含氧二萜化合物的制备
1.序列列表参考
2.本技术包含计算机可读形式的序列列表,其通过引用并入。
技术领域
3.本发明涉及在重组细胞如酵母细胞中生产含氧二萜化合物。含氧二萜化合物可用作合成有用生物活性化合物的中间或最终化合物,以用于例如药物治疗疾病(例如癌症)。本发明还涉及特别适合于生产这种化合物的基因、酶和细胞,例如酵母细胞。
背景技术:
4.萜烯是一组由基本的5碳结构异戊二烯(2-甲基-1,3-丁二烯)生成的多种化合物。二萜是具有20个碳结构的化合物,由一种酶二萜合酶的作用产生,该酶将化合物双(牻牛儿基)二磷酸盐(ggpp)转化为二萜结构,这通过进一步修饰可以形成广泛的diterpene或二萜化合物。
5.diterpene、二萜及其衍生物被广泛用作药物、化妆品、营养品、香料、香料和杀虫剂。在天然或工程细胞中增加这些化合物的产生的方法在本领域中是丰富的。
6.已知雷公藤这种中药植物能产生几种具有潜在药理性能的倍半萜、二萜和三萜,包括二萜化合物雷公藤内酯酮和雷公藤甲素。雷公藤甲素,一种含氧二萜化合物,及其衍生物已被确定为潜在的有价值的药理化合物,并正在作为免疫抑制剂和治疗癌症进行研究。雷公藤甲素可进一步用于治疗covid-19。雷公藤内酯酮可能用作男性避孕药。
7.使用工程微生物从可再生原料中生产有价值的分子是传统生产方式的理想选择。然而,实现经济上可行的产量、效价和生产率是工业化的主要障碍。
8.n l hansen等人(2017)在the plantjournal 2017,89,429-441中描述一种二萜合酶,能够将ggpp转化为dipterpene,即松香烷型二萜烯,这是雷公藤甲素的前体。p su等人(2018)在the plantjournal 2018,50-65:和j guo等人(2018)在pnas 2013,110,12108-12113中证实了这一发现。
9.将松香烷型二萜烯转化为其他二萜化合物(如雷公藤甲素)的完整途径尚未阐明。
10.细胞色素p450酶(cyp)参与terpenoid的生物合成,对于许多细胞色素p450酶来说,对于它们作用的底物、它们产生的化合物或它们在特定化合物生物合成中的作用一无所知。
11.us 20190270971a1公开了用于提高功能性表达p450酶的微生物宿主细胞的生产率的方法。该文件描述了如何修饰p450基因以提高微生物(如酵母)的性能,并提到与细胞色素p450还原酶的共表达有助于提高产量。提到雷公藤甲素可能是p450化学的主题,但该文件没有提供雷公藤甲素与任何特定p450酶或细胞色素之间的任何联系。
12.cn 108395997a描述了ggpp产量增加的酵母。用不同的二萜合酶和p450酶转化酵母以合成二萜化合物。该专利背后的科学家团队还支持更多的专利和专利申请,公开了使用合适的terpene合成酶和p450酶合成不同的二萜和三萜化合物,例如cn 108866029
(friedelin)、cn 107058419(kauren型)和wo 2020029564(fridelin和amyrins)。
13.cn 110747178a将p450基因twcyp728b70描述为编码在雷公藤甲素合成中具有作用的细胞色素p450酶。
技术实现要素:
14.本发明人解决了提供一种制备含氧二萜化合物如雷酚内酯、雷公藤内酯酮和雷公藤甲素的改进方法的问题。
15.在第一方面,本发明涉及一种制备含氧二萜化合物的方法,该方法包括以下步骤:
16.a.提供能够产生松香烷型二萜烯和/或脱氢松香二烯的宿主细胞;
17.b.用编码具有细胞色素p450活性的酶的第一基因转化宿主细胞;
18.c.在导致转化基因表达的条件下生长转化细胞;由此形成含氧二萜化合物;
19.其中:
20.编码具有细胞色素p450活性的酶的第一基因编码多肽,所述多肽包含与seq id no:1(twcyp82d274v1)、seq id no:2(twcyp82d274v2)、seq id no:74(twcyp82d274v3)或seq id no:75(twcyp82d274v4)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列,或其成熟多肽。
21.在第二方面,本发明涉及制备含氧二萜化合物的方法,并包括用编码具有细胞色素p450活性的酶的第一基因和进一步用编码具有细胞色素p450活性的第二酶的第二基因以及编码具有细胞色素p450活性的第三酶的第三基因转化宿主细胞,
22.其中:
23.编码具有细胞色素p450活性的酶的第二基因编码多肽,所述多肽包含与seq id no:3(twcyp71be85)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列,或其成熟多肽;和
24.编码具有细胞色素p450活性的酶的第三基因编码多肽,所述多肽包含与seq id no:4(twcyp71be85)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列,或其成熟多肽。
25.在第三方面,本发明涉及制备含氧二萜化合物的方法,其中所述方法包括用编码具有细胞色素p450活性的酶的第一、第二和第三基因转化宿主细胞,并且进一步用编码具有细胞色素p450活性的第四酶的第四基因转化宿主;
26.其中:
27.编码具有细胞色素p450活性的酶的第四基因编码多肽,所述多肽包含与seq id no:5(twcyp82d213v1)或seq id no:76(twcyp82d213v2)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列,或其成熟多肽。
28.根据本发明,可以提供有用的含氧二萜化合物雷酚内酯、雷公藤内酯酮或雷公藤甲素。
29.本发明还涉及用于本发明方法的多肽、多核苷酸、质粒和表达构建体以及重组宿主细胞。
附图说明
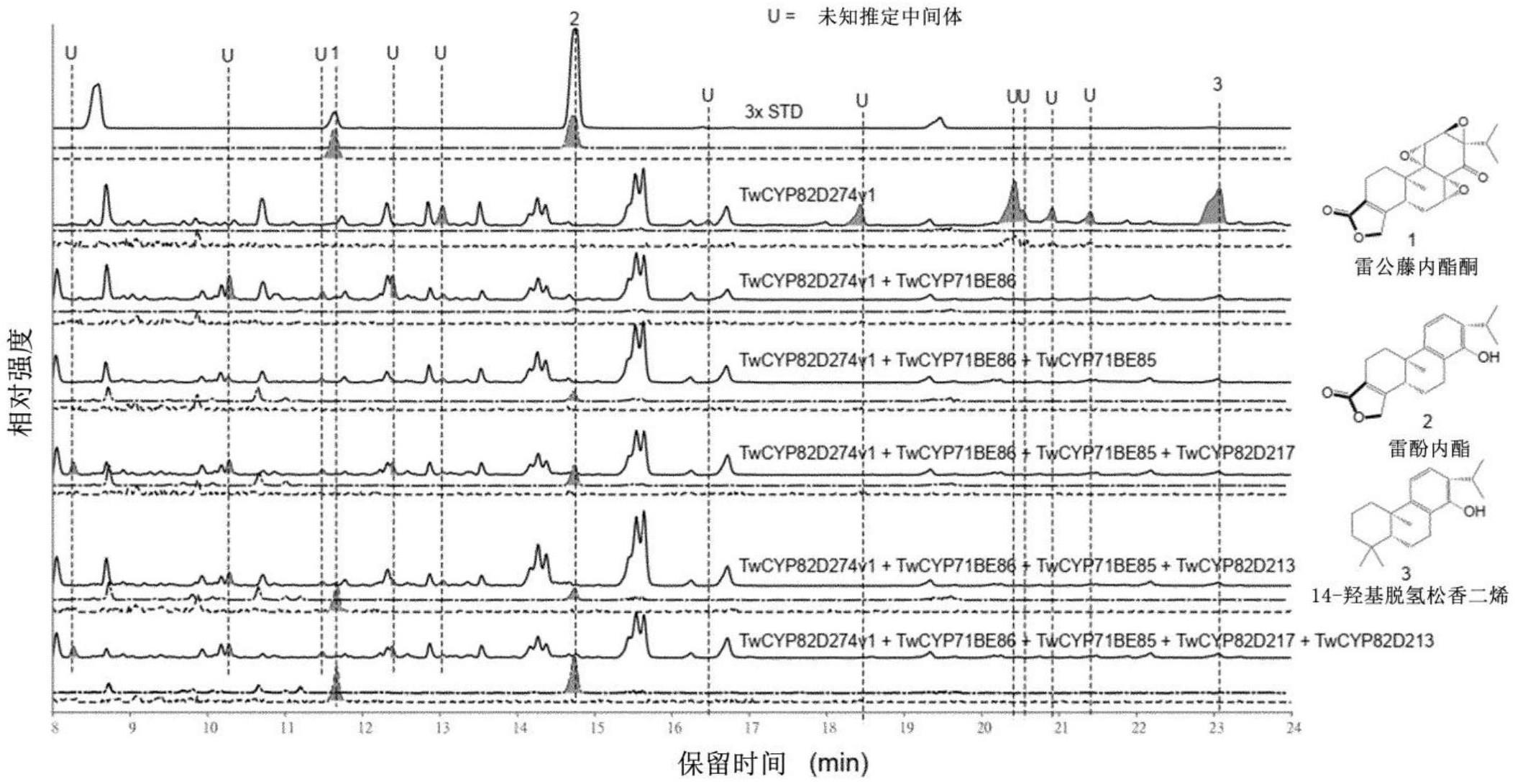
30.图1显示了表达miltiredene生物合成基因和选定的t.wilfordii cyp的n.benthamiana叶提取物的lcms谱。
31.twcyp与cfdxs、cfggpps、cftps1和cftps3共表达。“3xstd”代表三种混合真实标准品的lcms运行:雷公藤甲素、雷公藤内酯酮和雷酚内酯。实线表示m/z 280-380范围内的离子色谱图。虚线(
‑‑‑‑‑‑‑
)表示在m/z 313.1800
±
0.015处提取的离子色谱图,对应于雷酚内酯[m h]的亲代离子。虚线(
‑‑‑
)表示在m/z 359.1490
±
0.015处提取的离子色谱图,对应于雷公藤内酯酮[m h]的亲代离子。使用lc协议1。有关详细信息,请参见实施例1。
[0032]
图2显示了基因工程酿酒酵母(s.cerevisiae)菌株提取物的lcms图谱。
[0033]
在背景菌株(-)中,编码二萜生物合成酶spggpps7、cfttps1、cfttps3和twcpr1的基因被整合到野生型酿酒酵母的基因组中。在twcyp82d274v1菌株中,二萜生物合成酶用twcyp82d274v1表达,导致化合物(3)的形成,被鉴定为14-oh-脱氢松香二烯,标记为灰色。lc方法1用于分析。有关详细信息,请参见实施例3。
[0034]
图3显示了在599.85mhz下cdcl3中14-oh-脱氢松香二烯的1h nmr光谱。有关详细信息,请参见实施例4。
[0035]
图4显示了在150.83mhz下cdcl3中14-oh-脱氢松香二烯的
13
c nmr谱。有关详细信息,请参见实施例4。
[0036]
图5显示了整合了指定基因组合基因组的酵母提取物的lcms图谱。
[0037]
所有酵母菌株都具有基因组整合的spggpps7、cfttps1、cfttps3和twcpr1。“0.5ppm 3xstd”代表三种混合真实标准品的lcms运行:雷公藤甲素、雷公藤内酯酮和雷酚内酯。非虚线表示m/z 280-380范围内的离子色谱图。虚线表示在m/z 359.1490
±
0.015处提取的离子色谱图,对应于雷公藤内酯酮([m h]
)的亲代离子。使用lc协议2。有关详细信息,请参见实施例5。
[0038]
图6显示了从雷公藤中分离的twcyp和b5蛋白的不同变体的共表达
[0039]
从工程化酿酒酵母菌株的培养物中量化的雷酚内酯、雷公藤内酯酮和14-oh-脱氢松香二烯的水平。每一列代表工程酵母菌株及其所选化合物的产量。整合到每个单独菌株中的基因在下图中表示。定量基于代表每种感兴趣化合物的峰面积。每种化合物都有单独的刻度。有关详细信息,请参见实施例5。
[0040]
图7显示了雷公藤内酯酮拟议生物合成途径中关键中间体的相对数量(条形),当在体内建立时,通过在n.benthamiana(图a和d)和酿酒酵母(图b和e,表3中列出的菌株)中的异源基因表达。基因表达由左边的黑色方块表示,而相对数量由条形表示(3-4个生物复制的平均值;黑色菱形方块),白色和灰色填充颜色区分表达,并且twb5#1无表达。误差条表示标准偏差。“ditps”反映cfttps1和cfttps3。在峰面积的量化中,gcms(松香烷型二萜烯和14-oh-脱氢松香二烯)和lcms(所有其他化合物)的特征质量公差分别为
±
0.1m/z和
±
0.005m/z。图c:在n.benthamiana和酿酒酵母体内从松香烷型二萜烯到雷公藤内酯酮的假设生物合成途径,包括wagner-meerwein重排反应,以解释abietane碳骨架中c-19或c-18向
c-3的甲基转移。
[0041]
图8显示了在生物反应器中生长时,酵母菌株nvj8.15产生的雷酚内酯和雷公藤内酯酮在7天内的累积。雷公藤内酯酮(实线黑线)和雷酚内酯(虚线黑线)的水平显示了每天采集的培养物样品中的绝对量(ppm,w/v)。通过600nm处的吸光度(灰色虚线)定量生物质。
[0042]
图9显示了表达雷公藤内酯酮生物合成所需基因的酵母菌株,但其基因变体取代了twcyp82d274v1或twcyp82d113v1,与产生雷酚内酯(图a)和雷公藤内酯酮(图b)的能力有关,并导致类似的lcms谱(图c)。工程菌株中存在的基因用黑色方块表示。图a和b:条形图表示平均相对数量(2-3个生物重复,交叉),误差条形图显示标准误差。从左到右,条形代表酵母菌株:nvj10-1、nvj10-3、nvj10-6、nvj10-8(见表3)。图c:lcms分析酵母培养物的eic(m/z 280-360)。从上到下成对的色谱图表示酵母菌株nvj10-1、nvj10-3、nvj10-6和nvj10-8。
[0043]
序列的简短描述
[0044]
seq id no:1显示来源于雷公藤的细胞色素p450酶的氨基酸序列。该酶也称为twcyp82d274v1。
[0045]
seq id no:2显示来源于雷公藤的细胞色素p450酶的氨基酸序列。该酶也称为twcyp82d274v2。seq id no:2与seq id no:1仅在三个位置上不同,因此假设seq id no:1和seq id no:2代表同一基因的不同等位基因。
[0046]
seq id no:3显示来源于雷公藤的细胞色素p450酶的氨基酸序列。该酶也称为twcyp71be85。
[0047]
seq id no:4显示源自t.wilfordii的细胞色素p450酶的氨基酸序列。该酶也称为twcyp71be86。
[0048]
seq id no:5显示源自t.wilfordii的细胞色素p450酶的氨基酸序列。该酶也称为twcyp82d213v1。
[0049]
seq id no:6显示源自t.wilfordii的细胞色素p450酶的氨基酸序列。该酶也称为twcyp82d217。
[0050]
