包含dna分子混合物的组合物、其作为生物抑制剂的用途和生产方法
技术领域
1.本发明涉及适于抑制目标物种的包含dna分子混合物的组合物、使用所述组合物的方法和用途以及生产该组合物的方法。所述组合物尤其可用于人类和/或兽医医学、农业中的害虫和/或病害防治领域或在其中一种或多种目标物种的抑制是有益的其他领域中。
背景技术:
2.近年来,已有知识表明,一个物种的片段化dna对其来源的物种和具有相似基因组的系统发育相似的物种具有抑制作用。公开这种dna片段自抑制效应的第一批出版物是国际专利申请wo2014/020624及其意大利优先申请na2012a000046。这些文献主要关注新发现的自抑制性dna片段的效果,根据这些出版物,自抑制性dna片段是通过超声处理、热处理或热解从待抑制的物种(或从系统发育相似的物种)中分离的总dna,或通过从该物种(或从系统发育相似的物种)的总dna开始的随机dna片段合成而产生的。
3.为了更有效地利用dna片段来抑制生物物种,需要以有效形式生产它们的进一步方法。现已令人惊讶地发现,待抑制物种的dna片段在掺入非源自待抑制物种的dna后是有效的。基于这一令人惊讶的发现,本发明的发明人发现,在与待抑制物种无关的宿主物种中,也可以有效地产生该物种的抑制性dna片段。这为产生抑制性dna片段打开了新的发明可能性。
技术实现要素:
4.根据第一方面,本发明涉及一种组合物,所述组合物包含dna分子的混合物,所述dna分子的混合物具有来自不同来源的dna序列,所述混合物包含来自来源物种的多个第一dna序列以及来自宿主物种(不同于所述来源物种)的多个第二dna序列,所述第一dna序列优选来自染色体dna序列。在所述组合物中,所述dna分子的混合物任选地包含嵌合dna分子,其中来源物种dna序列在至少一端侧接非来源物种dna序列,所述非来源物种dna序列选自宿主物种dna序列或选自来自人工dna构建体的dna序列。这种组合物有利于抑制作为来源物种选择的目标物种,或选自系统发育上与来源物种相似的物种的目标物种,因为抑制性的dna片段可以在系统发育上远离目标物种的宿主物种中产生。
5.本发明的另一方面涉及用于医学的本发明的组合物。鉴于本发明的组合物可用于抑制寄生性和/或致病性生物的事实,其在人类和/或兽医医学中具有实用性。
6.本发明的又一方面涉及根据本发明的组合物作为抑制目标物种的产品的用途。这种用途可以是在人类和/或兽医医学中的用途,或者可以是在医学之外的用途。医学之外的用途的例子可以例如包括在农业中的用途,其中抑制生物可能是有益的,或否则会干扰农业生产(例如杂草),所述生物是寄生性和/或致病性生物、和/或是植物的害虫、和/或是家畜动物(如牛或家禽)的害虫。或者,该组合物也可用作防腐剂,例如用于易于微生物腐败的产品,如食品、个人健康护理产品或包含适合作为微生物基质的化合物的其他产品。
7.根据另一方面,本发明涉及一种抑制目标物种的方法,包括将所述目标物种暴露于根据本发明的组合物。该方法可以是非治疗方法或治疗方法。
8.根据另一方面,本发明涉及一种生产根据本发明的组合物的方法,包括:
9.‑
提供来自来源物种的dna源,任选包含片段化的来源物种dna;
10.‑
提供不同于来源物种的多种宿主物种的细胞;
11.‑
使所述宿主物种处于允许所述宿主物种以可复制的形式吸收来源物种dna的条件下;
12.‑
任选地,分离宿主物种细胞;
13.‑
任选地,从所述来源物种细胞中分离dna;
14.‑
任选地,将任选地从来源物种细胞中分离的dna片段化。
15.该方法是有益的,因为对目标物种的dna抑制现在可以在目标物种或系统发育相似的物种之外有效地产生。自抑制性dna现在尤其可以在与目标物种系统发育无关的物种中产生。
16.本发明的又一方面涉及一种生产农产品的方法。在该方法中,本发明的组合物用于抑制生物,所述生物是否则会干扰农业生产的感染和/或寄生性生物、和/或是植物或家畜动物(如牛)或家禽的害虫。通过抑制生物,所述生物是感染和/或寄生性生物和/或是植物或家畜动物的害虫,可以提高这些生产生物的农业产量。
附图说明
17.图1显示了实验1的抑制试验的结果。显示了拟南芥商业文库对同种幼苗存活的影响。数据指每个处理10个重复(培养皿有10个幼苗)的平均值和标准偏差。与对照相比的显著差异用星号标记(*,p<0.05;**,p<0.01;***,p<0.001)。
18.图2显示了实验2的抑制试验。显示了香附子基因组bac文库在两种不同基质类型上对同种幼苗生长的影响。数据指每个处理10个重复盆的平均值和标准偏差。与对照相比的显著差异用星号标记(*,p<0.05;**,p<0.01;***,p<0.001)。
19.图3显示了实验3的抑制试验的结果。显示了生长培养基中秀丽隐杆线虫基因组bac文库对同种胚胎致死率(顶部)和发育缺陷(底部)的影响,比较对象是包括异源dna但不包括所述自身dna基因组文库的大肠杆菌菌株(epi300)。数据指每个处理12个重复的平均值和标准偏差。
20.图4显示了实验4的抑制试验的结果。显示了海灰翅夜蛾商业文库对同种幼虫存活的影响。数据指每个处理10个重复的平均值和标准偏差。与对照相比的显著差异用星号标记(*,p<0.05;**,p<0.01;***,p<0.001)。
21.图5显示了实验9,调查结束时不同处理区块中马齿苋植物数量的总和。
22.图6显示了agp被子植物的系统发育树(2009年),用于实验10以确定系统发育距离。
23.图7显示了7个目标植物物种的根生长,这些植物物种用自身dna或从异源物种中提取的异源dna进行处理。每个条形指4次重复生物测定的平均值和标准偏差。在每一组中,对用于提取dna的植物物种根据与目标物种的系统发育距离(如实验10的方法中所述确定)进行排序。
具体实施方式
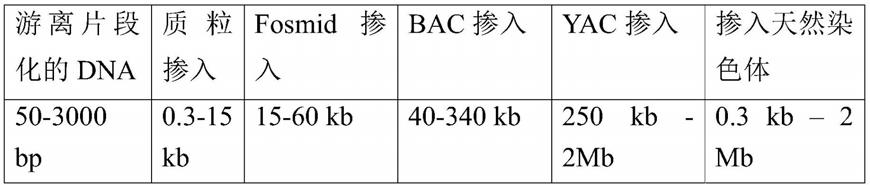
24.本发明的组合物包含dna分子的混合物,所述dna分子的混合物具有来自不同来源的dna序列。所述dna分子的混合物包括来源物种的多个第一dna序列和不同于所述来源物种的宿主物种的多个第二dna序列。正如技术人员将理解的,术语“物种”指的是抽象的概念,并且物种本身不能被抑制。因此,对物种的提及应理解为意指该物种的个体或生物体,例如该物种的多个个体或生物体,即种群。
25.正如术语混合物所暗示的,该组合物包含不同的dna分子。组合物中的dna分子在序列上不同,优选在大小上也不同。组合物中的dna分子的dna序列因来源不同而不同。特别地,第一组dna分子具有第一物种即来源物种的序列,第二组dna分子具有不同于所述来源物种的第二物种即宿主物种的序列。此外,来源物种dna序列的数量也不同。此外,宿主物种dna序列的数量也不同。因此,在来源物种dna序列群体和宿主物种dna序列群体中也存在差异。
26.在该组合物中,可以将来源物种的dna序列,优选作为染色体dna的部分序列(片段),掺入宿主物种天然基因组的dna分子中,例如来自宿主物种细胞天然基因组的染色体或天然质粒,或者可以掺入引入宿主物种细胞的人工dna构建体中,从而形成嵌合dna。或者,组合物中的来源物种dna序列已经从这种嵌合dna中破坏性地释放出来,例如通过断裂。因此,组合物中的dna分子可以包含一个或多个物种的全部(完整)天然染色体、物种的全部(完整)天然染色体外dna(如天然质粒)、全部(完整)人工dna构建体(如选自质粒、粘粒、fosmids或人工染色体)、物种的片段化天然染色体、物种的片段化天然染色体外dna(如天然质粒)、或片段化人工构建体(如选自质粒、粘粒、formids或人工染色体)。在本发明的组合物中,这样的dna分子也可以组合存在。本领域技术人员清楚这些替代的dna分子在序列和大小上可以不同以及如何不同。
27.包含来源物种dna序列的dna分子和包含宿主物种dna序列的dna分子优选是dna片段。本领域技术人员将理解,术语“片段化的dna”和相关术语如“dna片段”是指较大dna分子的部分链(或部分序列)。dna片段可以通过dna片段化(dna分子的分解)或从多核苷酸模板开始合成dna来获得。因此,dna片段是具有较大dna分子的部分序列的dna分子。在本发明的上下文中,抑制性dna片段优选通过dna片段化获得。在本发明的上下文中,术语“包括”和相关术语“例如包括”,包含“由...组成”。
28.可以通过任何合适的方法产生dna片段,特别是通过随机dna片段生成技术。wo2014/020624公开了许多用于产生自抑制性dna片段的合适技术。其中,公开了对来自来源物种的分离的dna进行超声波处理、热处理或热解。或者,从来源物种的dna开始的随机dna片段合成也可以用于随机dna片段的产生。这些和其他用于产生dna片段的技术是技术人员已知的,并且它们可基于可获得的普通常识被使用。可以进一步参考mann和krull(mann和krull 2004.biosensors and bioelectronics 20:945
–
955)、tong et al,(tong et al,2006.nat.protoc,1(2):729
‑
748)、fan et al(fan et al,(2008).nucleic acid research,36(19):e125)和用于pico的标准方案dna剪切(可通过超声波仪网站www.diagenode.com获得)。或者,核酸内切酶,如dnase i、核酸内切酶v或限制性内切酶,可用于片段化来源物种的dna。
29.dna片段化也经常用于(下一代)dna测序程序。在这种情况下,技术人员也可以从
与dna片段化相关的知识体系中获取知识。
30.此外,合成基因组序列部分的dna合成方法也可用于获得dna片段。例如,诸如随机pcr技术的pcr技术可能是合适的。此外,在本发明的上下文中,通过逆转录mrna获得的cdna分子可以被认为是dna片段,因为这种cdna分子代表较大的dna分子的部分序列。合适的cdna分子的合成在技术人员的知识范围内。
31.此外,诸如(生物)消化、腐烂和对包含dna的生物材料(例如细胞)作用的类似过程也可能导致生物材料中包含的dna的断裂。因此,不要求包含来源物种dna序列的dna分子是dna片段,因为dna片段也可以通过所述过程从更大的分子中释放出来。
32.所选择的用于产生dna片段的技术优选地是适用于随机dna片段生成的技术。随机dna片段应该理解为随机产生的dna片段。也就是没有预定模式所产生的dna片段。
33.从(下一代)dna测序背景下与dna片段化相关的研究中,已经了解到某些随机dna片段化技术,如(部分)限制性酶消化和超声波处理,可能对基因组的某些区域和/或序列有一定程度的偏好。本领域技术人员从wo2014/020624中公开的自身dna的抑制可以理解,这种偏好与随机产生的自身dna片段的抑制作用无关。
34.在本发明的组合物中,优选以双链dna分子的形式提供dna分子(dsdna)。这并不一定意味着dna分子以双链状态执行其生物抑制功能。不希望受这一理论的束缚,对于双链dna分子来说,来自(部分)解链的双链dna的单链dna序列参与了抑制作用似乎是合理的。因此,双链形式的dna分子可以通过(部分)dna解链机制提供抑制作用。
35.该组合物可以包含具有大小为约50个碱基(50bp)至约2mb(2*106bp)的来源物种dna序列的dna分子,例如50至3000bp、100至1500bp、或100至2000bp、100至2500bp、或可选地0.3
‑
15kb(例如1
‑
10kb、例如5
‑
10kb)、或15
‑
60kb(例如15
‑
40、25
‑
40kb)、24
‑
或者可选地40
‑
340kb(例如50
‑
200kb、例如120
‑
300kb、例如200
–
300kb)、或者可选地250kb
‑
2mb(例如400kb
‑
1500kb或400kb
‑
1000kb)、或者可选地0.3kb
–
2mb(例如1kb
–
2mb、例如10kb
–
2mb、例如100kb
–
2mb、例如360kb
‑
1mb。根据来源物种的dna序列是否是游离的片段dna序列,或者其是否包含在包含非来源物种dna序列的更大的dna分子中,其大小可能会有所不同。游离片段化的dna序列优选在较低范围的方向上具有更大的尺寸。整合到较大的dna分子中的来源物种dna序列将具有最大的尺寸范围,这取决于哪些可以稳定地整合到相应的dna分子中。技术人员将知道并理解不同的克隆和/或表达载体可以适应不同大小的dna插入物。表1提供了设想用于本发明某些实施方案的不同克隆/表达载体系统的概述,根据这些实施方案它们各自的插入片段大小。如果组合物包含人工dna构建体(载体,特别是克隆载体)的dna,优选人工dna构建体选自质粒、粘粒、fosmid或人工染色体,例如细菌人工染色体、酵母人工染色体或真菌人工染色体。
36.表1
[0037][0038]
该组合物优选包含来源物种和/或宿主物种的染色体dna片段。本领域技术人员将
知道术语“染色体dna”的范围,并将能够提供可从中衍生出dna片段的某一生物体的染色体dna。
[0039]
抑制性dna序列来源的物种在本说明书和权利要求书中被称为“来源物种”。很明显,术语“来源物种”被用作参考术语。该术语用于容易地区分来源物种与“宿主物种”和“目标物种”,这将在下面进一步讨论。
[0040]
根据某些实施方案,在本发明的组合物中,dna分子可以是游离的dna。术语“游离的dna”指的是dna片段没有被掺入到包膜(如细胞或病毒颗粒)中的状态。游离的dna片段可以以任何合适的形式提供。例如,游离的dna片段可以在合适溶剂的溶液中,例如水或含水混合物。溶剂可以是单相溶液或多相溶液的一部分,例如乳液或分散体。或者,游离的dna片段可以固体形式存在于本发明的组合物中。含有游离dna的固体可以是单相固体或多相固体,如分散体。
[0041]
根据某些实施方案,在本发明的组合物中,dna分子的混合物包含嵌合的dna分子,其中来源物种dna序列在至少一端侧接于非来源物种dna序列。本领域技术人员将知道术语“嵌合的dna分子”的含义,并将理解该术语是指包含至少两种不同来源的dna序列的dna分子,例如来自第一物种和至少一种不同来源,例如来自不同于第一物种的第二物种。因此,来源物种的dna序列和至少一个侧接非来源物种的dna序列处在单个dna分子中。嵌合的dna分子也可以称为杂交的dna分子。
[0042]
在嵌合的dna分子中,来源物种dna序列至少在一端侧接于非来源物种dna序列。在本发明的描述中,术语“至少一个”等同于“多个”,并且每次使用这些术语中的任何一个时,都意味着“一个或多个”,反之亦然。在某些优选实施方案中,这些术语表示多个,如2、3、4、5、6、7、8、9或10个,如果合适的话。本领域技术人员将会知道,由于dna分子的结构,在嵌合dna分子中,来源物种dna序列最多可以在两端侧接非来源物种dna序列。
[0043]
本领域技术人员将会理解,侧接dna序列占据相互侧的位置。因此,术语“侧接”在本发明的定义中用于表示“边界”或“邻接”或“毗连”,并且可以与这些术语互换。尽管在上下文中已经暗示了这一点,其中使用了术语“侧接”(及其任何等效术语),但是为了清楚起见,明确指出来源物种dna序列在单个dna分子中连接到至少一个侧接非来源物种。
[0044]
非来源物种dna序列是来源不同于来源物种的dna序列。非来源物种dna序列可以是来自不同于来源物种的物种的天然存在的dna序列,例如来自宿主物种的染色体dna序列。或者,非来源物种dna序列可以是人工dna序列,例如来自人工dna构建体,例如来自人工质粒、粘粒、fosmid或人工染色体的dna序列。在本发明的上下文中,人工染色体可以选自酵母或真菌人工染色体或细菌人工染色体。
[0045]
通过本领域技术人员已知的方法可以获得嵌合的dna分子,所述嵌合的dna分子包含在至少一端侧接非来源物种dna序列的来源物种dna序列。根据某些实施方案,其中来源物种dna序列是染色体dna序列,而非来源物种dna序列是来自人工质粒的序列,来源物种的染色体dna(可能与总基因组的其他部分的dna一起)可以被分离和(随机)片段化,例如通过(部分)限制性酶消化。消化的来源物种dna随后可以连接到用相容的限制性酶切割的人工质粒中,从而产生一群嵌合质粒,其中掺入了来源物种dna的不同部分序列(片段)。包含这种混合的质粒群和类似的或衍生的组合物的组合物,例如其中质粒被(随机)片段化成dna片段,是本发明组合物的实施方案。根据其他实施方案,其中来源物种dna序列是染色体dna
