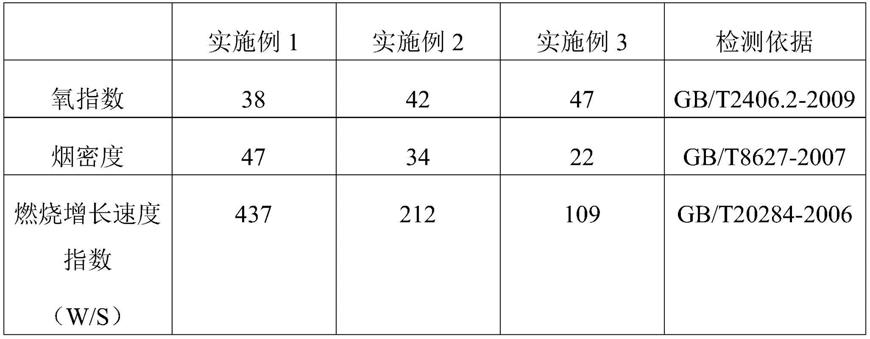
用于去抑制re1沉默转录因子靶基因的组合物和方法1.相关申请的交叉引用及优先权要求2.本技术要求2019年11月22日提交的美国临时专利申请no.62/939,149和2020年10月1日提交的美国临时专利申请no.63/086,248的优先权,它们公开的内容通过引用并入本文。
技术领域:
:3.本发明提供了用于去抑制re1沉默转录因子(rest)靶基因的方法、化合物和组合物。具体而言,本发明公开了一种具有序列tedleppepplpken(seqidno:1)和edleppepplpk(seqidno:15)的肽,或由d-氨基酸nekplppeppeldet(seqidno:16)和kplppeppelde(seqidno:17)组成的反向序列(反向倒置,retroinverted,ri)的肽,用于抑制rest活性。这些肽可用于治疗、预防或改善诸如创伤性脑损伤、癫痫、痴呆、亨廷顿病(hd)、慢性疼痛、脑癌(包括多形性胶质母细胞瘤)、胰腺癌;糖尿病和周围神经损伤等病变。
背景技术:
::4.阻遏因子1(re1)沉默转录因子(rest)是数百种神经元基因的阻遏因子1。其靶点代表终末分化神经元细胞表型所需的基因,包括编码电压和配体依赖性离子通道的基因、其受体、生长因子和轴突导向蛋白(bruceaw等人,procnatlacadsciusa101,10458-10463(2004);conacoc等人,procnatlacadsciusa103,2422-2427(2006);mortazavia等人,genomeres16,1208-1221(2006);和ottosj等人,jneurosci27,6729-6739(2007);所有这些都通过引用并入本文)。因此,在神经发生过程中,rest逐渐下调,以使成熟神经元表型得以确立(ballas等人,2005,上文(supra))。这一事件的重要性通过功能获得研究得到证实,这些研究表明,rest的持续存在会阻碍终末神经元分化(mandelg等人,procnatlacadsciusa108,16789-16794(2011)和gaoz等人,jneurosci31,9772-9786(2011);两者均通过引用并入本文)。5.对rest的转录或转录后调节知之甚少(ballasn等人2005,上文;ballasn等人,neuron31,353-365(2001);和kojimat等人,brainresmolbrainres90,174-186(2001);所有这些都通过引用并入本文)。然而,rest上的两个磷酸化位点(丝氨酸861和864)通过与c端结构域小磷酸酶-1(ctdsp1)的相互作用调节神经元分化2。当ctdsp1从丝氨酸861和864中去除磷酸盐时,rest蛋白被稳定,神经元分化受到抑制2-4。技术实现要素:6.本文公开了用于开发对ctdsp1具有高亲和力的肽的方法、化合物和组合物。7.本文公开了用于结合c端结构域小磷酸酶1(ctdsp1)的方法、化合物和组合物。8.本文公开了结合ctdsp1以抑制rest活性的rest类磷酸肽tedleppepplpken(seqidno:1)、edleppepplpk(seqidno:15)、nekplppeppeldet(seqidno:16)和kplppeppelde(seqidno:17)。小写字母代表d-氨基酸,其抗降解,因此增加了肽半衰期,而不影响结合亲和力5。9.本文公开了九十八(98)种rest类磷酸肽变体(rppv)(seqidno:18至117),其在不同程度上抑制rest上的ctdsp1活性(图25)。10.本文公开了细胞内转运肽,例如细胞穿透肽(cpp)和/或内涵体释放序列(seqidno:118至137和140至159),其n端或c端可融合至rpp(seqidno:1和15至17)或rppv(seqidno:18至117)以改善细胞内转运。还公开了可插入rpp或rppv与表7中所列肽之一之间以进一步改善细胞内转运的接头(seqidno:138、139、160和161)。11.本文公开的了n端或c端融合到细胞内转运肽(seqidno:118至137和140至159)并环化以进一步提高结合亲和力和稳定性(增加肽半衰期)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)。环化融合蛋白的示例为seqidno:2、5、12、13和14。12.本文公开了n端或c端融合到细胞内转运肽(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至137和140至159),促进rest蛋白质的降解。融合蛋白的示例为seqidno:2和4至14。13.本文公开了n端或c端融合到细胞内转运序列(seqidno:118至137和140-159)的如seqidno:1和15至17的rest类磷酸肽或seqidno:18至117的rppv,促进rest靶基因的表达。融合蛋白的示例为seqidno:2、4至14。14.本文公开了n或c端融合到细胞内转运肽(seqidno:118至137和140至159)的如seqidno:1和15至17的rpp或rppv(seqidno:18至117),通过向动物施用有效治疗量的化合物,以增加rest靶基因的表达或增加bdnf的表达,用于治疗患有与rest或ctdsp1相关疾病或病变的动物。融合蛋白的示例为seqidno:2和4至14。15.本文公开了融合到细胞内转运肽(seqidno:118至137和140至159)的rppseqidno:1和15至17或rppv(seqidno:18至117),可用于治疗动物的创伤性脑损伤、慢性疼痛、周围神经损伤、癫痫、糖尿病、阿尔茨海默病、亨廷顿病、脑肿瘤(包括多形性胶质母细胞瘤),或者胰腺癌。融合蛋白的示例为seqidno:2和4至14。附图说明16.随附附图包含在本说明书中,并构成本说明书的一部分。附图与上面给出的一般描述以及下面给出的示例性实施例和方法的详细描述一起,用于解释本发明的原理。在此类附图中:17.图1.(a)示出了ctdsp1结合位点位置的全长rest蛋白。(b)环状形式的与细胞穿透肽整合的反向倒置(ri)rest类磷酸肽。18.图2.使用monolith(nanotemper)测定其余类磷酸肽(rpp)对ctdsp1的结合亲和力。rpp(表4;seqidno:165)与ctdsp1(表2;seqidno:163)的亲和力约为130pm。用red-trisnta染料(20nm)荧光标记ctdsp1(40nm)上的his标签,并测定rpp(高达0.5μm)结合时荧光的变化,以评估结合。19.图3.使用biacore(gehealthcarelifesciences)测定rpp对ctdsp1的结合亲和力。将ctdsp1(表2;seqidno:163)固定在cm5传感器芯片(gehealthcarelifesciences)上,并测定(a)gst对照或(b)rpp(表4;seqidno:165)的结合亲和力。20.图4.rpp具有细胞渗透性,并在细胞核内聚集。(a-d)将100nmflag-rpp-cpp(caqkdykddddktedleppepplpkengrkkrrqrrrg(seqidno:4))添加到间充质祖细胞(mpcs)中并培养4小时。培养6天后,rpp集中在细胞核内。(e,f)在6天的时间内,肽从细胞质转移到细胞核(标准差,n=3)。(a)细胞骨架(phalloidin);(b)细胞核(hoechst染料);(c)rpp(flag抗体);(d)合成图像。21.图5.rpp积聚在脊髓坐骨神经核内。sprague-dawley(sd)雄性大鼠l3和l4区0.7cm的冠状脊髓切片,显示坐骨神经周围的细胞核,在大腿中部坐骨神经缺损损伤处注射1mg(1mg/ml)肽48小时后采集。a.hoechst染色显示细胞核。b.flag抗体示出了flag-rpp-cpp(seqidno:4)定位。22.图6.rpp降低rest蛋白水平。(左)示出了在用溶剂或100nmflag-rpp-cpp(seqidno:4)处理4小时后hek293细胞中外源性表达的rest蛋白水平的蛋白免疫印迹。用抗rest(milliporesigma,07-579)和抗gapdh检测印迹。(右)示出了100nmflag-rpp-cpp使rest蛋白水平降低58%的条形图。通过使用imagej测定凝胶峰面积来计算条带强度。23.图7.rpp增加bdnf表达。在培养24小时后,用flag-rpp-cpp(seqidno:4,100nm)或溶剂(pbs)培养4小时,用qpcr测定hek293细胞中的mrna水平。bdnf(2-ddct)表达的平均倍数变化为2.401±0.885(标准差,n=3)。平均倍数变化1=无变化。根据livak等,2001的描述计算2-ddct。24.图8.ctdsp1抑制后48小时,nbfl细胞中kv4.3的mrna水平。用表达带有gfp( )的rpp(seqidno:1)的质粒或带有gfp(-)的对照肽转染细胞。转染24小时后,根据荧光强度对细胞进行分选。误差棒表示标准偏差,n=2。25.图9.线性和环化rpp的比较。nbfl细胞被施用(1μm)rpp(线性_seqidno:4;环化_seqidno:2,表8)或对照肽(seqidno:3,表9)24或48小时。通过测定(a)bdnf、(b)ngf和(b)kv4.3转录水平的变化来评估疗效。n=1。26.图10.两名患者在用水(对照)或3μmrpp(seqidno:9)孵育48小时后,间充质祖细胞(mpc)中bdnf和ngfmrna的表达(归一化为肌动蛋白)。mrna水平相对于对照组示出(标准偏差,n=2)。27.图11.给ipsc(神经干细胞(nsc)-nl5)施用一条如下所示的rpp序列(3μm)或水对照。培养7天后,使用微管相关蛋白2(map2)神经元标记物(归一化为dapi)评估细胞的神经元分化。第3天更换培养基和变体。条形图中,map2水平为相对于对照(标准偏差n=4)。[0028][0029]图12.ipsc(神经干细胞(nsc)-nl5)采用1μmrpp(seqidno:9)或水对照。培养7天后,使用神经元标志物tuj1(iii类β-微管蛋白)和map2(均归一化为dapi)评估细胞的神经元分化。第3天更换培养基和rpp。(标准偏差,n=6)。[0030]图13.ipsc(神经干细胞(nsc)-nl5)采用1μmrpp(seqidno:13)或水对照。培养7天后,使用神经元标志物tuj1和map2(均归一化为dapi)评估细胞的神经元分化。第3天更换培养基和rpp。(标准偏差,n=6)。[0031]图14.rpp(seqidno:12)增加kv4.3、kv7.2、nav1.8和oprm1的表达。在成年sprague-dawley(sd)大鼠l5drg与不同浓度的rpp孵育48小时后,相对于水对照,对mrna表达进行qrt-pcr分析。*=p《0.05,***=p《0.01,***=p《0.001,单向方差分析和dunnett检验,误差棒=平均值的标准误差。[0032]图15.3μmrpp持续48小时(seqidno:13和14)可增加nav1.8的表达。rpp与成年sprague-dawley(sd)大鼠l5drg孵育后,对相对于水对照组的mrna表达进行qrt-pcr分析(归一化为β-肌动蛋白)。误差棒=平均值的标准误差。[0033]图16.rpp(seqidno:12)增加bdnf和ngf的表达。在成年sprague-dawley(sd)大鼠l5drg神经元与不同浓度的rpp孵育48小时后,对相对于水对照组的mrna表达(归一化为b-肌动蛋白)进行qrt-pcr分析。*=p《0.05,单因素方差分析和dunnett检验,误差棒=sem。[0034]图17.rpp(seqidno:12)不会导致drg神经元坏死。在用水、rpp(1、3或10μm)或triton培养48小时的sd大鼠中进行ldh毒检测。***=p《0.001,单因素方差分析和dunnett检验,误差棒=sd。[0035]图18.sni后慢性疼痛基因的rna水平。sni后第0天(假手术组,n=3)、第7天(n=5)或第28天(n=2)相对于β-肌动蛋白的rna倍数变化(平均值±sd,**p《0.01,***p《0.001,采用dunnett多重比较试验进行单因素方差分析。rna从坐骨神经drg(l5)中分离。[0036]图19.tbi后restmrna水平升高。(a)未受伤小鼠同侧皮质中restmrna表达的正常水平。(b)皮质控制性撞击伤后7天,小鼠同侧皮质的rest水平显著增加6。通过rnascope用dapi反染色示出的mrna。[0037]图20.(a)用于工程化ncrm-1ipsc的aavs1nanoluc-halotag敲入(ki)构建物示意图。将cmv驱动的构建物插入19q染色体的安全港(safeharbor)aavs1位点。通过转录激活物样效应器核酸酶(talens)。通过测序和连接pcr(未示出)证实aavs1ki。(b)用于工程化ncrm-1ipsc的map2nanoluc-halotag敲入(ki)构建物示意图。通过测序和连接pcr(未示出)证实,使用锌指核酸酶(zfn)和框架ki中的map2将纳米荧光素酶-halotag敲入map2转录起始位点(tss)(2号染色体)。cmv-nlht和map2-nlhtipsc表达多能性标记oct4/nanog/tra1-81/tra1-60(未示出)。(c)cmv-nlhtipsc和map2-nlht神经干祖细胞(nspc)和神经元的荧光素酶检测。注意,随着map2-nlht细胞的神经元分化,荧光素酶活性增加,这与随着分化,map2表达增加一致。[0038]图21.大鼠坐骨神经损伤模型:(左)坐骨神经切断手术过程。(右)坐骨神经切片免疫组化免疫荧光染色。对组织进行βiii微管蛋白、层粘连蛋白i和dapi染色。比例尺=100μm。[0039]图22.大鼠坐骨神经采集:(左)大鼠脊柱和相关周围神经的整体切片。(右)坐骨神经损伤部位与相关腰神经根的图像。[0040]图23.a.与基线(基础)相比,sni在损伤后4天(n=16)诱导机械痛阈(mt)致敏;配对t检验,*p《0.05,假手术组术后4天机械阈值无显著差异。b、单次皮下注射吗啡对创伤后mt的镇痛作用(sni mor,n=4)。平均值±sem;治疗前后单因素方差分析f=4.62,*p《0.05;经bonferroni多重比较检验,平均差和95%可信区间分别为-7.3、-14.5至-0.135,p《0.05。20mg/kgmor(n=4;f=19.03,**单因素方差分析p《0.001)。[0041]图24.在sni小鼠中,用生理盐水(对照组)和4剂量吗啡(mor)形成条件反射(condition)后的条件性位置偏好(cpp)。mor诱导的cpp呈剂量依赖性(单因素方差分析,f=4.167,p《0.05);通过tukey多重比较试验,10mg对对照组(p《0.001)和20mg(p《0.05)。[0042]图25.暴露于1μmrpp(seqidno:1)或其一种变体(表3)后,作为底物的rest肽(tedpspppspplpken)的体外ctdsp1磷酸酶活性(相对于gst对照)。[0043]图26.相对于水对照,在暴露于rpp(seqidno:12)后,磷酸化rest肽(tedpspppspplpken)作为底物的体外靶向(ctdsp1)和非靶向磷酸酶活性(1=ppa1,2=ppm1h,3=ppm1a,4=pp3ca,5=ppp1ca,6=pp5ca)。n=5,误差棒为标准偏差。[0044]图27.构建rpp库。a)实验方案;b)rest构建物的核苷酸序列(突变前);c)其余构建物的蛋白质序列(突变前)。rpp、cpp和his标签都有下划线。[0045]图28.rpp(seqidno:13)结构。沿着结构的底部是cpp,它含有一个多精氨酸序列和两个2-氨基十四烷酸,这些酸与其余的ri类磷酸结合(从结构右下方的赖氨酸开始)。具体实施方式[0046]在一个实施方案中,本发明提供了开发与ctdsp1具有高亲和力的肽的方法(如实施例1至3所述)。重要的是,该方法使用肽进化技术生成与ctdsp1具有高亲和力的肽。[0047]在一个实施方案中,本发明提供seqidno:1和15至17的肽或与seqidno:1和15至17具有至少50%、更优选至少60%、更优选至少70%、更优选至少80%、更优选至少90%或更优选至少95%相似性的肽。这些肽或其类似肽在本文中称为rest类磷酸肽(rpp)。然而,重要的是,必须保持rpp中的谷氨酸,尤其是seqidno:1的位置5和8、seqidno:15的位置4和7、seqidno:16的位置8和11以及seqidno:17的位置6和9处的谷氨酸。[0048]在一个实施方案中,本发明提供rppv(seqidno:18至117)或与rppv(seqidno:18至117)具有至少50%、更优选至少60%、更优选至少70%、更优选至少80%、更优选至少90%、更优选至少95%相似性的肽。所述肽或其类似肽在本文中称为rest类磷酸肽变体(rppv)(seqidno:18至117)。然而,重要的是,必须保持seqidno:18至67中的谷氨酸,尤其是seqidno:1中5和8位置的谷氨酸。对于反向倒置序列(rppvri)(seqidno:68至117),必须保持seqidno:16的位置8和11。[0049]在一个实施方案中,本发明是rpp(seqidno:1和15至17)或rppv(seqidno:18至117)以及细胞内转运肽(seqidno:118至137和140至159)(或与之具有至少50%、更优选至少60%、更优选至少70%、更优选至少80%的肽、更优选至少90%或更优选至少95%相似性的肽)的融合蛋白。[0050]两个或多个核苷酸序列或两个或多个氨基酸序列之间的同一性/相似性通过序列之间的同一性或相似性来表示。序列同一性可以用同一性百分比来衡量;百分比越高,序列越相同。序列相似性可以用相似性百分比来衡量(考虑保守的氨基酸替换);百分比越高,序列越相似。[0051]用于比较的序列比对方法在本领域是众所周知的。各种程序和比对算法如:smith&waterman,adv.appl.math.2:482,1981;needleman&wunsch,j.mol.biol.48:443,1970;pearson&lipman,proc.natl.acad.sci.usa85:2444,1988;higgins&sharp,gene,73:237-44,1988;higgins&sharp,cabios5:151-3,1989;corpet等人,nuc.acidsres.16:10881-90,1988;huang等人,computerappls.inthebiosciences8,155-65,1992;andpearson等人,meth.mol.bio.24:307-31,1994.altschul等人,j.mol.biol.215:403-10,1990详细介绍了序列比对方法和同源性计算。[0052]ncbi基本局部比对搜索工具(blast)(altschul等人,altschul等人.,j.mol.biol.215:403-10,1990)可从多个来源获得,包括国家生物信息中心(ncbi,国家医学图书馆,38a栋,8n805室,bethesda,md20894)和互联网,用于序列分析程序blastp,blastn,blastx,tblastn和tblastx。有关更多信息,请访问ncbi网站(www.ncbi.nlm.nih.gov)。[0053]blastn用于比较核苷酸序列,blastp用于比较氨基酸序列。如果两个比较的序列共享同源性,那么指定的输出文件将把这些同源区域表示为比对的序列。如果两个比较的序列没有同源性,那么指定的输出文件将不会显示比对的序列。[0054]比对时,匹配的数量通过计算两个序列中存在相同核苷酸或氨基酸残基的位置数量来确定。通过将匹配数除以已识别序列中所述序列的长度,或除以分节的长度(例如,已识别序列中所述序列的100个连续核苷酸或氨基酸残基),然后将所得值乘以100来确定序列一致性百分比。例如,当与具有1154个核苷酸的测试序列比对时,具有1166个匹配的核苷酸序列与测试序列(1166/1554*100=75.0)75.0%相同。百分比序列标识值四舍五入到最接近的十分位。例如,75.11、75.12、75.13和75.14四舍五入为75.1,而75.15、75.16、75.17、75.18和75.19四舍五入为75.2。长度值将始终为整数。在另一个示例中,包含15个核苷酸区域的靶序列,该区域与以下识别序列中的20个连续核苷酸比对,包含与该识别序列具有75%序列同源性的区域(即,15/20×100=75)。[0055]对于大于约30个氨基酸的氨基酸序列的比较,使用设置为默认参数的默认blosum62矩阵,使用blast2序列函数(空位(gap)存在代价(cost)为11,每个残基的空位代价为5/1)。同源物的典型特征是拥有至少70%的序列同一性,在与氨基酸序列的全长比对中使用ncbibasicblast2.0,具有nr或swissprot数据库等数据库的空位blastp。用blastn程序搜索的查询用dust过滤(hancock和armstrong,1994,comput.appl.biosci,10:67-70)。其他程序使用seg。此外,还可以执行手动比对。当用这种方法评估时,具有更大相似性的蛋白质将显示出越来越多的百分比同一性,例如与蛋白质至少有75%、80%、85%、90%、95%、98%或99%的序列同一性。当比对短肽(少于约30个氨基酸)时,使用blast2序列功能执行比对,使用pam30矩阵设置为默认参数(开头空位9,延长空位1罚分)。当用这种方法评估时,与参考序列更为相似的蛋白质将显示出越来越多的同一性,例如与蛋白质至少60%、70%、75%、80%、85%、90%、95%、98%或99%的序列同一性。当比较的序列同一性小于整个序列时,同源物通常在10-20个氨基酸的短窗口内拥有至少75%的序列同一性,并且可以拥有至少85%、90%、95%或98%的序列同一性,这取决于它们与参考序列的同一性。ncbi网站描述了在如此短的窗口中确定序列同一性的方法。[0056]两个核酸分子密切相关的一个指标是,如上所述,这两个分子在严格的条件下相互杂交。尽管如此,由于遗传密码的退化性,未示出高度同一性的核苷酸序列可能编码相同或相似(保守)的氨基酸序列。利用这种简并性可以改变核苷酸序列,从而产生多个核酸分子,这些核酸分子都编码基本相同的蛋白质。两个核苷酸序列基本相同的另一个(不一定是累积的)指标是,第一个核酸编码的多肽与第二个核酸编码的多肽在免疫上发生交叉反应。[0057]在某些实施方案中,rpp(seqidno:1和15至17)或rppv(seqidno:18至117)可与另一肽融合,以便例如允许细胞穿透。优选地,将rpp(seqidno:1和15至17)或rppv(seqidno:18至117)融合到允许细胞穿透和从内涵体逃逸的肽序列。更优选地,细胞穿透和逃逸肽序列是一个seqidno:118至137和140至159。rpp(seqidno:1和15至17)或rppv(seqidno:18至117)和细胞穿透肽(cpp)(seqidno:118至137和140至159)可直接融合,或包含连接它们的连接序列(例如,seqidno:138、139、160或161)。已经表明,当细胞或组织被rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽与cpp(seqidno:118至137和140至159)融合时,其定位于细胞核(图3,seqidno:4和图4_seqidno:4),导致rest降解(图5,seqidno:4),增加rest靶向基因的表达(图7、seqidno:4、图8、seqidno:1、图9、线性_seqidno:4和环化_seqidno:2、图10、seqidno:9、图14、seqidno:12、图15、seqidno:13和14以及图16、seqidno:12),并导致神经分化(图11、seqidno:5至11、图12、seqidno:9和图13、seqidno:13)。[0058]本发明人发现rpp(seqidno:1和15至17)结合ctdsp1以抑制rest活动。如图2和3所示的结合曲线所示,rpp对ctdsp1有较低的pm亲和力。在细胞和动物中,rpp与ctdsp1结合,从而阻止ctdsp1与rest结合。[0059]rest的增加和相关的神经基因抑制是以下疾病或病变的病理学的基础:[0060]创伤性脑损伤-创伤性脑损伤(tbi)导致神经元死亡导致认知能力丧失。为了改善tbi后的认知恢复,需要解决两个问题:1)减少损伤后持续的神经元死亡;2)使丢失的神经元再生。[0061]tbi后,大脑中的rest水平显著增加(图19)。几项研究一致表明,急性缺血引起的脑损伤诱导神经元表达rest,导致神经元死亡7-9。这些研究表明,当消除rest时,神经元存活率会提高7-9。其临床意义深远。例如,rest水平的提高与癫痫发作频率的增加相关,癫痫发作通常发生在脑损伤后10-12。在脑损伤的动物模型中,当rest受到抑制时,癫痫发作率显著降低13-14。此外,脑外伤后长期rest水平升高可能在脑损伤史引起的神经退行性疾病中起作用15-17。体外和体内研究都表明,消除rest可以提高神经元的存活率7-9、功能14,18和再生2,19-24。因此,tbi后靶向rest应该可以减轻脑损伤的影响,并抵消形成相关的年龄相关神经退行性疾病(如阿尔茨海默病)的风险17,25。[0062]神经发生改善tbi患者认知功能的基础是将神经发生的增加与学习、记忆和其他认知功能的改善联系起来的研究26-30,以及表明通过抗有丝分裂剂、辐射或基因操纵抑制神经发生会损害海马依赖型记忆的相反研究29-33。神经发生缺陷也与许多认知病因疾病有关,包括发育障碍(如小头畸形34、巨脑症35、自闭症35)和神经退行性疾病(如痴呆和阿尔茨海默病)36。实验治疗(包括输血、生长和神经营养因子以及干细胞移植10,37-49)专注于再生神经元。这些尝试未能刺激恢复认知功能所需的神经发生,至少部分是因为它们不能在tbi的情况下最终分化神经祖细胞26,28,50-57。re1沉默转录因子rest在单个检查点阻断了神经祖细胞向神经元的终末分化。[0063]周围神经损伤-周围神经损伤(pni)患者的神经元再生至少通过两种方式受到抑制。首先,支持发育和成熟神经元生长、存活和分化的神经营养因子(ntfs)减少58。其次,突触可塑性(包括轴突生长、囊泡转运和离子传导)所需基因的表达减少59。这两种现象与损伤诱导的rest表达一致60。因此,通过逆转神经再生障碍,消除rest可以加速pni的恢复。用rpp抑制ctdsp1可促进rest降解(图6),增加神经元分化(图11、12和13),并增加ntf表达(图7、9、10和16),这表明融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)刺激神经再生的潜力。[0064]慢性疼痛-慢性疼痛期间中枢和外周循环中被抑制的许多基因是抑制元件-1(re1)-沉默转录因子(rest)的直接或间接靶点,rest是干细胞、神经祖细胞和非神经细胞中神经元基因表达的抑制剂61-63。正常神经元的rest水平通常保持在较低水平,通过主动降解使染色质远离这种强大的神经元基因阻遏物64-69。在向神经病理性疼痛过渡之前,周围神经损伤后,小鼠和大鼠周围神经神经元的rest水平显著增加,随后中枢神经系统中处理和感受疼痛刺激的神经元出现峰值60,70,71。神经损伤后rest的激活导致感觉神经元正常兴奋性所需的几个基因,包括钾通道kv4.3(kcnd3)和kv7.2(kcnq2),钠通道nav1.8(scn10a)和μ阿片受体oprm1(图18)表达减少60,72-74。阻断rest以缓解慢性疼痛的基础发表于利用小鼠和大鼠周围神经损伤(pni)模型发表的研究60,72-74。在小鼠中,pni后感觉神经元中rest的基因敲除恢复了μ阿片受体60,72,73、nav1.860,72,73、kv4.372,73,75、kv7.272,73在背根神经节的基因表达,恢复c神经纤维的正常k m型电流60,减少痛觉过敏和痛觉超敏72-74,并恢复了慢性疼痛模型中的吗啡镇痛(表12)。使用msin3结合位点(msin3结合位点是一种适配器蛋白)的优化rest模拟物破坏rest抑制复合物,有助于许多基因的转录抑制,包括rest靶点,恢复c纤维功能,减少痛觉过敏和痛觉超敏,并恢复慢性疼痛小鼠模型中的吗啡镇痛(表12)74。在大鼠中,敲低rest控制了从急性疼痛到慢性疼痛的转变,表现为痛觉过敏和痛觉超敏的减少,并恢复了毒蕈碱镇痛(表12)72,73。[0065][0066][0067][0068]我们认识到,表12所示的研究表明,小鼠条件性敲除(cko)研究(完全敲除)与大鼠sirna敲除研究在降低超敏反应方面存在差异,后者显示损伤后没有超敏反应,也没有下调rest靶基因(表12),这只能部分减少损伤后的痛觉超敏(高达45%)和痛觉过敏(高达65%)。我们认为,敲除效应比sirna敲除更有效,因为sirna的生物利用度受到其大小(》13000kd)和因内涵体包埋导致的细胞运输不良的限制。[0069]我们知道,表12中的研究缺乏严谨性,因为没有进行功效分析来确定每组动物的数量,并且动物数量很低,这使得统计分析有问题。一些研究使用预测性较差的部分坐骨神经结扎(psnl)模型,而不是首选的备用神经损伤(sni)慢性疼痛模型。这些研究中的大多数只评估雄性。然而,尽管存在这些缺点,但研究结果是一致的,因此,支持我们提出的治疗慢性疼痛药物的体内疗效评估。此外,由于rest活动在哺乳动物中高度保守,我们相信这些研究为测试人类rest调节提供了强有力的概念验证支持。[0070]我们发现rest是通过其丝氨酸861和864的磷酸化来降解的。rest通过c端结构域小磷酸酶1(ctdsp1)使这些位点去磷酸化而稳定2-4。与rest一样,ctdsp1在非神经组织中表达,通过与rest的相互作用促进神经基因沉默。我们得出结论,抑制ctdsp1足以促进rest降解并刺激基因表达。为了支持这一结论,我们已经证明,间充质祖细胞(mpc)和神经中ctdsp1的敲除可导致rest靶基因的表达和轴突再生76。我们利用我们对rest-ctdsp1相互作用的机制理解,开发了一种包含rest相关调节区域的环化类磷酸肽77。其余的类磷酸肽(rpp)有几个药理学属性。由于其环化结构和d-氨基酸组成(例如,seqidno:9、12、13、14、16、17和68-117),它非常稳定。它具有超过大多数抗体的较低pm结合亲和力(图2和图3),表明发生脱靶效应的风险较低。其小尺寸(《3.5kd)表明其生物利用度应接近小分子药物。我们的初步数据表明,我们的模拟物抑制ctdsp1的活性,降低rest蛋白水平,并导致适当神经元活动所需的神经基因的表达77。[0071]最重要的是,我们的方法预计安全有效,原因如下:1)我们的药物对诸如神经损伤后的再生反应的核心靶点具有选择性。染色质的基本组织不允许表观遗传“书写”一组新的指令;然而,神经损伤会触发协同、广泛和瞬时的核小体重组,即基因组瞬时中间状态(gtis):一种有利于表观遗传重编程以获得必要反应的临时核小体结构78-81。2)我们的药物具有很高的结合亲和力(低pm),因此不会产生脱靶效应(图2和图3)。3)我们建议短治疗期少于1个月我们缩短治疗周期的基础是rest是神经元终末分化的守门员64,82(将细胞不可逆地提交给该谱系的过程83)。神经祖细胞,与慢性疼痛状态下的神经元具有表观遗传相似性63,因为它们也在gtis中,诱导几天后进行最终分化。与之一致地,我们证明,rpp可以在7天内诱导神经元分化(通过map2阳性细胞测定,map2是神经元分化的标志物)(图11、12和13)。一旦该过程开始并且细胞被提交(committed),就不需要进一步治疗。因此,我们期望通过这一早期治疗期实现的疼痛缓解能够持久。4)在测定坏死的离体drg神经元毒性检测中,我们已经证明我们的药物对神经元没有毒性(图17)。5)rest和ctdsp1主要参与神经发育,因此,我们预测抑制它们在受损的成年细胞中的活性将得到很好的耐受性。6)多肽药物的安全性使其成为一种有吸引力的治疗策略。目前,已上市的肽类药物超过68种,全球销售额超过147亿美元,140种正在临床开发中84,85。[0072]创新点[0073]一流的药物候选。我们开发了第一种靶向神经基因转录抑制物rest的药物。已发表的文献表明,rest表达导致慢性疼痛(表12)。pni后,外周神经系统(图18)和中枢神经系统的rest和ctdsp1表达均增加,导致功能障碍7-9。几项发表在啮齿动物身上的研究表明,阻断rest可以缓解慢性疼痛60,72-74。[0074]创新的药物设计。我们已经开发出一种环化类磷酸细胞穿透肽,它通过抑制ctdsp1(可保护其余部分不被降解一种磷酸酶)来破坏rest的稳定性3,从而允许神经特异性基因的表达。与rest一样,ctdsp1的表达仅限于非神经元细胞类型,神经元损伤后除外。我们的抑制剂是新颖的,因为它直接针对转录检查点,该检查点可立即调节诱导再生所需的基因。它克服了靶向转录因子和创建抗丝氨酸磷酸酶疗法的难题。关于后者,其他方法侧重于磷蛋白磷酸酶(ppp)和金属依赖性蛋白磷酸酶(ppp)86。然而,ctdsp1是一种卤酸脱卤酶(had),它使用两种催化型天冬氨酸,而不是仅仅依靠金属离子发挥活性87。这一催化位点能够受到特异性抑制,成功开发出针对类似酶(如hiv-1蛋白酶)的治疗药物证明了这一点88-91。最后,ctdsp1具有脯氨酸依赖性底物偏好,这在其他丝氨酸磷酸酶中没有发现,我们利用它来改善药物的结合和稳定性。[0075]初步数据。rest通过与神经元基因调节区附近的阻遏元件-1(re-1)位点的染色质结合来抑制基因表达,包括离子通道、生长因子和轴突导向蛋白1。因此,在干细胞如神经祖细胞(npc)能够最终分化之前,它们必须首先以rest为降解靶点,以表达所需的神经元基因64。ctdsp1保护rest免受降解,ctdsp1是阻止神经元基因表达所必需且充分的2。显性负性ctdsp1能结合磷酸化靶点,但不能催化去磷酸化,诱导p19干细胞的终末分化92。[0076]我们之前已经证明,ctdsp1和rest之间的相互作用依赖于map激酶erk2对rest丝氨酸861和864的磷酸化,并且这些丝氨酸突变为丙氨酸会增加rest稳定性2-4。我们推测,该rest调节区的一种不可水解的类磷酸化可以减缓ctdsp1活性并促进rest降解。为了验证我们的假设,我们开发了一种rest类磷酸肽或rpp77以评估rest和rest靶标的剂量依赖性影响。rpp含有丝氨酸861和864突变为谷氨酸的rest的氨基酸858至870,其可以模拟磷酸丝氨酸的形状和总电荷。在c端,我们融合了分别来自hiv-tat和ha2的富含精氨酸的细胞穿透肽和内涵体退出序列(exit序列),用于将肽和蛋白质输送到培养的哺乳动物细胞和活生物体中,并促进血脑屏障(bbb)的穿越93-96。[0077]我们研究了rpp是否可以降低rest蛋白(图6)。在左图中,蛋白免疫印迹(wb)显示rpp降低了hek细胞的rest(外源 内源)水平。在右图中,通过wb分析对总rest蛋白(外源 内源)水平进行量化显示,rpp给药的hek细胞的总rest蛋白水平下降了58.3%(图6)。[0078]在体外结合检测中,rpp对ctdsp1的pm亲和力较低,且下降速度较慢(图2和图3)。在磷酸酶抑制的体外试验中,rpp在低nm浓度下抑制其靶点ctdsp1,但在10μm时不抑制非靶点pp5、pp1、ppm1h、ppm1a或pp3ca活性(图26)。在mpcs的细胞通透性和稳定性分析中,rpp易位到细胞核中(图4和图5),这与rest和ctdsp1的定位图谱一致92,97。培养6天后,rpp水平保持稳定。[0079]通过在啮齿动物慢性疼痛模型中阻断rest,可以防止nav1.8、kv4.3、kv7.2和oprm1的下调,其被认为是痛觉过敏和痛觉超敏的原因(表12),这一点已经得到证实。基于这些研究,我们测试了阻断ctdsp1是否可以刺激与慢性疼痛相关的rest靶基因的表达。我们还表明,转染rpp(不含hivtat-ha2)或施用线性或环化rpp的nbfl细胞的kv4.3mrna增加(分别为2倍和4倍)(图8和9c)。[0080]我们发现,与对照组相比,3μmrpp(seqidno:5-11)使人类神经祖细胞(ipsc)的分化增加了2至2.7倍,如归一化为dapi(细胞核)的map2(成熟神经元标记)表达的测定所示(图11),1μmrpp(seqidno:9和13)增加了人类神经祖细胞(ipsc)的分化(图12和13),如归一化为dapi(细胞核)的tuj1(分别高达36%和3倍)和map2(分别高达33%和2倍)(神经元标记物)表达的测定所示。与对照组相比,细胞死亡没有增加(数据未示出)。使用该筛选的基础是,消除神经祖细胞中的rest已被证明可诱导神经元分化22,这是神经元基因整体去抑制的指标。[0081]在疗效和毒性的初步评估中,在整个drg神经元的离体培养中,在0、1、3或10μm下对rpp进行48小时的评估,以确定其诱导kv4.3(图14)、kv7.2(图14)、nav1.8(图14和15)和oprm1(图14)表达的潜力,并引起神经毒性(图17)。3μm时,seqidno:12诱导kv4.3,kv7.2,nav1.8和oprm1分别约为对照组的0、6、3和3倍(图14)。3μm时,seqidno:13和14诱导nav1.8分别约6和13倍(图15)。10μm时,seqidno:12感应kv4.3,kv7.2,nav1.8和oprm1分别约为2、7、5和4倍(图14)。在乳酸脱氢酶ldh细胞毒性检测中,rppseqidno:12在0、1、3或10μm时未示出毒性(图16)。正如所料,阳性对照(triton,n=6))是有毒的(图17)。[0082]癫痫-癫痫是大脑中不受控制的电活动,导致混乱、意识丧失和不受控制的运动98。它是离子通道和包括scn1a(nav1.1)、scn2a(nav1.2)、scn1b(navβ亚基1)、kcnq2(kv7.2)和kcnq3(kv7.3)的受体失调的结果,这些受体都是rest抑制的靶标14,99。癫痫的根本原因是神经元核内rest水平的增加100。[0083]糖尿病-胰腺β细胞和神经元在分化过程中具有相似的转录途径,涉及rest的消除101。β细胞过度表达rest下调rest靶基因降低胰岛素分泌102,103。[0084]阿尔茨海默病——在阿尔茨海默病中,乙酰胆碱和胆碱乙酰转移酶(chat)是合成乙酰胆碱所必需的转移酶,含量非常低。这种酶浓度的降低会导致与阿尔茨海默氏症相关的记忆和认知缺陷。大脑rest水平的增加会抑制chat16。[0085]亨廷顿病-在亨廷顿病中,rest区从胞质向神经元细胞核的移位被认为是与该病相关的神经元退化的原因104-106。[0086]脑癌,包括多形性胶质母细胞瘤-脑癌,包括多形性胶质母细胞瘤(gbm),起源于脑肿瘤起始细胞(btic),这是一种具有强大自我更新能力的癌干细胞,可成为癌性肿瘤。这些细胞对放疗和化疗有抗性,并在治疗数月后变成增殖性的107-109。融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽具有通过终末分化btic阻止肿瘤形成的潜力。一旦细胞最终分化,它们就不能再增殖并产生新的肿瘤110。必须指出,终末分化与通过其他方法(例如通过组蛋白脱乙酰酶抑制剂(hdac)靶向染色质重塑)实现的分化不同。抑制这些酶会随意改变基因表达,无法实现预防复发性gbm所需的永久性终末分化。btic向神经元的终末分化在单个检查点被rest阻断。通过观察到rest水平的增加与gbm的复发相关,强调了这种抑制因子在维持致癌性方面的重要性111-113,以及更短的无病生存期112-114。btic依赖rest维持其致癌性为治疗干预创造了机会。rest通过丝氨酸861和864的磷酸化进行降解,并且这些丝氨酸通过c端结构域小磷酸酶ctdsp1保持在去磷酸化状态2。因此,用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15-17)或rppv(seqidno:18至117)抑制ctdsp1应能释放终末分化的rest-制动(brake),并防止gbm和其他脑肿瘤的复发。[0087]胰腺癌-晚期、转移阳性胰腺癌细胞的rest水平较高115。胰腺癌患者肿瘤中的rest水平较高,生存率较低115。体外功能实验表明,rest基因敲除可抑制胰腺癌细胞(aspc-1和panc-1)的增殖、迁移、侵袭和上皮-间充质转化115。体内实验(皮下balb/c裸鼠模型和肠系膜上静脉注射balb/c裸鼠模型)表明,敲除rest抑制了异种移植瘤的生长和转移115。因此,抑制rest可以改善这些疾病的患者预后。因此,本发明还涉及抑制细胞内rest的方法。通常,该方法包括将细胞与融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽接触。例如,它可以将细胞暴露足够长的时间,以使融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)进入细胞并对rest活动产生影响。该方法可在体外或体内实施。在体外实施的情况下,该方法可用于研究rest的活性,以测试其他化合物对融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)对rest的影响的补充或拮抗能力,或出于对研究人员重要的任何其他原因。当在体内实施时,该方法可用作对受试者一种或多种与rest相关的疾病或病变进行治疗的方法。根据本发明这方面的方法,优选地,rest的活性降低。接触细胞的步骤可以是使药剂物理接触一个或多个靶标细胞的任何操作。因此,可以通过将药剂直接添加到要接触的细胞的体外培养物中,并允许药剂有足够的时间在培养基扩散并接触至少一个细胞。同样,也可以通过向水环境中的细胞中添加药剂来实现。或者,它可以通过任何可接受的给药途径向受试者施用药剂,并允许受试者的身体通过自然过程将药剂分配给目标细胞。因此,体内方法可以是将药剂局部或系统地递送到动物细胞的方法,尤其包括所有哺乳动物和人类。根据该方面,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)可用于治疗或预防性治疗受试者,并制备用于治疗的组合物。[0088]在另一个实施方案中,本发明提供了一种治疗患有或有可能患有涉及rest的疾病或病变的受试者的方法。通常,该方法包括施用足以影响受试者rest量或活性的剂量的、融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)。在某些方面,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)与ctdsp1的结合导致细胞中rest活性的抑制。