seq id no:7显示来源于雷公藤的细胞色素p450酶的氨基酸序列。该酶也称为twcyp82d275。
[0051]
seq id no:8显示来源于雷公藤的细胞色素b5酶的氨基酸序列。这种酶也被称为twb5#1。
[0052]
seq id no:9显示来源于雷公藤的细胞色素p450还原酶的氨基酸序列。这种酶也称为twcpr1。
[0053]
seq id no:10-66显示了实施例2中进一步描述的pcr引物。
[0054]
seq id no:67显示源自plectranthus barbatus的二萜合酶tps1的氨基酸序列。这种酶也称为cftps1。
[0055]
seq id no:68显示源自plectranthus barbatus的二萜合酶tps3的氨基酸序列。这种酶也称为cftps3。
[0056]
seq id no:69显示了萜合酶tps9的氨基酸序列,其来源于雷公藤。该酶也称为twtps9。
[0057]
seq id no:70显示了源自雷公藤的萜烯合酶的氨基酸序列。该酶也称为twtps27。
[0058]
seq id no:71显示了源自salvia miltiorrhiza的柯巴基焦磷酸合酶cps1的氨基
酸序列。这种酶也称为smcps。
[0059]
seq id no:72显示源自salvia miltiorrhiza的松香烷型二萜烯合酶ksl1的氨基酸序列。这种酶也称为smksl。
[0060]
seq id no:73显示了源自synechococcus sp的geranyl geranyl二磷酸合酶的氨基酸序列。该酶也称为spggpps7v1。
[0061]
seq id no:74显示源自雷公藤的细胞色素p450酶的氨基酸序列。该酶也被称为twcyp82d274v3。
[0062]
seq id no:75显示源自雷公藤的细胞色素p450酶的氨基酸序列。该酶也称为twcyp82d274v4。
[0063]
seq id no:76显示源自雷公藤的细胞色素p450酶的氨基酸序列。该酶也称为twcyp82d213v2。
[0064]
seq id no:77显示源自plectranthus barbatus的二萜合酶tps1的截断氨基酸序列。截断氨基酸序列以去除转运肽。这种酶也称为cfttps1。
[0065]
seq id no:78显示源自plectranthus barbatus的二萜合酶tps3的截断氨基酸序列。截断氨基酸序列以去除转运肽。这种酶也被称为cfttps3。
[0066]
seq id no:79显示源自plectranthus barbatus的dxs酶的氨基酸序列。这种酶也称为cfdxs。
[0067]
seq id no:80显示源自酿酒酵母的截断hmgr酶的氨基酸序列。这种酶也称为scthmgr。
[0068]
seq id no:81显示了源自synechococcus sp的geranyl geranyl二磷酸合酶的氨基酸序列。该酶也称为spggpps7v2。
具体实施方式
[0069]
根据本发明的第一方面,提供一种制备含氧二萜化合物的方法,该方法包括以下步骤:
[0070]
a.提供能够产生松香烷型二萜烯和/或脱氢松香二烯的宿主细胞;
[0071]
b.用编码具有细胞色素p450活性的酶的第一基因转化宿主细胞;
[0072]
c.在导致转化基因表达的条件下生长转化细胞;由此形成含氧二萜化合物;
[0073]
其中:
[0074]
编码具有细胞色素p450活性的酶的第一基因编码多肽,所述多肽包含或由下列组成:seq id no:1或与seq id no:1(twcyp82d274v1)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列,或其成熟多肽。
[0075]
具有seq id no:1的多肽是具有细胞色素p450活性的第一基因的优选实例,具有seq id no:2、seq id no:74和seq id no:55的多肽是这种多肽的其他实例。
[0076]
因此,由第一基因编码的酶具有通过在松香烷型二萜烯的二萜骨架的第14位插入oh基团将松香烷型二萜烯和/或脱氢松香二烯转化为14-oh-脱氢松香二烯的能力。
[0077]
在一些实施方案中,14-oh-脱氢松香二烯的合成通过化合物14-oh-松香烷型二萜烯进行,该化合物随后转化为14-oh-脱氢松香二烯。然而,本发明不限于将松香烷型二萜烯
转化为14-oh-脱氢松香二烯的任何特定机制。
[0078]
使用根据本发明第一方面的方法导致含氧二萜化合物14-oh-脱氢松香二烯的形成,
[0079]
14-oh-脱氢松香二烯
[0080]
这是合成药用含氧二萜化合物的有用中间体,包括众所周知的化合物,如雷酚内酯、雷公藤内酯酮和雷公藤甲素。
[0081]
在本发明的第二方面,方法步骤b.包括用编码具有细胞色素p450活性的酶的第一基因转化宿主细胞,并且进一步用编码具有具有细胞色素p450活性的第二酶的第二基因和编码具有细胞色素p450活性的第三酶的第三基因来转化宿主细胞,
[0082]
其中:
[0083]
编码具有细胞色素p450活性的酶的第二基因编码多肽,所述多肽包含或由下列组成:seq id no:4或与seq id no:4(twcyp71be86)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列,或其成熟多肽;和
[0084]
编码具有细胞色素p450活性的酶的第三基因编码多肽,所述多肽包含或由下列组成:seq id no:3或与seq id no:3(twcyp71be85)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列,或其成熟多肽。
[0085]
在该第二方面中,宿主细胞优选进一步制备含氧二萜化合物雷酚内酯(3br,9bs)-6-羟基-9b-甲基-7-丙-2-基-3,3b,4,5,10,11-六氢化萘并[2,1-e]异苯并呋喃-1-酮。
[0086]
雷酚内酯
[0087]
雷酚内酯是一种有价值的化合物,已被鉴定为抗雄激素。此外,它可以作为进一步修饰的起点,从而产生进一步的生物活性化合物。
[0088]
在本发明第二方面的优选实施方案中,用编码具有细胞色素b5活性的多肽的第五基因进一步转化宿主细胞,所述多肽包含或由下列组成:seq id no:8或与seq id no:8(twb5#1)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列,或其成熟多肽。令人惊讶地发现,在表达编码具有细胞色素p450活性的酶的第一、第二和第三基因的同一细胞中表达具有细胞色素b5活性的多肽,导致含氧二萜化合物的产量显著增加。与没有具有细胞色素b5活性的多肽的类似细胞的生产相比,生产增加至少50%,优选增加至少100%,优选至少200%或甚至更多。
[0089]
在本发明的第三方面中,用编码具有细胞色素p450活性的酶的第一、第二和第三基因转化宿主细胞,并且进一步用编码具有生物色素p450的活性的第四酶的第四基因来转化宿主细胞;
[0090]
其中:
[0091]
编码具有细胞色素p450活性的第四酶的第四基因编码多肽,所述多肽包含或由下列组成:seq id no:5或与seq id no:5(twcyp82d213v1)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列,或其成熟多肽。具有seq id no:5的多肽是具有细胞色素p450活性的第四基因的优选实例;具有seq id no:76的多肽是这种多肽的另一个例子。
[0092]
在本发明的第三方面,转化的真核细胞优选产生含氧二萜化合物雷公藤内酯酮。
[0093]
雷公藤内酯酮
[0094]
化合物雷公藤内酯酮据报道在癌症中具有强烈的抑制活性(fulu dong et al 2019,the prostate,volume 19,issue 11,第1284-1293页)。该化合物也可用作男性避孕药。此外,该化合物可用作进一步修饰的起点,从而产生进一步的生物活性化合物。
[0095]
在本发明第三方面的优选实施方案中,用编码具有细胞色素b5活性的多肽的第五基因进一步转化宿主细胞,并且所述多肽包含或由下列组成:seq id no:8或与seq id no:8(twb5#1)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列,或其成熟多肽。令人惊讶地发现,在表达编码具有细胞色素p450活性的酶的第一、第二和第三基因的同一细胞中表达具有细胞色素b5活性的多肽,导致含氧二萜化合物的产量显著增加。与没有具有细胞色素b5活性的多肽的类似细胞的生产相比,生产增加至少50%,优选增加至少100%,优选至少200%或甚至更多。
[0096]
在本发明第三方面的另一优选实施方案中,宿主细胞进一步用编码具有细胞色素p450活性的第五酶的第六基因和/或编码具有细胞细胞色素p45活性的第六酶的第七基因转化,
[0097]
其中:
[0098]
编码具有细胞色素p450活性的第五酶的第六基因编码多肽,所述多肽包含或由下列组成:seq id no:6或与seq id no:6(twcyp82d217)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列,或其成熟多肽;和
[0099]
编码具有细胞色素p450活性的第六酶的第七基因编码多肽,所述多肽包含或由下列组成:与seq id no:7(twcyp82d275)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列,或其成熟多肽。令人惊讶地发现,表达编码具有细胞色素p450活性的酶的第六
和/或第七基因,导致含氧二萜化合物的更高产量。优选地,与没有编码具有细胞色素p450活性的酶的第六和/或第七基因的类似细胞的生产相比,生产增加至少10%,优选增加至少20%,甚至更优选增加至少50%或甚至更多。
[0100]
第一、第二、第三、第四、第五、第六和第七基因可以包含在一种或多种核酸分子中,例如一种或多种异源核酸。编码具有细胞色素p450活性的第一酶的异源核酸在此可称为“第一异源核酸”。编码具有细胞色素p450活性的第二酶的异源核酸在此可称为“第二异源核酸”。编码具有细胞色素p450活性的第三酶的异源核酸在此可称为“第三异源核酸”。编码具有细胞色素p450活性的第四酶的异源核酸在此可称为“第四异源核酸”。编码具有细胞色素p450活性的第五酶的异源核酸在此可称为“第五异源核酸”。编码具有细胞色素p450活性的第六酶的异源核酸在此可称为“第六异源核酸”。编码具有细胞色素b5活性的酶的异源核酸在此可称为“第七异源核酸”。这并不意味着重组宿主细胞必须总共包含七种异源核酸;在一些实施方案中,细胞仅包含第一、第二、第三、第四、第五、第六和第七异源核酸中的一种或多种。
[0101]
根据本发明方法制备的含氧二萜化合物可通过生物或化学合成进一步修饰。与此相关,生物合成被理解为一种方法,其中包含本发明基因的宿主细胞进一步被提供有一个或多个额外的基因,所述基因编码具有修饰根据本发明方法制备的含氧二萜化合物的能力的其他酶。
[0102]
根据本发明方法制备的含氧二萜化合物的化学修饰可以在回收含氧二酚化合物之前直接在培养液上进行,或者可以在回收的含氧三萜化合物上进行。
[0103]
可以通过有机合成将雷公藤内酯酮还原为雷公藤甲素。这种合成的一个例子是通过氢化物对c-14酮的亲核攻击进行还原。对于该反应,硼氢化钠是在适当的溶剂例如水或甲醇中在中性ph下催化该反应的合适试剂。
[0104]
