18、lfa-l、itgb7、nkg2d、tnfr2、trance/rankl、dnam1(cd226)、slamf4(cd244、2b4)、cd84、cd96(tactile)、ceacam1、crt am、ly9(cd229)、cd 160(by55)、psgl1、cd100(sema4d)、cd69、slamf6(ntb-a、lyl08)、slam(slamf1、cd150、ipo-3)、blame(slamf8)、selplg(cd162)、ltbr、lat、gads、slp-76、pag/cbp或cdl9a。
[0129]
在一些实施方案中,第三多核苷酸编码来自以下的胞质共刺激信号传导结构域:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第三多核苷酸编码来自baffr的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自4-1bb的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自cd27的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自hvem的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自ox40的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自gitr的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自taci的胞质共刺激信号传导结构域。
[0130]
在一些实施方案中,第三多核苷酸编码具有根据seq id no:3的序列的寡肽。在一些实施方案中,多核苷酸编码具有与根据seq id no:3的序列有至少80%、85%、90%、95%或99%同一性的序列的寡肽。在一些实施方案中,第三多核苷酸编码具有根据seq id no:4的序列的寡肽。在一些实施方案中,多核苷酸编码具有与根据seq id no:4的序列有至少80%、85%、90%、95%或99%同一性的序列的寡肽。
[0131][0132]
在一些实施方案中,第二多核苷酸编码来自tnfr超家族成员的跨膜结构域。在一些实施方案中,第二多核苷酸编码来自以下的跨膜结构域:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,跨膜结构域与胞质共刺激信号传导结构域来源于不同的分子。在一些实施方案中,跨膜结构域与胞质共刺激信号传导结构域来源于同一分子。在一些实施方案中,跨膜结构域和胞质共刺激信号传导结构域来源于选自以下的同一共刺激分子:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy和tweak。
[0133]
在一些实施方案中,第二多核苷酸编码具有根据seq id no:5的序列的寡肽。在一些实施方案中,多核苷酸编码具有与根据seq id no:5的序列有至少80%、85%、90%、95%或99%同一性的序列的寡肽。在一些实施方案中,第二多核苷酸编码具有根据seq id no:6的序列的寡肽。在一些实施方案中,多核苷酸编码具有与根据seq id no:6的序列有至少80%、85%、90%、95%或99%同一性的序列的寡肽。
[0134]
seq id结构域名称aa序列seq id no:5tnfr1 tm结构域vllplviffglcllsllfigl
seq id no:6fas受体tm结构域lgwlcllllpiplivwv
[0135]
单分子
[0136]
在一些实施方案中,ccr是单分子。在一些实施方案中,第一多核苷酸和第二多核苷酸被直接或间接(例如,通过接头)连接至第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸被直接或间接(例如,通过接头)连接至第二多核苷酸。预期任何合适的接头用于与本文公开的分子一起使用。在一些实施方案中,接头是小分子。在一些实施方案中,接头是肽接头。
[0137]
在一些实施方案中,ccr具有根据seq id no:7的氨基酸序列。在一些实施方案中,多核苷酸编码具有与根据seq id no:7的序列有至少80%、85%、90%、95%或99%同一性的序列的寡肽。在一些实施方案中,ccr具有根据seq id no:8的氨基酸序列。在一些实施方案中,多核苷酸编码具有与根据seq id no:8的序列有至少80%、85%、90%、95%或99%同一性的序列的寡肽。在一些实施方案中,ccr具有根据seq id no:9的氨基酸序列。在一些实施方案中,多核苷酸编码具有与根据seq id no:9的序列有至少80%、85%、90%、95%或99%同一性的序列的寡肽。在一些实施方案中,ccr具有根据seq id no:10的氨基酸序列。在一些实施方案中,多核苷酸编码具有与根据seq id no:10的序列有至少80%、85%、90%、95%或99%同一性的序列的寡肽。
[0138][0139]
免疫细胞
[0140]
在某些实施方案中,本文公开了包含嵌合共刺激受体(ccr)核酸的免疫细胞,所述嵌合共刺激受体(ccr)核酸包含(a)编码肿瘤坏死因子受体超家族成员的胞外结构域的第一多核苷酸;(b)编码跨膜结构域多肽的第二多核苷酸;和(c)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸来源于tnfrsf的不同成员。在一些实施方案中,第一多核苷酸编码具有死亡结构域的tnfrsf成员的胞外结构域。在一些实施方案中,第一多核苷酸编码以下的胞外结构域:tnfr1、tnfr2、fas、dr4、dr5、dr3、dr6、edar、xedar、troy或ngfr。在一些实施方案中,第三多核苷酸编码来自以下的胞质共刺激信号传导结构域多肽:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,免疫细胞是t细胞(例如,细胞毒性t细胞、辅助性t细胞、调节性t细胞或γ-δt细胞)。在一些实施方案中,t细胞是工程化t细胞。在一些实施方案中,免疫细胞是自然杀伤细胞(nk细胞)。在一些实施方案中,免疫细胞是巨噬细胞。在一些实施方案中,免疫细胞是肿瘤
