基因组基因座的增强的重组
1.本技术是申请日为2016年8月19日、申请号为201680059466.6、发明名称为“基因组基因座的增强的重组”的发明专利申请的分案申请。
2.相关申请的交叉参考
3.本技术要求2015年8月21日提交的美国临时申请no.62/208,405的优先权,该临时申请以全文引用的方式并入本文中。
4.序列表的合并
5.本技术含有与其一起电子提交的序列表,所述序列表含有命名为“p34266wo00_seq.txt”的文件,所述文件大小为92,429个字符(在ms-中测量),创建于2016年8月18日,且以全文引用的方式并入本文中。
6.领域
7.本公开提供用于通过引入位点-特异性基因组修饰,允许重组发生并且选择在所选择的基因组基因座内具有分子变异的事件来增强预先选择的基因组基因座处的重组的组合物和方法。
8.背景
9.基因多样性为环境适应的基础。目前,植物育种者依赖于产生基因多样性的天然机制,所述天然机制依赖于罕见的随机突变或重组事件来产生具有诸如对新植物病原体生物型的抗性等所需性状的植株。然后,使用标准植物育种来将所需性状整合至所选优良种质系中,不过这是一种缓慢且成本高的方法,涉及多轮回交和选择。在一些情况下,标准育种技术不能克服与不利性状相关的某些遗传连锁(genetic linkage)。
10.因此,需要将有助于加速开发新的基因座来产生具有改善的环境适应和农艺学性状的植株的方法。在育种方案中还需要具有用于刺激单一细胞内的顺式染色体交换、姐妹染色体交换或多个染色体交换事件的方法。
11.概要
12.若干实施方案涉及一种产生具有新串联重复基因阵列的植株的方法,所述方法包括使植物细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一串联重复基因阵列的至少一个靶序列中引入基因组修饰,由此诱导与第二串联重复基因阵列的重组;以及选择包含新串联重复基因阵列的至少一个植株。在一些实施方案中,产生两种新串联重复基因阵列基因座。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列位于同源染色体上。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列位于非同源染色体上。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列位于部分同源染色体上。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组为不对称的,并且新串联重复基因阵列与第一串联重复基因阵列或第二串联重复基因阵列相比具有增加的基因数目。在一些实施方案中,重组为不对称的,并且新串联重复基因阵列与第一串联
重复基因阵列或第二串联重复基因阵列相比具有减少的基因数目。在一些实施方案中,重组为对称的。在一些实施方案中,重组为对称的,第一串联重复基因阵列和第二串联重复基因阵列为异源的,并且新串联重复基因阵列包含新的基因组合。在一些实施方案中,靶序列位于第一串联重复基因阵列的基因内。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与第二阵列的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,靶序列位于第一阵列的基因组基因座,所述基因组基因座与第二阵列的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的,其中第一阵列的基因组基因座和第一阵列的基因组基因座位于基因组中的对应位置。在一些实施方案中,靶序列位于第一阵列的基因组基因座中,所述基因组基因座与第二阵列的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的,其中第一阵列的基因组基因座和第一阵列的基因组基因座不位于基因组中的对应位置。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为同源的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为异源的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为部分同源的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为旁系同源的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为相同的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为不相同的。在一些实施方案中,第一串联重复基因阵列是定位于第一亲本基因组中,并且第二串联重复基因阵列是定位于第二亲本基因组中。在一些实施方案中,第一亲本基因组和第二亲本基因组在性别上不相容。在一些实施方案中,第一亲本基因组和第二亲本基因组为不同的物种。在一些实施方案中,第一亲本基因组为普通小麦(triticum aestivum)(小麦),并且第二亲本基因组是选自卵穗山羊草(aegilops ovate)、两芒山羊草(ae.biuncialis)、三芒山羊草(ae.triuncialis)、粗山羊草(ae.quarrosa)、黑麦(secale cereal)、野生二粒小麦(triticum dicoccoides)、二粒小麦(triticum dicoccum)以及硬粒小麦(triticum durum)。在一些实施方案中,第一亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦,并且第二亲本基因组为普通小麦(小麦)。在一些实施方案中,第一亲本基因组为陆地棉(gossypium hirsutum)(棉花),并且第二亲本基因组是选自史特啼棉(g.sturtii)、戴维逊氏棉(g.davidsonii)、亚洲棉(g.arboretum)以及雷蒙德氏棉(g.raimondii)。在一些实施方案中,第一亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉,并且第二亲本基因组为陆地棉(棉花)。在一些实施方案中,第一亲本基因组和/或第二亲本基因组为单倍体。在一些实施方案中,第一亲本基因组和/或第二亲本基因组为二倍体。在一些实施方案中,第一串联重复基因阵列、第二串联重复基因阵列以及新串联重复基因阵列编码nbs-lrr抗病蛋白、病原体识别受体(prr)蛋白、种子贮藏蛋白、细胞壁组分扩展蛋白、f-框蛋白、abc转运蛋白或丝氨酸-苏氨
酸/酪氨酸蛋白质激酶。在一些实施方案中,第一串联重复基因阵列、第二串联重复基因阵列以及新串联重复基因阵列编码核糖体rna。在一些实施方案中,至少一个包含新串联重复基因阵列的子代与包含第一串联重复基因阵列的植株、包含第二串联重复基因阵列的植株或包含第一串联重复基因阵列和第二串联重复基因阵列的植株相比展现改善的抗病性。在一些实施方案中,新串联重复基因阵列赋予对一种或多种选自以下的疾病的抗性:炭疽茎腐病(anthracnose stalk rot)(禾生刺盘孢(colletotrichum graminicola))、镰刀菌穗腐病(fusarium ear rot)(轮枝镰刀菌(fusarium verticillioides))、镰刀菌茎腐病(镰刀菌属(fusarium spp.))、赤霉穗腐病(gibberella ear rot)(念珠状赤霉(gibberella moniliformis))、赤霉茎腐病(玉米赤霉(gibberella zeae))、戈斯氏萎蔫病(goss's wilt)和叶枯病(苜蓿萎蔫病菌(clavibacter michiganensis))、灰色叶斑病(玉蜀黍尾孢菌(cercospora zeae-maydis)、玉米尾孢菌(c.zeina))、北方玉米叶枯病(大斑凸脐蠕孢(exserohilum turcicum))、猝死综合征(茄病镰刀菌(fusarium solani f.sp.glycines))、亚洲大豆锈病(豆薯层锈菌(phakopsora pachyrhizi))、疫霉根腐和茎腐病(phytophthora root and stem rot)(大豆疫霉(phytophthora sojae))、根结线虫病(root-knot nematode)(根结线虫属(meloidogyne spp.))、大豆胞囊线虫病(soybean cyst nematode)(大豆胞囊线虫(heterodera glycines))、肾形线虫病(reniform nematode)(肾状轮线虫(rotylenchulus reniformis))、根结线虫病(南方根结线虫(meloidogyne incognita))、镰刀菌萎蔫病(尖孢镰刀菌萎蔫专化型(fusarium oxysporurn f.sp.vasinfectum))、轮枝孢菌萎蔫病(verticillium wilt)(大丽轮枝孢菌(verticillium dahlia))、镰刀菌首枯病(禾谷镰刀菌(fusarium graminearum))、镰刀菌苗枯病(镰刀菌属、颖枯壳针孢(septoria nodorum))、镰刀菌叶斑病(雪腐小画线壳(monographella nivalis))以及茎锈病(禾柄锈菌(puccinia graminis))。在一些实施方案中,植株为玉米植株。在一些实施方案中,植株为大豆植株。在一些实施方案中,植株为棉花植株。在一些实施方案中,植株为小麦植株。在一些实施方案中,植株为高粱植株。在一些实施方案中,植株为芥花植株。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
13.若干实施方案涉及一种产生具有新串联重复基因阵列的植株的方法,所述方法包括使植物细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一串联重复基因阵列的至少一个靶序列中以及第二串联重复基因阵列的至少一个靶序列中引入基因组修饰,由此诱导第一和第二串联重复基因阵列之间的重组;以及选择包含新串联重复基因阵列的至少一个植株。在一些实施方案中,产生两种新串联重复基因阵列基因座。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列位于同源染色体上。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列位于非同源染色体上。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列位于部分同源染色体上。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组为不对称的,并且新串联重复基因阵列与第一串联重复基因阵列或第二串联重复基因阵列相比具有增加的基因数目。在一些实施方案中,重组为不对称的,并且新串联重复基因阵列与第一串联重复基因阵列或第二串联重复基因阵列相比具有减少的基因数目。在一些实施方案中,重组为对称的。在一些实施方案中,重组为对称的,第一串联重复基因阵列和第二串联重复基因阵列为异源的,并且新串联重复基因阵列包含新的基因组合。在一些实施方案中,靶序列为基因的。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,第一串联重复基因阵列中的靶序列位于基因组基因座中,所述基因组基因座与第二阵列的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,第一串联重复基因阵列中的靶序列位于基因组基因座中,所述基因组基因座与第二阵列的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的,其中第一阵列的基因组基因座和第一阵列的基因组基因座位于基因组中的对应位置。在一些实施方案中,第一串联重复基因阵列中的靶序列位于基因组基因座中,所述基因组基因座与第二阵列的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的,其中第一阵列的基因组基因座和第一阵列的基因组基因座不位于基因组中的对应位置。在一些实施方案中,第一串联重复基因阵列中的靶序列与第二串联重复基因阵列中的靶序列具有至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%或100%的序列同一性。在一些实施方案中,第一串联重复基因阵列与第二串联重复基因阵列为同源的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为异源的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为部分同源的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为旁系同源的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为相同的。
在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为不相同的。在一些实施方案中,第一串联重复基因阵列是定位于第一亲本基因组中,并且第二串联重复基因阵列是定位于第二亲本基因组中。在一些实施方案中,第一亲本基因组和第二亲本基因组在性别上不相容。在一些实施方案中,第一亲本基因组和第二亲本基因组为不同的物种。在一些实施方案中,第一亲本基因组为普通小麦(小麦),并且第二亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦。在一些实施方案中,第一亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦,并且第二亲本基因组为普通小麦(小麦)。在一些实施方案中,第一亲本基因组为陆地棉(棉花),并且第二亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉。在一些实施方案中,第一亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉,并且第二亲本基因组为陆地棉(棉花)。在一些实施方案中,第一亲本基因组和/或第二亲本基因组为单倍体。在一些实施方案中,第一亲本基因组和/或第二亲本基因组为二倍体。在一些实施方案中,第一串联重复基因阵列、第二串联重复基因阵列以及新串联重复基因阵列编码nbs-lrr抗病蛋白、病原体识别受体(prr)蛋白、种子贮藏蛋白、细胞壁组分扩展蛋白、f-框蛋白、abc转运蛋白或丝氨酸-苏氨酸/酪氨酸蛋白质激酶。在一些实施方案中,第一串联重复基因阵列、第二串联重复基因阵列以及新串联重复基因阵列编码核糖体rna。在一些实施方案中,至少一个包含新串联重复基因阵列的子代与包含第一串联重复基因阵列的植株、包含第二串联重复基因阵列的植株或包含第一串联重复基因阵列和第二串联重复基因阵列的植株相比展现改善的抗病性。在一些实施方案中,新串联重复基因阵列赋予对一种或多种选自以下的疾病的抗性:炭疽茎腐病(禾生刺盘孢)、镰刀菌穗腐病(轮枝镰刀菌)、镰刀菌茎腐病(镰刀菌属)、赤霉穗腐病(念珠状赤霉)、赤霉茎腐病(玉米赤霉)、戈斯氏萎蔫病和叶枯病(苜蓿萎蔫病菌)、灰色叶斑病(玉蜀黍尾孢菌、玉米尾孢菌)、北方玉米叶枯病(大斑凸脐蠕孢)、猝死综合征(茄病镰刀菌)、亚洲大豆锈病(豆薯层锈菌)、疫霉根腐和茎腐病(大豆疫霉)、根结线虫病(根结线虫属)、大豆胞囊线虫病(大豆胞囊线虫)、肾形线虫病(肾状轮线虫)、根结线虫病(南方根结线虫)、镰刀菌萎蔫病(尖孢镰刀菌萎蔫专化型)、轮枝孢菌萎蔫病(大丽轮枝孢菌)、镰刀菌首枯病(禾谷镰刀菌)、镰刀菌苗枯病(镰刀菌属、颖枯壳针孢)、镰刀菌叶斑病(雪腐小画线壳)以及茎锈病(禾柄锈菌)。在一些实施方案中,植株为玉米植株。在一些实施方案中,植株为大豆植株。在一些实施方案中,植株为棉花植株。在一些实施方案中,植株为小麦植株。在一些实施方案中,植株为高粱植株。在一些实施方案中,植株为芥花植株。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重
组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
14.若干实施方案涉及一种产生具有新串联重复基因阵列的植株的方法,所述方法包括使植物细胞与第一位点-特异性基因组修饰酶和第二位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一串联重复基因阵列的至少一个靶序列中引入基因组修饰,所述第二位点-特异性基因组修饰酶在第二串联重复基因阵列的至少一个靶序列中引入基因组修饰,由此诱导第一和第二串联重复基因阵列之间的重组;以及选择包含新串联重复基因阵列的至少一个植株。在一些实施方案中,产生两种新串联重复基因阵列基因座。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列位于同源染色体上。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列位于非同源染色体上。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列位于部分同源染色体上。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组为不对称的,并且新串联重复基因阵列与第一串联重复基因阵列或第二串联重复基因阵列相比具有增加的基因数目。在一些实施方案中,重组为不对称的,并且新串联重复基因阵列与第一串联重复基因阵列或第二串联重复基因阵列相比具有减少的基因数目。在一些实施方案中,重组为对称的。在一些实施方案中,重组为对称的,第一串联重复基因阵列和第二串联重复基因阵列为异源的,并且新串联重复基因阵列包含新的基因组合。在一些实施方案中,靶序列为基因的。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,第一串联重复基因阵列中的靶序列位于基因组基因座中,所述基因组基因座与第二阵列的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,第一串联重复基因阵列中的靶序列位于基因组基因座中,所述基因组基因座与第二阵列的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的,其中第一阵列的基因组基因座和第一阵列的基因组基因座位于基因组中的对应位置。在一些实施方案中,第一串联重复基因阵列中的靶序列位于基因组基因座中,所述基因组基因座与第二阵列的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的,其中第一阵列的基因组基因座和第一阵列的基因组基因座不位于基因组中的对应位置。在一些实施方案中,第一串联重复基因阵列中的靶序列与第二串联重复基因阵列中的靶序列具有至少80%、至少
81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%或100%的序列同一性。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为同源的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为异源的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为部分同源的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为旁系同源的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为相同的。在一些实施方案中,第一串联重复基因阵列和第二串联重复基因阵列为不相同的。在一些实施方案中,第一串联重复基因阵列是定位于第一亲本基因组中,并且第二串联重复基因阵列是定位于第二亲本基因组中。在一些实施方案中,第一亲本基因组和第二亲本基因组在性别上不相容。在一些实施方案中,第一亲本基因组和第二亲本基因组为不同的物种。在一些实施方案中,第一亲本基因组为普通小麦(小麦),并且第二亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦。在一些实施方案中,第一亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦,并且第二亲本基因组为普通小麦(小麦)。在一些实施方案中,第一亲本基因组为陆地棉(棉花),并且第二亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉。在一些实施方案中,第一亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉,并且第二亲本基因组为陆地棉(棉花)。在一些实施方案中,第一亲本基因组和/或第二亲本基因组为单倍体。在一些实施方案中,第一亲本基因组和/或第二亲本基因组为二倍体。在一些实施方案中,第一串联重复基因阵列、第二串联重复基因阵列以及新串联重复基因阵列编码nbs-lrr抗病蛋白、病原体识别受体(prr)蛋白、种子贮藏蛋白、细胞壁组分扩展蛋白、f-框蛋白、abc转运蛋白或丝氨酸-苏氨酸/酪氨酸蛋白质激酶。在一些实施方案中,第一串联重复基因阵列、第二串联重复基因阵列以及新串联重复基因阵列编码核糖体rna。在一些实施方案中,至少一个包含新串联重复基因阵列的子代与包含第一串联重复基因阵列的植株、包含第二串联重复基因阵列的植株或包含第一串联重复基因阵列和第二串联重复基因阵列的植株相比展现改善的抗病性。在一些实施方案中,新串联重复基因阵列赋予对一种或多种选自以下的疾病的抗性:炭疽茎腐病(禾生刺盘孢)、镰刀菌穗腐病(轮枝镰刀菌)、镰刀菌茎腐病(镰刀菌属)、赤霉穗腐病(念珠状赤霉)、赤霉茎腐病(玉米赤霉)、戈斯氏萎蔫病和叶枯病(苜蓿萎蔫病菌)、灰色叶斑病(玉蜀黍尾孢菌、玉米尾孢菌)、北方玉米叶枯病(大斑凸脐蠕孢)、猝死综合征(茄病镰刀菌)、亚洲大豆锈病(豆薯层锈菌)、疫霉根腐和茎腐病(大豆疫霉)、根结线虫病(根结线虫属)、大豆胞囊线虫病(大豆胞囊线虫)、肾形线虫病(肾状轮线虫)、根结线虫病(南方根结线虫)、镰刀菌萎蔫病(尖孢镰刀菌萎蔫专化型)、轮枝孢菌萎蔫病(大丽轮枝孢菌)、镰刀菌首枯病(禾谷镰刀菌)、镰刀菌苗枯病(镰刀菌属、颖枯壳针孢)、镰刀菌叶斑病(雪腐小画线壳)以及茎锈病(禾柄锈菌)。在一些实施方案中,植株为玉米植株。在一些实施方案中,植株为大豆植株。在一些实施方案中,植株为棉花植株。在一些实施方案中,植株为小麦植株。在一些实施方案中,植株为高粱植株。在一些实施方案中,植株为芥花植株。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、
rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
15.若干实施方案涉及一种产生具有rp1抗病性基因座的新等位基因的玉米植株的方法,所述方法包括使玉米细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一rp1抗病性基因座的至少一个靶序列中引入基因组修饰,由此诱导与第二rp1抗病性基因座的重组;以及选择包含rp1抗病性基因座的新等位基因的至少一个玉米植株。若干实施方案涉及一种产生具有rp1抗病性基因座的新等位基因的玉米植株的方法,所述方法包括使玉米细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一rp1抗病性基因座的至少一个靶序列中以及第一rp1抗病性基因座的至少一个靶序列中引入基因组修饰,由此诱导第一rp1抗病性基因座与第二rp1抗病性基因座之间的重组;以及选择包含rp1抗病性基因座的新等位基因的至少一个玉米植株。若干实施方案涉及一种产生具有rp1抗病性基因座的新等位基因的玉米植株的方法,所述方法包括使玉米细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一rp1抗病性基因座的至少一个靶序列中引入基因组修饰,并且使玉米细胞与第二位点-特异性基因组修饰酶接触,所述第二位点-特异性基因组修饰酶在第二rp1抗病性基因座的至少一个靶序列中引入基因组修饰,由此诱导第一rp1抗病性基因座与第二rp1抗病性基因座之间的重组;以及选择包含rp1抗病性基因座的新等位基因的至少一个玉米植株。在一些实施方案中,产生rp1抗病性基因座的两种新等位基因。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组为不对称的,并且rp1抗病性基因座的新等位基因与第一rp1抗病性基因座或第二rp1抗病性基因座相比具有增加的rp1基因数目。在一些实施方案中,重组为不对称的,并且rp1抗病性基因座的新等位基因与第一rp1抗病性基因座或第二rp1抗病性基因座相比具有减少的rp1基因数目。在一些实施方案中,重组为对称的。在一些实施方案中,重组为对称的,第一rp1抗病性基因座与第二rp1抗病性基因座为异源的,并且rp1抗病性基因座的新等位基因包含新的rp1基因组合。在一些实施方案中,靶序列位于基因内。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,靶序列是选自包含以下各项的组中的一种或多