在一个优选实施方案中,根据本发明方法生产的雷公藤内酯酮被转化为化合物雷公藤甲素,该化合物据报道为免疫抑制剂,正在研究用于癌症治疗。
[0105][0106]
宿主细胞
[0107]
能够产生松香烷型二萜烯和/或脱氢松香二烯的宿主细胞原则上可以是任何这样的细胞。该细胞可以是天然产生松香烷型二萜烯和/或脱氢松香二烯的细胞,也可以是经过工程设计产生一种或两种化合物的细胞。
[0108]
据信,至少在某些情况下,松香烷型二萜烯可以自发地转化为脱氢松香二烯,因此本发明可以使用生产松香烷型二萜烯的细胞进行,该细胞自发地转化成脱氢松香二烯,或者可以在含有促进松香烷型二萜烯转化为脱氢松香二烯的酶的细胞中进行(参见j.zi,et al.,organic&biomolecular chemistry 2013,11,7650-7652)。
[0109]
松香烷型二萜烯的合成通常始于ggpp的形成。ggpp可以通过一个二甲基烯丙基焦
12113,以及这些出版物中公开的程序和方法也可用于提供用于本发明的宿主细胞。
[0118]
宿主细胞可以是原核细胞,例如真细菌或古细菌细胞;或真核细胞,例如植物细胞、动物细胞、昆虫细胞、真菌细胞或酵母细胞。
[0119]
实际上,所有真核细胞都为其生物合成产生ggpp,但在一些实施方案中,与不产生增加量ggpp的类似真核细胞相比,真核细胞产生增加量的ggpp,这可能增加松香烷型二萜烯的产量。现有技术中还描述了增加真核细胞中ggpp产量的方法。
[0120]
宿主细胞可以是单细胞生物体,也可以包含在多细胞生物体内,例如植物。用作根据本发明的宿主细胞的合适植物或植物细胞的实例包括玉米(zea mays)、油菜(brassica napus,brassica rapa ssp.)、苜蓿(medicago sativa)、水稻(oryza sativa)、黑麦(s secale cerale)、高粱(sorghum bicolor,sorghum vulgare)、向日葵(helianthus annuas)、小麦(tritium aestivum和其他物种)、小黑麦、黑麦(secale)大豆(glycine max)、烟草(nicotiana tabacum)、马铃薯(solanum tuberosum)、花生(arachis hypogaea)、棉花(gossypium hirsutum)、甘薯(impomoea batatus)、木薯(manihotesculenta)、咖啡(coffea spp.)、椰子(cocos nucifera)、菠萝(anana comosus)、柑橘(citrus spp.)、可可(theobroma cacao)、茶(camellia senensis)、香蕉(musa spp)、鳄梨(persea americana)、无花果(ficus casica)、番石榴(psidium guajava)、芒果(mangiferindica)、橄榄(olea europae)、木瓜(carica papaya)、腰果(anacardium occidentale)、澳洲坚果(macadamia intergrifolia)、杏仁(primus amygdalus)、苹果(malus spp.)、梨(pyrus spp.)、李子和樱桃树(prunus spp.)、ribes(currant etc.)、葡萄、菊芋(helianthemum spp.)、非谷类草(禾本科)、糖和饲料甜菜(甜菜属)、菊苣、燕麦、大麦、蔬菜或观赏植物,农作物(例如,谷物和豆类、玉米、小麦、土豆、木薯、大米、高粱、小米、木薯、大麦、豌豆、甜菜、甘蔗、大豆、油菜、向日葵和其他根茎、块茎或种子作物)。其他重要的植物可能是果树、作物树、林木或用作香料或医药产品的植物(mentha spp.,clove,artemesia spp.,thymus spp.,lavendula spp.,allium spp.,hypericum,catharanthus spp.,vinca spp.,papaver spp.,digitalis spp.,rawolfia spp.,vanilla spp.,petrusilium spp.,eucalyptus,茶树,picea spp.,pinus spp.,abies spp.,juniperus spp)。可用于本发明的园艺植物可包括莴苣、菊苣和蔬菜芸苔,包括卷心菜、花椰菜和花椰菜、胡萝卜、康乃馨和天竺葵。
[0121]
植物也可以是烟草、葫芦、胡萝卜、草莓、向日葵、番茄、辣椒或菊花。
[0122]
植物的其它实例包括谷物植物,例如油籽植物或豆科植物。感兴趣的种子包括谷物种子,如玉米、小麦、大麦、高粱、黑麦等。油料种子植物包括棉花大豆、红花、向日葵、芸薹属植物、玉米、苜蓿、棕榈、椰子等。豆科植物包括豆类和豌豆。豆类包括瓜尔豆、刺槐豆、葫芦巴、大豆、菜豆、豇豆、绿豆、利马豆、蚕豆、扁豆、鹰嘴豆。
[0123]
特别优选的植物种类包括physcomitrella sp.,如p.patens;拟南芥属,如拟南芥;烟草属(nicotiana sp.),如n.benthamiana;衣藻属(chlamydomonas sp.),如c.reinhardtii;以及nannochloropsis sp.(n.oceanica)等南绿球藻(nannochloropsis sp.)。
[0124]
根据本发明使用的合适的真核细胞的实例包括真菌细胞,如agaricus、aspergillus、念珠菌、eremothecium、镰刀菌/赤霉素、克鲁维酵母、laetiporus、lentinus、
phaffia、phanerochaete、毕赤酵母、physcomitrella、rhodoturula、酿酒酵母、裂殖酵母、sphaceloma、xanthophyllomyces或耶罗维亚酵母。来自这些属的示例性物种包括lentinus tigrinus、硫黄菌、黄孢平革菌,pichia pastoris,cyberlindnera jadinii,physcomitrella patens,rhodoturula glutinis,粘液真菌,phaffia rhodozyma,红法夫酵母,fusariumfujikuroi/gibberellafujikuroi,candida utilis,candida glabrata,白色念珠菌和yarrowia lipolytica。
[0125]
在一些实施方案中,宿主细胞可以是子囊菌,例如赤霉素(gibberella fujikuroi)、乳酸克鲁维酵母(kluyveromyces lactis)、裂殖酵母(schizosaccharomyces pombe)、黑曲霉(aspergillus niger)、溶脂亚罗菌(yarrowia lipolytica)、棉花灰霉(ashbya gosspiii)或酿酒酵母。
[0126]
在一些实施方案中,宿主细胞可以是藻类细胞,例如三孢布拉克菌(blakeslea trispora)、杜氏盐藻(dunaliella salina)、雨生红球藻(haematoccus pluvialis)、小球藻(chlorella sp.)、裙带菜(裙带菜)、马尾藻(sargasum)、海带(laminaria japonica)、almeriensis scenedsmus。
[0127]
在一些实施方案中,宿主细胞可以是原核生物,例如芽孢杆菌细胞,例如枯草芽孢杆菌;大肠杆菌细胞,例如大肠杆菌细胞;乳杆菌细胞;乳球菌细胞;链霉菌细胞、链球菌细胞、角膜细菌细胞;醋酸杆菌细胞;不动杆菌细胞;或假单胞菌细胞。
[0128]
在一些实施方案中,宿主细胞可以是蓝藻细胞,例如聚囊藻属或聚球藻属。
[0129]
在一个实施方案中,选择适合在发酵罐中生长的宿主细胞。生长根据本发明的重组宿主细胞是生长宿主细胞以生产本发明的含氧二萜化合物的方便方法。
[0130]
在另一个实施方案中,宿主细胞是趋光细胞,并且细胞在温室或光生物反应器中培养。
[0131]
基因和酶
[0132]
本发明的重组宿主细胞能够产生能够产生松香烷型二萜烯和/或脱氢松香二烯。松香烷型二萜烯可以自发转化为脱氢松香二烯,也可以通过促进松香烷型二萜烯转化为脱氢松香二烯的酶进行转化。
[0133]
如上所述,松香烷型二萜烯的合成通常以一个二甲基烯丙基焦磷酸(dmap)分子和三个异戊烯基焦磷酸(ipp)分子通过双(牻牛儿基)二磷酸合酶缩合形成ggpp开始。
[0134]
重组宿主细胞和编码在重组宿主细胞中催化ggpp合成的酶的异源核酸在本领域中是公知的,参见例如wo 2015/113570。此外,许多宿主生物能够产生ggpp,因此异源核酸可能并不总是ggpp生产所必需的。
[0135]
在一些实施方案中,重组宿主细胞包含编码双(牻牛儿基)二磷酸合酶的异源核酸,如seq id no:73或seq id no:81中所述双(牻牛儿基)二磷酸合酶spggpps7,或与其具有至少80%、例如至少81%、例如至少82%、例如至少83%、例如至少84%、例如至少85%、例如至少86%、例如至少87%、例如至少88%、例如至少89%、例如至少90%、如至少91%、例如至少92%、例如至少93%、例如至少94%、例如至少95%、例如至少96%、例如至少97%、例如至少98%、例如至少99%的序列同一性的功能同源物,或其成熟多肽。
[0136]
随后,ggpp可通过一个或多个二萜合酶、柯巴基焦磷酸合酶和/或松香烷型二萜烯合酶的作用转化为松香烷型二萜烯。
[0137]
在一些实施方案中,重组宿主细胞包含一种或多种编码一种或多种二萜合酶的异源核酸,如二萜合酶cftps1(seq id no:67)和cftps3(seq id no:68)、或cfttps1(seq id no:77)和cfttps3(seq id no:78)、或与其具有至少80%、例如至少81%、例如至少82%、例如至少83%、例如至少84%、例如至少85%、例如至少86%、例如至少87%、例如至少88%、例如至少89%、例如至少90%、如至少91%、例如至少92%、例如至少93%、例如至少94%、例如至少95%、例如至少96%、例如至少97%、例如至少98%、例如至少99%的序列同一性的相应功能同源物,或其成熟多肽。
[0138]
在一些实施方案中,重组宿主细胞包含一种或多种编码一种或多种二萜合酶的异源核酸,如二萜合酶twtps9(seq id no:69)和twtps27(seq id no:70)、或与其具有至少80%、例如至少81%、例如至少82%、例如至少83%、例如至少84%、例如至少85%、例如至少86%、例如至少87%、例如至少88%、例如至少89%、例如至少90%、如至少91%、例如至少92%、例如至少93%、例如至少94%、例如至少95%、例如至少96%、例如至少97%、例如至少98%、例如至少99%的序列同一性的相应功能同源物,或其成熟多肽。
[0139]
在一些实施方案中,重组宿主细胞包含一种或多种柯巴基焦磷酸合酶和一种或多种松香烷型二萜烯合酶的组合,如柯巴基焦磷酸合酶smcps(seq id no:71)和松香烷型二萜烯合酶smksl(seq id no:72)的组合、或与其具有至少80%、例如至少81%、例如至少82%、例如至少83%、例如至少84%、例如至少85%、例如至少86%、例如至少87%、例如至少88%、例如至少89%、例如至少90%、如至少91%、例如至少92%、例如至少93%、例如至少94%、例如至少95%、例如至少96%、例如至少97%、例如至少98%、例如至少99%的序列同一性的相应功能同源物,或其成熟多肽。
[0140]