浸润淋巴细胞(til)。在一些实施方案中,免疫细胞是单核细胞。在一些实施方案中,免疫细胞是b细胞。
[0141]
t细胞
[0142]
在某些实施方案中,本文公开了包含嵌合共刺激受体(ccr)核酸的t细胞,所述嵌合共刺激受体(ccr)核酸包含(a)编码肿瘤坏死因子受体超家族成员的胞外结构域的第一多核苷酸;(b)编码跨膜结构域多肽的第二多核苷酸;和(c)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸来源于tnfrsf的不同成员。在一些实施方案中,第一多核苷酸编码具有死亡结构域的tnfrsf成员的胞外结构域。在一些实施方案中,第一多核苷酸编码以下的胞外结构域:tnfr1、tnfr2、fas、dr4、dr5、dr3、dr6、edar、xedar、troy或ngfr。在一些实施方案中,第三多核苷酸编码来自以下的胞质共刺激信号传导结构域多肽:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。
[0143]
在一些实施方案中,t细胞是细胞毒性t细胞、辅助性t细胞、调节性t细胞或γ-δt细胞。在一些实施方案中,t细胞包含编码可识别靶特异性配体的工程化t细胞受体(tcr)或合成抗原受体多肽的第二核酸。在一些实施方案中,合成抗原受体多核苷酸是嵌合抗原受体(car)。在一些实施方案中,合成抗原受体多核苷酸编码t细胞抗原偶联剂(tac)。
[0144]
在一些实施方案中,car包含抗原结合结构域、跨膜结构域和胞内信号传导结构域(例如,包括共刺激结构域和/或初级信号传导结构域的胞内信号传导结构域)。在一些实施方案中,跨膜结构域是选自由以下组成的组的跨膜结构域:t细胞受体的α或β、cd28、cd3ε、cd3δ、cd45、cd4、cd5、cd8、cd9、cd16、cd22、cd33、cd37、cd64、cd80、cd86、cd134、cd137和cd154。在一些实施方案中,胞内信号传导结构域是来源于以下的信号传导结构域:cd28、cd28t,ox-40、4-1bb/cd137、cd2、cd7、cd27、cd30、cd40、程序性死亡-1(pd-l)、诱导型t细胞共刺激物(icos)、淋巴细胞功能相关抗原-l(lfa-l、cd1-cd18)、cd3γ、cd3δ、cd3ε、cd247、cd276(b7-h3)、light(tnfsf14)、nkg2c、igα(cd79a)、dap-10、fcγ受体、mhc 1类分子、tnf受体蛋白、免疫球蛋白、细胞因子受体、整联蛋白、信号传导淋巴细胞活化分子(slam蛋白)、活化nk细胞受体、btla、toll配体受体、icam-l、b7-h3、cds、icam-l、gitr、baffr、light、hvem(lightr)、kirds2、slamf7、nkp80(klrf1)、nkp44、nkp30、nkp46、cd19、cd4、cd8α、cd8β、il-2rβ、il-2rγ、il-7rα、itga4、vla1、cd49a、itga4、ia4、cd49d、itga6、vla-6、cd49f、itgad、cd1 ld、itgae、cd 103、itgal、cd1 la、lfa-l、itgam、cd1 lb、itgax、cd1 lc、itgb1、cd29、itgb2、cd18、lfa-l、itgb7、nkg2d、tnfr2、trance/rankl、dnam1(cd226)、slamf4(cd244、2b4)、cd84、cd96(tactile)、ceacam1、crt am、ly9(cd229)、cd 160(by55)、psgl1、cd100(sema4d)、cd69、slamf6(ntb-a、lyl08)、slam(slamf1、cd150、ipo-3)、blame(slamf8)、selplg(cd162)、ltbr、lat、gads、slp-76、pag/cbp或cdl9a。
[0145]
在一些实施方案中,tac包含抗原结合结构域、结合与tcr复合物缔合的蛋白的结构域以及包含胞质结构域和跨膜结构域的t细胞共受体结构域。在一些实施方案中,与tcr复合物缔合的蛋白是cd3。在一些实施方案中,tac不包含共刺激结构域和/或活化结构域。在一些实施方案中,胞质结构域是cd4胞质结构域,并且跨膜结构域是cd4跨膜结构域。在一些实施方案中,tac包含抗原结合结构域、cd3结合结构域以及cd4胞质结构域和cd4跨膜结构域。在一些实施方案中,cd3抗原结合结构域来源于ucht1。
[0146]
在一些实施方案中,工程化tcr是t细胞受体(tcr)融合蛋白(tfp)。在一些实施方案中,tfp包含至少一个工程化cd3链,该工程化cd3链包含(a)胞外结构域的至少一部分、(b)包含来自胞内信号传导结构域的刺激结构域的胞内结构域;和(c)抗原结合域(例如,scfv)。在一些实施方案中,胞外结构域和胞内结构域来源于cd3α、cd3β、cd3γ、cd3δ或cd3ε。在一些实施方案中,工程化cd3链还包含跨膜结构域。在一些实施方案中,胞外结构域、跨膜结构域和胞内结构域来源于cd3ε。在一些实施方案中,工程化cd3链取代tcr的至少一个天然存在的cd3ε链。
[0147]
