西瓜clmir159-3p及其前体基因的应用
技术领域
1.本发明属于分子生物学技术领域,具体涉及西瓜clmir159-3p及其前体基因在提高植物耐冷性中的应用。
背景技术:
2.西瓜原产非洲热带地区,喜高温干燥,不耐低温。冬春低温不仅因能源投入增加设施西瓜的经济成本,而且易导致其光合速率下降、根系生长受阻,甚至植株死亡,进而影响产量和品质。
3.植物在长期的进化过程中,形成了对各种逆境条件的适应或抗性调控机制,从而在低温逆境条件下完成其生长发育历程。例如,植物感知低温后,以cbf 依赖型(c-repeat binding transcription factor)和非cbf依赖型的调控途径激活一个冷适应过程。cbf依赖型调控途径研究的较为深入,是c-repeat(crt)bindingfactor/dehydration-responsive element(dre)binding factor(cbf/dreb)介导的转录调控级联,它对诱导一系列冷响应基因(cor)至关重要。在这个途径中, cbfs/drebs会被低温迅速诱导,cbfs/drebs可以结合到cor基因启动子区域中的crt/dre顺式元件并激活它们的转录。作为现在研究比较清楚的的cbf转录激活子,ice1(myc型bhlh转录因子)在低温下可以直接结合cbf基因启动子激活它们的表达。sumoe3连接酶siz1和泛素e3连接酶hos1能够改变 ice1的sumo化和泛素化并影响ice1蛋白的稳定性和活性。低温还能够激活 snrk2.6/ost1,活化的snrk2.6通过磷酸化修饰ice1从而打破ice1与hos1的互作,抑制ice1降解,激活下游cbf-cor基因表达。然而,在以追求产量和品质为目的的长期人工驯化过程中,许多栽培品种的抗逆基因功能缺失或减弱,进而导致其抗性降低。因此,挖掘抗冷基因并验证其功能,对于耐冷植物新品种的选育具有重要意义。
技术实现要素:
4.有鉴于此,本发明提供了西瓜clmir159-3p及其前体基因clmir159在调控植物cbfs基因表达及低温抗性中的应用,通过对西瓜clmir159-3p前体基因 clmir159的克隆以及利用转基因技术构建过表达clmir159植物材料,可有效调控植物耐冷性。
5.为了实现上述目的,本发明采用如下技术方案:
6.西瓜clmir159-3p,核苷酸序列如seq id no.1所示。
7.西瓜clmir159-3p的前体clmir159,核苷酸序列如seq id no.2所示。
8.进一步地,构建携带clmir159的重组载体或重组菌。
9.西瓜clmir159-3p或clmir159或上述重组载体或上述重组菌可用于提高植物cbfs基因表达。
10.进一步地,通过过表达西瓜clmir159-3p或clmir159提高植物cbfs基因表达。
11.西瓜clmir159-3p或clmir159或上述重组载体或上述重组菌可用于提高植物低温抗性。
12.进一步地,通过过表达西瓜clmir159-3p或clmir159提高植物低温抗性。
13.西瓜clmir159-3p或clmir159或上述重组载体或上述重组菌可用于分子育种。
14.利用靶基因预测、降解组测序和qrt-pcr技术,鉴定出clmir159-3p的靶基因为cla97c08g151350(myb33)、cla97c11g224710(sporocyteless-like)。
15.经由上述的技术方案可知,与现有技术相比,本发明公开提供了西瓜 clmir159-3p及其前体基因clmir159在调控植物低温抗性中的应用,通过基因手段构建拟南芥异源过表达clmir159材料,调控clmir159-3p的表达水平发现, clmir159过表达可提高植物cbfs基因表达,提高植物的低温抗性。利用靶基因预测结合降解组测序和qrt-pcr技术,鉴定出clmir159-3p的靶基因为 cla97c08g151350(myb33)、cla97c11g224710(sporocyteless-like)。本发明提供的clmir159-3p/clmir159为创制耐冷植物种质及培育耐冷新品种提供了基因资源,具有较好的潜在应用价值。