baumannii)、或以色列放线菌(actinomyces israelii)、或戈氏放线菌(actinomyces gerencseriae)、或丙酸杆菌(propionibacterium propionicus)、或布氏锥虫(trypanosoma brucei)、或hiv(人类免疫缺陷病毒)、或溶组织内阿米巴(entamoeba histolytica)、无形体种(anaplasma species)、或管圆线虫种(angiostrongylus species)、或异尖线虫种(anisakis species)、或炭疽杆菌(bacillus anthracis)、或溶血隐秘杆菌(arcanobacterium haemolyticum)、或胡宁病毒(junin virus)、或蛔虫(ascaris lumbricoides)、或曲霉菌(aspergillus species)、或星状病毒科的物种、或巴贝虫属(babesia)物种、或蜡状芽孢杆菌(bacillus cereus)、或拟杆菌属物种(bacteroides species)、或大肠杆菌属(balantidium coli)、或巴尔通体(bartonella)、或蛔虫(baylisascaris)物种、或bk病毒、或何德毛结节菌(piedraia hortae)、或芽囊原虫属(blastocystis)的物种、或皮炎芽生菌(blastomyces dermatitidis)、或马丘坡病毒(machupo virus)、或肉毒梭菌属(clostridium botulinum)、或萨拜病毒(sabi
á
virus)、或布氏菌属的物种(brucella species)、或鼠疫杆菌属(yersinia pestis)、或洋葱伯克霍尔德氏菌(burkholderia cepacia)、或其它伯克霍尔德氏菌种、或溃疡分枝杆菌(mycobacterium ulcerans)、或杯状病毒科(caliciviridae family)、或弯曲杆菌属(campylobacter)物种、或白色念珠菌(candida albicans)、或其它念珠菌属、或菲律宾毛细管菌(capillaria philippinensis)、或嗜气毛细管菌(capillaria aerophila)、或杆状巴尔通体(bartonella bacilliformis)、或汉赛巴尔通体(bartonella henselae)、或a组链球菌属、或葡萄球菌属、或克氏锥虫、或杜克雷嗜血杆菌、或水痘带状疱疹病毒(vzv)、或α病毒、或沙眼衣原体、或肺炎衣原体、或霍乱弧菌、或壶菌、或华支睾吸虫、或艰难梭菌、或粗球孢子菌和波萨达斯球孢子菌、或科罗拉多蜱热病毒(ctfv)、或鼻病毒属、或冠状病毒、或prnp、或克里米亚
‑
刚果出血热病毒、或新型隐球菌、或隐孢子虫种、或巴西钩虫、或环孢子虫、或猪带绦虫、或巨细胞病毒属、或登革热病毒(den
‑
1、den
‑
2、den
‑
3和den
‑
4)或黄病毒属、或绿藻、或栅藻、或脆弱双核阿米巴、或白喉棒状杆菌、或裂头绦虫、或麦地那龙线虫、或埃博拉病毒(ebov)、或棘球绦虫、或埃立克体、或蛲虫、或肠球菌、或肠道病毒、或普氏立克次体、或细小病毒b19、或人类疱疹病毒6(hhv
‑
6)、或人类疱疹病毒7(hhv
‑
7)、或肝片形吸虫、或巨片形吸虫、或布氏姜片吸虫、或prnp、或丝状总科(filarioidea superfamily)、或产气荚膜梭菌、或多种、或梭杆菌种、或产气荚膜梭菌、或其它梭菌种、或白地霉、或蓝氏贾第鞭毛虫、或鼻疽伯克霍尔德氏菌、或棘颚口线虫、或刚颚口线虫、或淋病奈瑟菌、或肉芽肿克雷伯菌、或化脓性链球菌、或无乳链球菌、或流感嗜血杆菌、或肠道病毒、如柯萨奇a病毒和肠道病毒71型(ev71)、或辛诺柏病毒、或中原病毒、或幽门螺杆菌、或大肠杆菌o157:h7、o111和o104:h4、或来自布尼亚病毒科的物种、或亨德拉病毒、或甲型肝炎病毒、或乙型肝炎病毒、或丙型肝炎病毒、或丁型肝炎病毒、或戊型肝炎病毒、或单纯疱疹病毒1型和2型(hsv
‑
1和hsv
‑
2)、或荚膜组织胞浆菌、或十二指肠钩虫和美洲钩虫、或人博卡病毒(hbov)、或尤因埃立克体、或嗜吞噬细胞无形体、或人偏肺病毒(hmpv)、或查菲埃立克体、或人乳头瘤病毒之一、或人副流感病毒(hpiv)、或短膜壳绦虫和长膜壳绦虫、或爱泼斯坦
‑
巴尔病毒(ebv)、或正粘病毒科、或贝氏等孢球虫、或金格杆菌、或拉沙病毒、或嗜肺军团菌、或嗜肺军团菌、或利什曼原虫种、或麻风分枝杆菌、或瘤状分枝杆菌、或钩端螺旋体种、或单核细胞增生李斯特菌、或伯氏疏螺旋体、或伽氏疏螺旋体、或阿氏疏螺旋体、或班氏吴策线虫、或马来布鲁
nana)、或长膜壳绦虫(hymenolepis diminuta)、或牛带绦虫、或猪带绦虫、或锐突伯特绦虫、或司氏伯特绦虫、或迭宫绦虫、或erinaceieuropaei、或埃及血吸虫(schistosoma haematobium)、或日本血吸虫(schistosoma japonicum)、或湄公河血吸虫(schistosoma mekongi)、或echinostoma echinatum、或再生毛毕吸虫(trichobilharzia regenti)、或血吸虫科(schistosomatidae)、或钩虫(ancylostoma)、或管圆线虫(costaricensis)、或异尖线虫(anisakis)、或蛔虫属蛔虫(ascaris sp.ascaris)或蛔虫(lumbricoides)或贝氏蛔虫(baylisascaris)、或procyonis、或马来丝虫(brugia malayi)、或brugia、或timori、或巨肾虫(dioctophyme renale)或龙线虫属、或麦地那龙线虫、或蛲虫、或vermicularis、或enterobius gregorii、或棘颚口线虫、或spinigerum、或gnathostoma、或hispidum、或halicephalobus、或gingivalis、或loa loa filaria、或mansonella、或streptocerca、或onchocerca volvulus、或strongyloides、或stercoralis、或thelazia、或californiensis、或thelazia callipaeda、或toxocara canis、或toxocara cati、或trichinella spiralis。
[0052]
类似于就致病目标物种标注的内容,应当理解,寄生目标物种的抑制不需要在动物(包括人)体内或身体上。相反,这种抑制也可能超出了动物身体的范围。例如用于抑制培养物中的目标物种。优选从皮肤寄生虫和/或胃肠寄生虫和/或粘膜寄生虫中选择寄生目标物种,特别是从上面直接给出的名单中选择。从原生生物或线虫中选择寄生目标物种是优选的,特别是从上面直接给出的名单中选择原生生物和线虫。
[0053]
根据某些实施方案,目标物种可以选自对植物致病的物种,例如选自真菌或卵菌属或细菌或病毒或原生生物或镰孢属(fusarium spp.)、或蒂氏菌属(thielaviopsis spp.)、或轮枝菌属(verticillium spp.)、或者麦格纳波提种(magnaporthe spp.)、或稻瘟病菌(magnaporthe grisea)或菌核菌属(sclerotinia spp.)、或核盘菌属(sclerotinia sclerotiorum)、或植物盘菌属(phytophtora spp.)、或腐霉属(pythium spp.)、疟原虫属(plasmodiophora spp.)、或海绵孢属(spongospora spp.)、或植物致病杆菌(phytopathogenic bacilli)、或欧文氏菌属(erwinia spp.)、或土壤杆菌属(agrobacterium spp.)、或伯克霍尔德氏菌属(burkholderia spp.)、或变形菌(proteobacteria)、或黄单胞菌属、或假单胞菌属、或植原体属(phytoplasma spp.)、或螺原体属(spiroplasma spp.)的植物病原体。
[0054]
当靶物种被选择为侵染物种时,它可以选自农业害虫,例如农业害虫节肢动物,例如选自如下物种:条叶甲(acalymma)、或苜蓿无网长管蚜(acyrthosiphon kondoi)、或棉长管蚜(acyrthosiphon gossypii)、或豌豆蚜(acyrthosiphon pisum)、或非洲粘虫(african armyworm)、或非洲化蜜蜂(africanized bee)、或潜蝇科(agromyzidae)、或小地老虎(agrotis ipsilon)、或agrotis munda、或agrotis orthogonia(西部灰地老虎)、或agrotis porphyricollis、或akkaia taiwana、或aleurocanthus woglumi、或aleyrodes proletella、或alphitobius diaperinus、或alsophila aescularia、或葡萄跳甲(altica chalybea)、或南瓜缘蝽(anasa tristis)、或奥地利金龟子(anisoplia austriaca)、或anthonomus pomorum、或anthonomus signatus、或红蚧(aonidiella aurantii)、或黄圆蹄盾蚧(aonidiella citrina)、或木瓜圆蚧(aonidiella orientalis)、或apamea apamiformis、或apamea niveivenosa、或蚜虫(aphid)、或棉蚜虫(aphis gossypii)、或
aphis nasturtii、或苹果实蝇(apple maggot)、或阿根廷蚁(argentine ant)、或原切根虫(army cutworm)、或草地粘虫(fall armyworm)、或arotrophora arcuatalis、或灰粉虱(ash whitefly)、或竹叶扁蚜(astegopteryx bambusae)、或astegopteryx insularis、或小舞蚜(astegopteryx minuta)、或链蚧属(asterolecanium)、或asterolecanium coffeae、或中华毛庶芒蝇(atherigona reversura)、或athous haemorrhoidalis、或aulacophora、或茄粗额蚜(aulacorthum solani)、或澳洲疫蝗(australian plague locust)、或bactericera cockerelli、或实蝇(bactrocera)、或番石榴果实蝇(bactrocera correcta)、或bagrada hilaris、或knulliana、或甜菜夜蛾(beet armyworm)、或黑豆蚜虫(black bean aphid)、或绿盲蝽(blepharidopterus chlorionis)、或博贡蛾(bogong moth)、或棉铃象甲(boll weevil)、或棉铃虫(bollworm)、或菜蚜(brevicoryne brassicae)、或褐飞蝗(brown locust)、或臭大姐(brown marmorated stink bug)、或褐飞虱(brown planthopper)、或卷心菜蛾(cabbage moth)、或卷心菜虫(cabbage worm)、或四纹豆象(callosobruchus maculatus)、或胡萝卜茎蝇(carrot fly)、或坚蚜属(cerataphis brasiliensis)、或ceratitis aliena、或ceratitis andranotobaka、或地中海实蝇(ceratitis capitata)、或ceratitis flexuosa、或ceratitis grahami、或ceratitis ovalis、或ceratitis penicillata、或纳塔尔小条实蝇(ceratitis rosa)、或坚角蚜属(ceratoglyphina bambusae)、或无翅孤雌蚜(ceratopemphigus zehntneri)、或甘蔗绵蚜(ceratovacuna lanigera)、或黑角负泥虫(cereal leaf beetle)、或四红中瘤钉毛蚜(chaetosiphon tetrarhodum)、或大麦条足杆蝇(chlorops pumilionis)、或citrus long
‑
horned beetle、或褐软蜡蚧(coccus hesperidum)、或咖啡绿软蜡蚧(coccus viridis)、或苹果蠹蛾(codling moth)、或coffee borer beetle、或美洲苜蓿粉蝶(colias eurytheme)、或马铃薯甲虫(colorado potato beetle)、或common blossom thrips、或杂拟谷盗(confused flour beetle)、或谷实夜蛾(cotton bollworm)、或草螟属(crambus)、或巴西玉米根叶甲复合群(cucumber beetle)、或欧洲栗象(curculio elephas)、或榛子象(curculio nucum)、或美国榛实象(curculio occidentis)、或地老虎属(cutworm)、或cyclocephala borealis、或dargida diffusa、或油菜荚叶瘿蚊(dasineura brassicae)、或date stone beetle、或delia(fly)、或葱蝇(delia antiqua)、或萝卜地种蝇(delia floralis)、或灰地种蝇(delia platura)、或甘蓝种蝇(delia radicum)、或家黑皮蠹(dermestes ater)、或dermolepida albohirtum、或沙漠蝗(desert locust)、或瓜叶甲(diabrotica)、或带斑黄瓜叶甲(diabrotica balteata)、或南美叶甲(diabrotica speciosa)、或diamondback moth、或瓜绢野螟(diaphania indica)、或黄瓜绢野螟(diaphania nitidalis)、或柑桔木虱(diaphorina citri)、或蔗根非耳象(diaprepes abbreviatus)、或小蔗杆草螟(diatraea saccharalis)、或异黑蝗(differential grasshopper)、或苏丹棉铃虫(diparopsis castanea)、或摩洛哥蝗(dociostaurus maroccanus)、或樱桃果蝇(drosophila suzukii)、或板栗瘿蜂(dryocosmus kuriphilus)、或西圆尾蚜属(dysaphis crataegi)、或菠萝洁粉蚧(dysmicoccus brevipes)、或金钢钻属(earias perhuegeli)、或北美豆芫菁(epicauta vittata)、或食植瓢虫(epilachna)、或美国马铃薯跳甲(epitrix cucumeris)、或块茎跳甲(epitrix tuberis)、或香蕉弄蝶(erionota thrax)、或苹果绵蚜(eriosoma lanigerum)、或绵蚜亚科(eriosomatinae)、或
芹菜拟迈实蝇(euleia heraclei)、或eumetopina flavipes、或欧洲玉米螟(european corn borer)、或eurydema oleracea、或麦扁盾蝽(eurygaster integriceps)、或桔腺刺粉蚧(ferrisia virgata)、或forest bug、或麦花蓟马(frankliniella tritici)、或大蜡螟(galleria mellonella)、或黑切根虫(garden dart)、或geoica lucifuga、或假桃病毒叶蝉(glassy
