使用细小病毒治疗携带tp53基因突变和/或p53蛋白翻译后修饰的癌症
技术领域
1.本发明包括在肿瘤学领域内。特别地,本发明涉及属于细小病毒(parvoviridae)科,优选属于原细小病毒(protoparvovirus)属的病毒或此类病毒与化疗的组合在治疗癌症中的用途。特别地,癌症的特征在于呈现tp53基因突变和/或p53蛋白翻译后修饰。
背景技术:
2.p53蛋白和癌症
3.p53蛋白由tp53基因表达,通过非常不同的机制在控制细胞周期和基因组稳定性方面发挥重要作用,这就是为什么p53蛋白有时被称为“基因组守护者”。因此,tp53基因中的突变是导致人类多种不同类型癌症的主要机制之一也就不足为奇了。事实上,tp53是人类癌症中最常发生突变的基因,特别是,在没有有效治疗的破坏性疾病胶质母细胞瘤(glioblastoma,gbm)中经常发现基因tp53中的各种遗传改变(brennan,cw et al(2013)the somatic genomic landscape of glioblastoma,cell 155,462
‑
477)。tp53和其他基因中的遗传改变导致gbm肿瘤的生长受到功能冗余信号传导的控制,这使得它们响应于定向分子疗法而发生适应。因此应该强调的是,尽管tp53突变对于目前数百万人遭受的多种癌症的发生和进展具有巨大的重要性,但没有针对这些遗传病变的特异性有效治疗,因为现今tp53主要被认为是一种“非成药(non
‑
druggable)”基因(kastenhuber,er,ands.w.lowe.(2017),putting p53 in context,cell 170.1062
‑
1076)。
4.常规癌症化疗和p53
5.在目前的肿瘤门诊中,化疗方案经常使用(单独或组合)以下药物:
6.·
羟基脲(hydroxyurea,hu或oh
‑
u),其通过抑制核糖核苷酸还原酶来降低dntp(三磷酸核苷)的浓度,因此使dna的复制叉停止(吉西他滨或阿糖胞苷也是如此),从而引起可能包括p53在内的多种细胞反应。
7.·
顺铂(cisplatin,cispt),自40多年前就经常在临床中用于全身治疗,因为它对多种类型的实体癌具有活性[basu a,krishnamurthy s.2010.cellular responses to cisplatin induced dna damage.j nucleic acids doi:10.4061/2010/201367]。
[0008]
·5‑
氟尿嘧啶(5
‑
fluorouracil,5fu),其是一种类似于尿嘧啶的抗代谢物,通常用于对抗各种癌症,尽管对结直肠癌具有更好的功效。5
‑
氟尿嘧啶的作用机制复杂,包括异染色质和rna代谢的改变。
[0009]
溶瘤病毒
[0010]
多种病毒已在不同系统中表现出抗癌能力(溶瘤),作为天然毒株或经过基因改造而使用。在这些方面,至少应强调以下类型的具有一定溶瘤能力的病毒:细小病毒、麻疹病毒、呼肠孤病毒、腺病毒、疱疹病毒和痘病毒。然而,在现有技术中尚未发现选择性地作用于携带tp53突变和/或p53蛋白翻译后修饰的癌症的溶瘤病毒。换句话说,溶瘤病毒的能力到目前为止还没有与tp53基因突变和/或p53蛋白翻译后修饰具体联系起来。
[0011]
因此,本发明着眼于通过鉴定单独使用或与化疗联合使用的溶瘤病毒来解决上述技术问题,以特别靶向具有tp53基因突变和/或p53蛋白翻译后修饰的癌症。因此,本发明提供了有效的治疗窗,允许针对携带tp53基因遗传改变和/或p53蛋白翻译后修饰的肿瘤(例如在人原发性癌症中常见的那些)进行特异性和定制化的治疗。
技术实现要素:
[0012]
简介
[0013]
本发明涉及属于细小病毒科、特别属于原细小病毒属的病毒或这种病毒与化疗的组合在治疗癌症中的用途。值得注意的是,许多类型的癌症的特征为呈现tp53基因突变和/或p53蛋白翻译后修饰。此外,在一个优选的方面,本发明证明了,此类病毒在对癌细胞的毒性作用方面与常规化疗协同合作,前提是靶细胞携带tp53基因遗传改变和/或p53蛋白翻译后修饰,无论这些是组成型的或是由这些药物诱导的。
[0014]
因此,本发明打破了现有技术中的偏见,因为尽管每个病例所涉及的突变谱有待确定,但通常假设癌症患者中tp53突变的存在与化疗和放疗治疗的不良结果之间存在良好的相关性[tchelebi,l.,ashamalla,h.,and graves pr(2014)mutant p53 and the response to chemotherapy and radiation.in:deb s.,deb s.(eds)mutant p53 and mdm2 in cancer.subcellular biochemistry,vol 85.springer,dordrecht]。相反,本发明证明了,当本发明的细小病毒与基因毒性化疗药物联合使用时实现了协同作用,对特征为呈现tp53基因突变和/或p53蛋白翻译后修饰的各种类型癌症具有有效治疗。
[0015]
因此,本发明通过下述附图和实施例证明:
[0016]
·
细小病毒科的数种病毒株,特别是本发明中测试的原细小病毒属的数种病毒株,以毒性方式(通过表达病毒ns1蛋白)选择性感染来源非常不同的人癌细胞,包括来自胶质母细胞瘤患者的原代干细胞,前提是这些细胞携带tp53基因突变。
[0017]
·
p53蛋白的非生理性翻译后修饰,特别是残基丝氨酸15中的组成型或诱导型磷酸化,有利于本发明中测试的病毒株在包括具有胶质母细胞瘤的患者的原代干细胞在内的不同的人癌细胞中的毒性感染和复制。
[0018]
·
由原发性肿瘤中存在的或在干细胞和已建立的人胶质母细胞瘤细胞系中外源性表达的致瘤病毒蛋白诱导的p53蛋白的改变,确实促进了本发明中测试的细小病毒科病毒株的毒性感染和复制。
[0019]
·
当前临床癌症疗法的诱导了p53蛋白翻译后修饰(例如残基ser15的磷酸化)的化疗药物(如oh
‑
u、5fu或顺铂),与本发明中所测试的细小病毒科病毒株的毒性感染协同作用,非常显著地降低了这些癌细胞的活力。
[0020]
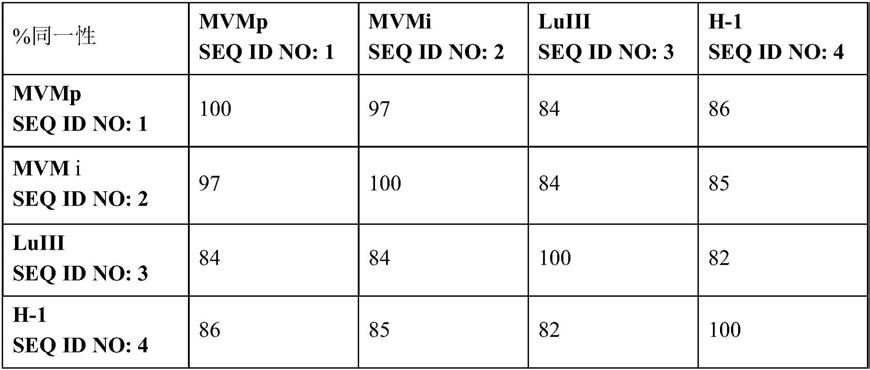
因此,本发明的第一方面涉及一种病毒颗粒,该病毒颗粒包含基本上由seq id no:1、seq id no:2、seq id no:3或seq id no:4组成的核苷酸序列,该病毒颗粒单独使用或与基因毒性化疗药物联合用于治疗癌症,其中癌症的特征为呈现tp53基因突变和/或p53蛋白翻译后修饰。此外,与序列seq id no:1、seq id no:2、seq id no:3或seq id no:4具有至少95%、至少96%、至少97%、至少98%或至少99%序列同一性的病毒颗粒被包括在本发明中。
[0021]
重要的是要强调包括在本发明中的属于细小病毒科的原细小病毒属的四种病毒
序列之间的高同源性百分比,如表1所示。值得注意的是,核苷酸的差异主要集中在基因组的非编码区,所以如果我们关注编码区,同源性百分比会更高。
[0022]
表1
[0023][0024]
本发明的第二方面涉及一种包含病毒颗粒的药物组合物,该病毒颗粒又包含基本上由seq id no:1、seq id no:2、seq id no:3或seq id no:4组成的核苷酸序列并且与基因毒性化疗药物联合。
[0025]
在本发明的优选方面,p53蛋白翻译后修饰是组成型的、由诱导adn损伤或基因毒性应激的化疗药物诱导的[kirkland,d.et al.updated recommended lists of genotoxic and non