在又一方面,本发明涉及具有细胞色素p450酶活性的多肽,并且所述多肽包含与seq id no:1、seq id no:2、seq id no:3、seq id no:4、seq id no:5、seq id no:6和seq id no:7中的一种具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、优选至少96%序列同一性、优选至少97%序列同一性、优选至少98%序列同一性、或甚至100%序列同一性的氨基酸序列,或其成熟多肽。
[0141]
在又一方面,本发明涉及具有细胞色素b5酶活性的多肽,并且所述多肽包含与seq id no:8具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、优选至少96%序列同一性、优选至少97%序列同一性、优选至少98%序列同一性、或甚至100%序列同一性的氨基酸序列,或其成熟多肽。
[0142]
本发明还涉及编码具有细胞色素p450酶活性的多肽的多核苷酸序列或基因,并且包含与seq id no:1、seq id no:2、seq id no:3、seq id no:4、seq id no:5、seq id no:6和seq id no:7中的一种具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、优选至少96%序列同一性、优选至少97%序列同一性、优选至少98%序列同一性、或甚至100%序列同一性的氨基酸序列,或其成熟多肽;或者编码具有细胞色素b5活性的多肽,所述多肽包含与seq id no:8具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、优选至少96%序列同一性、优选至少97%序列同一性、优选至少98%序列同一性、或甚至100%序列同一性的氨基酸序列,或其成熟多肽。
[0143]
在优选实施方案中,具有细胞色素p450活性的第一、第二、第三、第四、第五和第六
酶中的一种或多种包含或由根据以下任一项的氨基酸序列组成:seq id no:1(twcyp82d274v1)、seq id no:2(twcyp82d274v2)、seq id no:74(twcyp82d274v3)、seq id no:75(twcyp82d274v4)、seq id no:3(twcyp71be85)、seq id no:4(twcyp71be86)、seq id no:5(twcyp82d213v1)和seq id no:76(twcyp82d213v2),与其具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、优选至少96%序列同一性、优选至少97%序列同一性、优选至少98%序列同一性、优选至少99%序列同一性的相应功能同系物。
[0144]
在一些实施方案中,编码具有细胞色素p450活性的第一酶的第一异源核酸编码下列所述twcyp82d274:seq id no:1(twcyp82d274v1,seq id no:2(twcyp82d274v2),seq id no:74(twcyp82d274v3),seq id no:75(twcyp82d274v4),或与其具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物。
[0145]
在一些实施方案中,编码具有细胞色素p450活性的第二酶的第二异源核酸编码下列细胞色素p450酶twcyp71be86:seq id no:4,或与其具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物。
[0146]
在一些实施方案中,编码具有细胞色素p450活性的第三酶的第三异源核酸编码下列细胞色素p450酶twcyp71be85:seq id no:3,或与其具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物。
[0147]
在一些实施方案中,编码具有细胞色素p450活性的第四酶的第四异源核酸编码下列细胞色素p450酶:seq id no:5中所述twcyp82d213v1或seq id no:76(twcyp82d213v2)中所述twcyp82d213v2,或与其具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物。
[0148]
在一些实施方案中,编码具有细胞色素p450活性的第五酶的第五异源核酸编码下列细胞色素p450酶twcyp82d217:seq id no:6,或与其具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物。
[0149]
在一些实施方案中,编码具有细胞色素p450活性的第六酶的第六异源核酸编码下列细胞色素p450酶twcyp82d275:seq id no:7,或与其具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物。
[0150]
在一些实施方案中,编码具有细胞色素b5活性的酶的第七异源核酸编码下列细胞色素b5酶twb5#1:seq id no:8,或与其具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物。
[0151]
在一些实施方案中,提供了重组宿主细胞:
[0152]
i.其中所述宿主细胞能够产生松香烷型二萜烯和/或脱氢松香二烯;和
[0153]
ii.包含编码下列异源核酸:seq id no:1、seq id no:2、seq id no:74或seq id no:75的twcyp82d274,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽,
[0154]
其中所述细胞能够产生14-羟基脱氢松香二烯。
[0155]
在一些实施方案中,提供重组宿主细胞:
[0156]
i.其中所述宿主细胞能够产生松香烷型二萜烯和/或脱氢松香二烯;和
[0157]
ii.包含编码下列异源核酸:seq id no:1、seq id no:2、seq id no:74或seq id no:75的twcyp82d274,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0158]
iii.包含编码下列异源核酸:seq id no:4的twcyp71be86,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;
[0159]
其中所述细胞能够生产14-羟基脱氢松香二烯、3,14-二羟基脱氢松香二烯、3,14-dihydroxyabeodiene和/或14-羟基-18-醛-abeodiene。
[0160]
在一些实施方案中,提供了重组宿主细胞:
[0161]
i.其中所述宿主细胞能够产生松香烷型二萜烯和/或脱氢松香二烯;和
[0162]
ii.包含编码下列异源核酸:seq id no:1、seq id no:2、seq id no:74或seq id no:75的twcyp82d274,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0163]
iii.包含编码下列异源核酸:seq id no:4的twcyp71be86,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0164]
iv.包含编码下列异源核酸:seq id no:3的twcyp71be85,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;
[0165]
其中所述细胞能够生产14-羟基脱氢松香二烯,3,14-二羟基脱氢松香二烯,3,14-dihydroxyabeodiene,14-羟基-18-醛-abeodiene和/或雷酚内酯。
[0166]
在一些实施方案中,提供了重组宿主细胞:
[0167]
i.其中所述宿主细胞能够产生松香烷型二萜烯和/或脱氢松香二烯;和
[0168]
ii.包含编码下列异源核酸:seq id no:1、seq id no:2、seq id no:74或seq id no:75的twcyp82d274,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0169]
iii.包含编码下列异源核酸:seq id no:4的twcyp71be86,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0170]
iv.包含编码下列异源核酸:seq id no:3的twcyp71be85,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0171]
v.包含编码下列异源核酸:seq id no:5或seq id no:76的twcyp82d213,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;
[0172]
其中所述细胞能够生产14-羟基脱氢松香二烯,3,14-二羟基脱氢松香二烯,3,14-dihydroxyabeodiene,14-羟基-18-醛-abeodiene,雷酚内酯和/或雷公藤内酯酮。
[0173]
在一些实施方案中,提供了重组宿主细胞:
[0174]
i.其中所述宿主细胞能够产生松香烷型二萜烯和/或脱氢松香二烯;和
[0175]
ii.包含编码下列异源核酸:seq id no:1、seq id no:2、seq id no:74或seq id no:75的twcyp82d274,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0176]
iii.包含编码下列异源核酸:seq id no:4的twcyp71be86,或具有至少80%,如至
少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0177]
iv.包含编码下列异源核酸:seq id no:3的twcyp71be85,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0178]
v.包含编码下列异源核酸:seq id no:5或seq id no:76的twcyp82d213,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0179]
vi.包含编码下列异源核酸:seq id no:8的twb5#1,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;
[0180]
其中所述细胞能够产生效价比相同酵母细胞高至少2倍,例如至少3倍,例如高至少4倍,例如低至少5倍的雷公藤内酯酮,除非所述酵母不表达所述twb5#1或其功能同系物。
[0181]
在一些实施方案中,提供了重组宿主细胞:
[0182]
i.其中所述宿主细胞能够产生松香烷型二萜烯和/或脱氢松香二烯;和
[0183]
ii.包含编码下列异源核酸:seq id no:1、seq id no:2、seq id no:74或seq id no:75的twcyp82d274,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0184]
iii.包含编码下列异源核酸:seq id no:4的twcyp71be86,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0185]