‑
winged sharpshooter)、或温室白粉虱(greenhouse whitefly)、或greenidea artocarpi、或台湾毛管蚜(greenidea formosana)、或greenideoida ceyloniae、或东方蝼蛄(gryllotalpa orientalis)、或gryllotalpa vinae、或gypsy moths in the united states、或棉铃实夜蛾(helicoverpa armigera)、或helicoverpa gelotopoeon、或细点实夜蛾(helicoverpa punctigera)、或谷实夜蛾(helicoverpa zea)、或烟芽夜蛾(heliothis virescens)、或裂臀瓢虫(henosepilachna)、或马铃薯瓢虫(henosepilachna vigintioctomaculata)、或茄二十八星瓢虫(henosepilachna vigintioctopunctata)、或黑森瘿蚊(hessian fly)、或非洲独角仙(heteronychus arator)、或梅大尾蚜(hyalopterus pruni)、或李蔗锈一条蚜(hysteroneura setariae)、或吹绵蚧(icerya purchasi)、或ipuka dispersum、或茶小绿叶蝉(jacobiasca formosana)、或kaltenbachiella elsholtriae、或kaltenbachiella japonica、或谷斑皮蠹(khapra beetle)、或长尾里波灰蝶(lampides boeticus)、或潜叶蝇(leaf miner)、或lema daturaphila、或lepidiota consobrina、或紫牡蛎盾蚧(lepidosaphes beckii)、或榆蛎盾蚧(lepidosaphes ulmi)、或桉树枝瘿姬小蜂(leptocybe invasa)、或leptoglossus zonatus、或leptopterna dolabrata、或lesser wax moth、或银潜蛾属(鳞翅目)、或咖啡潜叶蛾(leucoptera caffeina)、或light brown apple moth、或苹果浅褐卷叶蛾(light brown apple moth controversy)、或萝卜蚜(lipaphis erysimi)、或拉美豌豆斑潜蝇(liriomyza huidobrensis)、或稻水象甲(lissorhoptrus oryzophilus)、或阿根廷茎象甲(listronotus bonariensis)、或长尾弄蝶(long
‑
tailed skipper)、或草盲蝽属(lygus)、或豆荚草盲蝽(lygus hesperus)、或蔷薇鳃角金龟(macrodactylus subspinosus)、或伪蒿小长管蚜(macrosiphoniella pseudoartemisiae)、或菊小长管蚜(macrosiphoniella sanborni)、或马铃薯长管蚜(macrosiphum euphorbiae)、或玉米象(maize weevil)、或烟草天蛾(manduca sexta)、或matsumuraja capitophoroides、或mayetiola hordei、或粉蚧科(mealybug)、或筛豆龟蝽(megacopta cribraria)、或高粱蚜(melanaphis sacchari)、或melittobia australica、或蛾蜡蝉(metcalfa pruinosa)、或墨西哥大豆瓢虫(mexican bean beetle)、或micromyzus judenkoi、或micromyzus kalimpongensis、或黑小瘤蚜(micromyzus niger)、或鳞翅目(moth)、或雕蛾(leek moth)、或美洲粘虫(mythimna unipuncta)、或冬葱瘤额蚜(myzus ascalonicus)、或苎麻白蚜(myzus boehmeriae)、或樱桃瘤蚜(myzus cerasi)、或myzus obtusirostris、或瘤蚜属(myzus ornatus)、或桃蚜(myzus persicae)、或丝角叶蜂属(nematus)、或鹅莓丝角叶蜂(nematus leucotrochus)、或茶藨子黄叶蜂(nematus ribesii)、或nematus spiraeae、或百合沟新瘤蚜(neomyzus circumflexus)、或金盏花新弓翅蚜(neotoxoptera oliveri)、或稻点绿蝽(nezara viridula)、或栎列队蛾(oak processionary)、或美洲稻缘蝽(oebalus pugnax)、或油橄榄果实蝇(olive fruit fly)、或石刀柏蛇潜蝇(ophiomyia simplex)、或椰蛀蛾(opisina arenosella)、或禾蝇属(opomyza)、或禾蝇(opomyza florum)、或禾蝇科(opomyzidae)、或
亚洲稻瘿蚊(orseolia oryzae)、或大眼锯谷盗(oryzaephilus mercator)、或瑞典麦秆蝇(oscinella frit)、或亚洲玉米螟(ostrinia furnacalis)、或棉籽尖长蝽(oxycarenus hyalinipennis)、或橙体凤蝶(papilio demodocus)、或木瓜秀粉蚧(paracoccus marginatus)、或椰子犀角金龟(paratachardina pseudolobata)、或paropsisterna selmani、或印度黄脊蝗(patanga succincta)、或甜菜棉蚜(pemphigus betae)、或香蕉交脉蚜(pentalonia nigronervosa)、或蝽总科(pentatomoidea)、或疆夜蛾(peridroma saucia)、或忽布疣蚜(phorodon humuli)、或马铃薯块茎蛾(phthorimaea operculella)、或食叶鳃金龟属(phyllophaga)、或大豆淡足跳甲(phyllotreta nemorum)、或根瘤蚜科(phylloxeridae)、或根瘤蚜总科(phylloxeroidea)、或豌豆潜叶蝇(phytomyza horticola)、或大菜粉蝶(pieris brassicae)、或棉红铃虫(pink bollworm)、或桔臀纹粉蚧(planococcus citri)、或苹果芽小卷蛾(platynota idaeusalis)、或李象(plum curculio)、或加州锯天牛(prionus californicus)、或锉叶蜂属(pristiphora)、或居竹伪角蚜(pseudoregma bambucicola)、或东非可可缘蝽(pseudotheraptus wayi)、或油菜金头跳甲(psylliodes chrysocephala)、或白斑蛛甲(ptinus fur)、或紫斑谷螟(pyralis farinalis)、或南瓜守瓜(raphidopalpa foveicollis)、或红火蚁(red imported fire ant)、或红蝗(red locust)、或欧洲樱桃实蝇(rhagoletis cerasi)、或西部樱桃绕实蝇(rhagoletis indifferens)、或越桔绕实蝇(rhagoletis mendax)、或黄蔷薇蚜(rhodobium porosum)、或马铃薯囊管蚜(rhopalosiphoninus latysiphon)、或玉米蚜(rhopalosiphum maidis)、或禾谷缢管蚜(rhopalosiphum padi)、或红腹缢管蚜(rhopalosiphum rufiabdominale)、或美杉梢小卷蛾(rhyacionia frustrana)、或红棕象甲(rhynchophorus ferrugineus)、或棕榈象甲(rhynchophorus palmarum)、或亚棕象甲(rhynchophorus vulneratus)、或谷蠹(rhyzopertha)、或米蛾(rice moth)、或俄国麦蚜(russian wheat aphid)、或橄榄黑盔蚧(saissetia oleae)、或梨圆蚧(san jose scale)、或蚧壳虫(scale insect)、或美洲沙漠蝗(schistocerca americana)、或麦二叉蚜(schizaphis graminum)、或超管二叉蚜(schizaphis hypersiphonata)、或小二叉蚜(schizaphis minuta)、或圆腹二叉蚜(schizaphis rotundiventris)、或黄刚毛蚜(schoutedenia lutea)、或尖眼蕈蚊科(sciaridae)、或茶黄蓟马(scirtothrips dorsalis)、或盾蝽科(scutelleridae)、或scutiphora pedicellata、或三叶斑潜蝇(serpentine leaf miner)、或八字地老虎(setaceous hebrew character)、或朴绵叶蚜(shivaphis celti)、或丫蚊夜蛾(silver)、或b型烟飞虱(silverleaf whitefly)、或樟修尾蚜(sinomegoura citricola)、或美甘蔗伪毛蚜(sipha flava)、或禾谷网蚜(sitobion avenae)、或sitobion lambersi、或sitobion leelamaniae、或荻草谷网蚜(sitobion miscanthi)、或sitobion pauliani、或sitobion phyllanthi、或sitobion wikstroemiae、或浅黄根瘤象(sitona lepidus)、或直条根瘤象(sitona lineatus)、或蜂巢小甲虫(small hive beetle)、或西南玉米杆草螟(southwestern corn borer)、或大豆蚜(soybean aphid)、或圆灰翅夜蛾(spodoptera cilium)、或斜纹夜蛾(spodoptera litura)、或黄瓜十一星叶甲(spotted cucumber beetle)、或斑点灯笼虫(spotted lanternfly)、或葫芦透翅蛾(squash vine borer)、或菜豆蛇潜蝇(stemborer)、或双斑纤盲蝽(stenotus binotatus)、或向日葵斯实蝇(strauzia longipennis)、或黄曲条菜跳甲(striped flea beetle)、或麦扁盾蝽(sunn pest)、或甘薯
ater、或桤木绿叶甲(leaf beetle)、或苜蓿叶蝉(leafhopper)、或萝卜蚜(lipaphis erysimi)、或四点叶蝉(macrosteles quadrilineatus)、或粉蚧属(mealybug)、或西瓜寡鬃实蝇(melon fly)、或象虫科(molytinae)、或藜泉蝇(pegomya hyoscyami)、或木蠹象属(pissodes)、或白松木蠹象(pissodes strobi)、或木蠹象族(pissodini)、或飞虱(planthopper)、或葡萄粉蚧(pseudococcus maritimus)、或暗色粉蚧(pseudococcus viburni)、或梨喀木虱(psylla pyri)、或木虱科(psyllidae)、或梢瘿蚊属(rabdophaga clavifex)、或棕榈象甲(rhynchophorus palmarum)、或葡萄带叶蝉(scaphoideus titanus)、或茶黄蓟马(scirtothrips dorsalis)、或银叶粉虱(silverleaf whitefly)、或实蝇科(tephritidae)、或蓟马科(thripidae)、或棕榈蓟马(thrips palmi)、或松小蠹(tomicus piniperda)、或大橘蚜(toxoptera citricida)、或角蝉(treehopper)、或个木虱科(triozidae)、或西花蓟马(western flower thrips)、或光材小蠹(xyleborus glabratus)。
[0057]
根据某些优选的实施方案,害虫物种被选择为寄生于植物的线虫物种,特别是选自根结线虫属,如花生根结线虫(m.arenaria)、南方根结线虫(m.incognita)、爪哇根结线虫(m.javanica)、北方根结线虫(m.hapla),或选自异皮线虫(hetrodera)属,如大豆胞囊线虫(heterodera glycines)、或燕麦胞囊线虫(heterodera avenae)和菲利普孢囊线虫(h.filipjevi),或选自球胞囊线虫属,例如马铃薯白线虫(globodera pallida)、或马铃薯金线虫(g.rostochiensis)、或选自短体线虫属,例如穿刺短体线虫(p.penetrans)、索氏短体线虫(p.thornei)、落选短体线虫(p.neglectus)、玉米短体线虫(p.zeae)、伤残根腐线虫(p.vulnus)或咖啡短体线虫(p.coffeae),或选自穿孔线虫属,例如香蕉穿孔线虫(radopholus similis)。
[0058]
根据某些实施方案,侵染物种可以选自杂草物种。在本发明中被认为是目标物种的杂草物种是例如来自如下的杂草物种:泽泻科(alismataceae)、或伞形科(apiaceae)、或菊科(asteraceae)、或苋科(amaranthaceae)、或仙人掌科(cactaceae)、或石竹科(caryophyllaceae)、或藜科(chenopodiaceae)、或蕨藻科(caulerpaceae)、或鸭跖草科(commelinaceae)、或禾本科(poaceae)、或马齿苋科(portulacaceae)、或大戟科(euphorbiaceae)、或豆科(fabaceae)(leguminosae)、或茜草科(rubiaceae)、或水鳖科(hydrocharitaceae)、或满江红科(azollaceae)、或槐叶苹科(salviniaceae)、或鸢尾科(iridaceae)、或百合科(liliaceae)、或雨久花科(pontederiaceae)、或野牡丹科(melastomataceae)、或桃金娘科(myrtaceae)、或蓼科(polygonaceae)、或海金沙科(lygodiaceae)、或蔷薇科(rosaceae)、或爵床科(acanthaceae)、或列当科(orobanchaceae)、或玄参科(scrophulariaceae)、或旋花科(convolvulaceae)、或菟丝子科(cuscutaceae)、或茄科(solanaceae)、或黑三棱科(sparganiaceae)。
[0059]
被认为目标物种的特定杂草物种可以选自慈菇(sagittaria sagittifolia linnaeus)、或大豕草(heracleum mantegazzianum sommier&levier)、或紫茎泽兰(ageratina adenophora(spreng.)king&h.e.robins.)、或ageratina riparia(regel)king&h.e.robins.、或赛金盏(arctotheca calendula(l.)levyns)、或尖刺刺苞木(carthamus oxyacanthus m.bieberstein)、或半毛菊(crupina vulgaris cass.)、或旋覆花(inula britannica l.)、或假泽兰(mikania cordata(burm.f.)b.l.robins.)、或薇甘