通常,该方法包括施用足量的融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)维持足够长的时间,以抑制rest活性。通常,给药量和时间足以看到疾病或病变的一个或多个临床症状的变化,或阻止疾病或病变进展到出现一个或多个临床症状的阶段。根据该方面,该药剂可用于治疗或预防性地治疗受试者,并制备用于治疗的组合物。[0089]在一个实施方案中,本发明提供了治疗、减轻或改善受试者的创伤性脑损伤、慢性疼痛、周围神经损伤、癫痫、糖尿病、阿尔茨海默病、亨廷顿病、脑肿瘤(包括多形性胶质母细胞瘤)或胰腺癌的方法。这里使用的术语“治疗”或“缓解”或“改善”以及类似术语包括预防和完全或部分治疗。这些术语还可以包括减轻症状、改善症状、降低症状的严重程度、降低疾病的发生率,或改善治疗结果的患者状况的任何其他变化。这些方法涉及对患有创伤性脑损伤、慢性疼痛、周围神经损伤、癫痫、糖尿病、阿尔茨海默病、亨廷顿病、脑肿瘤(包括多形性胶质母细胞瘤)或胰腺癌的动物受试者,或需要治疗创伤性脑损伤、慢性疼痛、周围神经损伤、癫痫、糖尿病、阿尔茨海默病、亨廷顿病、脑肿瘤(包括多形性胶质母细胞瘤)或胰腺癌的动物受试者,施用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)。[0090]可通过任何已知和可接受的途径向受试者施用融合至cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)。这些途径包括但不一定限于口服、通过粘膜(例如,经鼻、经吸入、经直肠、经子宫或经阴道、舌下)、经静脉(例如,静脉推注、静脉输注)、经腹腔和皮下。同样,可以通过直接注射到含有靶细胞(即待治疗细胞)的部位(例如,器官、组织)来施用。此外,施用可以遵循任何数量的方案。因此,它可以包括药物的单次剂量或给药,或一段时间内的多次剂量或给药。因此,治疗可涉及一次或多次重复施用步骤,直到达到预期结果。在实施方案中,治疗可以持续更长的时间段,例如周、月或年。本领域技术人员完全能够基于本领域的已知参数为个人轻松开发合适的给药方案。因此,这些方法也考虑控制但不一定消除疾病或病变。根据本发明的优选施用途径为口服和通过粘膜。[0091]根据受试者、疾病阶段、受试者年龄、受试者的一般健康状况以及医学领域技术人员已知并经常考虑的各种其他参数,施用量会有所不同。作为一般事项,将施用足量的药剂,以使受试者的症状发生可检测的变化。本文公开了合适的量,并且本领域技术人员可以在不进行过分(undue)或过度实验的情况下确定其他的合适量。[0092]融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)以受试者可接受、可耐受和有效的形式施用。本领域已知用于生物活性剂的多种药物形式和剂型,并且本发明设想了所有这些形式和剂型。因此,例如,该制剂可以配制成口服溶液、囊片(caplet)、胶囊、注射剂、不溶剂、栓剂、锭剂、片剂、乳膏或软膏、吸入剂等。[0093]本领域的普通技术人员将根据良好的医疗实践和个体受试者的临床状况,很容易优化有效剂量和施用方案。[0094]给药频率取决于化合物的药代动力学参数和施用途径。最佳药物配方将由本领域技术人员根据施用途径和所需剂量确定。此类制剂可能影响给药制剂的物理状态、稳定性、体内释放速率和体内清除速率。根据施用途径,根据体重、体表面积或器官大小计算合适的剂量。动物模型的可用性尤其有助于确定给定治疗药物的适当剂量。本领域的普通技术人员通常会进一步完善确定适当治疗剂量所需的计算,而无需进行不必要的实验,尤其是根据本文所公开的剂量信息和分析以及在动物或人类临床检测中观察到的药代动力学数据。[0095]通常,通过使用已建立的测定血液水平的分析以及相关剂量反应数据来确定适当的剂量。最终的给药方案将由主治医生决定,并考虑改变药物作用的因素,例如药物的比活性、损伤的严重程度和患者的反应性、患者的年龄、状况、体重、性别和饮食、任何感染的严重程度、施用时间和其他临床因素。随着研究的进行,将出现关于特定疾病和条件的适当剂量水平和治疗持续时间的进一步信息。然而,这些研究是常规的并且在本领域技术人员的水平之内。典型剂量可为每周(人体)约0.6mg(0.01mg/kg)至约60g(1g/kg),更优选每月。[0096]应了解,本发明的肽、组合物和治疗方法在人类医学和兽医学领域中有用。因此,要治疗的对象是哺乳动物,例如人类或其他哺乳动物。用于兽医目的,受试者包括例如农场动物,包括牛、羊、猪、马和山羊,伴侣动物,如狗和猫,外来动物和/或动物园动物,以及实验动物,包括小鼠、大鼠、兔子、豚鼠和仓鼠。[0097]融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)可作为药物或兽医学组合物(如片剂、胶囊、溶液或乳剂)施用给有需要的受试动物,优选哺乳动物,如人类。融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)可存在于含有其他成分的组合物中。适用于本发明的组合物的非限制性示例为药物组合物,例如片剂、丸剂、胶囊、囊片、多微粒(包括颗粒、珠子、球粒和微囊颗粒);粉末、酏剂、糖浆、悬浮液和溶液的形式。药物组合物通常包含医药上可接受的稀释剂或溶剂。药物组合物优选适于肠外给药(例如,口服)。可口服施用的组合物可以是固体或液体形式,可以是片剂、粉末、悬浮液和糖浆等形式。任选地,所述组合物包含一种或多种调味剂和/或着色剂。一般而言,治疗和营养组合物可包含不会显著干扰药剂对受试者的作用的任何物质。[0098]适用于此类组合物的药学上可接受的赋形剂或溶剂在药学领域是众所周知的。本发明组合物可含有0.01-99%(按重量计)的药剂。本发明的组合物通常以单位剂型制备。用于制备这些组合物的赋形剂在本领域是众所周知的。[0099]该组合物的产品形式的其他示例为食品补充剂,例如软凝胶或硬胶囊的形式,其包含选自明胶、淀粉、变性淀粉、淀粉衍生物(例如葡萄糖、蔗糖、乳糖和果糖)的封装材料。所述封装材料可任选地包含交联剂或聚合剂、稳定剂、抗氧化剂、用于保护光敏填料的吸光剂、防腐剂等。[0100]一般而言,在本技术中可使用溶剂一词来表示可与制剂混合的组合物,无论是药物溶剂、食品、营养补充剂或膳食辅助剂。就本发明而言,上述材料可被视为药剂的溶剂。在本发明的某些实施方案中,溶剂具有很少或没有生物活性,尤其是对rest。[0101]无需进一步描述,相信本领域普通技术人员可以使用前面的描述和下面的说明性实施例,制备和利用本发明的化合物,以及实施要求保护的方法。给出以下实施例以说明本发明。应当理解,本发明不限于实施例中描述的特定条件或细节。[0102]实施例1-his-ctdsp1质粒构建[0103]密码子优化(使用idt密码子优化工具)的ctdsp1基因被克隆到pbadhisa质粒中(thermofisherscientific)。首先,用引入hindiii和xhoi限制位点的引物p33和p34(表1)扩增pbad-hisa质粒。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸6分钟。扩增后,用qiagen凝胶条带纯化试剂盒对pcr片段进行凝胶纯化,并用hindiii和xhoi限制性内切酶消化。根据制造商的建议,使用qiagen试剂盒纯化所有消化片段,并使用t4dna连接酶(neb)以适当的组合连接。[0104]表1:用于质粒构建的引物[0105][0106][0107]根据制造商的方案,将连接片段转化到10g化学感受态细胞(lucigen)中。将转化细胞接种在含有50μg/ml氨苄西林的lb板上,并在37℃下培养过夜。通过菌落pcr检测菌落是否存在插入物。采集菌落并在20μl无菌0.9%氯化钠溶液中重新悬浮。将1μl该溶液转移到pcr管中,并用taq聚合酶(newenglandbiolabs,cat#m0482s)和30pm的侧翼引物进行扩增。最初将每个pcr反应(20μl)加热至95℃2.5分钟,然后94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸1分钟。琼脂糖电泳显示扩增产物。将具有正确插入物的克隆接种到含有5mllb和适当抗生素的培养管中,并在37℃下培养过夜。然后使用monarplasmidminiprep试剂盒(neb)纯化构建物。[0108]实施例2-his-ctdsp1的表达和纯化方法[0109]his-ctdsp1(表2;seqidno:162):将带有pbad-ctdsp1构建物的大肠杆菌(10g菌株lucigen)在37℃下剧烈摇动过夜。然后在1l烧瓶中,将4ml夜间培养物添加到500ml含有50μg/ml氨苄西林的lb培养基中,并在37℃下摇动培养。当培养物的od600达到0.4时,我们添加阿拉伯糖至最终浓度0.02%,并在30℃下摇动培养16小时。第二天早上,细胞以最大速度向下旋转(eppendorf离心机5810r),并在-80℃下冷冻。必要时,从冰箱中取出细胞颗粒,在室温下培养,用4mlbper蛋白裂解试剂(thermofisher)裂解,并如制造商所述用hispurcobalt纯化试剂盒(thermofisher,cat#90091)纯化蛋白。[0110]表2-ctdsp1核苷酸和蛋白质序列[0111][0112][0113]实施例3-开发对ctdsp1具有高亲和力的肽[0114]使用rna展示和蛋白质进化方法生成候选肽。[0115]rna展示文库的构建:我们已经构建了3个不同的rpp变体库。在所有文库中,丝氨酸861和864被谷氨酸盐取代。文库1是用引物p11到p14构建的(表1)。在文库1中,其余肽的每个位置都发生了突变。据估计有1×109个变体。在文库2中(引物p14至p18,表1),简并寡聚体未被谷氨酸盐修饰(touched)。该库预计有65×106个变体。在库3中,只有一个谷氨酸盐和每两个密码子发生突变。该库预计只生成65000个变体。[0116]在所有3个文库中,rpp序列(seqidno:1)通过合成简并寡核苷酸多样化。每个密码子只引入一个简并碱基:第一个或第二个位置,可以选择或4个氨基酸。其余盒(casssete)从pbad构建物中扩增为两个片段:左片段和右片段,通过连接重新结合(图27)。左侧片段用侧翼正向引物p23(表1)和一个反向引物(11r槽22r,表1)扩增。[0117]用一个正向引物(11f至22f,表1)和反向引物p24(表1)扩增右侧片段。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30循环,并在72℃下延伸40秒。扩增后,用qiagen凝胶条带纯化试剂盒对pcr片段进行凝胶纯化,并与t4dna连接酶混合和连接。连接反应包含20μl10x连接缓冲液、100ng片段混合物、0.5μl100mmatp、1μlt4dna连接酶(nebcat#m0202s)和1μlt4多核苷酸激酶。反应混合物在室温下孵育,并使用上述程序将其用作模板用侧翼引物p17和p20(表1)进行pcr。pcr片段通过qiagen凝胶条带纯化试剂盒进行凝胶纯化,并用作mrna展示实验的模板。[0118]出于质量控制目的,我们克隆并测序了该文库的一部分。用hindiii和xhoi限制性内切酶消化该文库的一小部分,用qiagen试剂盒纯化,并如上所述在pbad载体中克隆ctdsp1基因。通过genewiz对单个克隆进行测序,以确认rest盒成功突变,并且大多数克隆没有发生移码突变。[0119]体外转录:根据制造商协议,使用ribomax大规模rna生产系统t7(promega,cat#p1300)从扩增文库中翻译rna,并通过rneasy迷你试剂盒(qiagen,cat#74104)纯化。[0120]用嘌呤霉素将mrna连接至dna接头:采用idt法合成xl-pso寡核苷酸。寡核苷酸序列为:5’‑psoc6-(uagccggug)2’‑omc-aaaaaaaaaaaaaaa-spacer9-spaser9-acc-puro-3’。为了将该寡核苷酸连接到mrna,我们在pcr管中混合以下试剂:29.5μl无rna酶水。1μl1mhepes-koh,ph7.6,5μl1mkcl,2μl25mm亚精胺,0.5μl125mmedta,8μl上一步的mrna和4μl100mmxl-pso寡核苷酸。将pcr管置于pcr机中,加热至70℃5分钟,并以0.1℃/s的速度冷却至25℃。然后,我们将混合物转移到冰上的96孔板上,将365nm手持式紫外线灯放在顶部,并照射该板20分钟。之后,通过rneasymini试剂盒(qiagen,cat#74104)纯化交联rna。[0121]体外翻译:使用pureexpress体外蛋白质合成试剂盒(neb,cat#e6800s)进行翻译。我们在1.5ml试管中混合以下试剂:20μl溶液a、15μl溶液b、0.5μlrnasinplus、4.5μl水和10μl交联rna(1μg/l)。将混合物在37℃下培养2小时。[0122]用his标签纯化肽:我们使用ni-nta磁珠(qiagen,cat#36111)纯化rna肽复合物。用300μl洗涤缓冲液(50mmnah2po4,300mmnacl,10mm咪唑,0.005%吐温20)洗涤100μl珠子,在磁性支架上分离并悬浮在300μl洗涤缓冲液中。将前一步骤中的25μlrna肽复合物添加到洗涤过的珠子中,并在室温下在端对端摇床上孵育30分钟。用洗涤缓冲液洗涤珠子3次,然后在磁性支架上分离,并用50μl洗脱缓冲液(50mmnah2po4、300mmnacl、500mm咪唑、0.005%吐温20)洗脱。[0123]使用oligo-d(t)25磁珠进行纯化:oligo-d(t)25磁珠购自newenglandbiolabs(cat#s1419s)。用500μl洗涤缓冲液i(20mmtris-hcl,ph7.5,500mmnacl,1mmedta)洗涤100μl珠子悬浮液,将其置于缓冲液中并在磁性支架上分离,然后悬浮在50μl洗涤缓冲液i中。将前一步骤中的50μlrna肽复合物与50μl结合缓冲液(100mmtris-hcl,ph7.5,1mnacl,2mmedta)混合,在65℃下加热2分钟,放在冰上1分钟,并与洗过的珠子混合。该混合物在室温下孵育5分钟,然后用500μl洗涤缓冲液i(20mmtris-hcl,ph7.8,500mmnacl,1mmedta)洗涤两次,用500μl洗涤缓冲液ii(20mmtris-hcl,ph7.8,200mmnacl,1mmedta)洗涤一次。[0124]肽环化:在环化缓冲液(20mmtris-hcl,ph7.8,0.00mnacl,3mmα,α”‑二溴间二甲苯(sigma-aldrich),33%乙腈)中,将oligo-d(t)25磁珠结合的rna肽复合物孵育30分钟,并定期振荡。孵育后,用洗涤缓冲液iii(20mmtris-hcl,ph7.8,0.3mmnacl,5mm2-巯基乙醇)清洗珠子,然后用洗涤缓冲液iv(20mmtris-hcl,ph7.8,0.3mmnacl,0.5mmtcep)清洗珠子,然后用洗涤缓冲液i(20mmtris-hcl,ph7.8,500mmnacl,1mmedta)和洗涤缓冲液ii(20mmtris-hcl,ph7.8,200mmnacl,1mmedta)清洗珠子。通过添加30ul洗脱缓冲液(20mmtris-hcl,ph7.8)并在65℃下培养2分钟,然后立即在磁珠架上分离纯化产物,从磁珠中洗脱纯化产物。[0125]亲和性筛选:亲和性筛选在nuncmaxisorp板(thermofischerscientific)中进行。将his-ctdsp1(表2;seqidno:163)溶解于100μlpbs中,转移至maxisorp板孔中,并在室温下在轨道振动筛上孵育2小时。平板孔用pbs清洗两次,并在室温下用酪蛋白(pbsc缓冲液或含1%酪蛋白的pbs)封闭1h,摇动,然后用pbs清洗3次。阴性筛选孔仅通过与300μlpbsc溶液孵育并用pbs洗涤3次来包被酪蛋白。首先将纯化的rna肽复合物添加到这些含有100μlpbs的阴性筛选孔中,并在室温下摇动孵育20分钟,将该溶液转移到含有125μlpbsc的阳性筛选孔(由his-ctdsp1覆盖)中,并在室温下摇动培养1小时。通过向每个孔中添加25μl纯化的his-ctdsp1并孵育3分钟来进行脱靶筛选。随后用pbs清洗3次孵育孔并用于cdna合成。[0126]cdna合成和pcr:我们使用superscriptiii第一链合成系统(invitrogen,cat#18080-051)进行cdna合成。首先,将16μl水与2ul50mm引物p19(表1)和1μldntp溶液混合,并将该混合物添加到maxisorp板中的孔中。接下来,将该板在65℃下孵育5分钟,然后在4℃下冷却1分钟。然后我们将20μl混合物从板转移到pcr管中,并添加20μl反应混合物,其中含有4μl10x缓冲液、8μl25mmmgcl2、4μl0.1mdtt、2μlrna酶和2μlsuperscriptiii逆转录酶。该混合物在50℃下孵育50分钟。然后我们添加2μlrna酶h,并在37℃下孵育试管20分钟。[0127]使用引物p17和p20对在筛选中筛出的强结合物相对应的dna(表1)进行扩增。使用ventdna聚合酶进行扩增。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下20或变性15秒,55℃退火15秒,25个循环,并在72℃下延伸30秒。扩增后,在琼脂糖凝胶上运行pcr片段,根据制造商的方案,使用qiagen凝胶条带纯化试剂盒进行纯化。[0128]ngs数据分析。将每个周期的pcr反应进行下一代测序。我们使用了ampliconezserveicegenewiz。对unigene进行量化,并选择丰度最高的序列进行进一步测试(表3和3a)[0129]表3-rpp变体的蛋白质和核苷酸序列(seqidno在表19中标出)[0130][0131][0132][0133]表3b-rppri变体的蛋白质序列*(seqidno在表19中标出)[0134][0135][0136][0137]实施例4-肽-gst融合物的构建及其表达[0138]作为第一步,我们在pet29载体中克隆了gst蛋白。gst进行了密码子优化,两侧为hindiii和xhoi位点,由idt合成。用phusiondna聚合酶(neb,cat#m0530s)和引物p109和p108扩增(表1)。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸1分钟。扩增后,根据制造商的方案,使用qiagen凝胶条带纯化试剂盒对pcr片段进行凝胶纯化。[0139]使用引入hindiii和xhoi限制位点的引物p12和p14(表1)扩增pet29质粒。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸6分钟。扩增后,pcr片段用qiagen凝胶条带纯化试剂盒进行凝胶纯化,并用hindiii和xhoi限制性内切酶进行消化。所有消化片段用quiagen试剂盒进行纯化,并根据制造商的建议用t4dna连接酶(neb)以适当的组合进行连接。连接片段在10g化学感受态中转化,并如上所述进行测序。gst-rpp的表达构建物如表4所示(seqidno:164)[0140]表4-rpp-gst融合物的核苷酸和蛋白质序列[0141][0142]使用表5所示的引物,通过pcr重新创建所选序列,使用qiagen凝胶条带纯化试剂盒纯化凝胶,用hindiii和bamhi限制性内切酶消化,并在上述pet载体中克隆gst标签。根据制造商的方案,将连接片段转化到10g化学感受态细胞(lucigen)中。将转化细胞接种在含有25μg/ml卡那霉素的lb板上,并在37℃下培养过夜。第二天早上,通过菌落pcr检测菌落是否存在插入物。采集菌落并在20μl无菌0.9%氯化钠溶液中重新悬浮。将1μl该溶液转移到pcr管中,并用taq聚合酶(newenglandbiolabs,cat#m0482s)和30pmol的侧翼引物扩增。每个pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸1分钟。琼脂糖电泳显示扩增产物。将具有正确插入物的克隆接种到含有25μg/ml卡那霉素的培养管中,并在37℃下培养过夜。第二天早上,通过monarplasmidminiprep试剂盒(neb)纯化构建物。如上所述,将构建物转化为bl21(de3)感受态细胞(lucigen)。[0143]肽gst融合物的纯化:将含有所选构建物的大肠杆菌细胞(bl21菌株)在37℃下剧烈摇动过夜。第二天早上,我们在1l烧瓶中向100ml含25μg/ml卡那霉素的lb培养基中添加1ml夜间培养物,并在37℃下摇动培养。当培养物的od600达到0.4时,我们添加iptg至1mm的最终浓度,并在30℃下摇动培养16h。第二天早上,细胞在eppendorf离心机5810r中以最大速度旋转,并在-80℃下冷冻。必要时,从冰箱中取出细胞颗粒,在室温下培养,并用3mlbper蛋白裂解试剂(thermofisher)裂解。如制造商所述,使用谷胱甘肽琼脂糖(thermoscitificcat#16100)纯化肽gst融合物。[0144]表5-用于克隆框内带gst的rpp变体的引物[0145][0146][0147][0148][0149][0150][0151][0152][0153][0154]实施例5-rpp和rpp变体对ctdsp1磷酸酶活性的抑制[0155]结果:图25-在rna显示/蛋白质进化筛选中确定的前51个最丰富的rpp变体按照最丰富(v1)到最不丰富(v51)的肽顺序列在表3中。表5列出了用于制备这些肽的引物。磷酸酶活性筛选(表6,图25)用于评估前51位rppv抑制内源性磷酸化rest肽(tedpspppspplpken)氨基酸861和864处ctdsp1磷酸酶活性的能力。[0156]磷酸化rest肽(tedpspppspplpken)由genescript合成。在室温下,用10mmtrisph8、10mmmgcl2、100nmctdsp1、0.5μm磷酸盐传感器(phosphatesensor,thermofischer)、50μmrest肽和不同浓度的肽gst融合物进行磷酸酶反应10分钟。所有检测均在用水冲洗10次的96孔板(corningp/n3686)中进行。用bioteksynergyhtx在420/27nm激发和485/20nm发射下使用动力学读数进行荧光测定。[0157]筛选显示,与对照组(gst)相比,在1μm时,rpp(v1)抑制磷酸酶活性约60%(表6,图25)。一些变体抑制磷酸酶活性的效果优于rpp(表6,图25)。最有效的抑制剂是v33,其抑制率为100%;v35,其磷酸酶活性约为90%(表6,图25)。[0158]表6-rpp变体对ctdsp1磷酸酶活性的抑制[0159][0160][0161][0162]图26,表13-rpp,seqidno:12,抑制ctdsp1活性,ec50约为20nm,但不影响其他几种磷酸酶的活性。[0163]检测:如上文图26所述,评估rppseqidno:12抑制内源性磷酸化rest肽(tedpspppspplpken)氨基酸861和864处ctdsp1活性的能力。[0164][0165][0166]实施例6-使用monolith(nanotemper)和biacore(gehealthcarelifesciences)评估rpp的结合亲和力。[0167]如实施例2所述,构建并纯化his-ctdsp1(表2;seqidno:162)和gst-rpp(表4;seqidno:164)的表达构建物。ctdsp1(40nm)上的his标签用红色tris-nta染料(20nm)标记,染料与ctdsp1的比例为3:1,通过重力流尺寸排除去除未结合的染料。线性gst-rpp(表4;seqidno:165)与his-ctdsp1(表2;seqidno:163)的结合亲和力是根据制造商的建议,使用monolith(nanotemper)测定的。结合检测的结果如图2所示:在从低pm到0.5μm的几种浓度下,评估rpp(表4;seqidno:165)与his-ctdsp1的结合。对应于线性rpp与ctdsp1结合的kd在130pm下计算。[0168]使用gehealthcarelifesciences的biacore测定线性gst-rpp(表4;seqidno:165)与his-ctdsp1(表2;seqidno:163)的结合亲和力。按照biacore推荐的方案(cat#br-1000-50,gehealthcarelifescience),使用胺偶联将20μl(5g/ml)在10mm醋酸钠(ph值5.0)中的his标记的ctdsp1固定在cm5芯片上(gehealthcarelifescience)。这种纯化产生了约1300ru与cm5芯片结合的蛋白质。然后用hbs-ep缓冲液(ph7.4)清洗cm5芯片,在hbs-ep缓冲液(ph7.4)中以500nm的浓度以30μl/min的流速注入120μl分析物:gst-rpp或仅gst(阴性对照)。结合时间为2分钟,解离时间为3分钟。最后,用10mm甘氨酸hcl(ph1.5)(br-1003-54,gehealthscience)洗涤结合蛋白,以50μl/分钟的流速持续25秒。线性rpp与ctdsp1的结合导致:kd=1.7pm(图3b,阴性对照如图3a所示)。[0169]实施例7-与细胞穿透肽(cpp)和/或促进内涵体逃逸的肽融合的rpp和rppv[0170]合成了几种连接有细胞穿透肽(cpps)和/或促进内涵体逃逸的肽的rpp和rppv。我们使用的cpp和接头列表如表7所示。[0171]表7-允许细胞穿透和从内涵体逃逸的额外肽[0172][0173][0174]实施例8-rpp被间充质祖细胞和坐骨神经内化。[0175]间充质祖细胞(mpc)[0176]结果:在与rpp(seqidno:4)孵育4小时后,然后进行6天的培养,rpp在间充质祖细胞(mpc)中保持内化(图4、c和d),并在6天的培养期内定位在细胞核中(图4、e和f)。[0177]材料和方法[0178]给药和细胞培养:如前所述培养mpc116,将1.0×104个细胞/cm2铺板于24孔板中的玻璃盖玻片上,在37℃下培养过夜,加入100nmrpp(seqidno:4)4小时,然后更换培养基。细胞培养6天,每两天更换一次培养基。在24小时和72小时进行中期mpc采集,以评估细胞内rpp定位。[0179]免疫组化:phalloidin(invitrogen)用于细胞骨架染色,hoechst333429(calbiochem)染料用于显示细胞核,flag抗体(sigma-aldrich)用于显示rpp。图像用共焦激光扫描显微镜拍摄。[0180]坐骨神经[0181]结果:线性rpp积聚在坐骨神经组织的细胞核中(图5、a和b)。[0182]材料和方法[0183]给药和步骤:图5:对sprague-dawley大鼠的坐骨神经(l3和l4区域)(schmitz和beer,2001)117进行暴露,在神经残端回缩后,移除0.7cm的部分,形成约1cm的节段性缺损(hems和glasby,1993)118。将pbs中的1mgrpp(seqidno:4)注入受伤部位,并闭合手术切口。48小时后,处死动物,从l4到l6区域的脊髓冠状切片。[0184]合理剂量水平:最大可行剂量。[0185]供试品标识:rpp(seqidno:4)[0186]纯度:≥95%[0187]免疫组化:hoechst333429(calbiochem)染料用于显示细胞核,flag抗体(sigma-aldrich)用于显示rpp。图像用光学显微镜拍摄。[0188]实施例9-rpp诱导rest蛋白降解。[0189]结果:与溶剂对照组相比,rpp(seqidno:4)使rest蛋白水平降低了58%(图6)。[0190]材料和方法[0191]供试品标识:rpp(seqidno:4)[0192]纯度:≥95%[0193]给药和细胞培养:hek293细胞在添加10%小牛血清的dulbecco改良eagle培养基(dmem)中生长。用100nm全长人rest蛋白2,按照制造商的建议,使用lipofectamine2000(lifetechnologies)在35mm培养皿中80%至90%的汇合培养物中对hek293细胞进行转染。转染后24小时,用100nmrpp(seqidno:4)或溶剂对照(pbs)给细胞灌胃过夜。[0194]蛋白质印迹分析:按照ballas等人,2001的程序制备全细胞裂解物119。使用抗rest-c64,抗gapdh(abcam)及与红外染料结合的抗igg(thermofisher)的标准程序进行蛋白免疫印迹,并在odyssey红外荧光成像仪(licor)上进行分析。条形图是使用imagej对蛋白免疫印迹进行量化(https://imagej.nih.gov/ij)的结果。[0195]实施例10–rpp可用于诱导bdnf、ngf、kv4.3、kv7.2、nav1.8和oprm1mrna的表达,无神经毒性。[0196]结果.rpp增加bdnf、ngf、kv4.3、kv7.2、nav1.8和oprm1的表达(图7、8、9、10、14、15和16;表15)。rpp(seqidno:12)不会导致drg神经元坏死(图17),如在与不同浓度的rpp孵育48小时的drg神经元中进行ldh细胞毒性检测所示。***=p《0.001,n=4,单因素anova和dunnett试验,误差棒=sd。[0197][0198][0199][0200]图7:材料和方法[0201]供试品标识:rpp(seqidno:4),纯度:≥95%[0202]给药和细胞培养:100nmrpp;hek293细胞[0203]mrna提取和定量方案:细胞在qiazol(qiagen)中溶解,根据制造商的说明,使用rneasymidi试剂盒(qiagen)提取总rna。用nanodrop2000(thermofisher)对纯化的rna进行定量,并使用highcapacitycdna反转录试剂盒(appliedbiosystems)和每次反应2ng/l的rna进行反转录。对于qrt-pcr,将5μlcdna(相当于10ngrna)与10μlssoadvancedtmuniversalsupermix(biorad)和1μl每种引物(最终引物浓度各500nm)。在定量研究中,反应quatstudiotm7flex实时pcr系统(appliedbiosystems)重复三次。使用相对循环阈值(δdct)法和β-肌动蛋白作为校准物分析扩增数据。使用的引物如下:β-肌动蛋白正向:agagcacgagctgcctgac-3’,β-肌动蛋白反向:5'-ggatgccacaggactcca-3’,bdnf正向:5’tattagtgagtgggtaacggcg3’,bdnf反向:5’gaagtattgcttcagttggcctt3’。[0204]图8:材料和方法[0205]瞬时转染和细胞培养:nbfl细胞如前所述进行生长120。将0.5μmnfbl细胞接种在35mm培养皿上,在37℃培养过夜,并按照制造商的建议使用lipofectamine2000(lifetechnologies)转染2grest(seqidno:1)-ires-gfpcdna2。培养48小时后,通过荧光激活细胞分选(facs)对细胞进行分选.[0206]mrna提取和定量:如实施例10所述,从gfp 和–细胞中提取mrna;kv4.3的mrna水平采用实时rt-pcr检测116。使用β-肌动蛋白(actb)作为内参管家对照对基因表达归一化。使用的β-肌动蛋白引物与实施例10相同。kv4.3条引物为:正向:ctcactaccacctgctgctc,反向:tcagtccgtcgtctgctttc。[0207]图9:材料和方法[0208]质粒:rpp(seqidno:2和4):含有t7启动子、cpp和his标签的该构建物的一段通过idt合成为gblock(表8),并用表8所述的引物扩增。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸30秒。扩增后,根据制造商的方案,使用qiagen凝胶条带纯化试剂盒对pcr片段进行凝胶纯化。[0209]使用引入sbfi和xhoi限制位点的引物:正向:p12和p14(表1)扩增pet29质粒。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下进行次变性15秒,55℃退火15秒,30个循环,并在72℃下延伸6分钟。扩增后,用qiagen凝胶条带纯化试剂盒对pcr片段进行凝胶纯化,并用sbfi和xhoi限制性内切酶消化。根据制造商的建议,使用qiagen试剂盒纯化所有消化片段,并使用t4dna连接酶(neb)以适当的组合连接。连接片段在10g化学感受态细胞中转化并测序。[0210]rpp的核苷酸序列(表8)由hindiii和bamhi限制性内切酶组装和消化。该片段用cpp和his标签在上述pet载体中克隆。[0211]对照肽:将对照肽(cp)并入表9所示的gblock序列,然后克隆到pet载体中。cp序列如表9所示。[0212]体外表达:rpp(表8)或cp(表9)使用纯表达体外蛋白质合成试剂盒(newenglandbiolabs)在体外表达。使用表8中列出的引物从rpp(seqidno:2和4)或cp表达构建物(表8和9)中扩增rpp(seqidno:2和4)或cp序列,并使用qiagen凝胶条带纯化试剂盒进行凝胶纯化。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸30秒。然后,将15μl纯化的pcr片段与10μl纯表达体外蛋白质合成试剂盒(newenglandbiolabs)的溶液a和7.5μl溶液b混合,并在37℃下孵育2小时。接下来,添加2μldna酶i,并在37℃下孵育20分钟。[0213]表8-rpp-cpp-his_pet盒[0214][0215]agagctatgagctgcctgac,r-ggatgccacaggactcca;kv4.3:f-agctgtgcctcagaactaggcttt,r-taccagaaagacgcagggatgctt;kv7.2f-ccggcagaactcagaagaag,r-tttgaggccaggggtaagat;oprm1:f-ttcctggtcatgtatgtgattgta,r-gggcagtgtactggtcgctaa。[0227]图17:材料和方法[0228]细胞毒性检测:用ldh-glotm细胞毒性试验(promega),根据制造商的说明评估rpp(seqidno:12)的细胞毒性。用rpp处理48小时后收集培养基,或用2%tritonx-100孵育15分钟(可诱导坏死)。将培养基在ldh储存缓冲液(200mmtris-hcl(ph7.3)、10%甘油、1%bsa)中稀释1:50。为了验证检测的线性范围,还将ldh滴定曲线与实验样品一起运行。将50ml稀释培养基或ldh系列稀释液与50μlldh检测试剂(50μlldh检测酶混合物,0.25μl还原酶底物)孵育40分钟。使用infinitem200pro(tecan)仪器记录发光。[0229]实施例11–融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可用于诱导神经分化[0230]方法1:为了确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否最有效地促进人类神经发生和分化,将使用来自ncrm-1/xcl-1ips细胞系的神经干祖细胞(nspc)进行体外筛选检测。通过外体质粒重编程从cd34 人脐血细胞中获得ncrm-1/xcl-1ips细胞121,并通过类胚体(eb)方法分化为神经干祖细胞(nspc)122。ncrm-1/xcl-1神经干细胞是位置性原始神经干细胞,可快速分化为神经元123,使其成为基础广泛的毒理学和表型筛选平台的理想选择124-126。[0231]我们将使用两个工程化ncrm-1/xcl-1系在96孔板检测中进行高通量筛选。为了确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否具有细胞毒性,我们将使用在cmv启动子控制下组成性表达荧光素酶的ncrm-1/xcl-1系。在该系中,通过转录激活物样效应器核酸酶(talens),将cmv纳米荧光素酶halotag(cmv-nlht)构建物插入染色体19q的安全港aavs1位点(图20a)。我们将使用荧光素酶活性作为cmv-nlht细胞数的快速替代读数。使用这种方法,我们将收集时间过程数据,以测定长期暴露于药物的毒性。如果融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽表明毒性水平导致荧光素酶信号丢失≥10%,则这就会通过celltoxtmgreen毒性检测来确认。[0232]为了评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的促神经元分化作用,我们将使用第二个工程化ncrm-1/xcl-1系。在这个细胞系中,使用锌指核酸酶(zfn)将纳米荧光素酶halotag(nlht)敲入map2转录起始位点(tss)(2号染色体)(图20b)。在ncrm-1/xcl-1培养物中,map2表达随神经元分化而增加127,这是通过增加map2-nhlt培养基中的荧光素酶活性来复制的(图20c)。我们将使用培养基中的荧光素酶活性作为map2-nlht神经分化的替代读数,并收集时间过程数据,实时跟踪融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的分化效应。这种方法将允许我们快速比较用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽或对照肽处理的培养物中的神经分化率。将通过对两个同期组的细胞命运标记基因121;一组同期组的nspc和lin28a、rps27l、ifitm2、igfbp3、anxa1的多能标记基因;神经元标记基因c1orf61、iglon5、igsf11、chl1、sox9的qrt-pcr表达筛选,独立验证阳性神经分化效应。荧光素酶活性和qrt-pcr值将报告为平均值±平均值标准误差(sem)。通过单因素方差分析(anova)和tukey的多重比较检验,确定药物对神经元分化率的统计学显著影响。[0233]预测结果和备选策略:我们预测,用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对人nspc给药后,将加速神经元分化,如通过map2荧光素酶活性增加的测定所示的,与溶剂对照组相比,nspc/多能标记物的表达减少,神经元标记物的表达增加。[0234]如果融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽影响转变了ncrm-1/xcls标记时空表达谱,则可能需要在备用时间点收集数据。如果荧光素酶筛选缺乏评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的促分化作用的敏感性,则qrt-pcr数据可用于评估神经分化。[0235]材料和方法[0236]质粒:cmv纳米荧光素酶halotag(cmv-nlht)可从promaga购买。[0237]细胞系和细胞培养:如前所述通过外体质粒重编程从cd34 人脐血细胞中获得ncrm-1/xcl-1ips细胞121,并通过类胚体(eb)方法122分化为神经干祖细胞(nspc)和神经123。通过如papapetrou等人,2016先前描述的转录激活物样效应器核酸酶(talens)128将cmv纳米荧光素酶halotag(cmv-nlht)构建物插入染色体19q的安全港aavs1位点(ncrm-1/xcl-1细胞系)。使用锌指核酸酶(zfn)将纳米荧光素酶halotag(nlht)敲入ncrm-1/xcl-1细胞系的map2转录起始位点(tss)(2号染色体)的方法之前已经描述过127。[0238]荧光素酶检测:测定cmv-nlht细胞数量和神经分化(map2-nlht)的方法之前已由fritz等人,2017描述129。[0239]qrt-pcr筛选:用于筛选细胞命运标记基因的qrt-pcr方法已由chou等人,2011描述过121。[0240]方法2:rpp给药7天后诱导的多能干细胞(神经干细胞(nsc)-nl5)分化(图11(seqidno:5-11,3μm);图12(seqidno:9,1μm)图13(seqidno:13(图27),1μm),使用以下“神经元至空白(neurontoblank)”方案:[0241]1、24孔培养板(黑色visiplate)包被有matrigel(12ml/孔,37℃下30分钟)。[0242]2、nscs-nls(见下文培养基)按每孔0.5×105进行铺板。[0243]组分最终浓度量dmem/12培养基1x97mln2(100x)1x1mlb27添加2%2mlbfgf(制备为100μg/ml母液)10ng/ml10μl[0244]3、24小时后(细胞应》70%汇合),将培养基更换为神经元分化培养基(下文),添加不同浓度的测试肽或水对照(图11、12和13、表14)。[0245]每2天更换一次含有新测试肽和对照的神经元分化培养基,持续5天。[0246]组分最终浓度量d-mem/f-121x88.6mlglutamaxtm-i添加2mm1ml牛血清白蛋白(25%)1.8%7.2mlhesc添加2%2mlbdnf(制备为25ug/ml母液)10ng/ml40μlgdnf(制备微100ug/ml母液)10ng/ml10μl[0247]4、第7天,将细胞用1×pbs中的4%甲醛固定30分钟。[0248]5、用小鼠抗tuj1(1:1000)和兔抗map2(1:500)免疫标记细胞过夜。然后用二抗(抗鼠488、抗兔568和dapi)标记1小时。[0249]6、在biotek平板阅读器上收集数据[0250]a、dapi:激发360/40;发射460/20,增益35;[0251]b、alexa488:激发485/20;发射528/20,增益50;[0252]c、alexa568:激发560/20;发射620/10,增益75)。[0253]结果:在图11中,通过将map2(成熟神经元标记物)表达归一化为dapi(细胞核)并与对照组相比,在7天后,3μm的rppseqidno:5至11增加(1.7至2.7倍)了nsc-nl5的分化。在图12中,根据tuj1和map2神经元标记物(归一化为dapi(细胞核))测定,与对照组相比,1μm的rppseqidno:9在7天后使nsc-nl5分化增加约35%。在图13中,通过tuj1(分别增加60%和3倍)和map2(分别增加27%和2倍)神经元标记物(归一化为dapi(细胞核)并相对于对照组),7天后0.1和1μm的rppseqidno:13增加了nsc-nl5的分化。使用该筛选的基础是,神经祖细胞中rest的消除已被证明可诱导神经元分化22,是神经元基因整体去表达的读数。[0254]实施例12-融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可用于改善创伤性脑损伤后的运动和认知功能。