iv.包含编码下列异源核酸:seq id no:3的twcyp71be85,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0186]
v.包含编码下列异源核酸:seq id no:5或seq id no:76的twcyp82d213,或具有
至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;和
[0187]
vi.包含编码下列异源核酸:seq id no:8的twb5#1,或具有至少80%,如至少81%,如至少82%,如至少83%,如至少84%,如至少85%,如至少86%,如至少87%,如至少88%,如至少89%,如至少90%,如至少91%,如至少92%,如至少93%,如至少94%,如至少95%,如至少96%,如至少97%,如至少98%,如至少99%序列同一性的功能同系物,或其成熟多肽;
[0188]
其中所述细胞能够在发酵培养基中生长,并且其中发酵7天后所述发酵培养基包括:
[0189]-至少3ppm雷公藤内酯酮和/或
[0190]-至少1ppm雷酚内酯。
[0191]
上述重组宿主细胞可能由于几种不同的原因而能够产生松香烷型二萜烯和/或脱氢松香二烯。例如,宿主细胞可能内源性地能够产生松香烷型二萜烯。或者,重组宿主细胞可包含一个或多个异源核酸序列,其编码一种或多种参与松香烷型二萜烯生产的酶,例如seq id no:73或seq id no:81的二萜生物合成酶spggpps7、seq id no:67的cftps1、seq id no:77的cfttps1、seq id no:68的cfts3、seq id no:78的cfttps3和/或seq id no:9的twcpr1,与其具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、优选至少96%序列同一性、优选至少97%序列同一性、优选至少98%序列同一性、优选至少99%序列同一性的相应功能同系物,或其成熟多肽。
[0192]
具有细胞色素p450活性的第一(例如twcyp82d274)、第二(例如twcyp71be86)、第三(例如,twcyp71be85)、第四(例如:twcyp82d213)、第五(例如twcyp82d217)和第六(例如,twcyp82d275)酶以及具有细胞色素b5活性的酶(例如,twb5#1)的功能同系物可以通过在酵母细胞中表达相关蛋白并评估它们是否能够产生下文所述的特定化合物来验证。
[0193]
一种酵母细胞,表达twcyp82d274(seq id no:1、seq id no:2、seq id no:74或seq id no:75)的功能同系物,并且还表达:
[0194]
i.二萜生物合成酶spggpps7v2(seq id no:81)、cfttps1(seq id no:77)、cfttps3(seq id no:78)和twcpr1(seq id no:9),
[0195]
优选能够生产14-羟基脱氢松香二烯。
[0196]
一种酵母细胞,表达twcyp71be86(seq id no:4)的功能同系物,并且还表达:
[0197]
i.二萜生物合成酶spggpps7v2(seq id no:81)、cfttps1(seq id no:77)、cfttps3(seq id no:78)和twcpr1(seq id no:9);和
[0198]
ii.twcyp82d274(seq id no:1或seq id no:2),
[0199]
优选能够生产14-羟基脱氢松香二烯,3,14-二羟基脱氢松香二烯,3,14-dihydroxyabeodiene和14-羟基-18-醛-abeodiene。
[0200]
一种酵母细胞,表达twcyp71be85(seq id no:3)的功能同系物,并且还表达:
[0201]
i.二萜生物合成酶spggpps7v2(seq id no:81)、cfttps1(seq id no:77)、cfttps3(seq id no:78)和twcpr1(seq id no:9);和
[0202]
ii.twcyp82d274(seq id no:1或seq id no:2);和
[0203]
iii.twcyp71be86(seq id no:4),
[0204]
优选能够生产14-羟基脱氢松香二烯,3,14-二羟基脱氢松香二烯,3,14-dihydroxyabeodiene,14-羟基-18-醛-abeodiene和雷酚内酯。
[0205]
一种酵母细胞,表达twcyp82d213(seq id no:5或seq id no:76)的功能同系物,并且还表达:
[0206]
i.二萜生物合成酶spggpps7v2(seq id no:81)、cfttps1(seq id no:77)、cfttps3(seq id no:78)和twcpr1(seq id no:9);和
[0207]
ii.twcyp82d274(seq id no:1或seq id no:2);
[0208]
iii.twcyp71be86(seq id no:4);和
[0209]
iv.twcyp71be85(seq id no:3),
[0210]
优选能够生产14-羟基脱氢松香二烯,3,14-二羟基脱氢松香二烯,3,14-dihydroxyabeodiene,14-羟基-18-醛-abeodiene,雷酚内酯和雷公藤内酯酮。
[0211]
一种酵母细胞,表达twb5#1(seq id no:8)的功能同系物,并且还表达:
[0212]
i.二萜生物合成酶spggpps7v2(seq id no:81)、cfttps1(seq id no:77)、cfttps3(seq id no:78)和twcpr1(seq id no:9);
[0213]
ii.twcyp82d274(seq id no:1或seq id no:2);
[0214]
iii.twcyp71be86(seq id no:4);
[0215]
iv.twcyp71be85(seq id no:3);和
[0216]
v.twcyp82d213(seq id no:5或seq id no:76)
[0217]
优选能够产生效价比相同酵母细胞高至少2倍,例如至少3倍,例如,至少4倍,例如:至少5倍的雷公藤内酯酮,除非所述酵母不表达twb5#1的所述功能同系物。
[0218]
在优选实施方案中,具有细胞色素b5活性的酶包含或由下列组成:根据seq id no:8(twb5#1)的氨基酸序列,或与其具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、优选至少96%序列同一性、优选至少97%序列同一性、优选至少98%序列同一性、优选至少99%序列同一性的相应功能同系物,或其成熟多肽。
[0219]
本发明的多核苷酸可以通过从天然产生多肽的生物体(如植物雷公藤或近缘植物)中克隆来提供,或者可以通过基于本领域已知技术的多核苷酸序列的化学合成来提供,或者其可以具有自然界中未发现的序列,例如该序列可以针对特定选择的宿主细胞进行密码子优化。
[0220]
本发明的多肽可由天然产生多肽的生物提供,例如植物雷公藤或相关生物;或者它们可以通过将编码多肽的多核苷酸插入和表达到合适的宿主细胞中并从包含用相应基因转化的宿主细胞的培养液中回收多肽来提供。优选从合适的选择的重组宿主细胞提供本发明的多肽。
[0221]
为了在合适的宿主细胞中转化和表达基因,该基因通常与合适的调节元件可操作地连接,并插入适合于特定选择的宿主细胞的表达载体中。选择合适的调节元件、构建合适的表达载体和转化所选择的宿主细胞是普通从业者的技能范围内的,并且本发明不受这些元件的任何特定选择的限制。
[0222]
所产生的包含本发明基因的宿主细胞适合在容器中生长,例如发酵罐或摇瓶;在基因表达和含氧二萜化合物形成的条件下。当生长停止或培养液中积累了足够高量的含氧二萜化合物时,可进一步修饰含氧二酚化合物并从培养液中回收。
[0223]
序列同一性被理解为两个氨基酸或核苷酸序列之间相似性的度量。通过首先对齐两个序列来计算序列同一性,计算两个序列包含相同氨基酸残基或核苷酸的位置的数量,并计算同一性百分比,即具有相同氨基酸残基或核苷酸的位点的数量与整个对齐长度的同一性。
[0224]
已经开发了几种算法,并且可供技术人员使用。
[0225]
在本说明书和权利要求中,使用默认参数(blosum 62矩阵,间隙开放惩罚11;间隙扩展惩罚1,exp.thr10)使用ncbi blast 成对比对算法计算氨基酸序列的序列同一性,使用默认参数(匹配/不匹配分数1,-3;间隙打开罚分5;间隙扩展罚分2;exp.thr10)。ncbi blast 项目进一步描述于:madeira f el at(2019)nar 47:w636-w641。
[0226]
实施例
[0227]
材料和方法
[0228]
nicotiana benthamiana的基因工程
[0229]
从植物材料中克隆雷公藤cyp基因,并使用先前在(1-4)中描述的构建体和方法,在nicotiana benthamiana中与二萜生物合成基因cfdxs(seq id no:79)或scthmgr(seq id no:80)、cfggpps或spggpps7(seq id no:81)、cftps1(seq id no:67)或cfttps1(seq id no:77)、和cftps3(seq id no:68)或cfttps3(seq id no:78)共表达。基因沉默抑制剂p19也共同表达。简言之,将各自含有单独二萜生物合成基因或雷公藤cyp(twcyp)的二元载体转化为农杆菌。混合各自含有特定质粒的农杆菌液体培养物,以共同表达twcyp的特定组合。
[0230]
酿酒酵母的基因工程及工程化酿酒酵母的生长条件
[0231]
培养基
[0232]
ypd培养基:20g/l bacto
tm
蛋白胨,10g/l bacto
tm
酵母提取物,20g/l葡萄糖。
[0233]
不含尿嘧啶的合成完全(sc)meda:1.92g/l不含尿苷的酵母合成脱落培养基补充剂(sigma aldrich co.llc.目录号y1501)、6.7g/l不含氨基酸的酵母氮基(sigma-aldrich co.llc目录号y0626)、20g/l葡萄糖。进料时间(fit)基于enpump 200(enpresso gmbh),并根据产品随附的方案制造。琼脂平板:sc培养基,包括琼脂(15g/l)。
[0234]
通过在含有5-氟乳清酸(5-foa,0.74g/l)和尿嘧啶(30mg/l)的不含尿嘧啶的sc培养基的琼脂平板上选择ura3功能缺失,在亲本菌株中引入尿嘧啶营养缺陷型。
[0235]
在不含尿嘧啶琼脂平板的sc上分离酵母转化体。
[0236]
工程酿酒酵母菌株的分批补料发酵分离松香烷型二萜烯衍生二萜
[0237]
所有工程化酿酒酵母菌株均采用与前述类似的进料时间(fit;m2p实验室)方法在96个深井板中培养(插入参考forman等人2018)。为了分离和纯化雷公藤内酯酸途径中的关键中间体,使用2l a生物反应器(sartorius ag)在饲料分批发酵罐中培养选定的工程酿酒酵母菌株。通过向反应器罐(带叶轮)中加入100ml发酵剂培养物,开始补料分批发酵,然后通过高压灭菌提前制备,同时含有200ml分批葡萄糖和300ml分批盐混合物。还加入5ml维生素混合物、5ml微量元素和0.5ml微量元素。生物反应器中的培养在以下条件下开始