菊(mikania micrantha kunth)、或无茎大翅蓟(onopordum acaulon l.)、或onopordum illyricum l.、或窄叶黄菀(senecio inaequidens dc.)、或马达加斯加千里光(senecio madagascariensis poir.)、或(羽芒菊tridax procumbens l.)、或虾钳菜(alternanthera sessilis(l.)r.br.ex dc.)、或opuntia aurantiaca lindley、或drymaria arenarioides humboldt&bonpland、或salsola vermiculata l.、或caulerpa taxifolia(vahl)c.agardth、或圆叶鸭跖草(commelina benghalensis l.)、或不实野燕麦(avena sterilis linnaeus)、或竹节草(chrysopogon aciculatus(retz.)trin.)、或digitaria abyssinica(a.rich)stapf、或天鹅绒马唐(digitaria velutina(forsk.)beauv.)、或imperata brasiliensis trinius、或白茅(imperata cylindrica(l.)beauv.)、或田间鸭嘴草(ischaemum rugosum salisbury)、或千金子(leptochloa chinensis(l.)nees)、或nassella trichotoma hackel ex arech.、或oryza longistaminata a.chev.&roehr.、或野生稻(oryza punctata kotzchy ex steud.)、或oryza rufipogon griffiths、或鸭乸草(paspalum scrobiculatum linnaeus)、或pennisetum clandestinum hochst.ex chiov.、或大狼尾草(pennisetummacrourum trinius)、或花梗狼尾草(pennisetum pedicellatum trinius)、或牧地狼尾草(pennisetum polystachion(linnaeus)schultes)、或筒轴茅(rottboellia cochinchinensis(lour.)w.d.clayton)、或割手密(saccharum spontaneum l.)、或幽狗尾草(setaria pumila ssp.pallidefusca(schumacher)b.k.simon)、或尾稃草(urochloa panicoides beauvois)、或特拉西大戟(euphorbia terracina l.)、或阿拉伯金合欢(acacia nilotica(l.)willd.ex delile)、或山羊豆(galega officinalis l.)、或美洲含羞草(mimosa diplotricha c.wright ex sauvalle)、或含羞树(mimosa pigra l.)、或prosopis alpataco r.a.philippi、或prosopis argentina burkart、或prosopis articulata s.watson、或prosopis burkartii或prosopis caldenia burkart、或prosopis calingastana burkart、或prosopis campestris griesbach、或prosopis castellanosii burkart、或prosopis denudans bentham、或prosopis elata(burkart)burkart、或prosopis farcta(banks&soland.)j.f.macbr.、或prosopis ferox griesbach、或prosopis fiebrigii harms、或prosopis hassleri harms ex hassler、或prosopis humilis gillies ex hooker&arnott、或prosopis kuntzei harms ex hassler、或prosopis pallida(humb.&bonpl.ex willd.)kunth、或prosopis palmeri s.watson、或prosopis reptans benth.、或prosopis rojasiana burkart、或prosopis ruizlealii burkart、或prosopis ruscifolia griesbach、或prosopis sericantha gillies ex hook.&arn.、或prosopis strombulifera(lamarck)bentham、或prosopis torquata(cavan.ex lagasca y segura)dc.、或阔叶丰花草(spermacoce alata aubl.)、或黑藻(hydrilla verticillata(l.f.)royle)、或大卷蕴藻(lagarosiphon major(ridley)moss)、或龙舌草(ottelia alismoides(linnaeus)pers.)、或满江红(azolla pinnata r.brown)、或美洲槐叶萍(salvinia auriculata aublet)、或salvinia biloba raddi、或salvinia herzogii de la sota、或人厌槐叶萍(salvinia molesta d.s.mitchell)、或肖鸢尾(moraea collina thunb.)、或moraea flaccida(sweet)steud.、或moraea miniata andrews、或淡黄肖鸢尾(moraea ochroleuca(salisb.)drapiez)、或moraea pallida(baker)goldblatt、或葱叶阿福花(asphodelus fistulosus linnaeus)、
或eichhornia azurea(swartz)kunth、或箭叶雨久花(monochoria hastata(l.)solms)、或鸭舌草(monochoria vaginalis(burm.f.)k.presl ex kunth)、或印度野牡丹(melastoma malabathricum l.)、或五脉白千层(melaleuca quinquenervia(cav.)blake)、或角刺酸模(emex australis steinhall)、或emex spinosa(linnaeus)campdera、或曲轴海金沙(lygodium flexuosum(l.)sw.(1801)(mobot))、或小叶海金沙(lygodium microphyllum(cav.)r.br.)、或黑莓(rubus fruticosus l.)、或羽萼悬钩子(rubus moluccanus l.)、或小狮子草(hygrophila polysperma(roxb.)t.anders.)、或野菰属(aeginetia spp.l.)、或黑蒴属(alectra spp.thunb.)、或列当属(orobanche spp.(nonnative)l.)、或石龙尾(limnophila sessiliflora(vahl)blume)、或独脚金属(striga spp.lour.)、或蕹菜(ipomoea aquatica forssk.)、或菟丝子属(cuscuta spp.l.)、或非洲枸杞(lycium ferocissimum miers)、或solanum tampicense dunal、或托鲁巴姆(solanum torvum sw.)、或熱帶刺茄(solanum viarum dunal)、或黑三棱(sparganium erectum l.)。
[0060]
可以选作目标物种的引起产品变质的物种可以选自腐败微生物,例如选自细菌,例如革兰氏阴性杆菌(如假单胞菌属(pseudomonas spp.)、希瓦内拉菌属(shewanella spp.))、革兰氏阳性孢子形成菌(如芽孢杆菌属(bacillus spp.)、梭菌属(clostridium spp.)、乳酸菌(lactic acid bacteria)和其他革兰氏阳性菌(如丝裂菌属(brochothrix spp)、微球菌属(micrococcus spp.)、或肠杆菌科enterobacteriaceae(enterobacteriaceae)),真菌,例如接合菌纲,来自青霉属或曲霉属、或酵母如接合酵母属、酵母属、念珠菌属、德克拉酵母属(dekkera(brettanomyces)spp.)。或者,导致产品变质的目标物种可以选自储存的产品螨类,例如选自落新妇属(astigmata),例如选自糖螨科(glycyphagidae)或桃螨科(carpoglyphidae)。
[0061]
根据本发明组合物的某些优选实施方案,该组合物包含来自不同于来源物种的许多物种(宿主物种)的细胞,其中dna分子在宿主物种细胞中。
[0062]
如上所述,术语“宿主物种”仅用作参考术语。在本发明的上下文中,宿主物种通常是不同于来源物种的物种,优选是已在细胞内掺入了来源物种的dna序列、系统发育上不同的物种。根据某些实施方案,系统发育不同的(远缘)物种是来自不同分类目的物种,例如来自不同纲、不同门、不同界或不同域的物种。根据某些实施方案,系统发育上不同的物种是来自不同科的物种,例如来自不同目、不同纲、不同叶下珠、不同王国或不同领域的物种。宿主物种可以选自能够吸收和复制来源物种的外源dna(特别是来源物种dna的片段,例如掺入克隆和/或表达载体的来源物种的片段)的任何物种。从该描述中可以清楚地看出,不要求宿主物种的个体接受和复制来源物种的全部基因组。相反,自身dna对目标物种的抑制作用可以通过多种宿主物种细胞和/或个体来实现,其中不同的细胞和/或个体分别复制来源物种基因组的不同部分,如在dna文库中。因此,宿主物种细胞群体的单个细胞仅包含存在于组合物中的来源物种dna序列总数的一部分。宿主物种可以例如选自多细胞生物或微生物。例如,多细胞植物的细胞可以用作宿主物种细胞。本领域技术人员将会理解,可以通过各种技术将来源物种的dna引入多细胞植物的细胞中,包括例如胚胎细胞培养物的粒子轰击(生物枪法)、农杆菌介导的转化、病毒载体介导的转化。当选作植物时,宿主物种可以选自作物,优选粮食作物,例如选自谷类、玉米、甜菜、油菜籽、豌豆、大豆,油料作物,例如油菜籽、埃塞俄比亚芥菜、向日葵,淀粉作物,例如(甜)高粱,纤维作物,例如亚麻或大麻,木质纤
维素作物,例如芦苇加那利草、巨苇、柳枝稷、芒草、刺桐、短轮伐期林业作物,如柳树、杨树或桉树。
[0063]
或者,蠕虫治疗中使用的蠕虫,例如猪鞭毛虫(trichuris suis)、美洲钩虫(necator americanus)、毛首鞭虫(trichuris trichiura)、缩小膜壳绦虫(hymenolepis diminuta)、蛔虫(ascaris lumbricoides)、粪类圆线虫(strongyloides stercoralis)、蛲虫(enterobius vermicularis)或微小膜壳绦虫(hymenolepis nana)可用作宿主物种。用作宿主物种的蠕虫物种优选是人类的共生物种,特别是猪鞭毛虫、美洲钩虫、毛首鞭虫或缩小膜壳绦虫。已经观察到线虫物种的水平基因转移事件(参见plos negl trop dis.2007oct;1(1):e35;doi:10.1371/journal.pntd.0000035)并且对于治疗性蠕虫可以诱导水平基因转移以将来源物种dna纳入其基因组。
[0064]
根据其他替代实施方案,宿主物种选自微生物。正如本领域技术人员所知,微生物是一种微观生物,它可以是单细胞形式,也可以存在于细胞群体中。在本发明的上下文中,微生物优选选自原核生物,如古细菌或细菌,或真核生物,如真菌(包括酵母)或微植物(微藻)。选自微生物的宿主物种可以选自可以在液体培养物中生长的微生物,例如大肠杆菌,特别是菌株dh5或衍生菌株,或者菌株dh10b或衍生菌株,或者选自酵母属物种,例如酿酒酵母,例如菌株ab1380或衍生菌株,或者来自曲霉属物种,例如黑曲霉、米曲霉或构巢曲霉,或者来自芽孢杆菌属物种,或者来自假单胞菌属物种,或者来自乳酸菌,例如乳杆菌属、明串珠菌属、片球菌属、乳球菌属、肠球菌属、或链球菌属,或来自蓝藻,例如节旋藻属物种,特别是植物节旋藻(arthrospira plantenis)或极大节旋藻(arthrospira maxima)。根据某些实施方案,微生物可以是fda宣布的具有一般认为安全(gras)状态的生物体。或者,土壤微生物,优选不同土壤细菌和/或土壤真菌的混合培养物可以用作宿主物种。用作宿主物种的土壤微生物可以选自α变形菌属、β变形菌属、δ变形菌属、放线菌属、嗜热菌属、红色杆菌属、氯酸杆菌属、酸杆菌属、或solibacteres属。根据某些优选的实施方案,宿主物种选自生物防治剂,例如昆虫病原生物防治剂,例如微生物生物防治剂,特别是细菌生物防治剂,例如苏云金芽孢杆菌或枯草芽孢杆菌,或真菌生物防治剂,例如球孢白僵菌(beauveria bassiana)、玫烟色伊萨菌(isaria fumosorosea)、蜡蚧菌属(lecanicillium spp.)或者绿僵菌属(metarhizium spp.)。生物防治剂可选自例如木霉属(trichoderma spp.)或白粉寄生孢(ampelomyces quisqualis)。宿主物种还可以选自微生物植物生长刺激剂,例如芽孢杆菌物种、木霉物种或分枝杆菌物种。根据某些高度优选的实施方案,当宿主物种是微生物昆虫病原生物防治剂时,目标物种是节肢动物,优选选自农业害虫节肢动物或参与传播植物疾病的节肢动物媒介的节肢动物。上文已经介绍了这些群体中的生物的具体实例。
[0065]
如所解释的,在其中来源物种dna序列在宿主物种细胞中的实施方案中,来源物种dna序列或者被掺入到被引入到宿主物种细胞中的人工可复制dna构建体中,例如人工质粒或人工染色体中,或者被掺入到宿主物种细胞的天然基因组中。通过将来源物种的dna序列掺入到被引入到宿主细胞的人工dna构建体或宿主细胞的天然基因组中,来源物种的dna序列可以与宿主物种的dna一起被宿主物种细胞的dna复制机制复制。本发明的发明人令人惊讶地发现,当与(生产)宿主物种的dna结合时,物种的抑制性dna片段保持抑制性,这为产生和展示自抑制性dna片段开辟了新的可能性。
[0066]
将dna序列掺入dna构建体并将dna构建体引入宿主细胞已经成为生物科学各个领
域的标准做法。因此,本领域技术人员可以容易地在本发明中应用这种方法。此外,众所周知,各种生物可以从它们的环境中吸收dna分子,并可以将这些dna分子整合到它们的天然基因组中。特别是微生物,尤其是原核生物,天然地能够进行这种水平或侧向基因转移。此外,某些已知的基因工程技术,如生物弹道,依赖于在细胞(如植物细胞)的天然基因组中掺入dna片段。
[0067]
此外,转座元件(或转座子)可以促进不同物种的细胞之间的dna转移。
[0068]
优选地,dna分子混合物中来源物种的基因组的覆盖度在20
‑
100%之间,例如20
‑
90%、30
‑
100%、30
‑
90%、40
‑
100%、40
‑
90%、50
‑
100%、50
‑
90%,优选60
‑
100%,例如60
‑
90%、70
‑
100%、70
‑
90%、80
‑
100%、80
‑
90%。基于发明人的实验观察,可以得出在这些范围内,本发明组合物中的来源物种dna序列具有良好的抑制活性。本领域技术人员将知道并理解,dna分子混合物中来源物种基因组的覆盖度与存在于dna分子的整体(群体)中的来源物种dna序列(特别是染色体dna序列)的百分比相关,所述dna分子的整体(群体)存在于组合物中。鉴于实验结果表明总基因组的一部分是抑制性的,也可以预期(片段化的)cdna序列(其也代表基因组总dna序列的一部分)将对其来源的物种和系统发育相似的物种具有抑制活性。然而,优选的是,来源物种的dna序列包含非编码序列,如果来源物种的基因组包含大量的这种非编码序列,那么根据与染色体组织相关的现有知识,可以预期在非编码序列中存在更高的物种特异性。考虑到这一点,优选的是,来源物种的dna序列包括随机片段化的染色体dna。
[0069]