种:seq id no:3、seq id no:4、seq id no:5、seq id no:6、seq id no:7、seq id no:8以及seq id no:9。在一些实施方案中,第一rp1抗病性基因座中的靶序列与第二rp1抗病性基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,第一rp1抗病性基因座和第二rp1抗病性基因座为同源的。在一些实施方案中,第一rp1抗病性基因座和第二rp1抗病性基因座为异源的。在一些实施方案中,第一rp1抗病性基因座和第二rp1抗病性基因座为部分同源的。在一些实施方案中,第一rp1抗病性基因座和第二rp1抗病性基因座为相同的。在一些实施方案中,第一rp1抗病性基因座和第二rp1抗病性基因座为不相同的。在一些实施方案中,第一rp1抗病性基因座是定位于第一亲本基因组中,并且第二rp1抗病性基因座是定位于第二亲本基因组中。在一些实施方案中,rp1抗病性基因座的新等位基因赋予对一种或多种选自以下的疾病的抗性:炭疽茎腐病(禾生刺盘孢)、镰刀菌穗腐病(轮枝镰刀菌)、镰刀菌茎腐病(镰刀菌属)、赤霉穗腐病(念珠状赤霉)、赤霉茎腐病(玉米赤霉)、戈斯氏萎蔫病和叶枯病(苜蓿萎蔫病菌)、灰色叶斑病(玉蜀黍尾孢菌、玉米尾孢菌)以及北方玉米叶枯病(大斑凸脐蠕孢)。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
16.若干实施方案涉及一种产生具有rpp1抗病性基因座的新等位基因的大豆植株的方法,所述方法包括使大豆细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一rpp1抗病性基因座的至少一个靶序列中引入基因组修饰,由此诱导与第二rpp1抗病性基因座的重组;以及选择包含rpp1抗病性基因座的新等位基因的至少一个大豆植株。若干实施方案涉及一种产生具有rpp1抗病性基因座的新等位基因的大豆植株的方法,所述方法包括使大豆细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一rpp1抗病性基因座的至少一个靶序列中以及第一rpp1抗病性基因座的至少一个靶序列中引入基因组修饰,由此诱导第一rpp1抗病性基因座与第二rpp1抗病性基因座之间的重组;以及选择包含rpp1抗病性基因座的新等位基因的至少一个大豆植株。若干实施方案涉及一种产生具有rpp1抗病性基因座的新等位基因的大豆植株的
方法,所述方法包括使大豆细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一rpp1抗病性基因座的至少一个靶序列中引入基因组修饰,并且使大豆细胞与第二位点-特异性基因组修饰酶接触,所述第二位点-特异性基因组修饰酶在第二rpp1抗病性基因座的至少一个靶序列中引入基因组修饰,由此诱导第一rpp1抗病性基因座与第二rpp1抗病性基因座之间的重组;以及选择包含rpp1抗病性基因座的新等位基因的至少一个大豆植株。在一些实施方案中,产生rpp1抗病性基因座的两种新等位基因。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组为不对称的,并且rpp1抗病性基因座的新等位基因与第一rpp1抗病性基因座或第二rpp1抗病性基因座相比具有增加的rpp1基因数目。在一些实施方案中,重组为不对称的,并且rpp1抗病性基因座的新等位基因与第一rpp1抗病性基因座或第二rpp1抗病性基因座相比具有减少的rpp1基因数目。在一些实施方案中,重组为对称的。在一些实施方案中,重组为对称的,第一rpp1抗病性基因座和第二rpp1抗病性基因座为异源的,并且rpp1抗病性基因座的新等位基因包含新的rpp1基因组合。在一些实施方案中,靶序列位于基因内。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,靶序列是选自包含以下各项的组中的一种或多种:seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17以及seq id no:18。在一些实施方案中,第一rpp1抗病性基因座中的靶序列与第二rpp1抗病性基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,第一rpp1抗病性基因座和第二rpp1抗病性基因座为同源的。在一些实施方案中,第一rpp1抗病性基因座和第二rpp1抗病性基因座为异源的。在一些实施方案中,第一rpp1抗病性基因座和第二rpp1抗病性基因座为部分同源的。在一些实施方案中,第一rpp1抗病性基因座和第二rpp1抗病性基因座为相同的。在一些实施方案中,第一rpp1抗病性基因座和第二rpp1抗病性基因座为不相同的。在一些实施方案中,第一rpp1抗病性基因座是定位于第一亲本基因组中,并且第二rpp1抗病性基因座是定位于第二亲本基因组中。在一些实施方案中,rpp1抗病性基因座的新等位基因赋予对一种或多种选自以下的疾病的抗性:猝死综合征(sds)、疫霉根腐病、疫霉茎腐病、镰刀菌根腐病、丝核菌根腐病、炭腐病、大豆胞囊线虫病(scn)、核盘霉茎腐病(白霉)、褐茎腐病(bsr)、荚枯和茎枯病、茎溃疡、炭疽病、绿茎综合征、大豆锈病、壳针孢褐斑病、细菌性枯病、霜霉病、尾孢菌叶枯病、大豆灰斑病、白粉病、大豆花叶病毒以及豆荚斑驳病毒。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、
csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
17.若干实施方案涉及一种产生具有rps1抗病性基因座的新等位基因的大豆植株的方法,所述方法包括使大豆细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一rps1抗病性基因座的至少一个靶序列中引入基因组修饰,由此诱导与第二rps1抗病性基因座的重组;以及选择包含rps1抗病性基因座的新等位基因的至少一个大豆植株。若干实施方案涉及一种产生具有rps1抗病性基因座的新等位基因的大豆植株的方法,所述方法包括使大豆细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一rps1抗病性基因座的至少一个靶序列中以及第一rps1抗病性基因座的至少一个靶序列中引入基因组修饰,由此诱导第一rps1抗病性基因座与第二rps1抗病性基因座之间的重组;以及选择包含rps1抗病性基因座的新等位基因的至少一个大豆植株。若干实施方案涉及一种产生具有rps1抗病性基因座的新等位基因的大豆植株的方法,所述方法包括使大豆细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一rps1抗病性基因座的至少一个靶序列中引入基因组修饰,并且使大豆细胞与第二位点-特异性基因组修饰酶接触,所述第二位点-特异性基因组修饰酶在第二rps1抗病性基因座的至少一个靶序列中引入基因组修饰,由此诱导第一rps1抗病性基因座与第二rps1抗病性基因座之间的重组;以及选择包含rps1抗病性基因座的新等位基因的至少一个大豆植株。在一些实施方案中,产生rps1抗病性基因座的两种新等位基因。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组为不对称的,并且rps1抗病性基因座的新等位基因与第一rps1抗病性基因座或第二rps1抗病性基因座相比具有增加的rps1基因数目。在一些实施方案中,重组为不对称的,并且rps1抗病性基因座的新等位基因与第一rps1抗病性基因座或第二rps1抗病性基因座相比具有减少的rps1基因数目。在一些实施方案中,重组为对称的。在一些实施方案中,重组为对称的,第一rps1抗病性基因座和第二rps1抗病性基因座为异源的,并且rps1抗病性基因座的新等位基因包含新的rps1基因组合。在一些实施方案中,靶序列位于基因内。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,靶序列是选自包含以下各项的组中的一种或多种:seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26、seq id no:27、seq id no:28、seq id no:29、seq id no:30、seq id no:31以及seq id no:32。在一些实施方案中,第一rps1抗病性基因座中的靶序列与第二rps1抗病性基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约
600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,第一rps1抗病性基因座和第二rps1抗病性基因座为同源的。在一些实施方案中,第一rps1抗病性基因座和第二rps1抗病性基因座为异源的。在一些实施方案中,第一rps1抗病性基因座和第二rps1抗病性基因座为部分同源的。在一些实施方案中,第一rps1抗病性基因座和第二rps1抗病性基因座为相同的。在一些实施方案中,第一rps1抗病性基因座和第二rps1抗病性基因座为不相同的。在一些实施方案中,第一rps1抗病性基因座是定位于第一亲本基因组中,并且第二rps1抗病性基因座是定位于第二亲本基因组中。在一些实施方案中,rps1抗病性基因座的新等位基因赋予对一种或多种选自以下的疾病的抗性:猝死综合征(sds)、疫霉根腐病、疫霉茎腐病、镰刀菌根腐病、丝核菌根腐病、炭腐病、大豆胞囊线虫病(scn)、核盘霉茎腐病(白霉)、褐茎腐病(bsr)、荚枯和茎枯病、茎溃疡、炭疽病、绿茎综合征、大豆锈病、壳针孢褐斑病、细菌性枯病、霜霉病、尾孢菌叶枯病、大豆灰斑病、白粉病、大豆花叶病毒以及豆荚斑驳病毒。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
18.若干实施方案涉及一种产生具有rhg1大豆胞囊线虫抗性基因座的新等位基因的大豆植株的方法,所述方法包括使大豆细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一rhg1大豆胞囊线虫抗性基因座的至少一个靶序列中引入基因组修饰,由此诱导与第二rhg1大豆胞囊线虫抗性基因座的重组;以及选择包含rhg1大豆胞囊线虫抗性基因座的新等位基因的至少一个大豆植株。若干实施方案涉及一种产生具有rhg1大豆胞囊线虫抗性基因座的新等位基因的大豆植株的方法,所述方法包括使大豆细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一rhg1大豆胞囊线虫抗性基因座的至少一个靶序列中以及第一rhg1大豆胞囊线虫抗性基因座的至少一个靶序列中引入基因组修饰,由此诱导第一rhg1大豆胞囊线虫抗性基因座与第二rhg1大豆胞囊线虫抗性基因座之间的重组;以及选择包含rhg1大豆胞囊线虫抗性基因座的新等位基因的至少一个大豆植株。若干实施方案涉及一种产生具有rhg1大豆胞囊线虫抗性基因座的新等位基因的大豆植株的方法,所述方法包括使大豆细胞与第一位点-特异性基因组修饰酶接触,所述第一位点-特异性基因组修饰酶在第一rhg1大豆胞囊线虫抗性基
因座的至少一个靶序列中引入基因组修饰,并且使大豆细胞与第二位点-特异性基因组修饰酶接触,所述第二位点-特异性基因组修饰酶在第二rhg1大豆胞囊线虫抗性基因座的至少一个靶序列中引入基因组修饰,由此诱导第一rhg1大豆胞囊线虫抗性基因座与第二rhg1大豆胞囊线虫抗性基因座之间的重组;以及选择包含rhg1大豆胞囊线虫抗性基因座的新等位基因的至少一个大豆植株。在一些实施方案中,产生rhg1大豆胞囊线虫抗性基因座的两种新等位基因。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组为不对称的,并且rhg1大豆胞囊线虫抗性基因座的新等位基因与第一rhg1大豆胞囊线虫抗性基因座或第二rhg1大豆胞囊线虫抗性基因座相比具有增加的rhg1基因数目。在一些实施方案中,重组为不对称的,并且rhg1大豆胞囊线虫抗性基因座的新等位基因与第一rhg1大豆胞囊线虫抗性基因座或第二rhg1大豆胞囊线虫抗性基因座相比具有减少的rhg1基因数目。在一些实施方案中,重组为对称的。在一些实施方案中,重组为对称的,第一rhg1大豆胞囊线虫抗性基因座和第二rhg1大豆胞囊线虫抗性基因座为异源的,并且rhg1大豆胞囊线虫抗性基因座的新等位基因包含新的rhg1基因组合。在一些实施方案中,靶序列位于基因内。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,靶序列是选自包含以下各项的组中的一种或多种:seq id no:69、seq id no:70、seq id no:71、seq id no:72、seq id no:73、seq id no:74、seq id no:75以及seq id no:76。在一些实施方案中,第一rhg1大豆胞囊线虫抗性基因座中的靶序列与第二rhg1大豆胞囊线虫抗性基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,第一rhg1大豆胞囊线虫抗性基因座和第二rhg1大豆胞囊线虫抗性基因座为同源的。在一些实施方案中,第一rhg1大豆胞囊线虫抗性基因座和第二rhg1大豆胞囊线虫抗性基因座为异源的。在一些实施方案中,第一rhg1大豆胞囊线虫抗性基因座和第二rhg1大豆胞囊线虫抗性基因座为部分同源的。在一些实施方案中,第一rhg1大豆胞囊线虫抗性基因座和第二rhg1大豆胞囊线虫抗性基因座为相同的。在一些实施方案中,第一rhg1大豆胞囊线虫抗性基因座和第二rhg1大豆胞囊线虫抗性基因座为不相同的。在一些实施方案中,第一rhg1大豆胞囊线虫抗性基因座是定位于第一亲本基因组中,并且第二rhg1大豆胞囊线虫抗性基因座是定位于第二亲本基因组中。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案
中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
19.若干实施方案涉及一种提供具有改善的抗病性的植株的方法,所述方法包括:(a)向一个或多个植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在抗病性基因座中的至少一个靶序列中引入基因组修饰;(b)针对同源染色体上的抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择具有改善的抗病性的植株。若干实施方案涉及一种提供具有改善的抗病性的植株的方法,所述方法包括:(a)向一个或多个植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在第一抗病性基因座中的至少一个靶序列中引入基因组修饰;(b)针对第一抗病性基因座与第二抗病性基因座之间的重组进行筛选,其中第一和第二抗病性基因座位于非同源染色体上,以鉴定包含重组抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择具有改善的抗病性的植株。若干实施方案涉及一种提供具有改善的抗病性的植株的方法,所述方法包括:(a)向一个或多个植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在第一抗病性基因座中的至少一个靶序列中引入基因组修饰;(b)针对第一抗病性基因座与第二抗病性基因座之间的重组进行筛选,其中第一和第二抗病性基因座位于部分同源染色体上,以鉴定包含重组抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择具有改善的抗病性的植株。若干实施方案涉及一种提供具有改善的抗病性的植株的方法,所述方法包括:(a)向一个或多个植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在抗病性基因座中的第一靶序列和抗病性基因座中的第二靶序列中引入基因组修饰,其中第一靶序列和第二靶序列位于同源染色体上;(b)针对同源染色体上的抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择具有改善的抗病性的植株。若干实施方案涉及一种提供具有改善的抗病性的植株的方法,所述方法包括:(a)向一个或多个植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在第一抗病性基因座中的第一靶序列和第二抗病性基因座中的第二靶序列中引入基因组修饰,其中第一抗病性基因座和第二抗病性基因座位于非同源染色体上;(b)针对抗病性基因座之间的重组进行筛选,以鉴定包含重组抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择具有改善的抗病性的植株。若干实施方案涉及一种提供具有改善的抗病性的植株的方法,所述方法包括:(a)向一个或多个植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在第一抗病性基因座中的第一靶序列和第二抗病性基因座中的第二靶序列中引入基因组修饰,其中第一抗病性基因座和第二抗病性基因座位于部分同源染色体上;(b)针对抗病性基因座之间的重组进行筛选,以鉴定包含重组抗病性基因座的植物细胞;(c)针对改善
的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择具有改善的抗病性的植株。若干实施方案涉及一种提供具有改善的抗病性的植株的方法,所述方法包括:(a)向一个或多个植物细胞提供第一位点-特异性基因组修饰酶和第二位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在抗病性基因座中的第一靶序列中引入基因组修饰,所述第二位点-特异性基因组修饰酶在抗病性基因座中的第二靶序列中引入基因组修饰,其中第一靶序列和第二靶序列位于同源染色体上;(b)针对同源染色体上的抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择具有改善的抗病性的植株。若干实施方案涉及一种提供具有改善的抗病性的植株的方法,所述方法包括:(a)向一个或多个植物细胞提供第一位点-特异性基因组修饰酶和第二位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在第一抗病性基因座中的第一靶序列中引入基因组修饰,所述第二位点-特异性基因组修饰酶在第二抗病性基因座中的第二靶序列中引入基因组修饰,其中第一抗病性基因座和第二抗病性基因座位于非同源染色体上;(b)针对抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择具有改善的抗病性的植株。若干实施方案涉及一种提供具有改善的抗病性的植株的方法,所述方法包括:(a)向一个或多个植物细胞提供第一位点-特异性基因组修饰酶和第二位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在第一抗病性基因座中的第一靶序列中引入基因组修饰,所述第二位点-特异性基因组修饰酶在第二抗病性基因座中的第二靶序列中引入基因组修饰,其中第一抗病性基因座和第二抗病性基因座位于部分同源染色体上;(b)针对抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择具有改善的抗病性的植株。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组抗病性基因座与任一亲代中的抗病性基因座相比具有增加的基因数目。在一些实施方案中,重组抗病性基因座与任一亲代中的抗病性基因座相比具有减少的基因数目。在一些实施方案中,重组抗病性基因座与任一亲代中的抗病性基因座相比具有不同的基因组合。在一些实施方案中,靶序列为基因的。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与同源染色体上的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与同源染色体上不同位置的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,重组是在同源抗病性基因座之间。在一些实施方案中,重组是在异源抗病性基因座之间。在一些实施方案中,重组
是在部分同源抗病性基因座之间。在一些实施方案中,重组是在旁系同源抗病性基因座之间。在一些实施方案中,重组是在相同抗病性基因座之间。在一些实施方案中,同源染色体是来自性别上不相容的亲本基因组。在一些实施方案中,第一亲本基因组为普通小麦(小麦),并且第二亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦。在一些实施方案中,第一亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦,并且第二亲本基因组为普通小麦(小麦)。在一些实施方案中,第一亲本基因组为陆地棉(棉花),并且第二亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉。在一些实施方案中,第一亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉,并且第二亲本基因组为陆地棉(棉花)。在一些实施方案中,同源染色体为不同植物种类。在一些实施方案中,第一抗病性基因座为rgh1,并且第二抗病性基因座为rgh4。在一些实施方案中,抗病性基因座编码nbs-lrr抗病蛋白、病原体识别受体(prr)蛋白、种子贮藏蛋白、细胞壁组分扩展蛋白、f-框蛋白、abc转运蛋白或丝氨酸-苏氨酸/酪氨酸蛋白质激酶。在一些实施方案中,重组抗病性基因座赋予对一种或多种选自以下的疾病的抗性:炭疽茎腐病(禾生刺盘孢)、镰刀菌穗腐病(轮枝镰刀菌)、镰刀菌茎腐病(镰刀菌属)、赤霉穗腐病(念珠状赤霉)、赤霉茎腐病(玉米赤霉)、戈斯氏萎蔫病和叶枯病(苜蓿萎蔫病菌)、灰色叶斑病(玉蜀黍尾孢菌、玉米尾孢菌)、北方玉米叶枯病(大斑凸脐蠕孢)、猝死综合征(茄病镰刀菌)、亚洲大豆锈病(豆薯层锈菌)、疫霉根腐和茎腐病(大豆疫霉)、根结线虫病(根结线虫属)、大豆胞囊线虫病(大豆胞囊线虫)、肾形线虫病(肾状轮线虫)、根结线虫病(南方根结线虫)、镰刀菌萎蔫病(尖孢镰刀菌萎蔫专化型)、轮枝孢菌萎蔫病(大丽轮枝孢菌)、镰刀菌首枯病(禾谷镰刀菌)、镰刀菌苗枯病(镰刀菌属、颖枯壳针孢)、镰刀菌叶斑病(雪腐小画线壳)以及茎锈病(禾柄锈菌)。在一些实施方案中,植株为玉米植株。在一些实施方案中,植株为大豆植株。在一些实施方案中,植株为棉花植株。在一些实施方案中,植株为小麦植株。在一些实施方案中,植株为高粱植株。在一些实施方案中,植株为芥花植株。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
20.若干实施方案涉及一种提供具有改善的抗病性的玉米植株的方法,所述方法包括:(a)向一个或多个植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在rp1抗病性基因座中的至少一个靶序列中引入基因组修饰;(b)针对同源染色体上的rp1抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组rp1抗病性基因座的玉米细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的玉米细胞获得的玉米植株以及其子代进行测试;以及(d)选择对一种或多种选自以下的疾病具有改善的抗性的玉米植株:炭疽茎腐病(禾生刺盘孢)、镰刀菌穗腐病(轮枝镰刀菌)、镰刀菌茎腐病(镰刀菌属)、赤霉穗腐病(念珠状赤霉)、赤霉茎腐病(玉米赤霉)、戈斯氏萎蔫病和叶枯病(苜蓿萎蔫病菌)、灰色叶斑病(玉蜀黍尾孢菌、玉米尾孢菌)以及北方玉米叶枯病(大斑凸脐蠕孢)。若干实施方案涉及一种提供具有改善的抗病性的玉米植株的方法,所述方法包括:(a)向一个或多个玉米细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在rp1抗病性基因座中的第一靶序列以及rp1抗病性基因座中的第二靶序列中引入基因组修饰,其中第一靶序列和第二靶序列位于同源染色体上;(b)针对同源染色体上的rp1抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组rp1抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择对选自以下的疾病具有改善的抗性的玉米植株:炭疽茎腐病(禾生刺盘孢)、镰刀菌穗腐病(轮枝镰刀菌)、镰刀菌茎腐病(镰刀菌属)、赤霉穗腐病(念珠状赤霉)、赤霉茎腐病(玉米赤霉)、戈斯氏萎蔫病和叶枯病(苜蓿萎蔫病菌)、灰色叶斑病(玉蜀黍尾孢菌、玉米尾孢菌)以及北方玉米叶枯病(大斑凸脐蠕孢)。若干实施方案涉及一种提供具有改善的抗病性的玉米植株的方法,所述方法包括:(a)向一个或多个植物细胞提供第一位点-特异性基因组修饰酶和第二位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在rp1抗病性基因座中的第一靶序列中引入基因组修饰,所述第二位点-特异性基因组修饰酶在rp1抗病性基因座中的第二靶序列中引入基因组修饰,其中第一靶序列和第二靶序列位于同源染色体上;(b)针对同源染色体上的rp1抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组rp1抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的玉米细胞获得的玉米植株以及其子代进行测试;以及(d)选择对选自以下的疾病具有改善的抗性的玉米植株:炭疽茎腐病(禾生刺盘孢)、镰刀菌穗腐病(轮枝镰刀菌)、镰刀菌茎腐病(镰刀菌属)、赤霉穗腐病(念珠状赤霉)、赤霉茎腐病(玉米赤霉)、戈斯氏萎蔫病和叶枯病(苜蓿萎蔫病菌)、灰色叶斑病(玉蜀黍尾孢菌、玉米尾孢菌)以及北方玉米叶枯病(大斑凸脐蠕孢)。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组rp1抗病性基因座与任一亲代中的rp1抗病性基因座相比具有增加的基因数目。在一些实施方案中,重组rp1抗病性基因座与任一亲代中的rp1抗病性基因座相比具有减少的基因数目。在一些实施方案中,重组rp1抗病性基因座与任一亲代中的rp1抗病性基因座相比具有不同的基因组合。在一些实施方案中,靶序列为基因的。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,靶序列是选自包含以下各项的组中的一种或多种:seq id no:3、seq id no:4、seq id no:5、seq id no:6、seq id no:7、seq id no:8以及seq id no:9。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与同源染色体上的基因