附图说明
16.图1为本发明2种不同西瓜嫁接材料的耐冷性鉴定;
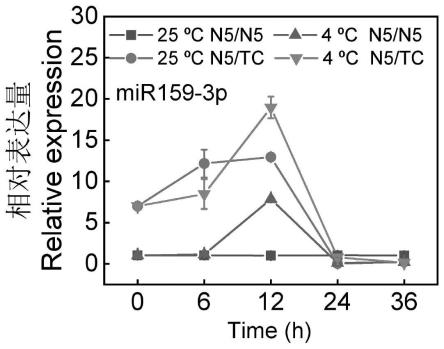
17.图2为本发明低温胁迫下耐冷和不耐冷西瓜材料中clmir159-3p的 qrt-pcr检测结果;
18.图3为本发明异源过表达拟南芥中clmir159的半定量pcr检测结果;
19.图4为本发明clmir159异源过表达拟南芥植株的低温抗性鉴定;
20.图5为本发明clmir159异源过表达拟南芥中cbfs基因的qrt-pcr 检测结果;
21.图6为本发明低温胁迫下耐冷和不耐冷西瓜材料中clmir159-3p靶基因cla97c08g151350(myb33)、cla97c11g224710(sporocyteless-like) 的qrt-pcr检测结果。
具体实施方式
22.下面对本发明实施例中的技术方案进行清楚、完整地描述,显然,所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有做出创造性劳动前提下所获得的所有其他实施例,都属于本发明保护的范围。
23.实施例1耐冷和不耐冷西瓜材料中clmir159-3p对低温胁迫的响应
24.为了改变西瓜的耐冷性,以西瓜栽培品种农科大5号(n5)为接穗,以耐冷野生西瓜种质tc为砧木,采用劈接的方法进行嫁接;并且设置自根嫁接的对照组n5/n5。嫁接苗长至四叶一心时,分别进行常温和低温处理,处理组合为: n5/n5 25℃、n5/tc 25℃、n5/n5 4℃、n5/tc 4℃。
25.各处理组处理36h后,分别测定净光合速率、psii最大光化学效率、丙二醛含量。结果如图1所示,4℃低温处理36h后,n5/tc植株的净光合速率和 fv/fm显著高于n5/n5植株,而其丙二醛(mda)含量显著低于n5/n5,表明 n5/tc植株的耐冷性高于n5/n5。
26.分别采集低温处理后第0、6、12、24、36h的叶片用液氮速冻后储存于-8 0℃超低温冰箱,用于clmir159-3p的测定,西瓜clmir159-3p的核苷酸序列如seq id no.1所示:uuuggauugaagggagcucua。
27.用mirna试剂盒(tiangen,beijing)提取西瓜叶片的mirna,将提取完成的mirna用
mircut plus mirna first-strand cdna kit进行反转录。 clmir159-3p扩增引物序列见表1,使用u6作为内参引物。使用mircute增强型mirna荧光定量检测试剂盒(sybrgreen)进行mirna荧光定量检测。
28.20μl反应体系中包括10μl sybr荧光染料、1μl cdna、0.4μl上游引物、0.4μl下游引物、8.2μl ddh2o。pcr反应条件为:95℃15min,94℃变性20 s、60℃退火延伸34s,40个循环。
29.表1qrt-pcr引物序列
[0030][0031]
结果见图2,正常温度条件和低温胁迫下,n5/tc植株clmir159-3p的相对表达量在0-24h显著高于n5/n5。低温胁迫下,n5/n5和n5/tc嫁接植株中的clmir159-3p均从第6h开始上调表达,在12h达到峰值,随后迅速下降,在24h恢复到初始水平。低温处理12h时,n5/tc植株clmir159-3p的相对表达量比n5/n5高1.43倍。上述结果表明,clmir159-3p可能参与耐冷西瓜材料对低温的响应与适应。
[0032]
实施例2异源过表达clmir159转基因拟南芥材料的构建与检测
[0033]
(1)查找西瓜基因组数据库,显示clmir159基因位于cla97chr04染色体的25169051-25169256bp,基因序列如下:
[0034]