‑
genotoxic chemicals for assessment of the performance of new or improved genotoxicity tests.mutation research 795(2016)7
‑
30][zhu,y.et al.cisplatin causes cell death via tab1 regulation of p53/mdm2/mdmx circuitry.(2013).genes and develop 27:1739
‑
1751]或由致瘤病毒诱导的。在本发明的优选方面,p53蛋白翻译后修饰在于ser15残基的磷酸化。在本发明的优选方面,tp53基因突变选自包含以下的组:r273h、p72r、e258k、g245s和/或v173l。在本发明的优选方面,tp53基因突变发现于编码的p53蛋白的dna结合结构域(dna
‑
binding domain,dbd)。在本发明的优选方面,基因毒性化疗药物选自包含以下的组:顺铂、羟基脲、5
‑
氟尿嘧啶、吉西他滨或阿糖胞苷。
[0026]
在本发明的优选方面,癌症是神经胶质瘤、肺癌、食道癌、肝癌、胰腺癌、膀胱癌、结直肠癌、前列腺癌、胶质母细胞瘤、神经胶质瘤、头颈癌、乳腺癌、胃癌、卵巢癌、子宫癌或黑色素瘤。在本发明的优选方面,所述病毒颗粒与基因毒性化疗药物联合使用,其中该病毒颗粒与基因毒性化疗药物同时施用、在基因毒性化疗药物之后或之前施用。
[0027]
本发明的第三方面涉及用于确定tp53基因突变和/或p53蛋白翻译后修饰的体外方法,该方法包括使用病毒颗粒,该病毒颗粒包含基本上由seq id no:1、seq id no:2、seq id no:3或seq id no:4组成的核苷酸序列。
[0028]
本发明的第四方面涉及一种用于体外诊断癌症或用于筛选癌症患者的方法,该方法包括通过使用病毒颗粒来确定tp53基因突变和/或p53蛋白翻译后修饰,该病毒颗粒包含基本上由seq id no:1、seq id no:2、seq id no:3或seq id no:4组成的核苷酸序列。
[0029]
本发明的第五方面涉及用于治疗癌症的tp53基因突变和/或p53蛋白翻译后修饰,其中tp53的突变选自包含以下的组:r273h、p72r、e258k、g245s或v173l,以及p53蛋白的翻
no:2、seq id no:3或seq id no:4组成的核苷酸序列,其中该癌症的特征为:呈现至少一种突变和/或p53蛋白的磷酸化,所述至少一种突变选自由以下组成的组:tp53基因中的r273h突变、tp53基因中的p72r突变、tp53基因中的e258k突变、tp53基因中的g245s突变、tp53基因中的v173l突变。
[0041]
在优选的实施方式中,p53蛋白的磷酸化是组成型的,或者是先前已被基因毒性化疗药物或致瘤病毒诱导的。
[0042]
在优选的实施方式中,p53蛋白的磷酸化由ser15残基的磷酸化组成。
[0043]
在优选的实施方式中,突变位于tp53基因的dbd区或p53蛋白的prd区。
[0044]
在优选的实施方式中,诱导p53蛋白磷酸化的基因毒性化疗药物选自包含以下的组:顺铂、羟基脲、5
‑
氟尿嘧啶、吉西他滨或阿糖胞苷。
[0045]
在优选的实施方式中,癌症选自包含以下的组:tp53基因中呈现r273h、p72r、e258k、g245s和/或v173l突变的神经胶质瘤、胶质母细胞瘤、急性髓性白血病、肺腺癌、膀胱癌或直肠腺癌。
[0046]
在优选的实施方式中,包含基本上由seq id no:1、seq id no:2、seq id no:3或seq id no:4组成的核苷酸序列的病毒颗粒与一种基因毒性化疗药物联合使用,其中该病毒颗粒在化疗药物之后施用。
[0047]
在优选的实施方式中,癌症的特征为在tp53基因中呈现r273h突变,该癌症选自包含以下的组:肺癌、食管癌、肝癌、胰腺癌、膀胱癌、结直肠癌、前列腺癌、胶质母细胞瘤、神经胶质瘤、头颈癌、乳腺癌、胃癌、卵巢癌、子宫癌或黑色素瘤。
[0048]
在优选的实施方式中,基因毒性化疗药物选自包含以下的组:顺铂、羟基脲、5
‑
氟尿嘧啶、吉西他滨或阿糖胞苷。
[0049]
在优选的实施方式中,本发明涉及一种用于治疗癌症的包含病毒颗粒的药物组合物,该病毒颗粒又包含基本上由seq id no:1、seq id no:2、seq id no:3或seq id no:4组成的核苷酸序列并且与基因毒性化疗药物联合,其中所述病毒颗粒在化疗药物之后施用。
[0050]
在优选的实施方式中,本发明涉及一种用于治疗一类癌症的方法,该癌症的特征为呈现至少一种突变和/或p53蛋白的磷酸化,所述至少一种突变选自包含以下的组或选自由以下组成的组:tp53基因中的r273h突变、tp53基因中的p72r突变、tp53基因中的e258k突变、tp53基因中的g245s突变、tp53基因中的v173l突变。该方法包括施用包含病毒颗粒的药物组合物,该病毒颗粒又包含基本上由seq id no:1、seq id no:2、seq id no:3或seq id no:4组成的核苷酸序列,在优选的实施方式中,该病毒颗粒与基因毒性化疗药物联合。
[0051]
为了本发明的目的,提供以下定义:
[0052]
·
术语“包括”意指其包括,但不限于跟随在词语“包括”之后的内容。因此,术语“包括”的使用表示所列元素是强制性的,但其他元素是可选的,并且可以存在或可以不存在。
[0053]
·“由...组成”意指包括且限于跟随在短语“由...组成”之后的内容。因此,短语“由......组成”表示所列元素是强制性的,并且不可以存在其他元素。
[0054]
·“药学上可接受的赋形剂”是指任选地可被包含在本发明的组合物中的赋形剂,并且它不会对患者造成显著的不利毒理学作用。
[0055]
本发明组合物的“治疗有效剂量或治疗有效量”应理解为施用时在患有癌症的对
象中引起阳性治疗反应的剂量。所需的确切量因对象而异,取决于对象的年龄、一般状况、所治疗疾病的严重程度、施用方式以及其他相关因素。合适的“有效”量可由本领域技术人员根据现有技术中提供的信息使用常规实验来确定。
附图说明
[0056]
图1.细小病毒,小鼠微小病毒(parvovirus minute virus of mice,mvm)在人胶质母细胞瘤干细胞(glioblastoma stem,gs或hgs)中诱导p53。用mvmp(原型株)和mvmi(免疫抑制株)病毒感染获得自胶质母细胞瘤患者的干细胞,并分析p53蛋白表达。左图:通过免疫荧光(immunofluorescence,if)在共聚焦显微镜下分析#5、#7和#8患者的gs细胞中病毒蛋白ns1和细胞p53的表达,这些gs细胞作为神经球生长。空白(mock):未感染细胞。右图:通过蛋白质印迹(western
‑
blot)在四名患者的gs中对相同蛋白质的诱导进行分析。巢蛋白(nestin):上样对照。分子质量(kda)显示在右侧;
[0057]
图2.容许细小病毒mvm的细胞毒性感染和复制的gs细胞中p53的翻译后修饰和tp53的遗传改变。a.通过if在共聚焦显微镜下分析来自四名患者(#5、#7、#8、#9)的胶质母细胞瘤干细胞(gs)中的ns1表达、ser15中p53磷酸化的诱导以及mvmp基因组复制之间的相关性。b.在两名患者的gs中,对mvmp基因组复制水平与ser15中p53磷酸化之间的相关性进行细胞计数分析。c.对mvmp感染的a9小鼠成纤维细胞进行细胞计数,并分析以下水平之间的相关性:ns1表达和病毒dna复制(上),s15中p53修饰和病毒dna复制(中),以及ns1表达和p53诱导(下)。d.图示通过大规模测序(massive sequencing,ngs)检测到的在先前感染的两名患者的gs中的tp53中的一些突变,并通过ns1( )/p53