通过在宿主物种细胞中的(嵌合)dna分子中复制来源物种dna序列,来源物种dna序列将与相当数量的宿主物种dna序列相关联,并且可能与来自人工dna构建体的相当数量的dna序列相关联。本发明的发明人惊奇地发现,当与较大量的这些非来源物种的dna序列结合时,来源物种的dna序列保持了其自抑制活性。因此,不需要从组合物中除去宿主物种的dna和/或人工构建体的dna序列。根据某些优选实施方案,本发明的组合物因此包含非来源物种的dna,其中所述非来源物种的dna超过来源物种的dna,优选使得非来源物种的dna与来源物种的dna的比例在10∶1和1000∶1之间,例如在20∶1和500∶1之间,在50∶1和500∶1之间,或在100∶1和500∶1之间。非来源物种dna包括宿主物种dna序列和任选地来自人工dna构建体的dna序列,并且优选基本上由宿主物种dna序列和任选地来自人工dna构建体的dna序列组成,所述人工dna构建体(例如克隆和/或表达载体)例如选自质粒、bac、yac或人工真菌染色体。
[0070]
如上所述,优选地选择这样的来源物种,即由其衍生的抑制性dna序列对目标物种具有抑制性。自wo2014/020624公开以来,对于本领域技术人员来说显而易见的是,抑制性dna序列可以来源于目标物种本身或系统发育相似的物种。在本发明的上下文中,来源物种因此可以选自目标物种或系统发育类似于目标物种的物种。
[0071]
在本发明的上下文中,系统发育相似的物种是具有相似基因组的物种。本领域技术人员将理解,在系统发育上密切相关的物种比在系统发育上较远的物种具有更相似的基因组。因此,系统发育相似意味着具有密切的系统发育关系。因此,可以基于已知的系统发育关系来确定系统发育相似性。因此,根据某些优选实施方案,系统发育上相似的物种是同一分类目内的物种。在某一目中,系统发育相似的物种优选来自同一单系群(进化支),例如来自同一科、同一亚科、同一族、同一亚族、同一属。最优选的是,系统发育相似的物种来自
同一分类科,例如同一亚科、同一族、同一亚族、同一属。
[0072]
此外,确定基因组相似性(或关联性)的技术也很容易获得。基因组相似性可以例如通过测定来自两个物种的基因组的单链dna(ssdna)片段的复性/再结合动力学来确定。替代地,或者另外,可以研究从两个物种的基因组的ssdna片段的混合物中复性的双链dna(dsdna)片段的变性(熔化)。后一种技术允许确定熔化温度t
m
(即一半的dna链处于ssdna状态的温度)和相关的t
50
h。涉及复性/变性动力学和评估熔解曲线的方法是在70年代早期引入的(参见de ley et al.eur j biochem.1970jan;12(1):133
‑
42)用于确定细菌的关联性,但是这些涉及熔化温度曲线分析的方法也已经用于确定真核物种的关联性(例如参见sibley和ahlquist,j mol evol(1984)20:2
‑
15)。
[0073]
如进一步所知,自wo2014/020624公开以来,物种的系统发育相似性可以基于来自一个物种的抑制性dna片段是否对另一个物种也是抑制性的来确定。根据某些其他实施方案,系统发育相似的物种因此是这样的物种,其通过提取的总dna的随机片段化或通过从总dna开始的随机片段合成获得的dna对目标物种具有抑制作用。对于本领域技术人员来说,很明显,基于这一功能定义,可以用类似于wo2014/020624和本文所附实验中的测试来确定系统发育。在同一分类目中,来源物种在系统发育上也与目标物种相似,因为通过提取的总dna的随机片段化或从总dna开始的随机片段合成而从来源物种获得的dna对目标物种具有抑制作用。
[0074]
本发明的组合物旨在抑制目标物种。所述目标物种可以与来源物种相同,或者可以是系统发育上与来源物种相似的物种。为了增强对目标物种的抑制,该组合物可以与多种其它(天然)杀生物剂结合使用,例如(天然)杀虫剂,例如杀真菌剂、杀昆虫剂、杀线虫剂、杀螨剂、杀真菌剂、杀细菌剂或杀藻剂。附加的(天然)杀生物剂可以是生物防治剂,例如微生物生物防治剂,例如细菌生物防治剂或真菌生物防治剂,特别是昆虫病原生物防治剂。根据某些实施方案,组合物中存在额外的(天然)杀生物剂。当额外的(天然)杀生物剂是微生物生物防治剂(例如昆虫病原生物防治剂)时,优选微生物生物防治剂的活细胞作为包含和复制来源物种dna的宿主细胞。例如,来源物种可以选自目标物种或系统发育上与目标物种相似的物种,其中,目标物种选自鳞翅目,特别是选自卷蛾科,例如选自卷蛾属,特别是春色卷蛾(choristoneura orae)、云杉色卷蛾(choristoneura fumiferana)、或西拟色卷蛾(choristoneura freemani),或者选自夜蛾科,例如甜菜夜蛾属,特别是草地贪夜蛾(spodoptera frugiperda)、斜纹夜蛾(spodoptera litura)、spodoptera litoralis、spodoptera cilium、或鸟灰翅夜蛾(spodoptera ornithogalli),或者选自螟蛾(pyralidae)科,如选自谷斑螟(plodia)属或粉斑螟(ephestia)属,或者选自双翅目(diptera)、鞘翅目(coleoptera)或膜翅目(hymenoptera),最优选选自雏蝗(choristoneura)属。然后,该组合物还可以包含针对目标物种的生物防治剂,例如细菌生物防治剂,例如苏云金芽孢杆菌或枯草芽孢杆菌,或者真菌生物防治剂,例如昆虫病原真菌,例如球孢白僵菌(beauveria bassiana)、玫烟色伊萨菌(isaria fumosorosea)、蜡蚧菌属(lecanicillium spp.)、或者绿僵菌属(metarhizium spp.)。生物防治剂可以是细菌生物防治剂或真菌生物防治剂的活性生物质,优选苏云金芽孢杆菌生物质。所述活性生物质,例如苏云金芽孢杆菌细胞,也可以具有以可复制形式掺入的来源物种dna,从而起到宿主物种细胞的作用。
[0075]
该组合物可以通过任何合适的方式施用于目标物种,例如表面接触、嗜细胞施用、全身施用(通过例如注射、摄入或吸入或吸附)。该组合物可以配制成用于干燥或液体处理的形式,所述形式选自分散体,例如气溶胶、悬浮液、可湿性或可溶性粉末、水或其它溶剂中的乳液、可分散颗粒、微胶囊悬浮液、可乳化浓缩物、流体糊、大乳液、油分散体、饵。可以使用包含水的溶剂系统或深共晶溶剂(des)系统,例如天然深共晶溶剂(nades)系统。
[0076]
本发明组合物的dna对目标物种具有抑制作用的浓度范围的确定在本领域技术人员的知识范围内。本领域技术人员将理解,所需浓度可能取决于多种因素,例如组合物中抑制目标物种的dna的效力、所需的抑制水平、是否施用额外的杀生物剂和/或对目标物种的施用途径。对于许多应用,合适的浓度可以在1
‑
1500ppm的范围内,例如2
‑
1300ppm、2
‑
1000ppm、5
‑
1000ppm、10
‑
1000ppm、50
‑
1000ppm、100
‑
1000ppm、200
‑
1000ppm、500
‑
1000ppm。对于其他应用,可能需要更高的浓度。
[0077]
从wo2014/020624中可以清楚地看出,抑制性dna片段可以用于医学。其中,wo2014/020624显示了对黑曲霉(aspergillus niger)的抑制,所述黑曲霉可能引起感染,如耳霉菌病。类似地,包含dna片段或其前体的本发明组合物也可用于抑制黑曲霉或其他寄生和/或致病生物(如线虫寄生虫)的药物中。在组合物的医学用途中,来源物种dna是抑制剂或其前体,因为当它处于未片段化的状态时,它可以原位转化,例如在胃肠道中转化为片段形式。因此,本发明的另一方面是本发明的组合物用于药物,或者换言之用作药物。本发明上下文中的医学用途涉及人类医学和/或兽医学,优选人类医学。在本发明的上下文中,提及动物包括提及人,除非与使用它的上下文有不同的含义。该组合物可以例如在药物中用作抗病原组合物,其能够抑制病原生物的扩散和/或生长,或者甚至用于杀死病原生物。可以用本发明的组合物抑制的病原靶生物是例如局部病原体、肠道病原体、肺部病原体或全身病原体。病原体可以是例如微生物病原体(例如选自细菌、真菌或病毒)、寄生虫(例如选自原生生物或选自蠕虫,例如线虫、绦虫、吸虫或蛔虫)。本发明组合物的医学用途特别是针对上文在本发明组合物的讨论中提出的病原和/或寄生目标物种,特别是与皮肤、指甲或粘膜相关的局部病原体和/或寄生虫。为了用作药物,该组合物是药学上可接受的形式。
[0078]
本发明的另一方面涉及本发明的组合物作为抑制目标物种的产品的用途。目标物种可以对应于来源物种,或者是在系统发育上与来源物种相似的物种。在产品中,本发明的组合物可以用作抑制目标物种的活性成分。目标物种的预期抑制可以在产品中,例如为了提高产品的生物稳定性,或者替代地,目标物种的预期抑制可以在目标表面上或者在产品将要应用的目标区域中。该产品可以包含其它成分,包括对寄生、致病和侵染物种有抑制作用的其它成分,例如选自杀生物剂、除草剂、杀真菌剂、杀虫剂、杀螨剂、杀线虫剂、抗寄生虫剂、杀藻剂、杀细菌剂中的一种或多种。该产品可以是医疗产品,也可以是非医疗产品。根据某些实施方案,所述产品是非医疗产品,例如农业产品、个人健康护理产品或卫生产品,例如肥皂或洗涤剂。
[0079]
在使用该组合物的某些优选实施方案中,来源物种dna序列可以在嵌合dna分子中在宿主物种细胞中。对于一些应用来说,在宿主物种细胞中存在嵌合的dna分子就足够了,因为它们可以通过宿主物种细胞所处的条件从宿主物种细胞中释放出来。例如,当产品是用于抑制目标物种的产品时,宿主物种细胞是目标物种合适的食物来源,当它们被目标物种食用时,包含来源物种dna序列的嵌合dna分子从目标物种肠内的宿主物种细胞中释放出
来。在这个过程中,来源物种的dna序列也可以从嵌合的dna分子中释放出来。同样在宿主物种细胞的自然衰退过程中,宿主物种细胞中包含的嵌合dna分子可以为宿主物种细胞释放,并且来源物种dna序列可以通过片段化从嵌合dna分子中释放。在本发明的另一种用途中,宿主细胞通过细胞破碎技术破碎,例如片段化dna的细胞破碎技术,来自破碎的宿主细胞的物质用于抑制目标物种。为此,可以任选地用已知方法从破碎的宿主细胞中分离出dna。或者,破碎的细胞团可以在不进一步分离dna的情况下用于抑制目标物种。
[0080]
本发明的另一方面涉及生产本发明组合物的方法。该方法包括以下步骤:
[0081]
‑
提供宿主物种的数量;
[0082]
‑
提供来自来源物种的dna源,其任选包含片段化的来源物种dna;
[0083]
‑
使宿主物种细胞和来自来源物种的dna源处于允许宿主物种细胞以可复制的形式吸收来源物种dna的条件下;
[0084]
‑
繁殖含有可复制形式的来源物种dna的宿主物种细胞;
[0085]
‑
任选地,分离含有可复制形式的来源物种dna的宿主物种细胞;
[0086]
‑
任选地,从宿主物种细胞中分离dna,特别是含有来源物种dna的dna;
[0087]
‑
任选地,片段化宿主物种细胞的dna,特别是含有来源物种dna的dna。
[0088]
在该方法的第一步中,提供宿主物种的数量。基于关于本发明的组合物、本发明的用途和可获得的公知常识所讨论的内容,技术人员将知道并理解如何提供合适的宿主物种细胞。
[0089]
在第二步中,提供来自来源物种的dna源,任选包含片段化的dna。来自来源物种的dna源可以是任何合适的来源。例如,来源物种的细胞可以这样使用。或者,可以使用从来源物种细胞释放的dna,例如从通过细胞破碎技术(例如片段化dna的细胞破碎技术)破碎的细胞中释放的dna。释放的dna可能被分离,也可能不被分离。分离的dna可能是也可能不是片段化的。来自来源物种的片段化的dna可以原样存在,或者可以掺入人工dna构建体中,例如在人工质粒、粘粒、人工染色体(如yac或bac)或真菌人工染色体中,或者在与转座元件结合的构建的嵌合dna分子中。
[0090]
在第三步中,宿主物种细胞和来自来源物种的dna源经受允许宿主物种细胞以可复制的形式吸收来源物种dna的条件。通过以可复制的形式吸收来源物种的dna,宿主物种细胞将利用其dna复制机制复制来源物种的dna。因此,条件必须是这样的,即所提供的来源物种dna被掺入宿主物种细胞中,使得它在功能上可用于待复制的宿主物种细胞的dna复制机制。技术人员将能够选择合适的条件,以允许宿主物种细胞以可复制的形式吸收来源物种的dna。
[0091]
例如,当提供的来源物种的dna掺入人工dna构建体中,如质粒、粘粒、人工染色体或与转座元件结合的构建的嵌合dna分子中时,技术人员将知道如何调节宿主物种细胞,使它们能够有效地吸收这些人工dna构建体,从而可以有效地复制它们。
[0092]
如果来源物种的dna是作为来源物种的细胞提供的。本领域技术人员将会知道,所需的条件必须允许来源物种细胞释放来源物种dna和将来源物种dna片段化。已经发现,在凋落物分解过程中,这些条件使得来自凋落物中植物材料的dna可能被转移到土壤微生物组中。基于这些发现,本领域技术人员将预期,来自包含在凋落物或类似的分解生物材料中的非植物来源的dna也可以结合到土壤微生物组中。因此,合适的条件可以是凋落物分解的
条件,特别是当宿主物种是土壤微生物时,例如选自α变形菌属、β变形菌属、δ变形菌属、放线菌属、嗜热菌属、红色杆菌属、氯酸杆菌属、酸菌属、或solibacteres属。
[0093]
在来源物种的dna是作为从来源物种细胞释放的dna提供的情况下,本领域技术人员将知道,释放的来源dna可以由宿主物种的合适细胞以天然过程吸收或通过强制方法如生物弹道(或基因枪法)吸收。
[0094]
第四步是任选的,包括分离宿主物种细胞的步骤。从包括实施例在内的整个说明书中,本领域技术人员将理解,并非所有应用都需要分离宿主细胞。当需要或期望分离宿主物种细胞时,技术人员将了解用于分离宿主物种细胞的合适技术。
[0095]
第五步也是任选的,包括从宿主细胞中分离dna的步骤。从包括实施例在内的整个说明书中,本领域技术人员将理解,并非所有应用都需要从宿主物种细胞中分离dna。当需要或期望从宿主物种细胞中分离dna时,技术人员将了解从宿主物种细胞中分离dna的合适技术。
[0096]
第六步是另一任选步骤,包括将宿主物种细胞的dna片段化的步骤。从包括实施例在内的整个说明书中,本领域技术人员将理解,并非所有应用都需要来自宿主物种细胞的dna片段化。当需要或期望宿主物种的dna片段化时,本领域技术人员将了解从宿主物种细胞中片段化dna的合适技术。在来自宿主物种细胞的dna被片段化的情况下,优选嵌合dna分子被片段化。优选使用随机片段化技术进行dna片段化。
[0097]
现在将通过以下非限制性实验进一步举例说明本发明。
[0098]
实验
[0099]
实验1
[0100]
拟南芥文库
[0101]
拟南芥的商业微生物基因组文库购自在酵母和大肠杆菌中工程化的tair(https://www.arabidopsis.org/)。
[0102]
yac文库cd4
‑
21p是pyac4载体中的混合酵母人工染色体(yac)基因组文库;从哥伦比亚植物细胞核中分离的dna;原始文库由1,152个克隆组成;平均插入大小=420kb;21%的克隆携带叶绿体dna;文库由81个混合池组成;克隆酶:ecori和ecori。
[0103]
poca18
‑
hyg载体中的bac文库cd4
‑
21p二元粘粒基因组文库;由用taqi部分消化形成的wassilewskija(ws)基因组dna片段产生;代表十个基因组等价物;载体在camv 35s启动子的控制下携带潮霉素抗性基因;克隆酶:clai和clai。
[0104]
每个文库用于获得如下制备的微生物溶液:
[0105]
yac文库cd4
‑
21p:
[0106]
将80份的每一份接种在含有ypd培养基(1%酵母提取物,2%细菌内毒素,1%葡萄糖)的试管中,并在28℃孵育48小时。孵育后,将试管合并,细胞悬浮液离心(4000rpm,5分钟),并重新悬浮在10ml 0.9%nacl(合并的酵母悬浮液)中。然后,在含有1升ypd培养基的台式搅拌反应器(new brunswick)中,以28℃、200rpm、1vvm通气速率进行混合的yac培养。向反应器中接种足量的等分混合酵母悬浮液,得到初始o.d.