(监测和自动控制):ph=5,温度=30℃,溶解氧(do)=20%。当通过供给氢氧化铵(32%)和硫酸(10%)来控制ph时,通过空气供给和搅拌来控制溶解氧。此外,通过添加抗泡沫乳液(35119,serva electrophology gmbh)来调节泡沫水平。在生物反应器中初始培养18小时后,开始以1.3%的速率用进料溶液进料。发酵过程持续7天,每天采样培养物。
[0238]
用于lc-ms分析的工程酿酒酵母的提取
[0239]
将基因工程酿酒酵母菌株转移到96孔板中的0.5ml培养基中,在30℃下以350转/分的轨道振荡生长3天。为了提取,将0.1ml酿酒酵母培养物转移到1.5ml玻璃瓶中。加入0.4ml meoh uhplc级。使用0:22μm 96孔滤板(merck millipore,darmstadt,德国)过滤酿酒酵母提取物,并在lc-ms分析之前在4℃下储存。
[0240]
用于lc-ms分析的二萜类代谢物的提取
[0241]
用于lcms分析的酵母培养物样品在1.5ml玻璃瓶中通过将酵母培养物和掺有5ppm穿心莲内酯(内标;fa17902,carbosynth)的甲醇以1:19(v/v)的比例混合(对于每日生物反应器样品)和1:4(v/v)的比例(对于96个深井培养物)制备。在室温下摇动混合30分钟。对于烟草样品,将2个叶盘置于1.5ml玻璃瓶中,用1ml甲醇提取溶液提取,在室温下振荡1小时。
[0242]
在lcms分析之前,样品通过0.22μm 96孔板过滤器(merck millipore,darmstadt,德国),并在5℃下储存。
[0243]
lc-ms分析
[0244]
使用与bruker compact esi-qtof-ms(bruker)系统耦合的ultimate3000uhplc 聚焦系统(dionex corporation,sunnyvale,ca)分析甲醇(meoh)提取物。样品在kinetex xb-c18柱(100x 2.1mm id,1:7μm粒径,100
°
a孔径;phenomenex inc.,torrance,ca)上分离,保持40℃,流速为0.3ml min-1,流动相包括0.05%(v/v)甲酸溶于水(溶剂a)和0.05%(v/v)甲酸溶于乙腈(溶剂b)。
[0245]
使用了两种lc协议:
[0246]
lc方法1:0-0.5分钟,10%b;0.5-21min,从10%b线性增加到80%b;21-31分钟,至90%b;31-34分钟,至100%b;34-39分钟100%b;39-40分钟线性从100%降至10%b。
[0247]
lc方法2:0-0.5分钟,20%b;0.5-11分钟,从20%b线性增加到80%b;11-20分钟,至90%b;20-22分钟,至100%b;22-27分钟100%b;27-28分钟线性从100%降至20%b。
[0248]
lc方法3:0-0.5分钟,20%b;0.5-9分钟,从20%b线性增加到100%b;9-11分钟,100%b;11-11.5分钟,从100%b线性下降至20%b;11.5-15分钟,20%b。
[0249]
用于gc-ms分析的二萜类代谢物的提取
[0250]
通过以1:4(v/v)的比例混合酵母培养物和纯甲醇,在1.5ml玻璃瓶中制备用于gcms分析的酵母培养物样品。短暂混合后,将非极性成分液-液萃取到己烷中,以1:1(v/v)的比例混合并振荡1小时,加入10ppm 1-二十碳烯。gcms分析之前,将己烷层转移到新的小瓶中。气相色谱-质谱(gc-ms)分析
[0251]
在岛津gcms-qp2010 ultra(岛津公司)上使用agilent hp-5ms柱(agilent technologies)20m x 0.18mm i.d.,0.18μm膜厚)进行gc-ms分析。以50cm s-1的恒定线速度使用氢气作为载气,在250℃下注入体积为1μl(无分流模式)。烤箱程序为80℃ 2分钟,以20℃/min的速率升温至180℃,以10℃/min速率升温至300℃,并以20
°
c/min速率升温至310
℃,保持3分钟。数据以.cdf格式存储并以mzmine2处理。
[0252]
松香烷型二萜烯衍生二萜类化合物的相对定量
[0253]
酵母培养物中的相对化合物量基于特征离子的归一化峰面积(使用mzmine2软件中的目标特征检测获得的数据)。对以下离子的信号进行了量化:1:松香烷型二萜烯m/z 91.1,2:14-羟基松香二烯m/z189.1,3:f15p1 m/z 303.2318,4:f20p2 m/z 283.2059,5:f15p2 m/z299.2002,6:雷酚内酯m/z 313.1794,8:雷公藤内酯酮m/z 359.1481。lcms和gcms数据的质量偏差分别为5ppm和100ppm。
[0254]
内标穿心莲内酯的基峰离子峰面积(m/z 315.1947)用于标准化。
[0255]
绝对量化
[0256]
通过共同分析甲醇制备的真实标准品和最终浓度为5ppm的内标物(穿心莲内酯),对雷酚内酯(ft65732,carbosynth)和雷公藤内酯酮(ft65197,carbosynth)进行了绝对定量。定量基于标准化峰面积,并根据标准反应曲线的线性外推斜率计算(雷酚内酯0.05、0.5、1、2ppm;雷公藤内酯酮0.5、1,2、10、20ppm)。
[0257]
用于核磁共振分析的酿酒酵母工程菌株中松香烷型二萜烯衍生物的分离和纯化
[0258]
本发明中的化合物从生物反应器培养物酵母菌株nvj8.15和nvj3.10中分离,并通过nmr进行结构鉴定。最初在celite (06858,sigma aldrich)存在下通过旋转蒸发干燥肉汤和甲醇裂解细胞(细胞:甲醇=1:4,v/v)的合并乙酸乙酯提取物。随后使用5.250(interchim,france)仪器通过连续分馏分离化合物,并通过紫外吸收和蒸发光散射检测(elsd)进行检测。该装置配备有(c1)pf-15sihp-f0025(ov002a,interchim)柱和(c2)us5c18hq-100/300(ssp750,interchin)柱,分别用于正相和反相分离。
[0259]
使用柱(参考文献9)对/粗提取物的干混合物进行初步预分级,并从手动填充的干负载柱加载。使用流动相己烷(a)和乙酸乙酯(b)(恒定流速为15ml/min)进行分离,然后用100%甲醇进行最终洗涤步骤。通过uv和elsd检测并收集感兴趣的化合物。在进一步分馏或nmr研究之前,使用lc-ms方法3和tlc分析通过lcms连续评估收集的馏分。通过使用c1的额外正相馏分或使用c2的反相柱分馏,从具有多种化合物的馏分中额外纯化感兴趣的化合物。
[0260]
对于c2的反相纯化,使用转子蒸发蒸发样品,并将其重新悬浮在2ml甲醇中。将样品直接注射到预处理柱c2上。c2的流动相由溶剂c:去离子水和溶剂d乙腈组成,各自用0.05%(v/v)甲酸酸化。使用32ml/min的恒定流速,随着溶剂d浓度的增加,具有线性溶剂梯度。通过elsd和uv检测并收集感兴趣的化合物。
[0261]
通过在岛津hplc(spd-m20a二极管阵列检测器、frc-10a馏分收集器、dgu-20a5脱气器、lc-20at泵、cbm-20a系统控制器、cto-10as vp柱烘箱、sil-10ap自动进样器)上多次注射100ul ontp半制备phenomenex luna 5μm c18(2)100?50x10mm(全多孔)柱(phenomenox,inc.,加利福尼亚州托伦斯,美国),完成额外的反相纯化。流动相在c和d之间呈线性梯度,d的量从50-100%增加。通过210nm的紫外吸收检测并收集感兴趣的化合物。
[0262]
质谱
[0263]
在m/z 50-1200的扫描范围内以正离子模式采集质谱,esi和ms设置如下:毛细管
电压,4000v;端板偏置,500v;干气温度,220c干气流量为8l min1;喷雾器压力,2巴;源cid能量为0ev;六极rf,50vpp;四极离子能量,4ev;碰撞电池能量,7ev。使用内部甲酸钠标准校准原始色谱数据,随后使用dataanalysis 4.3(build 110.102.1532)(64位),bruker导出为mzml格式。mzmine ver2.53用于可视化lc-ms色谱图。
[0264]
生物反应器起始培养基和进料培养基的培养基配方
[0265]
分批葡萄糖:
[0266]
葡萄糖一水合物55g/l
[0267]
分批盐混合物:
[0268]
硫酸铵25g/l
[0269]
磷酸二氢钾5g/l
[0270]
七水硫酸镁1.7g/l
[0271]
饲料葡萄糖:
[0272]
葡萄糖一水合物880g/l
[0273]
进料盐混合物:
[0274]
磷酸二氢钾21.6g/l
[0275]
七水硫酸镁24.24g/l
[0276]
硫酸钾8.4g/l
[0277]
硫酸钠0.672g/l
[0278]
准备说明:
[0279]
通过将组分溶解在milli-q水中并通过高压灭菌,在单独的bluecap瓶中制备批次和饲料盐混合物以及批次和饲料葡萄糖。
[0280]
进料溶液:
[0281]
通过将500ml饲料葡萄糖与500ml饲料盐混合物、10ml维生素混合物、10ml微量元素溶液和1ml微量元素溶液混合来制备饲料溶液。
[0282]
实施例1:在nicotiana benthamiana中的表达
[0283]
在农杆菌渗透后7天收获共表达感兴趣基因的特定组合(goi)的n.benthamiana叶片材料。将1ml甲醇(meoh)添加到2个叶盘(=2cm)中。萃取在室温下以200rpm的轨道振荡进行。使用0:22μm 96孔滤板(merck millipore,darmstadt,德国)过滤200μl提取物,并在4℃进行lc-ms分析。
[0284]
图1显示了获得的lcms配置文件。结果表明,用编码具有seq id no:1的酶的cyp82d274v1转化的n.benthamiana细胞导致产生14-oh-脱氢松香二烯;当用编码分别具有氨基酸序列seq id no:3和seq id no:4的酶的cyp71be85和cyp71be86进一步转化细胞时,形成雷酚内酯;并且当细胞进一步用编码具有seq id no:5的氨基酸序列的酶的cyp82d213转化时,形成雷公藤内酯酮。此外,可以看出,具有seq id no:6序列并由基因cyp82d217编码的酶增加雷酚内酯和雷公藤内酯酮的产量。
[0285]
实施例2:酿酒酵母菌株的构建。
[0286]
菌株构造
[0287]
母酵母菌株为酿酒酵母s288c(ncyc 3608;英国诺里奇酵母培养物国家保藏中心)。
[0288]
菌株的基因型和来源列于表3。
[0289]
使用乙酸锂转化法(8)制备构建的酵母菌株。没有功能ura3的亲本菌株通过以下程序变得合格:将甘油原液接种到5ml ypd培养基中,并在30℃o/n下生长。然后,将3ml o/n培养物转移到50mlypd培养基中,继续生长4-5小时,然后以4000rpm离心10分钟,然后丢弃上清液。然后,在无菌水中(第一次在25ml中,第二次在1ml中)洗涤2次后,将细胞准备好转化,并重新悬浮在0.4ml无菌水中。
[0290]
通过以下步骤转化有活力的酵母细胞:分别加入10μl有活力的noti消化质粒(各2μl)的混合物,并与60μl peg 3350(50%w/v)、9μl liac(1m)和12.5μl预浸鲑鱼精子dna(10mg/ml)混合。然后将所得混合物在42℃孵育40分钟,然后通过离心(3000rpm,5分钟)收集细胞并去除上清液。然后将细胞重新悬浮在100μl无菌水中,并在无尿嘧啶琼脂平板的sc上铺展。在30℃下孵育2天后,分离的转化体表现为单个菌落。使用表1中的基因和构建体特异性引物,通过菌落pcr确认基因构建体的插入。对于菌落pcr,将酵母菌落重新悬浮在50μl 20mm naoh中,并在99℃下孵育15分钟。将1μl菌落悬浮液用于pcr。