在一些实施方案中,工程化tcr是嵌合抗体-t细胞受体(tcr)构建体(catcr)。在一些实施方案中,catcr包含与靶抗原特异性结合的抗原结合模块和能够募集至少一种tcr相关信号传导分子的t细胞受体模块(tcrm)。在一些实施方案中,tcrm包括包含第一tcr跨膜结构域(tcr-tm)的第一tcr结构域(tcrd)和包含第二tcr-tm的第二tcrd,其中tcr-tm促进至少一种tcr相关信号传导分子的募集。在一些实施方案中,第一tcr-tm来源于t细胞受体的跨膜结构域之一(诸如αβtcr,或γδtcr),并且第二tcr-tm来源于t细胞受体的另一跨膜结构域。在一些实施方案中,tcr是αβtcr,并且第一tcr-tm和第二tcr-tm来源于tcrα和β亚基跨膜结构域。在一些实施方案中,tcr是γδtcr,并且第一tcr-tm和第二tcr-tm来源于tcrγ和δ亚基跨膜结构域。在一些实施方案中,tcrd和tcr-tm融合至来源于抗体的f(ab)。在一些实施方案中,tcrd和tcr-tm融合至来源于抗体的f(v)部分的单链抗体(scfv)。
[0148]
在一些实施方案中,抗原结合结构域结合癌细胞上的抗原。在一些实施方案中,抗原在癌细胞上差异地表达。在一些实施方案中,抗原在癌细胞上是上调的。在一些实施方案中,抗原是表面抗原,并且不由mhc呈递。在一些实施方案中,抗原是mhc/肽复合物。
[0149]
在一些实施方案中,抗原是her2(erbb-2)、b细胞成熟抗原(bcma)、cd19、甲胎蛋白(afp)、癌胚抗原(cea)、ca-125、muc-1、上皮肿瘤抗原(eta)、酪氨酸酶、黑素瘤相关抗原(mage)、前列腺特异性抗原(psa)、胶质瘤相关抗原、β-人类绒毛膜促性腺激素、甲状腺球蛋白、rage-1、mn-ca ix、人类端粒酶逆转录酶、ru1、ru2(as)、肠羧基酯酶、mut hsp70-2、m-csf、prostase、pap、ny-eso-1、lage-1a、p53、prostein、psma、存活蛋白和端粒酶、前列腺癌肿瘤抗原-1(pcta-1)、elf2 m、中性粒细胞弹性蛋白酶、cd22、胰岛素生长因子(igf)-i、igf-ii、igf-i受体和间皮素。在一些实施方案中,抗原是her2、bcma或cd19。在一些实施方案中,抗原是her2。在一些实施方案中,抗原是bcma。在一些实施方案中,抗原是cd19。
[0150]
使用方法
[0151]
在某些实施方案中,本文公开了治疗有相应需要的个体中的癌症的方法,所述方法包括向个体施用包含嵌合共刺激受体(ccr)核酸的免疫细胞,所述嵌合共刺激受体(ccr)核酸包含(a)编码肿瘤坏死因子受体超家族成员的胞外结构域的第一多核苷酸;(b)编码跨膜结构域多肽的第二多核苷酸;和(c)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸来源于tnfrsf的不同成员。在一些实施方案中,第一多核苷酸编码具有死亡结构域的tnfrsf成员的胞外结构域。在一些实施方案中,第一多核苷酸编码以下的胞外结构域:tnfr1、tnfr2、fas、dr4、dr5、dr3、dr6、edar、xedar、troy或ngfr。在一些实施方案中,第三多核苷酸编码来自以下的胞质共刺激信号传导结构域多肽:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,免疫细胞是t细
胞(例如,细胞毒性t细胞、辅助性t细胞、调节性t细胞或γ-δt细胞)。在一些实施方案中,t细胞是工程化t细胞。在一些实施方案中,免疫细胞是自然杀伤细胞(nk细胞)。在一些实施方案中,免疫细胞是巨噬细胞。在一些实施方案中,免疫细胞是肿瘤浸润淋巴细胞(til)。在一些实施方案中,免疫细胞是单核细胞。在一些实施方案中,免疫细胞是b细胞。
[0152]
在一些实施方案中,癌症是实体癌症或液体癌症(liquid cancer)。在一些实施方案中,癌症是上皮癌(carcinoma)、母细胞瘤、黑素瘤、肉瘤、血液学癌症或淋巴恶性肿瘤。
[0153]
在一些实施方案中,癌症是白血病或淋巴瘤。在一些实施方案中,癌症是混合谱系白血病(mll)、慢性淋巴细胞白血病(cll)、急性淋巴母细胞白血病(all)、大b细胞淋巴瘤、弥漫性大b细胞淋巴瘤、原发性纵隔b细胞淋巴瘤、高级别b细胞淋巴瘤或源于滤泡性淋巴瘤的大b细胞淋巴瘤。
[0154]
在一些实施方案中,癌症是肺癌、乳腺癌、结肠癌、多发性骨髓瘤、胶质母细胞瘤、胃癌、卵巢癌、胃癌、结肠直肠癌、尿路上皮癌、子宫内膜癌或黑素瘤。在一些实施方案中,癌症是肺癌。在一些实施方案中,癌症是乳腺癌。在一些实施方案中,癌症是结肠癌。在一些实施方案中,癌症是多发性骨髓瘤。在一些实施方案中,癌症是胶质母细胞瘤。在一些实施方案中,癌症是胃癌(gastric cancer)。在一些实施方案中,癌症是卵巢癌。在一些实施方案中,癌症是胃癌(stomach cancer)。在一些实施方案中,癌症是结肠直肠癌。在一些实施方案中,癌症是尿路上皮癌。在一些实施方案中,癌症是子宫内膜癌。在一些实施方案中,癌症是黑素瘤。
[0155]
药物组合物
[0156]