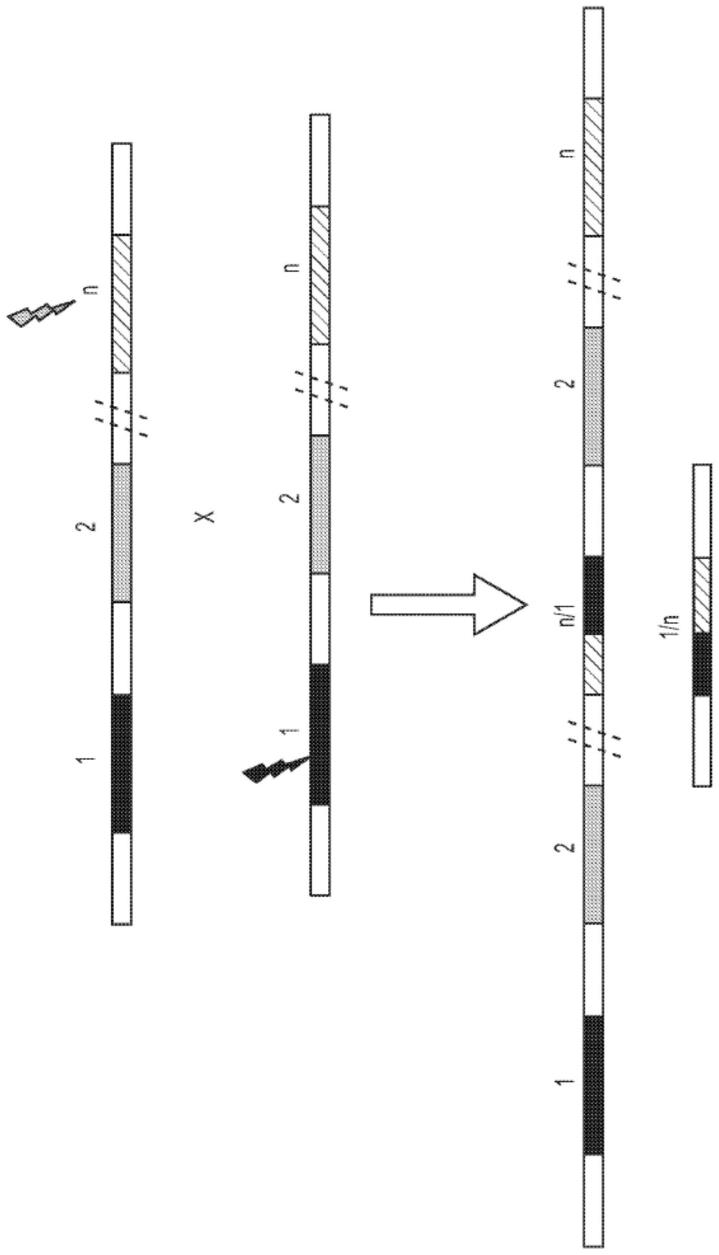
组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与同源染色体上不同位置的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,重组是在同源rp1抗病性基因座之间。在一些实施方案中,重组是在异源rp1抗病性基因座之间。在一些实施方案中,重组是在部分同源rp1抗病性基因座之间。在一些实施方案中,重组是在旁系同源rp1抗病性基因座之间。在一些实施方案中,重组是在相同rp1抗病性基因座之间。在一些实施方案中,同源染色体是来自性别上不相容的亲本基因组。在一些实施方案中,第一亲本基因组为普通小麦(小麦),并且第二亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦。在一些实施方案中,第一亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦,并且第二亲本基因组为普通小麦(小麦)。在一些实施方案中,第一亲本基因组为陆地棉(棉花),并且第二亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉。在一些实施方案中,第一亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉,并且第二亲本基因组为陆地棉(棉花)。在一些实施方案中,同源染色体为不同植物种类。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
21.若干实施方案涉及一种提供具有改善的抗病性的大豆植株的方法,所述方法包括:(a)向一个或多个植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在rpp1抗病性基因座中的至少一个靶序列中引入基因组修饰;(b)针对同源染色体上的rpp1抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组rpp1抗病性基因座的大豆细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的大豆细胞获得的大豆植株以及其子代进行测试;以及(d)选择对一种或多种选自以下的疾病具有改善的抗性的大豆植株:猝死综
合征(sds)、疫霉根腐病、疫霉茎腐病、镰刀菌根腐病、丝核菌根腐病、炭腐病、大豆胞囊线虫病(scn)、核盘霉茎腐病(白霉)、褐茎腐病(bsr)、荚枯和茎枯病、茎溃疡、炭疽病、绿茎综合征、大豆锈病、壳针孢褐斑病、细菌性枯病、霜霉病、尾孢菌叶枯病、大豆灰斑病、白粉病、大豆花叶病毒以及豆荚斑驳病毒。若干实施方案涉及一种提供具有改善的抗病性的大豆植株的方法,所述方法包括:(a)向一个或多个大豆细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在rpp1抗病性基因座中的第一靶序列以及rpp1抗病性基因座中的第二靶序列中引入基因组修饰,其中第一靶序列和第二靶序列位于同源染色体上;(b)针对同源染色体上的rpp1抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组rpp1抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择对选自以下的疾病具有改善的抗性的大豆植株:猝死综合征(sds)、疫霉根腐病、疫霉茎腐病、镰刀菌根腐病、丝核菌根腐病、炭腐病、大豆胞囊线虫病(scn)、核盘霉茎腐病(白霉)、褐茎腐病(bsr)、荚枯和茎枯病、茎溃疡、炭疽病、绿茎综合征、大豆锈病、壳针孢褐斑病、细菌性枯病、霜霉病、尾孢菌叶枯病、大豆灰斑病、白粉病、大豆花叶病毒以及豆荚斑驳病毒。若干实施方案涉及一种提供具有改善的抗病性的大豆植株的方法,所述方法包括:(a)向一个或多个植物细胞提供第一位点-特异性基因组修饰酶和第二位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在rpp1抗病性基因座中的第一靶序列中引入基因组修饰,所述第二位点-特异性基因组修饰酶在rpp1抗病性基因座中的第二靶序列中引入基因组修饰,其中第一靶序列和第二靶序列位于同源染色体上;(b)针对同源染色体上的rpp1抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组rpp1抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的大豆细胞获得的大豆植株以及其子代进行测试;以及(d)选择对选自以下的疾病具有改善的抗性的大豆植株:猝死综合征(sds)、疫霉根腐病、疫霉茎腐病、镰刀菌根腐病、丝核菌根腐病、炭腐病、大豆胞囊线虫病(scn)、核盘霉茎腐病(白霉)、褐茎腐病(bsr)、荚枯和茎枯病、茎溃疡、炭疽病、绿茎综合征、大豆锈病、壳针孢褐斑病、细菌性枯病、霜霉病、尾孢菌叶枯病、大豆灰斑病、白粉病、大豆花叶病毒以及豆荚斑驳病毒。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组rpp1抗病性基因座与任一亲代中的rpp1抗病性基因座相比具有增加的基因数目。在一些实施方案中,重组rpp1抗病性基因座与任一亲代中的rpp1抗病性基因座相比具有减少的基因数目。在一些实施方案中,重组rpp1抗病性基因座与任一亲代中的rpp1抗病性基因座相比具有不同的基因组合。在一些实施方案中,靶序列为基因的。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,靶序列是选自包含以下各项的组中的一种或多种:seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17以及seq id no:18。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与同源染色体上的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与同源染色体上不同位置的基因组基因座的至少约100bp、至少约
150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,重组是在同源rpp1抗病性基因座之间。在一些实施方案中,重组是在异源rpp1抗病性基因座之间。在一些实施方案中,重组是在部分同源rpp1抗病性基因座之间。在一些实施方案中,重组是在旁系同源rpp1抗病性基因座之间。在一些实施方案中,重组是在相同rpp1抗病性基因座之间。在一些实施方案中,同源染色体是来自性别上不相容的亲本基因组。在一些实施方案中,第一亲本基因组为普通小麦(小麦),并且第二亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦。在一些实施方案中,第一亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦,并且第二亲本基因组为普通小麦(小麦)。在一些实施方案中,第一亲本基因组为陆地棉(棉花),并且第二亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉。在一些实施方案中,第一亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉,并且第二亲本基因组为陆地棉(棉花)。在一些实施方案中,同源染色体为不同植物种类。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
22.若干实施方案涉及一种提供具有改善的抗病性的大豆植株的方法,所述方法包括:(a)向一个或多个植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在rps1抗病性基因座中的至少一个靶序列中引入基因组修饰;(b)针对同源染色体上的rps1抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组rps1抗病性基因座的大豆细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的大豆细胞获得的大豆植株以及其子代进行测试;以及(d)选择对一种或多种选自以下的疾病具有改善的抗性的大豆植株:猝死综合征(sds)、疫霉根腐病、疫霉茎腐病、镰刀菌根腐病、丝核菌根腐病、炭腐病、大豆胞囊线虫病(scn)、核盘霉茎腐病(白霉)、褐茎腐病(bsr)、荚枯和茎枯病、茎溃疡、炭疽病、绿茎综合征、大豆锈病、壳针孢褐斑病、细菌性枯病、霜霉病、尾孢菌叶枯病、大豆灰斑病、白粉病、大豆花叶病毒以及豆荚斑驳病毒。若干实施方案涉及一种提供具有改善的抗病性的大豆植株
的方法,所述方法包括:(a)向一个或多个大豆细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在rps1抗病性基因座中的第一靶序列以及rps1抗病性基因座中的第二靶序列中引入基因组修饰,其中第一靶序列和第二靶序列位于同源染色体上;(b)针对同源染色体上的rps1抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组rps1抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择对选自以下的疾病具有改善的抗性的大豆植株:猝死综合征(sds)、疫霉根腐病、疫霉茎腐病、镰刀菌根腐病、丝核菌根腐病、炭腐病、大豆胞囊线虫病(scn)、核盘霉茎腐病(白霉)、褐茎腐病(bsr)、荚枯和茎枯病、茎溃疡、炭疽病、绿茎综合征、大豆锈病、壳针孢褐斑病、细菌性枯病、霜霉病、尾孢菌叶枯病、大豆灰斑病、白粉病、大豆花叶病毒以及豆荚斑驳病毒。若干实施方案涉及一种提供具有改善的抗病性的大豆植株的方法,所述方法包括:(a)向一个或多个植物细胞提供第一位点-特异性基因组修饰酶和第二位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在rps1抗病性基因座中的第一靶序列中引入基因组修饰,所述第二位点-特异性基因组修饰酶在rps1抗病性基因座中的第二靶序列中引入基因组修饰,其中第一靶序列和第二靶序列位于同源染色体上;(b)针对同源染色体上的rps1抗病性基因座之间的不对称重组进行筛选,以鉴定包含重组rps1抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的大豆细胞获得的大豆植株以及其子代进行测试;以及(d)选择对选自以下的疾病具有改善的抗性的大豆植株:猝死综合征(sds)、疫霉根腐病、疫霉茎腐病、镰刀菌根腐病、丝核菌根腐病、炭腐病、大豆胞囊线虫病(scn)、核盘霉茎腐病(白霉)、褐茎腐病(bsr)、荚枯和茎枯病、茎溃疡、炭疽病、绿茎综合征、大豆锈病、壳针孢褐斑病、细菌性枯病、霜霉病、尾孢菌叶枯病、大豆灰斑病、白粉病、大豆花叶病毒以及豆荚斑驳病毒。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组rps1抗病性基因座与任一亲代中的rps1抗病性基因座相比具有增加的基因数目。在一些实施方案中,重组rps1抗病性基因座与任一亲代中的rps1抗病性基因座相比具有减少的基因数目。在一些实施方案中,重组rps1抗病性基因座与任一亲代中的rps1抗病性基因座相比具有不同的基因组合。在一些实施方案中,靶序列为基因的。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,靶序列是选自包含以下各项的组中的一种或多种:seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26、seq id no:27、seq id no:28、seq id no:29、seq id no:30、seq id no:31以及seq id no:32。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与同源染色体上的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与同源染色体上不同位置的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,重组是在同源rps1抗病性基因座之间。在一些实施方案中,重组是在异
源rps1抗病性基因座之间。在一些实施方案中,重组是在部分同源rps1抗病性基因座之间。在一些实施方案中,重组是在旁系同源rps1抗病性基因座之间。在一些实施方案中,重组是在相同rps1抗病性基因座之间。在一些实施方案中,同源染色体是来自性别上不相容的亲本基因组。在一些实施方案中,同源染色体为不同植物种类。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
23.若干实施方案涉及一种提供具有改善的线虫抗性的大豆植株的方法,所述方法包括:(a)向一个或多个植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在rhg1大豆胞囊线虫抗性基因座中的至少一个靶序列中引入基因组修饰;(b)针对同源染色体上的rhg1大豆胞囊线虫抗性基因座之间的不对称重组进行筛选,以鉴定包含重组rhg1大豆胞囊线虫抗性基因座的大豆细胞;(c)针对改善的线虫抗性对从步骤(b)中鉴定的大豆细胞获得的大豆植株以及其子代进行测试;以及(d)选择具有改善的线虫抗性的大豆植株。若干实施方案涉及一种提供具有改善的线虫抗性的大豆植株的方法,所述方法包括:(a)向一个或多个大豆细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶在rhg1大豆胞囊线虫抗性基因座中的第一靶序列和rhg1大豆胞囊线虫抗性基因座中的第二靶序列中引入基因组修饰,其中第一靶序列和第二靶序列位于同源染色体上;(b)针对同源染色体上的rhg1大豆胞囊线虫抗性基因座之间的不对称重组进行筛选,以鉴定包含重组rhg1大豆胞囊线虫抗性基因座的植物细胞;(c)针对改善的线虫抗性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择具有改善的线虫抗性的大豆植株。若干实施方案涉及一种提供具有改善的线虫抗性的大豆植株的方法,所述方法包括:(a)向一个或多个植物细胞提供第一位点-特异性基因组修饰酶和第二位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在rhg1大豆胞囊线虫抗性基因座中的第一靶序列中引入基因组修饰,所述第二位点-特异性基因组修饰酶在rhg1大豆胞囊线虫抗性基因座中的第二靶序列中引入基因组修饰,其中第一靶序列和第二靶序列位于同源染色体上;(b)针对同源染色体上的rhg1大豆胞囊线虫抗性基因座之间的不对称重组进行筛选,以鉴定包含重组rhg1大豆胞囊线虫抗性基因座的大豆细胞;(c)针对改善的线虫抗性对从步
骤(b)中鉴定的大豆细胞获得的大豆植株以及其子代进行测试;以及(d)选择具有改善的线虫抗性的大豆植株。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组rhg1大豆胞囊线虫抗性基因座与任一亲代中的rhg1大豆胞囊线虫抗性基因座相比具有增加的基因数目。在一些实施方案中,重组rhg1大豆胞囊线虫抗性基因座与任一亲代中的rhg1大豆胞囊线虫抗性基因座相比具有减少的基因数目。在一些实施方案中,重组rhg1大豆胞囊线虫抗性基因座与任一亲代中的rhg1大豆胞囊线虫抗性基因座相比具有不同的基因组合。在一些实施方案中,靶序列为基因的。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,靶序列是选自包含以下各项的组中的一种或多种:seq id no:69、seq id no:70、seq id no:71、seq id no:72、seq id no:73、seq id no:74、seq id no:75以及seq id no:76。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与同源染色体上的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与同源染色体上不同位置的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,重组是在同源rhg1大豆胞囊线虫抗性基因座之间。在一些实施方案中,重组是在异源rhg1大豆胞囊线虫抗性基因座之间。在一些实施方案中,重组是在部分同源rhg1大豆胞囊线虫抗性基因座之间。在一些实施方案中,重组是在旁系同源rhg1大豆胞囊线虫抗性基因座之间。在一些实施方案中,重组是在相同rhg1大豆胞囊线虫抗性基因座之间。在一些实施方案中,同源染色体是来自性别上不相容的亲本基因组。在一些实施方案中,同源染色体为不同植物种类。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
24.一种从近交系产生具有改变的抗病性基因座的植株的方法,所述方法包括向植物细胞提供位点-特异性基因组修饰酶,其中位点-特异性基因组修饰酶在抗病性基因座的至少一个靶序列中引入基因组修饰,由此在抗病性基因座内诱导同源染色体之间的不对称重组;以及从植物细胞生长出具有改变的抗病性基因座的植株。一种从近交系产生具有改变的抗病性基因座的植株的方法,所述方法包括向植物细胞提供第一位点-特异性基因组修饰酶和第二位点-特异性基因组修饰酶,其中第一位点-特异性基因组修饰酶在抗病性基因座的第一靶序列处引入基因组修饰,并且所述第二位点-特异性基因组修饰酶在抗病性基因座中的第二靶序列处引入基因组修饰,由此诱导同源染色体上的第一和第二靶序列之间的不对称重组;以及从植物细胞生长出具有改变的抗病性基因座的植株。一种从近交系产生具有改变的抗病性基因座的植株的方法,所述方法包括向植物细胞提供位点-特异性基因组修饰酶,其中位点-特异性基因组修饰酶在抗病性基因座的至少两个靶序列中引入基因组修饰,由此在抗病性基因座中诱导序列的缺失;以及从植物细胞生长出具有改变的抗病性基因座的植株。在一些实施方案中,具有改变的抗病性基因座的植株不需要回交来实现基因同一性。在一些实施方案中,具有改变的抗病性基因座的植株与不具有改变的抗病性基因座的近交系的植株相比具有改善的抗病性。在一些实施方案中,抗病性基因座编码一种或多种核苷酸结合位点富含亮氨酸重复序列(nbs-lrr)抗病蛋白。在一些实施方案中,植株为玉米植株并且抗病性基因座为rp1。在一些实施方案中,植株为大豆植株并且抗病性基因座为rpp1。在一些实施方案中,植株为大豆植株并且抗病性基因座为rps1。在一些实施方案中,植株为大豆植株并且抗病性基因座为rhg1。在一些实施方案中,植株为大豆植株并且抗病性基因座为rgh4。在一些实施方案中,具有改变的抗病性基因座的植株对一种或多种选自由以下组成的组的疾病具有改善的抗性:炭疽茎腐病(禾生刺盘孢)、镰刀菌穗腐病(轮枝镰刀菌)、镰刀菌茎腐病(镰刀菌属)、赤霉穗腐病(念珠状赤霉)、赤霉茎腐病(玉米赤霉)、戈斯氏萎蔫病和叶枯病(苜蓿萎蔫病菌)、灰色叶斑病(玉蜀黍尾孢菌、玉米尾孢菌)、北方玉米叶枯病(大斑凸脐蠕孢)、猝死综合征(茄病镰刀菌)、亚洲大豆锈病(豆薯层锈菌)、疫霉根腐和茎腐病(大豆疫霉)、根结线虫病(根结线虫属)、大豆胞囊线虫病(大豆胞囊线虫)、肾形线虫病(肾状轮线虫)、根结线虫病(南方根结线虫)、镰刀菌萎蔫病(尖孢镰刀菌萎蔫专化型)、轮枝孢菌萎蔫病(大丽轮枝孢菌)、镰刀菌首枯病(禾谷镰刀菌)、镰刀菌苗枯病(镰刀菌属、颖枯壳针孢)、镰刀菌叶斑病(雪腐小画线壳)以及茎锈病(禾柄锈菌)。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,改变的抗病性基因座与任一亲本基因组中的抗病性基因座相比具有增加的基因数目。在一些实施方案中,改变的抗病性基因座与任一亲本基因组中的抗病性基因座相比具有减少的基因数目。在一些实施方案中,改变的抗病性基因座与任一亲本基因组中的抗病性基因座相比具有新的基因组合。在一些实施方案中,亲本基因组中的抗病性基因座为相同的。在一些实施方案中,靶序列为基因的。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与同源染色体上的抗病性基因座中的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约
600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,靶序列位于基因组基因座中,所述基因组基因座与同源染色体上的抗病性基因座中的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的,其中靶序列和同源染色体上的抗病性基因座中的基因组基因座位于基因组中的不同位置。在一些实施方案中,植株为玉米植株。在一些实施方案中,植株为大豆植株。在一些实施方案中,植株为棉花植株。在一些实施方案中,植株为小麦植株。在一些实施方案中,植株为高粱植株。在一些实施方案中,植株为芥花植株。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。在一些实施方案中,抗病性基因座为rp1,并且靶序列是选自由以下组成的组中的一种或多种:seq id no:3、seq id no:4、seq id no:5、seq id no:6、seq id no:7、seq id no:8以及seq id no:9。在一些实施方案中,抗病性基因座为rpp1,并且靶序列是选自由以下组成的组中的一种或多种:seq id no:12、seq id no:13、seq id no:14、seq id no:15、seq id no:16、seq id no:17以及seq id no:18。在一些实施方案中,抗病性基因座为rps1,并且靶序列是选自由以下组成的组中的一种或多种:seq id no:21、seq id no:22、seq id no:23、seq id no:24、seq id no:25、seq id no:26、seq id no:27、seq id no:28、seq id no:29、seq id no:30、seq id no:31以及seq id no:32。在一些实施方案中,抗病性基因座为rhg1,并且靶序列是选自由以下组成的组中的一种或多种:seq id no:69、seq id no:70、seq id no:71、seq id no:72、seq id no:73、seq id no:74、seq id no:75以及seq id no:76。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
25.若干实施方案涉及一种增强所选基因组基因座处的重组的方法,所述方法包括向植物细胞提供至少一种位点-特异性基因组修饰酶,所述至少一种位点-特异性基因组修饰酶在第一基因组基因座中引入基因组修饰,由此诱导第一基因组基因座与第二基因组基因座之间的重组,其中所述至少一种位点-特异性基因组修饰酶不在第二基因组基因座处引入基因组修饰;以及选择包含第一基因组基因座与第二基因组基因座之间的重组事件的至少一个植物细胞。若干实施方案涉及一种增强所选基因组基因座处的重组的方法,所述方
法包括向植物细胞提供至少一种位点-特异性基因组修饰酶,所述至少一种位点-特异性基因组修饰酶在第一基因组基因座和第二基因组基因座处引入基因组修饰,由此诱导第一基因组基因座与第二基因组基因座之间的重组;以及选择包含第一基因组基因座与第二基因组基因座之间的重组事件的至少一个植物细胞。若干实施方案涉及一种增强所选基因组基因座处的重组的方法,所述方法包括向细胞提供第一位点-特异性基因组修饰酶和第二位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在第一基因组基因座处引入基因组修饰,所述第二位点-特异性基因组修饰酶在第二基因组基因座处引入基因组修饰,由此诱导第一基因组基因座与第二基因组基因座之间的重组;以及选择包含第一基因组基因座与第二基因组基因座之间的重组事件的至少一个子代。在一些实施方案中,第一和第二基因组基因座呈顺式。在一些实施方案中,第一和第二基因组基因座呈反式。在一些实施方案中,第一和第二基因组基因座为同源物。在一些实施方案中,第一和第二基因组基因座为旁系同源物。在一些实施方案中,第一和第二基因组基因座为部分同源物。在一些实施方案中,第一和第二基因组基因座为相同的。在一些实施方案中,第一基因组基因座和第二基因组基因座位于同源染色体上。在一些实施方案中,第一基因组基因座和第二基因组基因座位于非同源染色体上。在一些实施方案中,第一基因组基因座和第二基因组基因座位于部分同源染色体上。在一些实施方案中,第一和第二基因组基因座拥有至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%或100%的序列同一性。在一些实施方案中,第一基因组基因座和第二基因组基因座是定位于同源染色体上。在一些实施方案中,第一基因组基因座和第二基因组基因座是定位于非同源染色体上。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,重组为不对称的。在一些实施方案中,重组为对称的。在一些实施方案中,第一靶序列和/或第二靶序列为基因的。在一些实施方案中,第一靶序列和/或第二靶序列位于基因间区内。在一些实施方案中,第一靶序列位于基因组基因座中,所述基因组基因座与含有第二靶序列的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,第一靶序列位于基因组基因座中,所述基因组基因座与含有第二靶序列的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的,其中含有第一靶序列的基因组基因座和含有第二靶序列的基因组基因座位于基因组中的对应位置。在一些实施方案中,第一靶序列位于基因组基因座中,所述基因组基因座与含有第二靶序列的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的,其中含有第一靶序列的基因组基因座和含有第二靶序列的基因组基因座不位于基因组中的对应位置。在一些实施方案中,第一靶