590
=0.1。48小时后,收集酵母生物质,洗涤并重新悬浮在蒸馏水中,以达到每毫升50mg细胞干重的最终浓度。
[0107]
对于用作对照的酵母细胞,遵循相同的程序,但是使用ypd培养基中的酿酒酵母cen.pk113
‑
7d菌株的预培养物作为反应器的接种物。此外,在这种情况下,收集产生的生物
质,洗涤并重新悬浮在蒸馏水中,达到与上述相同的细胞密度。
[0108]
bac文库cd4
‑
21p:
[0109]
将80份的每一份接种在含有50mg ml
‑1潮霉素的胰蛋白胨大豆培养基(tsb)的试管中,并在37℃孵育48小时。孵育后,将试管合并,细胞悬浮液离心(10,000rpm,10分钟),并重新悬浮在10ml 0.9%nacl(合并的细菌悬浮液)中。然后,细菌文库的培养在含有1l包含潮霉素的tsb培养基的台式搅拌反应器(new brunswick)中进行,温度为37℃,转速为200rpm,通气速率为1vvm。向反应器中接种足量的等分混合细菌悬液,得到初始o.d.
590
=0.01。48小时后,收集细菌生物量,洗涤并重新悬浮在蒸馏水中,以获得每毫升50mg细胞干重的细胞密度。
[0110]
对于用作对照的细菌细胞,遵循相同的程序,但是使用在tsb的大肠杆菌atcc 10536的预培养物作为反应器的接种物。此外,在这种情况下,收集产生的生物质,洗涤并重新悬浮在蒸馏水中,达到与上述相同的细胞密度。
[0111]
抑制试验
[0112]
将所述两个文库的微生物溶液超声处理,所得细胞匀浆在室温有氧条件下孵育15天,通过加入蒸馏水补偿水分蒸发。用蒸馏水以1:1、1:10和1:100稀释匀浆的不同等分试样,并用于湿润培养皿中的滤纸,每个浓度水平用10株植物进行三次重复。拟南芥幼苗通过在培养皿中用滤纸上的标准营养液发芽种子来制备。选择至少一周大的健康幼苗,大小相似,用于抑制试验。通过将幼苗暴露于培养皿中的不同溶液中来进行实验(每个培养皿10粒种子,每个处理10个培养皿)。对照通过仅用蒸馏水在实验期间保持的幼苗来评估。处理包括暴露于文库或相应的没有基因组文库的微生物溶液中。10天后通过计数活植物的数量来评估幼苗的存活。实验在保持最佳温度和空气湿度的生长室中进行。
[0113]
结果
[0114]
抑制试验的结果通过测试拟南芥基因组的微生物文库(bac和yac)证明了异源和自身dna的混合物对同种幼苗存活的抑制作用。事实上,在暴露于基因组自身dna的yac和bac文库下,幼苗存活率显著降低(图1),比较对象是用蒸馏水处理的对照和包括异源dna但不包括所述自身dna基因组文库的相应微生物溶液。这种模式在稀释处理的三个水平中是一致的。此外,含有异源dna但不含所述自身dna的处理通常不影响幼苗存活,除了酿酒酵母材料,其与对照没有区别,其在最高浓度水平下显示出非特异性抑制作用。
[0115]
实验2
[0116]
香附子(cyperus aesculentus)文库
[0117]
bio s&t公司(加拿大蒙特利尔)利用载体psmart
‑
hindiii(氯霉素抗性)在大肠杆菌epi300中构建了香附子的基因组bac文库。将文库汇集在1
×
96孔板中,每个孔包含123个初级克隆,总共11,808个克隆(>2x基因组覆盖度)。平均插入片段大小(110kb)使用测试的随机克隆来确定,用noti消化。文库储存在
‑
80℃。
[0118]
所述文库用于获得如下制备的微生物溶液:
[0119]
为了扩增,每个孔从冷冻份接种在含有12.5mg/l氯霉素的胰蛋白胨大豆培养基(tsb)的试管中,并在37℃孵育48小时。孵育后,将试管合并,细胞悬浮液离心(10,000rpm,10分钟),并重新悬浮在10ml 0.9%nacl(合并的细菌悬浮液)中。然后,细菌文库的培养在含有1l包含氯霉素的tsb培养基的台式搅拌反应器(new brunswick)中进行,温度为37℃,
转速为200rpm,通气速率为1vvm。向反应器中接种足量的等分混合细菌悬液,得到初始o.d.
590
=0.01。48小时后,收集细菌生物量,洗涤并重新悬浮在蒸馏水中,以获得每毫升50mg细胞干重的细胞密度。
[0120]
对于用作对照的细菌细胞,遵循相同的程序,但是使用在tsb的大肠杆菌epi300的预培养物作为反应器的接种物。此外,在这种情况下,收集产生的生物质,洗涤并重新悬浮在蒸馏水中,达到与上述相同的细胞密度。
[0121]
抑制试验
[0122]
将微生物溶液超声处理,所得细胞匀浆在室温有氧条件下孵育15天,通过加入蒸馏水补偿水分蒸发。然后,用两种不同的用于植物生长的基质(石英砂和标准盆栽土)分别以1:10和1:100v/v稀释不同等份的匀浆。用滤纸上的标准营养液在培养皿中发芽种子,制备香附子幼苗。选择年龄至少为10天的类似大小的健康幼苗用于抑制试验。通过将幼苗暴露于15ml体积的小塑料盆中的不同基质混合物中来进行实验(每盆1株幼苗,每个处理20盆)。处理包括基质类型(两个水平,砂或盆栽土壤)、大肠杆菌材料类型(两个水平,有或没有bac库)和溶液浓度(两个水平,1:10或1:100)的因子组合。含有未混合基质的盆被视为对照(每种基质类型10罐)。自幼苗盆栽10天后,通过对植物材料进行破坏性取样,随后进行烘干(60℃
×
48小时)并称重总生物量,评估香附子在受控条件下(温度为22/16℃,光照时间为16:8小时,环境空气湿度,每天浇水至盆栽田间容量)的生长。
[0123]
结果
[0124]
结果表明,香附子的bac基因组文库对同种幼苗生长具有抑制作用。事实上,在暴露于基因组自身dna的bac文库下,幼苗生物量显著较低(图2),比较对象是用蒸馏水处理的对照和包括异源dna但不包括所述自身dna基因组文库的相应微生物材料。抑制作用是剂量依赖性的,在两个治疗浓度水平上是一致的。含有异源dna但不含所述自身dna的处理不影响幼苗生长,这与对照没有区别。
[0125]
实验3
[0126]
秀丽隐杆线虫(caenorhabtidis elegans)文库
[0127]
秀丽隐杆线虫的fosmid文库(商业化的)购自source bioscience(http://www.sourcebioscience.com/)。该文库包括41x 384平板,共包含15,744个克隆。大片段插入(平均大小43.3kb)文库利用fosmid拷贝控制载体pcc1fos(氯霉素抗性)在大肠杆菌epi300
‑
t1r中构建。
[0128]
本实验用到两种大肠杆菌菌株:op50和epi300
‑
t1r。大肠杆菌菌株在lb生长培养基中生长。在必要时,向生长培养基中加入终浓度为12.5μg/ml的氯霉素和最终浓度为0.01%重量体积比的阿拉伯糖。所用秀丽隐杆线虫是野生型bristol(n2)株。该秀丽隐杆线虫株在20℃在ngm琼脂平板(不含阿拉伯糖)上培养,并以平板上生长的大肠杆菌op50作为食物来源。使用基于pcc1fos的文库转化的大肠杆菌epi300
‑
t1r和epi300
‑
t1r作为指定的ngm平板(含阿拉伯糖)上的食物来源。
[0129]
为了扩增fosmid,将每个包含384个文库克隆的微孔板从冻存管接种到含氯霉素的lb琼脂平板(150mm培养皿)。将接种的41个平板于37℃孵育过夜。孵育后,将细菌收集在含有氯霉素的12ml的lb培养基中,用涂布棒涂布在每个平板上。将所得细菌混合物转移到15ml培养管(41管)中。取100μl接种在含有氯霉素和阿拉伯糖的4ml lb培养基中,并于37℃
孵育过夜(41管)。第二天,将100μl细菌培养物转移到2ml含有氯霉素和阿拉伯糖的lb培养基中,并于37℃孵育过夜(41管)。
[0130]
抑制试验
[0131]
将所得的41种细菌培养物混合,并将20μl这些混合的细菌用于每个含有阿拉伯糖(35mm)的ngm平板上。未转化的大肠杆菌epi300
‑
t1r用作阴性对照。在这种情况下,细菌以fosmids扩增所遵循的相同程序生长,但省略了氯霉素。为了筛选第一代,将单个l4期的秀丽隐杆线虫转移到每个含有阿拉伯糖的ngm平板上,并接种细菌,并保持在20℃。每12小时将该线虫转移到一个新鲜的相同平板上,以允许所有的卵沉积(3天)。
[0132]
监测以下参数:胚胎致死率(不能存活的卵与已产的卵的比率)和异常表型(异常表型与孵化的卵的比率)。同样的参数也被监测用于筛选第二代,对于第二代,来自第一代的单一的、无缺陷的l4期秀丽隐杆线虫被转移到每个含有阿拉伯糖的ngm平板上,并接种细菌,并如上用于筛选第一代所述进行处理。
[0133]
还测试了部分秀丽隐杆线虫基因组的抑制作用。为此,用秀丽隐杆线虫文库的克隆a01(wrm0610aa01)(染色体i;克隆开始:13249863
‑
克隆结束:13287543[大小37680])、秀丽隐杆线虫文库的克隆b01(wrm0610ca02)(x染色体;克隆开始:12671842
‑
克隆结束:12705840[大小33998])以及克隆a01和克隆b01的组合来饲养线虫。在第一代后代中测试胚胎致死率(不能存活的卵与已产的卵的比率)和异常表型(异常表型与已孵化的卵的比率)。
[0134]
结果
[0135]
在实验中,在第一代筛选中分析了以含有整个秀丽隐杆线虫文库的epi300
‑
t1r为食的11种线虫和以未转化的epi300
‑
t1r为食的12种线虫(分别为2574个和3055个卵)。饲喂含有整个秀丽隐杆线虫文库的epi300
‑
t1r的线虫显示出胚胎致死率的增加,与饲喂epi300
‑
t1r的线虫显著不同(分别为3.85%和0.82%,p<0.0001)。由于两个后代中都没有雄性,所以不同处理间雄性与孵化的卵的比例没有差异。与食用epi300
‑
t1r的线虫相比,食用含有整个秀丽隐杆线虫fosmid文库的epi300
‑
t1r的线虫的后代中观察到显著更高频率(6.32%)的发育缺陷(p<0.0001,图3)。
[0136]
值得注意的是,与第一代相比,整个秀丽隐杆线虫fosmid文库的负面影响在第二代中加剧(图3)。
[0137]
部分基因组的抑制结果列于下表2。
[0138][0139]
(1)饲养5条线虫,对1378个卵进行分析;(2)饲养5条线虫,对1299个卵进行分析;(3)饲养12条线虫,对2726个卵进行分析。%值与对照组相比有显著差异。
[0140]
结果表明,基因组的一小部分对胚胎致死率和异常表型的出现都有显著影响。组合克隆的效果增强(高于另外的克隆)。可以预期,通过与其他克隆组合,抑制作用可以逐渐增强。
[0141]
实验4
[0142]
海灰翅夜蛾(spodoptera littoralis)文库
[0143]
在大肠杆菌dh10b中,通过从实验室生长的海灰翅夜蛾幼虫中提取和消化(限制性酶hind iii)高分子量基因组dna,在hind iii位点使用载体copycontrol pcc1bach(bamh i hind iii,ecor i cloning ready vector
–
8128bp)构建了海灰翅夜蛾(棉叶波纹夜蛾)的基因组bac文库,然后冷冻。克隆在含有luria bertani(lb) 氯霉素固体基质的平板上生长并选择。筛选并挑选克隆,表达诱导至高拷贝数,然后转移到96深孔板中的孔中。每个孔包含290个初级克隆,总共27,840个克隆(>10倍基因组覆盖度)。平均插入片段大小(132kb)使用测试的随机克隆来确定,用noti消化。还制备了空载体对照,并在与文库载体相同的条件下生长。将文库和空载体储存在
‑
80℃。
[0144]
该文库用于如下生产质粒:将单个无菌牙签浸入并润湿每个单孔培养物中,然后转移并用于接种到含有1ml氯霉素lb(12.5μg/ml)的2ml微量离心管中。一旦接种,微离心培养物在轨道搅拌下于37℃孵育过夜。将所有96个起始培养物(96ml)混合在一起,以获得代表整个文库的起始接种物。制备10升含有氯霉素的lb,分置于玻璃锥形瓶中并灭菌。将每个基质受体用起始接种物接种,在37℃轨道搅拌下孵育过夜。
[0145]
为了从文库和空载体对照中提取质粒,按照制造商的说明,使用了qiagen质粒giga试剂盒。
[0146]
获得的文库的dna浓度为688ng/μl,在约800μl体积(dna总产量=550μg),并且空载体的浓度为268ng/μl。
[0147]
抑制试验
[0148]
在15厘米培养皿中,将三龄期的海灰翅夜蛾幼虫保持在受控条件下(温度22
‑
24℃,光周期16:8小时),并如前所述进行饲养(bergomaz和boppr
é
,1986)。处理包括如mazzoleni et al.(2015b)所述通过提取和超声处理从鲑鱼(大西洋鲑鱼(salmo salar))和海灰翅夜蛾获得的dna片段的溶液,以及如上所述通过从bac文库提取获得的自身和异源dna的混合物。
[0149]
三种处理溶液中的每一种都以200ng/μl的dna浓度稀释,每种溶液4ml包含在幼虫的日常饮食中。自生长实验开始以来,每个处理的20个重复(培养皿,每个有30个幼虫)在受控条件下保持20天。对照重复(n=10)也保持在相同的条件下,并且用标准的、未处理的饮食喂养。在20天生长期结束时评估所有处理的幼虫存活率。
[0150]
结果
[0151]
实验4中的测试结果表明,无论是在饲料颗粒中存在还是不存在异源dna(即用作bac文库载体的微生物菌株的dna)的情况下,通过摄入暴露于自身dna均会显著影响海灰翅夜蛾幼虫的存活。事实上,在这两种情况下,20天后观察到5%和0%的存活率(图4)。相反,在喂食仅含有异源dna(即鲑鱼dna)的颗粒的情况下,与未处理的对照相比,没有观察到显著的存活率降低,这表明单独的异源dna不会对幼虫产生抑制作用(图4)。
[0152]
实验5
[0153]
在一个证实性实验中,为了进一步证实上述实验的发现,即当片段化的自身dna与异源dna(例如用于dna扩增的宿主生物的dna)结合时,对物种的个体具有抑制作用,测试了在酵母中扩增获得的自身dna对黑腹果蝇(drosophila melanogaster)的抑制作用,酵母是黑腹果蝇的合适食物来源。
[0154]
自身dna扩增
[0155]
用标准饲养方法获得黑腹果蝇培养物。含有黑腹果蝇dna片段的酵母细胞在s&t bio(加拿大蒙特利尔)进行了工程改造。为此,使用sau3ai部分消化从黑腹果蝇培养物的个体中分离的总dna。使用bamhi克隆位点将消化的dna片段克隆到pgadt7载体中。用含有克隆的黑腹果蝇片段的pgadt7载体转化大肠杆菌dh10b宿主细胞,所得文库通过过夜培养在琼脂培养基上扩增。将约120,000个原始克隆置于半固体培养基上的大型培养皿(总共10个培养皿)上。然后洗涤菌落并收集到10个试管中。做了大量的准备工作来分离文库的dna。
[0156]
从大肠杆菌文库中分离dna然后用于转化酿酒酵母y187宿主细胞。通过过夜培养在琼脂培养基上扩增文库,并将每个大型培养皿(总共10个培养皿)约120,000个原始克隆铺在半固体培养基上。然后洗涤菌落并收集到10个试管中。通过凝胶色谱法对10个随机选择的克隆进行插入片段大小测定,显示平均大小为4k,范围为1
‑
6kb。
[0157]
抑制试验
[0158]
所述酵母文库被用作果蝇(sang(1978))从孵化到蛹阶段的标准饲料的组成部分。处理由如下组成:自身dna处理:用酵母文库饲养;对照1:饲喂无载体的相同酵母菌株作为对照;对照2:饲喂有空载体的相同的酵母菌株。结果列于下表3。
[0159]
表3.酵母文库中片段化的自身dna对黑腹果蝇早期生命阶段的抑制作用