在某些实施方案中,本文公开了药物组合物,所述药物组合物包含(a)免疫细胞,所述免疫细胞包含(i)编码肿瘤坏死因子受体超家族成员的胞外结构域的第一多核苷酸;(ii)编码跨膜结构域多肽的第二多核苷酸;和(iii)编码胞质共刺激信号传导结构域多肽(例如,来自肿瘤坏死因子受体超家族(tnfrsf)成员)的第三多核苷酸;以及(b)药学上可接受的载体。在一些实施方案中,第一多核苷酸和第三多核苷酸来源于tnfrsf的不同成员。在一些实施方案中,第一多核苷酸编码具有死亡结构域的tnfrsf成员的胞外结构域。在一些实施方案中,第一多核苷酸编码以下的胞外结构域:tnfr1、tnfr2、fas、dr4、dr5、dr3、dr6、edar、xedar、troy或ngfr。在一些实施方案中,第一多核苷酸编码tnfr1的胞外结构域。在一些实施方案中,第一多核苷酸编码tnfr2的胞外结构域。在一些实施方案中,第一多核苷酸编码fas的胞外结构域。在一些实施方案中,第三多核苷酸编码来自以下的胞质共刺激信号传导结构域:4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第三多核苷酸编码来自baffr的胞质共刺激信号传导结构域。在一些实施方案中,第三多核苷酸编码来自4-1bb的胞质共刺激信号传导结构域。在一些实施方案中,跨膜结构域和胞质共刺激信号传导结构域来自同一共刺激信号传导蛋白。在一些实施方案中,跨膜结构域和共刺激信号传导结构域来源于4-1bb、baffr、ox40、cd27、cd40、gitr、hvem、ox40、relt、taci、troy或tweak。在一些实施方案中,第一多核苷酸和第二多核苷酸被直接或间接(例如,通过接头)连接至第三多核苷酸。在一些实施方案中,第一多核苷酸和第三多核苷酸被直接或间接(例如,通过接头)连接至第二多核苷酸。在一些实施方案中,免疫细胞是t细胞(例如,细胞毒性t细胞、辅助性t细胞、调节性t细胞、γ-δt细胞)、自然杀伤细胞(nk细胞)、巨噬细胞、肿瘤浸润淋巴细胞(til)、单核细胞或b细胞。
[0157]
本文公开的药物组合物通过本身已知的制备向受试者施用的药学上可接受的组合物的方法进行制备,使得有效量的免疫细胞与药学上可接受的载体组合在混合物中。例如,合适的载体描述于remington'spharmaceutical sciences(remington's pharmaceutical sciences,第20版,mack publishing company,easton,pa.,usa,2000)中。在此基础上,组合物包含(尽管不仅仅)与一种或更多种药学上可接受的载体或稀释剂联合的物质的溶液,并且被包含在具有合适ph和与生理流体等渗的缓冲溶液中。
[0158]
合适的药学上可接受的载体包括不干扰药物组合物的生物活性的有效性的基本化学惰性和无毒的组合物。合适的药物载体的实例包括但不限于,水、盐水溶液、甘油溶液、n-(l(2,3-二油烯基氧基)丙基)n,n,n-三甲基氯化铵(dotma)、二油烯基磷酸酯基-乙醇胺(diolesylphosphotidyl-ethanolamine,dope)和脂质体。在一些实施方案中,这样的组合物包含治疗有效量的化合物连同合适量的载体,以提供向患者直接施用的形式。
[0159]
药物组合物以适于待治疗(或预防)的疾病的方式施用。施用的数量和频率由诸如患者的状况以及患者疾病的类型和严重程度等因素确定,尽管适当的剂量由临床试验确定。在指示“免疫有效量”、“抗肿瘤有效量”、“肿瘤抑制有效量”或“治疗有效量”时,待施用的本发明组合物的精确量由医师考虑年龄、体重、肿瘤尺寸、感染或转移程度以及患者(受试者)状况的个体差异来确定。
[0160]
药物组合物“大体上不含”指示,例如,不存在可检测水平的例如选自由以下组成的组的污染物:内毒素、支原体、有复制能力的慢病毒(rcl)、p24、vsv-g核酸、hiv gag、剩余抗cd3/抗cd28涂覆珠、小鼠抗体、汇集的人类血清、牛血清白蛋白、牛血清、培养基组分、载体包装细胞或质粒组分、细菌、真菌、支原体、il-2和il-7。
[0161]
在一些实施方案中,药物组合物以单次或多次,例如,每天、每周、每两周或每月、每小时施用,或者在被治疗的癌症重现(recurrence)、复发(relapse)或进展时施用。
[0162]
在一些实施方案中,本文公开的药物组合物通过任何合适的方法,包括但不限于静脉内或通过输注施用。在一些实施方案中,本文公开的药物组合物使用免疫疗法中通常已知的输注技术施用。在一些实施方案中,本文公开的药物组合物被直接注射到肿瘤、淋巴结或感染部位中。
实施例
[0163]
以下非限制性实施例用于说明本技术:
[0164]
实施例1.嵌合共刺激受体
[0165]
如先前陈述的,共刺激和共抑制与tcr信号传导并行作用,以调节t细胞的活化、增殖和持久性。通过tnfrsf信号传导调节t细胞响应对t细胞疗法是有吸引力的,因为每个受体都赋予独特的生物学作用。此外,鉴于tnfrsf在结构生物学上的相似性,通过产生基于tnfrsf的嵌合共刺激受体(ccr),通过tnfrsf将共抑制(即抑制性信号传导)重定向于共刺激(即刺激性信号传导)应该是可能的。事实上,先前的报道已经确定了,tnfrsf成员具有可以被交换以重定向信号传导的模块结构域。
[0166]
抗肿瘤t细胞产生高水平的tnfα,但实验数据表明,天然的tnfα信号传导限制t细胞免疫。tnfr1介导的tnfα信号传导引起细胞死亡,限制了t细胞的扩增和功能。因此,令人感兴趣的是,考虑通过嵌合tnfr1重定向tnfα生物学,该嵌合tnfr1在tnfα响应于抗原刺激
4-1bb ccr转导的jurkat细胞揭示出iκb的时间依赖性降解;与亲本非工程化(wt)jurkat细胞相比,降解速率增强,展示出tnfr1-4-1bb ccr的强有力的作用(图4)。具有截短的非信号传导胞质结构域的tnf-阻断剂受体消除了tnfα刺激的iκbα降解(图4),表明tnfr1-4-1bb ccr对iκb的增强的降解是由于胞质结构域。由于野生型tnfr1的毒性作用,野生型tnfr1在jurkat细胞中不能过表达。
[0174]