‑
s15( )的表达进行绘制。e.通过ngs检测到的三名感染患者的gs中tp53突变的出现频率,通过ns1( )/p53
‑
s15( )的表达进行绘制。f.表示通过ngs检测到的四名感染mvmp患者的gs细胞中p53蛋白结构域中的四种突变,并通过ns1( )/p53
‑
s15( )的表达绘制。还示出了v173l(以v170l存在于a9小鼠成纤维细胞)和r273h(存在于u373mg和u251mg人胶质母细胞瘤细胞系)的位置。示出了联盟tcga(癌症基因组图谱(the cancer genome atlas),美国国立卫生研究院(nih usa);https://portal.gdc.cancer.gov)和cosmic(英国桑格研究所(sanger institute of the uk);https://cancer.sangerac.uk/cosmic)报告的人gbm样品中的tp53结构域(tad:反式激活结构域(transactivation domain);prd:富含脯氨酸的结构域;dbd:dna结合结构域;od:寡聚化结构域(oligomerization domain))中所有描述的突变的累积绝对病例的类型和数量;
[0058]
图3.被细小病毒mvm毒株感染的已建立的人癌症细胞系和gs细胞系中的p53翻译后修饰和tp53的遗传改变。a.感染了细小病毒mvm的两个毒株(i、p)的并对p53、在ser15处磷酸化的p53(pp53)和细小病毒dna(vdna)进行染色的已建立的人和灵长类动物细胞系的共聚焦if图。b.不同的未感染mvm毒株(空白)或感染mvm毒株的细胞中的p53
‑
s15频率和表达水平的共聚焦if估计。c、d.gs(c)和不同的已建立的小鼠(a9)和人胶质母细胞瘤系(d)中ns1、p53和p53
‑
s15诱导的蛋白质印迹分析。e.通过基于ns1 和p53
‑
s15 的表达分选a9和gs细胞进行纯化。使用指定的群体对tp53进行遗传分析。显示了所选窗中标志物阳性的细胞百分比。f.通过针对不同细胞系中蛋白质的两个区域的特异性抗体对p53识别的蛋白质印迹分析。注意到,针对约20个结构域的特异性抗体缺乏对u87mg细胞系中p53的识别。
g.wttp53阻止病毒ns1蛋白的表达。用表达wtp53的质粒或载体dna转染的a9和nb324k细胞中p53和ns1表达的共聚焦if分析。h、i.色谱图示出了已建立的细胞和指定的合并群体(h)或gs(i)中p53区域的桑格测序。j.表示在指定类型的人类癌症中,本研究中检测的tp53突变与tcga(癌症基因组图谱,美国国立卫生研究院;https://portal.gdc.cancergov)和cosmic(英国桑格研究所;https://cancersangerac.uk/cosmic)联盟在tp53结构域(tad:反式激活结构域;prd:富含脯氨酸的结构域;dbd:dna结合结构域;od:寡聚化结构域)中描述的所有突变相关;该组合示出了不同类型癌症中的大多数突变是如何在tp53基因的dbd结构域中发生的,尽管在某种类型的癌症中特定突变的频率可能非常高;
[0059]
图4.转染了改变p53的腺病毒致瘤基因的小鼠和人细胞系以及患者的gs的细小病毒mvmp和mvmi感染。a.转染了表达致瘤基因的质粒的nih3t3细胞或(b)gs8细胞的共聚焦if分析显示出mvmp基因组的复制增加。c.gs8细胞中的类似研究显示,通过southern印迹分析的mvm基因组的主要复制中间体(main replicative intermediate,mrf)的积累水平。d.用表达致瘤基因的质粒进行转染对gs8细胞中mvmp基因组的复制的影响或(e)对u87mg中mvmi基因组的复制的影响,这通过if共聚焦(左)或细胞计数(右)来测量;
[0060]
图5.化疗药物(cd)对细小病毒mvmp感染u373mg胶质母细胞瘤细胞的影响。a.通过细胞计数和共聚焦if(b)分析三种cd对ns1和p53
‑
s15表达水平的影响。c.顺铂在ns1、p53
‑
s15和病毒基因组复制的水平上的剂量影响的if共聚焦分析。d.在类似条件下进行分析,通过sds印迹测量ns1和p53信号传导蛋白的水平,或(e)通过集落形成测量细胞活力;
[0061]
图6.cd对细小病毒mvmi感染u87mg胶质母细胞瘤细胞的影响。a、b.5fu和羟基脲(hu)对未感染的u87细胞中p53
‑
s15水平的剂量影响的if共聚焦分析。c.通过southern印迹分析的这些cd对病毒基因组的主要复制中间体(mrf)水平的剂量影响。d.这些cd对ns1、p53
‑
s15和病毒基因组复制的水平的影响的if共聚焦分析。印迹中测量的5fu对ns1和p53信号传导蛋白的水平的剂量影响(e),以及通过集落形成确定5fu对细胞活力的剂量影响(f)。通过印迹测量的hu对ns1和p53信号传导蛋白的水平的剂量影响(g),以及通过集落形成确定的hu对细胞活力的剂量影响(h);
[0062]
图7.cd对细小病毒mvmp感染u87mg细胞的影响。a.顺铂对ns1、p53
‑
s15和细小病毒基因组复制的水平的剂量影响的if共聚焦分析。b.通过测量ns1水平和数种p53信号传导细胞蛋白对等效样品进行蛋白质印迹分析。c、d和e.使用mvmp以不同感染复数(pfu/细胞)感染并联合指定剂量的不同cd的细胞存活率(通过集落形成测量)。
具体实施方式
[0063]
实施例1.材料和方法
[0064]
实施例1.1.细胞和细胞培养。
[0065]
人胶质母细胞瘤干细胞获得自马德里ram
ó
n y caial医院神经外科服务提供的肿瘤外植体。外植体被同一家医院的病理学服务进行的组织化学诊断为iv级胶质母细胞瘤。所有情况都获得了患者的知情同意,并获得了马德里ram
ó
n y caial医院机构伦理委员会(institutional ethics committee of the ram
ó
n y caial hospital)的批准。组织培养的进一步研究分别得到了马德里大学伦理委员会和severo ochoa分子生物学中心(centro de biolog
í
a molecular severo ochoa,csic
‑
uam)的授权。在手术室底部采集的活检经机
械和酶促分解,最后将细胞悬液过滤以在补充有各种因子的dmem培养基:f12(1∶1)中培养。
[0066]
另一方面,已建立的小鼠、人癌症和其他哺乳动物的细胞系在0.3%nahco3缓冲的杜氏改良伊格尔培养基(dulbecco
′
s modified eagle medium,dmem)和5%co2的气氛中培养。培养基中补充有抗生素(75u/ml链霉素、75μg/ml青霉素g)、2mm l
‑
谷氨酰胺和在56℃下灭活30分钟的5%胎牛血清(fetal bovine serum,fcs)。使用了以下细胞系:
[0067]
·
人胶质母细胞瘤:
[0068]
‑
u87
‑
mg:胶质母细胞瘤
‑
人星形细胞瘤(atcc:htb14)
[0069]
‑
u373
‑
mg:胶质母细胞瘤
‑
人星形细胞瘤(rrid:cvcl_2219)
[0070]
‑
u251
‑
mg:胶质母细胞瘤
‑
人星形细胞瘤(rrid:cvcl_0021)(ecacc09063001)