序列与第二靶序列具有至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%或100%的序列同一性。在一些实施方案中,第一基因组基因座和第二基因组基因座中的一个或多个包含一个或多个独立地选自由以下组成的组的基因组区:基因、串联重复基因阵列、增强子、抑制子、启动子、终止序列、剪接受体序列、剪接供体序列、内含子、外显子、sirna以及数量性状基因座(qtl)。在一些实施方案中,包含第一基因组基因座与第二基因组基因座之间的重组事件的一个植物细胞的子代展现对一种或多种选自以下的疾病的抗性:炭疽茎腐病(禾生刺盘孢)、镰刀菌穗腐病(轮枝镰刀菌)、镰刀菌茎腐病(镰刀菌属)、赤霉穗腐病(念珠状赤霉)、赤霉茎腐病(玉米赤霉)、戈斯氏萎蔫病和叶枯病(苜蓿萎蔫病菌)、灰色叶斑病(玉蜀黍尾孢菌、玉米尾孢菌)、北方玉米叶枯病(大斑凸脐蠕孢)、猝死综合征(茄病镰刀菌)、亚洲大豆锈病(豆薯层锈菌)、疫霉根腐和茎腐病(大豆疫霉)、根结线虫病(根结线虫属)、大豆胞囊线虫病(大豆胞囊线虫)、肾形线虫病(肾状轮线虫)、根结线虫病(南方根结线虫)、镰刀菌萎蔫病(尖孢镰刀菌萎蔫专化型)、轮枝孢菌萎蔫病(大丽轮枝孢菌)、镰刀菌首枯病(禾谷镰刀菌)、镰刀菌苗枯病(镰刀菌属、颖枯壳针孢)、镰刀菌叶斑病(雪腐小画线壳)以及茎锈病(禾柄锈菌)。在一些实施方案中,植株为玉米植株。在一些实施方案中,植株为大豆植株。在一些实施方案中,植株为棉花植株。在一些实施方案中,植株为小麦植株。在一些实施方案中,植株为高粱植株。在一些实施方案中,植株为芥花植株。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
26.若干实施方案涉及一种将感兴趣的基因组基因座种质渗入至所选种质中的方法,所述方法包括产生包含第一亲本基因组和第二亲本基因组的植物细胞,所述第一亲本基因组包含感兴趣的基因组基因座,所述第二亲本基因组包含所选种质;向植物细胞提供第一位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在第一亲本基因组中在邻近感兴趣的基因组基因座的靶序列处引入基因组修饰,由此诱导第一亲本基因组与第二亲本基因组之间的重组;以及选择包含至少一个重组染色体的至少一个子代,所述至少一个重组染色体包含所选种质和感兴趣的基因组基因座。若干实施方案涉及一种将感兴趣的基
因组基因座种质渗入至所选种质中的方法,所述方法包括产生包含第一亲本基因组和第二亲本基因组的植物细胞,所述第一亲本基因组包含感兴趣的基因组基因座,所述第二亲本基因组包含所选种质;向植物细胞提供第一位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在第一亲本基因组中在邻近感兴趣的基因组基因座的靶序列处引入基因组修饰并且在第二亲本基因组中的靶位点处引入基因组修饰,由此诱导第一亲本基因组与第二亲本基因组之间的重组;以及选择包含至少一个重组染色体的至少一个子代,所述至少一个重组染色体包含所选种质和感兴趣的基因组基因座。若干实施方案涉及一种将感兴趣的基因组基因座种质渗入至所选种质中的方法,所述方法包括产生包含第一亲本基因组和第二亲本基因组的植物细胞,所述第一亲本基因组包含感兴趣的基因组基因座,所述第二亲本基因组包含所选种质;向植物细胞提供第一位点-特异性基因组修饰酶和第二位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在第一亲本基因组中在邻近感兴趣的基因组基因座的靶序列处引入基因组修饰,所述第二位点-特异性基因组修饰酶在第一亲本基因组中在邻近所述基因组基因座的第二靶序列处引入基因组修饰,其中第二靶序列相较于第一位点-特异性基因组修饰酶的靶序列位于基因组中感兴趣的基因组基因座的相对侧,由此诱导第一亲本基因组与第二亲本基因组之间的重组;以及选择包含至少一个重组染色体的至少一个植物细胞,所述至少一个重组染色体包含所选种质和感兴趣的基因组基因座。若干实施方案涉及一种将感兴趣的基因组基因座种质渗入至所选种质中的方法,所述方法包括产生包含第一亲本基因组和第二亲本基因组的植物细胞,所述第一亲本基因组包含感兴趣的基因组基因座,所述第二亲本基因组包含所选种质;向植物细胞提供第一位点-特异性基因组修饰酶和第二位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在第一亲本基因组中在邻近感兴趣的基因组基因座的靶序列处引入基因组修饰并且在第二亲本基因组中的靶位点处引入基因组修饰,并且进一步向植物细胞中引入第二位点-特异性基因组修饰酶,所述第二位点-特异性基因组修饰酶在第一亲本基因组中在邻近所述基因组基因座的第二靶序列处引入基因组修饰,其中第二靶序列相较于第一位点-特异性基因组修饰酶的靶序列位于基因组中感兴趣的基因组基因座的相对侧,由此诱导第一亲本基因组与第二亲本基因组之间的重组;以及选择包含至少一个重组染色体的至少一个植物细胞,所述至少一个重组染色体包含所选种质和感兴趣的基因组基因座。在一些实施方案中,第二位点-特异性基因组修饰酶在第二亲本基因组中的靶序列处引入基因组修饰。在一些实施方案中,重组为不对称的。在一些实施方案中,重组为对称的。在一些实施方案中,感兴趣的基因组基因座包含一个或多个独立地选自由以下组成的组的基因组区:基因、串联重复基因阵列、多基因家族、增强子、抑制子、启动子、终止序列、剪接受体序列、剪接供体序列、内含子、外显子、sirna、编码非编码rna的序列、微rna、转基因以及数量性状基因座(qtl)。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,靶序列为基因的。在一些实施方案中,靶序列位于基因间区内。在一些实施方案中,靶序列位于第一亲本基因组的基因组基因座中,所述基因组基因座与第二亲本基因组的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约
500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的。在一些实施方案中,靶序列位于第一亲本基因组的基因组基因座中,所述基因组基因座与第二亲本基因组的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的,其中第一亲本基因组的基因组基因座和第二亲本基因组的基因组基因座是定位于对应位置。在一些实施方案中,靶序列位于第一亲本基因组的基因组基因座中,所述基因组基因座与第二亲本基因组的基因组基因座的至少约100bp、至少约150bp、至少约200bp、至少约250bp、至少约300bp、至少约350bp、至少约400bp、至少约450bp、至少约500bp、至少约600bp、至少约700bp、至少约800bp、至少约900bp或至少约1000bp为同源的,其中第一亲本基因组的基因组基因座和第二亲本基因组的基因组基因座不定位于对应位置,从而产生不对称重组。在一些实施方案中,第一亲本基因组和第二亲本基因组在性别上不相容。在一些实施方案中,第一亲本基因组和第二亲本基因组为不同的物种。在一些实施方案中,第一亲本基因组为普通小麦(小麦),并且第二亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦。在一些实施方案中,第一亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦,并且第二亲本基因组为普通小麦(小麦)。在一些实施方案中,第一亲本基因组为陆地棉(棉花),并且第二亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉。在一些实施方案中,第一亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉,并且第二亲本基因组为陆地棉(棉花)。在一些实施方案中,第一亲本基因组和/或第二亲本基因组为单倍体。在一些实施方案中,第一亲本基因组和/或第二亲本基因组为二倍体。在一些实施方案中,感兴趣的基因组基因座为rp1抗病性基因座。在一些实施方案中,感兴趣的基因组基因座为rpp1抗病性基因座。在一些实施方案中,感兴趣的基因组基因座为rps1抗病性基因座。在一些实施方案中,感兴趣的基因组基因座为rhg1抗病性基因座。在一些实施方案中,感兴趣的基因组基因座为rgh4抗病性基因座。在一些实施方案中,植株为玉米植株。在一些实施方案中,植株为大豆植株。在一些实施方案中,植株为棉花植株。在一些实施方案中,植株为小麦植株。在一些实施方案中,植株为高粱植株。在一些实施方案中,植株为芥花植株。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案
中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
27.若干实施方案涉及一种去除连锁累赘(linkage drag)的方法,所述方法包括产生包含第一亲本基因组和第二亲本基因组的植物细胞,其中第一亲本基因组包含与不想要的基因组基因座顺式连接的感兴趣的基因组基因座;向细胞提供第一位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在感兴趣的基因组基因座与不想要的基因组基因座之间引入基因组修饰,由此诱导第一亲本基因组与第二亲本基因组之间的重组,并且使感兴趣的基因组基因座与不想要的基因座分开;以及选择包含感兴趣的基因组基因座的至少一个子代。若干实施方案涉及一种去除连锁累赘的方法,所述方法包括产生包含第一亲本基因组和第二亲本基因组的植物细胞,其中第一亲本基因组包含与不想要的基因组基因座顺式连接的感兴趣的基因组基因座;向细胞提供第一位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶在感兴趣的基因组基因座与不想要的基因组基因座之间引入第一基因组修饰,并且相较于第一基因组修饰在不需要的基因组基因座的相对侧引入第二基因组修饰,由此诱导第一亲本基因组与第二亲本基因组之间的重组,并且去除不需要的基因座,而保留第二基因组修饰远端的第一亲本基因组的种质;以及选择包含感兴趣的基因组基因座的至少一个子代。在一些实施方案中,第二位点-特异性基因组修饰酶在第二亲本基因组中的靶序列处引入基因组修饰。在一些实施方案中,重组为不对称的。在一些实施方案中,重组为对称的。在一些实施方案中,感兴趣的基因组基因座包含一个或多个独立地选自由以下组成的组的基因组区:基因、串联重复基因阵列、多基因家族、增强子、抑制子、启动子、终止序列、剪接受体序列、剪接供体序列、内含子、外显子、sirna、编码非编码rna的序列、微rna、转基因以及数量性状基因座(qtl)。在一些实施方案中,基因组修饰为双链断裂(dsb)。在一些实施方案中,基因组修饰为单链断裂。在一些实施方案中,基因组修饰为重组酶介导的dna交换反应。在一些实施方案中,基因组修饰为转座酶介导的dna交换反应。在一些实施方案中,在减数分裂开始时发生基因组修饰。在一些实施方案中,第一亲本基因组和第二亲本基因组在性别上不相容。在一些实施方案中,第一亲本基因组和第二亲本基因组为不同的物种。在一些实施方案中,第一亲本基因组为普通小麦(小麦),并且第二亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦。在一些实施方案中,第一亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦,并且第二亲本基因组为普通小麦(小麦)。在一些实施方案中,第一亲本基因组为陆地棉(棉花),并且第二亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉。在一些实施方案中,第一亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉,并且第二亲本基因组为陆地棉(棉花)。在一些实施方案中,第一亲本基因组和/或第二亲本基因组为单倍体。在一些实施方案中,第一亲本基因组和/或第二亲本基因组为二倍体。在一些实施方案中,感兴趣的基因组基因座为rp1抗病性基因座。在一些实施方案中,感兴趣的基因组基因座为rpp1抗病性基因座。在一些实施方案中,感兴趣的基因组基因座为rps1抗病性基因座。在一些实施方案中,感兴趣的基因组基因座为rhg1抗病性基因座。在一些实施方案中,感兴趣的基因组基因座为rhg4抗病性基因座。在一些实施方案中,植株为玉米植株。在一些实施方案中,
植株为大豆植株。在一些实施方案中,植株为棉花植株。在一些实施方案中,植株为小麦植株。在一些实施方案中,植株为高粱植株。在一些实施方案中,植株为芥花植株。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。若干实施方案涉及一种将相排斥的基因组基因座偶联的方法,所述方法包括产生包含第一亲本基因组和第二亲本基因组的植物细胞,所述第一亲本基因组包含第一基因组基因座,所述第二亲本基因组包含第二基因组基因座,其中第一基因组基因座和第二基因位点为相排斥的;向细胞提供第一位点-特异性基因组修饰酶,所述第一位点-特异性基因组修饰酶靠近第一基因组基因座引入基因组修饰,由此诱导第一亲本基因组与第二亲本基因组之间的重组;以及选择在相同染色体上包含第一基因组基因座和第二基因组基因座的至少一个植物细胞。在一些实施方案中,第一基因组基因座和第二基因组基因座是定位于同源染色体上。在一些实施方案中,第一亲本基因组和第二亲本基因组在性别上不相容。在一些实施方案中,第一亲本基因组和第二亲本基因组为不同的物种。在一些实施方案中,感兴趣的第一基因组基因座和/或感兴趣的第二基因组基因座包含一个或多个独立地选自由以下组成的组的基因组区:基因、串联重复基因阵列、增强子、抑制子、启动子、终止序列、剪接受体序列、剪接供体序列、内含子、外显子、sirna以及数量性状基因座(qtl)。在一些实施方案中,第一亲本基因组和/或第二亲本基因组为单倍体。在一些实施方案中,第一亲本基因组和/或第二亲本基因组为二倍体。在一些实施方案中,第一亲本基因组为普通小麦(小麦),并且第二亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦。在一些实施方案中,第一亲本基因组是选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦,并且第二亲本基因组为普通小麦(小麦)。在一些实施方案中,第一亲本基因组为陆地棉(棉花),并且第二亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉。在一些实施方案中,第一亲本基因组是选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉,并且第二亲本基因组为陆地棉(棉花)。在一些实施方案中,感兴趣的基因组基因座为rp1抗病性基因座。在一些实施方案中,感兴趣的第一基因组基因座和/或感兴趣的第二基因组基因座为rpp1抗病性基因座。在一些实施方案中,感兴趣的第一基因
组基因座和/或感兴趣的第二基因组基因座为rps1抗病性基因座。在一些实施方案中,感兴趣的第一基因组基因座和/或感兴趣的第二基因组基因座为rhg1抗病性基因座。在一些实施方案中,感兴趣的第一基因组基因座和/或感兴趣的第二基因组基因座为rhg4抗病性基因座。在一些实施方案中,感兴趣的第一基因组基因座为rhg1,并且感兴趣的第二基因组基因座为rhg4。在一些实施方案中,植株为玉米植株。在一些实施方案中,植株为大豆植株。在一些实施方案中,植株为棉花植株。在一些实施方案中,植株为小麦植株。在一些实施方案中,植株为高粱植株。在一些实施方案中,植株为芥花植株。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为选自以下的核酸内切酶:大范围核酸酶、锌指核酸酶、转录激活因子样效应物核酸酶(talen)、argonaute、rna引导性核酸内切酶、i型crispr-cas系统、ii型crispr-cas系统或iii型crispr-cas系统。在一些实施方案中,位点-特异性基因组修饰酶为选自包含以下各项的组的crispr相关蛋白:cpf1、cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3以及csf4核酸酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为rna引导性重组酶。在一些实施方案中,位点-特异性基因组修饰酶为包含重组酶和crispr相关蛋白的融合蛋白。在一些实施方案中,重组酶为与dna识别基序连接的酪氨酸重组酶,或与dna识别基序连接的丝氨酸重组酶。在一些实施方案中,重组酶为cre重组酶、flp重组酶、tnp1重组酶、phic31整合酶、r4整合酶或tp-901整合酶。在一些实施方案中,位点-特异性基因组修饰酶为与dna结合域连接的转座酶。在一些实施方案中,第一亲本基因组和第二亲本基因组中的一种或多种是来自优良种质系。若干实施方案涉及通过根据上述方法所产生的植株、植物细胞或植物种子。
28.若干实施方案涉及一种产生新串联重复基因阵列的方法,所述方法包括使细胞与位点-特异性基因组修饰酶接触,所述位点-特异性基因组修饰酶裂解第一串联重复基因阵列中的至少一个靶序列,由此诱导与第二串联重复基因阵列的同源序列的不对称重组;以及选择包含新串联重复基因阵列的至少一个子代。在一些实施方案中,第一和第二串联重复基因阵列为相同的。在其他实施方案中,第一和第二串联重复基因阵列为不同的。在一些实施方案中,视重组位点而定,不对称重组产生两种新串联重复基因阵列。在一些实施方案中,不对称重组在串联重复基因中的至少一种中产生缺失。在一些实施方案中,细胞为植物细胞。在另一实施方案中,从选自近交植株或杂交植株的植株获得植物细胞。在其他实施方案中,细胞为哺乳动物细胞。
29.在一个方面中,位点-特异性基因组修饰酶是选自核酸内切酶、重组酶、转座酶、解旋酶或其任何组合。在另一方面中,核酸内切酶是选自大范围核酸酶、锌指核酸酶、talen、核酸引导性核酸内切酶、argonaute、crispr/cpf1系统以及crispr/cas9系统。
30.在一个方面中,串联重复基因编码选自以下的蛋白质:nbs-lrr抗病蛋白、病原体识别受体(prr)蛋白、种子贮藏蛋白、细胞壁组分扩展蛋白、f-框蛋白、abc转运蛋白、免疫球蛋白、丝氨酸-苏氨酸/酪氨酸蛋白质激酶以及核糖体rna。在rhg1的情况下,串联重复序列由三种基因组成:推定氨基酸转运蛋白、α-snap蛋白以及创伤诱导蛋白。
31.本公开还是关于一种改变植株的抗病性的方法,所述方法包括向植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶裂解一个或多个抗病性基因座中的保守区;以及从植物细胞生长出植株。在一个实施方案中,抗病性基因座包含nbs-lrr类别的抗病性基因。
32.本公开进一步有关于一种提供具有改善的抗病性的植株的方法,所述方法包括:(a)向一个或多个植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶裂解抗病性基因座中的靶序列;(b)针对同源染色体上的抗病性基因座之间的不对称重组对一个或多个植物细胞进行筛选,以鉴定包含重组抗病性基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择具有改善的抗病性的植株。此外,本公开提供一种通过此方法产生的植株、植物细胞、植物种子或植物部分。在一些实施方案中,植株具有至少一个重组抗病性基因座。在一些实施方案中,植株在抗病性基因座中的至少一个中具有至少一个缺失。在一个方面中,所述植株与具有抗病性基因座的亲本等位基因或抗病性基因座中的一个或多个基因的缺失的植株相比具有改善的抗病性。在一个实施方案中,抗病性基因座包含nbs-lrr类别的抗病性基因。
33.此外,本公开提供一种检测两个亲本染色体之间的同源重组的方法,所述方法包括:a.鉴定侧接两个亲本染色体中的每一个上的所靶向的感兴趣的基因座的限制核酸酶位点;b.提供对第一亲本染色体具特异性的pcr引物以及对第二亲本染色体具特异性的另一pcr引物;以及c.使用被设计成特定地识别第一染色体上的感兴趣的基因座的5
’‑
侧接区以及第二染色体上的感兴趣的基因座的3'-侧接区的独特接点的探针来鉴定指示发生重组的pcr产物。
34.附图简述
35.图1说明在串联重复基因阵列之间所诱导的不对称重组。在此图示中,基因阵列具有1、2、
…
、n个串联重复基因,并且在第一亲本染色体上的基因n与第二亲本染色体上的基因1之间发生不对称重组,从而使得形成新阵列,所述新阵列包含来自第一重组染色体上的亲代染色体(在此图示中为1、2、2、n)与第二重组染色体上的单个新基因(以1/n形式说明)的基因。闪电形状指示基因组修饰的位点。
36.图2说明在两个亲本染色体(左侧的一对)上反式排斥布置的两个基因组基因座之间所诱导的重组,并且在基因组修饰之后,在子代染色体(右侧的一对)上基因座呈顺式。闪电形状指示基因组修饰的位点。
37.图3说明通过反向pcr鉴定两个亲本染色体之间的重组的方法。在亲本染色体中的每一个上鉴定出侧接所靶向的感兴趣的基因组基因座的限制性核酸酶位点(由三角形指示)。对第一亲本染色体(
‘
a’)具特异性的pcr引物由黑色箭头指示。对第二亲本染色体(
‘
a’)具特异性的pcr引物由灰色箭头指示。诱导的双链断裂(由闪电形状指示)促进亲本染色体之间的重组,从而将限制性核酸内切酶位点与引物结合位点两者带到相同重组染色体上。对
‘
a’染色体上的所靶向的感兴趣的基因组基因座的5'-侧接区和
‘
a’染色体上的所靶向的感兴趣的基因组基因座的3'-侧接区的独特接点具特异性的探针由棒状物加上各末端的星形来指示。仅在发生重组的情况下观测到pcr产物。
38.图4说明关于在同源基因阵列中经历所诱导的不对称重组的转化体的高-通量基因筛选。在此实例中,亲本基因组中的每一个中存在含有3个基因的基因阵列。所述分析鉴
manual;animal cell culture(r.i.freshney编(1987));recombinant protein purification:principles and methods,18-1142-75,ge healthcare life sciences;c.n.stewart,a.touraev,v.citovsky,t.tzfira编(2011)plant transformation technologies(wiley-blackwell);以及r.h.smith(2013)plant tissue culture.techniques and experiments(academic press,inc.)。
46.本文中所引用的任何参考文献以全文引用的方式并入本文中。
47.除非上下文另外明确指示,否则如本文所用,单数形式“一个(种)(a/an)”和“所述”包括复数参考物。举例来说,术语“一种化合物”或“至少一种化合物”可包括多种化合物,包括其混合物。
48.如本文所用,术语“约”表明一个值包括用于测定一个值的方法的固有误差变异,或实验之间所存在的变异。
49.如本文所用,术语“位点-特异性基因组修饰酶”是指可以位点-特异性方式修饰核苷酸序列的任何酶。在本公开中,位点-特异性基因组修饰酶包括核酸内切酶、重组酶、转座酶、解旋酶以及其任何组合。
50.如本文所用,术语“重组”是指使得两个dna分子交换核苷酸序列的过程。在一些实施方案中,重组发生在两组亲本染色体之间。在一些实施方案中,重组发生在两个同源染色体之间。在一些实施方案中,重组发生在非同源染色体之间。在一些实施方案中,重组发生在部分同源染色体之间。在一些实施方案中,重组使得产生新的基因序列、基因数目、基因排列、等位基因或等位基因组合。
51.如本文所用,术语“重组事件”是指两个dna分子之间重组的情况。
52.如本文所用,术语“同源重组”是指由两个基因组基因座共同拥有的保守区处的核苷酸序列交换。同源重组包括对称同源重组和不对称同源重组。不对称同源重组也可称为不等重组。
53.用于检测重组的许多方法为本领域中已知的并且包括但不限于1)表型筛选,2)分子标记物技术,诸如通过或illumina/infinium技术进行的单核苷酸多态性-snp分析,3)南方墨点法(southern blot),以及4)测序。图3中说明了通过反向pcr鉴定两个亲本染色体之间的重组的方法的一个实例。在此方法中,在两个亲本染色体中的每一个上鉴定出侧接所靶向的感兴趣的基因的限制性核酸酶位点。这些限制性核酸酶位点可为相同或不同的。设计了对第一亲本染色体具特异性的pcr引物和对第二亲本染色体具特异性的另一pcr引物。诱导的双链断裂促进两个亲本染色体之间的重组,从而将限制性核酸内切酶位点与引物结合位点两者带到相同重组染色体上。仅在发生重组的情况下观测到pcr产物。