已经显示出4-1bb信号传导活化mapk信号传导途径。p38 mapk的磷酸化和活化导致下游转录因子的活化。评价tnfr1-4-1bb ccr和tnf-阻断剂工程化jurkat细胞的tnfα刺激后的磷酸化p38蛋白(phospho-p38protein)水平。用tnfα刺激诱导了磷酸化p38蛋白水平在非工程化wt细胞和用tnfr1-41bb ccr工程化的细胞二者中的产生。磷酸化p38水平在刺激后5min达到其峰。与非工程化(wt)jurkat细胞相比,用tnfr1-4-1bb ccr工程化的jurkat细胞中峰响应更高(图5)。与用iκb观察的结果相似,tnf-阻断剂受体tnf-阻断剂受体的表达消除了p38磷酸化(图5)。
[0175]
这些结果清楚地展示出在t细胞中tnfr1-4-1bb ccr能够将tnfα信号传导重定向于增强的nfκb信号传导。
[0176]
使用第三代自失活慢病毒表达系统工程化原代t细胞以稳定地表达图2a-图2c中的受体之一。对转导的t细胞通过染色和流式细胞术评价受体的表面表达。tnfr1-4-1bb ccr的表达在cd8 和cd4 t细胞子集二者中均是明显的,其中在cd4 t细胞中注意到较高的转导率(图6a-图6b)。在活化和转导t细胞后,对原代培养物的生长进行追踪达14天。tnfr1-4-1bb ccr在t细胞中的表达不影响细胞的体外生长,表明转导的t细胞对tnfr1-4-1bb ccr的引入耐受(图7)。
[0177]
已显示出tnfrsf共刺激增强t细胞中的存活信号传导,以改进持久性。预期由tnfr1-4-1bb ccr提供的4-1bb共刺激向转导的t细胞提供增强的存活特征。将原代人类t细胞用tnfr1-4-1bb ccr转导,并且在il2和il7生长因子中生长14天。在第14天,将细胞从细胞因子中取出,并且用信号1(抗cd3)或信号2(tnfα)刺激,并且对活细胞数目进行追踪持续4天。从t细胞中去除生长因子导致对照t细胞培养物的凋亡和死亡。然而,用tnfr1-4-1bb ccr转导的t细胞在两种刺激条件下都显示出更好的存活(图8a-8b),表明tnfr1-4-1bb ccr的tnfα刺激增强工程化t细胞的存活信号传导,上调抗凋亡途径。这些数据也表明抗cd3刺激的t细胞分泌足够水平的tnfα,以刺激tnfr1-4-1bb ccr存活信号传导。
[0178]
tnfr1-4-1bb ccr转导的t细胞在信号1(抗cd3)刺激下变成活化的,以分泌tnfα和ifnγ二者。与对照细胞相比,ccr t细胞的细胞因子谱朝向ifnγ产生偏倚(图9)。
[0179]
鉴于tnfr1-4-1bb ccr的有希望的功能,然后确定ccr的表达是否会通过合成抗原受体在t细胞活化的背景下提供共刺激活性。为此,将ccr与对bcma有特异性的tac受体(bcma-tac)共表达。构建表达tac受体和由2a肽分开的tnfr1-4-1bb ccr或tnf-阻断剂的慢病毒(图10)。
[0180]
为了确保bcma-tac和tnfr1-4-1bb ccr在2a表达系统下的表达不干扰tac的功能,进行功能测定以评价抗肿瘤活性。将tac和tac ccr工程化t细胞与表达萤光素酶的bcma阳性kms11细胞共培养24小时。杀伤活性通过kms11靶的发光降低来测量。tac ccr t细胞显示出与tac t细胞相似的杀伤活性(图11),表明2a表达系统导致功能受体表达。由于t细胞活化和细胞毒性通过单独的tac受体介导,预期由4-1bb-ccr提供的另外的信号不影响体外杀
伤测定。
[0181]
活化的t细胞通过几种可溶性介导物(mediators)包括分泌的细胞因子来介导细胞毒性和促炎信号传导。与抗原阳性肿瘤细胞靶共培养后,在体外胞内细胞因子测定中评价ifnγ和tnfα的表达。在抗原衔接(antigen engagement)后,tac-工程化t细胞和tac ccr-工程化t细胞二者都表达ifnγ和tnfα。有趣的是,在cd4 和cd8 tac ccr t细胞二者中存在观察到的细胞因子表达降低(图12)。在临床上,使用car工程化t细胞表现出对患者的高毒性风险,部分通过细胞因子风暴(cytokine storm)介导。抗原衔接后t细胞分泌的细胞因子连同ccr表达的降低可能是减轻car t细胞毒性同时维持抗肿瘤活性的有效手段。
[0182]
为了确定ccr表达在tac工程化t细胞中的功能性结果,在与抗原阳性的肿瘤靶共培养后,进行增殖测定以追踪t细胞分裂。在抗原衔接和t细胞活化后,t细胞经历快速分裂和增殖,产生子细胞以介导抗肿瘤活性。在增殖测定中比较用单独的tac、tac ccr和tac tnf-阻断剂工程化的t细胞。在7天增殖测定后,用4-1bb-ccr工程化的cd4 和cd8 tac t细胞二者都显示出增强的增殖(图13a-13b)。与cd8 t细胞相比,cd4 t细胞中作用更明显。与单独的tac相比,用tnf-阻断剂工程化的t细胞负面地影响增殖。数据表明,ccr的表达是增强tac t细胞增殖能力的介导信号。比较tac与tac ccr工程化之间的增殖和分裂指数,观察到tac ccr增强tac t细胞的增殖。这些值还揭示,平均而言,tac ccr产生比单独的tac t细胞多30%-50%的正增殖的细胞。
[0183]
tnfrsf包含已知共刺激t细胞的能够增强增殖、存活和记忆发育的数种分子,包括4-1bb、ox40和cd27。鉴于原型tnfr1-4-1bb ccr的有希望的结果,试图确定其他共刺激结构域是否会具有同等或更大的活性。另选的tnfrsf共刺激信号可以诱导具有独特特征的tac工程化t细胞以改进抗肿瘤活性。选择18种胞内结构域进行评价(列于表1中),并且设计了一系列tnfrsf ccr,其中跨膜结构域来源于tnfr1或各自的tnfrsf(如表1中示出的)。
[0184]
为了筛选tnfrsf ccr的实用性,评价ccr增强工程化t细胞在用板结合的抗cd3(okt3)刺激后增殖的能力。
[0185]
为了确认筛选的实用性,将t细胞用原始tnfr1-4-1bb、tnf-阻断剂或对照受体(截短的ngfr)工程化,并且用板结合的抗cd3活化(图14)。与先前的数据一致,筛选鉴定了相对于对照(截短的ngfr)的tnfr1-4-1bb ccr的增殖增强和tnf-阻断剂的抑制作用。因此,板结合的cd3是筛选tnfrsf共刺激受体的有效测定。