[0160][0161]
结果表明,含有果蝇dna的酵母文库对幼虫有明显的抑制作用。在幼虫生长期间,观察到发育时间(对照时间的两倍)和死亡率(三龄阶段40%和蛹83%)显著延长。在使用标准饮食的对照1和使用空文库载体的酵母的对照2之间没有观察到差异,因此证明抑制(根据生命阶段持续时间和死亡率的水平确定)是由于酵母文库中的果蝇dna插入导致的。
[0162]
实验6
[0163]
地中海松(pinus halepensis)和黑杨(populus nigra)的抑制
[0164]
基于在宿主生物中复制的人工dna构建体中掺入的自身dna片段(一部分)对目标物种的有效抑制,可以预期掺入宿主物种天然基因组中的自身dna同样将有效抑制目标物种。已知横向基因转移广泛存在于自然界,包括土壤微生物群。
[0165]
为了证实引入宿主物种天然基因组的自身dna对目标物种的抑制作用,在该土壤微生物实验中,用两种不同的凋落物材料(地中海松和黑杨的叶子)接种了来自橡树林(所述两种物种都不存在)土壤的土壤微生物。如mazzoleni et al.2015所述,分解的凋落物被用于抑制试验。显示出同种抑制的分解的凋落物材料被用于微生物组dna的宏基因组分析,以证实植物dna在土壤微生物组dna中的整合。
[0166]
具体而言,从田间收集地中海松和黑杨的鲜叶,并以下列方式用于分解实验。通过
在随机选择的植物(n>20)下放置网,在自然群落中收集刚脱落的叶子,在通风室中室温下干燥,直到达到恒重,然后在室温下储存。分解实验在具有最佳水分利用率和温度条件的生长室中进行:每隔7天用蒸馏水浇灌凋落物至保持能力,温度为夜间18
±
2℃,白天24
±
2℃。干落叶(每种100克,重复三次)放入塑料托盘(尺寸30
×
50
×
50cm)中。根据bonanomi等人(2011),制备并分发了从所述两个物种都不存在的橡树林收集的微生物接种物。接种120天后收集凋落物,并在纸袋中干燥(在40℃下直至达到恒重)。
[0167]
为了证实分解的凋落物材料的物种特异性抑制效果,进行了如下生物测定试验:将8厘米宽的无菌滤纸条置于方形培养皿(尺寸为12
×
12
×
1.5cm)中。将每个物种的预发芽种子(每个盘子五粒)放在之前用不同的分解的凋落物材料修改过的纸条的顶部。将根观察板置于45
°
并覆盖不透明的薄片。28天后评估根长。
[0168]
为了从分解的材料中分离微生物群,将1g凋落物样品放入5ml试管中。向每个样品中加入3ml磷酸盐缓冲盐水(pbs)。通过涡旋60秒使样品均质化,随后以10000rpm离心1分钟,以从凋落物碎片中分离微生物细胞。离心后,将分离的微生物细胞的上清液转移到新的2ml试管中。在合并阶段后,用ctab法从微生物细胞中提取dna。在提取步骤结束时,样品用聚乙烯吡咯烷酮(pvp)处理以除去酚,以避免对pcr的可能抑制。提取的dna用乙醇进一步纯化,然后在微生物层流室中干燥,并在无核酸酶pcr级水中洗脱。
[0169]
通过标准的鸟枪法测序技术对分解的凋落物进行宏基因组学分析。然后根据下面描述的生物信息学程序,分析宏基因组学的成对末端读段,以找到属于微生物物种和来自分解凋落物的植物物种的dna的重组片段。
[0170]
为了找到属于上述植物和相应微生物组物种的可能的重组dna片段,遵循以下程序。使用bbmap的bbmerged函数(bushnell 2016)合并宏基因组学实验的成对末端读段,应用kmers选项以降低假阳性(extend2=20,iterations=5)。未合并的成对末端使用bbmap的fuse.sh函数进行融合(bushnell 2016),在正向和反向读段之间插入0n。然后使用blastn将合并和融合的读段与宿主参考基因组或相关物种(如果基因组不可用)进行比对(camacho等人,2009)。使用了以下参考基因组:火炬松(pinus taeda)(1760464骨架~22gb)和毛果杨(populus trichocarpa)(1694骨架,~500mb)。所有与宿主基因组至少有一次匹配的读段都被认为是匹配的。使用blastn(camacho等人,2009)对照ncbi的非冗余数据库,对宿主基因组进行独立比对,以确认与参考物种的匹配。
[0171]
结果
[0172]
生物测定实验证实了分解的凋落物对地中海松和黑杨的物种特异性抑制作用。地中海松的发芽种子在自己的凋落物上的根生长比在黑杨凋落物上的生长少50%。与地中海松相比,黑杨根在自己的分解的凋落物上的生长减少了70%。
[0173]
对分解的凋落物进行宏基因组学数据分析清楚地表明,植物物种和微生物之间存在几种重组dna片段。
[0174]
从44,431,532个合并读段开始,108,288个读段匹配火炬松基因组骨架。然后将这108,288个读段与ncbi核苷酸数据库(nt)进行比较,100%的读段在nt数据库中存在至少一个匹配,具有41,533个唯一id。在所述唯一id中,我们随机选择了10,000个id的子集来获得详细的分类单元注释。从匹配序列的总数(108,288)来看,对29,149个序列的子集确认了注释,其中4,595个序列被确认与nt中的火炬松序列匹配。在这4,595个序列中,1,359个具有
低于80%的覆盖度,这意味着该序列可以被认为是松树和微生物物种之间的重组序列。为了证实松树凋落物中的微生物对松树基因组摄取的特异性,还检查了与毛果杨(p.trichocarpa)的匹配,仅发现139个匹配。
[0175]
在黑杨凋落物中也发现了类似的结果。注释结果总结在表4和表5中。
[0176]
通过鸟枪法进行的分析清楚地证实了宏基因组学数据中与分解植物凋落物的参考基因组部分相似的片段的富集。换句话说,结果显示了来自凋落物基质的dna片段的特异性内含物,从而产生了由显示抑制作用的植物标记的微生物组(microbiome)。
[0177]
表4.地中海松凋落物宏基因组学分析结果。
[0178][0179]
表5黑杨凋落物宏基因组学分析结果。
[0180][0181]
为了证实这些显著的发现,重复实验并扩展分析。叶子的收集、凋落物的产生和材料的处理如上所述。凋落物在最佳条件下孵育和分解120天,30天后和实验结束时收集样品。根据mazzoleni et al.(2015)的描述,使用标准商业试剂盒对30天和120天的新鲜凋落物和分解材料进行dna提取,并对提取的dna进行分析。我们将通过鸟枪法测序方法从宿主植物物种(松属和杨属)的凋落物(分解0、30、120天)中获得的读段与指定的参考基因组进行了比较。
[0182]
根据以下程序进行生物信息学分析:
[0183]
使用bbmap的bbmerged函数(bushnell(2016))合并宏基因组学实验的成对端读段,应用kmers选项来减少假阳性(extend2=20,iterations=5)。
[0184]
然后使用blastn将合并的读段与宿主基因组进行比对(camacho等人(2009))。所有至少有一个命中的读段,其中lister作为具有匹配的总读段,并且它们根据其覆盖范围被分组列出。可能的重组片段是那些匹配20
‑
80%或90%覆盖度的片段。还考虑了以90%同一性进行过滤。关于生物信息学程序的更多细节可在bushnell(2016年)和camacho等人
(2009年)中找到。
[0185]
结果如表6所示。他们表明,样品中识别的植物全dna片段的频率随着时间的推移而降低,证明随着分解的进行,凋落物中游离植物dna的数量减少。另一方面,随着分解的进行,与植物基因组匹配的部分片段(意味着它们是由部分植物dna和部分其他物质的dna组成的片段)的数量增加(表7),在120天后达到峰值。需要注意的是,分解120天的凋落物对同一物种(即松属和杨属)种子的萌发抑制作用最大。
[0186]
根据每个样品中测序匹配的数量(表6)和基因组部分每个读段的相对覆盖度(表7)分析数据。部分覆盖的读段被假定为反映读取样本中的嵌合体。
[0187]
表6在比对过的读段中匹配的频率
[0188][0189]
表6显示,分别考虑来自爆炸搜索分析与松属和杨属基因组的匹配频率,在0天凋落物中比在120天凋落物中产生更高的数字。这意味着由于序列衰减,与宿主植物的相似性降低。
[0190]
表7.每个覆盖读段的相对频率
[0191]
[0192]
表7显示了与宿主植物相比,读段的高覆盖度的相对降低(覆盖度值下降99%至100%)和随着凋落物老化,部分覆盖序列的增加(20%至90%覆盖度)。
[0193]
对宿主基因组的相似性也进行了90%同一性的研究,以排除与宿主植物的可能不可靠的相似性。覆盖度降低表明在从凋落物中测序的读段中存在嵌合体。
[0194]
一个非常有趣的观点是,一方面,与植物基因组匹配的数量显示杨属的快速分解凋落物的减少非常高,而松属凋落物的分解较慢对应于松树序列的更慢的消失。另一方面,两种物种材料都显示出混合片段数量的不断增加,其中仍然可以识别出物种,反映了天然掺入到微生物有机体中。
[0195]
这些结果支持天然文库的形成,类似于本技术中进一步描述的微藻的单一物种培养(见下文)。在这种情况下,天然文库形成现象发生在天然土壤微生物组中,该微生物组富含其周围植物凋落物的dna,从而对产生凋落物的植物产生物种特异性抑制作用。
[0196]
实验7
[0197]
钝顶节旋藻(arthrospira platensis)天然文库
[0198]
为了进一步证实宿主物种天然基因组中掺入的自身dna对目标物种的抑制作用,还通过将属于蓝细菌的钝顶节旋藻与从拟南芥中提取的dna一起孵育来诱导对dna的天然摄取,并且使用转化的钝顶节旋藻细胞来抑制拟南芥。
[0199]
在培养前,蓝细菌的纯培养物在zarrouk生长培养基上保持在受控条件下(温度22
‑
24℃,光周期12:12小时,辐照度20μmol光子m
‑2s
‑1),在孵育前至少4小时不吹气。
[0200]
用于孵育的拟南芥dna是通过从新鲜叶子中提取全基因组的标准方法制备的,并通过超声破碎法随机破碎(mazzoleni et al.(2015a)报道了方案的细节)。样品在vc 505sonics超声波处理器中进行超声波处理,微尖浸入1.5ml微管中,用冷水水浴30分钟,以20%的振幅进行45秒/15秒的开/关循环。
[0201]
钝顶节旋藻与拟南芥dna的孵育
[0202]
在培养之前,将钝顶节旋藻培养在缺乏mg
的生长培养基中,以降低蓝细菌的细胞内和细胞外dna酶活性,并促进外源dna的持久性和掺入(cao等人,1999)
[0203]
在培养过程中,将300μl浓缩的钝顶节旋藻细胞(通过温和离心获得最多5
×
107个细胞)加入到含有300mg玻璃珠(直径500μ)的15ml离心管中。然后,在培养管中加入100μl先前通过超声破碎片段化的拟南芥dna和100μl的mg
缺乏的培养基,随后在涡流混合器中以最高速度搅拌15s。
[0204]
在同一试管中用zarrouk的mg
缺乏培养基将样品体积增加到2.5ml,并在振荡器中在ld光照循环下培养24小时。然后在光生物反应器中用完全zarrouk培养基将体积逐渐增加到10升,达到每升1克细胞干重的细胞密度。90天后,将生物质离心并收集,得到细胞密度为每升100克细胞干重的材料沉淀。
[0205]
抑制试验
[0206]
将蓝细菌沉淀超声处理,所得细胞匀浆在室温下温育15天,通过加入蒸馏水补偿水分蒸发。然后,用蒸馏水以1:1、1:10和1:100稀释不同等分的匀浆,并用于在培养皿中润湿滤纸,每个浓度水平用10株植物进行三次重复。制备拟南芥幼苗,并进行与实验1相同的实验。
[0207]
结果
[0208]
观察到与拟南芥dna孵育后钝顶节旋藻培养物的颜色变化,间接表明拟南芥dna掺入到钝顶节旋藻细胞中。事实上,这种观察到的颜色变化表明了基因表达的变化,这不太可能是由于拟南芥dna掺入而导致的钝顶节旋藻基因组变化的结果。
[0209]
抑制试验的结果通过测试拟南芥基因组的蓝细菌天然文库证明了异源和自身dna的混合物对同种幼苗存活的抑制作用。在高浓度下,与不含拟南芥dna(即含有异源dna但不含植物dna)的微藻溶液的对照相比,暴露于吸收拟南芥dna的钝顶节旋藻天然文库下的幼苗存活率显著降低(表8)。
[0210]
虽然最可能的是只有一部分拟南芥dna被整合到钝顶节旋藻基因组中(并因此被钝顶节旋藻细胞复制),但对拟南芥幼苗有相当大的抑制作用。鉴于对秀丽隐杆线虫基因组的一部分的抑制的测试结果(见实验3),可以预期通过将抑制性转化的钝顶节旋藻培养物与通过类似转化程序获得的进一步培养物结合,抑制效果可以增加。
[0211]
表8.转化的钝顶节旋藻对拟南芥的抑制作用
[0212][0213]
死亡率%(培养皿上移植的10株幼苗/处理)。
[0214]
实验8
[0215]
进行验证性实验,以进一步证实宿主物种天然基因组中掺入的自身dna的抑制作用。在该验证性实验中,两种不同物种的蓝细菌(光合微藻)在生物反应器中生长,向生长培养基中加入片段化的拟南芥dna,并测试蓝细菌被破坏的生物量的抑制。
[0216]
选择的蓝细菌是钝顶节旋藻(如实施例7)和集胞藻(synechocystis sp.)pcc6803。在孵育前,所有培养物保持在表1所示的培养条件下,在孵育前至少4小时不吹气。
[0217]
表9孵育的物种和相应的培养条件(stein)
[0218][0219]
为收获的细胞分离的dna通过超声破碎随机片段化。样品在vc505sonics超声波处理器中进行超声波处理,微尖浸入1.5ml微管中,用冷水水浴30分钟,以20%的振幅进行45秒/15秒的开/关循环。
[0220]
钝顶节旋藻与拟南芥dna孵育
[0221]
在培养之前,将钝顶节旋藻培养在缺乏mg
的生长培养基中,以降低蓝细菌的细胞内和细胞外dna酶活性,并促进外源dna的持久性和掺入(cao等人,1999)
[0222]
如dunahay等人(1997)所述,在培养过程中,将300ul浓缩的钝顶节旋藻细胞(通过温和离心获得高达5
×
107个细胞)加入到含有300mg玻璃珠(500μm直径)的15ml离心管中。然后,在孵育管中加入100ul先前通过超声破碎片段化的拟南芥dna(约10ug)和100ul的mg
缺乏的培养基,并使用涡流混合器以最高速度搅拌该管15秒。
[0223]
在同一试管中用zarrouk的mg
缺乏培养基将样品体积增加到2.5ml,并在振荡器中在ld光照循环下培养24小时。然后用完全zarrouk培养基逐渐增加体积。
[0224]
用拟南芥dna孵育集胞藻pcc6803
[0225]
在培养之前,细胞已经在用edta(2mm)修饰的bg11培养基中培养两天,以降低钙离子依赖性dna酶活性,如zang等人(2007)所述。
[0226]
在孵育期间,将400ul的浓缩的集胞藻pcc6803细胞(通过温和离心获得高达5*107个细胞,并用无edta的bg11培养基洗涤)加入到15ml离心管中,该离心管含有100ul的先前通过超声破碎片段化的拟南芥dna(约10ug)。然后将试管在振荡器中在ld光照下孵育5小时,然后转移到烧瓶中,用bg11培养基稀释至2.5ml,然后逐渐增加体积。
[0227]
实验
[0228]
根据上述方案暴露于拟南芥dna的微藻在两个5升生物反应器中用作接种物,并在标准生长条件下生长,直到达到最大细胞密度。然后,从培养基中提取两种藻类的生物量以形成沉淀,仔细洗涤,然后干燥。
[0229]