tagggtttgaggcagagctccttgaagtccaatagaggtttatgccaggttgattaagctgctaagctatggatcccacagcccctattgctaatctcataaatcaaaattaagctaataggcctgtggcttgcatatctcaggagcttcatcacctgtttttg ttagatccttgtttggattgaagggagctctacatcttctct;seq id no.2。
[0035]
按照构建mirna前体的要求,在clmir159基因所在位置上下游各延长约 150bp,即基因组中定位于4号染色体25168917-25169399bp的片段,得到如下序列:
[0036][0037]
序列中黑体加粗序列为与上下游引物匹配的序列部分;单下划线序列为 clmir159对应的基因组序列部分;双下划线序列为成熟clmir159-3p所对应的基因组序列部分。
[0038]
(2)pcr扩增目的基因
[0039]
从西瓜栽培品种
‘
农科大5号’(n5)的cdna中利用pcr技术将上述片段扩增下来,所用pcr引物为:(5’端分别带有限制性内切酶bamhi和ecori 的识别序列)
[0040]
ttttgctcttc-3’;seq id no.4;
[0041]
mir159-r:5
’‑
atcccccgggctgcaggaattcgatatc-ggttaagaaaa tgggataatggg-3’;seq id no.5;
[0042]
pcr扩增得到片段长度为539bp、含有pgreen载体同源臂的clmir159 片段。20μl反应体系中包括10μl高保真酶、1μl cdna、0.5μl上游引物、0.5μl 下游引物、8μl ddh2o。pcr反应条件为:98℃10s,55℃变性5s、72℃退火延伸35s,36个循环。
[0043]
(3)将步骤(2)扩增获得的clmir159片段与xhoi和ecorv限制性内切酶双酶切过的pgreen线性化载体进行连接,获得重组质粒 pgreen-clmir159。
[0044]
(4)将pgreen-clmir159通过热激法转化根癌农杆菌菌株gv3101。
[0045]
(5)将含有pgreen-clmir159质粒的农杆菌摇菌至od为1.5-2.0,用蘸花法侵染没有开放的拟南芥花序。收获的t0代拟南芥利用basta筛选抗性植株。采集野生型columbia和具有basta抗性植株的叶片提取dna,利用pcr 鉴定clmir159过表达阳性转基因植株(oe#1-#3),pcr所用引物序列如下:
[0046]
mir159-2f:5
’‑
catttggagaggacagccca-3’;seq id no.6;
[0047]
mir159-2r:5
’‑
ctccagtgaaaagttcttctcc-3’;seq id no.7;
[0048]
20μl反应体系中包括10μl taq酶、1μl cdna、0.5μl上游引物、0.5μl下游引物、8μl ddh2o。pcr反应条件为:94℃3min,94℃变性30s、60℃退火 30s,72℃延伸40s,35个循环,72
℃5min。
[0049]
结果见图3,野生型没有clmir159的dna条带,而过表达株系oe#1-oe#3 有clmir159的dna条带。
[0050]
获得的阳性clmir159基因过表达株系进行单株收种,并连续自交3代进行纯化。
[0051]
实施例3clmir159异源过表达拟南芥植株的低温抗性鉴定
[0052]
将clmir159异源过表达拟南芥种子消毒后播至1/2ms培养基中正常培养,培养至拟南芥4片真叶。对野生型拟南芥和clmir159过表达植株进行-10℃低温处理1.5h,然后正常温度条件下恢复5d;并设置未经低温处理的对照组。
[0053]
结果见图4,在正常情况下,野生型拟南芥和过表达拟南芥oe#1、oe#2、 oe#3植株表型基本无差异。低温处理后,野生型拟南芥植株矮小,并且有一部分枯死,叶片大部分枯黄;而oe#1植株较大,叶片微有枯黄,植株死亡较少; oe#2和oe#3拟南芥植株叶片微有枯黄,植株死亡较少。