[0255]方法:我们将确定在tbi大鼠模型中,用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽治疗是否会改变神经元存活率和神经发生量。我们将这些指标与组织病理学和功能恢复结果相关联,以确定tbi后rest肽的有效性。我们已经表明,损伤后病变周围的restmrna增加(图19)。我们将使用具有良好特征的受控皮质撞击损伤(cci)大鼠模型6,130-134。[0256]雄性和雌性sprague-dawley大鼠(约250-300g雄性,150-200g雌性)将被cci单侧损伤右侧顶叶皮质(直径5mm,速度4m/s,深度2mm)。假手术动物将进行相同的程序,不会产生影响。cci后6小时,大鼠将在其右心室植入一个套管,该套管将连接至渗透微型泵,用于脑室内导管(icv)输送对照或融合至cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽(0.17μg/h)。六小时是一个具有转化相关性的现实初始治疗窗口,用于确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的疗效,直至处死。我们将在两个不同的时间点处死大鼠–第一组在3dpi时,检查融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对神经元死亡和增殖的急性影响,第二组在30dpi时测定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对功能和形态恢复及神经发生的影响。对于每个同期组,将有12组肽3(对照肽、两个剂量的融合到cpp(seqidno:118至137和159至143)的rpp(seqidno:1和15至17))或rppv(seqidno:18至117))×2(假手术组,cci)×2(雄性/雌性)设计中。雄性和雌性大鼠将在不同的天数进行跑步,以确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的在创伤、行为或反应中的任何性别特异性差异。大鼠在第一周每天腹腔注射brdu(50mg/kg),以确定增殖和成熟细胞的命运(第一同期组-每天注射1-3dpi,最后注射后2小时处死;第二同期组-每天1-7dpi)。我们之前的经验表明,我们需要16只大鼠/组获得重要的行为数据(第二同期组),8只大鼠/组获得免疫组化(第一同期组)。[0257]在第二个同期组中,为了确定与对照组相比,用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽治疗的功能结果,我们将检查cci或假损伤后不同时间点的运动和认知行为。我们将在大鼠受伤前对其进行训练,以在平衡木行走(beamwalk)、滚轴(rotorod)和旷场试验中获得运动功能的基线读数。大鼠将在伤后1、3、7和10天(dayspost-injury,dpi)在平衡木行走和滚轴上测试运动功能的恢复情况。旷场试验将在的第4、12和22dpi进行。识别记忆将通过在20dpi下进行的新型物体识别测试进行评估。数据将通过重复测定双因素方差分析和dunnets事后校正进行分析。空间记忆和学习将由morris水迷宫(mwm)检测,在25dpi开始。所有试验都将记录游泳速度和找到隐藏平台的时延。在训练的第五天进行的探测试验将确定大鼠在之前包含隐藏平台的象限中所花费的时间。还将对每只大鼠进行可见平台测试,以确保动物之间的视力差异很小。带有tukey多重比较校正的双因素方差分析将确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否与对照肽相比显著改变假手术或受伤大鼠mwm中的认知功能135。[0258]功能评估完成后,经心灌注处死8只大鼠/组,取脑进行处理。我们将通过甲酚紫染色切片,并测定以500μm为间隔的病灶的12个切片,来确定3和30dpi时病灶的大小。病灶体积将表示为同侧半球体积的百分比。将检查穿过额顶皮质和背侧海马(包括脑室下区(svz)和齿状回(dg))的一系列冠状脑切片。将使用体视学对不同海马区域和svz中的brdu阳性细胞总数(用抗brdu检测)进行计数,以确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否会改变损伤后新增殖细胞的存活率。切片将与不同的细胞特异性标记物共同染色,以确定3dpi时增殖细胞的特性及其在30dpi时的成熟命运。细胞特异性标记物包括neun(成熟神经元)、sox2(神经干细胞)、双皮质素(doublecortin,dcx)(神经元祖细胞)、gfap或aldh1l1(星形胶质细胞)、iba1(小胶质细胞)、ng2(少突胶质前体细胞,opc)和apc或gstpi(成熟少突胶质细胞)。将比较不同处理组之间在30dpi时dg和嗅球中brdu/neun双阳性细胞的数量,以确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否增加损伤后神经发生。我们还将确定是否存在brdu 神经元祖细胞或成熟神经元向海马不同区域的迁移-远离dg,或从svz向病灶区或吻侧迁移流(rms)。除神经源性凹部(nich)外,检查病灶周围区域内每种brdu 细胞类型的总数,将确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否会改变损伤后的病理学。由于我们预计融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽将增加神经元存活率,因此我们将量化两个时间点病变周围区域中退化神经元(flurojadec)的数量和存活神经元(neun)的数量。我们将用luxolfastblue检查胼胝体,以显示髓鞘数量的任何差异。无偏光学分馏器法将用于对神经发生区、病变周围区、吻侧迁移流和嗅球的细胞进行计数,以获得准确的细胞特异性和增殖细胞计数。所有数据将通过dunnetts事后校正的双因素方差分析进行分析。将通过flag抗血清染色,在切片中检查融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的分布。每组剩下的8只大鼠(第二同期组)将被处死,并迅速移除它们的大脑。病灶同侧和对侧的大脑区域将被打孔并快速冷冻以分离rna和蛋白质。我们将通过蛋白免疫印迹检测rest水平,并通过qrt-pcr检测病灶周围或神经源性凹部中bdnf的表达,与对照组大鼠大脑进行比较。[0259]预测结果和备选策略:我们预计融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽将通过提高神经元存活率来减少3dpi和30dpicci后的病灶体积。将在两个时间点检查退化和存活的神经元,以直接评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽处理对神经元存活的影响。融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽在病灶周围区域早期神经保护作用可能会在第一周改善运动功能。虽然我们之前已经发现小鼠cci后3dpi时神经元死亡最多136,我们可能需要检查大鼠体内的不同时间点。我们还预计,与对照肽相比,用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽治疗的动物将显示出更多的神经发生。我们的数据将表明,在3dpi时,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否会导致不同脑区dcx 前体的增加,或在30dpi时海马、嗅球或病变周围的brdu /neun 成熟神经元的增加。我们将能够将这些数据与行为结果相关联,以观察使用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的治疗是否会提高mwm检测或新物体识别检测的表现135。通过这种方式,我们将能够确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否对炎症或细胞增殖有任何负面影响。[0260]材料和方法[0261]测试系统[0262]物种:雄性和雌性sprague-dawley大鼠。[0263]研究开始时的年龄:约为22至29周。[0264]研究开始时体重:雄性约250至300克,雌性约150至200克。[0265]研究的设计:对于每个同期组(1和2),将有12组3(对照肽、两个剂量的融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽)×2(假手术组,cci)×2(雄性/雌性)设计中。在同期组1(免疫组化)中,将有8只大鼠/组,在同期组2(行为)中,将有16只大鼠/组。[0266]合理剂量水平:高剂量将是最大可行剂量(约0.17g/h),低剂量将是基于脑脊髓液(csf)药物水平的估计有效剂量,其近似于实施例13中确定的有效剂量。[0267]施用途径:植入右心室的套管与渗透微型泵相连,用于脑室内导管(icv)输送。[0268]施用频率:持续输液直至处死。[0269]给药周期:同期组1:3dpi;同期组2:30dpi。[0270]环境条件:2只大鼠/笼,随意进食饮水。[0271]供试品标识:融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽[0272]纯度:≥95%[0273]剂量配方的制备:每4天一批(bulk)。[0274]剂量配方检测和稳定性:将在研究第一周的第1天和第7天评估剂量配方的稳定性和浓度。可接受的浓度范围为标称浓度的±10%。[0275]受控皮质撞击(cci)模型:cci模型之前已被描述6,130-134。融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽或对照(仅溶剂)的施用将在cci后6小时开始。[0276]免疫组化:xiong,y等人,2007和2008对这些方法进行了描述135-137。[0277]morris水迷宫:choi等人,2006之前描述了这些方法138。[0278]实施例13-融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可用于改善周围神经损伤后的再生。[0279]方法:周围神经损伤(pni)后的功能恢复需要营养因子(ntf),它们的存在或缺乏是再生反应强度的生物标志物139,140。已知几种ntf基因,包括脑源性神经营养因子(bdnf)、神经生长因子(ngf)、pleitrophin(ptn)和神经营养因子-3(ntf-3),被损伤诱导的rest表达所抑制,我们已经证明rpp(seqidno:2)可以逆转这种转录抑制,至少在bdnf和ngf的情况下是如此(图7、9、10和16)141-142。[0280]1)实时定量逆转录pcr(qrt-pcr)分析表明,nbfl细胞注射1μm线性(seqidno:4)或环化(seqidno:2)rpp16或48小时后,引起bdnf和ngfmrna表达增加(图9a和b及下表)。[0281][0282]2)48小时后,与水对照相比,rpp(seqidno:9,3μm)使ntfsbdnf和ngf分别增加约2倍和3倍。这一发现证实了rpp在mpc中是活跃的(图10)。如gerevasi等人,2020所述,将mpc在通用培养基中培养76。[0283]3)我们将对融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对从雄性和雌性肌肉骨骼创伤患者分离的间充质祖细胞(mpc)中ngf、bdnf、ptn和ntf-3表达的影响进行量化。mpc存在于周围神经损伤部位,被广泛表征143。mpc将使用leonnesti博士制定的标准分离方案,在正常和预先计划的手术处理过程中采集116,143。来自3个不同受试者的mpc将通过培养进行扩增。首先,我们将在无神经元诱导的情况下传代细胞四次,以通过qrt-pcr和蛋白免疫印迹(wb)建立rest和ntf转录物和蛋白质水平表达的基线。接下来,我们将重复bulkenhoover等人所描述的神经元诱导扩张144。简而言之,mpc在预诱导培养基中培养2天,然后添加全反式维甲酸(ra)1天,然后在神经诱导培养基中培养7天。神经诱导培养基每三天更换一次。诱导后第7天,将评估rest和ntf水平。我们预计,神经元诱导应导致rest表达减少,ntfs表达相应增加。[0284]评估融合到cpp(seqidno:118至137和159至143)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽诱导mpc中ntf表达的效力,我们首先需要通过评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的细胞毒性来确定剂量。简言之,mpc将以4个剂量水平(最高10μm)在用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽替代ra的改良预诱导培养基中或在预诱导培养基(含ra)中进行铺板1天。每个给药组将有三个技术和三个生物重复,并将使用mtt检测来测定细胞毒性145。为了确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的最佳浓度,以降低rest水平和增加ntf水平,我们将在处理后1、3和6天(dayspost-treatment,dpt)用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽替代ra的神经诱导培养基中进行剂量反应(3个剂量水平)。融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的剂量将基于mtt分析的结果145。我们的阴性和阳性对照将分别为神经诱导培养基或 ra。我们将用qrt-pcr和wb对rest和ntfs的基因表达和蛋白水平进行定量139,146。将在1、4和7dpt收获细胞,并从细胞裂解物中分离rna和蛋白质。将进行双因素anova分析,然后进行tukey的事后检验,以在指定时间点比较–和 对照组与融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽给药组。总体平均值将报告为平均值±标准差,p值为0.05或更小,具有统计显著性。将使用sas统计软件包(sasinstitute,carync)进行统计分析。[0285]4)在整个drg神经元(l5,来自雄性sd大鼠)的离体培养物中,在0、0.3、1、3、10μm处评估rpp(seqidno:12)诱导bdnf和ngf表达的潜力(图16)。rpp在0.3、1和3μm时增加bdnf表达约25%,在10m时增加75%。rpp在3和10μm时分别增加ngf约50%和2倍(图15)。在1μm处未观察到任何影响,0.3μm因n=1而不确定(图16)。[0286]5)在体外培养的整个drg神经元(l5,来自雄性sd大鼠)上进行的ldh细胞毒性检测中评估rpp(seqidno:12)。rpp在0、1、3或10μm时没有显示毒性(图17)。正如预期的那样,阳性对照组(2%triton)具有神经毒性,增加了90000rlu。[0287]6)我们将评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽通过分别使用脊髓和分离的背根神经节(drg)外植体,在运动和感觉离体神经模型中诱导ntf表达并刺激再生的能力147。这两种模型广泛用于研究生长因子的神经保护和营养特性148。将从脊柱两侧(l4-l6)提取(6只大鼠)一段10mm的完整drg并进行培养(图21和22)149,150。剩余的脊柱材料将被保存下来,用于运动神经元连接的脊髓切片培养。这些脊柱将用切片机以300μm的间隔横向切片,切片将被转移到带有半透膜的培养插入物中,并在培养条件下适应一周149,150。插入一周龄的器官型脊髓切片的培养物(具有稳定的存活运动神经元群),之后将被转移到6孔板中再培养7天。[0288]两种离体外植体培养物将在neurobasal/b27培养基中保持2周,以允许神经元过程的广泛延长147。为了诱发物理损伤,将使用玻璃巴斯德吸管在距离drg神经节周长6mm处和脊髓腹侧切片的灰白质交界处的精细神经元突起上划伤。体外培养物将在损伤后24小时开始用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽(0、1、3或10μm)处理。[0289]受损神经突(运动和感觉)的再生能力将通过监测细胞活力和测定轴突生长来确定。通过分别用钙黄绿素am和溴乙啡锭二聚体1标记活细胞和死细胞来评估7dpt时的细胞活力。神经突延长是神经元再生的一个指标,将在1、4和7dpt时进行测定。将使用与单色数码相机连接的zeissaxioobserver显微镜,获取划痕区域的灰度显微照片151。将使用imagej分析划痕区域内再生芽的数量、长度和总面积152。[0290]ngf通过其受体,即drg中的trka受体发挥其大部分功能活性153,ptn主要在运动神经元中引起轴突生长增加150-154。bdnf和ntf-3存在于运动神经元和感觉神经元中。我们将用qrt-pcr和wb对ntf的基因表达和蛋白水平进行定量139,146。在1、4和7dpi采集每个离体系统的组织,并分离rna和蛋白质裂解物。将进行双因素anova分析,然后进行tukey的事后检验,以在指定时间点比较对照组和融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的给药组。总体平均值将报告为平均值±标准差,p值为0.05或更小,具有统计显著性。将使用sas软件(sasinstitute,carync)进行统计分析。[0291]7)将在100只大鼠体内坐骨神经缺损后,评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽改善再生和功能恢复的能力。我们已经证明,在坐骨神经损伤部位注射rpp48小时后,rpp在脊髓坐骨神经神经元的细胞核中积聚(图4)。将通过静脉(iv)施用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,这将允许我们测试比肌肉内(im)或皮下(sc)施用更高的最大可行剂量(mfd)。静脉注射也可避免与口服和腹腔注射(ip)相关的首过代谢。再生和功能恢复研究的剂量浓度将基于sprague-dawley大鼠一个月单次iv剂量范围发现毒性和毒代动力学研究的结果。将在完成1)后启动剂量范围判定的毒性研究。简而言之,该研究将使用四种剂量水平,最高1000mg/kg或最大耐受剂量或mfd,如fda指南(m3(r2),2009)所建议。每组分配的动物数量如下:主要研究:10只/性别/组;tk:3只/性别/对照组和9只/性别/给药组。将评估动物的体重、食物消耗、临床化学、血液学、尿液分析、器官重量、组织病理学(8个核心组织)和毒代动力学。[0292]在再生和功能恢复研究中,spraguedawley大鼠将每周使用长期静脉导管155,通过静脉注射融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,3个给药组(tbd) 对照组(10个/性别/组)治疗6周。所有动物在暴露坐骨神经前都将被麻醉156,在神经残端回缩后,移除0.7cm的部分,以形成约1cm的节段性缺损157。神经将使用脱细胞神经移植修复。然后闭合手术切口,让神经再生6周。只有给药组施用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽。在6周期间,我们将评估功能恢复和对融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的剂量耐受性。每周对大鼠称重,记录温度,并进行脚趾伸展反射检测(以确定受伤腿部的最大足迹宽度,测试运动神经功能)156。步态分析(digigait)将每2周进行一次,并进行电生理研究158。简而言之,将对大鼠坐骨神经进行电生理测试。采用单脉冲电击(1ma,0.1ms)对移植物缝合点附近5mm处的本地坐骨神经干施加电刺激。记录腓肠肌腹部的复合肌肉动作电位(cmap),从1v到12v,或直到达到超最大cmap。还记录了未手术对侧坐骨神经的正常cmap进行比较。恢复率将由受伤后肢的cmap与对侧正常后肢的cmap的比率决定159,160。[0293]在第6周结束时,将收集坐骨神经、脊髓腹角(vh)以及与l4至l6神经根相关的drg161,162。将对神经再生进行组织学评估。将移植物的近端、远端和中心进行准备以使透射电子显微镜(tem)通过横截面,评估移植物沿线的髓鞘形成163,164。简而言之,将使用超薄切片机从神经上切下约70nm的超薄切片,然后用醋酸铀酰和柠檬酸铅染色。将从随机切片中选择10-15个场进行分析,量化有髓纤维的数量和大小。平均纤维密度的计算方法如之前所述73。除tem外,还将准备更多来自同一神经的切片,用于ihc可视化β-微管蛋白iii以测定轴突的直接生长,用s100以测定雪旺细胞对移植物的渗透和生长,用血管性血友病因子以评估愈合神经的血管生成和毛细血管浸润,以及用luxolblue染色165,166测定移植物内髓鞘的形成。将评估受损运动神经和感觉神经细胞体内腹角和drg与融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽相关变化。将使用光学显微镜拍摄染色组织的图像,并使用imagej软件对解剖匹配组织中阳性反应的强度和面积比例进行量化(https://imagej.nih.gov/ij),比较各组之间的神经再生率165。将如之前所述,对受损神经的drg神经元进行计数,并与对侧未受损drg进行比较167。ntfs(bdnf、ngf、ptn和ntf-3)、ctdsp1和rest将通过qpcr和wb进行检查。[0294]预测结果和替代策略:我们预计,与神经诱导培养基ra相比,经融合至cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽 神经诱导培养基(-维甲酸,ra)处理的间充质祖细胞(mpc)将增加神经营养因子(ntf)的表达并降低rest水平,但与神经诱导培养基 ra类似。我们预计,与未治疗组相比,经rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽融合到cpp(seqidno:118至137和140至159)治疗的离体模型,由于rest水平降低,将显示神经突生长增加。在缺乏支持细胞的情况下,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的再生效应可能不会显著,支持细胞可在神经元损伤后在体内分泌神经营养因子。另一种实验将包括在受伤之前以及随后的融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的治疗之前,神经支持细胞(如雪旺细胞和间充质干细胞)与背根神经节(drg)和脊髓外植体(富含运动和感觉神经元)的共培养。接下来,我们将分析融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽帮助大鼠模型中切断的神经再生的能力。我们预计,与对照组大鼠相比,经组织学、电生理学和功能评估确定,经融合至cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽治疗的大鼠的恢复情况将有所改善。我们还期望融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽增加ntf表达并降低rest蛋白水平。如果我们没有观察到rest阻断恢复和解除的迹象,我们将通过直接注射到靶点(即移植物部位或鞘内),增加融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的局部浓度。[0295]材料和方法[0296]细胞培养:mpc的获取、培养和神经营养诱导之前已有描述168。简而言之,mpc在预诱导培养基中培养2天,然后添加全反式维甲酸(ra)1天,然后在神经诱导培养基中培养7天。神经诱导培养基每三天更换一次。入职后第7天,将评估rest和ntf水平。[0297]qrt-pcr:示例10中描述的方法。[0298]蛋白质印迹分析:按照ballas等人2001年119的程序制备全细胞裂解物。使用抗rest-c的标准程序进行蛋白质印迹64,抗ntf(使用市售抗体)、抗gapdh(abcam)和与红外染料结合的抗igg(thermofisher),并在奥德赛红外荧光成像仪(licor)上进行分析。[0299]供试品标识:rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽融合至cpp(seqidno:118至137和140至159),纯度≥95%[0300]剂量配方的制备:每4天一批。[0301]剂量配方和稳定性:将在研究第一周的第1天和第7天评估剂量配方的稳定性和浓度。可接受的浓度范围为标称浓度的±10%。[0302]脊椎动物:雄性和雌性成年sprague-dawley大鼠(约250-300g雄性,150-200g雌性)将用于确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽在改善pni恢复方面的功效。这项研究将需要100只8-11周大的大鼠。大鼠将从charlesriverlaboratories获得。[0303]步骤说明:[0304]手术:在所有手术之前,用异氟烷(4%诱导,3%维持)麻醉大鼠。在2)器官型脊髓培养物将使用前述技术从出生后第8天大鼠(sprague-dawley)的腰脊髓制备(rothstein等人,1993;corse等人,1999)。简而言之,将迅速处死大鼠,移除腰骶脊髓,并将其置于含有葡萄糖(6.4mg/l)的gey平衡盐溶液(gibco)中。在使用无菌技术的层流罩中,在放大镜下仔细切除脑膜,并切断腰脊神经根。将脊髓(cord)放置在aclar膜上,并用mcilwain组织切割器从l2到l5以300μm的间隔进行切片。将使用gey的平衡盐/葡萄糖溶液,小心地将单个脊髓切片转移到6孔培养板中的millicellcm(微孔)渗透膜上。将在每个膜上放置五段线。每个孔含有1ml培养基,由50%最低基本培养基加25mmhepes组成;含d-葡萄糖的25%hanks平衡盐溶液(25.6mg/l);25%热灭活马血清;和2mml-谷氨酰胺。培养物将在37℃、5%co2的加湿培养箱中保持一周,每3天更换一次培养基。一周后,切片将被转移到另一个6孔培养板上,并在任何治疗之前在培养中稳定和延长神经突起7天。[0305]在3)中我们将暴露坐骨神经117(图21和22)并移除0.7cm的部分,以在神经残端收缩后形成约1cm的节段性缺损(hems和glasby,1993)。神经将使用脱细胞神经移植修复。然后闭合手术切口,让神经再生6周。第一组(对照组)将进行约1cm的坐骨神经切除,并将移植1cm脱细胞同种异体神经。第二组至第五组将接受1cm坐骨神经切除术,移植1cm脱细胞同种异体移植物,每周静脉注射融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,药物浓度由剂量范围判定确定。所有动物将在第6周处死。[0306]行为评估:我们将对动物进行长达6周的监测,以评估功能恢复和对肽治疗的耐受性。每周对大鼠称重,记录温度,并进行脚趾伸展反射检测(以确定受伤腿部测试运动神经功能的最大足迹宽度)117。步态分析将每2周进行一次。6周处死前进行电生理检查。[0307]电生理学评估:电生理学测试将按照之前的方法进行158。简言之,重新暴露大鼠坐骨神经,并在移植物缝合点附近5mm处对天然坐骨神经干施加电刺激(单脉冲电击,1ma,0.1ms)。在腓肠肌腹部记录1v至12v的cmap,或直到达到超最大cmap。还记录了未手术对侧坐骨神经的正常cmap进行比较。grasstechs88x刺激器(astromedinc.)用于测试,polyviwe16数据采集软件(astromed,inc.)用于记录。恢复率是大鼠受伤后肢的cmap与对侧正常后肢的cmap之比158。[0308]处死:大鼠于伤后6周处死。处死后,我们将用组织学方法评估轴突通过移植物的生长、再髓鞘化、损伤部位的mpc细胞活性、前角细胞和drg活性以及再生神经的血管化。我们将在每只大鼠处死后采集腓肠肌和胫骨前肌,以评估总重量来量化萎缩。[0309]合理解释:pni后,脉管系统、免疫系统和周围神经系统内所有细胞和分子之间存在复杂的相互作用。我们不完全了解创伤性损伤后不同细胞之间、信号分子、细胞-细胞相互作用和回路的顺序或相互作用,因此目前无法在体外对其进行建模。此外,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可能会影响神经前体细胞、成年神经元以及其他细胞类型,因此在pni的情况下对其进行体内评估至关重要。fda还要求在允许生物制剂的ind状态之前提供体内疗效证明,因此这为这些实验的体内性质提供了进一步的理由。之所以使用大鼠,是因为在pni表明其在这些类型的实验中的可靠性后,进行了大量的行为研究。因此,有大量文献为实验的设计和执行提供了详细的框架。大鼠比小鼠更聪明,体型也更大,能够更好地辨别受伤后功能恢复的变化。与小鼠相比,大鼠生理学与人类有更多的相似之处,从而使实验数据具有更好的转化相关性。此外,我们针对的rest通路在大鼠中是保守的。[0310]最大限度地减少疼痛和痛苦:用异氟醚(3-4%)麻醉大鼠,以治疗节段性周围神经缺损。手术后,动物将被允许在加热垫上恢复,直到移动。手术后,将给大鼠服用对乙酰氨基酚(饮用水中6毫克/毫升),持续2天,此外,如果观察到疼痛或痛苦的迹象。实验室和兽医人员每天都会对老鼠进行观察,并根据兽医的建议对被认为处于困境的动物进行治疗或安乐死。[0311]安乐死:使用的安乐死符合美国兽医协会动物安乐死指南的建议。[0312]实施例14-融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可用于预防和治疗慢性疼痛。[0313]完成的评估:[0314]神经损伤后rest的激活导致感觉神经元正常兴奋性所需的几个基因表达减少,包括钾通道kv4.3(kcnd3)和kv7.2(kcnq2),钠通道nav1.8(scn10a)和μ阿片受体oprm160,72-74。阻断rest以缓解慢性疼痛的基础是利用小鼠和大鼠周围神经损伤(pni)模型发表的公开研究60,72-74。[0315]1)实时定量逆转录pcr(qrt-pcr)分析表明,nbfl细胞注射1μm线性(seqidno:4)或环化(seqidno:2)rpp16或48小时后,会引起kv4.3mrna表达增加(下图9c和表16)。[0316][0317]4)rpp(seqidno:12;0(水)、1、3或10μm)在整个drg(l5,来自雄性sd大鼠)的体外培养物中孵育48小时,并评估其诱导慢性疼痛相关基因kv4.3、kv7.2、nav1.8和oprm1表达的潜力(图14)。在10μm时,与对照组相比,rpp增加kv4.3表达2倍;在3μm和10μm,分别增加kv7.2表达7.5倍和9倍;在3μm和10μm,分别增加nav1.8的表达2.9和4.8倍;在3μm和10μm,分别增加了oprm1的表达(图14)。在整个drg(l5,来自雄性sd大鼠)的离体培养物中,在3μm的48小时给药期后评估rppseqidno:13和14,并评估nav1.8。序列13和14增加nav1.8表达分别为对照组的6倍和13倍(图15)。[0318]5)在体外培养的整个drg神经元(l5,来自雄性sd大鼠)上进行的ldh检测中,rpp(seqidno:12)没有神经毒性。与对照组相比,rpp并没有增加rlu水平。正如预期的那样,阳性对照组(2%)的rlu增加了90000倍(图17)。[0319]计划评估:[0320]目标1:确定融合至cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽药代动力学、分布、以及剂量[0321]子目标1.1:用于检测大鼠和猴血浆、脑脊液和组织中融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽(范围在10至10000ng/ml之间)的液相色谱-串联质谱(lc-ms/ms)方法的开发和验证。[0322]子目标1.2:为了确定体内动物研究和临床中的施用途径,将在sd大鼠中进行探索性药代动力学(pk)研究。在静脉注射、sc注射或po注射1000mg/kgrpp(seqidno:1和15至17)或rppv(seqidno:18至117)后,在5分钟、30分钟、4小时、8小时、24小时和48小时(2个性别/时间点)采集血浆样本,并评估峰值药物浓度(cmax)、曲线下面积(auc)和半衰期(t1/2)。为了评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对中枢和外周神经系统的渗透性,还将在安乐死时收集靶组织(csf、脑、腰脊髓和背根神经节(drg)),并测定药物浓度。将使用经验证的ls-ms/ms方法(子目标1.1)检测融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽。[0323]子目标1.3:为了确定大鼠疗效研究(目标2)中的剂量,将使用子目标1.1中确定的最佳施用途径对sd大鼠进行单剂量范围判定。根据子目标1.1中的pk数据,将选择四个剂量水平 溶剂对照(3个/性别/组)。高剂量(hd)应确定极限剂量(最大可行剂量(mfd))、最大耐受剂量(mtd)和/或暴露饱和)。较低剂量将以1/3的增量间隔。将每天评估动物的存活率和临床症状(异常和疼痛或窘迫症状)。将在给药后的6个时间点采集血液,这些时间点是根据子目标1.1的pk数据选择的,并评估cmax、auc和t1/2。将在安乐死时收集将在疗效研究中评估基因变化的组织(drg、腰脊髓、大脑),并评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的浓度。[0324]目标2:评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对慢性疼痛的影响。[0325]我们已经证明,在坐骨神经切断部位给药48小时后,rpp在大鼠腰椎(l4–l6)脊髓运动神经元的细胞核中累积(图4)。为了评估我们的药物对疼痛的影响,我们将使用坐骨神经的临床前sni模型。sni是一种公认的神经病理性疼痛动物模型,可产生强烈和持久的热敏感性变化、外周和中枢吗啡镇痛以及c纤维感觉减退60,72,74。此外,正如我们预测的那样,我们观察到ctdsp1水平增加(图18)。[0326]融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽减少慢性疼痛的功效将在第1阶段完成,疼痛减轻的持久性将在第2阶段确定。sni坐骨神经大鼠模型将用于测试融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否能减少慢性神经病理性疼痛。将施用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,或用于治疗神经病理性疼痛的溶剂或标准护理药物羟考酮。下表17总结了第1阶段的实验步骤:[0327]表17[0328][0329][0330]为了实现这一目标,总共需要312只2个月大的sd大鼠(雄性和雌性数量相等)。每组12只大鼠的样本量将有80%的效力检测疼痛评估中标准差1.20的差异。效力分析基于独立样本的t检验,具有5%的双侧显著性水平。对于疗效研究,将大鼠随机分配到右侧表格中指定的实验组(n=12/组)。对10个随机分组的大鼠进行刺激诱发和非刺激诱发行为测试、形态学、免疫组化分析、定量rt-pcr(qrt-pcr)分析和wb分析。安乐死时,将在开始给药后的2个时间点(14天和28天)抽取血液,并评估标准pk参数。提取drg和大脑以确定oprm1、nav1.8,kv4.3和kv7.2水平。此外,将收集组织进行lc-ms/ms分析,以确定rpp浓度。[0331]手术:sni或假手术(暴露神经分支而不损伤神经的手术)将按照之前所述进行169。简而言之,将麻醉大鼠,并结扎腓总神经和胫总神经。将切断并移除结扎远端各神经的2-4mm段,保留坐骨神经的第三支腓肠神经完整。腓肠神经在损伤后4天开始对同侧神经支配区的伤害性和非伤害性刺激产生增强的反应,7天稳定,维持6个月。损伤后7天是慢性神经病理性疼痛发展的公认时间点,并且具有可转化相关性。[0332]药物/溶剂:手术后第5天开始施用。对于融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,测试的途径、频率和药物浓度将基于特定目标1中的pk研究。对于羟考酮,大鼠的给药途径和有效药物暴露(接近人类的有效暴露):(0.56mg/kg剂量的羟考酮和0.2ml溶剂)是已知的170。[0333]行为测试:将在手术前一天和从术后第4天开始每隔一天评估对疼痛传递和运动功能的影响。与羟考酮和溶剂相比,刺激诱发和非刺激诱发行为试验将用于确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的功能有效性。近年来,对一些刺激诱发行为测试转化性的关注导致刺激诱发、非刺激诱发和操作性或自愿性行为测试的使用越来越多。对于这一提议,我们将使用最广泛使用和可转化的刺激诱发行为测试:电子vonfrey和热痛觉过敏测试、非刺激诱发步态分析和条件性逃避/回避试验(placeescape/avoidanceparadigm)来测定疼痛的厌恶状态171。最近的研究表明,在动物模型中加入步态分析来检测细微的改善和恶化是有价值的172。进行行为测试的大鼠处理人员和评估人员将被随机分配给对应的实验/对照组。每只动物都有一个唯一的动物编号。将使用秘密颜色代码指定对照组和实验组。只有在分析之后,才会显示此代码。[0334]刺激诱发行为测试:对机械刺激的超敏反应:将使用一种电子vonfrey(bioseb,查维尔,法国),由一个配有塑料尖的手持式力传感器组成,用于测试左、右后爪的机械后退阈值。力传感器的塑料尖将应用于爪子的内侧足底表面以测试隐神经,并应用于爪子的外侧足底表面以测试坐骨神经(腓神经)。[0335]热痛觉过敏:将根据一种已公布的方法进行测定173,我们将该方法修改为使用激光作为热源。将使用808nm波长的激光器作为热源(2w输出功率,皮肤上的光束直径=3mm),并将其应用于被测神经的相关皮肤。小光束尺寸允许相关皮肤小体被专门定位。对于坐骨神经和隐神经,将以10秒为分界点记录热刺激引起的肢体退缩时延。[0336]非刺激诱发行为测试:步态分析:使用digigait和坐骨神经指数评估坐骨神经运动功能172。简而言之,利用rodentbehavioralcore中可用的digigait设备(我们的实验室已对其进行了表征和优化),将在损伤前和术后第7、14和28天测定坐骨神经功能。将大鼠放置在digigait支架内的电动跑步机上,并设置20cm/s的速度记录所有动物。坐骨神经指数程序将用于分析受影响后肢的功能。[0337]条件性逃避/回避:该步骤旨在验证以下假设,即大鼠将避免与对痛觉过敏区域的机械刺激相关的环境。据报道,在使用止痛药治疗后,受伤大鼠对逃避/回避行为的变化具有敏感性(baastrup等人,2010)174。labuda和fuchs(2000)171描述了大脑依赖的条件性逃避/回避试验测试,该测试依赖于对新厌恶环境的逃避/回避学习。受伤后3至5周,由同一名受训研究人员仅对动物进行一次测试。测试将在usurodentbehavioralcore中进行。将允许动物在试验室中适应环境光线和噪音条件至少1小时。在本试验中,大鼠可自由进入封闭室的“非厌恶性”黑暗侧和“厌恶性”光明侧,封闭室具有网状地板,可通过vonfrey丝从下方轻松进入。如果大鼠分别处于黑暗或光明区域,受伤或未受伤的后爪都会受到常规刺激。每15秒,根据动物在此期间的位置,将足以在受伤的爪子上引发撤退反应的机械刺激(vonfrey丝)施加到动物一只后爪的外侧足底表面,持续30分钟。逃逸/回避行为将定义为从黑暗区域向光明区域的转移。将记录在方框白色侧花费的时间百分比以及黑白侧之间的交叉次数。白色侧的累积时间和交叉总数将用作逃避/回避学习的指示。[0338]安乐死:为了进行形态学和免疫组化分析,将用氯胺酮/甲苯噻嗪(80至100mg/kg 10mg/kg,i.p.,21号针)麻醉大鼠,然后用300ml磷酸盐缓冲盐水(ph7.4),然后是0.1m磷酸盐缓冲液中的300ml4%多聚甲醛经心灌注。灌注后,解剖神经、drg和脊髓,在4%多聚甲醛中固定24小时,在30%蔗糖中冷冻24小时。使用低温恒温器(leicacm3050s,leicabiosystems,wetzlar,germany)将组织样品切割成10μm的切片。[0339]组织学:安乐死和神经取样后,切片将用苏木精和曙红染色。其余靶基因oprm1、nav1.8、kv4.3和kv7.2的表达水平和分布将通过免疫荧光进行评估。[0340]基因表达分析:对于坐骨神经及其相关drg和脊髓节段的基因表达分析,rest靶基因oprm1、nav1.8,kv4.3和kv7.2和对照基因(hprt、gapd、rn18s)将通过qrt-pcr进行评估。[0341]蛋白质表达分析:坐骨神经相关drg的蛋白质表达分析将通过oprm1、nav1.8、kv4.3和kv7.2的sds-page免疫印迹进行评估。gapdh将用作加载对照。[0342]统计方法:对于行为数据,结果将以平均值±平均值标准误差(sem)表示。基于治疗后的时间,将使用双因素方差分析来评估各组动物。bonferroni-holm方法将用于调整多重比较。双尾统计显著性将被确定为p《0.05。对于免疫组化分析,将使用双因素anova来比较交互作用的平均值以及组和时间的影响。对于qrt-pcr,值将报告为平均值的平均值±标准误差(sem)。通过单因素方差分析(anova)和tukey的多重比较检验,确定药物对基因表达的统计显著影响。[0343]下表18概述了第2阶段的实验步骤:[0344]表18[0345][0346][0347]在第2阶段,将检查融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽减轻疼痛的持久性。在本研究中,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽剂量(低、中或高),以及在第1阶段确定为最有效的给药期(14或28天)将被使用。将遵循上述第1阶段疗效研究的相同方法。