将藻类沉淀接种到1g的森林土壤,并每周保持在室温和潮湿条件下,以保持自然分解长至60天。每隔15天对腐烂的材料取样,并在实验室最佳条件下添加到在培养皿中的滤纸上,以评估平板上拟南芥植物的萌发和根生长。
[0230]
结果
[0231]
未分解的沉淀对拟南芥的种子萌发和根伸长没有显著的抑制作用,而在分解30和60天后,在用来自两个物种的分解沉淀的处理中观察到对萌发的显著抑制。分解90天后,分解沉淀的抑制效果略有下降。这表明蓝藻沉淀的分解对植物的萌发具有明显的抑制作用,因为包含在藻类基因组中的植物的dna被分解过程释放,并最终被分解的土壤微生物进一步扩增。
[0232]
表10
–
通过暴露于两种钝顶节旋藻(左)和集胞藻pcc6803(右)的(分解的)生物质对拟南芥进行抑制试验的结果。
[0233][0234]
生物信息学分析
[0235]
对在上述种子萌发实验中用作天然文库的两种蓝细菌的基因组进行了测序和分析。使用bbmap的bbmerged函数(bushnell(2016))合并宏基因组学实验的成对端读段,应用kmers选项来减少假阳性(extend2=20,iterations=5)。
[0236]
然后使用blastn将合并的读段与宿主基因组进行比对(camacho等人(2009))。所有至少有一个命中的读段,其中lister列为具有匹配的总读段,并且它们根据其覆盖范围被分组列出。可能的重组片段是那些匹配20
‑
80%或90%覆盖度的片段。还考虑了以90%同一性进行过滤。关于生物信息学程序的细节可在bushnell(2016年)和camacho等人(2009年)中找到。
[0237]
表11.生物信息学分析的结果
[0238][0239]
表11显示,在来自蓝细菌样品的dna中发现的与拟南芥基因组匹配的序列计数不仅在样品之间有很大差异,而且在根据植物和蓝细菌基因组的覆盖度定义的不同匹配类别
之间也有很大差异。
[0240]
类别1和类别2显示了非常低的计数,这可能与测序和生物信息学方法固有的不确定性有关。类别3显示了最高的样本间差异,可能与两种蓝细菌之间基因组大小、染色体拷贝数和参考基因组数据库中用于比对的数据可用性的差异有关。类别4数据表明,在两种样品类别中,可能存在与植物和蓝细菌基因组部分匹配的嵌合序列。
[0241]
类别5显示空值,表明两种样品类别都不包括嵌合序列之外的游离拟南芥dna。
[0242]
值得注意的是,与钝顶节旋藻样品相比,集胞藻pcc 6803样品显示出对拟南芥幼苗更大的抑制作用,也显示出与拟南芥基因组匹配的更高数量的dna序列,无论是考虑所有混合类别的序列计数,还是单独考虑每个有意义的类别。这证实了抑制作用与自身dna片段有关,并且当与宿主物种dna结合时,自身dna片段的抑制作用得以维持。在该实验的基础上,进一步证实了宿主物种的细胞可以在其天然基因组中有效地复制系统发育远缘的目标物种的抑制性自身dna。
[0243]
实验9
[0244]
通过测试被土壤微生物分解和吸收的片段化自身dna对杂草马齿苋(portulaca oleracea)的抑制作用,进行了进一步的验证性实验。该试验是在一个农场的菠菜种植试验地进行的,在那里马齿苋侵染是一个反复出现的问题。
[0245]
马齿苋的bac文库首先由bio s&t公司(加拿大蒙特利尔)用与实验2中讨论的香附子文库相似的方法生成。将文库的过夜培养物(terrific broth(tb)作为培养基,250rpm搅拌,温度=37℃)用作60升的生物反应器(有效体积为40升)发酵的起始培养物(生物反应器有效体积的1%)。
[0246]
使用营养肉汤(nb)作为大规模液体发酵的基质。
[0247]
此外,在培养基的总体积中加入0.1%的左旋阿拉伯糖来诱导psmart
‑
hindlll质粒。液体发酵在温度、搅拌、通气、ph的受控的条件下进行2天,以获得具有高细胞密度并刺激质粒dna产生的发酵液。根据od600估算,达到的浓度约为5.6
×
108个细胞/ml。
[0248]
堆肥的接种和成熟
[0249]
将如前所述获得的发酵液煮沸约15分钟,随后加入三种不同浓度的泥炭中:
[0250]
1.30ml/kg(1)
[0251]
2.3ml/kg(稀释1:10)
[0252]
3.0.3ml/kg(稀释1:100)
[0253]
接种后的土壤在空气中成熟约20天。如此获得的产品用于田间试验。
[0254]
田间试验
[0255]
田间试验是在salerno(it)地区的一个农场进行的,该农场经常出现马齿苋侵染的问题。
[0256]
菠菜播种前两天(7月中旬),产品按照以下方案进行分配:
[0257]
田间实验(尺寸地块约2x2.4m)
[0258]
cnt1cnt1:10cnt1:1001:100cnt1:10cnt1cnt
[0259]
(cnt=对照(未添加煮沸发酵液的泥炭))
[0260]
在为实验测试准备的隧道附近的隧道中,对抗马齿苋杂草的斗争通过常规农艺措
施和使用根据当前立法允许的除草剂来进行,构成进一步的对照,以便在测试中评估产品的功效。
[0261]
特别是,农民通常采用的斗争方式规定使用威百亩,这是一种农业通用杀虫剂,主要用作广谱种植前土壤熏蒸剂,以防治杂草、杂草种子、真菌、线虫和土壤昆虫。用威百亩处理是昂贵的,并且对土壤有害,导致根际生物多样性的损失。
[0262]
在菠菜发芽时(播种后约10天),进行了第一次调查,从这里开始,每周进行其他评估,直到栽培菠菜的生命周期结束。为了确定该产品对马齿苋发芽率的抑制作用,在田间评估期间,根除马齿苋植物,计数并检查根部是否存在坏死。
[0263]
结果
[0264]
如图5所示,如调查结束时所确定的,区块处理1中马齿苋植物的数量显著减少。此外,如所预料的,在马齿苋植物较少的地区,发芽菠菜的数量显著增加。
[0265]
该实验进一步证实了抑制作用与自身dna片段有关,并且当自身dna片段在宿主物种中扩增并与宿主物种dna结合时,自身dna片段的这些抑制作用得以维持。考虑到所使用的实验方案和上述实验的结果,可以合理地预期,泥炭微生物组对马齿苋dna片段的扩增一定有助于抑制作用。
[0266]
实验10
[0267]
系统发育相似物种的dna片段的抑制作用
[0268]
根据wo2014/020624(与宿主物种dna序列无关),用片段化dna测试了来自系统发育相似物种的dna片段的抑制作用。从其他实验的结果来看,可以预期,在对根据wo2014/020624的dna片段观察到抑制作用的情况下,在根据本发明的组合物中也将观察到dna片段化时的作用。
[0269]
来自植物材料的dna的提取和定量
[0270]
使用制造商(qiagen,valencia,ca)描述的dneasy plant maxi试剂盒从植物叶片材料中提取dna,以使用fulton等人(1995)的方案制备处理溶液(附件e)。收获100克新鲜重量的起始材料,并在
‑
20℃下冷冻。然后,将样品混合,浸入200ml新鲜微量制备的缓冲液中,然后在奶酪纱布中过滤。将dna重新悬浮在纯无菌水中。从叶子中提取的dna通过超声片段化为200
‑
500bp的目标大小范围。这是由bandelin sonopulse(bandelin,berlin,de)以90%的功率和0.9秒的脉冲持续12分钟完成的。safe(invitrogen)在3%metaphor
tm
琼脂糖凝胶(lonza scientific,nj)上对超声处理的条带大小进行了目测验证。
[0271]
用纯化的dna对目标植物进行生物测定
[0272]
从植物叶片中提取的纯化dna对八种目标物种的自毒性和植物毒性效应进行了评估(图7)。实验设计如下:1.无菌蒸馏水作为对照;2.添加总提取的和随机片段化的自身dna;3.添加来自单一异源物种的总提取的和随机片段化的dna(表1);以200μg ml
‑1的浓度应用dna。生物测定在体外进行,通过将每个目标物种(4个重复)的表面无菌种子(每个板中n=20)放置在9cm培养皿中的吸收4ml测试溶液的无菌滤纸上。根据目标物种,在实验开始后可变天数后测量幼苗根长(图7)。根据完全随机的设计,培养皿被安排在生长室中(温度22
±
2℃,每两天用蒸馏水浇灌一次)。
[0273]
[0274][0275]
对于每个生物测定,分离目标物种和用作dna提取来源的植物物种的系统发育距离以下列方式评估:
[0276]
‑
指定零距离
[0277]
‑
在距离=1处属于同一科的物种
[0278]
‑
在距离=2处属于同一目不同科的物种
[0279]
‑
在距离=2 n处的不同目的物种,其中n等于参考系统发育树(apg 2009,图6)中分离植物目的树节点数,该系统发育树报告了被子植物(即开花植物)分类目的系统发育。
[0280]
生物测定中使用的所有植物的种、科和目,以及所有目标和dna来源植物对的相应
系统发育距离报告于表12中。
[0281]
生物测定结果(即目标物种的根生长)表示为用蒸馏水处理的对照的百分比,并绘制为分离dna来源和目标物种的系统发育距离的函数。
[0282]
结果
[0283]
生物测定结果显示,目标物种自身dna对其根系生长有明显的抑制作用(图8)。对于来自与目标物种系统发育相似(在同一分类目内)的物种的dna,当测试时也观察到相当大的抑制作用,尽管不如来自目标物种本身的dna的作用大。来自远离目标物种的物种的异源dna的影响不显著。
[0284]
参考文献
[0285]
a simple insect diet for rearing arctiidae and other moths.journal of the lepidopterists’society,40,131
–
137.
[0286]
bonanomi g,incerti g,barile e,capodilupo m,antignani v,mingo a,lanzotti v,scala f,mazzoleni s.2011.phytotoxicity,not nitrogen immobilization,explains plant litter inhibitory effects:evidence from solid
‑
state 13c nmr spectroscopy.new phytologist,191:1018
‑
1030.
[0287]
bushnell b,2016.bbmap short read aligner.university of california,berkeley,california.url http://sourceforge.net/projects/bbmap.
[0288]
camacho c,coulouris g,avagyan v,ma n,papadopoulos j,bealer k,madden tl.2009.blast :architecture and applications.bmc bioinformatics,10(1),421.
[0289]
cao,j.,xu,z.,qiu,g.and li,b.,1999.effects of mg2 on the growth and dnase activity of spirulina platensis,a cyanobacterium.bioresource technology,67(3),pp.287
‑
290.
[0290]
dunahay,terri g.,sally a.adler,and jonathan w.jarvik."transformation of microalgae using silicon carbide whiskers."recombinant gene expression protocols.humana press,1997.503
‑
509.
[0291]
mazzoleni,s.,bonanomi,g.,incerti,g.,chiusano,m.l.,termolino,p.,mingo,a.,senatore,m.,giannino,f.,carten
ì
,f.,rietkerk,m.and lanzotti,v.,2015a.inhibitory and toxic effects of extracellular self
‑
dnain litter:a mechanism for negative plant
–
soil feedbacks?new phytologist,205(3),pp.1195
‑
1210.
[0292]
mazzoleni,s.,carten
ì
,f.,bonanomi,g.,senatore,m.,termolino,p.,giannino,f.,incerti,g.,rietkerk,m.,lanzotti,v.and chiusano,m.l.,2015b.inhibitory effects of extracellular self
‑
dna:a general biological process?new phytologist,206(1),pp.127
‑
132.
[0293]
smit af,hubley r,green p.1996.repeatmasker.url:http://www.repeatmasker.org.
[0294]
sang jh(1978)the nutritional requirements of drosophila.the genetics and biology of drosophila,vol.2(ed.by m.ashburner&trf wright),pp.159
–
192.academic press,new york,ny,usa.
[0295]
stein,j.(ed.)handbook of phycological methods.culture methods and growth measurements.cambridge university press.448 pp.
[0296]
zang,x.,liu,b.,liu,s.,arunakumara,k.k.i.u.,&zhang,x.(2007).optimum conditions for transformation of synechocystis sp.pcc 6803.the journal of microbiology,45(3),241
‑
245.
[0297]
angiosperm phylogeny group(agp),2009.an update of the angiosperm phylogeny group classification for the orders and families of flowering plants.botanical journal of the linnean society 161(2):105
–
121,doi:10.1111/j.1095
‑
8339.2009.00996.x.
[0298]
fulton tm,chunwongse j,tanksley sd.1995.microprep protocol for extraction of dna from tomato and other herbaceous plants.plant molecular biology reporter 13:207
‑
209.
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。