[0071]
·
其他人癌症:
[0072]
‑
hela:人宫颈癌(atcc
‑
ccl
‑
2)
[0073]
‑
nb324k:用大t抗原1多瘤病毒(polyomavirus)sv40转化的人新生儿肾成纤维细胞(rrid:cvcl_u409)。
[0074]
·
其他哺乳动物的致癌细胞系:
[0075]
‑
cos
‑
1:由多瘤病毒sv40转化的灵长类肝成纤维细胞(atcccrl
‑
1650)
[0076]
‑
a9:源自通过hgprt表型选择的小鼠l成纤维细胞的系。其被注射到小鼠中时显示出一定的致癌能力(atcc ccl
‑
1.4)。
[0077]
实施例1.2.流式细胞术。
[0078]
在所有实验中,在facs cantoii细胞仪(bd biosciences)中获得至少30000个事件/样品,这些事件在一个区域中基于其大小和复杂性(分别为fsc和ssc参数)进行预先选择,以排除细胞碎屑和其他污染颗粒。使用cellquest软件(bd biosciences)获取事件,并使用flowjo软件(tree star)来分析数据。细胞周期分析根据[gil
‑
ranedo,j.,hernando,e.,riolobos,l.,dominguez,c.,kann,m.,and jos
é
m.almendral.2015.the mammalian cell cycle regulates nuclear parvovirus capsid assembly.plospathogens,11;11(6):e1004920]。对于染色,将细胞在室温下用pbs 0.1%tritonx
‑
100透化10分钟,然后用补充有1%fcs的相同缓冲液封闭20分钟。将细胞重新悬浮在pbs(ph7.2) 0.5%bsa中,添加图中所示的一抗,然后在37℃下搅拌1小时。将细胞在pbs中洗涤,添加二抗并进行类似的孵育。在流式细胞仪中分析样品之前,在pbs中再进行两次洗涤。
[0079]
在通过流式细胞术(facs)纯化细胞以对tp53进行遗传分析时(如图3e所示),所有溶液均使用不含核酸酶的蒸馏水(gifco,不含dnase/rnase的蒸馏水10977
‑
035)用焦碳酸二乙酯(diethyl pyrocarbonate,dpc)来制备。将细胞透化并与上述抗体一起孵育,但pbs缓冲液补充有钒酸盐(rnase抑制剂)、明胶而非bsa(允许高压灭菌),并进行过滤。分选前,用在4℃预冷的高压灭菌的pbs
‑
dep彻底洗涤细胞仪设备(aria,bd),将避开(circumvented)的细胞收集在冰上的无菌管(f15,falcon)上,并立即加入trizol以抑制降解,然后进行rna纯化。
[0080]
实施例1.3.细胞活力的集落形成分析。
[0081]
基于[rubio,mp,guerra,s.,and almendral,jm 2001.genome replication and postencapsidation functions mapping to the nonstructural gene restrict the host range of a murine parvovirus in human cells.j virol 75(23):11573
‑
11582]
parvovirus capsidassembly.plospathogens,11;11(6):e1004920]。简而言之,培养基中存在的病毒通过在4℃下用3.4%聚乙二醇6000和0.5m nacl沉淀过夜来回收,然后用sorvallgsa斜角转子以5000rpm离心30分钟。为了回收细胞内病毒,将细胞沉淀在50mm tris
‑
hcl ph7.5,1mm edta(te)中重悬,并进行三个连续的冻融循环,之后添加0.2%sds并在8000rpm、4℃下用sorvall hb4摆动转子澄清10分钟。将从培养基中回收的病毒和细胞内病毒合并并通过在50mm tris
‑
hcl ph8.0,1mm edta,0.1m nacl和0.2%sds中的20%蔗糖垫(merck)用tst28.38转子以16000rpm离心18小时。将沉淀在含有0.2%sarkosyl(sigma)的te中重悬,并在cscl梯度(ni=1.371)中以50000rpm在15℃下用tft80.13转子运行24小时以达到平衡。通过注射器从梯度项部收集0.5ml级分,并通过与小鼠红细胞的血凝反应来确定空衣壳和装有dna的病毒的存在。最后,合并含有病毒的级分并用pbs透析。在
‑
70℃下等分储存纯化的病毒。
[0094]
实施例1.8.血凝反应。
[0095]
该方法用于估计病毒颗粒的数量和后续纯化。它是基于[s
á
nchez
‑
martinez,c.,grueso,e.,carroll,m.,rommelaere,j.and almendral,jm(2012)essential role of the unordered vp2 n
‑
terminal domain of the parvovirus mvm capsid in nuclear assembly and endosomal enlargement of the virionfivefold channel for cell entry.virology,432(1),pp.45
‑
56]和[hernando,e.,llamas
‑
saiz,al,foces
‑
foces,c.,mckenna,r.,portman,i.,agbandje
‑
mckenna,m.and almendral,jm(2000)biochemical and physical characterization of parvovirus minute virus of mice virus
‑
like particles,virology,267(2),pp.299
‑
309]。对于血凝(hemagglutination,ha)测定,将成年小鼠血液用磷酸盐盐水(phosphate saline,pbs)洗涤3次,通过在台式离心机中以1500rpm离心5分钟来收集红细胞。洗涤数次后,将红细胞沉淀物以50%(v/v)在pbs中重悬,并保持在4℃直至使用。ha在u型微量测试板(nunc)中进行。待评估的样品以在pbs中100μl的终体积应用,并在pbs中按1∶2进行系列稀释。最后,向每孔添加50pl 2%红细胞的pbs溶液,轻轻摇动板并避光保持在4℃下至少2小时。由保持血凝能力的最高稀释度的倒数中获得的滴度。
[0096]
实施例1.9.通过空斑形成进行细小病毒mvm滴定。
[0097]
基于[gil
‑
ranedo,j.,hernando,e.,valle,n.,riolobos,l.,maroto,b.andalmendral,jm(2018)differential phosphorylation and n
‑
terminal configuration of capsid subunits in parvovirus assembly and viral trafficking,virology,518,pp.184
‑
194]。将nb324k细胞以2.2
×
10e5个细胞/板的密度接种在p60板中,24小时后使用。除去培养基,在含有ca 和mg 的pbs(完全或pbsc)中洗涤细胞,并以每个p60 400μl添加病毒接种物,该病毒接种物是在补充有0.1%fcs的pbsc中稀释的。在37℃温和搅拌下吸附1小时后,除去接种物并添加7ml在37℃平衡的接种培养基(dmem,10%fcs和0.6%琼脂糖lm