不同之处在于,施用期(14或28天)后,每隔30天对大鼠进行一次疼痛反应和运动功能的监测和行为评估。大鼠将在第30天被安乐死。本研究共需要72只2个月大的sd大鼠(雄性和雌性数量相等)。每组12只大鼠的样本量将有80%的效力检测疼痛评估中标准差1.20的差异。效力分析基于独立样本的t检验,具有5%的双侧显著性水平。将大鼠随机分配到右侧表格中指定的实验组。对10个随机分组的大鼠进行刺激诱发和非刺激诱发行为测试、形态学、免疫组化分析、定量rt-pcr(qrt-pcr)分析和wb分析。[0348]目标3:融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的毒理学和安全性药理学评估。为了支持首次在人体内给药,我们将根据fda指南m3(r2)、s2b、s6(r1)、s7a和s7b的建议175-179,评估两种融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的体外心脏和遗传毒性以及体内毒理学、安全药理学和毒代动力学。在这两个物种中,将根据目标1中的pk评估结果选择给药途径。[0349]子目标3.1:猴子的单剂量范围判定:将在3只猴子/性别/组中评估四个剂量水平 溶剂对照。hd应确定mtd、mfd和/或暴露饱和度。较低剂量将以1/3的增量间隔。将在6个时间点采集血液(根据目标1中收集的pk数据确定),并评估cmax、auc和t1/2。将每日评估生存率和临床症状。[0350]子目标3.2和3.3:大鼠(3.2)和猴子(3.3)的毒理学研究。这两个物种的给药期均为4周,以支持相同时间的人类给药 6周的恢复期,其由于在细胞检测中观察到rpp的高稳定性而被选择77。将有三个剂量水平 溶剂对照。高剂量将是剂量范围判定(大鼠目标1.2和猴子目标3.1)中确定的最大耐受剂量。根据范围判定中的auc,md将比hd低1/3。ld将接近体内药效研究中的有效剂量(目标2)。给药间隔为每个物种中与融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的t1/2。在大鼠研究中,将有15只/性别/组。10只/性别/组将在给药期结束时被处死,5只/性别/溶剂将在恢复期结束时被处死。此外,9只/性别/rpp组和3只/性别/溶剂组将被分配到研究的毒代动力学部分。在猴子研究中,将对所有动物进行毒理学和tk评估(主要研究中为3只/性别/组,恢复期为2只/性别/组)。将对这两个物种的以下毒理学参数进行评估:临床观察、体重、食物消耗、眼底镜检查(眼底镜检查和裂隙灯检查),并将评估一组标准的临床化学、器官重量和组织学参数。猴子将接受使用非手术遥测功能测定的呼吸(例如潮气量和血红蛋白血氧饱和度)和心脏心电图记录评估179,按照fda指南s7a的建议179。心血管评估将在治疗前、第一次给药后和恢复期结束时(如需要)进行两次。这些记录将由顾问委员会认证的心脏病专家进行定性评估。对所有波形进行定性评估,以检测心律或传导障碍,包括pr和qrs间期的评估。将使用改良的irwin功能观察组合(battery)对大鼠的中枢神经系统功能进行评估179,180。将在6个时间点(由范围判定研究确定)进行血浆tk评估(cmax、auc和t1/2)。[0351]子目标3.4:体外herg检测177。herg通道抑制是长qt综合征的常见原因,也与可导致心室颤动和心源性猝死的心律失常相关。为了测试融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否抑制herg通道电流,我们将对用高浓度融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽(10μm)或阳性对照(西沙必利,0.03μm)处理的cho细胞中的herg通道电流进行电生理学评估。每个细胞将作为自身的阳性对照。全细胞记录将使用传统的电压钳技术进行。[0352]子目标3.5:遗传毒性。将在符合oecd的体外ames和体外染色体畸变检测(s2b,1997年11月)176中评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,以确定其遗传毒性潜力。[0353]目标4:融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的滥用可能性评估。由于阿片受体基因表达被rest抑制,因此融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可能增强内源性阿片受体配体(如内啡肽181,182)的作用。此外,最近的研究确实表明,在涉及疼痛的情况下,drg和导水管周围灰质(pag)中rest的表达增加183,184。为了确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否产生奖赏寻求行为,我们将使用条件性位置偏爱(cpp)评估其对大鼠的影响,这是一种广泛使用的行为检测,用于测试药物的奖赏特性185-189。cpp是一种成熟的方法,其已在sni动物模型中证明了吗啡的奖赏寻求行为。[0354]首先,将融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽施用给未受伤的大鼠(途径tbd),然后进行cpp。为了与目标2保持一致,将使用羟考酮代替吗啡作为阳性对照,并将生理盐水注射到单独的大鼠体内作为阴性对照。[0355]设备由三个隔室组成:两个外部隔室设计为具有不同的特性(例如白色和黑色墙壁),中央隔室没有特殊特性。每个实验包括三个阶段。i期预试验(第1天):施用生理盐水后,将大鼠放置在中心隔室中,打开闸门,以便进入其他隔室。将对大鼠进行15分钟的监测,通过记录在每个小室中的时间来确定基线偏好。将确定隔室的平均自发偏好时间。在任何一个试验室中花费超过60%试验时间的大鼠将被视为有偏好,并将从试验中移除。ii期条件反射形成(第2-7天):给大鼠注射溶剂或rpp(seqidno:1和15至17)或rppv(seqidno:18至117)融合到cpp(seqidno:118至137和140至159)的肽,然后,他们将被随机分配/调节,以在一个隔室和另一个隔室中接受与cpp(seqidno:118至137和140至159)融合的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽。与cpp(seqidno:118至137和140至159)融合的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的剂量将基于对目标2中确定的机械和热痛觉过敏产生可测定影响的浓度。治疗室以及药物和溶剂的给药顺序将对所有组进行平衡,以避免偏见。该阶段包括六个连续的30分钟的调节期。对于单独的一组,也将使用羟考酮进行调节。iii期试验(第8天):将向大鼠注射生理盐水,并将其放置在cpp箱的中心,并监测15分钟。后处理期间在药物对隔室中花费的时间与预处理期间花费的时间之间的差异将用于评估rpp(seqidno:1和15至17)或rppv(seqidno:18至117)引起的位置调节程度融合到cpp的肽(seqidno:118至137和140至159)190。[0356]将使用8组12只受伤大鼠(sni和假手术组,用羟考酮、融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽形成条件反射,对融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,和生理盐水)进行评估。每组12只大鼠的样本量将有80%的效力检测疼痛评估中1.20个标准差的差异。效力分析基于独立样本的t检验,具有5%的双侧显著性水平。将大鼠随机分配到实验组。将随机分配大鼠进行sni或假手术。vonfrey试验将在受伤前一天、受伤后4天和8天进行,以监测疼痛状态。受伤后8天,cpp测试将如前所述进行。进行行为测试的研究助理将不知道大鼠被随机分配到哪个实验组/对照组。每只动物都有一个唯一的动物编号。将使用秘密彩色代码指定对照组和实验组。只有在分析之后,才会显示此代码。将对动物实施安乐死,并按照目标2所述对样本进行分析。[0357]预测结果和替代策略:我们预测,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽将降低受损神经元的rest水平,并恢复兴奋性所需的离子通道的正确表达。在行为水平上,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽应使sni大鼠的刺激或非刺激诱发行为测试恢复到基线水平。此外,鉴于其作用模式,我们不期望融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽具有成瘾性。然而,如果药物管理显示出滥用可能性,我们将通过融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽施用期间的渐进比率(pr)强化表来确认这一阳性结果。[0358]实施例15-融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可用于预防多形性胶质母细胞瘤(gbm)的复发[0359]评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对脑肿瘤初始细胞(btic)和体内的神经发生和抗癌效力。将使用我们建立的方法,在mayoclinic获得的10名iv级gbm患者的原代培养btc中,评估与融合到cpp(seqidno:118至137和140至159)的供试品rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的活性191-193。将细胞(0.25×104)铺板于(3.8cm2tc皿)培养基(250μl)中,所述培养基以最佳剂量范围(在实施例13所述的神经元分化研究中确定)添加5μl溶剂或3个剂量水平的供试品,持续0、2、4或8天(主要研究),然后在洗出供试品8天后恢复(恢复期)。通过wb和rt-pcr,在给药期间(主要研究)的第0、2、4和8天以及恢复期间(每天3个/tc孔/组)的第10、13和16天监测多能性(sox2和巢蛋白)、增殖(ki67)、细胞死亡(caspase3)和分化(map2、神经元和gfap、胶质细胞)的标志物。[0360]来自10名iv级gbm患者的原代培养btc将用于我们和其他人之前描述的颅内异种成瘤裸鼠模型191,192,194。简而言之,将细胞(5×104)悬浮在5μlpbs(对照组)或含有3tbd(基于上述btic神经发生效力检测)供试品剂量水平之一的pbs中,并使用带导螺杆系统的自动注射泵输送112,195。注入坐标为x:1.5;y:1.34;z:-3.5靶向小鼠纹状体;高度可靠的肿瘤移植部位196。使用kaplan-meier方法197对小鼠在第4、8、16和20周的存活率进行检测,然后进行死后组织病理学检查,以评估肿瘤大小和侵袭性(h&e染色)、细胞死亡(tunel)和增殖(抗人细胞核)(所有步骤均为3个/性别/组)。[0361]统计方法:如图所示,数值将报告为平均值±平均值标准误差(sem)或标准偏差(sd)。统计显著性将通过应用学生t检验的适当参数或非参数变化或单因素方差分析(anova)来确定。简而言之,将使用kruskal-wallis检验对不同对照和候选药物治疗的总rest与磷酸化rest的比率进行统计检验。将使用tukey多重比较试验对btic对照和候选药物培养物以及原位异种移植体之间标记物表达的差异进行统计检验。注射注射btic的用对照品或供试品治疗的小鼠之间的肿瘤大小和侵袭性差异将使用mann-whitneyu检验进行统计检验,生存率差异将使用对数秩检验和cox比例风险检验198进行统计检验。[0362]预测结果:与对照组相比,在每个时间点,我们预计有btic(给药供试品)异种移植物的小鼠会出现无瘤或侵袭性较小的肿瘤,细胞死亡(tunel)更多,gbm增殖(抗人细胞核)更少。此外,我们预计注射供试品和btic的小鼠与单独注射btic的小鼠相比,存活时间会延长。[0363]材料和方法[0364]测试系统[0365]细胞:原代培养的脑肿瘤起始细胞(btic)来自10例iv级gbm患者。alfredoquinoneshinojosa博士及其研究团队之前已经描述了提取和繁殖btic的方法191-193。[0366]物种/菌株:nu/nu-harlansprague-dawley小鼠[0367]年龄:研究开始时:6周[0368]体重:研究开始时:雄性:20-30克;雌性:18-35g[0369]供试品标识。[0370]融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽[0371]纯度:≥95%hplc[0372]剂型制备和稳定性:每周将批量制备融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽和溶剂对照品。将在每周开始时评估剂型浓度。可接受的浓度范围为标称浓度的±10%。[0373]溶剂:水[0374]研究设计[0375]供试品的神经发生效力研究:[0376]剂量:3个供试品剂量水平 溶剂对照。将根据实施例11中所述神经元分化研究的结果确定供试品剂量。[0377]施用:每2天给药一次,并更换培养基。[0378]持续时间:主要研究:0、2、4或8天;恢复期:2、5或8天。[0379]重复数:每组3个组织培养(tc)孔。[0380]给药体积:5μl[0381]分析参数:将在给药期间(主要研究)的第0、2、4和8天以及恢复期间(每天3个/tc孔/组)的第10、13和16天(表10),通过wb和rt-pcr监测多能性(sox2和nestin)、增殖(ki67)、细胞死亡(caspase3)和分化(map2、神经元和gfap、胶质细胞)的标记物。[0382][0383][0384]供试品对小鼠btic的抗癌效力:[0385]剂量:3剂供试品 溶剂对照。供试品剂量将基于神经源性效力研究的结果。[0386]施用频率:单剂量[0387]途径:带导向螺杆系统的自动注射泵112,195。注入坐标为x:1.5;y:1.34;z:-3.5靶向小鼠纹状体;高度可靠的肿瘤移植部位196。[0388]溶剂:磷酸盐缓冲盐水(pbs)[0389]动物数量:3只/性别/组[0390]给药体积:5μl[0391]参数分析:将使用kaplan-meier方法197对小鼠在第4、8、16和20周的存活率进行检测,然后进行死后组织病理学检查,以评估肿瘤大小和侵袭性(h&e染色)、细胞死亡(tunel)和增殖(抗人细胞核)(表11)。[0392][0393][0394]环境条件[0395]居住条件:2只动物/笼,光/暗循环12小时。[0396]饮食:随意进食进水。[0397]下表19示出了seqidno:1至324的序列:[0398][0399][0400][0401][0402][0403][0404][0405][0406][0407][0408][0409][0410][0411][0412][0413][0414][0415][0416][0417][0418]尽管本发明的某些当前优选实施例已在本文中具体描述,但对于本发明所属领域的技术人员来说,显而易见的是,可以在不脱离本发明的精神和范围的情况下对本文所示和描述的各种实施方案进行变更和修改。[0419]参考文献(说明书,实施例除外)sci5,573-587,doi:10.1002/wcs.1304(2014).[0445]26samuels,i.s.etal.deletionoferk2mitogen-activatedproteinkinaseidentifiesitskeyrolesincorticalneurogenesisandcognitivefunction.jneurosci28,6983-6995,doi:28/27/6983[pii]10.1523/jneurosci.0679-08.2008(2008).[0446]27shors,t.j.etal.neurogenesisintheadultisinvolvedintheformationoftracememories.nature410,372-376,doi:10.1038/35066584(2001).[0447]28villeda,s.a.etal.theageingsystemicmilieunegativelyregulatesneurogenesisandcognitivefunction.nature477,90-94,doi:10.1038/nature10357(2011).[0448]29bird,c.m.&burgess,n.thehippocampussupportsrecognitionmemoryforfamiliarwordsbutnotunfamiliarfaces.currbiol18,1932-1936,doi:10.1016/j.cub.2008.10.046(2008).[0449]30t.j.shors,g.m.,a.beylin,m.zhao,t.rydel,e.gould.neurogenesisintheadultisinvolvedintheformationoftracememories.nature410,372-376(2001).[0450]31vanelzakker,m.,fevurly,r.d.,breindel,t.&spencer,r.l.environmentalnoveltyisassociatedwithaselectiveincreaseinfosexpressionintheoutputelementsofthehippocampalformationandtheperirhinalcortex.learnmem15,899-908,doi:10.1101/lm.1196508(2008).[0451]32thornton,g.k.&woods,c.g.primarymicrocephaly:doallroadsleadtorome?trendsgenet25,501-510,doi:10.1016/j.tig.2009.09.011(2009).[0452]33ghaziuddin,m.,zaccagnini,j.,tsai,l.&elardo,s.ismegalencephalyspecifictoautism?jintellectdisabilres43(pt4),279-282(1999).[0453]34kuhn,h.g.,cooper-kuhn,c.m.,boekhoorn,k.&lucassen,p.j.changesinneurogenesisindementiaandalzheimermousemodels:aretheyfunctionallyrelevant?eurarchpsychiatryclinneurosci257,281-289,doi:10.1007/s00406-007-0732-4(2007).[0454]35baker,m.tumourssparkstem-cellreview.nature457,941,doi:10.1038/457941a(2009).[0455]36brederlau,a.etal.transplantationofhumanembryonicstemcell-derivedcellstoaratmodelofparkinson'sdisease:effectofinvitrodifferentiationongraftsurvivalandteratomaformation.stemcells24,1433-1440,doi:2005-0393[pii]10.1634/stemcells.2005-0393(2006).[0456]37burns,t.c.,verfaillie,c.m.&low,w.c.stemcellsforischemicbraininjury:acriticalreview.jcompneurol515,125-144,doi:10.1002/cne.22038(2009).[0457]38carlson,a.p.,schermer,c.r.&lu,s.w.retrospectiveevaluationofanemiaandtransfusionintraumaticbraininjury.jtrauma61,567-571,doi:theregulationofneuronalexcitability.jneurosci30,13235-13245,doi:10.1523/jneurosci.1981-10.2010(2010).[0484]65escayg,a.etal.mutationsofscn1a,encodinganeuronalsodiumchannel,intwofamilieswithgefs 2.natgenet24,343-345,doi:10.1038/74159(2000).[0485]66martin,d.etal.restrepressesasubsetofthepancreaticendocrinedifferentiationprogram.devbiol405,316-327,doi:10.1016/j.ydbio.2015.07.002(2015).[0486]67abderrahmani,a.etal.neuronaltraitsarerequiredforglucose-inducedinsulinsecretion.febslett565,133-138,doi:10.1016/j.febslet.2004.04.002(2004).[0487]68martin,d.etal.functionalsignificanceofrepressorelement1silencingtranscriptionfactor(rest)targetgenesinpancreaticbetacells.diabetologia51,1429-1439,doi:10.1007/s00125-008-0984-1(2008).[0488]69rigamonti,d.etal.lossofhuntingtinfunctioncomplementedbysmallmoleculesactingasrepressorelement1/neuronrestrictivesilencerelementsilencermodulators.jbiolchem282,24554-24562,doi:10.1074/jbc.m609885200(2007).[0489]70sipione,s.etal.earlytranscriptionalprofilesinhuntingtin-induciblestriatalcellsbymicroarrayanalyses.hummolgenet25,210,doi:10.1093/hmg/ddv416(2016).[0490]71zuccato,c.etal.huntingtininteractswithrest/nrsftomodulatethetranscriptionofnrse-controlledneuronalgenes.natgenet35,76-83,doi:10.1038/ng1219(2003).[0491]72bao,s.etal.targetingcancerstemcellsthroughl1camsuppressesgliomagrowth.cancerres68,6043-6048,doi:10.1158/0008-5472.can-08-1079(2008).[0492]73jackson,m.,hassiotou,f.&nowak,a.glioblastomastem-likecells:attherootoftumorrecurrenceandatherapeutictarget.carcinogenesis36,177-185,doi:10.1093/carcin/bgu243(2015).[0493]74liu,g.etal.analysisofgeneexpressionandchemoresistanceofcd133 cancerstemcellsinglioblastoma.molcancer5,67,doi:10.1186/1476-4598-5-67(2006).[0494]75persano,l.,rampazzo,e.,basso,g.&viola,g.glioblastomacancerstemcells:roleofthemicroenvironmentandtherapeutictargeting.biochempharmacol85,612-622,doi:10.1016/j.bcp.2012.10.001(2013).[0495]76conti,l.etal.restcontrolsself-renewalandtumorigeniccompetenceofhumanglioblastomacells.plosone7,e38486,doi:10.1371/journal.pone.0038486(2012).[0496]77kamal,m.m.etal.restregulatesoncogenicpropertiesofglioblastomastemcells.stemcells30,405-414,doi:10.1002/stem.1020(2012).[0497]78wagoner,m.p.&roopra,a.arestderivedgenesignaturestratifiesglioblastomasintochemotherapyresistantandresponsivedisease.bmcgenomics13,686,doi:10.1186/1471-2164-13-686(2012).[0498]79taylor,p.etal.restisanovelprognosticfactorandtherapeutictargetformedulloblastoma.molcancerther11,1713-1723,doi:10.1158/1535-7163.mct-11-0990(2012).[0499]80jin,h.etal.identificationofre1-silencingtranscriptionfactorasapromoterofmetastasisinpancreaticcancer.frontoncol9,291,doi:10.3389/fonc.2019.00291(2019).[0500]参考文献(仅针对实施例)[0501]1bruce,a.w.etal.genome-wideanalysisofrepressorelement1silencingtranscriptionfactor/neuron-restrictivesilencingfactor(rest/nrsf)targetgenes.procnatlacadsciusa101,10458-10463,doi:10.1073/pnas.04018271010401827101[pii](2004).[0502]2nesti,e.,corson,g.m.,mccleskey,m.,oyer,j.a.&mandel,g.c-terminaldomainsmallphosphatase1andmapkinasereciprocallycontrolreststabilityandneuronaldifferentiation.procnatlacadsciusa111,e3929-3936,doi:1414770111[pii]10.1073/pnas.1414770111(2014).[0503]3nesti,e.methodsandcompositionsusefulinmanipulatingthestabilityofre1silencingtranscriptionfactor.us,ptcpatent(2014,2015).[0504]4nesti,e.harnessingthemastertranscriptionalrepressorresttoreciprocallyregulateneurogenesis..neurogenesis2,doi:10.1080/23262133.2015.1055419(2015).[0505]5arranz-gibert,p.etal.immunosilencingpeptidesbystereochemicalinversionandsequencereversal:retro-d-peptides.scirep8,6446,doi:10.1038/s41598-018-24517-6(2018).[0506]6hall,e.d.,bryant,y.d.,cho,w.&sullivan,p.g.evolutionofpost-traumaticneurodegenerationaftercontrolledcorticalimpacttraumaticbraininjuryinmiceandratsasassessedbythedeolmossilverandfluorojadestainingmethods.jneurotrauma25,235-247,doi:10.1089/neu.2007.0383(2008).[0507]7calderone,a.etal.ischemicinsultsderepressthegenesilencerrestinneuronsdestinedtodie.jneurosci23,2112-2121,doi:23/6/2112[pii](2003).[0508]8kaneko,n.,hwang,j.y.,gertner,m.,pontarelli,f.&zukin,r.s.caseinkinase1suppressesactivationofrestininsultedhippocampalneuronsandhaltsischemia-inducedneuronaldeath.jneurosci34,6030-6039,doi:34/17/6030[pii]10.1523/jneurosci.4045-13.2014(2014).[0509]9noh,k.m.etal.repressorelement-1silencingtranscriptionfactor(rest)-dependentepigeneticremodelingiscriticaltoischemia-inducedneuronaldeath.procnatlacadsciusa109,e962-971,doi:1121568109[pii]10.1073/pnas.1121568109(2012).[0510]10abe,k.therapeuticpotentialofneurotrophicfactorsandneuralstemcellsagainstischemicbraininjury.jcerebbloodflowmetab20,1393-1408,doi:10.1097/00004647-200010000-00001(2000).[0511]11kanding,p.k.g.,andramondiaz-arrastia.intranslationalresearchintraumaticbraininjury(edgrantglaskowitzd)ch.14,(taylorandfrancis,2016).[0512]12pasquina,p.,kirtley,r.&ling,g.moderate-to-severetraumaticbraininjury.seminneurol34,572-583,doi:10.1055/s-0034-1396010(2014).[0513]13hwang,j.y.,kaneko,n.,noh,k.m.,pontarelli,f.&zukin,r.s.thegenesilencingtranscriptionfactorrestrepressesmir-132expressioninhippocampalneuronsdestinedtodie.jmolbiol426,3454-3466,doi:10.1016/j.jmb.2014.07.032(2014).[0514]14mcclelland,s.etal.thetranscriptionfactornrsfcontributestoepileptogenesisbyselectiverepressionofasubsetoftargetgenes.elife3,e01267(2014).[0515]15lu,t.etal.restandstressresistanceinageingandalzheimer'sdisease.nature507,448-454,doi:nature13163[pii]10.1038/nature13163(2014).[0516]16orta-salazar,e.etal.rest/nrsf-inducedchangesofchatproteinexpressionintheneocortexandhippocampusofthe3xtg-admousemodelforalzheimer'sdisease.lifesciences116,83-89,doi:10.1016/j.lfs.2014.09.013(2014).[0517]17sharp,d.j.,scott,g.&leech,r.networkdysfunctionaftertraumaticbraininjury.natrevneurol10,156-166,doi:10.1038/nrneurol.2014.15(2014).[0518]18roopra,a.,dingledine,r.&hsieh,j.epigeneticsandepilepsy.epilepsia53suppl9,2-10,doi:10.1111/epi.12030(2012).[0519]19bergsland,m.,covacu,r.,perezestrada,c.,svensson,m.&brundin,l.nitricoxide-inducedneuronaltogliallineagefate-changedependsonnrsf/restfunctioninneuralprogenitorcells.stemcells32,2539-2549,doi:10.1002/stem.1749(2014).[0520]20conaco,c.,otto,s.,han,j.j.&mandel,g.reciprocalactionsofrestandamicrornapromoteneuronalidentity.procnatlacadsciusa103,2422-2427,doi:0511041103[pii]10.1073/pnas.0511041103(2006).[0521]21covey,m.v.,streb,j.w.,spektor,r.&ballas,n.restregulatesthepoolsizeofthedifferentneurallineagesbyrestrictingthegenerationofneuronsandoligodendrocytesfromneuralstem/progenitorcells.development139,2878-2890,doi:dev.074765[pii]10.1242/dev.074765(2012).[0522]22gao,z.etal.themasternegativeregulatorrest/nrsfcontrolsadultneurogenesisbyrestrainingtheneurogenicprograminquiescentstemcells.jneurosci31,9772-9786,doi:31/26/9772[pii]10.1523/jneurosci.1604-11.2011(2011).[0523]23kohyama,j.etal.bmp-inducedrestregulatestheestablishmentandmaintenanceofastrocyticidentity.jcellbiol189,159-170,doi:jcb.200908048[pii]10.1083/jcb.200908048(2010).[0524]24mandel,g.etal.repressorelement1silencingtranscriptionfactor(rest)controlsradialmigrationandtemporalneuronalspecificationduringneocorticaldevelopment.procnatlacadsciusa108,16789-16794,doi:1113486108[pii]10.1073/pnas.1113486108(2011).[0525]25vincent,a.s.,roebuck-spencer,t.m.&cernich,a.cognitivechangesanddementiariskaftertraumaticbraininjury:implicationsforagingmilitarypersonnel.alzheimersdement10,s174-187,doi:10.1016/j.jalz.2014.04.006(2014).[0526]26kirn,j.r.therelationshipofneurogenesisandgrowthofbrainregionstosonglearning.brainlang115,29-44,doi:s0093-934x(09)00135-7[pii]10.1016/j.bandl.2009.09.006(2010).[0527]27oomen,c.a.,bekinschtein,p.,kent,b.a.,saksida,l.m.&bussey,t.j.adulthippocampalneurogenesisanditsroleincognition.wileyinterdisciprevcognsci5,573-587,doi:10.1002/wcs.1304(2014).[0528]28samuels,i.s.etal.deletionoferk2mitogen-activatedproteinkinaseidentifiesitskeyrolesincorticalneurogenesisandcognitivefunction.jneurosci28,6983-6995,doi:28/27/6983[pii]10.1523/jneurosci.0679-08.2008(2008).[0529]29shors,t.j.etal.neurogenesisintheadultisinvolvedintheformationoftracememories.nature410,372-376,doi:10.1038/35066584(2001).[0530]30villeda,s.a.etal.theageingsystemicmilieunegativelyregulatesneurogenesisandcognitivefunction.nature477,90-94,doi:10.1038/nature10357(2011).[0531]31bird,c.m.&burgess,n.thehippocampussupportsrecognitionmemoryforfamiliarwordsbutnotunfamiliarfaces.currbiol18,1932-1936,doi:10.1016/j.cub.2008.10.046(2008).[0532]32t.j.shors,g.m.,a.beylin,m.zhao,t.rydel,e.gould.neurogenesisintheadultisinvolvedintheformationoftracememories.nature410,372-376(2001).[0533]33vanelzakker,m.,fevurly,r.d.,breindel,t.&spencer,r.l.environmentalnoveltyisassociatedwithaselectiveincreaseinfosexpressionintheoutputelementsofthehippocampalformationandtheperirhinalcortex.learnmem15,899-908,doi:10.1101/lm.1196508(2008).[0534]34thornton,g.k.&woods,c.g.primarymicrocephaly:doallroadsleadtorome?trendsgenet25,501-510,doi:10.1016/j.tig.2009.09.011(2009).[0535]35ghaziuddin,m.,zaccagnini,j.,tsai,l.&elardo,s.ismegalencephalyspecifictoautism?jintellectdisabilres43(pt4),279-282(1999).[0536]36kuhn,h.g.,cooper-kuhn,c.m.,boekhoorn,k.&lucassen,p.j.changesinneurogenesisindementiaandalzheimermousemodels:aretheyfunctionallyrelevant?eurarchpsychiatryclinneurosci257,281-289,doi:10.1007/s00406-007-0732-4(2007).[0537]37baker,m.tumourssparkstem-cellreview.nature457,941,doi:10.1038/457941a(2009).[0538]38brederlau,a.etal.transplantationofhumanembryonicstemcell-derivedcellstoaratmodelofparkinson'sdisease:effectofinvitrodifferentiationongraftsurvivalandteratomaformation.stemcells24,1433-1440,doi:2005-0393[pii]10.1634/stemcells.2005-0393(2006).[0539]39burns,t.c.,verfaillie,c.m.&low,w.c.stemcellsforischemicbraininjury:acriticalreview.jcompneurol515,125-144,doi:10.1002/cne.22038(2009).[0540]40carlson,a.p.,schermer,c.r.