54.如本文所用,术语“重组率”是指两个基因组基因座之间将发生重组事件的概率。重组率可能会受许多因素的影响,包括但不限于两个基因组基因座之间的距离、出现基因座的染色体区(例如着丝粒区、端粒区)、转录活性、染色体倒位的存在以及其他因素。用于测量重组的方法包括但不限于作图群体中的连锁分析;以及如本公开中所描述的定量技术,诸如定量pcr(qpcr)或微滴数字pcr(ddpcr)。
55.如本文所用,术语“基因组基因座”是指染色体上的特定位置。基因组基因座可包含单个核苷酸、几个核苷酸、很多核苷酸、基因、基因的一部分、基因簇、多基因家族或基因组区中的基因阵列。在一些实施方案中,基因组基因座可包含串联重复基因阵列。在一些实
施方案中,基因组基因座可包含qtl。可根据特定序列来定义基因组基因座,或者可根据侧接标记物来定义基因组基因座。还可根据连锁图来定义基因组基因座。
56.如本文所用,术语“靶序列”是指位点-特异性基因组修饰酶所结合和/或对其发挥裂解、切口酶、重组酶或转座酶活性的核苷酸序列。靶序列可为基因的或非基因的。
57.如本文所用,术语“基因”意指基因组序列中对应于遗传单位的可定位区。基因可包括调控区,诸如启动子、增强子、5
’‑
未翻译区、内含子区、外显子区、3'-未翻译区、转录区以及其他功能序列区,这些区可以天然基因或转基因形式存在于植物或哺乳动物基因组中。视情况而定,术语“靶基因”可指被靶向而用于结合和/或裂解的基因的全长核苷酸序列或被靶向而用于结合和/或裂解的基因的一部分的核苷酸序列。靶基因可为内源性基因或转基因。
58.如本文所用,术语“事件”是指由细胞基因组dna的分子重组产生的基因组序列。重组包括同源重组、非同源重组、顺式重组、姐妹染色单体交换、多个染色体重排、对称和不对称重组。事件可发生在基因序列中或者事件可发生在基因间序列中。在一些实施方案中,事件可为新颖基因组序列。
59.如本文所用,“优良系”为已由育种和选择优良农艺学性能所产生的任何系。
60.如本文所用,术语“近交种”意指已关于遣传同质性进行育种的系。
61.如本文所用,术语“杂交种”意指至少两个在基因上不同的亲代之间交配的子代。交配方案的实例包括但不限于单交、改良的单交、加倍改良单交、三源杂交、改良的三源杂交以及双交,其中改良的杂交中的至少一个亲代为姐妹系之间杂交的子代。在一些实施方案中,可通过使普通小麦(小麦)与选自卵穗山羊草、两芒山羊草、三芒山羊草、粗山羊草、黑麦、野生二粒小麦、二粒小麦以及硬粒小麦的植株杂交来产生杂交种。在一些实施方案中,可通过使陆地棉(棉花)与选自史特啼棉、戴维逊氏棉、亚洲棉以及雷蒙德氏棉的植株杂交来形成杂交种。
62.如本文所用,术语“标记物”是指可用于辩别有机体、基因组、染色体、基因组基因座、基因或基因部分的可检测特征。此类特征的实例可包括基因标记物、蛋白质组成、蛋白质水平、油组成、油水平、碳水化合物组成、碳水化合物水平、脂肪酸组成、脂肪酸水平、氨基酸组成、氨基酸水平、生物聚合物、药物、淀粉组成、淀粉水平、可发酵淀粉、发酵产率、发酵效率、能量产率、二级化合物、代谢产物、形态特征以及农艺学特征。
63.如本文所用,术语“基因标记物”是指可用于辩别核酸的多态核酸序列或核酸特征。“多态性”为个体间在序列(特别是dna序列)或特征(诸如转录型态或甲基化模式)方面的变异。适用的多态性可包含但不限于一种或多种碱基变化、一种或多种核苷酸的插入或一种或多种核苷酸的缺失、单核苷酸多态性(snp)、简单序列重复(ssr)以及插入缺失(插入和缺失)。多态性可由核酸复制中的随机过程、通过诱变、由于移动基因组元件、由拷贝数变异以及在减数分裂(诸如不等交换)、基因组复制以及染色体断裂和融合过程期间产生。
64.如本文所用,术语“连锁”是指一种现象,其中相同染色体上的基因比如果其传递为独立的那么预期会偶然发生的更经常倾向于一起分离。举例来说,如果那么基因组基因座a具有基因“a”或“a”并且基因组基因座b具有基因“b”或“b”,那么具有aabb的第一亲本基因组与具有aabb的第二亲本基因组之间的杂交将产生四种可能的配子,其中基因被分离至ab、ab、ab以及ab中。零期望为将不存在向四种可能的基因型中的每一种中的独立相等分
离,即没有连锁,4个配子中每1个将具有各基因型。在此情况下,配子分离至不同于1/4的基因型中是归因于连锁。
65.如本文所用,“植物”是指整个植株、植物细胞、植物组织或植物种子。来源于植物的细胞或组织培养物可包含任何植物组件或植物器官(例如叶、茎、根等)、植物组织、种子、植物细胞和/或其子代。子代植株可以是来自任何后代,例如f1、f2、f3、f4、f5、f6、f7等。植物细胞为植物的生物细胞,所述生物细胞取自植物或通过培养来源于取自植物的细胞。
66.如本文所用,“植物基因组”是指植物细胞的核基因组、线粒体基因组或质体(例如叶绿体)基因组。在一些实施方案中,植物基因组可包含由雄性贡献的亲本基因组和由雌性贡献的亲本基因组。在一些实施方案中,植物基因组可仅包含一种亲本基因组。
67.术语“保守区”是指由具有至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%或100%的序列同一性的两个或更多个dna序列共同拥有的连续多核苷酸序列。保守区的长度可为至少10bp、至少20bp、至少30bp、至少40bp、至少50bp、至少60bp、至少70bp、至少80bp、至少90bp、至少100bp、至少200bp、至少300bp、至少400bp、至少500bp、至少600bp、至少700bp、至少800bp、至少900bp、至少1,000bp或更长。如本文所用,术语“同一性”当结合核酸使用时描述两个或更多个核苷酸序列之间的相似度。两个序列之间的“序列同一性”百分比可通过在比较窗上比较两个最佳比对序列,使得与用于最佳比对两个序列的参考序列(它不包含添加或缺失)相比在比较窗中的序列部分可包含添加或缺失(空位)来确定。如下计算所述百分比:确定在两个序列中出现相同核酸碱基或氨基酸残基的位置的数目以得到匹配位置的数目,用匹配位置的数目除以比较窗中的位置的总数目,并且将结果乘以100以得到序列同一性百分比。与参考序列相比在每个位置均相同的序列被认为与参考序列同一并且反之亦然。可使用任何适合的计算机程序来进行两个或更多个序列的比对。举例来说,广泛使用并被接受的用于进行序列比对的计算机程序为clustalw v1.6(thompson等(1994)nucl.acids res.,22:4673-4680)。
68.如本文所用,术语“旁系同源物”是指因基因组内的复制而相关的基因。在一些实施方案中,旁系同源物位于相同基因组中。在一些实施方案中,旁系同源物位于不同基因组中。在一些实施方案中,旁系同源物为同功能的。在一些实施方案中,旁系同源物为异功能的。
69.如本文所用,术语“同源物”是指与凭世系来自共同的祖先dna序列的第二基因有关的基因。术语同源物可适用于因物种形成事件而分离的基因之间的关系(直系同源物)或因基因复制事件而分离的基因之间的关系(旁系同源物)。在一些实施方案中,同源物位于相同基因组中。在一些实施方案中,同源物位于不同基因组中。在一些实施方案中,同源物为同功能的。在一些实施方案中,同源物为异功能的。
70.如本文所用,术语“部分同源物”是指因物种形成而产生并且通过异源多倍化在相同基因组中回到一起的一对基因。
71.如本文所用,术语“部分同源染色体”是指不同物种的拥有共同的祖先来源的染色体。
72.如本文所用,术语“串联重复”是指两个相同序列任何一次一个接着另一个地出现在染色体区段中。
73.如本文所用,术语“基因复制”是指含有基因的dna区的任何复制。
74.基因簇为在染色体上如邻居一般键联的两个或更多个基因的组。基因簇中的基因常常编码类似的多肽或蛋白质,它们全体共同拥有一般化的功能并且常常定位于距离彼此几千个碱基对以内。
75.基因家族为通过单个原始基因的复制而形成的若干类似基因的集合。基因家族可包含2个或更多个基因、3个或更多个基因、4个或更多个基因、5个或更多个基因、6个或更多个基因、7个或更多个基因、8个或更多个基因、9个或更多个基因、10个或更多个基因、15个或更多个基因、20个或更多个基因、25个或更多个基因、30个或更多个基因、35个或更多个基因、40个或更多个基因、45个或更多个基因、50个或更多个基因、55个或更多个基因、60个或更多个基因、65个或更多个基因、70个或更多个基因、75个或更多个基因、80个或更多个基因、90个或更多个基因、100个或更多个基因、150个或更多个基因、200个或更多个基因、250个或更多个基因、300个或更多个基因、350个或更多个基因、400个或更多个基因、450个或更多个基因或500个或更多个基因。在一些实施方案中,基因家族可包含串联重复基因阵列。
76.如本文所用,术语“串联重复基因阵列”是指因含有一个或多个基因的染色体区段的串联重复而形成的基因簇。在一些实施方案中,串联重复基因阵列可包含2个或更多个基因、3个或更多个基因、4个或更多个基因、5个或更多个基因、6个或更多个基因、7个或更多个基因、8个或更多个基因、9个或更多个基因、10个或更多个基因、15个或更多个基因、20个或更多个基因、25个或更多个基因、30个或更多个基因、35个或更多个基因、40个或更多个基因、45个或更多个基因、50个或更多个基因、55个或更多个基因、60个或更多个基因、65个或更多个基因、70个或更多个基因、75个或更多个基因、80个或更多个基因、90个或更多个基因、100个或更多个基因、150个或更多个基因、200个或更多个基因、250个或更多个基因、300个或更多个基因、350个或更多个基因、400个或更多个基因、450个或更多个基因或500个或更多个基因。串联重复基因阵列的实例包括但不限于编码nbs-lrr或prr抗病蛋白、种子贮藏蛋白、细胞壁组分扩展蛋白、f-框蛋白、abc转运蛋白、丝氨酸-苏氨酸/酪氨酸蛋白质激酶以及核糖体rna的基因。
77.如本文所用,术语“等位基因”是指相同基因或相同基因位点的许多替代形式中的一种。
78.如本文所用,术语“抗病性基因座”是指与植物中的疾病或病原体抗性相关的基因组区。抗病性基因座可包含一个或多个基因、基因家族、基因阵列或qtl,其编码赋予植株对至少一种疾病或病原体的抗性的蛋白质。在一个实施方案中,抗病性基因座包含一个或多个nbs-lrr抗病性基因,也称为nb-lrr基因、r基因、lrr基因。在另一实施方案中,抗病性基因座包含一个或多个prr抗病性基因。抗病性基因座可涵盖已知赋予病原体抗性的特定基因、基因簇、基因阵列和/或基因家族,例如rp1或rpp1或rps1。在另一实施方案中,抗病性基因座包含rgh1基因座。在另一实施方案中,抗病性基因座包含rgh4基因座。或者,抗病性基因座可涵盖基因组区,但赋予抗病性的实际基因/元件组成为未知的。
79.如本文所用,术语“免疫球蛋白基因”是指编码免疫球蛋白重链或轻链的区(例如vh、vl、ch区;铰链区;可变(v)区段、多样性(d)区段、连接(j)区段或其部分)的任何基因。术语免疫球蛋白包括任何免疫球蛋白类别,即igm、igg、igd、iga以及ige,以及任何同型。
80.如本文所用,“植物病原体”为导致植物或植物组织感染的任何有机体或药剂。常
见病原体包括病毒、细菌、真菌、昆虫以及线虫。
81.如本文所用,术语“数量性状基因座”或“qtl”是指与至少一种遗传背景中(例如至少一个育种群体中)的表型性状的差异表达相关的dna的区。qtl与构成所讨论的性状的基础的基因紧密相关。在一些情况下,构成性状的基础的元件、基因或基因集合的身份为未知的。与病原体抗性相关的qtl的实例为rhg4。
82.如本文所用,称为“单倍体”的细胞或基因组具有单个染色体集合,并且单倍体植株中的减少的染色体数目(n)与配子相同。
83.如本文所用,称为“二倍体”的细胞或基因组具有两组染色体并且染色体数目(2n)与接合子相同。
84.串联基因重复在重复基因簇的累积中起作用,重复基因簇的累积又有助于基因家族的扩大。串联重复基因座的实例包括编码以下各项的那些串联重复基因座:nbs-lrr(核苷酸结合位点富含亮氨酸重复序列)抗病蛋白、病原体识别受体(prr)蛋白、种子贮藏蛋白、细胞壁组分扩展蛋白、f-框蛋白、abc转运蛋白、丝氨酸-苏氨酸/酪氨酸蛋白质激酶以及核糖体rna。另一实例包括rhg1基因座,rhg1基因座包含至少三个单独的基因,并且此基因座的拷贝数变异(cnv)与大豆中的线虫抗性有关。串联重复基因阵列或多基因家族内的不等重组可提供多样性。举例来说,不等重组可在单个nbs-lrr基因簇中如每几千个配子中发生一次那样经常发生。然而,对于大多数基因簇,不等重组的发生频率较低。有时,出现新植株病原体生物型,这要求推广抗病性基因座的新抗性旁系同源物、基因变异或改变的cnv。当前,育种者依赖于产生基因多样性的天然机制,所述天然机制依赖于罕见随机突变事件来产生对新的植物病原体生物型具有抗性的植株。
85.一种具有特定经济价值的串联重复基因组基因座涵盖nbs-lrr抗病性基因家族。基于自然界中的不等重组率以及当前技术促进植株中的同源重组的能力,估计具有定制核酸内切酶(例如talen)的所有转化体中高达百分之十将在单个基因簇中诱导新的nbs-lrr变异。这些可编码针对具有农艺学重要性的植物病原体的新抗性表型。在转化之后,在生物分析中对r0植株或其子代进行表型分型以识别新的生物抗性性状。
86.在一个方面中,本公开中所描述的技术将有助于开发高-通量且廉价的性状开发平台以加速基因组基因座,尤其是包含串联阵列基因组区的基因组基因座中的分子变异。在一个实施方案中,性状开发是从与生物应力抗性相关的基因组区入手。在另一实施方案中,性状开发是从与非生物应力抗性相关的基因组区入手。在另一实施方案中,性状开发是从与组成质量相关的基因组区入手。在另一实施方案中,性状开发是从与株型相关的基因组区入手。在另一实施方案中,性状开发是从与成熟组相关的基因组区入手。在另一实施方案中,性状开发是从顺式染色体交换的基因组区入手。在另一实施方案中,性状开发是从姐妹染色体臂交换入手。在另一实施方案中,性状开发是从非同源染色体之间的染色体臂交换入手。在另一实施方案中,性状开发是从部分同源染色体之间的染色体臂交换入手。在另一实施方案中,性状开发是从单个核内染色体间的多个基因组区交换入手。
87.本文所描述的组合物和方法涉及使用位点-特异性基因组修饰酶通过刺激在基因组基因座中所选靶序列处的重组来产生新颖等位基因。在一些实施方案中,可使用本文所描述的组合物和方法在存在于植物或动物基因组中的基因簇产生新颖变异。在一些实施方案中,基因簇可包含串联重复基因阵列。在一些实施方案中,基因簇可包含基因家族。在一
些实施方案中,基因簇可为抗病性基因座。在若干实施方案中,本文所描述的组合物和方法刺激来自一种种质系的所选基因组基因座与第二种质系的顺式重组。在若干实施方案中,本文所描述的组合物和方法刺激姐妹染色体交换。在若干实施方案中,本文所描述的组合物和方法刺激同源染色体之间的基因组基因座交换。在若干实施方案中,本文所描述的组合物和方法刺激非同源染色体之间的基因组基因座交换。在若干实施方案中,本文所描述的组合物和方法刺激部分同源染色体之间的基因组基因座交换。在若干实施方案中,本文所描述的组合物和方法刺激基因组基因座中所选靶序列之间的不等重组。在若干实施方案中,本文所描述的组合物和方法刺激单个细胞内的多个基因组交换事件。
88.若干实施方案涉及一种产生基因组基因座的新等位基因的方法,所述方法包括使细胞与位点-特异性基因组修饰酶接触,所述位点-特异性基因组修饰酶裂解基因组基因座中的至少一个靶序列,由此诱导重组;以及选择包含基因组基因座的新等位基因的至少一个子代。在一个方面中,本公开提供产生基因组基因座的新等位基因的方法,所述方法包括使细胞与位点-特异性基因组修饰酶接触,所述位点-特异性基因组修饰酶裂解基因组基因座中的基因阵列中的至少一个靶序列,由此诱导与基因组基因座中的第二基因阵列的重组;以及选择包含基因组基因座的新等位基因的至少一个子代。在一些实施方案中,第一和第二基因阵列为相同的。在一些实施方案中,第一和第二基因阵列为同源的。在一些实施方案中,第一和第二基因阵列为部分同源的。在一些实施方案中,第一和第二基因阵列为串联重复基因阵列。在其他实施方案中,第一和第二基因阵列为异源的。在一些实施方案中,视重组位点而定因不对称重组而产生两种新等位基因。在一些实施方案中,不对称重组在等位基因中的至少一种中产生缺失。
89.在另一方面中,本公开提供一种诱导植物细胞或哺乳动物细胞中的至少两个靶基因组dna序列之间的重组或增加其重组率的方法,所述方法包括用位点-特异性基因组修饰酶转化植物细胞或哺乳动物细胞,所述位点-特异性基因组修饰酶特定地在所述至少两个靶dna序列之间保守的dna区内部的至少一个靶序列中诱导基因组修饰。
90.在一些实施方案中,细胞为植物细胞。在另一实施方案中,从选自近交植株和杂交植株的植株获得植物细胞。在其他实施方案中,细胞为哺乳动物细胞。在另一实施方案中,哺乳动物细胞为人类细胞或例如来自啮齿动物(例如小鼠、大鼠、仓鼠、豚鼠)、兔、猪、非人类灵长类动物(例如猴、黑猩猩、猕猴)物种或任何其他哺乳动物的细胞。
91.若干实施方案涉及一种改变植物或哺乳动物的抗病性的方法,所述方法包括用位点-特异性基因组修饰酶转化植物或哺乳动物,所述位点-特异性基因组修饰酶特定地裂解一种或多种基因组基因座中的保守区。在一些实施方案中,本公开提供一种改变植物的抗病性的方法,所述方法包括向植物细胞提供位点-特异性核酸酶,所述位点-特异性核酸酶裂解一个或多个抗病性基因座中的保守区;以及从植物细胞生长出植株。
92.本公开进一步提供一种提供具有改善的抗病性的植株的方法,所述方法包括:(a)向一个或多个植物细胞提供位点-特异性基因组修饰酶,所述位点-特异性基因组修饰酶裂解基因组基因座中的靶序列;(b)针对重组事件对一个或多个植物细胞进行筛选,以鉴定包含重组基因组基因座的植物细胞;(c)针对改善的抗病性对从步骤(b)中鉴定的植物细胞获得的植株以及其子代进行测试;以及(d)选择具有改善的抗病性的植株。在一些实施方案中,所靶向的基因组基因座包含串联重复基因组区。在一些实施方案中,所靶向的基因组基
因座包含相同的串联重复序列阵列。在一些实施方案中,所靶向的基因组基因座包含异源的串联重复序列阵列。在一些实施方案中,重组是在近交植株中。在一些实施方案中,重组是在杂交植株中。在一些实施方案中,重组是在相同染色体上。在一些实施方案中,重组是在两个染色体之间。在一些实施方案中,染色体为同源的。在一些实施方案中,染色体为非同源的。在一些实施方案中,染色体为部分同源的。在某些实施方案中,所靶向的基因组基因座与抗病性有关。在某些实施方案中,抗病性基因座包括编码nbs-lrr基因的基因。在某些实施方案中,抗病性基因座与对大豆胞囊线虫的抗性有关,例如为rhg1基因座或rhg4基因座。
93.在一些实施方案中,所靶向的基因组基因座的重组为不对称重组。在一些实施方案中,所靶向的基因组基因座的重组为对称重组。在一些实施方案中,位点-特异性基因组修饰酶使所靶向的基因组基因座处的重组率与所靶向的基因组基因座的天然存在的重组率相比增加至少2倍、至少3倍、至少4倍、至少5倍、至少6倍、至少7倍、至少8倍、至少9倍或至少10倍。
94.位点-特异性基因组修饰酶
95.若干实施方案涉及通过提供位点-特异性基因组修饰酶来促进重组。如本文所用,术语“位点-特异性酶”是指可以位点-特异性方式修饰核苷酸序列的任何酶。在一些实施方案中,通过提供单链断裂诱导剂来促进重组。在一些实施方案中,通过提供双链断裂诱导剂来促进重组。在一些实施方案中,通过提供链分离诱导试剂来促进重组。在一个方面中,位点-特异性基因组修饰酶是选自核酸内切酶、重组酶、转座酶、解旋酶或其任何组合。
96.在一个方面中,核酸内切酶是选自大范围核酸酶、锌指核酸酶(zfn)、转录激活因子样效应物核酸酶(talen)、argonaute(argonaute蛋白的非限制性实例包括嗜热栖热菌(thermus thermophilus)argonaute(ttago)、强烈炽热球菌(pyrococcus furiosus)argonaute(pfago)、格氏嗜盐碱杆菌(natronobacterium gregoryi)argonaute(ngago)、rna引导性核酸酶,诸如crispr相关核酸酶(crispr相关核酸酶的非限制性实例包括cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4、cpf1、其同源物或其修饰型式)。
97.重组酶的非限制性实例包括与dna识别基序连接的酪氨酸重组酶,本文所提供的与dna识别基序连接的酪氨酸重组酶选自由以下各项组成的组:cre重组酶、gin重组酶、flp重组酶以及tnp1重组酶。在一个方面中,本文所提供的cre重组酶或gin重组酶是拴系至锌指dna-结合域或tale dna-结合域或cas9核酸酶。在另一方面中,本文所提供的与dna识别基序连接的丝氨酸重组酶是选自由以下各项组成的组:phic31整合酶、r4整合酶以及tp-901整合酶。在另一方面中,本文所提供的与dna结合域连接的dna转座酶是选自由以下各项组成的组:tale-piggybac和tale-mutator。
98.诸如大范围核酸酶、zfn、talen、argonaute蛋白(argonaute蛋白的非限制性实例包括嗜热栖热菌argonaute(ttago)、强烈炽热球菌argonaute(pfago)、格氏嗜盐碱杆菌argonaute(ngago)、其同源物或其修饰型式)、rna引导性核酸酶(rna引导性核酸酶的非限制性实例包括crispr相关核酸酶,诸如cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、
cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4、cpf1、其同源物或其修饰型式)以及工程化的rna引导性核酸酶(rgn)等位点-特异性基因组修饰酶在基因组序列的靶位点处诱导诸如双链dna断裂(dsb)或单链dna断裂等基因组修饰,然后所述基因组序列通过同源重组(hr)或非同源末端-连接(nhej)的自然过程而修复。然后在裂解位点发生序列修饰,所述序列修饰可包括在nhej的情况下产生基因破坏的缺失或插入或因同源重组而发生的外源性序列整合。
99.在本公开的一个方面中,对位点-特异性基因组修饰酶加以选择以在基因组基因座中的一个、几个或许多个别靶序列中诱导基因组修饰。在本公开的另一方面中,基于对通过定量pcr方法所鉴定的串联重复序列的检测对位点-特异性基因组修饰酶加以选择以在基因组区中诱导基因组修饰。在本公开的另一个方面中,对位点-特异性基因组修饰酶加以选择以在已知与拷贝数变异有关的基因组区中诱导基因组修饰。基因组修饰刺激dna修复,从而可通过诸如以下的机制引起对基因组基因座的改变:基因组区的移除、基因组区的插入/缺失突变(插入缺失)、基因区内干扰基因功能的插入缺失、基因转换、串联重复阵列的非平行拷贝之间的不等重组、同源染色体之间的重组、非同源染色体之间的重组、部分同源染色体之间的重组以及其他类型的重排。在暴露于位点-特异性基因组修饰酶之后,可以各种方式鉴定所得改变,包括用于鉴定新颖变异的表型筛选、测序或分子方法。由于串联重复基因座的丰度,可将位点-特异性基因组修饰酶设计成切割许多拷贝,并且在植株中表达以刺激许多不同的改变。可使位点-特异性基因组修饰酶在植株中表达,使得基因组基因座内发生一种或多种基因组修饰,并且针对分子变化对所得子代进行筛选。随后,针对诸如以下的新颖表型对具有被证实的分子重排的子代进行表型筛选:改善的产量、改善的组成质量、改善的对非生物应力的抗性、改变的株型以及对特定植物病原体的抗性。
100.zfn
101.锌指核酸酶(zfn)以融合至foki限制性核酸内切酶的裂解结构域的工程化锌指dna-结合域为特征的合成蛋白。可将zfn设计成裂解几乎任何长段双链dna,用于修饰锌指dna-结合域。zfn从由融合至经过工程化以结合靶dna序列的锌指阵列的foki核酸内切酶的非特异性dna裂解结构域组成的单体形成二聚体。
102.zfn的dna-结合域典型地由3至4个锌指阵列组成。相对于锌指∞-螺旋起点在位置-1、 2、 3以及 6处的氨基酸有助于位点-特异性结合于靶dna,可对其进行改变和定制以适合特定靶序列。其他氨基酸形成共同的主链以产生具有不同序列特异性的zfn。用于选择zfn的靶序列的规则为本领域中已知的。
103.foki核酸酶结构域需要发生二聚化以裂解dna,并且因此两个zfn的c端区需要结合具有裂解位点的相对的dna链(相隔5-7bp)。如果两个-zf-结合位点呈回文结构,那么zfn单体可切割靶位点。如本文所用,术语zfn为广泛的并且包括单体zfn,所述单体zfn可在没有另一zfn帮助的情况下裂解双链dna。还使用术语zfn来指经过工程化以在相同位点合作裂解dna的一对zfn的一个或两个成员。
104.因为原则上可使用各种方法中的一种对锌指结构域的dna-结合特异性进行再工程化,所以理论上可将定制的zfn构建成靶向几乎任何基因序列。公共可获得的工程化锌指
结构域的方法包括上下文依赖组装(context-dependent assembly,coda)、寡聚体库工程化(oligomerized pool engineering,open)以及模块组装(modular assembly)。
105.talen
106.可将转录激活因子样效应物(tale)工程化以结合实际上任何dna序列。tale蛋白为来源于黄单孢菌属(xanthomonas)的各种植物细菌病原体的dna-结合域。在感染期间x病原体将tale分泌至宿主植物细胞中。tale移动至核,在此处它识别宿主基因组中特定基因的启动子区中的特定dna序列并且与其结合。tale具有中心dna-结合域,所述中心dna-结合域由13-28个具有33-34个氨基酸的重复单体组成。各单体的氨基酸为高度保守的,除了位置12和13处的高变氨基酸残基。这两个可变氨基酸称为重复-可变双残基(rvd)。rvd的氨基酸对ni、ng、hd以及nn分别优先识别腺嘌呤、胸腺嘧啶、胞嘧啶以及鸟嘌呤/腺嘌呤,并且rvd的调节可识别连续dna碱基。通过选择含有适当的rvd的重复区段的组合,氨基酸序列与dna识别之间的这种简单的关系已允许特定dna结合域的工程化。可将转录激活因子样效应物(tale)dna结合域融合至功能结构域,诸如重组酶、核酸酶、转座酶或解旋酶,由此赋予序列对功能结构域的特异性。