‑
gqt pronadisa)。孵育6天后,将板在10%甲醛(panreac)的pbs溶液中固定,并用0.2%紫色晶体(panreac)在用pbs制备的10%甲醛中的溶液进行染色。裂解空斑数的计数乘以稀释度使得获得以每毫升空斑形成单位(pfu/ml)为单位的病毒感染滴度。
[0098]
实施例1.10.获取病毒dna。
[0099]
来自用质粒电穿孔或用mvm感染的细胞的低分子量dna是通过改进的hirt方法[segovia,jc,gallego,jm,bueren,ja and almendral,jm(1999)severe leukopenia and dysregulated erythropoiesis in scid mice persistently infected with the parvovirus minute virus of mice.,journal of virology,73(3),pp.1774
‑
1784]获得的。简而言之,将转染的细胞在补充有20μg/ml trna载体的hirt溶液(50mm tris ph7.5,0.5%sds,10mm edta)中裂解以确保恢复,并用蛋白酶k(100μg/ml)(merck)在37℃消化2小时。将反应调整至1mnacl,并使基因组dna在4℃下沉淀过夜。将样品在微量离心机(eppendorff)中于4℃下以14k rpm离心30分钟后,从上清液中获得富集的低分子量病毒dna的级分。将该dna在
‑
20℃下用0.3mnacl和2.5倍体积的无水乙醇沉淀,用70%乙醇洗涤以除去盐分,然后重悬于水或重悬于50mm tris ph7.5和1mm edta中。
[0100]
实施例1.11.dna电泳并转移到膜上。
[0101]
在tris
‑
硼酸盐
‑
edta缓冲液(45mm tris
‑
硼酸盐,1mm edta)中的含有5μg/ml溴化乙锭(boehringer)的0.8%琼脂糖凝胶(gibco)中,将样品连同分子量标记(噬菌体的hindiii消化)以及mvm的复制形式和ssdna的对照以60v进行2
‑
3小时电泳。在0.4m naoh中过夜转移至尼龙膜(hybond
‑
n ,amersham,pharmacia)上。最后,将膜在室温下在2
×
ssc溶液中洗涤10分钟(20
×
ssc:3m nacl、0.3m柠檬酸钠)并在78℃下干燥2小时。
[0102]
实施例1.12.与特异性mvm探针杂交。
[0103]
将膜在预杂交溶液(5
×
ssc,5
×
denhardts溶液[ficoll(ty400),聚乙烯吡咯烷酮,bsa],10mmtris
‑
hclph7.5,0.5%sds,50%甲酰胺)中在42℃下孵育四小时,以消除可能的非特异性结合。接下来,将溶液替换为杂交溶液,该杂交溶液由预杂交溶液的相同组分连同变性探针一起形成。探针是全长mvm基因组,通常使用dctp
‑
α
32
p通过“随机引发”用
32
p在体外标记为高特异性活性,并通过sephadex g
‑
50离心柱纯化。42℃下进行杂交1天或2天,最后用0.1
×
ssc和0.5%sds的溶液在50℃下洗涤膜3小时,然后再暴露于x射线胶片。
[0104]
实施例1.13.抗体
[0105]
免疫学技术中使用的一抗和二抗是:
[0106]
[0107][0108]
实施例1.14.化学试剂。
[0109]
羟基脲从calbiochem(羟基脲货号400046
‑
5gm)获得。5
‑
氟尿嘧啶(5fu)从sigma(参考号f
‑
6627
‑
1g)获得。顺铂从emc millipore(232120
‑
50mg)获得。
[0110]
实施例1.15.蛋白分析:sds
‑
page并转移到膜上。将蛋白质样品在上样缓冲液(10%甘油(merck),5%β
‑
巯基乙醇(merck),0.002%溴酚蓝(merck),0.5mtris
‑
hclph6.3,0.4%sds)中煮沸5分钟后变性后,通过8%聚丙烯酰胺的变性凝胶电泳分离。在tris
‑
甘氨酸缓冲液(25mm tri
‑
hcl(serva),192mm甘氨酸(gibco),0.1%sds)中以100v在微型凝胶(10cm
×
10cm
×
0.1cm)中进行电泳2小时至4小时,并与分子质量标记(“预染sds
‑
page标准品,宽范围”(biorad)或“蛋白质分子量标准品,宽范围”,amersham)同时跑胶。将样品在转移缓冲液(25mmtris碱,192mm甘氨酸,0.1%sds,20%甲醇)中以100v(trans
‑
blot电泳转移盒,biorad)转移到硝酸纤维素膜(schleicher和schuell)中,进行一小时。
[0111]
实施例1.16.检测印迹到膜上的蛋白质(“蛋白质印迹”)。
[0112]
使膜在tbs
‑
t缓冲液(20mmtrisph7.5,140mmnacl,0.1%吐温20)中水合,并在含有10%胎牛血清(fbs)的tbs
‑
t中在4℃下振荡孵育1小时。用tbs
‑
t洗涤后,将膜与在含1%fbs和1%np40的tbs
‑
t中稀释的一抗在4℃下一起孵育24小时。彻底洗涤后,添加二抗在rt下孵育1小时。最后,用tbs
‑
t和tbs(不含吐温20)洗涤膜,用ecl系统(
″
enhanced chemiluminiscence”,amersham)显影并以放射自显影术(kodak)曝光。
[0113]
实施例1.17.质粒。
[0114]
为了在细胞培养物中表达p53蛋白,使用了pcmv
‑
neo
‑
p53质粒[baker sj,markowitz s,fearon er,willson jk,and vogelstein b.suppression of human colorectal carcinoma cell growth by wild
‑
typep53.science 1990,249(4971):912
‑
5],该质粒由j.paramio(ciemat,madrid)友情提供。名为“辅助子(helper)”的质粒和表达腺病毒e4orf6致瘤基因的质粒[winter;k.,von kietzell,k.,heilbronn,r.,pozzuto,t.,fechner,h.,ands.weger(2012).roles of e4orf6 and va i rna inadenovirus
‑
mediesated stimulation of human parvovirus b19 dna replication and structural gene expression.j.virol.86,5099
‑
5109]由s.weger(charit
é
,berlin)友情捐赠。根据细胞类型,通过化学(jetpaid)或物理(电穿孔)方法将质粒转染到细胞中。
[0115]
实施例1.18.tp53基因测序。
[0116]
使用trizol和常规方案处理细胞样品以获得总rna。然后使用逆转录酶和随机引物将rna复制为cdna。使用tp53序列和附录(annex)中列出的引物将由此获得的cdna用于对人或小鼠tp53基因进行测序。所采用的方法是对从facs分选的gs中获得的rna样品进行常规桑格测序或大规模测序(ngs)(参见图2和图3)。两种测序方法均由马德里科学园(parque cient
í
fico de madrid,pcm)使用该设施中可用的设备和方法来进行。
[0117]
实施例2.结果.