&lu,s.w.retrospectiveevaluationofanemiaandtransfusionintraumaticbraininjury.jtrauma61,567-571,doi:10.1097/01.ta.0000231768.44727.a200005373-200609000-00007[pii](2006).[0541]41carmeliet,p.&storkebaum,e.vascularandneuronaleffectsofvegfinthenervoussystem:implicationsforneurologicaldisorders.semincelldevbiol13,39-53,doi:10.1006/scdb.2001.0290s1084952101902903[pii](2002).[0542]42choe,y.,kozlova,a.,graf,d.&pleasure,s.j.bonemorphogenicproteinsignalingisamajordeterminantofdentatedevelopment.jneurosci33,6766-6775,doi:33/16/6766[pii]10.1523/jneurosci.0128-13.2013(2013).[0543]43erdo,f.etal.host-dependenttumorigenesisofembryonicstemcelltransplantationinexperimentalstroke.jcerebbloodflowmetab23,780-785,doi:10.1097/01.wcb.0000071886.63724.fb(2003).[0544]44greig,n.h.etal.incretinmimeticsaspharmacologictoolstoelucidateandasanewdrugstrategytotreattraumaticbraininjury.alzheimersdement10,s62-75,doi:s1552-5260(13)02925-7[pii]10.1016/j.jalz.2013.12.011(2014).[0545]45kaplan,g.b.,vasterling,j.j.&vedak,p.c.brain-derivedneurotrophicfactorintraumaticbraininjury,post-traumaticstressdisorder,andtheircomorbidconditions:roleinpathogenesisandtreatment.behavpharmacol21,427-437,doi:10.1097/fbp.0b013e32833d8bc9(2010).[0546]46knoepfler,p.s.deconstructingstemcelltumorigenicity:aroadmaptosaferegenerativemedicine.stemcells27,1050-1056,doi:10.1002/stem.37(2009).dysfunctionsinneuropathicpain.journalofneuroscience30,4806-4814,doi:10.1523/jneurosci.5541-09.2010(2010).[0561]61alvarado,s.etal.peripheralnerveinjuryisaccompaniedbychronictranscriptome-widechangesinthemouseprefrontalcortex.molpain9,21,doi:10.1186/1744-8069-9-21(2013).[0562]62hammer,p.etal.mrna-seqwithagnosticsplicesitediscoveryfornervoussystemtranscriptomicstestedinchronicpain.genomeres20,847-860,doi:10.1101/gr.101204.109(2010).[0563]63willis,d.e.,wang,m.,brown,e.,fones,l.&cave,j.w.selectiverepressionofgeneexpressioninneuropathicpainbytheneuron-restrictivesilencingfactor/repressorelement-1silencingtranscription(nrsf/rest).neuroscienceletters625,20-25,doi:10.1016/j.neulet.2015.12.003(2016).[0564]64ballas,n.,grunseich,c.,lu,d.d.,speh,j.c.&mandel,g.restanditscorepressorsmediateplasticityofneuronalgenechromatinthroughoutneurogenesis.cell121,645-657,doi:s0092-8674(05)00285-0[pii]10.1016/j.cell.2005.03.013(2005).[0565]65chong,j.a.etal.rest:amammaliansilencerproteinthatrestrictssodiumchannelgeneexpressiontoneurons.cell80,949-957,doi:0092-8674(95)90298-8[pii](1995).[0566]66mortazavi,a.,leeperthompson,e.c.,garcia,s.t.,myers,r.m.&wold,b.comparativegenomicsmodelingofthenrsf/restrepressornetwork:fromsingleconservedsitestogenome-widerepertoire.genomeres16,1208-1221,doi:gr.4997306[pii]10.1101/gr.4997306(2006).[0567]67otto,s.j.etal.anewbindingmotifforthetranscriptionalrepressorrestuncoverslargegenenetworksdevotedtoneuronalfunctions.jneurosci27,6729-6739,doi:27/25/6729[pii]10.1523/jneurosci.0091-07.2007(2007).[0568]68schoenherr,c.j.&anderson,d.j.silencingisgolden:negativeregulationinthecontrolofneuronalgenetranscription.curropinneurobiol5,566-571,doi:0959-4388(95)80060-3[pii](1995).[0569]69schoenherr,c.j.,paquette,a.j.&anderson,d.j.identificationofpotentialtargetgenesfortheneuron-restrictivesilencerfactor.procnatlacadsciusa93,9881-9886(1996).[0570]70rose,k.etal.transcriptionalrepressionofthemchannelsubunitkv7.2inchronicnerveinjury.pain152,742-754,doi:10.1016/j.pain.2010.12.028(2011).[0571]71costigan,m.etal.multiplechronicpainstatesareassociatedwithacommonaminoacid-changingalleleinkcns1.brain133,2519-2527,doi:10.1093/brain/awq195(2010).[0572]72zhang,f.etal.repressorelement1-silencingtranscriptionfactordrivesthedevelopmentofchronicpainstates.pain,doi:10.1097/j.pain.0000000000001633(2019).[0573]73zhang,j.,chen,s.r.,chen,h.&pan,h.l.re1-silencingtranscriptionfactorcontrolstheacute-to-chronicneuropathicpaintransitionandchrm2receptorgeneexpressioninprimarysensoryneurons.jbiolchem293,19078-19091,doi:10.1074/jbc.ra118.005846(2018).[0574]74ueda,h.etal.amimeticofthemsin3-bindinghelixofnrsf/restamelioratesabnormalpainbehaviorinchronicpainmodels.bioorgmedchemlett27,4705-4709,doi:10.1016/j.bmcl.2017.09.006(2017).[0575]75uchida,h.,sasaki,k.,ma,l.&ueda,h.neuron-restrictivesilencerfactorcausesepigeneticsilencingofkv4.3geneafterperipheralnerveinjury.neuroscience166,1-4,doi:10.1016/j.neuroscience.2009.12.021(2010).[0576]76gervasinm.,d.a.,clarkdm.,dinglem,,pisarchikav.,nestilj.c-terminaldomainsmallphosphatase1(ctdsp1)regulatesgrowthfactorexpressionandaxonalregeneration.journaloftranslationalmedicinemanuscriptedsubmittedforpublication.(2020).[0577]77nesti,e.,pisarchik,a.compositionsandmethodsforderepressingre1silencingtranscriptionfactortargetgenes.usapatent(2019).[0578]78cole,l.a.,kurscheid,s.,nekrasov,m.,domaschenz,r.,dennis,j.h.,tremethick,d.j.redefiningthenucleosomalarchitectureofactiveandinactivepromotersinthecontextofcellularplasticityandcancer.naturecommunicationsrevisionreturned(2020).[0579]79ren,y.,vera,d.l.,hughes,k.a.&dennis,j.h.stimulationofthedrosophilaimmunesystemaltersgenome-widenucleosomeoccupancy.genomdata3,146-147,doi:10.1016/j.gdata.2015.01.001(2015).[0580]80sexton,b.s.etal.thespring-loadedgenome:nucleosomeredistributionsarewidespread,transient,anddna-directed.genomeres24,251-259,doi:10.1101/gr.160150.113(2014).[0581]81sexton,b.s.etal.hierarchicalregulationofthegenome:globalchangesinnucleosomeorganizationpotentiategenomeresponse.oncotarget7,6460-6475,doi:10.18632/oncotarget.6841(2016).[0582]82lunyak,v.v.&rosenfeld,m.g.norestforrest:rest/nrsfregulationofneurogenesis.cell121,499-501,doi:10.1016/j.cell.2005.05.003(2005).[0583]83dirks,p.b.makingacommitment:neuronsrefusecancer'sadvances.natneurosci22,507-508,doi:10.1038/s41593-019-0373-8(2019).[0584]84kaspar,a.a.&reichert,j.m.futuredirectionsforpeptidetherapeuticsdevelopment.drugdiscovtoday18,807-817,doi:10.1016/j.drudis.2013.05.011(2013).[0585]85hamzeh-mivehroud,m.,alizadeh,a.a.,morris,m.b.,bretchurch,w.&dastmalchi,s.phagedisplayasatechnologydeliveringonthepromiseofpeptidedrugdiscovery.drugdiscoverytoday18,1144-1157,doi:10.1016/j.drudis.2013.09.001(2013).[0586]86hoekstra,e.,peppelenbosch,m.p.&fuhler,g.m.meetingreporteurophosphatase2015:phosphatasesasdrugtargetsincancer.cancerresearch76,193-196,doi:10.1158/0008-5472.can-15-2091(2016).[0587]87shi,y.serine/threoninephosphatases:mechanismthroughstructure.cell139,468-484,doi:10.1016/j.cell.2009.10.006(2009).[0588]88sussman,f.,villaverde,m.c.,l.dominguez,j.&danielson,u.h.ontheactivesiteprotonationstateinasparticproteases:implicationsfordrugdesign.currentpharmaceuticaldesign,volume19,number2319,4257-4275,[0589]doi:10.1002/(sici)1520-667x(1998)10:1《19::aid-mcs3》3.0.co;2-1(2013).[0590]89anobom,c.d.etal.fromstructuretocatalysis:recentdevelopmentsinthebiotechnologicalapplicationsoflipases.biomedresearchinternational2014,doi:10.1155/2014/684506(2014).[0591]90blundell,t.l.,jhoti,h.&abell,c.high-throughputcrystallographyforleaddiscoveryindrugdesign.naturereviewsdrugdiscovery1,45-54,doi:10.1038/nrd706(2002).[0592]91weir,m.r.renininhibitors:novelagentsforrenoprotectionorabetterangiotensinreceptorblockerforbloodpressurelowering?currentopinioninnephrologyandhypertension16,416-421,doi:10.1097/mnh.0b013e328209fe00(2007).[0593]92yeo,m.etal.smallctdphosphatasesfunctioninsilencingneuronalgeneexpression.science307,596-600,doi:307/5709/596[pii]10.1126/science.1100801(2005).[0594]93fulda,s.,wick,w.,weller,m.&debatin,k.m.smacagonistssensitizeforapo2l/trail-oranticancerdrug-inducedapoptosisandinduceregressionofmalignantgliomainvivo.naturemedicine8,808-815,doi:10.1038/nm735(2002).[0595]94noguchi,h.etal.anewcell-permeablepeptideallowssuccessfulallogeneicislettransplantationinmice.naturemedicine10,305-309,doi:10.1038/nm994(2004).[0596]95pooga,m.etal.cellulartranslocationofproteinsbytransportan.thefasebjournal:officialpublicationofthefederationofamericansocietiesforexperimentalbiology15,1451-1453,doi:10.1096/fj.00-0780fje(2001).[0597]96schwarze,s.r.,ho,a.,vocero-akbani,a.&dowdy,s.f.invivoproteintransduction:deliveryofabiologicallyactiveproteinintothemouse.science(newyork,n.y.)285,1569-1572,doi:10.1126/science.285.5433.1569(1999).2398(2015).[0661]160a.etal.perineuraldexmedetomidineeffectsonsciaticnerveinrat.brazilianjournalofanesthesiology(englishedition)67,57-66,doi:10.1016/j.bjane.2015.08.012(2017).[0662]161rigaud,m.etal.speciesandstraindifferencesinrodentsciaticnerveanatomy:implicationsforstudiesofneuropathicpain.pain136,188-201,doi:10.1016/j.pain.2008.01.016(2008).[0663]162sleigh,j.n.,weir,g.a.&schiavo,g.asimple,step-by-stepdissectionprotocolfortherapidisolationofmousedorsalrootganglia.bmcresnotes9,82,doi:10.1186/s13104-016-1915-8(2016).[0664]163ronchi,g.etal.discrepanciesinquantitativeassessmentofnormalandregeneratedperipheralnervefibersbetweenlightandelectronmicroscopy.jperiphernervsyst19,224-233,doi:10.1111/jns.12090(2014).[0665]164varejao,a.s.etal.functionalandmorphologicalassessmentofastandardizedratsciaticnervecrushinjurywithanon-serratedclamp.jneurotrauma21,1652-1670,doi:10.1089/neu.2004.21.1652(2004).[0666]165carriel,v.,garzon,i.,alaminos,m.&cornelissen,m.histologicalassessmentinperipheralnervetissueengineering.neuralregenres9,1657-1660,doi:10.4103/1673-5374.141798(2014).[0667]166hazari,a.,wiberg,m.,johansson-ruden,g.,green,c.&terenghi,g.aresorbablenerveconduitasanalternativetonerveautograftinnervegaprepair.brjplastsurg52,653-657,doi:10.1054/bjps.1999.3184(1999).[0668]167shi,t.j.etal.effectofperipheralnerveinjuryondorsalrootganglionneuronsinthec57bl/6jmouse:markedchangesbothincellnumbersandneuropeptideexpression.neuroscience105,249-263(2001).[0669]168jackson,w.m.etal.mesenchymalprogenitorcellsderivedfromtraumatizedmuscleenhanceneuritegrowth.jtissueengregenmed7,443-451,doi:10.1002/term.539(2013).[0670]169kobielaketza,b.k.,grunbergne,kasperce,osbornel,pryorb,tosininl,wux,andersjj.characterizationofmacrophage/microglialactivationandeffectofphotobiomodulationinthesparednerveinjurymodelofneuropathicpain..painmedicine5,932-946,doi:10.1093/pm/pnw144(2016).[0671]170whiteside,g.t.,adedoyina,leventhall.predictivevalidityofanimalpainmodels?acomparisonofthepharmacokinetic-pharmacodynamicrelationshipforpaindrugsinratsandhumans.neuropharm,767-775,doi:10.1016/j.neuropharm.2008.01.001(2008).[0672]171labuda,c.j.&fuchs,p.n.abehavioraltestparadigmtomeasuretheaversivequalityofinflammatoryandneuropathicpaininrats.expneurol163,490-494,doi:10.1006/exnr.2000.7395(2000).behaviourbydopamineinhibition.nature248,257-258(1974).[0687]186spyraki,c.,fibiger,h.c.&phillips,a.g.attenuationofheroinrewardinratsbydisruptionofthemesolimbicdopaminesystem.psychopharmacology79,278-283(1983).[0688]187phillips,a.g.&lepiane,f.g.reinforcingeffectsofmorphinemicroinjectionintotheventraltegmentalarea.pharmacology,biochemistry,andbehavior12,965-968(1980).[0689]188phillips,a.g.,lepiane,f.g.&fibiger,h.c.dopaminergicmediationofrewardproducedbydirectinjectionofenkephalinintotheventraltegmentalareaoftherat.lifesciences33,2505-2511(1983).[0690]189prusaj,j.j.,rosecransja.inmethodsofbehavioranalysisinneuroscience(edbuccafuscojj)ch.4,(crcpress/taylor&francis,2009).[0691]190carr,k.d.,bak,t.h.,simon,e.j.&portoghese,p.s.effectsoftheselectivekappaopioidantagonist,nor-binaltorphimine,onelectrically-elicitedfeedingintherat.lifesciences45,1787-1792(1989).[0692]191galli,r.etal.isolationandcharacterizationoftumorigenic,stem-likeneuralprecursorsfromhumanglioblastoma.cancerres64,7011-7021,doi:10.1158/0008-5472.can-04-1364(2004).[0693]192garzon-muvdi,t.etal.regulationofbraintumordispersalbynkcc1throughanovelroleinfocaladhesionregulation.plosbiol10,e1001320,doi:10.1371/journal.pbio.1001320(2012).[0694]193guerrero-cazares,h.,chaichana,k.l.&quinones-hinojosa,a.neurospherecultureandhumanorganotypicmodeltoevaluatebraintumorstemcells.methodsmolbiol568,73-83,doi:10.1007/978-1-59745-280-9_6(2009).[0695]194tilghman,j.etal.regulationofglioblastomatumor-propagatingcellsbytheintegrinpartnertetraspanincd151.neoplasia18,185-198,doi:10.1016/j.neo.2016.02.003(2016).[0696]195lal,s.etal.animplantableguide-screwsystemforbraintumorstudiesinsmallanimals.jneurosurg92,326-333,doi:10.3171/jns.2000.92.2.0326(2000).[0697]196manton,c.a.etal.inductionofcelldeathbythenovelproteasomeinhibitormarizomibinglioblastomainvitroandinvivo.scirep6,18953,doi:10.1038/srep18953(2016).[0698]197kaplan,e.l.m.,p.nonparametricestimationfromincompleteobservations.journaloftheamericanstatisticalassociation53,457-481,doi:10.2307/2281868(1958).[0699]198marubinie,v.m.c.estimationofsurvivalprobabilities.analysingsurvivaldatafromclinicaltrialsandobservationalstudies.41–81(johnwileyandsons,1995).当前第1页12当前第1页12
技术领域:
:3.本发明提供了用于去抑制re1沉默转录因子(rest)靶基因的方法、化合物和组合物。具体而言,本发明公开了一种具有序列tedleppepplpken(seqidno:1)和edleppepplpk(seqidno:15)的肽,或由d-氨基酸nekplppeppeldet(seqidno:16)和kplppeppelde(seqidno:17)组成的反向序列(反向倒置,retroinverted,ri)的肽,用于抑制rest活性。这些肽可用于治疗、预防或改善诸如创伤性脑损伤、癫痫、痴呆、亨廷顿病(hd)、慢性疼痛、脑癌(包括多形性胶质母细胞瘤)、胰腺癌;糖尿病和周围神经损伤等病变。
背景技术:
::4.阻遏因子1(re1)沉默转录因子(rest)是数百种神经元基因的阻遏因子1。其靶点代表终末分化神经元细胞表型所需的基因,包括编码电压和配体依赖性离子通道的基因、其受体、生长因子和轴突导向蛋白(bruceaw等人,procnatlacadsciusa101,10458-10463(2004);conacoc等人,procnatlacadsciusa103,2422-2427(2006);mortazavia等人,genomeres16,1208-1221(2006);和ottosj等人,jneurosci27,6729-6739(2007);所有这些都通过引用并入本文)。因此,在神经发生过程中,rest逐渐下调,以使成熟神经元表型得以确立(ballas等人,2005,上文(supra))。这一事件的重要性通过功能获得研究得到证实,这些研究表明,rest的持续存在会阻碍终末神经元分化(mandelg等人,procnatlacadsciusa108,16789-16794(2011)和gaoz等人,jneurosci31,9772-9786(2011);两者均通过引用并入本文)。5.对rest的转录或转录后调节知之甚少(ballasn等人2005,上文;ballasn等人,neuron31,353-365(2001);和kojimat等人,brainresmolbrainres90,174-186(2001);所有这些都通过引用并入本文)。然而,rest上的两个磷酸化位点(丝氨酸861和864)通过与c端结构域小磷酸酶-1(ctdsp1)的相互作用调节神经元分化2。当ctdsp1从丝氨酸861和864中去除磷酸盐时,rest蛋白被稳定,神经元分化受到抑制2-4。技术实现要素:6.本文公开了用于开发对ctdsp1具有高亲和力的肽的方法、化合物和组合物。7.本文公开了用于结合c端结构域小磷酸酶1(ctdsp1)的方法、化合物和组合物。8.本文公开了结合ctdsp1以抑制rest活性的rest类磷酸肽tedleppepplpken(seqidno:1)、edleppepplpk(seqidno:15)、nekplppeppeldet(seqidno:16)和kplppeppelde(seqidno:17)。小写字母代表d-氨基酸,其抗降解,因此增加了肽半衰期,而不影响结合亲和力5。9.本文公开了九十八(98)种rest类磷酸肽变体(rppv)(seqidno:18至117),其在不同程度上抑制rest上的ctdsp1活性(图25)。10.本文公开了细胞内转运肽,例如细胞穿透肽(cpp)和/或内涵体释放序列(seqidno:118至137和140至159),其n端或c端可融合至rpp(seqidno:1和15至17)或rppv(seqidno:18至117)以改善细胞内转运。还公开了可插入rpp或rppv与表7中所列肽之一之间以进一步改善细胞内转运的接头(seqidno:138、139、160和161)。11.本文公开的了n端或c端融合到细胞内转运肽(seqidno:118至137和140至159)并环化以进一步提高结合亲和力和稳定性(增加肽半衰期)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)。环化融合蛋白的示例为seqidno:2、5、12、13和14。12.本文公开了n端或c端融合到细胞内转运肽(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至137和140至159),促进rest蛋白质的降解。融合蛋白的示例为seqidno:2和4至14。13.本文公开了n端或c端融合到细胞内转运序列(seqidno:118至137和140-159)的如seqidno:1和15至17的rest类磷酸肽或seqidno:18至117的rppv,促进rest靶基因的表达。融合蛋白的示例为seqidno:2、4至14。14.本文公开了n或c端融合到细胞内转运肽(seqidno:118至137和140至159)的如seqidno:1和15至17的rpp或rppv(seqidno:18至117),通过向动物施用有效治疗量的化合物,以增加rest靶基因的表达或增加bdnf的表达,用于治疗患有与rest或ctdsp1相关疾病或病变的动物。融合蛋白的示例为seqidno:2和4至14。15.本文公开了融合到细胞内转运肽(seqidno:118至137和140至159)的rppseqidno:1和15至17或rppv(seqidno:18至117),可用于治疗动物的创伤性脑损伤、慢性疼痛、周围神经损伤、癫痫、糖尿病、阿尔茨海默病、亨廷顿病、脑肿瘤(包括多形性胶质母细胞瘤),或者胰腺癌。融合蛋白的示例为seqidno:2和4至14。附图说明16.随附附图包含在本说明书中,并构成本说明书的一部分。附图与上面给出的一般描述以及下面给出的示例性实施例和方法的详细描述一起,用于解释本发明的原理。在此类附图中:17.图1.(a)示出了ctdsp1结合位点位置的全长rest蛋白。(b)环状形式的与细胞穿透肽整合的反向倒置(ri)rest类磷酸肽。18.图2.使用monolith(nanotemper)测定其余类磷酸肽(rpp)对ctdsp1的结合亲和力。rpp(表4;seqidno:165)与ctdsp1(表2;seqidno:163)的亲和力约为130pm。用red-trisnta染料(20nm)荧光标记ctdsp1(40nm)上的his标签,并测定rpp(高达0.5μm)结合时荧光的变化,以评估结合。19.图3.使用biacore(gehealthcarelifesciences)测定rpp对ctdsp1的结合亲和力。将ctdsp1(表2;seqidno:163)固定在cm5传感器芯片(gehealthcarelifesciences)上,并测定(a)gst对照或(b)rpp(表4;seqidno:165)的结合亲和力。20.图4.rpp具有细胞渗透性,并在细胞核内聚集。(a-d)将100nmflag-rpp-cpp(caqkdykddddktedleppepplpkengrkkrrqrrrg(seqidno:4))添加到间充质祖细胞(mpcs)中并培养4小时。培养6天后,rpp集中在细胞核内。(e,f)在6天的时间内,肽从细胞质转移到细胞核(标准差,n=3)。(a)细胞骨架(phalloidin);(b)细胞核(hoechst染料);(c)rpp(flag抗体);(d)合成图像。21.图5.rpp积聚在脊髓坐骨神经核内。sprague-dawley(sd)雄性大鼠l3和l4区0.7cm的冠状脊髓切片,显示坐骨神经周围的细胞核,在大腿中部坐骨神经缺损损伤处注射1mg(1mg/ml)肽48小时后采集。a.hoechst染色显示细胞核。b.flag抗体示出了flag-rpp-cpp(seqidno:4)定位。22.图6.rpp降低rest蛋白水平。(左)示出了在用溶剂或100nmflag-rpp-cpp(seqidno:4)处理4小时后hek293细胞中外源性表达的rest蛋白水平的蛋白免疫印迹。用抗rest(milliporesigma,07-579)和抗gapdh检测印迹。(右)示出了100nmflag-rpp-cpp使rest蛋白水平降低58%的条形图。通过使用imagej测定凝胶峰面积来计算条带强度。23.图7.rpp增加bdnf表达。在培养24小时后,用flag-rpp-cpp(seqidno:4,100nm)或溶剂(pbs)培养4小时,用qpcr测定hek293细胞中的mrna水平。bdnf(2-ddct)表达的平均倍数变化为2.401±0.885(标准差,n=3)。平均倍数变化1=无变化。根据livak等,2001的描述计算2-ddct。24.图8.ctdsp1抑制后48小时,nbfl细胞中kv4.3的mrna水平。用表达带有gfp( )的rpp(seqidno:1)的质粒或带有gfp(-)的对照肽转染细胞。转染24小时后,根据荧光强度对细胞进行分选。误差棒表示标准偏差,n=2。25.图9.线性和环化rpp的比较。nbfl细胞被施用(1μm)rpp(线性_seqidno:4;环化_seqidno:2,表8)或对照肽(seqidno:3,表9)24或48小时。通过测定(a)bdnf、(b)ngf和(b)kv4.3转录水平的变化来评估疗效。n=1。26.图10.两名患者在用水(对照)或3μmrpp(seqidno:9)孵育48小时后,间充质祖细胞(mpc)中bdnf和ngfmrna的表达(归一化为肌动蛋白)。mrna水平相对于对照组示出(标准偏差,n=2)。27.图11.给ipsc(神经干细胞(nsc)-nl5)施用一条如下所示的rpp序列(3μm)或水对照。培养7天后,使用微管相关蛋白2(map2)神经元标记物(归一化为dapi)评估细胞的神经元分化。第3天更换培养基和变体。条形图中,map2水平为相对于对照(标准偏差n=4)。[0028][0029]图12.ipsc(神经干细胞(nsc)-nl5)采用1μmrpp(seqidno:9)或水对照。培养7天后,使用神经元标志物tuj1(iii类β-微管蛋白)和map2(均归一化为dapi)评估细胞的神经元分化。第3天更换培养基和rpp。(标准偏差,n=6)。[0030]图13.ipsc(神经干细胞(nsc)-nl5)采用1μmrpp(seqidno:13)或水对照。培养7天后,使用神经元标志物tuj1和map2(均归一化为dapi)评估细胞的神经元分化。第3天更换培养基和rpp。(标准偏差,n=6)。[0031]图14.rpp(seqidno:12)增加kv4.3、kv7.2、nav1.8和oprm1的表达。在成年sprague-dawley(sd)大鼠l5drg与不同浓度的rpp孵育48小时后,相对于水对照,对mrna表达进行qrt-pcr分析。*=p《0.05,***=p《0.01,***=p《0.001,单向方差分析和dunnett检验,误差棒=平均值的标准误差。[0032]图15.3μmrpp持续48小时(seqidno:13和14)可增加nav1.8的表达。rpp与成年sprague-dawley(sd)大鼠l5drg孵育后,对相对于水对照组的mrna表达进行qrt-pcr分析(归一化为β-肌动蛋白)。误差棒=平均值的标准误差。[0033]图16.rpp(seqidno:12)增加bdnf和ngf的表达。在成年sprague-dawley(sd)大鼠l5drg神经元与不同浓度的rpp孵育48小时后,对相对于水对照组的mrna表达(归一化为b-肌动蛋白)进行qrt-pcr分析。*=p《0.05,单因素方差分析和dunnett检验,误差棒=sem。[0034]图17.rpp(seqidno:12)不会导致drg神经元坏死。在用水、rpp(1、3或10μm)或triton培养48小时的sd大鼠中进行ldh毒检测。***=p《0.001,单因素方差分析和dunnett检验,误差棒=sd。[0035]图18.sni后慢性疼痛基因的rna水平。sni后第0天(假手术组,n=3)、第7天(n=5)或第28天(n=2)相对于β-肌动蛋白的rna倍数变化(平均值±sd,**p《0.01,***p《0.001,采用dunnett多重比较试验进行单因素方差分析。rna从坐骨神经drg(l5)中分离。[0036]图19.tbi后restmrna水平升高。(a)未受伤小鼠同侧皮质中restmrna表达的正常水平。(b)皮质控制性撞击伤后7天,小鼠同侧皮质的rest水平显著增加6。通过rnascope用dapi反染色示出的mrna。[0037]图20.(a)用于工程化ncrm-1ipsc的aavs1nanoluc-halotag敲入(ki)构建物示意图。将cmv驱动的构建物插入19q染色体的安全港(safeharbor)aavs1位点。通过转录激活物样效应器核酸酶(talens)。通过测序和连接pcr(未示出)证实aavs1ki。(b)用于工程化ncrm-1ipsc的map2nanoluc-halotag敲入(ki)构建物示意图。通过测序和连接pcr(未示出)证实,使用锌指核酸酶(zfn)和框架ki中的map2将纳米荧光素酶-halotag敲入map2转录起始位点(tss)(2号染色体)。cmv-nlht和map2-nlhtipsc表达多能性标记oct4/nanog/tra1-81/tra1-60(未示出)。(c)cmv-nlhtipsc和map2-nlht神经干祖细胞(nspc)和神经元的荧光素酶检测。注意,随着map2-nlht细胞的神经元分化,荧光素酶活性增加,这与随着分化,map2表达增加一致。[0038]图21.大鼠坐骨神经损伤模型:(左)坐骨神经切断手术过程。(右)坐骨神经切片免疫组化免疫荧光染色。对组织进行βiii微管蛋白、层粘连蛋白i和dapi染色。比例尺=100μm。[0039]图22.大鼠坐骨神经采集:(左)大鼠脊柱和相关周围神经的整体切片。(右)坐骨神经损伤部位与相关腰神经根的图像。[0040]图23.a.与基线(基础)相比,sni在损伤后4天(n=16)诱导机械痛阈(mt)致敏;配对t检验,*p《0.05,假手术组术后4天机械阈值无显著差异。b、单次皮下注射吗啡对创伤后mt的镇痛作用(sni mor,n=4)。平均值±sem;治疗前后单因素方差分析f=4.62,*p《0.05;经bonferroni多重比较检验,平均差和95%可信区间分别为-7.3、-14.5至-0.135,p《0.05。