[0186]
表1中描述的合成tnfr1-ccr的cdna通过genscript合成并亚克隆到慢病毒载体中。将用抗cd3/抗cd28珠刺激的pbmc用ccr进行慢病毒工程化,并且在il2和il7中生长14天,在该点,评价t细胞的总体生长、工程化效率、受体表达和用板结合的抗cd3刺激后的增殖。人类pbmc的工程化效率范围(用ccr)为15%-85%,如通过包含在所有慢病毒中的转导标志物(ngfr)评价的(图15)。将工程化细胞用针对胞外tnfr1结构域的抗体进行染色,以评价表面上受体的量。ccr的表面表达范围为高于背景的0%-1500%(图16)。主体ccr工程化培养物的倍数扩增范围为对照的50%-450%(图17)。第14天工程化t细胞在抗cd3刺激后的5天增殖测定中进行评价。ccr工程化t细胞在cd4(图18)和cd8(图19)t细胞子集二者中的增殖范围为1(无增殖)至15(由单细胞产生平均15个细胞)。ccr增殖筛选的阳性击中阈值设为未收到共刺激信号的ngfr转导对照的增殖指数。用ccr工程化的cd4 t细胞的增殖显示出比对cd8 t细胞的增殖更明显的作用。最具有增殖性的ccr cd4细胞比ngfr工程化细胞增殖多
75%,而最具有增殖性的ccr cd8 t细胞比对应的ngfr工程化细胞增殖多60%。
[0187]
来自设计的构建体的多因素筛选结果使用聚类算法来分析,以评价ccr工程化t细胞之间的相似性。聚类算法中包含的数据是表面表达、工程化效率和cd4 和cd8 t细胞子集二者的增殖。数据维数(data dimensionality)首先使用主成分分析来降低,并且中心使用k-均值聚类方法来计算。相关热图描述了构建体之间的相似性(图20)。树状图的分组鉴定具有相似属性的受体。上组包含4-1bb和baffr ccr,所述4-1bb和baffr ccr已经显示出更高的表面表达、改进的培养物生长和增强的增殖。中间分组包含ngfr转导的对照,该对照无ccr表达、对生长的影响最小、对增殖无增强。中间组还包含原始4-1bb ccr,所述4-1bb ccr具有很少的ccr表达、对生长的改进最小以及对增殖的一定增强。下组包含tnf-阻断剂、light和fas共刺激结构域,其已展示出减慢培养物生长并且负面地影响增殖。有趣的是,这些组基于多参数聚类形成,这提供了对共刺激结构域的洞察,并且突出了上组中的从增殖筛选的阳性击中聚类在一起的成员。虽然该组中不是所有受体都提供对增殖的增强,但它们可能仍然值得追踪。共刺激受体不仅增强增殖,还增强细胞毒性功能和存活以及持久性。该共刺激分组中的受体可能具有除增殖之外的其他共刺激特性。
[0188]
对由聚类中心产生的tnfrsf ccr分组进行评价,揭示了具有可期望功能属性的工程化t细胞。具有4-1bb和baffr共刺激信号传导结构域的ccr显示出改进的pbmc扩增、高工程化效率和改进的增殖能力。为了验证4-1bb和baffr结构域的实用性,将tnfr1-4-1bb和tnfr-baffr构建体在增殖测定中以3个pbmc供体运行(图21a-图21b)。所选择的构建体的表现与筛选结果可比较,显示出相比于对照受体增强的增殖。这些数据提供了筛选数据的置信度,并且确认使用4-1bb和baffr信号传导结构域用于共刺激工程化t细胞的潜力。细胞毒性t细胞的重要功能是活化后分泌细胞因子。评价tnfr1-4-1bb和tnfr-baffr工程化t细胞在抗cd3刺激后的细胞因子产生(图22a-图22b),其中观察到可比较水平的ifnγ产生细胞,但观察到减少水平的tnfα产生细胞,可能是因为tnfα与ccr结合。
[0189]
讨论
[0190]
t细胞活化是复杂的、良好协调的信号传导级联的结果。tcr复合物、共受体、衔接体分子的完全募集以及共刺激和共抑制性分子的整合导致受体信号强度的总和,导致诱导活化的t细胞编程。tcr是该回路的关键组分。tcr将t细胞毒性的焦点定向于病原体的独特表位。合成抗原受体,包括t细胞抗原偶联剂(tac)的开发,导致了将t细胞的细胞毒性天赋定向于新靶的能力;以增强的肿瘤监视和抗肿瘤活性来重利用人类免疫系统。tac分子递送双信号假说的信号1。在工程化细胞内衔接信号2可能诱导t细胞活化的更强刺激。t细胞活化嵌合受体的成功设想了开发新的嵌合共刺激受体以诱导工程化细胞中的独特特征的想法。共刺激受体包括cd28、4-1bb和ox40是良好描述的受体,它们为活化的t细胞提供第二信号,促进活化、增殖和细胞毒性功能。如以上描述的,开发了能够在活化的t细胞分泌配体tnfα的控制下提供共刺激的嵌合共刺激受体(ccr)。该设计方法背后的灵感是仅在t细胞活化后提供共刺激。这些研究已经发现活化的t细胞分泌丰富的tnfα。原则上,分泌的tnfα应该结合并刺激ccr。初始ccr构建体设计包含tnfrsf的共刺激分子4-1bb。ccr将共刺激信号作为单独的受体工程化,导致其跨越许多细胞疗法的实用性,包括所有形式的t细胞疗法(例如,工程化t细胞,til)、nk细胞应用和工程化单核细胞疗法。本项目中的ccr显示出增强的增殖能力和改变的细胞因子产生二者,无论是单独表达还是与tac受体组合表达。在ccr
工程化t细胞中观察到的增强增殖可能提供了克服持续对过继性t细胞转移领域提出挑战的根深蒂固的肿瘤块的抑制性微环境所需的适应性边缘(adaptive edge)。改变的ccr t细胞的细胞因子产生谱看起来不会导致对增殖、细胞毒性或存活的损伤。降低的细胞因子产生甚至可能证明对施用工程化t细胞后的患者较小的毒性。目前,car t细胞在患者中的输注和扩增导致由细胞因子风暴引起的毒性,这种毒性可能通过用具有改变的细胞因子产生谱的工程化t细胞来减轻。
[0191]