[0118]
实施例2.1.细小病毒mvm主要ns1细胞毒性蛋白在人胶质母细胞瘤干细胞(gs)中的表达与p53诱导相关。
[0119]
图1(左)示出了来自被mvmp或mvmi菌株感染的三名患者(#5、#7和#8)的gs中的p53反应。在未感染的神经球中,p53染色较弱且均匀。感染后,在gs中检测到主要的非结构细胞毒性病毒蛋白ns1的表达,gs的百分比取决于患者而不同,但在所有三个病例中,ns1 细胞中都强烈且特异性地诱导了p53。当在蛋白质印迹中分析来自四名患者的gs感染时(图1,右),从来自大多数不表达ns1的细胞的信号背景不能明显看出p53的p53诱导。只有在对mvm更宽容的gs7中,这种诱导是显著的。因此,响应于mvm感染,gs细胞诱导真正的涉及p53的dna损伤反应(dnadamage response,ddr)。
[0120]
实施例2.2.容许细小病毒mvm基因组复制的那些gs诱导在丝氨酸15残基处通过磷酸化所修饰的p53。
[0121]
我们接下来研究了gs响应于细小病毒mvm而发生的ddr是否涉及细胞类型依赖性和/或病毒株依赖性的、通过磷酸化在p53的丝氨酸15残基处的修饰(pp53
‑
s15)。在未感染的gs神经球中观察到pp53
‑
s15的基础表达,其中在gs5和gs7中有一些突出的细胞,而当被mvmp感染时,在表达ns1的细胞的很大一部分中明显诱导了pp53
‑
s15(图2a)。图3b示出了在gs中及在各种已建立的系中响应于mvm菌株感染的基础表达百分比和pp53
‑
s15诱导的估计值。然而,神经球中的病毒基因组复制优先发生在那些pp53
‑
s15 gs细胞中(下图2a)。这种ns1 /pp53
‑
s15 相关性已通过流式细胞术定量证实(图2b),因为在来自两名受mvmp感染的患者的ns1 gs群中,病毒dna(vdna )的合成优选在表达高水平pp53
‑
s15的细胞中进行。
[0122]
实施例2.3.容许ns1表达并且有时容许mvm基因组复制的不同来源(包括各种类型的人类癌症)的转化细胞系携带通常通过ser15处的磷酸化而突变的和/或表型改变的p53。
[0123]
为了将之前的研究扩展到其他类型的癌症以及用于培养这些病毒的细胞,用两种病毒株(mvmp、mvmi)感染数种人和其他哺乳动物细胞系,并分析了ns1表达和病毒基因组复
制与tp53基因突变的存在和p53蛋白翻译后修饰的关系。在这些分析中获得的结果是:
[0124]
·
永生化小鼠成纤维细胞的a9细胞系通常用于培养mvmp病毒[gardiner,em and tattersall,p.1988.mapping of the fibrotropic and lymphotropic host range determinants of the parvovirus minute virus of mice.j virol 62(8):2605
‑
2613][gil
‑
ranedo,于,hernando,e.,riolobos,l.,dominguez,c.,kann,m.,and jos
éꢀ
m.almendral.2015.the mammalian cell cycle regulates nuclear parvovirus capsid assembly.plospathogens,11;11(6):e1004920],当病毒的大分子合成达到其最大值时,将感染了mvmp并用胸苷同步化的a9细胞系在该逮捕(arrest)释放9小时后进行分析[gil
‑
ranedo,j.,hernando,e.,riolobos,l.,dominguez,c.,kann,m.,and jos
éꢀ
m.almond tree 2015.the mammalian cell cycle regulates nuclear parvovirus capsid assembly.plospathogens,11;11(6):e1004920]。据观察,未感染的a9细胞的细胞周期在这些条件下在g2累积(图2c,上),而感染细胞中的很大一部分停留在g2之前。尽管细胞群中接近40%不表达ns1,但所有表达ns1的细胞都已发展到g2。更重要的是,那些ns1表达细胞中只有一半容许合成病毒dna(vdna)。p53是响应于感染而被诱导的,并且合成水平与ns1的水平相对应(图2c,下)。在这些细胞中,pp53
‑
s15组成型水平是不可检测的(也参见图3c),但响应于感染而被强烈诱导,并且那些vdna 细胞具有更高的p53
‑
s15诱导水平(图2c,中间图)。
[0125]
为了分析允许或不允许vdna合成的ns1 细胞之间的显著差异,我们继续对这两个细胞群进行分选并表征它们的tp53遗传。分选后的a9 ns1 细胞再次表现出分布在大小相似的两个vdna /
‑
群中(图3e,左)。值得注意的是,在两个a9群以及初始未分选的a9群中表达的mrna中检测到了tp53
‑
g665c突变(其决定v170l的氨基酸改变),但在不容许mvm感染的未转化的对照小鼠成纤维细胞中没有检测到(图3h)。因此,a9细胞具有无义v170l(对应于人tp53基因中的v173l)突变,该突变可能是ns1的表达所必需的(参见下文)。mvmp基因组复制主要限于感染诱导了ser15处p53磷酸化的a9细胞亚群。
[0126]
·
人和灵长类细胞系由致瘤病毒转化(图3a,上),该致瘤病毒表达使p53失活或功能改变的致瘤基因。这些是三种细胞系:nb324k,由致瘤多瘤病毒sv40 t抗原转化的人新生儿肾细胞[shein,hm,enders,jf and levinthal,jd(1962).transformation induced by simian virus 40in human renal cell cultures.ii.cell
‑
virus relationships.proceedings of the national academy of sciences of the united states of america,48,pp.1350
‑
7]。从cdna确定的这些细胞中tp53基因的序列没有突变(数据未显示),但它们确实显示了pp53
‑
s15组成型改变、与ns1表达相关的p53诱导以及大多数细胞中高水平的病毒基因组复制(图3a和图3d),尽管在病毒复制细胞中pp53
‑
s15表达水平有所不同。cos
‑
1细胞(也由sv40转化的灵长类成纤维细胞)和hela(从由乳头瘤病毒感染诱导的人宫颈癌中获得)中的结果相似(图3a),尽管病毒基因组复制和pp53
‑
s15组成型水平通常较低。
[0127]
·
从胶质母细胞瘤患者建立的细胞系。
[0128]
‑
u373
‑
mg和u251
‑
mg。这些细胞携带有编码p53中r273h氨基酸改变的tp53
‑
g849a突变,该突变在文献中有详细记载,我们通过其mrna的桑格测序证实了这一突变(数据未显示)。这些细胞对两种mvm(p、i)毒株的容许性很差,因为群中只有一小部分以低水平诱导
ns1,这与p53上升一致(图3a和图3d)。这些胶质母细胞瘤细胞系显示组成型pp53
‑
s15改变(尽管以基础水平改变),并且它们仅在那些低数量的pp53
‑
s15 细胞中维持mvm(p、i)菌株基因组复制(图3a)。
[0129]
‑
u87
‑
mg。如文献中先前所述,在该细胞系中未能发现tp53 mrna的遗传改变(数据未显示),它们也没有显示出pp53
‑
s15组成型修饰(参见图4、图6和图7)。然而,u87
‑
mg表达的p53蛋白必须具有其他类型的翻译后修饰,因为针对第212至217位氨基酸结构域的商品化抗体(invitrogen)确实识别了它(以及在其他细胞类型中的其他p53种类),而针对来自残基20的环境中的肽而获得的具有不同特异性的其他商品化抗体(cell signaling)不识别p53(图3f)。值得注意的是,u87
‑
mg细胞大多容许mvmi的ns1蛋白表达,但不允许mvmp的ns1蛋白表达,尽管mvmi不复制其基因组,也不诱导pp53
‑
s15 (图4和图6)。
[0130]
实施例2.4.细小病毒mvm的细胞毒性ns1蛋白的表达需要tp53突变或p53蛋白修饰。
[0131]
尽管tp53的遗传改变和/或p53蛋白的修饰与mvm的ns1蛋白的表达之间的相关性在数种转化细胞系中是一致的(图1
‑
图3),但该证据没有证明这两种现象之间存在因果关系。为了解决这个重要问题,我们继续在具有tp53