20mg/kgmor(n=4;f=19.03,**单因素方差分析p《0.001)。[0041]图24.在sni小鼠中,用生理盐水(对照组)和4剂量吗啡(mor)形成条件反射(condition)后的条件性位置偏好(cpp)。mor诱导的cpp呈剂量依赖性(单因素方差分析,f=4.167,p《0.05);通过tukey多重比较试验,10mg对对照组(p《0.001)和20mg(p《0.05)。[0042]图25.暴露于1μmrpp(seqidno:1)或其一种变体(表3)后,作为底物的rest肽(tedpspppspplpken)的体外ctdsp1磷酸酶活性(相对于gst对照)。[0043]图26.相对于水对照,在暴露于rpp(seqidno:12)后,磷酸化rest肽(tedpspppspplpken)作为底物的体外靶向(ctdsp1)和非靶向磷酸酶活性(1=ppa1,2=ppm1h,3=ppm1a,4=pp3ca,5=ppp1ca,6=pp5ca)。n=5,误差棒为标准偏差。[0044]图27.构建rpp库。a)实验方案;b)rest构建物的核苷酸序列(突变前);c)其余构建物的蛋白质序列(突变前)。rpp、cpp和his标签都有下划线。[0045]图28.rpp(seqidno:13)结构。沿着结构的底部是cpp,它含有一个多精氨酸序列和两个2-氨基十四烷酸,这些酸与其余的ri类磷酸结合(从结构右下方的赖氨酸开始)。具体实施方式[0046]在一个实施方案中,本发明提供了开发与ctdsp1具有高亲和力的肽的方法(如实施例1至3所述)。重要的是,该方法使用肽进化技术生成与ctdsp1具有高亲和力的肽。[0047]在一个实施方案中,本发明提供seqidno:1和15至17的肽或与seqidno:1和15至17具有至少50%、更优选至少60%、更优选至少70%、更优选至少80%、更优选至少90%或更优选至少95%相似性的肽。这些肽或其类似肽在本文中称为rest类磷酸肽(rpp)。然而,重要的是,必须保持rpp中的谷氨酸,尤其是seqidno:1的位置5和8、seqidno:15的位置4和7、seqidno:16的位置8和11以及seqidno:17的位置6和9处的谷氨酸。[0048]在一个实施方案中,本发明提供rppv(seqidno:18至117)或与rppv(seqidno:18至117)具有至少50%、更优选至少60%、更优选至少70%、更优选至少80%、更优选至少90%、更优选至少95%相似性的肽。所述肽或其类似肽在本文中称为rest类磷酸肽变体(rppv)(seqidno:18至117)。然而,重要的是,必须保持seqidno:18至67中的谷氨酸,尤其是seqidno:1中5和8位置的谷氨酸。对于反向倒置序列(rppvri)(seqidno:68至117),必须保持seqidno:16的位置8和11。[0049]在一个实施方案中,本发明是rpp(seqidno:1和15至17)或rppv(seqidno:18至117)以及细胞内转运肽(seqidno:118至137和140至159)(或与之具有至少50%、更优选至少60%、更优选至少70%、更优选至少80%的肽、更优选至少90%或更优选至少95%相似性的肽)的融合蛋白。[0050]两个或多个核苷酸序列或两个或多个氨基酸序列之间的同一性/相似性通过序列之间的同一性或相似性来表示。序列同一性可以用同一性百分比来衡量;百分比越高,序列越相同。序列相似性可以用相似性百分比来衡量(考虑保守的氨基酸替换);百分比越高,序列越相似。[0051]用于比较的序列比对方法在本领域是众所周知的。各种程序和比对算法如:smith&waterman,adv.appl.math.2:482,1981;needleman&wunsch,j.mol.biol.48:443,1970;pearson&lipman,proc.natl.acad.sci.usa85:2444,1988;higgins&sharp,gene,73:237-44,1988;higgins&sharp,cabios5:151-3,1989;corpet等人,nuc.acidsres.16:10881-90,1988;huang等人,computerappls.inthebiosciences8,155-65,1992;andpearson等人,meth.mol.bio.24:307-31,1994.altschul等人,j.mol.biol.215:403-10,1990详细介绍了序列比对方法和同源性计算。[0052]ncbi基本局部比对搜索工具(blast)(altschul等人,altschul等人.,j.mol.biol.215:403-10,1990)可从多个来源获得,包括国家生物信息中心(ncbi,国家医学图书馆,38a栋,8n805室,bethesda,md20894)和互联网,用于序列分析程序blastp,blastn,blastx,tblastn和tblastx。有关更多信息,请访问ncbi网站(www.ncbi.nlm.nih.gov)。[0053]blastn用于比较核苷酸序列,blastp用于比较氨基酸序列。如果两个比较的序列共享同源性,那么指定的输出文件将把这些同源区域表示为比对的序列。如果两个比较的序列没有同源性,那么指定的输出文件将不会显示比对的序列。[0054]比对时,匹配的数量通过计算两个序列中存在相同核苷酸或氨基酸残基的位置数量来确定。通过将匹配数除以已识别序列中所述序列的长度,或除以分节的长度(例如,已识别序列中所述序列的100个连续核苷酸或氨基酸残基),然后将所得值乘以100来确定序列一致性百分比。例如,当与具有1154个核苷酸的测试序列比对时,具有1166个匹配的核苷酸序列与测试序列(1166/1554*100=75.0)75.0%相同。百分比序列标识值四舍五入到最接近的十分位。例如,75.11、75.12、75.13和75.14四舍五入为75.1,而75.15、75.16、75.17、75.18和75.19四舍五入为75.2。长度值将始终为整数。在另一个示例中,包含15个核苷酸区域的靶序列,该区域与以下识别序列中的20个连续核苷酸比对,包含与该识别序列具有75%序列同源性的区域(即,15/20×100=75)。[0055]对于大于约30个氨基酸的氨基酸序列的比较,使用设置为默认参数的默认blosum62矩阵,使用blast2序列函数(空位(gap)存在代价(cost)为11,每个残基的空位代价为5/1)。同源物的典型特征是拥有至少70%的序列同一性,在与氨基酸序列的全长比对中使用ncbibasicblast2.0,具有nr或swissprot数据库等数据库的空位blastp。用blastn程序搜索的查询用dust过滤(hancock和armstrong,1994,comput.appl.biosci,10:67-70)。其他程序使用seg。此外,还可以执行手动比对。当用这种方法评估时,具有更大相似性的蛋白质将显示出越来越多的百分比同一性,例如与蛋白质至少有75%、80%、85%、90%、95%、98%或99%的序列同一性。当比对短肽(少于约30个氨基酸)时,使用blast2序列功能执行比对,使用pam30矩阵设置为默认参数(开头空位9,延长空位1罚分)。当用这种方法评估时,与参考序列更为相似的蛋白质将显示出越来越多的同一性,例如与蛋白质至少60%、70%、75%、80%、85%、90%、95%、98%或99%的序列同一性。当比较的序列同一性小于整个序列时,同源物通常在10-20个氨基酸的短窗口内拥有至少75%的序列同一性,并且可以拥有至少85%、90%、95%或98%的序列同一性,这取决于它们与参考序列的同一性。ncbi网站描述了在如此短的窗口中确定序列同一性的方法。[0056]两个核酸分子密切相关的一个指标是,如上所述,这两个分子在严格的条件下相互杂交。尽管如此,由于遗传密码的退化性,未示出高度同一性的核苷酸序列可能编码相同或相似(保守)的氨基酸序列。利用这种简并性可以改变核苷酸序列,从而产生多个核酸分子,这些核酸分子都编码基本相同的蛋白质。两个核苷酸序列基本相同的另一个(不一定是累积的)指标是,第一个核酸编码的多肽与第二个核酸编码的多肽在免疫上发生交叉反应。[0057]在某些实施方案中,rpp(seqidno:1和15至17)或rppv(seqidno:18至117)可与另一肽融合,以便例如允许细胞穿透。优选地,将rpp(seqidno:1和15至17)或rppv(seqidno:18至117)融合到允许细胞穿透和从内涵体逃逸的肽序列。更优选地,细胞穿透和逃逸肽序列是一个seqidno:118至137和140至159。rpp(seqidno:1和15至17)或rppv(seqidno:18至117)和细胞穿透肽(cpp)(seqidno:118至137和140至159)可直接融合,或包含连接它们的连接序列(例如,seqidno:138、139、160或161)。已经表明,当细胞或组织被rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽与cpp(seqidno:118至137和140至159)融合时,其定位于细胞核(图3,seqidno:4和图4_seqidno:4),导致rest降解(图5,seqidno:4),增加rest靶向基因的表达(图7、seqidno:4、图8、seqidno:1、图9、线性_seqidno:4和环化_seqidno:2、图10、seqidno:9、图14、seqidno:12、图15、seqidno:13和14以及图16、seqidno:12),并导致神经分化(图11、seqidno:5至11、图12、seqidno:9和图13、seqidno:13)。[0058]本发明人发现rpp(seqidno:1和15至17)结合ctdsp1以抑制rest活动。如图2和3所示的结合曲线所示,rpp对ctdsp1有较低的pm亲和力。在细胞和动物中,rpp与ctdsp1结合,从而阻止ctdsp1与rest结合。[0059]rest的增加和相关的神经基因抑制是以下疾病或病变的病理学的基础:[0060]创伤性脑损伤-创伤性脑损伤(tbi)导致神经元死亡导致认知能力丧失。为了改善tbi后的认知恢复,需要解决两个问题:1)减少损伤后持续的神经元死亡;2)使丢失的神经元再生。[0061]tbi后,大脑中的rest水平显著增加(图19)。几项研究一致表明,急性缺血引起的脑损伤诱导神经元表达rest,导致神经元死亡7-9。这些研究表明,当消除rest时,神经元存活率会提高7-9。其临床意义深远。例如,rest水平的提高与癫痫发作频率的增加相关,癫痫发作通常发生在脑损伤后10-12。在脑损伤的动物模型中,当rest受到抑制时,癫痫发作率显著降低13-14。此外,脑外伤后长期rest水平升高可能在脑损伤史引起的神经退行性疾病中起作用15-17。体外和体内研究都表明,消除rest可以提高神经元的存活率7-9、功能14,18和再生2,19-24。因此,tbi后靶向rest应该可以减轻脑损伤的影响,并抵消形成相关的年龄相关神经退行性疾病(如阿尔茨海默病)的风险17,25。[0062]神经发生改善tbi患者认知功能的基础是将神经发生的增加与学习、记忆和其他认知功能的改善联系起来的研究26-30,以及表明通过抗有丝分裂剂、辐射或基因操纵抑制神经发生会损害海马依赖型记忆的相反研究29-33。神经发生缺陷也与许多认知病因疾病有关,包括发育障碍(如小头畸形34、巨脑症35、自闭症35)和神经退行性疾病(如痴呆和阿尔茨海默病)36。实验治疗(包括输血、生长和神经营养因子以及干细胞移植10,37-49)专注于再生神经元。这些尝试未能刺激恢复认知功能所需的神经发生,至少部分是因为它们不能在tbi的情况下最终分化神经祖细胞26,28,50-57。re1沉默转录因子rest在单个检查点阻断了神经祖细胞向神经元的终末分化。[0063]周围神经损伤-周围神经损伤(pni)患者的神经元再生至少通过两种方式受到抑制。首先,支持发育和成熟神经元生长、存活和分化的神经营养因子(ntfs)减少58。其次,突触可塑性(包括轴突生长、囊泡转运和离子传导)所需基因的表达减少59。这两种现象与损伤诱导的rest表达一致60。因此,通过逆转神经再生障碍,消除rest可以加速pni的恢复。用rpp抑制ctdsp1可促进rest降解(图6),增加神经元分化(图11、12和13),并增加ntf表达(图7、9、10和16),这表明融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)刺激神经再生的潜力。[0064]慢性疼痛-慢性疼痛期间中枢和外周循环中被抑制的许多基因是抑制元件-1(re1)-沉默转录因子(rest)的直接或间接靶点,rest是干细胞、神经祖细胞和非神经细胞中神经元基因表达的抑制剂61-63。正常神经元的rest水平通常保持在较低水平,通过主动降解使染色质远离这种强大的神经元基因阻遏物64-69。在向神经病理性疼痛过渡之前,周围神经损伤后,小鼠和大鼠周围神经神经元的rest水平显著增加,随后中枢神经系统中处理和感受疼痛刺激的神经元出现峰值60,70,71。神经损伤后rest的激活导致感觉神经元正常兴奋性所需的几个基因,包括钾通道kv4.3(kcnd3)和kv7.2(kcnq2),钠通道nav1.8(scn10a)和μ阿片受体oprm1(图18)表达减少60,72-74。阻断rest以缓解慢性疼痛的基础发表于利用小鼠和大鼠周围神经损伤(pni)模型发表的研究60,72-74。在小鼠中,pni后感觉神经元中rest的基因敲除恢复了μ阿片受体60,72,73、nav1.860,72,73、kv4.372,73,75、kv7.272,73在背根神经节的基因表达,恢复c神经纤维的正常k m型电流60,减少痛觉过敏和痛觉超敏72-74,并恢复了慢性疼痛模型中的吗啡镇痛(表12)。使用msin3结合位点(msin3结合位点是一种适配器蛋白)的优化rest模拟物破坏rest抑制复合物,有助于许多基因的转录抑制,包括rest靶点,恢复c纤维功能,减少痛觉过敏和痛觉超敏,并恢复慢性疼痛小鼠模型中的吗啡镇痛(表12)74。在大鼠中,敲低rest控制了从急性疼痛到慢性疼痛的转变,表现为痛觉过敏和痛觉超敏的减少,并恢复了毒蕈碱镇痛(表12)72,73。[0065][0066][0067][0068]我们认识到,表12所示的研究表明,小鼠条件性敲除(cko)研究(完全敲除)与大鼠sirna敲除研究在降低超敏反应方面存在差异,后者显示损伤后没有超敏反应,也没有下调rest靶基因(表12),这只能部分减少损伤后的痛觉超敏(高达45%)和痛觉过敏(高达65%)。我们认为,敲除效应比sirna敲除更有效,因为sirna的生物利用度受到其大小(》13000kd)和因内涵体包埋导致的细胞运输不良的限制。[0069]我们知道,表12中的研究缺乏严谨性,因为没有进行功效分析来确定每组动物的数量,并且动物数量很低,这使得统计分析有问题。一些研究使用预测性较差的部分坐骨神经结扎(psnl)模型,而不是首选的备用神经损伤(sni)慢性疼痛模型。这些研究中的大多数只评估雄性。然而,尽管存在这些缺点,但研究结果是一致的,因此,支持我们提出的治疗慢性疼痛药物的体内疗效评估。此外,由于rest活动在哺乳动物中高度保守,我们相信这些研究为测试人类rest调节提供了强有力的概念验证支持。[0070]我们发现rest是通过其丝氨酸861和864的磷酸化来降解的。rest通过c端结构域小磷酸酶1(ctdsp1)使这些位点去磷酸化而稳定2-4。与rest一样,ctdsp1在非神经组织中表达,通过与rest的相互作用促进神经基因沉默。我们得出结论,抑制ctdsp1足以促进rest降解并刺激基因表达。为了支持这一结论,我们已经证明,间充质祖细胞(mpc)和神经中ctdsp1的敲除可导致rest靶基因的表达和轴突再生76。我们利用我们对rest-ctdsp1相互作用的机制理解,开发了一种包含rest相关调节区域的环化类磷酸肽77。其余的类磷酸肽(rpp)有几个药理学属性。由于其环化结构和d-氨基酸组成(例如,seqidno:9、12、13、14、16、17和68-117),它非常稳定。它具有超过大多数抗体的较低pm结合亲和力(图2和图3),表明发生脱靶效应的风险较低。其小尺寸(《3.5kd)表明其生物利用度应接近小分子药物。我们的初步数据表明,我们的模拟物抑制ctdsp1的活性,降低rest蛋白水平,并导致适当神经元活动所需的神经基因的表达77。[0071]最重要的是,我们的方法预计安全有效,原因如下:1)我们的药物对诸如神经损伤后的再生反应的核心靶点具有选择性。染色质的基本组织不允许表观遗传“书写”一组新的指令;然而,神经损伤会触发协同、广泛和瞬时的核小体重组,即基因组瞬时中间状态(gtis):一种有利于表观遗传重编程以获得必要反应的临时核小体结构78-81。2)我们的药物具有很高的结合亲和力(低pm),因此不会产生脱靶效应(图2和图3)。3)我们建议短治疗期少于1个月我们缩短治疗周期的基础是rest是神经元终末分化的守门员64,82(将细胞不可逆地提交给该谱系的过程83)。神经祖细胞,与慢性疼痛状态下的神经元具有表观遗传相似性63,因为它们也在gtis中,诱导几天后进行最终分化。与之一致地,我们证明,rpp可以在7天内诱导神经元分化(通过map2阳性细胞测定,map2是神经元分化的标志物)(图11、12和13)。一旦该过程开始并且细胞被提交(committed),就不需要进一步治疗。因此,我们期望通过这一早期治疗期实现的疼痛缓解能够持久。4)在测定坏死的离体drg神经元毒性检测中,我们已经证明我们的药物对神经元没有毒性(图17)。5)rest和ctdsp1主要参与神经发育,因此,我们预测抑制它们在受损的成年细胞中的活性将得到很好的耐受性。6)多肽药物的安全性使其成为一种有吸引力的治疗策略。目前,已上市的肽类药物超过68种,全球销售额超过147亿美元,140种正在临床开发中84,85。[0072]创新点[0073]一流的药物候选。我们开发了第一种靶向神经基因转录抑制物rest的药物。已发表的文献表明,rest表达导致慢性疼痛(表12)。pni后,外周神经系统(图18)和中枢神经系统的rest和ctdsp1表达均增加,导致功能障碍7-9。几项发表在啮齿动物身上的研究表明,阻断rest可以缓解慢性疼痛60,72-74。[0074]创新的药物设计。我们已经开发出一种环化类磷酸细胞穿透肽,它通过抑制ctdsp1(可保护其余部分不被降解一种磷酸酶)来破坏rest的稳定性3,从而允许神经特异性基因的表达。与rest一样,ctdsp1的表达仅限于非神经元细胞类型,神经元损伤后除外。我们的抑制剂是新颖的,因为它直接针对转录检查点,该检查点可立即调节诱导再生所需的基因。它克服了靶向转录因子和创建抗丝氨酸磷酸酶疗法的难题。关于后者,其他方法侧重于磷蛋白磷酸酶(ppp)和金属依赖性蛋白磷酸酶(ppp)86。然而,ctdsp1是一种卤酸脱卤酶(had),它使用两种催化型天冬氨酸,而不是仅仅依靠金属离子发挥活性87。这一催化位点能够受到特异性抑制,成功开发出针对类似酶(如hiv-1蛋白酶)的治疗药物证明了这一点88-91。最后,ctdsp1具有脯氨酸依赖性底物偏好,这在其他丝氨酸磷酸酶中没有发现,我们利用它来改善药物的结合和稳定性。[0075]初步数据。rest通过与神经元基因调节区附近的阻遏元件-1(re-1)位点的染色质结合来抑制基因表达,包括离子通道、生长因子和轴突导向蛋白1。因此,在干细胞如神经祖细胞(npc)能够最终分化之前,它们必须首先以rest为降解靶点,以表达所需的神经元基因64。ctdsp1保护rest免受降解,ctdsp1是阻止神经元基因表达所必需且充分的2。显性负性ctdsp1能结合磷酸化靶点,但不能催化去磷酸化,诱导p19干细胞的终末分化92。[0076]我们之前已经证明,ctdsp1和rest之间的相互作用依赖于map激酶erk2对rest丝氨酸861和864的磷酸化,并且这些丝氨酸突变为丙氨酸会增加rest稳定性2-4。我们推测,该rest调节区的一种不可水解的类磷酸化可以减缓ctdsp1活性并促进rest降解。为了验证我们的假设,我们开发了一种rest类磷酸肽或rpp77以评估rest和rest靶标的剂量依赖性影响。rpp含有丝氨酸861和864突变为谷氨酸的rest的氨基酸858至870,其可以模拟磷酸丝氨酸的形状和总电荷。在c端,我们融合了分别来自hiv-tat和ha2的富含精氨酸的细胞穿透肽和内涵体退出序列(exit序列),用于将肽和蛋白质输送到培养的哺乳动物细胞和活生物体中,并促进血脑屏障(bbb)的穿越93-96。[0077]我们研究了rpp是否可以降低rest蛋白(图6)。在左图中,蛋白免疫印迹(wb)显示rpp降低了hek细胞的rest(外源 内源)水平。在右图中,通过wb分析对总rest蛋白(外源 内源)水平进行量化显示,rpp给药的hek细胞的总rest蛋白水平下降了58.3%(图6)。[0078]在体外结合检测中,rpp对ctdsp1的pm亲和力较低,且下降速度较慢(图2和图3)。在磷酸酶抑制的体外试验中,rpp在低nm浓度下抑制其靶点ctdsp1,但在10μm时不抑制非靶点pp5、pp1、ppm1h、ppm1a或pp3ca活性(图26)。在mpcs的细胞通透性和稳定性分析中,rpp易位到细胞核中(图4和图5),这与rest和ctdsp1的定位图谱一致92,97。培养6天后,rpp水平保持稳定。[0079]通过在啮齿动物慢性疼痛模型中阻断rest,可以防止nav1.8、kv4.3、kv7.2和oprm1的下调,其被认为是痛觉过敏和痛觉超敏的原因(表12),这一点已经得到证实。基于这些研究,我们测试了阻断ctdsp1是否可以刺激与慢性疼痛相关的rest靶基因的表达。我们还表明,转染rpp(不含hivtat-ha2)或施用线性或环化rpp的nbfl细胞的kv4.3mrna增加(分别为2倍和4倍)(图8和9c)。[0080]我们发现,与对照组相比,3μmrpp(seqidno:5-11)使人类神经祖细胞(ipsc)的分化增加了2至2.7倍,如归一化为dapi(细胞核)的map2(成熟神经元标记)表达的测定所示(图11),1μmrpp(seqidno:9和13)增加了人类神经祖细胞(ipsc)的分化(图12和13),如归一化为dapi(细胞核)的tuj1(分别高达36%和3倍)和map2(分别高达33%和2倍)(神经元标记物)表达的测定所示。与对照组相比,细胞死亡没有增加(数据未示出)。使用该筛选的基础是,消除神经祖细胞中的rest已被证明可诱导神经元分化22,这是神经元基因整体去抑制的指标。[0081]在疗效和毒性的初步评估中,在整个drg神经元的离体培养中,在0、1、3或10μm下对rpp进行48小时的评估,以确定其诱导kv4.3(图14)、kv7.2(图14)、nav1.8(图14和15)和oprm1(图14)表达的潜力,并引起神经毒性(图17)。3μm时,seqidno:12诱导kv4.3,kv7.2,nav1.8和oprm1分别约为对照组的0、6、3和3倍(图14)。3μm时,seqidno:13和14诱导nav1.8分别约6和13倍(图15)。10μm时,seqidno:12感应kv4.3,kv7.2,nav1.8和oprm1分别约为2、7、5和4倍(图14)。在乳酸脱氢酶ldh细胞毒性检测中,rppseqidno:12在0、1、3或10μm时未示出毒性(图16)。正如所料,阳性对照(triton,n=6))是有毒的(图17)。[0082]癫痫-癫痫是大脑中不受控制的电活动,导致混乱、意识丧失和不受控制的运动98。它是离子通道和包括scn1a(nav1.1)、scn2a(nav1.2)、scn1b(navβ亚基1)、kcnq2(kv7.2)和kcnq3(kv7.3)的受体失调的结果,这些受体都是rest抑制的靶标14,99。癫痫的根本原因是神经元核内rest水平的增加100。[0083]糖尿病-胰腺β细胞和神经元在分化过程中具有相似的转录途径,涉及rest的消除101。β细胞过度表达rest下调rest靶基因降低胰岛素分泌102,103。[0084]阿尔茨海默病——在阿尔茨海默病中,乙酰胆碱和胆碱乙酰转移酶(chat)是合成乙酰胆碱所必需的转移酶,含量非常低。这种酶浓度的降低会导致与阿尔茨海默氏症相关的记忆和认知缺陷。大脑rest水平的增加会抑制chat16。[0085]亨廷顿病-在亨廷顿病中,rest区从胞质向神经元细胞核的移位被认为是与该病相关的神经元退化的原因104-106。[0086]脑癌,包括多形性胶质母细胞瘤-脑癌,包括多形性胶质母细胞瘤(gbm),起源于脑肿瘤起始细胞(btic),这是一种具有强大自我更新能力的癌干细胞,可成为癌性肿瘤。这些细胞对放疗和化疗有抗性,并在治疗数月后变成增殖性的107-109。融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽具有通过终末分化btic阻止肿瘤形成的潜力。一旦细胞最终分化,它们就不能再增殖并产生新的肿瘤110。必须指出,终末分化与通过其他方法(例如通过组蛋白脱乙酰酶抑制剂(hdac)靶向染色质重塑)实现的分化不同。抑制这些酶会随意改变基因表达,无法实现预防复发性gbm所需的永久性终末分化。btic向神经元的终末分化在单个检查点被rest阻断。通过观察到rest水平的增加与gbm的复发相关,强调了这种抑制因子在维持致癌性方面的重要性111-113,以及更短的无病生存期112-114。btic依赖rest维持其致癌性为治疗干预创造了机会。rest通过丝氨酸861和864的磷酸化进行降解,并且这些丝氨酸通过c端结构域小磷酸酶ctdsp1保持在去磷酸化状态2。因此,用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15-17)或rppv(seqidno:18至117)抑制ctdsp1应能释放终末分化的rest-制动(brake),并防止gbm和其他脑肿瘤的复发。[0087]胰腺癌-晚期、转移阳性胰腺癌细胞的rest水平较高115。胰腺癌患者肿瘤中的rest水平较高,生存率较低115。体外功能实验表明,rest基因敲除可抑制胰腺癌细胞(aspc-1和panc-1)的增殖、迁移、侵袭和上皮-间充质转化115。体内实验(皮下balb/c裸鼠模型和肠系膜上静脉注射balb/c裸鼠模型)表明,敲除rest抑制了异种移植瘤的生长和转移115。因此,抑制rest可以改善这些疾病的患者预后。因此,本发明还涉及抑制细胞内rest的方法。通常,该方法包括将细胞与融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽接触。例如,它可以将细胞暴露足够长的时间,以使融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)进入细胞并对rest活动产生影响。该方法可在体外或体内实施。在体外实施的情况下,该方法可用于研究rest的活性,以测试其他化合物对融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)对rest的影响的补充或拮抗能力,或出于对研究人员重要的任何其他原因。当在体内实施时,该方法可用作对受试者一种或多种与rest相关的疾病或病变进行治疗的方法。根据本发明这方面的方法,优选地,rest的活性降低。接触细胞的步骤可以是使药剂物理接触一个或多个靶标细胞的任何操作。因此,可以通过将药剂直接添加到要接触的细胞的体外培养物中,并允许药剂有足够的时间在培养基扩散并接触至少一个细胞。同样,也可以通过向水环境中的细胞中添加药剂来实现。或者,它可以通过任何可接受的给药途径向受试者施用药剂,并允许受试者的身体通过自然过程将药剂分配给目标细胞。因此,体内方法可以是将药剂局部或系统地递送到动物细胞的方法,尤其包括所有哺乳动物和人类。根据该方面,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)可用于治疗或预防性治疗受试者,并制备用于治疗的组合物。[0088]在另一个实施方案中,本发明提供了一种治疗患有或有可能患有涉及rest的疾病或病变的受试者的方法。通常,该方法包括施用足以影响受试者rest量或活性的剂量的、融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)。在某些方面,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)与ctdsp1的结合导致细胞中rest活性的抑制。通常,该方法包括施用足量的融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)维持足够长的时间,以抑制rest活性。通常,给药量和时间足以看到疾病或病变的一个或多个临床症状的变化,或阻止疾病或病变进展到出现一个或多个临床症状的阶段。根据该方面,该药剂可用于治疗或预防性地治疗受试者,并制备用于治疗的组合物。[0089]在一个实施方案中,本发明提供了治疗、减轻或改善受试者的创伤性脑损伤、慢性疼痛、周围神经损伤、癫痫、糖尿病、阿尔茨海默病、亨廷顿病、脑肿瘤(包括多形性胶质母细胞瘤)或胰腺癌的方法。这里使用的术语“治疗”或“缓解”或“改善”以及类似术语包括预防和完全或部分治疗。这些术语还可以包括减轻症状、改善症状、降低症状的严重程度、降低疾病的发生率,或改善治疗结果的患者状况的任何其他变化。这些方法涉及对患有创伤性脑损伤、慢性疼痛、周围神经损伤、癫痫、糖尿病、阿尔茨海默病、亨廷顿病、脑肿瘤(包括多形性胶质母细胞瘤)或胰腺癌的动物受试者,或需要治疗创伤性脑损伤、慢性疼痛、周围神经损伤、癫痫、糖尿病、阿尔茨海默病、亨廷顿病、脑肿瘤(包括多形性胶质母细胞瘤)或胰腺癌的动物受试者,施用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)。[0090]可通过任何已知和可接受的途径向受试者施用融合至cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)。这些途径包括但不一定限于口服、通过粘膜(例如,经鼻、经吸入、经直肠、经子宫或经阴道、舌下)、经静脉(例如,静脉推注、静脉输注)、经腹腔和皮下。同样,可以通过直接注射到含有靶细胞(即待治疗细胞)的部位(例如,器官、组织)来施用。此外,施用可以遵循任何数量的方案。因此,它可以包括药物的单次剂量或给药,或一段时间内的多次剂量或给药。因此,治疗可涉及一次或多次重复施用步骤,直到达到预期结果。在实施方案中,治疗可以持续更长的时间段,例如周、月或年。本领域技术人员完全能够基于本领域的已知参数为个人轻松开发合适的给药方案。因此,这些方法也考虑控制但不一定消除疾病或病变。根据本发明的优选施用途径为口服和通过粘膜。[0091]根据受试者、疾病阶段、受试者年龄、受试者的一般健康状况以及医学领域技术人员已知并经常考虑的各种其他参数,施用量会有所不同。作为一般事项,将施用足量的药剂,以使受试者的症状发生可检测的变化。本文公开了合适的量,并且本领域技术人员可以在不进行过分(undue)或过度实验的情况下确定其他的合适量。[0092]融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)以受试者可接受、可耐受和有效的形式施用。本领域已知用于生物活性剂的多种药物形式和剂型,并且本发明设想了所有这些形式和剂型。因此,例如,该制剂可以配制成口服溶液、囊片(caplet)、胶囊、注射剂、不溶剂、栓剂、锭剂、片剂、乳膏或软膏、吸入剂等。[0093]本领域的普通技术人员将根据良好的医疗实践和个体受试者的临床状况,很容易优化有效剂量和施用方案。[0094]给药频率取决于化合物的药代动力学参数和施用途径。最佳药物配方将由本领域技术人员根据施用途径和所需剂量确定。此类制剂可能影响给药制剂的物理状态、稳定性、体内释放速率和体内清除速率。根据施用途径,根据体重、体表面积或器官大小计算合适的剂量。动物模型的可用性尤其有助于确定给定治疗药物的适当剂量。本领域的普通技术人员通常会进一步完善确定适当治疗剂量所需的计算,而无需进行不必要的实验,尤其是根据本文所公开的剂量信息和分析以及在动物或人类临床检测中观察到的药代动力学数据。[0095]通常,通过使用已建立的测定血液水平的分析以及相关剂量反应数据来确定适当的剂量。最终的给药方案将由主治医生决定,并考虑改变药物作用的因素,例如药物的比活性、损伤的严重程度和患者的反应性、患者的年龄、状况、体重、性别和饮食、任何感染的严重程度、施用时间和其他临床因素。随着研究的进行,将出现关于特定疾病和条件的适当剂量水平和治疗持续时间的进一步信息。然而,这些研究是常规的并且在本领域技术人员的水平之内。典型剂量可为每周(人体)约0.6mg(0.01mg/kg)至约60g(1g/kg),更优选每月。[0096]应了解,本发明的肽、组合物和治疗方法在人类医学和兽医学领域中有用。因此,要治疗的对象是哺乳动物,例如人类或其他哺乳动物。用于兽医目的,受试者包括例如农场动物,包括牛、羊、猪、马和山羊,伴侣动物,如狗和猫,外来动物和/或动物园动物,以及实验动物,包括小鼠、大鼠、兔子、豚鼠和仓鼠。[0097]融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)可作为药物或兽医学组合物(如片剂、胶囊、溶液或乳剂)施用给有需要的受试动物,优选哺乳动物,如人类。融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)可存在于含有其他成分的组合物中。适用于本发明的组合物的非限制性示例为药物组合物,例如片剂、丸剂、胶囊、囊片、多微粒(包括颗粒、珠子、球粒和微囊颗粒);粉末、酏剂、糖浆、悬浮液和溶液的形式。药物组合物通常包含医药上可接受的稀释剂或溶剂。药物组合物优选适于肠外给药(例如,口服)。可口服施用的组合物可以是固体或液体形式,可以是片剂、粉末、悬浮液和糖浆等形式。任选地,所述组合物包含一种或多种调味剂和/或着色剂。一般而言,治疗和营养组合物可包含不会显著干扰药剂对受试者的作用的任何物质。[0098]适用于此类组合物的药学上可接受的赋形剂或溶剂在药学领域是众所周知的。本发明组合物可含有0.01-99%(按重量计)的药剂。本发明的组合物通常以单位剂型制备。用于制备这些组合物的赋形剂在本领域是众所周知的。[0099]该组合物的产品形式的其他示例为食品补充剂,例如软凝胶或硬胶囊的形式,其包含选自明胶、淀粉、变性淀粉、淀粉衍生物(例如葡萄糖、蔗糖、乳糖和果糖)的封装材料。所述封装材料可任选地包含交联剂或聚合剂、稳定剂、抗氧化剂、用于保护光敏填料的吸光剂、防腐剂等。[0100]一般而言,在本技术中可使用溶剂一词来表示可与制剂混合的组合物,无论是药物溶剂、食品、营养补充剂或膳食辅助剂。就本发明而言,上述材料可被视为药剂的溶剂。在本发明的某些实施方案中,溶剂具有很少或没有生物活性,尤其是对rest。[0101]无需进一步描述,相信本领域普通技术人员可以使用前面的描述和下面的说明性实施例,制备和利用本发明的化合物,以及实施要求保护的方法。给出以下实施例以说明本发明。应当理解,本发明不限于实施例中描述的特定条件或细节。[0102]实施例1-his-ctdsp1质粒构建[0103]密码子优化(使用idt密码子优化工具)的ctdsp1基因被克隆到pbadhisa质粒中(thermofisherscientific)。首先,用引入hindiii和xhoi限制位点的引物p33和p34(表1)扩增pbad-hisa质粒。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸6分钟。扩增后,用qiagen凝胶条带纯化试剂盒对pcr片段进行凝胶纯化,并用hindiii和xhoi限制性内切酶消化。根据制造商的建议,使用qiagen试剂盒纯化所有消化片段,并使用t4dna连接酶(neb)以适当的组合连接。[0104]表1:用于质粒构建的引物[0105][0106][0107]根据制造商的方案,将连接片段转化到10g化学感受态细胞(lucigen)中。将转化细胞接种在含有50μg/ml氨苄西林的lb板上,并在37℃下培养过夜。通过菌落pcr检测菌落是否存在插入物。采集菌落并在20μl无菌0.9%氯化钠溶液中重新悬浮。将1μl该溶液转移到pcr管中,并用taq聚合酶(newenglandbiolabs,cat#m0482s)和30pm的侧翼引物进行扩增。最初将每个pcr反应(20μl)加热至95℃2.5分钟,然后94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸1分钟。琼脂糖电泳显示扩增产物。将具有正确插入物的克隆接种到含有5mllb和适当抗生素的培养管中,并在37℃下培养过夜。然后使用monarplasmidminiprep试剂盒(neb)纯化构建物。[0108]实施例2-his-ctdsp1的表达和纯化方法[0109]his-ctdsp1(表2;seqidno:162):将带有pbad-ctdsp1构建物的大肠杆菌(10g菌株lucigen)在37℃下剧烈摇动过夜。然后在1l烧瓶中,将4ml夜间培养物添加到500ml含有50μg/ml氨苄西林的lb培养基中,并在37℃下摇动培养。当培养物的od600达到0.4时,我们添加阿拉伯糖至最终浓度0.02%,并在30℃下摇动培养16小时。第二天早上,细胞以最大速度向下旋转(eppendorf离心机5810r),并在-80℃下冷冻。必要时,从冰箱中取出细胞颗粒,在室温下培养,用4mlbper蛋白裂解试剂(thermofisher)裂解,并如制造商所述用hispurcobalt纯化试剂盒(thermofisher,cat#90091)纯化蛋白。[0110]表2-ctdsp1核苷酸和蛋白质序列[0111][0112][0113]实施例3-开发对ctdsp1具有高亲和力的肽[0114]使用rna展示和蛋白质进化方法生成候选肽。[0115]rna展示文库的构建:我们已经构建了3个不同的rpp变体库。在所有文库中,丝氨酸861和864被谷氨酸盐取代。文库1是用引物p11到p14构建的(表1)。在文库1中,其余肽的每个位置都发生了突变。据估计有1×109个变体。在文库2中(引物p14至p18,表1),简并寡聚体未被谷氨酸盐修饰(touched)。该库预计有65×106个变体。在库3中,只有一个谷氨酸盐和每两个密码子发生突变。该库预计只生成65000个变体。[0116]在所有3个文库中,rpp序列(seqidno:1)通过合成简并寡核苷酸多样化。每个密码子只引入一个简并碱基:第一个或第二个位置,可以选择或4个氨基酸。其余盒(casssete)从pbad构建物中扩增为两个片段:左片段和右片段,通过连接重新结合(图27)。左侧片段用侧翼正向引物p23(表1)和一个反向引物(11r槽22r,表1)扩增。[0117]用一个正向引物(11f至22f,表1)和反向引物p24(表1)扩增右侧片段。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30循环,并在72℃下延伸40秒。扩增后,用qiagen凝胶条带纯化试剂盒对pcr片段进行凝胶纯化,并与t4dna连接酶混合和连接。连接反应包含20μl10x连接缓冲液、100ng片段混合物、0.5μl100mmatp、1μlt4dna连接酶(nebcat#m0202s)和1μlt4多核苷酸激酶。反应混合物在室温下孵育,并使用上述程序将其用作模板用侧翼引物p17和p20(表1)进行pcr。pcr片段通过qiagen凝胶条带纯化试剂盒进行凝胶纯化,并用作mrna展示实验的模板。[0118]出于质量控制目的,我们克隆并测序了该文库的一部分。用hindiii和xhoi限制性内切酶消化该文库的一小部分,用qiagen试剂盒纯化,并如上所述在pbad载体中克隆ctdsp1基因。通过genewiz对单个克隆进行测序,以确认rest盒成功突变,并且大多数克隆没有发生移码突变。[0119]体外转录:根据制造商协议,使用ribomax大规模rna生产系统t7(promega,cat#p1300)从扩增文库中翻译rna,并通过rneasy迷你试剂盒(qiagen,cat#74104)纯化。[0120]用嘌呤霉素将mrna连接至dna接头:采用idt法合成xl-pso寡核苷酸。寡核苷酸序列为:5’‑psoc6-(uagccggug)2’‑omc-aaaaaaaaaaaaaaa-spacer9-spaser9-acc-puro-3’。为了将该寡核苷酸连接到mrna,我们在pcr管中混合以下试剂:29.5μl无rna酶水。1μl1mhepes-koh,ph7.6,5μl1mkcl,2μl25mm亚精胺,0.5μl125mmedta,8μl上一步的mrna和4μl100mmxl-pso寡核苷酸。将pcr管置于pcr机中,加热至70℃5分钟,并以0.1℃/s的速度冷却至25℃。然后,我们将混合物转移到冰上的96孔板上,将365nm手持式紫外线灯放在顶部,并照射该板20分钟。之后,通过rneasymini试剂盒(qiagen,cat#74104)纯化交联rna。[0121]体外翻译:使用pureexpress体外蛋白质合成试剂盒(neb,cat#e6800s)进行翻译。我们在1.5ml试管中混合以下试剂:20μl溶液a、15μl溶液b、0.5μlrnasinplus、4.5μl水和10μl交联rna(1μg/l)。将混合物在37℃下培养2小时。[0122]用his标签纯化肽:我们使用ni-nta磁珠(qiagen,cat#36111)纯化rna肽复合物。用300μl洗涤缓冲液(50mmnah2po4,300mmnacl,10mm咪唑,0.005%吐温20)洗涤100μl珠子,在磁性支架上分离并悬浮在300μl洗涤缓冲液中。将前一步骤中的25μlrna肽复合物添加到洗涤过的珠子中,并在室温下在端对端摇床上孵育30分钟。用洗涤缓冲液洗涤珠子3次,然后在磁性支架上分离,并用50μl洗脱缓冲液(50mmnah2po4、300mmnacl、500mm咪唑、0.005%吐温20)洗脱。[0123]使用oligo-d(t)25磁珠进行纯化:oligo-d(t)25磁珠购自newenglandbiolabs(cat#s1419s)。用500μl洗涤缓冲液i(20mmtris-hcl,ph7.5,500mmnacl,1mmedta)洗涤100μl珠子悬浮液,将其置于缓冲液中并在磁性支架上分离,然后悬浮在50μl洗涤缓冲液i中。将前一步骤中的50μlrna肽复合物与50μl结合缓冲液(100mmtris-hcl,ph7.5,1mnacl,2mmedta)混合,在65℃下加热2分钟,放在冰上1分钟,并与洗过的珠子混合。该混合物在室温下孵育5分钟,然后用500μl洗涤缓冲液i(20mmtris-hcl,ph7.8,500mmnacl,1mmedta)洗涤两次,用500μl洗涤缓冲液ii(20mmtris-hcl,ph7.8,200mmnacl,1mmedta)洗涤一次。[0124]肽环化:在环化缓冲液(20mmtris-hcl,ph7.8,0.00mnacl,3mmα,α”‑二溴间二甲苯(sigma-aldrich),33%乙腈)中,将oligo-d(t)25磁珠结合的rna肽复合物孵育30分钟,并定期振荡。