在包含4-1bb ccr时发现的t细胞增殖的改进导致对来源于广泛tnfrsf家族的其他共刺激结构域在工程化t细胞共刺激中的实用性的分析。在tnfr1-嵌合体平台内对tnfrsf共刺激信号传导结构域的增殖筛选揭示了ccr具有增强t细胞功能的潜力的各种特性。值得注意的是,该筛选揭示了通常不被认为是t细胞的共刺激物的baff-r共刺激结构域的实用性。重要的是,这些不同ccr的实用性可能根据应用而不同,这为产生的ccr的整个集合创造价值。目的是产生具有增强的体内抗肿瘤活性的tac ccr t细胞产物。未来的方向将包括继续评价ccr对工程化t细胞的作用。事实上,这些数据的结果表明,本文描述的ccr设计可以用于通过任何介导t细胞的抑制和/或死亡信号的tnfrsf将信号传导重定向。作为进一步迭代的实例,可以设计采用以下的ccr:采用fas(tnfrsf6)的胞外结构域来破坏来自fasl(tnfsf6)的死亡信号传导,或者采用dr4/dr5(tnfrsf10a/b)的胞外结构域来破坏来自trail(tnfsf10)的死亡信号传导,因为fasl和trail二者都可以促进活化后的t细胞死亡。已经显示出ccr工程化t细胞具有增强的增殖;未来的工作将确定这些共刺激的t细胞的细胞因子产生谱。研究由ccr信号传导改变的基因途径并且发现人类t细胞中增殖的调节物将是令人感兴趣的。
[0192]
实验方法
[0193]
t细胞生长
[0194]
从健康供体接收外周血单个核细胞,并且在补充有il-2(100u/ml)和il-7(10ng/ml)的rpmi培养基中,用抗cd3和抗cd28磁珠进行刺激。1天之后,将细胞使用第三代慢病毒载体和包装系统进行转导。将t细胞培养物以1
×
106个细胞/ml维持,每两天添加il-2和il-7。培养14天之后,对t细胞的cd4/cd8、嵌合受体、ngfr(转导标志物)进行表征。
[0195]
慢病毒产生
[0196]
慢病毒通过用包装质粒prsv-rev、pmdlg-prre、pmd2.g和pccl转移质粒通过lipofectamine 2000转染试剂(life technologies)转染hek293t来制备。颗粒通过以28,000rpm超速离心进行浓缩。病毒滴度通过稀释病毒和转导hek293t细胞来确定。通过流式细胞术定量转导的hek293t细胞的%ngfr (转导标志物),并以转导单位(tu/ml)计算滴定。
[0197]
确定蛋白水平(蛋白印迹)
[0198]
将t细胞在冰上使用组织匀浆器在具有蛋白酶抑制剂混合物(1:100)(sigma aldrich)的裂解缓冲液(150mm nacl、0.5%脱氧胆酸钠、0.1%sds、50mm tris、1%triton x100,ph8.0)中匀质化,随后以9,000
×
g在4℃离心15分钟。样品蛋白浓度通过二喹啉甲酸测定(sigma aldrich)确定。将具有2-巯基乙醇的laemmli缓冲液添加至样品,随后在95℃热变性5min。将样品通过10%sds-page凝胶电泳,并且在恒定350ma湿转移至硝化纤维素膜持续90min。将膜用在tbst中的5%(w/v)脱脂乳封闭剂在4℃封闭过夜。将膜在室温与一级抗体(1:1000,在具有2%封闭剂的tbst中)摇动孵育2h。将膜在tbst中洗涤3次,并且与萦光
缀合的二级抗体(1:5000,在具有2%脱脂乳封闭剂的tbst中)孵育1h。将膜用tbst洗涤3次。
[0199]
流式细胞术表面染色
[0200]
收获t细胞,以1500rpm沉淀,并且与针对质膜蛋白的萦光标记物抗体孵育30min。将t细胞在facs缓冲液(1%bsa、1
×
pbs、2.5mm edta)中洗涤并沉淀。将t细胞悬浮于300μl facs缓冲液中并过滤,然后在流式细胞仪上运行。流式细胞仪记录标记的抗体的前向散射、侧向散射和适当的萦光通道。将细胞计数器数据使用flowjo v10软件分析,并且以散点图显示。
[0201]
体外胞内细胞因子产生
[0202]
收获t细胞,并且用抗原阳性的肿瘤细胞系在37℃刺激4小时。添加golgiplug和golgistop试剂(bd bioscience)以阻止t细胞分泌细胞因子。4小时之后,刺激通过添加0.02m edta来停止,并且在室温孵育15min。收集细胞并洗涤随后离心并染色。胞内细胞因子通过固定和透化细胞,随后用萦光标记的针对tnf-α、ifn-γ和il-2的抗体染色来评价。萦光通过流式细胞术评价,并且在flowjo v10软件上进行分析。
[0203]
体外细胞毒性
[0204]
收获t细胞,并且以8至0.25的效应物与靶比例与萤光素酶表达抗原阳性肿瘤细胞系共孵育16-18小时。孵育后,将d-萤光素添加至培养物中,并且在室温孵育10min。亮度在板读取仪上读取。亮度的量与孔中肿瘤细胞的存活力相关。跨效应物与靶比例绘制数据。
[0205]
增殖测定
[0206]
收获t细胞,以1500rpm沉淀,并且重悬于在pbs中的1:1000celltrace
tm violet染色,在37℃持续20min;以1
×
106个细胞/ml染色细胞。将t细胞培养基以4:1染色体积添加,并且在37℃孵育,以猝灭剩余的celltrace
tm
染料。将染色的细胞以1500rpm沉淀,并且重悬于t细胞培养基中用于在无细胞因子情况下在24孔板中以0.5
×
106个细胞/ml铺板。肿瘤靶以1:2的效应物:靶比例添加至孔中。对于单个ccr的表达,将细胞用涂覆有10μg/ml抗cd3(okt3 clone)的板刺激。对t细胞追踪5天或7天,记录细胞数目和存活力。在第5天或第7天收获细胞并进行流式细胞术染色。celltrace
tm violet稀释峰表示分裂的t细胞,并且由此可以计算增殖的度量。
[0207]
实施例3:fas嵌合共刺激受体
[0208]