107.转录激活因子样效应物核酸酶(talen)为通过将转录激活因子样效应物(tale)dna结合域融合至核酸酶结构域而产生的人造限制酶。如本文所用,术语talen为广泛的并且包括单体talen,所述单体talen可在没有另一talen帮助的情况下裂解双链dna。还使用术语talen来指在相同位点合作裂解dna的一对talen的一个或两个成员。在一些实施方案中,核酸酶是选自由以下各项组成的组:pvuii、muth、tevi、foki、alwi、mlyi、sbfi、sdai、stsi、cledorf、clo051以及pept071。当foki融合至tale结构域时,talen对的各成员结合于侧接靶位点的dna位点,foki单体发生二聚化并且在靶位点处引起dsb。
108.除野生型foki裂解结构域之外,已设计具有突变的foki裂解结构域的变体以改善裂解特异性和裂解活性。foki结构域如二聚体一般发挥作用,需要针对靶基因组中的位点具有独特dna结合域的两个构建体具有适当的取向和间隔。talen dna结合域与foki裂解结构域之间的氨基酸残基数目以及两个个别talen结合位点之间的碱基数目两者均为实现高水平活性的参数。pvuii、muth以及tevi裂解结构域为用于tale的foki和foki变体的适用替代物。pvuii当偶联至tale时充当高度特异性裂解结构域(参见yank等2013.plos one.8:e82539)。muth能够在dna中引入链-特异性缺口(参见gabsalilow等2013.nucleic acids research.41:e83)。tevi在dna中在所靶向的位点处引入双链断裂(参见beurdeley等,2013.nature communications.4:1762)。
109.tale结合域的氨基酸序列以及dna识别之间的关系允许产生可被设计的蛋白质。可使用诸如dna works等软件程序来设计tale构建体。设计tale构建体的其他方法为本领域技术人员已知的。doyle等(2012)tal effector-nucleotide targeter(tale-nt)2.0:tools for tal effector design and target prediction.nucleic acids res.40(w1):w117-w122;cermak(2011).efficient design and assembly of custom talen and other tal effector-based constructs for dna targeting.nucleic acids res.39(12):e82。
110.大范围核酸酶
111.大范围核酸酶通常在微生物中被鉴定出,是具有高活性和长识别序列(>14bp)从
而引起靶dna的位点-特异性消化的独特的酶。天然存在的大范围核酸酶的工程化型式典型地具有延长的dna识别序列(例如14
–
40bp)。
112.大范围核酸酶的工程化比zfn和talen更具挑战性,因为大范围核酸酶的dna识别和裂解功能发生在单个结构域中。已使用专门的诱变和高-通量筛选方法来形成新颖的大范围核酸酶变体,所述新颖的大范围核酸酶变体识别独特的序列并且具有改善的核酸酶活性。
113.argonaute
114.argonaute蛋白家族为dna引导性核酸内切酶。已报道从格氏嗜盐碱杆菌分离的argonaute适合用于人类细胞中的dna引导性基因组编辑(gao等dna-guided genome editing using the natronobacterium gregoryi argonaute.nature biotechnology 34:768-773(2016)。已鉴定出来自其他物种的argonaute核酸内切酶(argonaute蛋白的非限制性实例包括嗜热栖热菌argonaute(ttago)、强烈炽热球菌argonaute(pfago)、格氏嗜盐碱杆菌argonaute(ngago)、其同源物或其修饰型式)。已将这些独特的argonaute核酸内切酶中的每一种与编码dna向导的序列相关联。
115.crispr
116.crispr(成簇的有规律间隔开的短回文重复序列)/cas(crispr相关的)系统为dna-结合域使得能够在特定序列处修饰基因组dna的合成蛋白(例如zfn和talen)的替代物。crispr/cas系统的特异性是基于使用互补碱基配对来识别靶dna序列的rna向导。在一些实施方案中,位点-特异性基因组修饰酶为crispr/cas系统。在一个方面中,本文所提供的位点-特异性基因组修饰酶可包含任何rna引导性cas核酸酶(rna引导性核酸酶的非限制性实例包括cas1、cas1b、cas2、cas3、cas4、cas5、cas6、cas7、cas8、cas9(也称为csn1和csx12)、cas10、csy1、csy2、csy3、cse1、cse2、csc1、csc2、csa5、csn2、csm2、csm3、csm4、csm5、csm6、cmr1、cmr3、cmr4、cmr5、cmr6、csb1、csb2、csb3、csx17、csx14、csx10、csx16、csax、csx3、csx1、csx15、csf1、csf2、csf3、csf4、cpf1、其同源物或其修饰型式);并且任选地,包含靶向相应核酸酶所必需的向导rna。
117.crispr/cas系统为细菌和古生菌的适应性免疫系统的一部分,通过以序列-依赖性方式裂解外来dna保护它们不受诸如病毒等入侵核酸的影响。通过将被称为间隔序列的短入侵dna片段整合在crispr基因座近端的两个邻近的重复序列之间来获得免疫性。crispr阵列(包括间隔序列)在后来遇到入侵性dna期间进行转录并且被加工成长度为约40nt的小干扰crispr rna(crrna),小干扰crispr rna与反式活化crispr rna(tracrrna)组合以活化并引导cas9核酸酶。此举裂解入侵dna中称为前间隔序列(protospacer)的同源双链dna序列。裂解的先决条件为在靶dna下游存在保守的前间隔序列邻近基序(pam),它通常具有序列5
′‑
ngg-3
′
,但不太经常具有nag。特异性是由pam上游约12个碱基处的所谓的“种子序列”提供,所述种子序列必须在rna与靶dna之间相匹配。cpf1以类似于cas9的方式起作用,但cpf1不需要tracrrna。
118.靶基因
119.可将本公开应用于任何基因组基因座以产生遗传变异或重组。在一个实施方案中,基因组基因座具有基因组序列的串联重复拷贝。在一些实施方案中,将基因组序列的重复拷贝组织在基因簇中,例如呈串联阵列形式。此类基因组区的实例包括但不限于涵盖编
码以下各项的基因的基因组基因座:nbs-lrr(核苷酸结合位点富含亮氨酸重复序列)抗病蛋白、病原体识别受体(prr)蛋白、种子贮藏蛋白、细胞壁组分扩展蛋白、f-框蛋白、abc转运蛋白、免疫球蛋白、丝氨酸-苏氨酸/酪氨酸蛋白质激酶以及核糖体rna。在另一方面中,可将基因组区与拷贝数变异(cnv)与抗性(诸如对大豆胞囊线虫的抗性)有关的基因座(例如rhg1)相关联。在一个方面中,使用本公开来靶向和修饰nbs-lrr抗病性基因座,以产生赋予植株改善的抗病性的新nbs-lrr变体。在另一方面中,使用本公开来靶向和修饰具有编码免疫球蛋白的基因的基因座,以在哺乳动物细胞中产生新免疫球蛋白变体。
120.在一个方面中,本公开提供一种产生新串联重复基因阵列的方法,所述方法包括使细胞与位点-特异性基因组修饰酶接触,所述位点-特异性基因组修饰酶在第一串联重复基因阵列中的至少一个靶序列处修饰基因组,由此诱导与第二串联重复基因阵列的重组;以及选择包含新串联重复基因阵列的至少一个子代。在一些实施方案中,通过不对称重组来产生新串联重复基因阵列。在一些实施方案中,基于周围序列与第二串联重复基因阵列中的序列的同源性来选择靶序列。在一些实施方案中,靶序列和第二串联重复基因阵列中的序列位于串联重复基因阵列内的不同位置。
121.在一些实施方案中,靶序列位于基因组的基因区中。在其他实施方案中,靶序列位于基因组的基因间区中。
122.在一个实施方案中,位点-特异性基因组修饰酶的靶序列位于所选基因组基因座的基因区内。在另一实施方案中,位点-特异性基因组修饰酶的靶序列位于所选基因组基因座的基因间区中。在一些实施方案中,所选基因组基因座可包含qtl。在一些实施方案中,所选基因组基因座可包含两个或更多个串联阵列基因单元。在一个实施方案中,串联阵列基因单元位于相同染色体上。在另一实施方案中,串联阵列基因单元位于不同染色体上。在一些实施方案中,串联阵列基因单元位于同源染色体上。在一些实施方案中,串联阵列基因单元位于非同源染色体上。在一些实施方案中,串联阵列基因单元位于部分同源染色体上。在另一实施方案中,串联阵列基因单元为旁系同源物。在一些实施方案中,串联阵列基因单元为同源物。在一些实施方案中,串联阵列基因单元为部分同源物。在另一实施方案中,串联阵列基因单元为不同的,更具体地说一个基因单元与另一基因单元不相同。
123.为促进所选基因组基因座中的不对称重组或为促进不同基因组基因座之间的重组,或为促进同源基因组基因座之间的重组,或为促进旁系同源基因组基因座之间的重组,或为促进部分同源基因组基因座之间的重组,或为促进同源染色体上的基因组基因座之间的重组,或为促进非同源染色体上的基因组基因座之间的重组,或为促进部分同源染色体上的基因组基因座之间的重组,以一种方式对位点-特异性基因组修饰酶的靶序列加以选择,使得周围基因组区的序列与选择用于重组的基因组基因座中的序列因至少50bp、至少100bp、至少150bp、至少200bp、至少250bp、至少300bp、至少350bp、至少400bp、至少450bp、至少500bp、至少550bp、至少600bp、至少650bp、至少700bp、至少750bp、至少800bp、至少850bp、至少900bp、至少950bp或至少1,000bp而高度类似(所选基因组基因座可例如位于与靶序列的相同基因组基因座中、与靶序列不同的基因组基因座中、基因组基因座内与靶序列不同的位置、旁系同源基因组基因座中、同源基因组基因座中、部分同源基因组基因座中、同源染色体上、部分同源染色体上或旁系同源染色体上)。在一些实施方案中,所选基因组基因座中的序列与靶序列周围的基因组区的序列可在至少50bp、至少100bp、至少150bp、
至少200bp、至少250bp、至少300bp、至少350bp、至少400bp、至少450bp、至少500bp、至少550bp、至少600bp、至少650bp、至少700bp、至少750bp、至少800bp、至少850bp、至少900bp、至少950bp或至少1,000bp上具有至少85%同一性、至少90%同一性、至少91%同一性、至少92%同一性、至少93%同一性、至少94%同一性、至少95%同一性、至少96%同一性、至少97%同一性、至少98%同一性、至少99%同一性或100%同一性。在一些实施方案中,所选基因组基因座可含有用于相同位点-特异性基因组修饰酶或不同位点-特异性基因组修饰酶的靶序列。靶序列周围的基因组区中的这种高水平的同源性有助于与所选基因组基因座的重组。在一些实施方案中,靶序列周围的基因组区中的高水平的同源性有助于不对称重组。在一些实施方案中,归因于所选基因组基因座内的多态性,新组装的基因组基因座促进形成新颖拷贝数变体和/或新颖基因。在包括在具有同源性的区中的基因组区或基因座的情况下,通过同源基因组区的随机匹配形成的变异数增加有较高的概率。如果靶序列存在于多个基因组区中,那么可不仅因不等重组,而且因缺失而引起新变体的产生。在相同基因组基因座内两个靶序列彼此远远分开,缺失的几率降低,因此为不等重组提供更多的机会。
124.植物免疫系统具有两种不同而又高度相关联的路径来识别和抵御病原攻击。病原细胞的诸如细胞壁或鞭毛组分等一些必需的表面分子可由多种跨膜蛋白,主要是激酶识别。这些病原信号统称为病原体相关分子模式(pamp),而其受体被称为病原体识别受体(prr)。针对它们的防御机制被称为pamp-触发免疫性(pti),所述pamp-触发免疫性被认为是植物中分子防御的第一线。然而,一些病原体具有完善的机制来应付此第一线防御,并且能够主动将效应分子运送至植物细胞中,从而对细胞功能进行再编程以有利于病原体。这些效应物触发另一主要防御路径,即效应物触发的免疫性(eti)。eti的识别蛋白构成具有保守结构域结构的大超家族,它们包括核苷酸结合位点(nbs)和富含亮氨酸重复序列(lrr)结构域。这些大的丰富的nbs-lrr蛋白参与检测多种多样的病原体,包括细菌、病毒、真菌、线虫、昆虫以及卵菌。pti与eti之间的主要差异在于虽然pamp受体在大类群中相当保守,但nbs-lrr蛋白甚至在物种内也是高度可变的。这种卓越的可变性反映了植株与其在极其多样化的选择压力下保持nbs-lrr基因的病原环境之间的激烈竞争。
125.典型植物种类可具有几百个nbs-lrr基因,它们常常被组织在大的串联重复基因单元簇中。举例来说,水稻带有约580个nbs-lrr基因。其中大多数被分配至串联重复簇中。约有130个此类簇分散在水稻基因组中。每个簇的基因数目从两个变化到十八个,在连续统下端频率更高。这种基因组组织有助于在细胞减数分裂期间在减数母细胞中、在细胞有丝分裂期间在体细胞中或在dna修复期间在体细胞中串联阵列基因拷贝之间的频繁不对称比对。在相同等位基因之间可发生不对称重组,从而可如图1中所说明在基因阵列内产生新重组的基因单元的新的等位基因和等位基因组合。不等比对还允许非平行基因拷贝之间发生交换事件,从而可产生新的等位基因和等位基因组合。如上文所描述的不等(不对称)重组被广泛视为促成nbs-lrr基因座的卓越多样性的主要机制。在自然界中,在单个nbs-lrr基因簇中不等重组可如每几千个配子中发生一次那样经常发生。然而,对于大部分nbs-lrr基因簇来说,其发生频率较低。
126.在一个方面中,本公开描述使用位点-特异性基因组修饰酶来诱导染色体dna中的基因组修饰以加快新颖等位基因开发的速率,特别是加快nbs-lrr基因组基因座中的不等重组的方法。这些基因组修饰可产生增加的不对称重组率、对称重组率或插入缺失、缺失或
倒位产生率,这些又可产生新的疾病识别特异性。
127.在一些实施方案中,nbs-lrr抗病性基因编码赋予对一种或多种选自以下的疾病的抗性的蛋白质:玉米或小麦的各种真菌锈病;诸如大豆、玉米以及小麦等各种物种的镰刀菌属疾病;玉米戈斯氏萎蔫病、灰色叶斑病和锈病、亚洲大豆锈病;大豆和棉花根结线虫病;棉花肾形线虫病;以及小麦茎锈和叶锈病。在另一实施方案中,nbs-lrr抗病性基因编码赋予对一种或多种选自以下的玉米疾病的抗性的蛋白质;炭疽茎腐病(禾生刺盘孢)、曲霉穗腐病(曲霉属(aspergillus spp.))、常见玉米锈病(高粱柄锈菌(puccinia sorghi))、色二孢穗腐病(干腐色二孢(diplodia frumenti)、玉蜀黍色二孢(d.maydis))、色二孢条斑病(大孢色二孢(diplodia macrospora))、色二孢茎腐病(干腐色二孢、玉蜀黍色二孢)、眼斑病(玉米短梗霉(aureobasidium zeae))、镰刀菌穗腐病(轮枝镰刀菌)、镰刀菌茎腐病(镰刀菌属)、赤霉穗腐病(念珠状赤霉)、赤霉茎腐病(玉米赤霉)、戈斯氏萎蔫病和叶枯病(苜蓿萎蔫病菌)、灰色叶斑病(玉蜀黍尾孢菌、玉米尾孢菌)、黑穗病(丝轴黑粉菌(sphacelotheca reiliana))、北方玉米叶枯病(大斑凸脐蠕孢)、腐霉病(腐霉属(pythium spp.))、南方叶枯病(异旋孢腔菌(cochliobolus heterostrophus))。在另一实施方案中,nbs-lrr抗病性基因编码赋予对一种或多种选自以下的大豆疾病的抗性的蛋白质:镰刀菌根腐病(镰刀菌属)、猝死综合征(茄病镰刀菌)、亚洲大豆锈病(豆薯层锈菌)、疫霉根腐和茎腐病(大豆疫霉)、根结线虫病(根结线虫属)、大豆胞囊线虫病(大豆胞囊线虫)、肾形线虫病(肾状轮线虫)、茎溃疡(菜豆间座壳)。在另一实施方案中,nbs-lrr抗病性基因编码赋予对一种或多种选自以下的棉花疾病的抗性的蛋白质:黑根腐病(根串珠霉(thielaviopsis basicola))、铃腐病(镰刀菌属、刺盘孢属、疫霉属、立枯丝核菌(rhizoctonia solani))、叶斑病(链格孢属(alternaria spp.)、棉尾孢菌(cercospora gossypina)、立枯丝核菌)、白粉病(棉拟粉孢霉(oidiopsis gossypii))、棉锈病(柄锈菌属(puccinia spp.)、棉层锈菌(phakopsora gossypii))、肾形线虫病(肾状轮线虫)、根结线虫病(南方根结线虫)、链格孢叶斑病(大孢链格孢(alternaria macrospora))、镰刀菌萎蔫病(尖孢镰刀菌萎蔫专化型)、轮枝孢菌萎蔫病(大丽轮枝孢菌)。在另一实施方案中,nbs-lrr抗病性基因编码赋予对一种或多种选自以下的小麦疾病的抗性的蛋白质:镰刀菌首枯病(禾谷镰刀菌)、镰刀菌苗枯病(镰刀菌属、颖枯壳针孢)、镰刀菌叶斑病(雪腐小画线壳)、叶锈病(小麦柄锈菌(puccinia triticina))、茎锈病(禾柄锈菌)、黄锈病(条形柄锈菌)、白粉病(禾白粉病菌)、小麦壳针孢斑块病(小麦壳针孢)、颖枯壳针孢斑块病(颖枯壳针孢)、小麦瘿蚊(黑森瘿蚊(mayetiola destructor))。
128.在一些实施方案中,nbs-lrr抗病性基因编码赋予对一种或多种由包括但不限于以下各项的病毒引起的疾病的抗性的蛋白质:胡椒斑驳病毒、胡椒轻型斑驳病毒、黄瓜花叶病毒、番茄黄曲叶病毒、黄瓜绿斑驳花叶病毒、马铃薯病毒y、小西葫芦黄花叶病毒、芜菁花叶病毒以及水稻条纹病毒。
129.在一些实施方案中,如本文所描述产生的抗病性基因座的新等位基因与亲代等位基因相比赋予对一种或多种选自以下的疾病的改善的抗性:玉米或小麦的各种真菌锈病;诸如大豆、玉米以及小麦等各种物种的镰刀菌属疾病;玉米戈斯氏萎蔫病、灰色叶斑病和锈病、亚洲大豆锈病;大豆和棉花根结线虫病;棉花肾形线虫病;以及小麦茎锈和叶锈病。在一些实施方案中,如本文所描述产生的抗病性基因座的新等位基因与亲代等位基因相比赋予
对一种或多种选自以下的玉米疾病的改善的抗性;炭疽茎腐病(禾生刺盘孢)、曲霉穗腐病(曲霉属)、常见玉米锈病(高粱柄锈菌)、色二孢穗腐病(干腐色二孢、玉蜀黍色二孢)、色二孢条斑病(大孢色二孢)、色二孢茎腐病(干腐色二孢、玉蜀黍色二孢)、眼斑病(玉米短梗霉)、镰刀菌穗腐病(轮枝镰刀菌)、镰刀菌茎腐病(镰刀菌属)、赤霉穗腐病(念珠状赤霉)、赤霉茎腐病(玉米赤霉)、戈斯氏萎蔫病和叶枯病(苜蓿萎蔫病菌)、灰色叶斑病(玉蜀黍尾孢菌、玉米尾孢菌)、黑穗病(丝轴黑粉菌)、北方玉米叶枯病(大斑凸脐蠕孢)、腐霉病(腐霉属)、南方叶枯病(异旋孢腔菌)。在一些实施方案中,如本文所描述产生的抗病性基因座的新等位基因与亲代等位基因相比赋予对一种或多种选自以下的大豆疾病的改善的抗性:镰刀菌根腐病(镰刀菌属)、猝死综合征(茄病镰刀菌)、亚洲大豆锈病(豆薯层锈菌)、疫霉根腐和茎腐病(大豆疫霉)、根结线虫病(根结线虫属)、大豆胞囊线虫病(大豆胞囊线虫)、肾形线虫病(肾状轮线虫)、茎溃疡(菜豆间座壳)。在一些实施方案中,如本文所描述产生的抗病性基因座的新等位基因与亲代等位基因相比赋予对一种或多种选自以下的棉花疾病的改善的抗性:黑根腐病(根串珠霉)、铃腐病(镰刀菌属、刺盘孢属、疫霉属、立枯丝核菌)、叶斑病(链格孢属、棉尾孢菌、立枯丝核菌)、白粉病(棉拟粉孢霉)、棉锈病(柄锈菌属、棉层锈菌)、肾形线虫病(肾状轮线虫)、根结线虫病(南方根结线虫)、链格孢叶斑病(大孢链格孢)、镰刀菌萎蔫病(尖孢镰刀菌萎蔫专化型)、轮枝孢菌萎蔫病(大丽轮枝孢菌)。在一些实施方案中,如本文所描述产生的抗病性基因座的新等位基因与亲代等位基因相比赋予对一种或多种选自以下的小麦疾病的改善的抗性:镰刀菌首枯病(禾谷镰刀菌)、镰刀菌苗枯病(镰刀菌属、颖枯壳针孢)、镰刀菌叶斑病(雪腐小画线壳)、叶锈病(小麦柄锈菌)、茎锈病(禾柄锈菌)、黄锈病(条形柄锈菌)、白粉病(禾白粉病菌)、小麦壳针孢斑块病(小麦壳针孢)、颖枯壳针孢斑块病(颖枯壳针孢)、小麦瘿蚊(黑森瘿蚊)。
130.在一些实施方案中,如本文所描述产生的抗病性基因座的新等位基因与亲代等位基因相比赋予对一种或多种由包括但不限于以下各项的病毒引起的疾病的改善的抗性:胡椒斑驳病毒、胡椒轻型斑驳病毒、黄瓜花叶病毒、番茄黄曲叶病毒、黄瓜绿斑驳花叶病毒、马铃薯病毒y、小西葫芦黄花叶病毒、芜菁花叶病毒以及水稻条纹病毒。
131.在一些实施方案中,如本文所描述产生的抗病性基因座的新等位基因与亲代等位基因相比赋予对一种或多种选自由以下各项组成的组的疾病的改善的抗性:炭疽茎腐病(禾生刺盘孢)、镰刀菌穗腐病(轮枝镰刀菌)、镰刀菌茎腐病(镰刀菌属)、赤霉穗腐病(念珠状赤霉)、赤霉茎腐病(玉米赤霉)、戈斯氏萎蔫病和叶枯病(苜蓿萎蔫病菌)、灰色叶斑病(玉蜀黍尾孢菌、玉米尾孢菌)、北方玉米叶枯病(大斑凸脐蠕孢)、猝死综合征(茄病镰刀菌)、亚洲大豆锈病(豆薯层锈菌)、疫霉根腐和茎腐病(大豆疫霉)、根结线虫病(根结线虫属)、大豆胞囊线虫病(大豆胞囊线虫)、肾形线虫病(肾状轮线虫)、根结线虫病(南方根结线虫)、镰刀菌萎蔫病(尖孢镰刀菌萎蔫专化型)、轮枝孢菌萎蔫病(大丽轮枝孢菌)、镰刀菌首枯病(禾谷镰刀菌)、镰刀菌苗枯病(镰刀菌属、颖枯壳针孢)、镰刀菌叶斑病(雪腐小画线壳)以及茎锈病(禾柄锈菌)。
132.其他所选病毒或真菌病诱导有机体可选自包括以下各项的列表:枝顶孢属(acremonium)、苜蓿花叶病毒(alfamovirus)、青葱x病毒(allexivirus)、链格孢属、互隔链格孢(alternaria alternata)、葡萄卷叶病毒(ampelovirus)、曲霉属、米曲霉(aspergillus oryzae)、杂色曲霉(aspergillus versicolor)、出芽短梗霉
(aureobasidium pullulans)、菜豆金黄花叶病毒(begomovirus)、平脐蠕孢属(bipolaris)、小麦平脐蠕孢(bipolaris sorokiniana)(长蠕孢枯病(helminthosporium blight))、盘梗霉属(bremia)、莴苣盘梗霉(bremia lactucae)、大麦黄花叶病毒(bymovirus)、毛状病毒(capillovirus)、香石竹潜病毒(carlavirus)、香石竹斑驳病毒(carmovirus)、花椰菜花叶病毒(caulimovirus)、枝孢属(cladosporium)、草本枝孢(cladosporium herbarum)、线形病毒(closterovirus)、豇豆花叶病毒(comovirus)、毛形病毒(crinivirus)、南瓜花叶病毒(cucumovirus)、质型弹状病毒(cytorhabdovirus)、葡萄白粉菌(erisphe necator)、白粉菌属(erysiphe)、蚕豆萎蔫病毒(fabavirus)、弯曲病毒科(flexiviridae)、凹陷病毒(foveavirus)、真菌传棒状病毒(furovirus)、双生病毒(geminivirus)、大麦条纹花叶病毒(hordeivirus)、等轴不稳环斑病毒(ilarvirus)、黄症病毒(luteovirus)、葡萄斑点病毒(maculavirus)、稻瘟病菌(magnaporthe grisea)(灰色叶斑病)、微座孢属(microdochium)、雪霉微座孢(microdochium nivale)(粉雪霉病)、线虫传多面体病毒(nepovirus)、青霉属(penicillium)、茎点霉属(phoma)、植物呼肠孤病毒(phytoreovirus)、黄斑叉丝单囊壳(podosphaero macularis)、马铃薯卷叶病毒(polerovirus)、马铃薯帚顶病毒(pomovirus)、马铃薯x病毒(potexvirus)、马铃薯y病毒(potyvirus)、梨孢属(pyricularia)、丝核菌属(rhizoctonia)、稻丝核菌(rhizoctonia oryzae)(丝核菌鞘斑病(rhizoctonia sheath spot))、温州蜜柑矮缩病毒(sadwavirus)、核盘霉属(sclerotinia)、壳针孢属(septoria)、芹菜壳针孢(septoria apiicola)、单丝壳菌(spatherotheca fuliginea)、单丝壳属(sphaerotheca)、葡萄穗霉属(stachybotrys)、黑葡萄穗霉(stachybotrys chartarum)、taastrupvirus、纤细病毒(tenuivirus)、烟草花叶病毒(tobamovirus)、烟草脆裂病毒(tobravirus)、番茄丛矮病毒(tombusvirus)、番茄斑萎病毒(tospovirus)、毛癣菌属(trichophyton)、红色毛癣菌(trichophyton rubrum)、发状病毒(trichovirus)。
133.在本公开的一个方面中,植株为玉米并且nbs-lrr为rp1。在另一方面中,植株为大豆并且nbs-lrr为rpp1。在另一方面中,植株为大豆并且nbs-lrr为rps1。在另一方面中,植株为大豆并且串联重复阵列为赋予线虫抗性的rhg1基因座。在另一方面中,植株为大豆并且抗病性基因座为rhg4基因座。
134.大豆锈病(sbr)为大豆的最具破坏性的疾病之一。已在各种种质中针对sbr鉴定出若干抗性基因座,其中定位于染色体18上的rpp1为最有效的。已对基因座进行作图,达到1cm间隔(kim等2012theor.appl.genet.125:1339-1352)。在物理上,对应染色体区段除其他基因外具有nbs-lrr基因的几个串联重复阵列,它们很可能是rpp1的候选物。不幸地,最近已出现针对rpp1具毒性的新sbr生物型,从而需要将rpp1的新抗性等位基因以及其他抗性基因座的新抗性等位基因推广到商业种质中。当前,育种者必须依赖于在这些基因座处基因多样性的天然机制,并且随后对通过这些罕见随机突变而开发的大豆新品种进行选择。或者,与在此所描述的系统不相关的转基因方法可提供对sbr具有抗性的大豆事件。使用本公开中所提出的方法,将由位点-特异性基因组修饰酶特异性地靶向存在于rpp1区中的nbs-lrr基因家族以引起新对偶构型的快速开发。将通过下文提出的高-通量分子法对这些个别事件进行基因分型。将针对抗病性对包含重组事件的个体进行表型分析。