[0054]
在正常条件下,野生型拟南芥和过表达拟南芥oe#1、oe#2、oe#3成活率都为100%,三个株系的相对电导率基本无差异。低温处理后,过表达拟南芥成活率明显高于野生型拟南芥,并且三个株系oe#1、oe#2、oe#3分别比wt高 33.84%、46.84%、27.65%;过表达拟南芥相对电导率明显低于野生型拟南芥, oe#1、oe#2、oe#3分别比wt降低了19.08%、15.35%、22.44%。
[0055]
上述结果说明,过表达clmir159可以提高拟南芥的低温抗性。
[0056]
实施例4拟南芥中clmir159异源过表达对cbfs基因表达的调控
[0057]
为了探究拟南芥中clmir159异源表达对cbfs基因表达的调控,低温处理后0h、1h、12h、1d、3d、5d取野生型拟南芥和过表达拟南芥叶片。用rna 提取试剂盒(tiangen,beijing)提取拟南芥叶片的总rna。总rna使用fastkingcdna第一链合成试剂盒(tiangen,beijing)反转成cdna。
[0058]
atcbfs基因的扩增引物序列的见表1;以atactin为内参基因。
[0059]
荧光定量pcr利用美国bio-rad公司的iq5实时定量仪进行分析,并使用extaqtm ii(2
×
)的试剂盒(takara,tokyo,japan)。20μl反应体系中包括10μl sybr荧光染料、1μl cdna、1μl上游引物、1μl下游引物、 7μl ddh2o。pcr反应条件为:95℃3min;95℃变性30s、58℃退火30s、 72℃延伸30s,40个循环。荧光定量pcr得到的ct值采用2-δδct
法对cbf相对基因的表达水平进行计算。
[0060]
由图5可知,低温处理后,wt和oe#1、oe#2、oe#3叶片中atcbf1、 atcbf2、atcbf3的表达量均上调表达,且三者表达量的变化趋势基本相同。 wt和oe#2、oe#3叶片中atcbf1、atcbf2、atcbf3的表达量均在低温1h 后达到峰值,oe#2则在12h最高。相比于野生型拟南芥,oe#2在1h时叶片 atcbf1、atcbf2、atcbf3表达量分别提高了97.21%、80.27%、47.88%,oe#3 则分别提高了76.28%、48.55%、102.23%。
[0061]
上述结果表明,clmir159过表达诱导的低温抗性与其调控cbfs基因表达有关。
[0062]
实施例5西瓜clmir159-3p的靶基因鉴定
[0063]
为了确定clmir159-3p的靶基因,通过psrnatarget网站 (https://www.zhaolab.org/psrna target/)和降解组测序鉴定clmir159-3p的靶基因。
[0064]
由图6可知,靶基因的预测剪切位点位于clmir159-3p的第10位点。以β
ꢀ‑
actin为
内参基因,通过qrt-pcr分析可知,cla97c08g151350(myb33)、 cla97c11g224710(sporocyteless-like)靶基因表达趋势基本与clmir159-3p 表达相反,表明这2个基因可能受clmir159-3p调控。
[0065]
本说明书中各个实施例采用递进的方式描述,每个实施例重点说明的都是与其他实施例的不同之处,各个实施例之间相同相似部分互相参见即可。
[0066]
对所公开的实施例的上述说明,使本领域专业技术人员能够实现或使用本发明。对上述实施例的多种修改对本领域的专业技术人员来说将是显而易见的,本文中所定义的一般原理可以在不脱离本发明的精神或范围的情况下,在其它实施例中实现。因此,本发明将不会被限制于本文所示的这些实施例,而是要符合与本文所公开的原理和新颖特点相一致的最宽的范围。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。