孵育后,用洗涤缓冲液iii(20mmtris-hcl,ph7.8,0.3mmnacl,5mm2-巯基乙醇)清洗珠子,然后用洗涤缓冲液iv(20mmtris-hcl,ph7.8,0.3mmnacl,0.5mmtcep)清洗珠子,然后用洗涤缓冲液i(20mmtris-hcl,ph7.8,500mmnacl,1mmedta)和洗涤缓冲液ii(20mmtris-hcl,ph7.8,200mmnacl,1mmedta)清洗珠子。通过添加30ul洗脱缓冲液(20mmtris-hcl,ph7.8)并在65℃下培养2分钟,然后立即在磁珠架上分离纯化产物,从磁珠中洗脱纯化产物。[0125]亲和性筛选:亲和性筛选在nuncmaxisorp板(thermofischerscientific)中进行。将his-ctdsp1(表2;seqidno:163)溶解于100μlpbs中,转移至maxisorp板孔中,并在室温下在轨道振动筛上孵育2小时。平板孔用pbs清洗两次,并在室温下用酪蛋白(pbsc缓冲液或含1%酪蛋白的pbs)封闭1h,摇动,然后用pbs清洗3次。阴性筛选孔仅通过与300μlpbsc溶液孵育并用pbs洗涤3次来包被酪蛋白。首先将纯化的rna肽复合物添加到这些含有100μlpbs的阴性筛选孔中,并在室温下摇动孵育20分钟,将该溶液转移到含有125μlpbsc的阳性筛选孔(由his-ctdsp1覆盖)中,并在室温下摇动培养1小时。通过向每个孔中添加25μl纯化的his-ctdsp1并孵育3分钟来进行脱靶筛选。随后用pbs清洗3次孵育孔并用于cdna合成。[0126]cdna合成和pcr:我们使用superscriptiii第一链合成系统(invitrogen,cat#18080-051)进行cdna合成。首先,将16μl水与2ul50mm引物p19(表1)和1μldntp溶液混合,并将该混合物添加到maxisorp板中的孔中。接下来,将该板在65℃下孵育5分钟,然后在4℃下冷却1分钟。然后我们将20μl混合物从板转移到pcr管中,并添加20μl反应混合物,其中含有4μl10x缓冲液、8μl25mmmgcl2、4μl0.1mdtt、2μlrna酶和2μlsuperscriptiii逆转录酶。该混合物在50℃下孵育50分钟。然后我们添加2μlrna酶h,并在37℃下孵育试管20分钟。[0127]使用引物p17和p20对在筛选中筛出的强结合物相对应的dna(表1)进行扩增。使用ventdna聚合酶进行扩增。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下20或变性15秒,55℃退火15秒,25个循环,并在72℃下延伸30秒。扩增后,在琼脂糖凝胶上运行pcr片段,根据制造商的方案,使用qiagen凝胶条带纯化试剂盒进行纯化。[0128]ngs数据分析。将每个周期的pcr反应进行下一代测序。我们使用了ampliconezserveicegenewiz。对unigene进行量化,并选择丰度最高的序列进行进一步测试(表3和3a)[0129]表3-rpp变体的蛋白质和核苷酸序列(seqidno在表19中标出)[0130][0131][0132][0133]表3b-rppri变体的蛋白质序列*(seqidno在表19中标出)[0134][0135][0136][0137]实施例4-肽-gst融合物的构建及其表达[0138]作为第一步,我们在pet29载体中克隆了gst蛋白。gst进行了密码子优化,两侧为hindiii和xhoi位点,由idt合成。用phusiondna聚合酶(neb,cat#m0530s)和引物p109和p108扩增(表1)。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸1分钟。扩增后,根据制造商的方案,使用qiagen凝胶条带纯化试剂盒对pcr片段进行凝胶纯化。[0139]使用引入hindiii和xhoi限制位点的引物p12和p14(表1)扩增pet29质粒。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸6分钟。扩增后,pcr片段用qiagen凝胶条带纯化试剂盒进行凝胶纯化,并用hindiii和xhoi限制性内切酶进行消化。所有消化片段用quiagen试剂盒进行纯化,并根据制造商的建议用t4dna连接酶(neb)以适当的组合进行连接。连接片段在10g化学感受态中转化,并如上所述进行测序。gst-rpp的表达构建物如表4所示(seqidno:164)[0140]表4-rpp-gst融合物的核苷酸和蛋白质序列[0141][0142]使用表5所示的引物,通过pcr重新创建所选序列,使用qiagen凝胶条带纯化试剂盒纯化凝胶,用hindiii和bamhi限制性内切酶消化,并在上述pet载体中克隆gst标签。根据制造商的方案,将连接片段转化到10g化学感受态细胞(lucigen)中。将转化细胞接种在含有25μg/ml卡那霉素的lb板上,并在37℃下培养过夜。第二天早上,通过菌落pcr检测菌落是否存在插入物。采集菌落并在20μl无菌0.9%氯化钠溶液中重新悬浮。将1μl该溶液转移到pcr管中,并用taq聚合酶(newenglandbiolabs,cat#m0482s)和30pmol的侧翼引物扩增。每个pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸1分钟。琼脂糖电泳显示扩增产物。将具有正确插入物的克隆接种到含有25μg/ml卡那霉素的培养管中,并在37℃下培养过夜。第二天早上,通过monarplasmidminiprep试剂盒(neb)纯化构建物。如上所述,将构建物转化为bl21(de3)感受态细胞(lucigen)。[0143]肽gst融合物的纯化:将含有所选构建物的大肠杆菌细胞(bl21菌株)在37℃下剧烈摇动过夜。第二天早上,我们在1l烧瓶中向100ml含25μg/ml卡那霉素的lb培养基中添加1ml夜间培养物,并在37℃下摇动培养。当培养物的od600达到0.4时,我们添加iptg至1mm的最终浓度,并在30℃下摇动培养16h。第二天早上,细胞在eppendorf离心机5810r中以最大速度旋转,并在-80℃下冷冻。必要时,从冰箱中取出细胞颗粒,在室温下培养,并用3mlbper蛋白裂解试剂(thermofisher)裂解。如制造商所述,使用谷胱甘肽琼脂糖(thermoscitificcat#16100)纯化肽gst融合物。[0144]表5-用于克隆框内带gst的rpp变体的引物[0145][0146][0147][0148][0149][0150][0151][0152][0153][0154]实施例5-rpp和rpp变体对ctdsp1磷酸酶活性的抑制[0155]结果:图25-在rna显示/蛋白质进化筛选中确定的前51个最丰富的rpp变体按照最丰富(v1)到最不丰富(v51)的肽顺序列在表3中。表5列出了用于制备这些肽的引物。磷酸酶活性筛选(表6,图25)用于评估前51位rppv抑制内源性磷酸化rest肽(tedpspppspplpken)氨基酸861和864处ctdsp1磷酸酶活性的能力。[0156]磷酸化rest肽(tedpspppspplpken)由genescript合成。在室温下,用10mmtrisph8、10mmmgcl2、100nmctdsp1、0.5μm磷酸盐传感器(phosphatesensor,thermofischer)、50μmrest肽和不同浓度的肽gst融合物进行磷酸酶反应10分钟。所有检测均在用水冲洗10次的96孔板(corningp/n3686)中进行。用bioteksynergyhtx在420/27nm激发和485/20nm发射下使用动力学读数进行荧光测定。[0157]筛选显示,与对照组(gst)相比,在1μm时,rpp(v1)抑制磷酸酶活性约60%(表6,图25)。一些变体抑制磷酸酶活性的效果优于rpp(表6,图25)。最有效的抑制剂是v33,其抑制率为100%;v35,其磷酸酶活性约为90%(表6,图25)。[0158]表6-rpp变体对ctdsp1磷酸酶活性的抑制[0159][0160][0161][0162]图26,表13-rpp,seqidno:12,抑制ctdsp1活性,ec50约为20nm,但不影响其他几种磷酸酶的活性。[0163]检测:如上文图26所述,评估rppseqidno:12抑制内源性磷酸化rest肽(tedpspppspplpken)氨基酸861和864处ctdsp1活性的能力。[0164][0165][0166]实施例6-使用monolith(nanotemper)和biacore(gehealthcarelifesciences)评估rpp的结合亲和力。[0167]如实施例2所述,构建并纯化his-ctdsp1(表2;seqidno:162)和gst-rpp(表4;seqidno:164)的表达构建物。ctdsp1(40nm)上的his标签用红色tris-nta染料(20nm)标记,染料与ctdsp1的比例为3:1,通过重力流尺寸排除去除未结合的染料。线性gst-rpp(表4;seqidno:165)与his-ctdsp1(表2;seqidno:163)的结合亲和力是根据制造商的建议,使用monolith(nanotemper)测定的。结合检测的结果如图2所示:在从低pm到0.5μm的几种浓度下,评估rpp(表4;seqidno:165)与his-ctdsp1的结合。对应于线性rpp与ctdsp1结合的kd在130pm下计算。[0168]使用gehealthcarelifesciences的biacore测定线性gst-rpp(表4;seqidno:165)与his-ctdsp1(表2;seqidno:163)的结合亲和力。按照biacore推荐的方案(cat#br-1000-50,gehealthcarelifescience),使用胺偶联将20μl(5g/ml)在10mm醋酸钠(ph值5.0)中的his标记的ctdsp1固定在cm5芯片上(gehealthcarelifescience)。这种纯化产生了约1300ru与cm5芯片结合的蛋白质。然后用hbs-ep缓冲液(ph7.4)清洗cm5芯片,在hbs-ep缓冲液(ph7.4)中以500nm的浓度以30μl/min的流速注入120μl分析物:gst-rpp或仅gst(阴性对照)。结合时间为2分钟,解离时间为3分钟。最后,用10mm甘氨酸hcl(ph1.5)(br-1003-54,gehealthscience)洗涤结合蛋白,以50μl/分钟的流速持续25秒。线性rpp与ctdsp1的结合导致:kd=1.7pm(图3b,阴性对照如图3a所示)。[0169]实施例7-与细胞穿透肽(cpp)和/或促进内涵体逃逸的肽融合的rpp和rppv[0170]合成了几种连接有细胞穿透肽(cpps)和/或促进内涵体逃逸的肽的rpp和rppv。我们使用的cpp和接头列表如表7所示。[0171]表7-允许细胞穿透和从内涵体逃逸的额外肽[0172][0173][0174]实施例8-rpp被间充质祖细胞和坐骨神经内化。[0175]间充质祖细胞(mpc)[0176]结果:在与rpp(seqidno:4)孵育4小时后,然后进行6天的培养,rpp在间充质祖细胞(mpc)中保持内化(图4、c和d),并在6天的培养期内定位在细胞核中(图4、e和f)。[0177]材料和方法[0178]给药和细胞培养:如前所述培养mpc116,将1.0×104个细胞/cm2铺板于24孔板中的玻璃盖玻片上,在37℃下培养过夜,加入100nmrpp(seqidno:4)4小时,然后更换培养基。细胞培养6天,每两天更换一次培养基。在24小时和72小时进行中期mpc采集,以评估细胞内rpp定位。[0179]免疫组化:phalloidin(invitrogen)用于细胞骨架染色,hoechst333429(calbiochem)染料用于显示细胞核,flag抗体(sigma-aldrich)用于显示rpp。图像用共焦激光扫描显微镜拍摄。[0180]坐骨神经[0181]结果:线性rpp积聚在坐骨神经组织的细胞核中(图5、a和b)。[0182]材料和方法[0183]给药和步骤:图5:对sprague-dawley大鼠的坐骨神经(l3和l4区域)(schmitz和beer,2001)117进行暴露,在神经残端回缩后,移除0.7cm的部分,形成约1cm的节段性缺损(hems和glasby,1993)118。将pbs中的1mgrpp(seqidno:4)注入受伤部位,并闭合手术切口。48小时后,处死动物,从l4到l6区域的脊髓冠状切片。[0184]合理剂量水平:最大可行剂量。[0185]供试品标识:rpp(seqidno:4)[0186]纯度:≥95%[0187]免疫组化:hoechst333429(calbiochem)染料用于显示细胞核,flag抗体(sigma-aldrich)用于显示rpp。图像用光学显微镜拍摄。[0188]实施例9-rpp诱导rest蛋白降解。[0189]结果:与溶剂对照组相比,rpp(seqidno:4)使rest蛋白水平降低了58%(图6)。[0190]材料和方法[0191]供试品标识:rpp(seqidno:4)[0192]纯度:≥95%[0193]给药和细胞培养:hek293细胞在添加10%小牛血清的dulbecco改良eagle培养基(dmem)中生长。用100nm全长人rest蛋白2,按照制造商的建议,使用lipofectamine2000(lifetechnologies)在35mm培养皿中80%至90%的汇合培养物中对hek293细胞进行转染。转染后24小时,用100nmrpp(seqidno:4)或溶剂对照(pbs)给细胞灌胃过夜。[0194]蛋白质印迹分析:按照ballas等人,2001的程序制备全细胞裂解物119。使用抗rest-c64,抗gapdh(abcam)及与红外染料结合的抗igg(thermofisher)的标准程序进行蛋白免疫印迹,并在odyssey红外荧光成像仪(licor)上进行分析。条形图是使用imagej对蛋白免疫印迹进行量化(https://imagej.nih.gov/ij)的结果。[0195]实施例10–rpp可用于诱导bdnf、ngf、kv4.3、kv7.2、nav1.8和oprm1mrna的表达,无神经毒性。[0196]结果.rpp增加bdnf、ngf、kv4.3、kv7.2、nav1.8和oprm1的表达(图7、8、9、10、14、15和16;表15)。rpp(seqidno:12)不会导致drg神经元坏死(图17),如在与不同浓度的rpp孵育48小时的drg神经元中进行ldh细胞毒性检测所示。***=p《0.001,n=4,单因素anova和dunnett试验,误差棒=sd。[0197][0198][0199][0200]图7:材料和方法[0201]供试品标识:rpp(seqidno:4),纯度:≥95%[0202]给药和细胞培养:100nmrpp;hek293细胞[0203]mrna提取和定量方案:细胞在qiazol(qiagen)中溶解,根据制造商的说明,使用rneasymidi试剂盒(qiagen)提取总rna。用nanodrop2000(thermofisher)对纯化的rna进行定量,并使用highcapacitycdna反转录试剂盒(appliedbiosystems)和每次反应2ng/l的rna进行反转录。对于qrt-pcr,将5μlcdna(相当于10ngrna)与10μlssoadvancedtmuniversalsupermix(biorad)和1μl每种引物(最终引物浓度各500nm)。在定量研究中,反应quatstudiotm7flex实时pcr系统(appliedbiosystems)重复三次。使用相对循环阈值(δdct)法和β-肌动蛋白作为校准物分析扩增数据。使用的引物如下:β-肌动蛋白正向:agagcacgagctgcctgac-3’,β-肌动蛋白反向:5'-ggatgccacaggactcca-3’,bdnf正向:5’tattagtgagtgggtaacggcg3’,bdnf反向:5’gaagtattgcttcagttggcctt3’。[0204]图8:材料和方法[0205]瞬时转染和细胞培养:nbfl细胞如前所述进行生长120。将0.5μmnfbl细胞接种在35mm培养皿上,在37℃培养过夜,并按照制造商的建议使用lipofectamine2000(lifetechnologies)转染2grest(seqidno:1)-ires-gfpcdna2。培养48小时后,通过荧光激活细胞分选(facs)对细胞进行分选.[0206]mrna提取和定量:如实施例10所述,从gfp 和–细胞中提取mrna;kv4.3的mrna水平采用实时rt-pcr检测116。使用β-肌动蛋白(actb)作为内参管家对照对基因表达归一化。使用的β-肌动蛋白引物与实施例10相同。kv4.3条引物为:正向:ctcactaccacctgctgctc,反向:tcagtccgtcgtctgctttc。[0207]图9:材料和方法[0208]质粒:rpp(seqidno:2和4):含有t7启动子、cpp和his标签的该构建物的一段通过idt合成为gblock(表8),并用表8所述的引物扩增。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸30秒。扩增后,根据制造商的方案,使用qiagen凝胶条带纯化试剂盒对pcr片段进行凝胶纯化。[0209]使用引入sbfi和xhoi限制位点的引物:正向:p12和p14(表1)扩增pet29质粒。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下进行次变性15秒,55℃退火15秒,30个循环,并在72℃下延伸6分钟。扩增后,用qiagen凝胶条带纯化试剂盒对pcr片段进行凝胶纯化,并用sbfi和xhoi限制性内切酶消化。根据制造商的建议,使用qiagen试剂盒纯化所有消化片段,并使用t4dna连接酶(neb)以适当的组合连接。连接片段在10g化学感受态细胞中转化并测序。[0210]rpp的核苷酸序列(表8)由hindiii和bamhi限制性内切酶组装和消化。该片段用cpp和his标签在上述pet载体中克隆。[0211]对照肽:将对照肽(cp)并入表9所示的gblock序列,然后克隆到pet载体中。cp序列如表9所示。[0212]体外表达:rpp(表8)或cp(表9)使用纯表达体外蛋白质合成试剂盒(newenglandbiolabs)在体外表达。使用表8中列出的引物从rpp(seqidno:2和4)或cp表达构建物(表8和9)中扩增rpp(seqidno:2和4)或cp序列,并使用qiagen凝胶条带纯化试剂盒进行凝胶纯化。pcr反应(20μl)最初在95℃下加热2.5分钟,然后在94℃下变性15秒,55℃退火15秒,30个循环,并在72℃下延伸30秒。然后,将15μl纯化的pcr片段与10μl纯表达体外蛋白质合成试剂盒(newenglandbiolabs)的溶液a和7.5μl溶液b混合,并在37℃下孵育2小时。接下来,添加2μldna酶i,并在37℃下孵育20分钟。[0213]表8-rpp-cpp-his_pet盒[0214][0215]agagctatgagctgcctgac,r-ggatgccacaggactcca;kv4.3:f-agctgtgcctcagaactaggcttt,r-taccagaaagacgcagggatgctt;kv7.2f-ccggcagaactcagaagaag,r-tttgaggccaggggtaagat;oprm1:f-ttcctggtcatgtatgtgattgta,r-gggcagtgtactggtcgctaa。[0227]图17:材料和方法[0228]细胞毒性检测:用ldh-glotm细胞毒性试验(promega),根据制造商的说明评估rpp(seqidno:12)的细胞毒性。用rpp处理48小时后收集培养基,或用2%tritonx-100孵育15分钟(可诱导坏死)。将培养基在ldh储存缓冲液(200mmtris-hcl(ph7.3)、10%甘油、1%bsa)中稀释1:50。为了验证检测的线性范围,还将ldh滴定曲线与实验样品一起运行。将50ml稀释培养基或ldh系列稀释液与50μlldh检测试剂(50μlldh检测酶混合物,0.25μl还原酶底物)孵育40分钟。使用infinitem200pro(tecan)仪器记录发光。[0229]实施例11–融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可用于诱导神经分化[0230]方法1:为了确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否最有效地促进人类神经发生和分化,将使用来自ncrm-1/xcl-1ips细胞系的神经干祖细胞(nspc)进行体外筛选检测。通过外体质粒重编程从cd34 人脐血细胞中获得ncrm-1/xcl-1ips细胞121,并通过类胚体(eb)方法分化为神经干祖细胞(nspc)122。ncrm-1/xcl-1神经干细胞是位置性原始神经干细胞,可快速分化为神经元123,使其成为基础广泛的毒理学和表型筛选平台的理想选择124-126。[0231]我们将使用两个工程化ncrm-1/xcl-1系在96孔板检测中进行高通量筛选。为了确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否具有细胞毒性,我们将使用在cmv启动子控制下组成性表达荧光素酶的ncrm-1/xcl-1系。在该系中,通过转录激活物样效应器核酸酶(talens),将cmv纳米荧光素酶halotag(cmv-nlht)构建物插入染色体19q的安全港aavs1位点(图20a)。我们将使用荧光素酶活性作为cmv-nlht细胞数的快速替代读数。使用这种方法,我们将收集时间过程数据,以测定长期暴露于药物的毒性。如果融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽表明毒性水平导致荧光素酶信号丢失≥10%,则这就会通过celltoxtmgreen毒性检测来确认。[0232]为了评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的促神经元分化作用,我们将使用第二个工程化ncrm-1/xcl-1系。在这个细胞系中,使用锌指核酸酶(zfn)将纳米荧光素酶halotag(nlht)敲入map2转录起始位点(tss)(2号染色体)(图20b)。在ncrm-1/xcl-1培养物中,map2表达随神经元分化而增加127,这是通过增加map2-nhlt培养基中的荧光素酶活性来复制的(图20c)。我们将使用培养基中的荧光素酶活性作为map2-nlht神经分化的替代读数,并收集时间过程数据,实时跟踪融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的分化效应。这种方法将允许我们快速比较用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽或对照肽处理的培养物中的神经分化率。将通过对两个同期组的细胞命运标记基因121;一组同期组的nspc和lin28a、rps27l、ifitm2、igfbp3、anxa1的多能标记基因;神经元标记基因c1orf61、iglon5、igsf11、chl1、sox9的qrt-pcr表达筛选,独立验证阳性神经分化效应。荧光素酶活性和qrt-pcr值将报告为平均值±平均值标准误差(sem)。通过单因素方差分析(anova)和tukey的多重比较检验,确定药物对神经元分化率的统计学显著影响。[0233]预测结果和备选策略:我们预测,用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对人nspc给药后,将加速神经元分化,如通过map2荧光素酶活性增加的测定所示的,与溶剂对照组相比,nspc/多能标记物的表达减少,神经元标记物的表达增加。[0234]如果融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽影响转变了ncrm-1/xcls标记时空表达谱,则可能需要在备用时间点收集数据。如果荧光素酶筛选缺乏评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的促分化作用的敏感性,则qrt-pcr数据可用于评估神经分化。[0235]材料和方法[0236]质粒:cmv纳米荧光素酶halotag(cmv-nlht)可从promaga购买。[0237]细胞系和细胞培养:如前所述通过外体质粒重编程从cd34 人脐血细胞中获得ncrm-1/xcl-1ips细胞121,并通过类胚体(eb)方法122分化为神经干祖细胞(nspc)和神经123。通过如papapetrou等人,2016先前描述的转录激活物样效应器核酸酶(talens)128将cmv纳米荧光素酶halotag(cmv-nlht)构建物插入染色体19q的安全港aavs1位点(ncrm-1/xcl-1细胞系)。使用锌指核酸酶(zfn)将纳米荧光素酶halotag(nlht)敲入ncrm-1/xcl-1细胞系的map2转录起始位点(tss)(2号染色体)的方法之前已经描述过127。[0238]荧光素酶检测:测定cmv-nlht细胞数量和神经分化(map2-nlht)的方法之前已由fritz等人,2017描述129。[0239]qrt-pcr筛选:用于筛选细胞命运标记基因的qrt-pcr方法已由chou等人,2011描述过121。[0240]方法2:rpp给药7天后诱导的多能干细胞(神经干细胞(nsc)-nl5)分化(图11(seqidno:5-11,3μm);图12(seqidno:9,1μm)图13(seqidno:13(图27),1μm),使用以下“神经元至空白(neurontoblank)”方案:[0241]1、24孔培养板(黑色visiplate)包被有matrigel(12ml/孔,37℃下30分钟)。[0242]2、nscs-nls(见下文培养基)按每孔0.5×105进行铺板。[0243]组分最终浓度量dmem/12培养基1x97mln2(100x)1x1mlb27添加2%2mlbfgf(制备为100μg/ml母液)10ng/ml10μl[0244]3、24小时后(细胞应》70%汇合),将培养基更换为神经元分化培养基(下文),添加不同浓度的测试肽或水对照(图11、12和13、表14)。[0245]每2天更换一次含有新测试肽和对照的神经元分化培养基,持续5天。[0246]组分最终浓度量d-mem/f-121x88.6mlglutamaxtm-i添加2mm1ml牛血清白蛋白(25%)1.8%7.2mlhesc添加2%2mlbdnf(制备为25ug/ml母液)10ng/ml40μlgdnf(制备微100ug/ml母液)10ng/ml10μl[0247]4、第7天,将细胞用1×pbs中的4%甲醛固定30分钟。[0248]5、用小鼠抗tuj1(1:1000)和兔抗map2(1:500)免疫标记细胞过夜。然后用二抗(抗鼠488、抗兔568和dapi)标记1小时。[0249]6、在biotek平板阅读器上收集数据[0250]a、dapi:激发360/40;发射460/20,增益35;[0251]b、alexa488:激发485/20;发射528/20,增益50;[0252]c、alexa568:激发560/20;发射620/10,增益75)。[0253]结果:在图11中,通过将map2(成熟神经元标记物)表达归一化为dapi(细胞核)并与对照组相比,在7天后,3μm的rppseqidno:5至11增加(1.7至2.7倍)了nsc-nl5的分化。在图12中,根据tuj1和map2神经元标记物(归一化为dapi(细胞核))测定,与对照组相比,1μm的rppseqidno:9在7天后使nsc-nl5分化增加约35%。在图13中,通过tuj1(分别增加60%和3倍)和map2(分别增加27%和2倍)神经元标记物(归一化为dapi(细胞核)并相对于对照组),7天后0.1和1μm的rppseqidno:13增加了nsc-nl5的分化。使用该筛选的基础是,神经祖细胞中rest的消除已被证明可诱导神经元分化22,是神经元基因整体去表达的读数。[0254]实施例12-融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可用于改善创伤性脑损伤后的运动和认知功能。[0255]方法:我们将确定在tbi大鼠模型中,用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽治疗是否会改变神经元存活率和神经发生量。我们将这些指标与组织病理学和功能恢复结果相关联,以确定tbi后rest肽的有效性。我们已经表明,损伤后病变周围的restmrna增加(图19)。我们将使用具有良好特征的受控皮质撞击损伤(cci)大鼠模型6,130-134。[0256]雄性和雌性sprague-dawley大鼠(约250-300g雄性,150-200g雌性)将被cci单侧损伤右侧顶叶皮质(直径5mm,速度4m/s,深度2mm)。假手术动物将进行相同的程序,不会产生影响。cci后6小时,大鼠将在其右心室植入一个套管,该套管将连接至渗透微型泵,用于脑室内导管(icv)输送对照或融合至cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽(0.17μg/h)。六小时是一个具有转化相关性的现实初始治疗窗口,用于确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的疗效,直至处死。我们将在两个不同的时间点处死大鼠–第一组在3dpi时,检查融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对神经元死亡和增殖的急性影响,第二组在30dpi时测定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对功能和形态恢复及神经发生的影响。对于每个同期组,将有12组肽3(对照肽、两个剂量的融合到cpp(seqidno:118至137和159至143)的rpp(seqidno:1和15至17))或rppv(seqidno:18至117))×2(假手术组,cci)×2(雄性/雌性)设计中。雄性和雌性大鼠将在不同的天数进行跑步,以确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的在创伤、行为或反应中的任何性别特异性差异。大鼠在第一周每天腹腔注射brdu(50mg/kg),以确定增殖和成熟细胞的命运(第一同期组-每天注射1-3dpi,最后注射后2小时处死;第二同期组-每天1-7dpi)。我们之前的经验表明,我们需要16只大鼠/组获得重要的行为数据(第二同期组),8只大鼠/组获得免疫组化(第一同期组)。[0257]在第二个同期组中,为了确定与对照组相比,用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽治疗的功能结果,我们将检查cci或假损伤后不同时间点的运动和认知行为。我们将在大鼠受伤前对其进行训练,以在平衡木行走(beamwalk)、滚轴(rotorod)和旷场试验中获得运动功能的基线读数。大鼠将在伤后1、3、7和10天(dayspost-injury,dpi)在平衡木行走和滚轴上测试运动功能的恢复情况。旷场试验将在的第4、12和22dpi进行。识别记忆将通过在20dpi下进行的新型物体识别测试进行评估。数据将通过重复测定双因素方差分析和dunnets事后校正进行分析。空间记忆和学习将由morris水迷宫(mwm)检测,在25dpi开始。所有试验都将记录游泳速度和找到隐藏平台的时延。在训练的第五天进行的探测试验将确定大鼠在之前包含隐藏平台的象限中所花费的时间。还将对每只大鼠进行可见平台测试,以确保动物之间的视力差异很小。带有tukey多重比较校正的双因素方差分析将确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否与对照肽相比显著改变假手术或受伤大鼠mwm中的认知功能135。[0258]功能评估完成后,经心灌注处死8只大鼠/组,取脑进行处理。我们将通过甲酚紫染色切片,并测定以500μm为间隔的病灶的12个切片,来确定3和30dpi时病灶的大小。病灶体积将表示为同侧半球体积的百分比。将检查穿过额顶皮质和背侧海马(包括脑室下区(svz)和齿状回(dg))的一系列冠状脑切片。将使用体视学对不同海马区域和svz中的brdu阳性细胞总数(用抗brdu检测)进行计数,以确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否会改变损伤后新增殖细胞的存活率。切片将与不同的细胞特异性标记物共同染色,以确定3dpi时增殖细胞的特性及其在30dpi时的成熟命运。细胞特异性标记物包括neun(成熟神经元)、sox2(神经干细胞)、双皮质素(doublecortin,dcx)(神经元祖细胞)、gfap或aldh1l1(星形胶质细胞)、iba1(小胶质细胞)、ng2(少突胶质前体细胞,opc)和apc或gstpi(成熟少突胶质细胞)。将比较不同处理组之间在30dpi时dg和嗅球中brdu/neun双阳性细胞的数量,以确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否增加损伤后神经发生。我们还将确定是否存在brdu 神经元祖细胞或成熟神经元向海马不同区域的迁移-远离dg,或从svz向病灶区或吻侧迁移流(rms)。除神经源性凹部(nich)外,检查病灶周围区域内每种brdu 细胞类型的总数,将确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否会改变损伤后的病理学。由于我们预计融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽将增加神经元存活率,因此我们将量化两个时间点病变周围区域中退化神经元(flurojadec)的数量和存活神经元(neun)的数量。我们将用luxolfastblue检查胼胝体,以显示髓鞘数量的任何差异。无偏光学分馏器法将用于对神经发生区、病变周围区、吻侧迁移流和嗅球的细胞进行计数,以获得准确的细胞特异性和增殖细胞计数。所有数据将通过dunnetts事后校正的双因素方差分析进行分析。将通过flag抗血清染色,在切片中检查融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的分布。每组剩下的8只大鼠(第二同期组)将被处死,并迅速移除它们的大脑。病灶同侧和对侧的大脑区域将被打孔并快速冷冻以分离rna和蛋白质。我们将通过蛋白免疫印迹检测rest水平,并通过qrt-pcr检测病灶周围或神经源性凹部中bdnf的表达,与对照组大鼠大脑进行比较。[0259]预测结果和备选策略:我们预计融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽将通过提高神经元存活率来减少3dpi和30dpicci后的病灶体积。将在两个时间点检查退化和存活的神经元,以直接评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽处理对神经元存活的影响。融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽在病灶周围区域早期神经保护作用可能会在第一周改善运动功能。虽然我们之前已经发现小鼠cci后3dpi时神经元死亡最多136,我们可能需要检查大鼠体内的不同时间点。我们还预计,与对照肽相比,用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽治疗的动物将显示出更多的神经发生。我们的数据将表明,在3dpi时,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否会导致不同脑区dcx 前体的增加,或在30dpi时海马、嗅球或病变周围的brdu /neun 成熟神经元的增加。我们将能够将这些数据与行为结果相关联,以观察使用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的治疗是否会提高mwm检测或新物体识别检测的表现135。通过这种方式,我们将能够确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否对炎症或细胞增殖有任何负面影响。[0260]材料和方法[0261]测试系统[0262]物种:雄性和雌性sprague-dawley大鼠。[0263]研究开始时的年龄:约为22至29周。[0264]研究开始时体重:雄性约250至300克,雌性约150至200克。[0265]研究的设计:对于每个同期组(1和2),将有12组3(对照肽、两个剂量的融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽)×2(假手术组,cci)×2(雄性/雌性)设计中。在同期组1(免疫组化)中,将有8只大鼠/组,在同期组2(行为)中,将有16只大鼠/组。[0266]合理剂量水平:高剂量将是最大可行剂量(约0.17g/h),低剂量将是基于脑脊髓液(csf)药物水平的估计有效剂量,其近似于实施例13中确定的有效剂量。[0267]施用途径:植入右心室的套管与渗透微型泵相连,用于脑室内导管(icv)输送。[0268]施用频率:持续输液直至处死。[0269]给药周期:同期组1:3dpi;同期组2:30dpi。[0270]环境条件:2只大鼠/笼,随意进食饮水。[0271]供试品标识:融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽[0272]纯度:≥95%[0273]剂量配方的制备:每4天一批(bulk)。[0274]剂量配方检测和稳定性:将在研究第一周的第1天和第7天评估剂量配方的稳定性和浓度。可接受的浓度范围为标称浓度的±10%。[0275]受控皮质撞击(cci)模型:cci模型之前已被描述6,130-134。融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽或对照(仅溶剂)的施用将在cci后6小时开始。[0276]免疫组化:xiong,y等人,2007和2008对这些方法进行了描述135-137。[0277]morris水迷宫:choi等人,2006之前描述了这些方法138。[0278]实施例13-融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可用于改善周围神经损伤后的再生。[0279]方法:周围神经损伤(pni)后的功能恢复需要营养因子(ntf),它们的存在或缺乏是再生反应强度的生物标志物139,140。已知几种ntf基因,包括脑源性神经营养因子(bdnf)、神经生长因子(ngf)、pleitrophin(ptn)和神经营养因子-3(ntf-3),被损伤诱导的rest表达所抑制,我们已经证明rpp(seqidno:2)可以逆转这种转录抑制,至少在bdnf和ngf的情况下是如此(图7、9、10和16)141-142。[0280]1)实时定量逆转录pcr(qrt-pcr)分析表明,nbfl细胞注射1μm线性(seqidno:4)或环化(seqidno:2)rpp16或48小时后,引起bdnf和ngfmrna表达增加(图9a和b及下表)。[0281][0282]2)48小时后,与水对照相比,rpp(seqidno:9,3μm)使ntfsbdnf和ngf分别增加约2倍和3倍。这一发现证实了rpp在mpc中是活跃的(图10)。如gerevasi等人,2020所述,将mpc在通用培养基中培养76。[0283]3)我们将对融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对从雄性和雌性肌肉骨骼创伤患者分离的间充质祖细胞(mpc)中ngf、bdnf、ptn和ntf-3表达的影响进行量化。mpc存在于周围神经损伤部位,被广泛表征143。mpc将使用leonnesti博士制定的标准分离方案,在正常和预先计划的手术处理过程中采集116,143。来自3个不同受试者的mpc将通过培养进行扩增。首先,我们将在无神经元诱导的情况下传代细胞四次,以通过qrt-pcr和蛋白免疫印迹(wb)建立rest和ntf转录物和蛋白质水平表达的基线。接下来,我们将重复bulkenhoover等人所描述的神经元诱导扩张144。简而言之,mpc在预诱导培养基中培养2天,然后添加全反式维甲酸(ra)1天,然后在神经诱导培养基中培养7天。神经诱导培养基每三天更换一次。诱导后第7天,将评估rest和ntf水平。我们预计,神经元诱导应导致rest表达减少,ntfs表达相应增加。[0284]评估融合到cpp(seqidno:118至137和159至143)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽诱导mpc中ntf表达的效力,我们首先需要通过评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的细胞毒性来确定剂量。简言之,mpc将以4个剂量水平(最高10μm)在用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽替代ra的改良预诱导培养基中或在预诱导培养基(含ra)中进行铺板1天。每个给药组将有三个技术和三个生物重复,并将使用mtt检测来测定细胞毒性145。为了确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的最佳浓度,以降低rest水平和增加ntf水平,我们将在处理后1、3和6天(dayspost-treatment,dpt)用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽替代ra的神经诱导培养基中进行剂量反应(3个剂量水平)。融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的剂量将基于mtt分析的结果145。我们的阴性和阳性对照将分别为神经诱导培养基或 ra。我们将用qrt-pcr和wb对rest和ntfs的基因表达和蛋白水平进行定量139,146。将在1、4和7dpt收获细胞,并从细胞裂解物中分离rna和蛋白质。将进行双因素anova分析,然后进行tukey的事后检验,以在指定时间点比较–和 对照组与融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽给药组。