135.rp1为一种针对玉米真菌锈病的主要抗性基因座。它定位于染色体10上并且由串
联重复的nbs-lrr基因单元组成。在玉米基因型间其拷贝数为高度可变的,表明在所述区中频繁发生本公开中所描述的减数分裂重排。因此,存在若干rp1单倍型,其中的每一种均有效抵抗许多锈病生物型。另一方面,鉴于病原体新毒性小种的典型高发展速率,当前广泛用于商业玉米种质中的rp1单倍型集合或许不久就会被新的病原体打垮。如本公开中所描述的在rp1基因座中所靶向的重组可以与产生新毒性病原体相同的步调或甚至更快地产生新等位基因构型,并且因此育种者可通过将这些新的基因推广到商业种质中来使锈病被控制住。
136.在美国最有破坏性的大豆根腐和茎腐病之一是由卵菌纲病原体大豆疫霉引起的。虽然存在对致病性大豆疫霉(p.sojae)剂具有抗性的大豆栽培物,但是有易感性和毒力转化的记录,表明需要继续开发在抗性基因座方面具有多样性的大豆以预防广泛的作物损失。
137.在美国大豆胞囊线虫(scn)为在经济上最具破坏性的大豆病原体(cook,2012)。大豆中的数量性状基因座rhg1是定位至染色体18并且被发现赋予对scn的抗性,不过scn抗性的分子基础为不清楚的。已将scn抗性基因座的位置缩小至包含三个单独基因的31-kb区。已报导rhg1基因座的3或10个拷贝的串联重复序列与scn抗性有关联。
138.在本公开的一个方面中,针对nbs-lrr基因簇的保守区所设计的位点-特异性基因组修饰酶(例如核酸内切酶)可显著增加nbs-lrr基因簇的重组率。当通过定制的核酸内切酶将双链断裂引入植物基因组中时,观测到重组频率增加约1000倍。相应地,依据靶向的核酸酶活性可假定不等重组率可以类似比率增加。假设关于多基因家族所观测到的不等交换的天然比率为1/10,000至1/100,000,将靶向双链断裂引入新nbs-lrr等位基因中可产生1000倍的频率增加。具体地说,这可产生每几十个转化体或几百个转化体发生一次的增加的频率。这将极大地扩大nbs-lrr基因中的遗传型变异并且因此扩大其针对各种疾病的识别特异性。
139.在一些实施方案中,新nbs-lrr等位基因可赋予使用常规育种方法所不能实现的对疾病的抗性。在一些实施方案中,新等位基因可扩展现有抗病性基因座对广泛范围的引起疾病的生物型的识别能力。
140.在一些实施方案中,靶基因为免疫球蛋白基因。在一些实施方案中,靶序列位于重链的基因编码部分(例如vh、ch1、ch2、ch3、ch4、铰链区或其部分)上。在其他实施方案中,靶序列位于轻链的基因编码部分(例如vl和cl或其部分)上。在一些实施方案中,靶序列位于编码可变(v)区段、多样性(d)区段、连接(j)区段或其部分或任何组合的基因上。在一些实施方案中,靶序列位于侧接免疫球蛋白基因的基因间区中。已知人类免疫球蛋白基因以基因区段的串联阵列的形式存在,在b-淋巴细胞发育期间经历体细胞重组和超突变。这些方法使得能够产生大量的能够识别和清除感染剂的免疫球蛋白分子。
141.使用位点-特异性基因组修饰酶产生新串联重复基因阵列的一般策略
142.下文说明使用位点-特异性基因组修饰酶(诸如位点-特异性核酸内切酶)产生基因组基因座(诸如抗病性基因座)的新等位基因的一般策略。nbs-lrr抗病性基因座为非限制性实例。
143.在一个实施方案中,首先在植株中进行dna序列分析以鉴定根据两个或更多个所靶向的基因组基因座具保守性的短序列基序。在一些实施方案中,所靶向的基因组基因座
包含串联重复基因、基因家族或基因簇。在某些实施方案中,植株是选自玉米、大豆、棉花、小麦以及芥花。在某些实施方案中,所靶向的基因组基因座包含nbs-lrr基因。在一些实施方案中,短序列基序的长度为至少50bp、至少100bp、至少150bp、至少200bp、至少250bp、至少300bp、至少350bp、至少400bp、至少450bp、至少500bp、至少600bp、至少700bp、至少800bp、至少900bp、至少1000bp、至少1100bp、至少1200bp、至少1300bp、至少1400bp、至少1500bp、至少1600bp、至少1700bp、至少1800bp、至少1900bp或至少2000bp。在一些实施方案中,在所鉴定的短序列基序中所述两个或更多个所靶向的基因组基因座拥有至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%或100%的序列同一性。可将位点-特异性基因组修饰酶设计成靶向所鉴定的短序列基序。
144.在一些实施方案中,相似性检索以及位点-特异性基因组修饰酶的后续设计集中于nbs lrr基因座的与nbs结构域相对的lrr结构域。由于lrr结构域直接参与病原体识别,如果lrr结构域为内部重排的,而不仅仅是作为完整结构域与另一nbs结构域“对换”,那么更可能产生新的识别特异性。玉米与大豆均被认为带有几十种保守lrr标签。大豆还含有若干可开发的具有抗病性的岛,因此在已知与抗病性有关的区中形成更大的基因多样性。棉花和小麦为真正的异源多倍体物种,可能具有甚至更多的此类序列。
145.将若干定制的位点-特异性基因组修饰酶设计成靶向所鉴定的短序列基序。在一些实施方案中,位点-特异性基因组修饰酶为核酸内切酶。在一些实施方案中,位点-特异性基因组修饰酶为重组酶。在一些实施方案中,位点-特异性基因组修饰酶为转座酶。在一些实施方案中,位点-特异性基因组修饰酶为解旋酶。在一些实施方案中,核酸酶为转座酶。在某些实施方案中,核酸内切酶是选自大范围核酸酶、zfn、talen以及cripr/cas系统。
146.在一些实施方案中,在使用定量平台转化之前测试位点-特异性基因组修饰酶的活性。可用于本公开中的定量平台的一个实例包括但不限于基于原生质体的寡核苷酸整合分析继之以数字pcr以检测寡核苷酸整合。然后在植株中对有效的位点-特异性基因组修饰酶进行大规模稳定转化实验。
147.图3中说明了在原生质体分析中检测同源重组的方法。在此方法中,在两个亲本染色体中的每一个上鉴定出侧接所靶向的感兴趣的基因的靶序列,诸如限制性核酸酶位点。在一些实施方案中,靶序列为相同的。在其他实施方案中,靶序列为不同的。在一些实施方案中,通过相同的位点-特异性基因组修饰酶对靶序列进行修饰。在其他实施方案中,通过不同的位点-特异性基因组修饰酶对靶序列进行修饰。如箭头所示设计出对第一亲本染色体具特异性的pcr引物和对第二亲本染色体具特异性的另一pcr引物。诱导的双链断裂促进两个亲本染色体之间的重组,将靶序列与引物结合位点两者带到相同重组染色体上。将探针设计成特异性地识别第一染色体上的基因或阵列的5'-侧接区与第二染色体上的基因或阵列的3'-侧接区的独特接点。仅在发生重组的情况下观测到pcr产物。
148.在某些实施方案中,植株是选自:苜蓿、小茴香、苹果、杏、菊芋、芝麻菜、芦笋、鳄梨、香焦、大麦、菜豆、甜菜、黑莓、蓝莓、芸苔、花椰菜、抱子甘蓝、卷心菜、芥花、哈蜜瓜、胡萝卜、木薯、菜花、芹菜、樱桃、芜荽、柑桔、克莱门氏小柑橘、咖啡、玉米、棉花、黄瓜、花旗松、茄子、菊苣、苦苣、桉树、茴香、无花果、葫芦、葡萄、葡萄柚、蜜瓜、豆薯、奇异果、莴苣、韭葱、柠
檬、莱姆酸橙、火炬松、芒果、玉米、甜瓜、蘑菇、坚果、燕麦、秋葵、洋葱、橙子、观赏植物、番木瓜、欧芹、豌豆、桃子、花生、梨、胡椒、柿子、松树、菠萝、车前草、李子、石榴、白杨、马铃薯、南瓜、榅桲、辐射松、红菊苣、萝卜、覆盆子、水稻、黑麦、高粱、南方松、大豆、菠菜、笋瓜、草莓、甜菜、甘蔗、向日葵、甘薯、枫香、甜玉米、红桔、茶、烟草、番茄、草皮、藤本植物、西瓜、小麦、山药以及小西葫芦植株。
149.在某些实施方案中,植株是选自:六出花属(alstroemeria)(例如巴西六出花(alstoemeria brasiliensis))、紫菀、映山红(例如杜鹃花属(rhododendron sp.))、秋海棠(例如秋海棠属(begonia sp.))、风铃草、叶子花、仙人掌(例如仙人掌科蟹爪兰(cactaceae schlumbergera truncata))、山茶、康乃馨(例如香石竹(dianthus caryophyllus))、菊花(例如菊花属(chrysanthemum sp.))、铁线莲(例如铁线莲属(clematis sp.))、鸡冠花、耧斗菜、仙客来(例如仙客来属(cyclamen sp.))、黄水仙(例如水仙属(narcissus sp.))、假扁柏、小苍兰(例如香雪兰(freesia refracta))、老鹳草、大丁草、唐菖蒲(例如唐菖蒲属(gladiolus sp.))、冬青、木槿(例如朱槿(hibiscus rosasanensis))、绣球花(例如八仙花(macrophylla hydrangea))、桧柏、百合(例如百合属(lilium sp.))、木兰、迷你玫瑰、兰花(例如兰科(orchidaceae)的成员)、矮牵牛(例如杂种撞羽朝颜(petunia hybrida))、猩猩木(例如一品红(euphorbia pulcherima))、报春花、杜鹃花、蔷薇科(rosaceae)、玫瑰(例如玫瑰属(rosa sp.))、金鱼草(例如金鱼草属(antirrhinum sp.))、灌木、树(诸如林木(阔叶树和常绿树,诸如针叶树))以及郁金香(例如郁金香属(tulipa sp.))。
150.在一些实施方案中,植株为玉米、玉米、大豆、棉花、小麦、高粱或芥花。在一些实施方案中,植株为近交系或杂交系。
151.在一些实施方案中,使用近交系来进行转化。在其他实施方案中,使用杂交系来进行转化。在杂交种中,进行一次基因组修饰,不仅是不等重组而且甚至是常规同源重组都将在基因组基因座(例如异合子型nbs-lrr基因簇)处形成新等位基因。因此,在转化不严格地为基因型依赖性转化的情况下,具有多态性抗病性基因的杂交种将产生最大频率的新颖变异。在仅某些系容易转化(诸如玉米)的情况下,可形成近等基因系,所述近等基因系因抗病性基因簇而变化,但具有相似转化特性。此类近等基因系的杂交将产生可转化的系,所述可转化的系关于抗病性基因为多种多样的。
152.在一些实施方案中,位点-特异性基因组修饰酶在植株中为稳定转化的。在其他实施方案中,位点-特异性基因组修饰酶在植株中为瞬时转化的。在一些实施方案中,位点-特异性基因组修饰酶在植株中组成性表达。在其他实施方案中,位点-特异性基因组修饰酶在组织特异性启动子的控制下在植株中表达。在其他实施方案中,位点-特异性基因组修饰酶在本领域中已知的可调控启动子的控制下在植株中表达。在一些实施方案中,可调控启动子为化学诱导型启动子。
153.在一些实施方案中,位点-特异性基因组修饰酶在热休克启动子、组织特异性启动子或化学诱导型启动子的控制下在植株中表达。
154.然后在生物分析中例如针对新抗病性表型对r0植株或其自交和/或回交子代进行表型分型。关于对一组疾病的抗性的高-通量生物分析(例如对叶穿孔物或非常幼小的苗的测试)允许在相当短的时间内在对数千个植株中进行筛选。
155.在一些实施方案中,使用叶圆片分析来进行早期、高-通量筛选。在一些实施方案中,收集叶圆片并且放置于皮式培养皿(petri dish)中并且使用本领域中已知的方案接种病原体。将培养皿在适当条件下孵育以促进病原体生长,并且使用视觉检查来确定植株圆片对由病原体引起的损害的抗性。
156.在另一实施方案中,使用叶接种分析来进行高-通量表型筛选,其中从所选植株去除叶子,并且放置于皮式培养皿中,接种病原体,并且将培养皿在适当条件下孵育以促进病原体生长。使用视觉检查来确定叶子对由病原体引起的损害的抗性。
157.在另一实施方案中,使个别植株在温室中在盆中生长,接种病原体并且监测植株生长的表型参数。在一些实施方案中,所跟踪的植株生长参数为植株高度、株冠发育、根构型、鲜重、茎强度以及任何其他植物健康参数。人工地或使用自动化温室的系统来进行植株生长参数数据的收集。在另一实施方案中,在琼脂培养基中或转动过滤分析中对来自植株的个别种子进行评估以评估根和下胚轴伸长(chon等,2000)。琼脂或转动的纸巾可为在测试病原体存在下吸胀的。
158.使用关于田间或温室条件最佳化的若干成像系统在人工方法所需时间的一部分时间内对大量植株进行快速且准确的表型分型。实例包括在自动化或传统温室或田间环境中使用诸如荧光、热或红外照相机等多种技术对整个植株或所选器官(例如叶)的2d或3d成像。
159.然后在田间试验中针对生物应力对已通过高-通量叶圆片分析、叶接种分析、根或胚轴伸长分析以及温室分析的植株进行筛选。为此,以完全区组设计将来自具有所鉴定的新基因座的植株的种子种植在田间,植株可接种或可不接种病原体,并且对植株生长参数进行评分并且测定产量。在育种计划中使用对生物应力具有抗性的植株来将新基因型种质渗入至优良种质中。
160.在一些实施方案中,使用位点-特异性基因组修饰酶的组成性表达,因为它可产生稳定水平的随机改变。在有可能产生许多转化体的情况下,可使用组成性表达,因为各事件将经历独立切割和后续随机修复结果。因此,作为调控位点-特异性基因组修饰酶活性的替代方式,可对许多独立事件进行筛选。然而,在调控基因组暴露于定制核酸酶(例如talen)的时间安排的其他情况下可使所需改变的频率最大化。
161.应以产生并且允许回收许多独立改变的方式来进行位点-特异性基因组修饰酶活性的时间安排。对位点-特异性基因组修饰酶活性进行正确的时间安排预防在细胞谱系中早期的一种或几种改变(诸如插入缺失)的“固定”,从而然后预防在所述基因处的后续切割,由此使在所述位置将回收的变异量最小化。
162.某些组织或细胞类型可具有不同的各种dna修复路径相对水平,使得某一组织或细胞类型中的活化可引起较高丰度的所需类型的dna修复和所得改变。举例来说,减数分裂开始时的核酸内切酶活性可使簇中基因之间或来自不同亲代的各种簇之间的重组事件的可能性最大化。
163.在一些实施方案中,通过以具有可调控启动子的基因盒的形式递送位点-特异性基因组修饰酶来实现对位点-特异性基因组修饰酶活性的调控。此类启动子将为本领域技术人员已知的并且包括热休克启动子、组织特异性启动子、化学诱导型启动子或其他环境调控启动子。用于调控位点-特异性基因组修饰酶活性的其他方法包括rna稳定性、蛋白质
稳定性、蛋白质定位、配体诱导蛋白活化、条件性内含肽中断(conditional intein disruption)以及其他方法。
164.在减数分裂开始时基因组暴露于位点-特异性基因组修饰酶且因此对其进行修饰对刺激减数分裂重组和不等交换可为最佳的。另外,此阶段的改变将产生具有在下一代中可回收的不同改变的许多独立配子。关于位点-特异性基因组修饰酶活性的时间安排的其他选择可为在植株发育的早期,使得各种子有机会产生不同的改变;或在发生分生组织分枝时的一个时间,诸如在玉米中在雄穗发育期间。
165.在一些实施方案中,在减数分裂开始时靶基因组基因座中的保守区裂解。
166.除不等同源重组之外,在形成双链断裂后还可发生若干其他类型的突变。举例来说,称为合成-依赖性链退火的机制可引起不对称基因转换,从而像不对称同源重组一样也将形成所涉及基因的新功能变体。另一机制非同源末端连接将形成点突变或短插入缺失,从而将在dsb周围引起氨基酸变化,或通过移码突变或通过引入终止密码子而赋予nbs-lrr基因非功能性。
167.如果在相同染色体中的两个串联阵列nbs-lrr基因中同时发生dsb,那么切割物之间相对短的片段可被清除,而不是再整合至基因组中。在此情况下,将通过单链退火或非同源末端连接使在缺失片段外的染色体末端再连结。此情况的净结果将为新的nbs-lrr等位基因以及损失两个或更多个大基因片段。基因转换还可引起所有或部分的基因用来自相关基因的序列置换。此情况的影响可能是通过将来自一种基因的编码区的部分用另一种置换或甚至置换整个编码区,使得第一基因的调控部分现在控制第二基因的拷贝而产生新颖的等位基因。
168.在这些许多可能的dna修复机制中,形成新的功能性等位基因的那些(例如不等同源重组)对于开发新的抗病特异性来说为重要的。然而,赋予nbs-lrr基因非功能性的突变可能自相矛盾地也非常适合用于开发针对某些类型的病原体的新抗性表型。nbs-lrr基因触发程序化细胞死亡(pcd)的路径,所述路径杀死植株中的感染细胞,因此在病原攻击位点周围形成局部损害。存在越来越多的证据证明一些死体营养型或半活体营养型病原体可破坏此过程而有利于病原体。能够分泌宿主特异性毒素的死体营养型或半活体营养型病原体可在pcd将破坏植物组织的大区段而不是形成小损害的程度上刺激nbs-lrr基因。这些衰弱性组织又可被病原体入侵。在此类型的发病机理的所有已知情况下,功能性nbs-lrr基因引起疾病易感性,而致病性nbs-lrr基因的缺失或截短产生抗性。在一些极端情况下,例如高粱的“买罗(milo)”病,此类缺失引起完全且不可逆的免疫性,因此所述疾病不再成为经济问题。已针对许多重要的疾病,包括大豆中由镰刀菌属物种引起的猝死综合征(sds)假定了此发病机制。
169.本公开还提供一种通过本文所公开的方法产生的植株。在一些实施方案中,位点-特异性基因组修饰酶修饰至少两个基因组基因座。在一些实施方案中,位点-特异性基因组修饰酶修饰至少两个抗病性基因或其他串联重复基因。在一个方面中,植株具有至少一个重组基因组基因座或其他串联基因组基因座。在一个方面中,植株在所选基因组基因座中的一种中具有至少一个缺失。在一个方面中,植株具有至少一个重组抗病性基因或其他串联重复基因。在一个方面中,植株在抗病性基因或其他串联重复基因中的一种中具有至少一个缺失。在另一方面中,植株与不具有重组基因组基因座或在所选基因组基因座中的一
种中不具有缺失的植株相比具有改善的抗病性。在另一方面中,植株与在所选基因组基因座中的一种中不具有重复的植株相比具有改善的抗病性。在另一方面中,植株与不具有重组抗病性基因或在抗病性基因中的一种中不具有缺失的植株相比具有改善的抗病性。在一个方面中,植株在其基因组中不具有位点-特异性基因组修饰酶。
170.在一个方面中,在本公开所描述的方法中转化的仅有的外源性基因为位点-特异性基因组修饰酶和任何必需的可选标记物。因为在引起对基因组的改变之后转基因将被隔离掉,所以它将不是最终产物的一部分。通常将具有转基因的多个拷贝的事件以及整合至内源性基因中的事件弃去。然而,在一些实施方案中,只要位点-特异性基因组修饰酶为有效的,就可以使用它们。这将允许操作高-通量转化和筛选平台来快速鉴定新的性状,诸如新的抗病性性状。这还将允许形成改善的抗病性性状,其中改善的抗病性性状在面临改变的病原体群体时维持稳健的抗性,或其中改善的性状提供对更广谱的病原体小种的抗性。此外,它将节省大部分事件表征的成本。
171.以下实施例是出于说明目的而呈现,并且不应被视为限制。
实施例
172.实施例1:双链断裂靶位点选择
173.为加速所选基因组基因座中的重组,在一个或多个所选种质系中的所选基因组基因座中进行综合序列分析,以鉴定所选基因组基因座的基因区与基因间区两者之中的多态性。通过本领域技术人员已知的方法进行测序。测序方法的非限制性实例包括bac克隆测序、深度测序、“鸟枪法(shot-gun)”测序、随机测序、直接测序、下一代测序法等。然后使用生物信息学工具来鉴定诸如以下各项的位点-特异性基因组修饰酶的特异性靶序列:大范围核酸酶、锌指核酸酶、锌指重组酶、argonaute、talen、taler、rna引导性核酸酶或crispr相关蛋白。
174.部分基于周围基因组区的序列来选择靶序列,使得靶序列周围的基因组区与所选基因组基因座内的其他区在至少100bp上具有高水平的同源性。在靶序列周围的基因组区中的这种高水平的同源性有助于不等交换。归因于所选基因组基因座内的多态性,新组装的基因组基因座促进形成新颖变体。在基因组基因座内具有同源性的区包括多个串联重复基因或多基因家族的多个成员的情况下,因同源区的随机匹配而形成的变异数增加的概率较高。如果靶序列存在于基因组基因座内的多个位置,那么可不仅因不等重组,而且因缺失而形成新变体。在两个靶序列彼此以不断增加的距离定位的情况下,靶序列之间缺失的几率降低,因此为不等重组提供更多的机会。
175.除使用测序数据来选择感兴趣的基因组基因座内的基因组修饰酶靶序列之外,还可进一步使用测序数据来选择种质以用于转化。举例来说,可使用测序数据来告知用于位点-特异性基因组修饰酶的转化的特定优良种质或杂交种质的选择。可通过在代表不同种质的个别植株的集合中进行再测序,并且鉴定所选基因组基因座的基因区与基因间区两者之中的多态性来实现针对所选基因组基因座处的重组对种质进行鉴定。
176.实施例2:原生质体分析
177.使用原生质体分析作为用于测试所选基因组基因座内核酸酶诱导的重组的快速分析。从来自如实施例1中所描述而鉴定的所选种质的玉米植株的叶肉细胞制备原生质体,
并且在所选基因组基因座处为异合子型。类似地,从来自如实施例1中所描述而鉴定的所选种质的大豆植株的子叶制备原生质体,并且在所选基因组基因座处为异合子型的。对于原生质体分析,选择在所选基因组基因座的侧接侧具有多态性的个别植株以有助于根据一种或多种pcr分析配置进行筛选。
178.将针对如实施例1中所描述而鉴定的特定靶序列而选择和/或设计的位点-特异性基因组修饰酶克隆至具有植株-特异性表达元件的表达盒中。表达元件包括增强子、启动子、内含子、5
’‑
未翻译前导序列(5
’‑
utr)以及3'-未翻译聚腺苷酸化序列(3’utr)。使用本领域中已知的方法将表达盒进一步密码子最佳化,例如用于玉米中的表达盒使用单子叶密码子最佳化,而用于大豆中的表达盒使用双子叶密码子最佳化。将表达盒合并至适合用于原生质体分析的转化载体中。
179.使用含有编码位点-特异性基因组修饰酶的至少一个表达盒的转化载体使用标准原生质体转化方案(例如电穿孔或聚乙二醇(peg))转化原生质体。在一个实施方案中,位点-特异性基因组修饰酶为crispr/cas9核酸酶,至少一个向导rna靶向基因组基因座中的所选靶序列中的一个或多个。在另一实施方案中,位点-特异性基因组修饰酶为talen。在另一实施方案中,位点-特异性基因组修饰酶为连接至重组酶的crispr相关蛋白。在另一实施方案中,位点-特异性基因组修饰酶为argonaute。在另一实施方案中,位点-特异性基因组修饰酶为重组酶。在另一实施方案中,位点-特异性基因组修饰酶为大范围核酸酶。在另一实施方案中,位点-特异性基因组修饰酶为tale重组酶。
180.将含有位点-特异性基因组修饰酶表达盒的转化载体转化至原生质体中。原生质体分析的设计使得位点-特异性基因组修饰酶在所选基因组基因座中引入基因组修饰,例如双链断裂(dsb)、单链断裂、转座酶介导的dna交换反应或重组酶介导的dna交换反应,以促进重组。在48至72小时后收获转化的原生质体并且提取基因组dna。在一种pcr分析配置中,以片段在消化和自我连结后将形成用于探针的独特模板的方式在基因的侧接区中选择限制性核酸酶位点。此独特模板不因任一亲本染色体的自我连结而存在,而是对通过重组染色体的自我连结而产生的模板具特异性(参见图3)。使用探针来定量重组染色体。此分析不需要在产生原生质体种质的亲代之间在感兴趣的基因组基因座中具多态性,仅需要在其更广泛的基因环境中具多态性。
181.图3中说明了在原生质体分析中检测重组的方法。在该图中,限制性核酸酶位点由在“a”旁系同源物的5'-侧翼、
‘
a’旁系同源物的3'-侧翼下方以及在新的重组“a/a”旁系同源物的5'-侧翼与3'-侧翼两者处的三角形表示。在基因组dna的限制性核酸酶消化之后,允许基因组片段自我连结以形成圆形分子。当在基因座处发生同源重组时,那么“a”旁系同源物的5'-侧翼与“a”旁系同源物的3'-侧翼连接。使用反向pcr加上液滴数字pcr(ddpcr)技术来检测这些罕见的重组事件。具体地说,将一种pcr引物设计成专属于“a”旁系同源物的5'-侧翼,将第二pcr引物设计成专属于“a”旁系同源物的3'-侧翼,并且将pcr探针设计成专属于通过“a”旁系同源物的5'-侧翼与“a”旁系同源物的3'-侧翼的自我连结所形成的接点。仅在发生重组的情况下观测到pcr产物。
182.图5中呈现了用于检测同源重组的另一pcr分析配置。在此分析中,对pcr引物组合加以选择以扩增所选基因组基因座内的一个或多个个别基因组区,使得个别pcr扩增子的
长度有所不同。在重组后,使用相同引物对产生新尺寸的pcr扩增子。通过凝胶电泳或通过毛细管电泳对pcr扩增子进行拆分。
183.实施例3:用于检测大豆rps1基因座中的重组的原生质体分析
184.使用含有用于大豆密码子最佳化cas9(seq id no:77)的表达盒的载体以及由大豆u6i(seq id no:80)或u6c(seq id no:81)启动子驱动的一个或两个单向导rna(sgrna)构建体对大豆种质a3555的子叶原生质体进行共转化。针对由seq id no:21至seq id no:32表示的6个所鉴定的rpsi靶序列(ts)中的一个对sgrna进行设计。对于这六个rpsi靶序列中的每一个,存在因1个核苷酸而不同的两种变体(参见表4)。使用海肾(renilla)荧光素酶构建体作为转化对照。在两天孵育期之后,从转化的原生质体分离全部基因组dna。如实施例1中所描述使用pcr分析来鉴定重组rps1旁系同源物,所述pcr分析利用个别rps1旁系同源物间的序列长度多态性。虽然rps1旁系同源物为相当保守的,但归因于沿它们的长度分布的多个短插入缺失在它们之间存在显著的长度多态性。因此,两个保守引物之间的pcr扩增子的尺寸将在广泛范围内变化。举例来说,在大豆williams82(w82)参考大豆基因组的21个单独rps1旁系同源物中的两个高度保守区之间产生的pcr扩增子各自具有独特的pcr扩增子长度。为分析从此实施例的大豆原生质体提取的dna,在从转化的大豆原生质体提取的dna的pcr扩增中使用35种pcr引物(seq id no:33-67)。这些pcr引物代表163种不同的引物对,73种独特的引物组合(独特引物组合)如表1中详细描述。使用pcr反应的毛细管电泳,鉴定出24种新颖扩增子。通过呈现于图6、7以及8中的来自pcr扩增子的毛细管电泳分析的电泳型态说明了序列长度变异结果的实例。序列长度变体预期代表重组rpsi基因座。通过pcr扩增子的序列分析来鉴定新颖rpsi变体。
185.表1.rps1原生质体分析引物矩阵.