[0291]
表1:使用引物列表
[0292]
[0293][0294]
酿酒酵母基因组工程基因构建体的组装
[0295]
质粒名称和编码的基因构建体列于表2中。如前所述(5),通过user克隆产生所有质粒。此外,如前所述(6),为user克隆制备了名为装配器-1、-2和-3的亲本载体,用于同时整合多达六个基因构建体,并含有asisi/nb.bsmi user盒。用于与user兼容的pfux7聚合酶(7)进行pcr扩增的引物列于表1中。表3列出了本工作中使用和生成的载体。
[0296]
酿酒酵母的密码子优化基因来自美国旧金山twist biosciences。下表中前缀为“co_”的所有基因都经过密码子优化。使用与上述表1中描述的引物相同的引物扩增密码子优化的基因,不同之处在于引物被修改以适应密码子优化基因中任何核苷酸变化的杂交。用于密码子优化基因扩增的引物也公开于j.andersen-ranberg et al.,expanding the landscape of diterpene structural diversity through stereochemically controlled combinatorial biosynthesis.angewandte chemie international edition,n/a(2016)。
[0297]
表2:产生和使用的载体和质粒
[0298]
[0299][0300]
表3:使用和产生的酿酒酵母菌株列表
[0301]
[0302]
[0303][0304]
实施例3:酵母、酿酒酵母中的表达
[0305]
提取和代谢物分析
[0306]
将基因工程酿酒酵母菌株转移到96孔板中的0.5ml培养基中,并在30℃下以350rpm的轨道振荡培养3天。为了提取,将0.1ml酿酒酵母培养物转移到1.5ml玻璃瓶中。加入0.4ml metoh uhplc级。使用0:22μm 96孔滤板(merck millipore,darmstadt,德国)过滤酿酒酵母提取物提取物,并在lc-ms分析之前在4℃下储存。
[0307]
提取物的lcms谱可以在图2中看到,其中可以观察到用编码具有seq id no:1氨基酸序列的酶的twcyp82d274v1转化背景菌株导致14-oh-脱氢松香二烯的形成。
[0308]
实施例4:通过nmr分析检测14-oh-脱氢松香二烯
[0309]
通过nmr分析实施例3中鉴定为14-oh-脱氢松香二烯的化合物以确认其特性。
[0310]
核磁共振纯化
[0311]
工程酵母中雷公藤甲素中间体的纯化
[0312]
将产生所需目标化合物的工程酵母从sc琼脂接种到10ml ydp中,并在30℃条件下培养。将5ml on培养物接种在500ml fit培养基中,并在30℃下生长5天。用500ml etac从培养物中提取目标化合物。通过转子蒸发除去溶剂,并将分析物重新悬浮在己烷中。重复提取3次。己烷提取物用于supelclean
tm
/na2so4spe管(sigma-aldrich)和分析物使用1:99-5:95etac:己烷的阶梯梯度从柱中洗脱。用lc-ms或gc-ms分析每个馏分,并选择含有感兴趣化合物的馏分进行nmr分析。
[0313]
nmr分析
[0314]
nmr数据在配备有针对13c和1h优化的5mm低温冷却dch探针的bruker avance iii hd 600mhz nmr光谱仪(1h工作频率599.85mhz)上获得(bruker biospin,karlsruhe,germany)。nmr数据记录在cdcl3(euriso top,99.8原子%d)中的5mm管中,温度平衡至300k,锁定参数优化,梯度匀场,接收器增益设置,所有这些都由topspin 3.2版和iconnmr 4.7.5版(德国卡尔斯鲁厄bruker biospin)自动控制。1h和13c化学位移分别参考ph 7.26ppm和pc77.16ppm的残留溶剂信号。用30
°
脉冲和64k数据点以及零填充到256k数据点,以12khz的谱宽、1s的弛豫延迟和2.7s的采集时间采集1h光谱。使用waltz-16复合脉冲去耦方案对13c光谱进行1h去耦。使用4096(hmbc)、2048(dqf-cosy和roesy)或1024(多重编辑的hsqc)数据点(直接维度)和256(dqf-cosy、hmbc和roesy)或128(多重编辑hsqc)个数据点(间接维度)获取2d同核和异核实验。2d nmr数据在f1中被零填充到1k,在f2中被零填补到两倍的点数,在f1中采用正向线性预测(lpbin=0)。nmr数据的处理使用topspin 4.0.9版(德国卡尔斯鲁厄bruker biospin)完成。
[0315]
14-oh-脱氢松香二烯的nmr光谱数据如表4所示。
[0316]
表41(14-oh-脱氢松香二烯)的1h和
13
c nmr光谱数据
[0317]
[0318][0319]
a1
h nmr(599.85mhz)和
13
c nmr(150.83mhz)数据获得于cdcl3.bnh=氢的数量。多重性报告为明显分裂:s=单重,d=双重,t=三重,sep=七重,m=多重(也适用于重叠),br
=宽a’表示最高化学位移值,b’表示最低化学位移值。
[0320]
图3显示了在599.85mhz下cdcl3中14-oh-脱氢松香二烯的1h nmr谱,图4显示了在150.83mhz下cdcl3中14-oh-脱氢松香二烯的
13
c nmr谱,证实了该化合物的同一性。
[0321]
实施例5:导致产生雷酚内酯和雷公藤内酯酮的基因在酿酒酵母中的表达
[0322]
这是一项初步研究,旨在评估导致啤酒酵母产生雷酚内酯和雷公藤内酯酮的基因表达的影响。
[0323]
用各自含有单独二萜生物合成基因或雷公藤cyps(twcyps)的载体进一步转化实施例2中产生的背景酵母菌株
[0324]
提取物的lcms谱可以在图5中看到,其中可以看到,用编码分别具有seq id no:1、seq id no:3和seq id no:4氨基酸序列的酶的twcyp82d274v1、twcyp71be85和twcyp71be 86转化背景菌株;导致雷酚内酯的形成;并且用编码具有seq id no:5的氨基酸序列的酶的cyp82d213进一步转化导致雷公藤内酯酮的形成。
[0325]
图6显示了在生成的转化体的提取物中检测到的含氧二萜化合物的含量概况。
[0326]
左侧面板显示雷酚内酯和雷公藤内酯酮的含量,右侧面板显示了14-oh-脱氢松香二烯的含量。表达编码具有seq id no:8的氨基酸序列的酶的基因twb5#1导致显著更高的雷酚内酯和雷公藤内酯酮的产生。
[0327]
twb5#2-6基因是不增加雷酚内酯或雷公藤内酯酮产生的其他雷公藤细胞色素b5基因(未提供序列)。
[0328]
实施例6:在酿酒酵母和n.benthamiana中生产含氧二萜化合物
[0329]
按照上文“材料和方法”一节所述,培养所有工程酿酒酵母菌株和本氏芽胞杆菌。类似地,提取二萜类代谢物,通过lc-ms、gc-ms和nmr进行分析,并如上所述进行定量。
[0330]
优选实验生物体是酵母,并且异源基因已经在生物体中稳定转染,因为这给出了最精确和可重复的结果。
[0331]
结果如图7-9所示。从图中可以清楚地看出,不同于酵母细胞和烟草植物的生物都能够根据本发明的方法以高滴度生产所提出的雷公藤内酯酮生物合成途径中所要求的关键中间体。
[0332]
图10-26显示了其他关键化合物的核磁共振波谱。所产生的化合物的nmr光谱数据显示在下表5-21中。
[0333]
表5.f1-14的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0334][0335]
表6.f1-15的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0336][0337]
表7.f1-18的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0338]
[0339][0340]
表8.f1-23(f15p1)的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0341][0342]
表9.f1-31的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0343]
[0344][0345]
表10.f2-x的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0346][0347]
表11.f2-10的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0348][0349]
表12.f20p1的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0350]
[0351][0352]
表13.f20p2的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0353][0354]
表14.f20p3的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0355]
[0356][0357]
表15.f20p4的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0358][0359][0360]
表16.f15p2的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0361][0362]
表17.f55p2的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0363]
[0364][0365]
表18.f55p3的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0366][0367]
5表19.f15p4的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0368][0369][0370]
表20.f20p5的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0371][0372]
表21.f60p1的1h和
13
c nmr数据和2d hmbc和roesy相关性
[0373][0374]
a1
h nmr(600.13)和
13
c nmr(150.90mhz)数据获得于cdcl3中的样品。b基于hsqc和
hmbc实验的分配。c以明显分裂形式报告的多重性:s=单重态,d=双重态,t=三重态,sext=六重态,m=多重态(包括重叠共振),br=宽。α表示me指向平面,并且β表示我指向平面外。a表示最低化学位移值,b表示最高化学位移值
[0375]
参考文献
[0376]
1.j.andersen-ranberg et al.,expanding the landscape of diterpene structural diversity through stereochemically controlled combinatorial biosynthesis.angewandte chemie international edition,n/a(2016).