‑
v173l突变(实际上相当于v170l小鼠突变)的小鼠a9细胞和缺乏tp52突变但显示出高度磷酸化的p53
‑
s15的nb324k细胞(图3a和图3c)中从pcmv
‑
neo
‑
p53质粒过表达p53蛋白[baker sj,markowitz s,fearon er,willson jk,vogelstein b.suppression of human colorectal carcinoma cell growth by wild
‑
type p53.science 1990,249(4971):912
‑
5]。如图3g所示,在两种细胞类型中,用载体dna(从鲑鱼精子制备)转染时ns1的表达都非常高,而p53水平处于基础水平或是不可检测的。然而,当通过pcmv
‑
neo
‑
p53转染过表达p53时,会引起ns1表达的绝对抑制。那些以高于特征组成型水平过表达转染的wtp53的小鼠a9和人nb324k细胞完全排除了病毒ns1蛋白的积累(参见图3g中的ptp53的图)。因此,ns1的表达需要由tp53基因突变或p53蛋白翻译后修饰诱导的p53的功能改变。
[0132]
实施例2.5.细小病毒mvm表达细胞毒性蛋白并复制其基因组,优选在携带tp53遗传改变的人胶质母细胞瘤干细胞(gs)中进行。
[0133]
对已建立的细胞系的分析支持在pp53
‑
s15修饰与gs细胞中mvm感染的容许性之间发现的关联(图2a),并进一步表明这种关系可能来自遗传基础。因此,为了深入解决这个问题,对四名患者的gs中的tp53的遗传状态进行了分析。为此,获得了来自未感染对照、mvmp或mvmi感染的对照以及受mvmp感染并针对ns1和pp53
‑
s15表达进一步分选的gs的样品(facs,参见材料和方法)。图3e说明了提交进行遗传分析的ns1 /pp53
‑
s15 和ns1 /pp53
‑
s15细胞群的限制性窗(门)选择(以确保纯度)。从所有这些样品中纯化polya mrna,复制为cdna并通过pcr跨tp53基因进行扩增。通过新一代测序对这些扩增子进行测序(ngs,图2d和图2e),并通过传统的桑格测序方法进行确认(图3i)。检测到的tp53中的各种突变描述如下(从p53蛋白的n到c方向上进行讨论):
[0134]
·
p72r突变。c215g突变涉及p53的富含脯氨酸的结构域(prd)中的氨基酸残基p72r改变(无义突变)(参见图2f和图3j),该c215g突变在被感染和经facs分选( / 和 /
‑
)的gs7群中是很明显的,并且在很大比例的同样分选的gs8群和非常低比例的允许ns1表达( /
‑
)的gs5细胞中也是很明显的。因此,p53
‑
p72r的改变有利于mvmp的转录和复制,尽管在
不同程度上取决于gbm患者的gs背景。有趣的是,cosmic数据库描述了单个gbm样品中的这种突变(图2f),但它是造血肿瘤和淋巴肿瘤中的常见突变,尤其是p72r是迄今为止急性髓性白血病(acute myeloid leukemia,aml)中最常见的tp53突变(图3j)。
[0135]
·
v173l突变对应于我们已经在a9小鼠成纤维细胞系中鉴定的v170l改变。该突变已在两个gbm样品中被描述(图2f),它位于p53的dbd结构域的“热点(hotspot)”区域,在该区域已发现更多的突变(图3j)。v173l改变涉及多种癌症,但在直肠腺癌和肺腺癌中的频率尤其高(图3j)。
[0136]
·
g245s突变。通过ngs发现,来自患者8和患者9的gs的tp53基因在位置g733a处具有突变,这导致这些患者的所有细胞和所有样品(无论mvm感染的分选群和总群)的g245s氨基酸改变(无义突变)(图2d、图2e)。通过桑格测序在相同的样品中确认了突变(图3i)。突变的普遍存在表明它本身并不有益于病毒基因表达。然而,这种tp53
‑
g733a突变也出现在低比例的gs5细胞中(图2e),且仅在那些感染和分选的细胞中( / 和 /
‑
),这表明p53
‑
g245s的改变可能有助于这些gs5细胞中的mvm转录和复制。值得注意的是,这种突变已经在更新的gbm遗传数据库中高频描述(图2f),与之一致的是其在p53的“热点”区域dbd结构域内定位。此外,p53
‑
g245s改变是针对tp53遗传学进行分析的多种类型的人癌症中最常见的突变之一(图3j),例如膀胱癌、直肠腺癌和肺腺癌(图3j)。
[0137]
·
e258k突变。通过ngs在低百分比的表达ns1的感染和分选的gs7群体中观察到导致e258k氨基酸改变(错义突变)的g772a突变,强烈提示该g772a突变有助于这些gbm干细胞中的病毒转录。之前没有在当前的gbm遗传数据库中描述过该突变(图2f),尽管该突变也位于发现了更多突变的“热点”p53 dbd结构域中。事实上,e258k突变在许多类型的癌症中都是常见的(图2f,下),它是膀胱癌中非常常见的突变(图3j)。
[0138]
·
r273h突变。在我们分析的四名患者的原发性gs中没有发现这种无义突变(图2和图3),尽管它在gbm遗传数据库中非常常见(图2f)。值得注意的是,p53
‑
r273h突变目前被强调为所研究的数千种人癌症中最常见的突变(参见图3j中的示例)。尽管没有识别出四种gs中的r273h突变可能是由于样品数量有限,但我们的研究或多或少地表明,这种突变本身并不有益于mvm感染。这一假设得到以下事实的支持:携带有这种突变的u373mg和u251mg胶质母细胞瘤细胞系对mvmp和mvmi感染的容许性很低(图3a和图5)。然而,mvmp在u373mg胶质母细胞瘤系的细胞毒性感染中与化疗药物产生非常好的协同作用(图5,参见下文),强烈提示gbm和其他携带p53
‑
r273h突变的癌症的基因毒性化疗与mvm联合可能带来非常重要的临床益处。
[0139]
·
s366a突变。通过ngs在不取决于mvm感染的gs5和gs7的总体群和分选群中观察到导致s366a氨基酸改变(无义突变)的t1096g突变。这表明这种突变本身既不利于也不抑制细小病毒mvm感染。这种突变与其他突变不太可能在某种程度上与gs7和gs5对mvm感染的容许性有关,尽管可获得的数据不能排除这种可能性。突变s366a位于p53羧基末端,其在gbm样品中没有被描述(图2f)。事实上,它在人类癌症中的整体作用似乎不太相关(图3j,下)。
[0140]
综上所述,通过该分析可以得出以下相关结论:
[0141]
·
可能将mvmp基因组复制到不同的水平的表达ns1细胞毒性蛋白的所有gs群,以一定百分比携带一些tp53突变(单独或伴随其他突变)。
[0142]
·
分别有利于mvm在gs细胞系和胶质母细胞瘤细胞系中感染的g245s突变和r273h突变,是在gbm和其他具有遗传特征的人类癌症中发现的最常见的p53突变,这强调了我们分析的重要性。
[0143]
·
p72r、v173l和e258k突变在gbm中先前没有被描述过或者被发现的频率非常低。然而,它们在其他癌症类型中确实出现频率高,这可能允许将基于mvm的治疗扩展到其他癌症。
[0144]
·
p72r突变(除了一个病例)和e258k突变在gbm中先前没有被描述过,但在mvmp感染后通过ngs在gs中检测到它们,这可能允许将mvm用于诊断肿瘤中微小但临床相关的p53突变。
[0145]
·
携带与ns1表达相关的tp53突变的gs也诱导pp53
‑
s15磷酸化,这应该促进病毒转录和复制。
[0146]
·
因此,在gs群的巨大遗传异质性中,即使大多数细胞是wttp53,mvm也在细胞毒性感染中,有时也在病毒基因组复制中,特异性地靶向携带遗传改变的tp53的低比例gs。
[0147]
·
人类癌症中最常见的tp53突变r273h本身不会使细胞容许mvm,但它确实促进了病毒与化疗的重要的协同细胞毒性(更多参见下文)。
[0148]
实施例2.6.改变p53的致瘤基因的外源表达增加了gs和胶质母细胞瘤细胞系对mvm感染的容许性。
[0149]
具有组成型pp53
‑
s15染色和表达病毒致瘤基因的癌症细胞系中对ns1表达和mvm基因组复制的高度容许性(图3a,上)促使我们研究这两种特征之间可能的因果关系,即通过外源地功能性改变p53是否可以使wttp53细胞对mvm易感。图4a示出了不容许mvmp感染的nih3t3小鼠成纤维细胞在通过所谓的“辅助子”质粒转染后,大大增加了ns1表达和病毒基因组复制,该质粒通过e1a、e1b和e4orf6腺病毒致瘤基因的表达介导的降解而使p53失活,并通过va