186.187.188.189.[0190][0191]
实施例4:靶基因座处的植物原位(in planta)重组
[0192]
为实现植物原位重组,如实施例2中所描述,将针对在原生质体分析中所鉴定的特定靶序列而设计的位点-特异性基因组修饰酶克隆至转化载体中,所述转化载体含有具有植株-特异性表达元件并且关于植物表达进行密码子最佳化的表达盒。将表达盒合并至土壤杆菌属(agrobacterium)转化载体中。使用土壤杆菌属的植物转化方法为本领域中已知的。或者,通过生物弹射击转化法(biolistic transformation method)将转化载体引入植物细胞,所述生物弹射击转化方法也是本领域中熟知的。在转化之后,使用本领域中熟知的方法选择稳定植株。
[0193]
可使用含有组织-特异性启动子和/或化学诱导型启动子(诸如乙醇诱导型启动子)的表达盒来调控位点-特异性基因组修饰酶的表达。在使用化学诱导型启动子的情况下,可在应用化学诱导剂之后在所需生长期诱导位点-特异性基因组修饰酶的表达。任选地,可使用具有组织-特异性与化学诱导型启动子两者的表达盒,使得在应用化学诱导剂的时间和生长期仅在所需组织(例如花粉)中发生位点-特异性基因组修饰酶的表达。
[0194]
可进行所选基因组基因座的植物原位靶向,使得位点-特异性基因组修饰酶在感兴趣的基因组基因座内的至少一个靶序列内诱导基因组修饰,例如双链断裂(dsb)、单链断裂、转座酶介导的dna交换反应或重组酶介导的dna交换反应。基因组修饰的引入将增加感兴趣的基因组基因座内的重组率。重组事件可为对称或不等的并且可在感兴趣的基因组基因座内保守或相异的区之间发生。从如实施例1中所描述而鉴定的植株获得用于转化的植物细胞。另外,可至少部分基于有利的再生特性来选择用于转化的植株的种质。被选择用于转化的植物细胞在感兴趣的基因组基因座处可为纯合子型或异合子型的。举例来说,可选择包含在rp1基因座处不相同的亲本基因组的玉米植株或包含在rpp1基因座处不相同的亲本基因组的大豆植株。在感兴趣的基因组基因座内,无论感兴趣的基因组基因座为纯合子型还是异合子型,均可对位点-特异性基因组修饰酶的靶序列加以选择以诱导感兴趣的基因组基因座的不相同区之间的重组。
[0195]
在稳定植株的转化和选择之后,获得各植株的叶样品,提取基因组dna,并且使用droplet digital
tm pcr(ddpcr)技术进行高-通量分析,以鉴定在所选基因组基因座处具有重组的个别植株。图4中说明了pcr分析的设计。通过检测基因组基因座中改变的基因拷贝数目来鉴定不等重组事件。在图4中所说明的实例中,各亲本基因型贡献三种旁系同源物,因此被选择用于转化的细胞具有六种旁系同源物(参见图4中顶部的“r0亲本”)。在发生重组的事件中,说明了一种可能的实例,其中在不等重组之后,一个配子在基因组基因座中具有五种旁系同源物,而另一个配子在基因组基因座中具有一种旁系同源物(参见图4中的“配子1”和“配子2”)。在回交之后,
‘
配1’的子代的基因型将在基因组基因座处具有八种旁系同源物;等位基因上的五种旁系同源物来自配子1,并且三种旁系同源物来自野生型亲代等位基因。类似地,由对
‘
配2’的回交产生的子代的基因型将在基因组基因座处具有四种旁系同源物;等位基因上的1种旁系同源物来自配子1,并且三种旁系同源物来自野生型亲代等位基因。因为pcr探针是针对基因座内所有旁系同源物中的共同区而设计,所以ddpcr分析结果反映旁系同源物的数目。使用旁系同源物数目相较于亲本旁系同源物数目的变化来检测基因座处的重组事件。举例来说,如果亲本等位基因各自在基因组基因座中具有三种旁系同源物(如图4中所说明),那么在同源不对称重组后,所得配子在基因座处可具有五种旁系同源物、四种旁系同源物、三种旁系同源物、两种旁系同源物或一种旁系同源物。在回交或自交后,这些配子将产生具有改变的拷贝数的植株,所述改变的拷贝数是通过ddpcr技术来检测。
[0196]
实施例5:对具有重组的植株进行表型筛选以鉴定新颖抗病性性状
[0197]
使用在实施例3中所鉴定的在所选基因组基因座内具有在分子上得到证实的重组的植株来进行表型筛选,以识别对生物应力具有新的或增强的耐受性的植株。
[0198]
对于早期、高-通量筛选,使用叶圆片分析。在这些分析中,收集叶圆片并且放置于皮式培养皿中并且使用本领域中已知的方案接种病原体。将培养皿在适当条件下孵育以促进病原体生长,并且使用视觉检查来确定植株圆片对由病原体引起的损害的抗性。
[0199]
用于鉴定对病原体具有增强的耐受性的植株的另一种高-通量筛选分析为叶接种分析,其中从所选植株去除叶子,并且放置于皮式培养皿中,接种病原体,并且将培养皿在适当的条件下孵育以促进病原体生长。使用视觉检查来确定叶子对由病原体引起的损害的抗性。
[0200]
用于鉴定具有增强的疾病耐受性的植株的另一种高-通量筛选分析为根伸长和胚轴伸长分析。在此分析中,将种子铺在琼脂上或转动的滤纸中并且在多种条件下孵育以允许根和胚轴伸长。将琼脂或滤纸接种感兴趣的病原体。测量根和胚轴的长度作为疾病耐受性的指示。
[0201]
用于鉴定具有增强的病原体耐受性的植株的另一种表型筛选分析为使个别植株在温室中在盆中生长,将植株接种病原体并且监测植株生长的表型参数。所跟踪的植株生长参数为植株高度、株冠发育、根构型、鲜重、茎强度以及任何其他植物健康参数。人工地或使用自动化温室的系统来进行植株生长参数数据的收集。
[0202]
使用关于田间或温室条件最佳化的若干成像系统在人工方法所需时间的一部分时间内对大量植株进行快速且准确的表型分型。实例包括在自动化或传统温室或田间环境中使用诸如荧光、热或红外照相机等多种技术对整个植株或所选器官(例如叶)的2d或3d成像。
[0203]
然后在田间试验中针对生物应力对已通过高-通量叶圆片分析或叶接种分析以及温室分析的植株进行筛选。为此,以完全区组设计将来自具有所鉴定的重组等位基因的植株的种子种植在田间,植株可接种或可不接种病原体,并且对植株生长参数进行评分并且测定产量。在育种计划中使用对生物应力具有抗性的植株来将新等位基因种质渗入至优良种质中。
[0204]
实施例6.玉米的rp1锈病抗性基因座中的加速重组
[0205]
玉米的rp1锈病抗性基因座为具有高农艺学价值的nbs-lrr基因簇的一个实例。在新nbs-lrr等位基因的开发中,重组事件可以是在nbs-lrr簇的匹配拷贝之间或在nbs-lrr簇的错配拷贝之间。在rp1基因座中对b73玉米基因组进行再测序。使用公共可获得的rp1基因模型进行诠释,总的来说,在染色体10上的rp1基因座中鉴定出16种rp1旁系同源物。在这16种rp1旁系同源物中,14种聚集在一起,而另外两种旁系同源物,虽然也定位于染色体10上,但由较大的染色体区段隔开。
[0206]
为鉴定适合用于在rp1基因座中化脓性链球菌(streptococcus pyogenes)cas9介导的双链断裂(dsb)诱导的重组的靶序列,使用生物信息学工具对涵盖这16种所鉴定的rp1旁系同源物中的每一种的基因组基因座进行分析。以seq id no:1和seq id no:2形式呈现rp1旁系同源物的两个代表性实例。使用用于多序列比对的clustal w算法来比对rp1旁系同源物(higgins等1994)。搜索在大多数rp1旁系同源物间保守的序列的区的
‘
最佳’(g(n)
18
gngg;seq id no:78)和
‘
最差’((n
21
)gg;seq id no:79)crispr/cas9靶序列(ts)。根据保守程度进一步按优先次序列出包括此类靶序列的区,以使切割效率最大化:根据经验,所有旁系同源物中至少一半必定为可由一种或最多两种同源向导-rna(grna)构建体裂解的。在满足上述所有准则的区之中,定位于rp1旁系同源物的c端富含亮氨酸重复序列(lrr)结构域中的区获得靶向优先权,以在crispr/cas9核酸酶存在下诱导双链断裂。与nbs-lrr类别的基因的nbs结构域相比,lrr结构域在旁系同源物间倾向于更不同。此外,具有抗性特异性的大多数决定簇是定位于lrr结构域中。因为这个原因,lrr结构域中的不等重组预期以比nbs结构域中的不等重组更高的频率加速新颖抗病性等位基因的产生。
[0207]
由此分析,鉴定出5种分开的共同序列。对于靶序列(ts)1和2,存在共同序列的因1个核苷酸而不同的两种变体。以seq id no:3-9形式呈现共同rp1靶序列。对于两种代表性
rp1旁系同源物(seq id no:1和seq id no:2),表2中呈现了不同crispr/cas9靶序列的位置。
[0208]
表2.代表性rp1旁系同源物中rp1 crispr/cas9靶序列的位置.
[0209][0210]
在16种单独rp1旁系同源物中评估保守crispr/cas9靶序列的存在,12种rp1旁系同源物具有关于ts-1(ts-1a:seq id no:3或ts-1b:seq id no:4)的crispr/cas9靶序列;在16种rp1旁系同源物中有10种具有关于ts-2(ts-2a:seq id no:5,ts-2b:seq id no:6)的crispr/cas9靶序列;16种rp1旁系同源物中有8种具有关于ts-3(seq id no:7)的crispr/cas9靶序列;16种rp1旁系同源物中有10种具有关于ts-4(seq id no:8)的crispr/cas9靶序列,并且16种rp1旁系同源物中有8种具有关于ts-5(seq id no:9)的crispr/cas9靶序列。
[0211]
设计植物转化载体以递送crispr/cas9核酸酶组分。从化脓性链球菌得到cas9序列并且关于单子叶表达对核苷酸序列进行密码子最佳化。另外,cas9序列在蛋白质的5’与3'末端均含有核靶向序列(seq id no:77)。在相同转化载体中,存在一个或两个向导-rna(grna)编码盒:两个grna盒用于rp1 ts-1(ts-1a和ts-1b);两个grna盒用于rp1 ts-2(ts-2a和ts-2b);并且各载体的一个grna盒被设计成靶向rp1 ts-3,或ts-4,或ts-5。转化载体还含有赋予对草甘膦(glyposate)的耐受性的选择盒。如实施例4中所描述将转化载体引入植株中。
[0212]
用于玉米组织的dna提取方法为本领域中已知的。用于鉴定rp1基因座的方法在本领域中已知是通过使用基因组标记物来进行。深度测序的方法为本领域中已知的并且指以高精度对dna区进行多次(成百上千次)测序以检测单核苷酸多态性(snp)。进行测序的方法的非限制性实例包括测序平台(illumina,san diego,ca)、roche454测序系统(roche,branford,ct)、单分子实时测序技术(pacific biosciences,menlo park,ca)测序平台以及本领域中已知的其他方法。
[0213]
除r0和r1植株的分子筛选之外,如实施例5中所描述针对抗病性对r1或其他近交或杂交子代植株进行筛选。
[0214]
实施例7:rpp1大豆锈病抗性基因座中的加速重组
[0215]
大豆的rpp1病抗性基因座为具有高农艺学价值的nbs-lrr基因簇的一个实例。在新nbs-lrr等位基因的开发中,重组事件可为“相等”的,例如在nbs-lrr簇的对应区之间,或
为“不等”的,例如在nbs-lrr簇的非对应区之间。在rpp1基因座中对不同种质的大豆基因组进行测序。使用公共可获得的rpp1基因模型进行诠释,总的来说,在rpp1基因座中鉴定出三种rpp1旁系同源物。
[0216]
为鉴定适合用于在rpp1基因座中化脓性链球菌cas9介导的双链断裂(dsb)诱导的重组的靶位点,使用生物信息学工具对涵盖所鉴定的rpp1旁系同源物中的每一种的基因组区进行分析。以seq id no:10和seq id no:11形式呈现在williams 82种质(w82)中所鉴定的rpp1旁系同源物的两个代表性实例。使用用于多序列比对的clustal w算法来比对rpp1旁系同源物(higgins等1994)。搜索在大多数rpp1旁系同源物间保守的序列的区的
‘
最佳’(g(n)
18
gngg;seq id no:57)和
‘
最差’((n
21
)gg;seq id no:58)crispr/cas9靶序列(ts)。如实施例6中所描述根据保守程度进一步按优先次序列出包括此类靶序列的区以使切割效率最大化。
[0217]
由此分析,鉴定出5种单独的共同序列。对于靶序列(ts)2和5,存在共同序列的因1个核苷酸而不同的两种变体。以seq id no:12-18形式呈现共同rpp1靶序列。对于两种代表性rpp1旁系同源物(seq id no:10和seq id no:11),表3中呈现了不同crispr/cas9靶序列的位置。注意,关于ts-4(seq id no:16、ts-5a(seq id no:17以及ts-5b(seq id no:18)的靶序列(ts)位置如由相应seq id no:10和seq id no:11的起点和终点位置所指示是位于逆向链上。
[0218]
表3.代表性rpp1旁系同源物中rpp1 crispr/cas9靶序列的位置.
[0219][0220]
在3种单独rpp1旁系同源物中评估保守crispr/cas9靶序列的存在,所有3种rpp1旁系同源物均具有关于ts-1(ts-1:seq id no:12)、ts-3(seq id no:15)以及ts-4(seq id no:16)的crispr/cas9靶序列。一种旁系同源物具有关于ts-2a(seq id no:13)与ts-2b(seq id no:14)的crispr/cas9靶序列;rpp1旁系同源物中有两种具有关于ts-5a(seq id no:17)的crispr/cas9靶序列,并且旁系同源物中有一种具有关于ts-5b(seq id no:18)的crispr/cas9靶序列。
[0221]
构建植物转化载体以递送crispr/cas9核酸酶组分。从化脓性链球菌得到cas9序列,并且关于单子叶表达对核苷酸序列进行密码子最佳化,并且进行13种氨基酸变化以降低变应原性(e24d、d54g、v143a、t191i、g205r、k234r、t310s、t593a、e630g、h723q、v743i、
r753、l847i)。另外,cas9序列在蛋白质的5’与3'末端均含有核靶向序列(seq id no:77)。在相同转化载体中,存在一个或两个向导-rna(grna)编码盒:两个grna盒用于rpp1 ts-2(ts-2a和ts-2b);两个grna盒用于rpp1 ts-5(ts-5a和ts-5b);并且各载体的一个grna盒被设计成靶向rpp1 ts-1,或ts-3,或ts-4。转化载体还含有赋予对草甘膦的耐受性的选择盒。
[0222]
使被鉴定为具有含有适合于重组的多态性的基因型的植株杂交以形成杂种群体。使用近交亲本种质或f1杂交植株或对于整个感兴趣的区来说为异合子型的子代作为起始物质来进行植株转化。使用本领域中已知的方法将植株转化载体引入玉米细胞中,例如使用土壤杆菌属转化或生物弹射击转化。使用本领域中已知的方法使用草甘膦对转化的愈伤组织进行选择。如实施例4中所描述将转化载体引入植株中。
[0223]
用于玉米组织的dna提取方法为本领域中已知的。用于鉴定rp1基因座的方法在本领域中已知是通过使用基因组标记物来进行。深度测序的方法为本领域中已知的并且指以高精度对dna区进行多次(成百上千次)测序以检测单核苷酸多态性(snp)。进行测序的方法的非限制性实例包括测序平台(illumina,san diego,ca)、roche454测序系统(roche,branford,ct)、单分子实时测序技术(pacific biosciences,menlo park,ca)测序平台以及本领域中已知的其他方法。
[0224]
除r0和r1植株的分子筛选之外,如实施例5中所描述针对抗病性对r1或其他近交或杂交子代植株进行筛选。
[0225]
实施例8:rps1大豆疫霉属抗性基因座中的加速重组
[0226]
大豆的rps1疫霉属抗性基因座为具有高农艺学价值的nbs-lrr基因簇的一个实例。在新nbs-lrr基因型的开发中,重组事件可为“相等”的,例如在nbs-lrr簇的对应区之间,或为“不等”的,例如在nbs-lrr簇的非对应区之间。在rps1基因座中对不同种质的大豆基因组进行测序。使用公共可获得的rps1基因模型进行诠释,总的来说,在rps1大豆疫霉属抗性基因座中鉴定出23种rps1旁系同源物。
[0227]
由此分析,鉴定出6种单独的共同序列,每一种具有共同序列的因1个核苷酸而不同的两种变体。以seq id no:21-32形式呈现共同rps1靶序列。对于两种代表性rps1旁系同源物(seq id no:19和seq id no:20),表4中呈现了不同crispr/cas9靶序列的位置。注意,关于ts-6a(seq id no:31)和ts-6b(seq id no:32)的靶序列(ts)位置如由相应seq id no:19和seq id no:20的起点和终点位置所指示是位于逆向链上。
[0228]
表4.代表性rps1旁系同源物中rps1 crispr/cas9靶位点的位置.
id no:77)中的rhg1基因座的公共可获得的基因组序列来鉴定双链断裂诱导crispr/cas9核酸酶的靶序列。由此分析,鉴定出八个靶序列(ts):ts1(seq id no:69);ts2(seq id no:70);ts3(seq id no:71);ts4(seq id no:72);ts5(seq id no:73);ts6(seq id no:74);ts7(seq id no:75);以及ts8(seq id no:76)。crispr/cas9靶序列中有六个是定位于rgh1基因座的基因区内,并且crispr/cas9靶序列中有两个是定位于rhg1基因座的基因间区中(参见表5)。
[0238]
表5.rhg1靶位点和其在靶序列中的坐标
[0239][0240]
如实施例1中所描述构建植株转化载体以递送crispr/cas9核酸酶组分。
[0241]
使用本领域中已知的方法并且如实施例6中所描述转化被鉴定为具有含有适合于重组的多态性的基因型的植株。
[0242]
dna提取方法和用于鉴定rhg1基因座的方法为本领域中已知的。使用深度测序来鉴定rhg1基因座内的修饰。
[0243]
除r0和r1植株的分子筛选之外,如实施例5中所描述针对抗病性对r1或其他近交或杂交子代植株进行筛选。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。