tnfrsf、fas或cd95是t细胞凋亡的重要调节物。已经展示出fasl在t细胞和肿瘤上表达。fas-fasl相互作用可以通过表达fasl的t细胞与表达fas的t细胞的相互作用而促进活化t细胞的死亡,从而导致自相残杀。肿瘤上的fasl与表达fas的t细胞之间的相互作用也与肿瘤特异性t细胞的细胞死亡相关。死亡信号通过fas受体胞质结构域介导,fas受体胞质结构域与衔接体蛋白(adapter protein)fadd或具有死亡结构域的fas相关蛋白相互作用,其募集胱天蛋白酶8和胱天蛋白酶10以形成死亡诱导信号传导复合体(disc)。disc裂解并活化胱天蛋白酶8/胱天蛋白酶10,以触发效应物胱天蛋白酶从而介导凋亡。
[0209]
去除fas的胞质区会得到显性负性受体,其被预期减弱细胞死亡信号。用4-1bb或baffr的胞质结构域替代fas胞质结构域被预期抑制死亡信号,并且用可以增强t细胞存活的共刺激信号替代它。
[0210]
产生fas嵌合体,其中fas受体的胞外结构域与4-1bb或baffr的胞质结构域结合,如图2b中示出的。
[0211]
fas嵌合共刺激受体的工程化及表达
[0212]
将原代人类t细胞用表达fas-嵌合体或fas-trunc的慢病毒转导。作为对照,将原代人类t细胞用编码非fas受体(ngfr)的对照慢病毒转导。所有慢病毒载体都表达ngfr,使得能够基于ngfr的表达来确定转导效率。基于ngfr转导标志物表达,所有用于工程化t细胞的构建体显示出在40%-50%之间的效率,这与对照病毒可比较(图24)。
[0213]
在细胞表面上用针对天然fas的抗体检测到fas-嵌合体。如以上注意的,非工程化t细胞在培养时间段的第14天表达高水平的fas。被转导以表达包含4-1bb和baffr共刺激结构域的fas-嵌合体的t细胞显示出高于天然fas水平的fas表达水平,如以平均萦光强度评价的(图25)。与ngfr对照中的1.35
×
104相比,fas-4-1bb和fas-baffr嵌合体的细胞表面表达为4.9
×
104和7.9
×
104,如以平均萦光强度评价的。有趣的是,fas-trunc受体几乎不以高于ngfr水平表达(mfi=2.0
×
104)。
[0214]
被工程化以表达fas-嵌合体的t细胞在制造过程期间以与ngfr对照相比可比较的速率生长,表明修饰的fas受体的表达不影响制造时间段期间的t细胞生长(图26)。
[0215]
修饰的fas受体增强增殖
[0216]
为了确定修饰的fas受体的表达是否影响t细胞增殖,将原代人类t细胞用表达fas-trunc、fas-4-1bb或fas-baffr的慢病毒工程化。将对照t细胞用仅表达ngfr的慢病毒工程化。将工程化t细胞用激动剂cd3抗体作为t细胞受体的替代刺激物进行刺激。所有用修饰的fas受体工程化的t细胞都显示出相对于对照ngfr t细胞增强的增殖。用fas-4-1bb或fas-baffr工程化的t细胞与fas-trunc等同增殖,表明抑制fas信号足以增强增殖,而不需要另外的共刺激信号传导(图27)。
[0217]
fas-嵌合体克服外源fasl信号传导
[0218]
fas嵌合体被设计为重定向fasl的凋亡信号。为了确定fas嵌合体是否能够阻断来自fasl的凋亡,将工程化t细胞用可溶性三聚体fasl刺激,并且48小时后使用代谢染料alamarblue评价存活力。对照细胞暴露于fasl,随着浓度的增加,产生降低的t细胞存活力。在存在外源fasl的情况下,对照t细胞(ngfr)显示出剂量依赖性的存活力损失。在存在外源fasl的情况下,所有用修饰的fas受体工程化的t细胞都显示出与对照细胞相比增强的存活力。用fas-baffr工程化的t细胞显示出对外源fasl的最大耐受性,表明共刺激结构域对t细胞存活的益处(图28)。
[0219]
fas-嵌合体在存在fasl的情况下增强增殖
[0220]
由于用修饰的fas受体工程化的t细胞在细胞因子停用后免受fasl介导的细胞死亡,试图确定外源fasl对用修饰的fas受体工程化的t细胞的增殖的影响。将用修饰的fas受体工程化的t细胞用抗cd3活化,并且3天后通过alamarblue评价细胞密度。外源fasl的存在导致对照t细胞密度降低(图29)。用fas-trunc受体工程化的t细胞也观察到相似的细胞密度下降。然而,在存在500ng/ml和1000ng/ml fasl的情况下,被工程化以表达包含共刺激信号结构域的fas-嵌合体的t细胞显示出与ngfr对照工程化的t细胞相比增强的增殖(图29)。被工程化以表达fas-baffr的t细胞随着fasl浓度的增加显示出增强增殖的能力,而用fas-4-1bb工程化的t细胞不受外源fasl的影响(图29)。
[0221]
虽然已经参考实施例描述了本技术,但应当理解,权利要求书的范围不应受到实施例中所述实施方案的限制,而是应给予与作为整体的描述一致的最宽泛解释。
[0222]
所有出版物、专利和专利申请通过引用以其整体并入本文,其程度如同每一个单独的出版物、专利或专利申请被具体和单独地指明通过引用以其整体并入。在发现本技术中的术语在通过引用并入本文的文档中定义不同的情况下,本文提供的定义将用作该术语的定义。
[0223]
表1.tnfrsf ccr构建体设计
[0224][0225]
表1描绘了嵌合共刺激受体筛选的构建体设计。tnfr-融合受体显示出对体外工程化的t细胞的功能增强,因此,在tnfr超家族中寻找新的共刺激结构域是有吸引力的。共刺激筛选被设计为评价tnfr1胞外结构域与tnfrsf成员共刺激信号传导结构域之间融合的t细胞共刺激功能。该表描述了用于筛选中每个构建体的跨膜结构域和共刺激结构域。该表还列出了设计的ccr是否包含已知发送胱天蛋白酶级联的信号的经典死亡结构域。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。