[0377]
2.i.pateraki et al.,total biosynthesis of the cyclic amp booster forskolin from coleus forskohlii.elife 6,e23001(2017).
[0378]
3.i.pateraki et al.,manoyl oxide(13r),the biosynthetic precursor of forskolin,is synthesized in specialized root cork cells in coleus forskohlii.plant physiology 164,1222-1236(2014).
[0379]
4.n.l.hansen et al.,the terpene synthase gene family in tripterygium wilfordii harbors a labdane-type diterpene synthase among the monoterpene synthase tps-b subfamily.the plant journal 89,429-441(2017).
[0380]
5.h.h.nour-eldin,b.g.hansen,m.h.h.j.k.jensen,b.a.halkier,advancing uracil-excision based cloning towards an ideal technique for cloning pcr fragments.nucleic acids research 34,e122(2006).
[0381]
6.n.b.jensen et al.,easyclone:method for iterative chromosomal integration of multiple genes in saccharomyces cerevisiae.fems yeast research 14,238-248(2014).
[0382]
7.m.h.h.a mutant pfu dna polymerase designed for advanced uracil-excision dna engineering.bmc biotechnology 10,21(2010).
[0383]
8.r.d.gietz,r.h.schiestl,high-efficiency yeast transformation using the liac/ss carrier dna/peg method.nature protocols 2,31-34(2007).
[0384]
9.hansen,n.l.,et al.,integrating pathway elucidation with yeast engineering to produce polpunonic acid the precursor ofthe anti-odesity agent celastrol.microd cell fact,2020.19(1):p.15.
[0385]
10.voinnet o,rivas s,mestre p,baulcombe d.an enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus[retracted in:plant j.2015 nov;84(4):846].plant j.2003;33(5):949-956.doi:10.1046/j.1365-313x.2003.01676.x
[0386]
项目
[0387]
1.一种能够产生含氧二萜化合物的重组宿主细胞,其中所述宿主细胞能够产生松香烷型二萜烯和/或脱氢松香二烯,并且已经用编码具有细胞色素p450活性的酶的第一基因转化,该酶能够将松香烷型二萜烯和/或脱氢松香二烯转化为14-羟基脱氢松香二烯。
[0388]
2.根据项目1所述的重组宿主细胞,其中所述第一基因编码多肽,所述多肽包含与seq id no:1(twcyp82d274v1)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性、优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序
列、或其成熟多肽。
[0389]
3.根据项目1或2所述的重组宿主细胞,其中所述重组宿主细胞进一步包含:
[0390]
编码具有细胞色素p450活性的第二酶的第二基因和编码具有细胞色素p450活性第三酶的第三基因,其中:
[0391]
所述第二基因编码多肽,所述多肽包含与seq id no:4(twcyp71be86)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性、优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列、或其成熟多肽;
[0392]
所述第三基因编码多肽,所述多肽包含与seq id no:3(twcyp71be85)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性、优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列、或其成熟多肽。
[0393]
4.根据项目3所述的重组宿主细胞,其中所述宿主细胞还包含编码具有细胞色素b5活性的多肽的基因,所述多肽包含与seq id no:8(twb5#1)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性、优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列、或其成熟多肽。
[0394]
5.根据项目3或4所述的重组宿主细胞,其中所述重组宿主细胞进一步包含:
[0395]
编码具有细胞色素p450活性的第四酶的第四基因,其中:
[0396]
所述第四基因编码多肽,所述多肽包含与seq id no:5(twcyp82d213)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性、优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列、或其成熟多肽。
[0397]
6.根据项目5所述的重组宿主细胞,其中所述重组宿主细胞进一步包含:
[0398]
编码具有细胞色素p450活性的第五酶的第五基因和编码具有细胞色素p450活性第六酶的第六基因,其中:
[0399]
所述第五基因编码多肽,所述多肽包含与seq id no:6(twcyp82d217)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性、优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列、或其成熟多肽;
[0400]
所述第六基因编码多肽,所述多肽包含与seq id no:7(twcyp82d275)具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性、优选至少95%序列同一性、更优选至少98%序列同一性的氨基酸序列、或其成熟多肽。
[0401]
7.根据前述项目中任一项所述的重组宿主细胞,其中能够产生松香烷型二萜烯和/或脱氢松香二烯的宿主细胞是用一个或多个编码以下基因的基因转化的重组细胞:
[0402]
a双(牻牛儿基)二磷酸合酶;
[0403]
b能够将ggpp转化为松香烷型二萜烯的二萜合酶;
[0404]
c联合能够ggpp转化为松香烷型二萜烯的两种或多种二萜合酶的组合;或
[0405]
d柯巴基焦磷酸合酶和松香烷型二萜烯合酶。
[0406]
8.根据项目7所述的重组宿主细胞,其中所述双(牻牛儿基)二磷酸合酶是包含seq id no:73或seq id no:81的氨基酸序列的多肽。
[0407]
9.根据项目7所述的重组宿主细胞,其中能够将ggpp转化为松香烷型二萜烯的两种或多种二萜合酶的组合是包含seq id no:67的氨基酸序列的多肽和包含seq id no:68的氨基酸序列的组合;或是包含seq id no:69的氨基酸序列的多肽和包含seq id no:70的
氨基酸序列的组合。
[0408]
10.根据项目7所述的重组宿主细胞,其中柯巴基焦磷酸合酶和松香烷型二萜烯合酶的组合是包含seq id no:71的氨基酸序列的多肽和包含seq id no:72的多肽肽的组合。
[0409]
11.根据前述项目中任一项所述的重组宿主细胞,其中所述重组宿主细胞选自原核细胞或真核细胞。
[0410]
12.根据项目11所述的重组宿主细胞,是选自大肠杆菌、芽孢杆菌、乳杆菌和棒状杆菌的原核细胞。
[0411]
13.根据项目11所述的重组宿主细胞,是选自酿酒酵母、scizosaccha romyces、klyveromyces、pichia、candida和yarrowia物种的真核细胞。
[0412]
14.根据项目11所述的重组宿主细胞,其中所述细胞是酿酒酵母细胞。
[0413]
15.根据项目中任一项所述的重组宿主细胞用于制备含氧二萜化合物中的用途。
[0414]
16.权利要求15的用途,其中所述含氧二萜化合物选自14-oh-脱氢松香二烯、雷酚内酯和雷公藤内酯酮。
[0415]
17.如权利要求16所述的用途,其中所述含氧二萜化合物是雷公藤内酯酮,其中雷公藤内酯酮进一步转化为雷公藤甲素。
[0416]
18.根据权利要求15至17中任一项所述的用途,其中使用一个或多个分离和/或色谱步骤回收所述含氧二萜化合物。
[0417]
19.具有细胞色素p450酶活性的多肽,所述多肽包含与序列seq id no:1、seq id no:2、seq id no:3、seq id no:4、seq id no:5、seq id no:6、seq id no:7、seq id no:8中的一种具有至少80%序列同一性、优选至少85%序列同一性、优选至少90%序列同一性,优选至少95%序列同一性、优选至少96%序列同一性、优选至少97%序列同一性、优选至少98%序列同一性、或甚至100%序列同一性的氨基酸序列,或其成熟多肽。
[0418]
20.编码项目19所述的多肽的多核苷酸。
[0419]
21.包含项目20所述的多核苷酸的质粒、表达载体、表达构建体或重组宿主细胞。
[0420]
22.化合物14-oh-脱氢松香二烯。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。