‑
rna i而使pkr失活[winter,k.,von kietzell,k.,heilbronn,r.,pozzuto,t.fechner,h.,and s.weger(2012).roles of e4orf6 and va i rna inadenovirus
‑
mediesated stimulation of human parvovirus b19 dna replication and structural gene expression.j.virol.86,5099
‑
5109]。同时(图4d),如果用“辅助子”质粒或仅表达e4orf6致瘤基因的另一种质粒转染响应于ns1表达发生高ddr的mvmp感染的gs5细胞,则该gs5细胞会显著增加病毒基因组复制[wintcr,k.,von kietzell,k.,heilbronn,r.,pozzuto,t.,fechner,h.,and s.weger.(2012).roles of e4orf6 and va i rna in adenovirus
‑
mediated stimulation of human parvovirus b19 dna replicationandstructuralgeneexpression.j.virol.86,5099
‑
5109]。结果在gs8细胞中得到证实,尽管gs8细胞在ns1表达细胞中诱导p53,但gs8细胞对mvmp复制非常限制(图4b),但是,当通过转染表达致瘤基因时,观察到mvm复制显著增加(图4c)。该实验策略被转化到mvmi感染u87
‑
mg胶质母细胞瘤细胞上,u87
‑
mg胶质母细胞瘤细胞具有wttp53序列但表达翻译后修饰的p53蛋白(图3)。一致地,通过if共聚焦(图4e,左)和细胞计数(图4e,右)进行测量,致瘤基因表达也导致病毒基因组复制的显著增加。因此,使p53功能性失活的腺病毒致瘤基因增加了gs和具有wttp53的胶质母细胞瘤细胞系中的mvm复制。
[0150]
实施例2.7.改变p53磷酸化的化疗药物增加mvmp基因表达、复制以及杀死携带在多种类型的人类癌症中非常常见的tp53
‑
r273h突变的u373
‑
mg胶质母细胞瘤细胞的能力。
[0151]
由于mvm感染与pp53
‑
s15表达相关(图2和图3),因此我们研究了病毒感染是否可以从与临床方案中常用的化疗药物(chemotherapeutic drug,cd)(5fu、hu和cis
‑
pt)的联合治疗中受益。例如,cispt是临床方案中多种人类癌症的常用药物(dasari,s.,andtchounwou,pb.cisplatin in cancer therapy:molecular mechanisms of action.eur j pharmacol.2014october 5;0:364
‑
378)。在该研究中,我们使用了具有两种遗传背景的人胶质母细胞瘤细胞系。在第一项目研究中,携带r273h突变的u373mg经常存在于多种人类癌症中[muller,pa,and vousden,kh 2013.p53 mutations in cancer.nat.cell biol.13,2
‑
8][brennan,cw et al.the somatic genomic landscape of glioblastoma.cell 155,462
‑
477],此外,在我们对当前数据库进行的深度搜索中,u373mg似乎在gbm(图2f)和一般性癌症(图3j)中最常见。图5a通过流式细胞术分析mvmp感染的u373mg(允许以定量方式确定细胞中的蛋白质表达水平)显示,相比于单纯感染,在与单剂量cd的联合治疗中,表达ns1蛋白的细胞百分比以及表达水平得以增加。cispt处理决定了ns1表达的两个参数的最高增加。这种益处很可能与由cispt引起的所有细胞中普遍的pp53
‑
s15诱导有关,这也在处理的和未感染的细胞中观察到。这种效果得到了if共聚焦的证实(图5b)。
[0152]
因此,分析了cispt剂量对不同mvmp生命周期参数以及对u373
‑
mg细胞中p53磷酸化和功能信号传导的影响。为此,对在37℃下用顺铂剂量(10μm
‑
120μm)处理1小时的u373
‑
mg细胞接种mvmp进行吸附,然后使感染进展24小时。对细胞进行取样和处理,以用于病毒大分子标志物和pp53
‑
s15测定。通过if
‑
共聚焦观察到pp53
‑
s15水平在所有测定的5μm至20μm cis
‑
pt剂量范围内显著增加(图5c),这与与基础感染相比ns1表达和病毒基因组(vdna)复制的增加(%)是协调的。值得注意的是,病毒基因组复制水平受益于10μm cispt,但不会受益于更高剂量,更高剂量似乎会破坏vdna合成机制。通过蛋白质印迹证实了在5μm至20μm cispt剂量范围内ns1水平的增加(图5d),蛋白质印迹还反映出与p53功能信号传导相关的p21和mdm2因子水平的改变。
[0153]
我们最终分析了这些mvm/cispt协同效应是否影响癌细胞的活力,以及它们是否因此暗示了治疗意义。如图5e所示,使用cispt进行的简单的1小时处理逐渐降低了u373
‑
mg细胞的活力(克隆生成试验),这与其在肿瘤门诊中作为化疗药物的常见用途一致。此外,以两个mo1单独接种mvmp也会在一定程度上降低细胞活力。然而,当cispt和mvmp联合应用时,细胞活力以累加方式显著降低(图5e,5μm和10μm cispt)。这种药物与病毒在杀死携带tp53
‑
r273h突变的人u373
‑
mg胶质母细胞瘤细胞中的共同合作,可以在对携带该突变的多种人类癌症的治疗中产生重要的治疗益处(参见图3j)。
[0154]
实施例2.8.两种细小病毒株mvmp和mvmi与cd联合治疗增加了它们的基因表达和复制以及它们杀死携带p53翻译后修饰的u87
‑
mg人胶质母细胞瘤细胞的能力。
[0155]
在第二个研究中,将对cd/mvm联合治疗的这种分析扩展到缺乏tp53突变但表达翻译后修饰p53蛋白形式的u87
‑
mg人胶质母细胞瘤细胞系(图3f)。在第一个分析中,研究了可能的cd/mvmi协作。在基础状态下,u87
‑
mg细胞没有显示出显著的pp53
‑
s15信号,mvmi表达高ns1水平但不能复制其基因组(图6d)。然而,hu和5fu处理未感染的u87
‑
mg细胞本身显著增加了ser15残基处的p53磷酸化(图6a、图6b)。与这种效应一致,当以选定的剂量施用这些cd时,它们会成比例地增加这些细胞中病毒基因组的复制水平(图6c、图6d)。如图6f和图6h
所示,这种vdna合成增加对应于mvmi与5fu(其效果达到协同水平)以及与oh
‑
u(附加效果)联合杀死u87
‑
mg细胞的更大能力。图6e和图6g显示出,伴随感染而增加的pp53
‑
s15磷酸化(相对于未感染细胞)在为病毒带来最佳益处的cd剂量下更高,这也决定了p21水平(p53的功能性标志物)更明显的下降。
[0156]
在第二个分析中,分析了cd/mvmp协作。u87
‑
mg的mvmp感染(图7a)即使在高感染复数(moi 10)下也会产生非常低比例的ns1 细胞。然而,当细胞在感染前用不同的cispt剂量处理时,观察到ns1 细胞的百分比非常显著地增加(图7a)。这种增加与pp53
‑
s15水平的增加一致,而pp53
‑
s15水平与使用的cispt剂量相关。通过蛋白质印迹也证明了ns1/pp53
‑
s15协同增加(图7b),蛋白质印迹也反映了p21改变。如上文对u373mg所描述的,mvmp基因组复制(vdna)也受益于10μm或20μm cispt处理,但不会受益于更高剂量,表明病毒复制机制相比于病毒转录机制对cispt的敏感性增加(图7a)。ns1
细胞%的增加对应于mvmp与cispt联合杀死u87
‑
mg细胞的更大能力,其在10μm和20μm剂量下达到协同水平(图7c)。有趣的是,在这些剂量下,感染细胞中p21水平(p53的功能性标志物)的降低更为明显(图7b)。最后,使mvmp与5fu(图7d)和hu(图7e)联合,在这些cd在未感染细胞中诱导更高pp53
‑
s15水平的剂量下,也在u87
‑
mg细胞杀伤中也产生了一致的累加效应(分别参见图6a和图6b)。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。