总体平均值将报告为平均值±标准差,p值为0.05或更小,具有统计显著性。将使用sas统计软件包(sasinstitute,carync)进行统计分析。[0285]4)在整个drg神经元(l5,来自雄性sd大鼠)的离体培养物中,在0、0.3、1、3、10μm处评估rpp(seqidno:12)诱导bdnf和ngf表达的潜力(图16)。rpp在0.3、1和3μm时增加bdnf表达约25%,在10m时增加75%。rpp在3和10μm时分别增加ngf约50%和2倍(图15)。在1μm处未观察到任何影响,0.3μm因n=1而不确定(图16)。[0286]5)在体外培养的整个drg神经元(l5,来自雄性sd大鼠)上进行的ldh细胞毒性检测中评估rpp(seqidno:12)。rpp在0、1、3或10μm时没有显示毒性(图17)。正如预期的那样,阳性对照组(2%triton)具有神经毒性,增加了90000rlu。[0287]6)我们将评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽通过分别使用脊髓和分离的背根神经节(drg)外植体,在运动和感觉离体神经模型中诱导ntf表达并刺激再生的能力147。这两种模型广泛用于研究生长因子的神经保护和营养特性148。将从脊柱两侧(l4-l6)提取(6只大鼠)一段10mm的完整drg并进行培养(图21和22)149,150。剩余的脊柱材料将被保存下来,用于运动神经元连接的脊髓切片培养。这些脊柱将用切片机以300μm的间隔横向切片,切片将被转移到带有半透膜的培养插入物中,并在培养条件下适应一周149,150。插入一周龄的器官型脊髓切片的培养物(具有稳定的存活运动神经元群),之后将被转移到6孔板中再培养7天。[0288]两种离体外植体培养物将在neurobasal/b27培养基中保持2周,以允许神经元过程的广泛延长147。为了诱发物理损伤,将使用玻璃巴斯德吸管在距离drg神经节周长6mm处和脊髓腹侧切片的灰白质交界处的精细神经元突起上划伤。体外培养物将在损伤后24小时开始用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽(0、1、3或10μm)处理。[0289]受损神经突(运动和感觉)的再生能力将通过监测细胞活力和测定轴突生长来确定。通过分别用钙黄绿素am和溴乙啡锭二聚体1标记活细胞和死细胞来评估7dpt时的细胞活力。神经突延长是神经元再生的一个指标,将在1、4和7dpt时进行测定。将使用与单色数码相机连接的zeissaxioobserver显微镜,获取划痕区域的灰度显微照片151。将使用imagej分析划痕区域内再生芽的数量、长度和总面积152。[0290]ngf通过其受体,即drg中的trka受体发挥其大部分功能活性153,ptn主要在运动神经元中引起轴突生长增加150-154。bdnf和ntf-3存在于运动神经元和感觉神经元中。我们将用qrt-pcr和wb对ntf的基因表达和蛋白水平进行定量139,146。在1、4和7dpi采集每个离体系统的组织,并分离rna和蛋白质裂解物。将进行双因素anova分析,然后进行tukey的事后检验,以在指定时间点比较对照组和融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的给药组。总体平均值将报告为平均值±标准差,p值为0.05或更小,具有统计显著性。将使用sas软件(sasinstitute,carync)进行统计分析。[0291]7)将在100只大鼠体内坐骨神经缺损后,评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽改善再生和功能恢复的能力。我们已经证明,在坐骨神经损伤部位注射rpp48小时后,rpp在脊髓坐骨神经神经元的细胞核中积聚(图4)。将通过静脉(iv)施用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,这将允许我们测试比肌肉内(im)或皮下(sc)施用更高的最大可行剂量(mfd)。静脉注射也可避免与口服和腹腔注射(ip)相关的首过代谢。再生和功能恢复研究的剂量浓度将基于sprague-dawley大鼠一个月单次iv剂量范围发现毒性和毒代动力学研究的结果。将在完成1)后启动剂量范围判定的毒性研究。简而言之,该研究将使用四种剂量水平,最高1000mg/kg或最大耐受剂量或mfd,如fda指南(m3(r2),2009)所建议。每组分配的动物数量如下:主要研究:10只/性别/组;tk:3只/性别/对照组和9只/性别/给药组。将评估动物的体重、食物消耗、临床化学、血液学、尿液分析、器官重量、组织病理学(8个核心组织)和毒代动力学。[0292]在再生和功能恢复研究中,spraguedawley大鼠将每周使用长期静脉导管155,通过静脉注射融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,3个给药组(tbd) 对照组(10个/性别/组)治疗6周。所有动物在暴露坐骨神经前都将被麻醉156,在神经残端回缩后,移除0.7cm的部分,以形成约1cm的节段性缺损157。神经将使用脱细胞神经移植修复。然后闭合手术切口,让神经再生6周。只有给药组施用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽。在6周期间,我们将评估功能恢复和对融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的剂量耐受性。每周对大鼠称重,记录温度,并进行脚趾伸展反射检测(以确定受伤腿部的最大足迹宽度,测试运动神经功能)156。步态分析(digigait)将每2周进行一次,并进行电生理研究158。简而言之,将对大鼠坐骨神经进行电生理测试。采用单脉冲电击(1ma,0.1ms)对移植物缝合点附近5mm处的本地坐骨神经干施加电刺激。记录腓肠肌腹部的复合肌肉动作电位(cmap),从1v到12v,或直到达到超最大cmap。还记录了未手术对侧坐骨神经的正常cmap进行比较。恢复率将由受伤后肢的cmap与对侧正常后肢的cmap的比率决定159,160。[0293]在第6周结束时,将收集坐骨神经、脊髓腹角(vh)以及与l4至l6神经根相关的drg161,162。将对神经再生进行组织学评估。将移植物的近端、远端和中心进行准备以使透射电子显微镜(tem)通过横截面,评估移植物沿线的髓鞘形成163,164。简而言之,将使用超薄切片机从神经上切下约70nm的超薄切片,然后用醋酸铀酰和柠檬酸铅染色。将从随机切片中选择10-15个场进行分析,量化有髓纤维的数量和大小。平均纤维密度的计算方法如之前所述73。除tem外,还将准备更多来自同一神经的切片,用于ihc可视化β-微管蛋白iii以测定轴突的直接生长,用s100以测定雪旺细胞对移植物的渗透和生长,用血管性血友病因子以评估愈合神经的血管生成和毛细血管浸润,以及用luxolblue染色165,166测定移植物内髓鞘的形成。将评估受损运动神经和感觉神经细胞体内腹角和drg与融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽相关变化。将使用光学显微镜拍摄染色组织的图像,并使用imagej软件对解剖匹配组织中阳性反应的强度和面积比例进行量化(https://imagej.nih.gov/ij),比较各组之间的神经再生率165。将如之前所述,对受损神经的drg神经元进行计数,并与对侧未受损drg进行比较167。ntfs(bdnf、ngf、ptn和ntf-3)、ctdsp1和rest将通过qpcr和wb进行检查。[0294]预测结果和替代策略:我们预计,与神经诱导培养基ra相比,经融合至cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽 神经诱导培养基(-维甲酸,ra)处理的间充质祖细胞(mpc)将增加神经营养因子(ntf)的表达并降低rest水平,但与神经诱导培养基 ra类似。我们预计,与未治疗组相比,经rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽融合到cpp(seqidno:118至137和140至159)治疗的离体模型,由于rest水平降低,将显示神经突生长增加。在缺乏支持细胞的情况下,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的再生效应可能不会显著,支持细胞可在神经元损伤后在体内分泌神经营养因子。另一种实验将包括在受伤之前以及随后的融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的治疗之前,神经支持细胞(如雪旺细胞和间充质干细胞)与背根神经节(drg)和脊髓外植体(富含运动和感觉神经元)的共培养。接下来,我们将分析融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽帮助大鼠模型中切断的神经再生的能力。我们预计,与对照组大鼠相比,经组织学、电生理学和功能评估确定,经融合至cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽治疗的大鼠的恢复情况将有所改善。我们还期望融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽增加ntf表达并降低rest蛋白水平。如果我们没有观察到rest阻断恢复和解除的迹象,我们将通过直接注射到靶点(即移植物部位或鞘内),增加融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的局部浓度。[0295]材料和方法[0296]细胞培养:mpc的获取、培养和神经营养诱导之前已有描述168。简而言之,mpc在预诱导培养基中培养2天,然后添加全反式维甲酸(ra)1天,然后在神经诱导培养基中培养7天。神经诱导培养基每三天更换一次。入职后第7天,将评估rest和ntf水平。[0297]qrt-pcr:示例10中描述的方法。[0298]蛋白质印迹分析:按照ballas等人2001年119的程序制备全细胞裂解物。使用抗rest-c的标准程序进行蛋白质印迹64,抗ntf(使用市售抗体)、抗gapdh(abcam)和与红外染料结合的抗igg(thermofisher),并在奥德赛红外荧光成像仪(licor)上进行分析。[0299]供试品标识:rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽融合至cpp(seqidno:118至137和140至159),纯度≥95%[0300]剂量配方的制备:每4天一批。[0301]剂量配方和稳定性:将在研究第一周的第1天和第7天评估剂量配方的稳定性和浓度。可接受的浓度范围为标称浓度的±10%。[0302]脊椎动物:雄性和雌性成年sprague-dawley大鼠(约250-300g雄性,150-200g雌性)将用于确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽在改善pni恢复方面的功效。这项研究将需要100只8-11周大的大鼠。大鼠将从charlesriverlaboratories获得。[0303]步骤说明:[0304]手术:在所有手术之前,用异氟烷(4%诱导,3%维持)麻醉大鼠。在2)器官型脊髓培养物将使用前述技术从出生后第8天大鼠(sprague-dawley)的腰脊髓制备(rothstein等人,1993;corse等人,1999)。简而言之,将迅速处死大鼠,移除腰骶脊髓,并将其置于含有葡萄糖(6.4mg/l)的gey平衡盐溶液(gibco)中。在使用无菌技术的层流罩中,在放大镜下仔细切除脑膜,并切断腰脊神经根。将脊髓(cord)放置在aclar膜上,并用mcilwain组织切割器从l2到l5以300μm的间隔进行切片。将使用gey的平衡盐/葡萄糖溶液,小心地将单个脊髓切片转移到6孔培养板中的millicellcm(微孔)渗透膜上。将在每个膜上放置五段线。每个孔含有1ml培养基,由50%最低基本培养基加25mmhepes组成;含d-葡萄糖的25%hanks平衡盐溶液(25.6mg/l);25%热灭活马血清;和2mml-谷氨酰胺。培养物将在37℃、5%co2的加湿培养箱中保持一周,每3天更换一次培养基。一周后,切片将被转移到另一个6孔培养板上,并在任何治疗之前在培养中稳定和延长神经突起7天。[0305]在3)中我们将暴露坐骨神经117(图21和22)并移除0.7cm的部分,以在神经残端收缩后形成约1cm的节段性缺损(hems和glasby,1993)。神经将使用脱细胞神经移植修复。然后闭合手术切口,让神经再生6周。第一组(对照组)将进行约1cm的坐骨神经切除,并将移植1cm脱细胞同种异体神经。第二组至第五组将接受1cm坐骨神经切除术,移植1cm脱细胞同种异体移植物,每周静脉注射融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,药物浓度由剂量范围判定确定。所有动物将在第6周处死。[0306]行为评估:我们将对动物进行长达6周的监测,以评估功能恢复和对肽治疗的耐受性。每周对大鼠称重,记录温度,并进行脚趾伸展反射检测(以确定受伤腿部测试运动神经功能的最大足迹宽度)117。步态分析将每2周进行一次。6周处死前进行电生理检查。[0307]电生理学评估:电生理学测试将按照之前的方法进行158。简言之,重新暴露大鼠坐骨神经,并在移植物缝合点附近5mm处对天然坐骨神经干施加电刺激(单脉冲电击,1ma,0.1ms)。在腓肠肌腹部记录1v至12v的cmap,或直到达到超最大cmap。还记录了未手术对侧坐骨神经的正常cmap进行比较。grasstechs88x刺激器(astromedinc.)用于测试,polyviwe16数据采集软件(astromed,inc.)用于记录。恢复率是大鼠受伤后肢的cmap与对侧正常后肢的cmap之比158。[0308]处死:大鼠于伤后6周处死。处死后,我们将用组织学方法评估轴突通过移植物的生长、再髓鞘化、损伤部位的mpc细胞活性、前角细胞和drg活性以及再生神经的血管化。我们将在每只大鼠处死后采集腓肠肌和胫骨前肌,以评估总重量来量化萎缩。[0309]合理解释:pni后,脉管系统、免疫系统和周围神经系统内所有细胞和分子之间存在复杂的相互作用。我们不完全了解创伤性损伤后不同细胞之间、信号分子、细胞-细胞相互作用和回路的顺序或相互作用,因此目前无法在体外对其进行建模。此外,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可能会影响神经前体细胞、成年神经元以及其他细胞类型,因此在pni的情况下对其进行体内评估至关重要。fda还要求在允许生物制剂的ind状态之前提供体内疗效证明,因此这为这些实验的体内性质提供了进一步的理由。之所以使用大鼠,是因为在pni表明其在这些类型的实验中的可靠性后,进行了大量的行为研究。因此,有大量文献为实验的设计和执行提供了详细的框架。大鼠比小鼠更聪明,体型也更大,能够更好地辨别受伤后功能恢复的变化。与小鼠相比,大鼠生理学与人类有更多的相似之处,从而使实验数据具有更好的转化相关性。此外,我们针对的rest通路在大鼠中是保守的。[0310]最大限度地减少疼痛和痛苦:用异氟醚(3-4%)麻醉大鼠,以治疗节段性周围神经缺损。手术后,动物将被允许在加热垫上恢复,直到移动。手术后,将给大鼠服用对乙酰氨基酚(饮用水中6毫克/毫升),持续2天,此外,如果观察到疼痛或痛苦的迹象。实验室和兽医人员每天都会对老鼠进行观察,并根据兽医的建议对被认为处于困境的动物进行治疗或安乐死。[0311]安乐死:使用的安乐死符合美国兽医协会动物安乐死指南的建议。[0312]实施例14-融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可用于预防和治疗慢性疼痛。[0313]完成的评估:[0314]神经损伤后rest的激活导致感觉神经元正常兴奋性所需的几个基因表达减少,包括钾通道kv4.3(kcnd3)和kv7.2(kcnq2),钠通道nav1.8(scn10a)和μ阿片受体oprm160,72-74。阻断rest以缓解慢性疼痛的基础是利用小鼠和大鼠周围神经损伤(pni)模型发表的公开研究60,72-74。[0315]1)实时定量逆转录pcr(qrt-pcr)分析表明,nbfl细胞注射1μm线性(seqidno:4)或环化(seqidno:2)rpp16或48小时后,会引起kv4.3mrna表达增加(下图9c和表16)。[0316][0317]4)rpp(seqidno:12;0(水)、1、3或10μm)在整个drg(l5,来自雄性sd大鼠)的体外培养物中孵育48小时,并评估其诱导慢性疼痛相关基因kv4.3、kv7.2、nav1.8和oprm1表达的潜力(图14)。在10μm时,与对照组相比,rpp增加kv4.3表达2倍;在3μm和10μm,分别增加kv7.2表达7.5倍和9倍;在3μm和10μm,分别增加nav1.8的表达2.9和4.8倍;在3μm和10μm,分别增加了oprm1的表达(图14)。在整个drg(l5,来自雄性sd大鼠)的离体培养物中,在3μm的48小时给药期后评估rppseqidno:13和14,并评估nav1.8。序列13和14增加nav1.8表达分别为对照组的6倍和13倍(图15)。[0318]5)在体外培养的整个drg神经元(l5,来自雄性sd大鼠)上进行的ldh检测中,rpp(seqidno:12)没有神经毒性。与对照组相比,rpp并没有增加rlu水平。正如预期的那样,阳性对照组(2%)的rlu增加了90000倍(图17)。[0319]计划评估:[0320]目标1:确定融合至cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽药代动力学、分布、以及剂量[0321]子目标1.1:用于检测大鼠和猴血浆、脑脊液和组织中融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽(范围在10至10000ng/ml之间)的液相色谱-串联质谱(lc-ms/ms)方法的开发和验证。[0322]子目标1.2:为了确定体内动物研究和临床中的施用途径,将在sd大鼠中进行探索性药代动力学(pk)研究。在静脉注射、sc注射或po注射1000mg/kgrpp(seqidno:1和15至17)或rppv(seqidno:18至117)后,在5分钟、30分钟、4小时、8小时、24小时和48小时(2个性别/时间点)采集血浆样本,并评估峰值药物浓度(cmax)、曲线下面积(auc)和半衰期(t1/2)。为了评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对中枢和外周神经系统的渗透性,还将在安乐死时收集靶组织(csf、脑、腰脊髓和背根神经节(drg)),并测定药物浓度。将使用经验证的ls-ms/ms方法(子目标1.1)检测融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽。[0323]子目标1.3:为了确定大鼠疗效研究(目标2)中的剂量,将使用子目标1.1中确定的最佳施用途径对sd大鼠进行单剂量范围判定。根据子目标1.1中的pk数据,将选择四个剂量水平 溶剂对照(3个/性别/组)。高剂量(hd)应确定极限剂量(最大可行剂量(mfd))、最大耐受剂量(mtd)和/或暴露饱和)。较低剂量将以1/3的增量间隔。将每天评估动物的存活率和临床症状(异常和疼痛或窘迫症状)。将在给药后的6个时间点采集血液,这些时间点是根据子目标1.1的pk数据选择的,并评估cmax、auc和t1/2。将在安乐死时收集将在疗效研究中评估基因变化的组织(drg、腰脊髓、大脑),并评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的浓度。[0324]目标2:评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对慢性疼痛的影响。[0325]我们已经证明,在坐骨神经切断部位给药48小时后,rpp在大鼠腰椎(l4–l6)脊髓运动神经元的细胞核中累积(图4)。为了评估我们的药物对疼痛的影响,我们将使用坐骨神经的临床前sni模型。sni是一种公认的神经病理性疼痛动物模型,可产生强烈和持久的热敏感性变化、外周和中枢吗啡镇痛以及c纤维感觉减退60,72,74。此外,正如我们预测的那样,我们观察到ctdsp1水平增加(图18)。[0326]融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽减少慢性疼痛的功效将在第1阶段完成,疼痛减轻的持久性将在第2阶段确定。sni坐骨神经大鼠模型将用于测试融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否能减少慢性神经病理性疼痛。将施用融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,或用于治疗神经病理性疼痛的溶剂或标准护理药物羟考酮。下表17总结了第1阶段的实验步骤:[0327]表17[0328][0329][0330]为了实现这一目标,总共需要312只2个月大的sd大鼠(雄性和雌性数量相等)。每组12只大鼠的样本量将有80%的效力检测疼痛评估中标准差1.20的差异。效力分析基于独立样本的t检验,具有5%的双侧显著性水平。对于疗效研究,将大鼠随机分配到右侧表格中指定的实验组(n=12/组)。对10个随机分组的大鼠进行刺激诱发和非刺激诱发行为测试、形态学、免疫组化分析、定量rt-pcr(qrt-pcr)分析和wb分析。安乐死时,将在开始给药后的2个时间点(14天和28天)抽取血液,并评估标准pk参数。提取drg和大脑以确定oprm1、nav1.8,kv4.3和kv7.2水平。此外,将收集组织进行lc-ms/ms分析,以确定rpp浓度。[0331]手术:sni或假手术(暴露神经分支而不损伤神经的手术)将按照之前所述进行169。简而言之,将麻醉大鼠,并结扎腓总神经和胫总神经。将切断并移除结扎远端各神经的2-4mm段,保留坐骨神经的第三支腓肠神经完整。腓肠神经在损伤后4天开始对同侧神经支配区的伤害性和非伤害性刺激产生增强的反应,7天稳定,维持6个月。损伤后7天是慢性神经病理性疼痛发展的公认时间点,并且具有可转化相关性。[0332]药物/溶剂:手术后第5天开始施用。对于融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,测试的途径、频率和药物浓度将基于特定目标1中的pk研究。对于羟考酮,大鼠的给药途径和有效药物暴露(接近人类的有效暴露):(0.56mg/kg剂量的羟考酮和0.2ml溶剂)是已知的170。[0333]行为测试:将在手术前一天和从术后第4天开始每隔一天评估对疼痛传递和运动功能的影响。与羟考酮和溶剂相比,刺激诱发和非刺激诱发行为试验将用于确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的功能有效性。近年来,对一些刺激诱发行为测试转化性的关注导致刺激诱发、非刺激诱发和操作性或自愿性行为测试的使用越来越多。对于这一提议,我们将使用最广泛使用和可转化的刺激诱发行为测试:电子vonfrey和热痛觉过敏测试、非刺激诱发步态分析和条件性逃避/回避试验(placeescape/avoidanceparadigm)来测定疼痛的厌恶状态171。最近的研究表明,在动物模型中加入步态分析来检测细微的改善和恶化是有价值的172。进行行为测试的大鼠处理人员和评估人员将被随机分配给对应的实验/对照组。每只动物都有一个唯一的动物编号。将使用秘密颜色代码指定对照组和实验组。只有在分析之后,才会显示此代码。[0334]刺激诱发行为测试:对机械刺激的超敏反应:将使用一种电子vonfrey(bioseb,查维尔,法国),由一个配有塑料尖的手持式力传感器组成,用于测试左、右后爪的机械后退阈值。力传感器的塑料尖将应用于爪子的内侧足底表面以测试隐神经,并应用于爪子的外侧足底表面以测试坐骨神经(腓神经)。[0335]热痛觉过敏:将根据一种已公布的方法进行测定173,我们将该方法修改为使用激光作为热源。将使用808nm波长的激光器作为热源(2w输出功率,皮肤上的光束直径=3mm),并将其应用于被测神经的相关皮肤。小光束尺寸允许相关皮肤小体被专门定位。对于坐骨神经和隐神经,将以10秒为分界点记录热刺激引起的肢体退缩时延。[0336]非刺激诱发行为测试:步态分析:使用digigait和坐骨神经指数评估坐骨神经运动功能172。简而言之,利用rodentbehavioralcore中可用的digigait设备(我们的实验室已对其进行了表征和优化),将在损伤前和术后第7、14和28天测定坐骨神经功能。将大鼠放置在digigait支架内的电动跑步机上,并设置20cm/s的速度记录所有动物。坐骨神经指数程序将用于分析受影响后肢的功能。[0337]条件性逃避/回避:该步骤旨在验证以下假设,即大鼠将避免与对痛觉过敏区域的机械刺激相关的环境。据报道,在使用止痛药治疗后,受伤大鼠对逃避/回避行为的变化具有敏感性(baastrup等人,2010)174。labuda和fuchs(2000)171描述了大脑依赖的条件性逃避/回避试验测试,该测试依赖于对新厌恶环境的逃避/回避学习。受伤后3至5周,由同一名受训研究人员仅对动物进行一次测试。测试将在usurodentbehavioralcore中进行。将允许动物在试验室中适应环境光线和噪音条件至少1小时。在本试验中,大鼠可自由进入封闭室的“非厌恶性”黑暗侧和“厌恶性”光明侧,封闭室具有网状地板,可通过vonfrey丝从下方轻松进入。如果大鼠分别处于黑暗或光明区域,受伤或未受伤的后爪都会受到常规刺激。每15秒,根据动物在此期间的位置,将足以在受伤的爪子上引发撤退反应的机械刺激(vonfrey丝)施加到动物一只后爪的外侧足底表面,持续30分钟。逃逸/回避行为将定义为从黑暗区域向光明区域的转移。将记录在方框白色侧花费的时间百分比以及黑白侧之间的交叉次数。白色侧的累积时间和交叉总数将用作逃避/回避学习的指示。[0338]安乐死:为了进行形态学和免疫组化分析,将用氯胺酮/甲苯噻嗪(80至100mg/kg 10mg/kg,i.p.,21号针)麻醉大鼠,然后用300ml磷酸盐缓冲盐水(ph7.4),然后是0.1m磷酸盐缓冲液中的300ml4%多聚甲醛经心灌注。灌注后,解剖神经、drg和脊髓,在4%多聚甲醛中固定24小时,在30%蔗糖中冷冻24小时。使用低温恒温器(leicacm3050s,leicabiosystems,wetzlar,germany)将组织样品切割成10μm的切片。[0339]组织学:安乐死和神经取样后,切片将用苏木精和曙红染色。其余靶基因oprm1、nav1.8、kv4.3和kv7.2的表达水平和分布将通过免疫荧光进行评估。[0340]基因表达分析:对于坐骨神经及其相关drg和脊髓节段的基因表达分析,rest靶基因oprm1、nav1.8,kv4.3和kv7.2和对照基因(hprt、gapd、rn18s)将通过qrt-pcr进行评估。[0341]蛋白质表达分析:坐骨神经相关drg的蛋白质表达分析将通过oprm1、nav1.8、kv4.3和kv7.2的sds-page免疫印迹进行评估。gapdh将用作加载对照。[0342]统计方法:对于行为数据,结果将以平均值±平均值标准误差(sem)表示。基于治疗后的时间,将使用双因素方差分析来评估各组动物。bonferroni-holm方法将用于调整多重比较。双尾统计显著性将被确定为p《0.05。对于免疫组化分析,将使用双因素anova来比较交互作用的平均值以及组和时间的影响。对于qrt-pcr,值将报告为平均值的平均值±标准误差(sem)。通过单因素方差分析(anova)和tukey的多重比较检验,确定药物对基因表达的统计显著影响。[0343]下表18概述了第2阶段的实验步骤:[0344]表18[0345][0346][0347]在第2阶段,将检查融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽减轻疼痛的持久性。在本研究中,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽剂量(低、中或高),以及在第1阶段确定为最有效的给药期(14或28天)将被使用。将遵循上述第1阶段疗效研究的相同方法。不同之处在于,施用期(14或28天)后,每隔30天对大鼠进行一次疼痛反应和运动功能的监测和行为评估。大鼠将在第30天被安乐死。本研究共需要72只2个月大的sd大鼠(雄性和雌性数量相等)。每组12只大鼠的样本量将有80%的效力检测疼痛评估中标准差1.20的差异。效力分析基于独立样本的t检验,具有5%的双侧显著性水平。将大鼠随机分配到右侧表格中指定的实验组。对10个随机分组的大鼠进行刺激诱发和非刺激诱发行为测试、形态学、免疫组化分析、定量rt-pcr(qrt-pcr)分析和wb分析。[0348]目标3:融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的毒理学和安全性药理学评估。为了支持首次在人体内给药,我们将根据fda指南m3(r2)、s2b、s6(r1)、s7a和s7b的建议175-179,评估两种融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的体外心脏和遗传毒性以及体内毒理学、安全药理学和毒代动力学。在这两个物种中,将根据目标1中的pk评估结果选择给药途径。[0349]子目标3.1:猴子的单剂量范围判定:将在3只猴子/性别/组中评估四个剂量水平 溶剂对照。hd应确定mtd、mfd和/或暴露饱和度。较低剂量将以1/3的增量间隔。将在6个时间点采集血液(根据目标1中收集的pk数据确定),并评估cmax、auc和t1/2。将每日评估生存率和临床症状。[0350]子目标3.2和3.3:大鼠(3.2)和猴子(3.3)的毒理学研究。这两个物种的给药期均为4周,以支持相同时间的人类给药 6周的恢复期,其由于在细胞检测中观察到rpp的高稳定性而被选择77。将有三个剂量水平 溶剂对照。高剂量将是剂量范围判定(大鼠目标1.2和猴子目标3.1)中确定的最大耐受剂量。根据范围判定中的auc,md将比hd低1/3。ld将接近体内药效研究中的有效剂量(目标2)。给药间隔为每个物种中与融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的t1/2。在大鼠研究中,将有15只/性别/组。10只/性别/组将在给药期结束时被处死,5只/性别/溶剂将在恢复期结束时被处死。此外,9只/性别/rpp组和3只/性别/溶剂组将被分配到研究的毒代动力学部分。在猴子研究中,将对所有动物进行毒理学和tk评估(主要研究中为3只/性别/组,恢复期为2只/性别/组)。将对这两个物种的以下毒理学参数进行评估:临床观察、体重、食物消耗、眼底镜检查(眼底镜检查和裂隙灯检查),并将评估一组标准的临床化学、器官重量和组织学参数。猴子将接受使用非手术遥测功能测定的呼吸(例如潮气量和血红蛋白血氧饱和度)和心脏心电图记录评估179,按照fda指南s7a的建议179。心血管评估将在治疗前、第一次给药后和恢复期结束时(如需要)进行两次。这些记录将由顾问委员会认证的心脏病专家进行定性评估。对所有波形进行定性评估,以检测心律或传导障碍,包括pr和qrs间期的评估。将使用改良的irwin功能观察组合(battery)对大鼠的中枢神经系统功能进行评估179,180。将在6个时间点(由范围判定研究确定)进行血浆tk评估(cmax、auc和t1/2)。[0351]子目标3.4:体外herg检测177。herg通道抑制是长qt综合征的常见原因,也与可导致心室颤动和心源性猝死的心律失常相关。为了测试融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否抑制herg通道电流,我们将对用高浓度融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽(10μm)或阳性对照(西沙必利,0.03μm)处理的cho细胞中的herg通道电流进行电生理学评估。每个细胞将作为自身的阳性对照。全细胞记录将使用传统的电压钳技术进行。[0352]子目标3.5:遗传毒性。将在符合oecd的体外ames和体外染色体畸变检测(s2b,1997年11月)176中评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,以确定其遗传毒性潜力。[0353]目标4:融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的滥用可能性评估。由于阿片受体基因表达被rest抑制,因此融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可能增强内源性阿片受体配体(如内啡肽181,182)的作用。此外,最近的研究确实表明,在涉及疼痛的情况下,drg和导水管周围灰质(pag)中rest的表达增加183,184。为了确定融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽是否产生奖赏寻求行为,我们将使用条件性位置偏爱(cpp)评估其对大鼠的影响,这是一种广泛使用的行为检测,用于测试药物的奖赏特性185-189。cpp是一种成熟的方法,其已在sni动物模型中证明了吗啡的奖赏寻求行为。[0354]首先,将融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽施用给未受伤的大鼠(途径tbd),然后进行cpp。为了与目标2保持一致,将使用羟考酮代替吗啡作为阳性对照,并将生理盐水注射到单独的大鼠体内作为阴性对照。[0355]设备由三个隔室组成:两个外部隔室设计为具有不同的特性(例如白色和黑色墙壁),中央隔室没有特殊特性。每个实验包括三个阶段。i期预试验(第1天):施用生理盐水后,将大鼠放置在中心隔室中,打开闸门,以便进入其他隔室。将对大鼠进行15分钟的监测,通过记录在每个小室中的时间来确定基线偏好。将确定隔室的平均自发偏好时间。在任何一个试验室中花费超过60%试验时间的大鼠将被视为有偏好,并将从试验中移除。ii期条件反射形成(第2-7天):给大鼠注射溶剂或rpp(seqidno:1和15至17)或rppv(seqidno:18至117)融合到cpp(seqidno:118至137和140至159)的肽,然后,他们将被随机分配/调节,以在一个隔室和另一个隔室中接受与cpp(seqidno:118至137和140至159)融合的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽。与cpp(seqidno:118至137和140至159)融合的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的剂量将基于对目标2中确定的机械和热痛觉过敏产生可测定影响的浓度。治疗室以及药物和溶剂的给药顺序将对所有组进行平衡,以避免偏见。该阶段包括六个连续的30分钟的调节期。对于单独的一组,也将使用羟考酮进行调节。iii期试验(第8天):将向大鼠注射生理盐水,并将其放置在cpp箱的中心,并监测15分钟。后处理期间在药物对隔室中花费的时间与预处理期间花费的时间之间的差异将用于评估rpp(seqidno:1和15至17)或rppv(seqidno:18至117)引起的位置调节程度融合到cpp的肽(seqidno:118至137和140至159)190。[0356]将使用8组12只受伤大鼠(sni和假手术组,用羟考酮、融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽形成条件反射,对融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽,和生理盐水)进行评估。每组12只大鼠的样本量将有80%的效力检测疼痛评估中1.20个标准差的差异。效力分析基于独立样本的t检验,具有5%的双侧显著性水平。将大鼠随机分配到实验组。将随机分配大鼠进行sni或假手术。vonfrey试验将在受伤前一天、受伤后4天和8天进行,以监测疼痛状态。受伤后8天,cpp测试将如前所述进行。进行行为测试的研究助理将不知道大鼠被随机分配到哪个实验组/对照组。每只动物都有一个唯一的动物编号。将使用秘密彩色代码指定对照组和实验组。只有在分析之后,才会显示此代码。将对动物实施安乐死,并按照目标2所述对样本进行分析。[0357]预测结果和替代策略:我们预测,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽将降低受损神经元的rest水平,并恢复兴奋性所需的离子通道的正确表达。在行为水平上,融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽应使sni大鼠的刺激或非刺激诱发行为测试恢复到基线水平。此外,鉴于其作用模式,我们不期望融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽具有成瘾性。然而,如果药物管理显示出滥用可能性,我们将通过融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽施用期间的渐进比率(pr)强化表来确认这一阳性结果。[0358]实施例15-融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽可用于预防多形性胶质母细胞瘤(gbm)的复发[0359]评估融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽对脑肿瘤初始细胞(btic)和体内的神经发生和抗癌效力。将使用我们建立的方法,在mayoclinic获得的10名iv级gbm患者的原代培养btc中,评估与融合到cpp(seqidno:118至137和140至159)的供试品rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽的活性191-193。将细胞(0.25×104)铺板于(3.8cm2tc皿)培养基(250μl)中,所述培养基以最佳剂量范围(在实施例13所述的神经元分化研究中确定)添加5μl溶剂或3个剂量水平的供试品,持续0、2、4或8天(主要研究),然后在洗出供试品8天后恢复(恢复期)。通过wb和rt-pcr,在给药期间(主要研究)的第0、2、4和8天以及恢复期间(每天3个/tc孔/组)的第10、13和16天监测多能性(sox2和巢蛋白)、增殖(ki67)、细胞死亡(caspase3)和分化(map2、神经元和gfap、胶质细胞)的标志物。[0360]来自10名iv级gbm患者的原代培养btc将用于我们和其他人之前描述的颅内异种成瘤裸鼠模型191,192,194。简而言之,将细胞(5×104)悬浮在5μlpbs(对照组)或含有3tbd(基于上述btic神经发生效力检测)供试品剂量水平之一的pbs中,并使用带导螺杆系统的自动注射泵输送112,195。注入坐标为x:1.5;y:1.34;z:-3.5靶向小鼠纹状体;高度可靠的肿瘤移植部位196。使用kaplan-meier方法197对小鼠在第4、8、16和20周的存活率进行检测,然后进行死后组织病理学检查,以评估肿瘤大小和侵袭性(h&e染色)、细胞死亡(tunel)和增殖(抗人细胞核)(所有步骤均为3个/性别/组)。[0361]统计方法:如图所示,数值将报告为平均值±平均值标准误差(sem)或标准偏差(sd)。统计显著性将通过应用学生t检验的适当参数或非参数变化或单因素方差分析(anova)来确定。简而言之,将使用kruskal-wallis检验对不同对照和候选药物治疗的总rest与磷酸化rest的比率进行统计检验。将使用tukey多重比较试验对btic对照和候选药物培养物以及原位异种移植体之间标记物表达的差异进行统计检验。注射注射btic的用对照品或供试品治疗的小鼠之间的肿瘤大小和侵袭性差异将使用mann-whitneyu检验进行统计检验,生存率差异将使用对数秩检验和cox比例风险检验198进行统计检验。[0362]预测结果:与对照组相比,在每个时间点,我们预计有btic(给药供试品)异种移植物的小鼠会出现无瘤或侵袭性较小的肿瘤,细胞死亡(tunel)更多,gbm增殖(抗人细胞核)更少。此外,我们预计注射供试品和btic的小鼠与单独注射btic的小鼠相比,存活时间会延长。[0363]材料和方法[0364]测试系统[0365]细胞:原代培养的脑肿瘤起始细胞(btic)来自10例iv级gbm患者。alfredoquinoneshinojosa博士及其研究团队之前已经描述了提取和繁殖btic的方法191-193。[0366]物种/菌株:nu/nu-harlansprague-dawley小鼠[0367]年龄:研究开始时:6周[0368]体重:研究开始时:雄性:20-30克;雌性:18-35g[0369]供试品标识。[0370]融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽[0371]纯度:≥95%hplc[0372]剂型制备和稳定性:每周将批量制备融合到cpp(seqidno:118至137和140至159)的rpp(seqidno:1和15至17)或rppv(seqidno:18至117)肽和溶剂对照品。将在每周开始时评估剂型浓度。可接受的浓度范围为标称浓度的±10%。[0373]溶剂:水[0374]研究设计[0375]供试品的神经发生效力研究:[0376]剂量:3个供试品剂量水平 溶剂对照。将根据实施例11中所述神经元分化研究的结果确定供试品剂量。[0377]施用:每2天给药一次,并更换培养基。[0378]持续时间:主要研究:0、2、4或8天;恢复期:2、5或8天。[0379]重复数:每组3个组织培养(tc)孔。[0380]给药体积:5μl[0381]分析参数:将在给药期间(主要研究)的第0、2、4和8天以及恢复期间(每天3个/tc孔/组)的第10、13和16天(表10),通过wb和rt-pcr监测多能性(sox2和nestin)、增殖(ki67)、细胞死亡(caspase3)和分化(map2、神经元和gfap、胶质细胞)的标记物。[0382][0383][0384]供试品对小鼠btic的抗癌效力:[0385]剂量:3剂供试品 溶剂对照。供试品剂量将基于神经源性效力研究的结果。[0386]施用频率:单剂量[0387]途径:带导向螺杆系统的自动注射泵112,195。注入坐标为x:1.5;y:1.34;z:-3.5靶向小鼠纹状体;高度可靠的肿瘤移植部位196。[0388]溶剂:磷酸盐缓冲盐水(pbs)[0389]动物数量:3只/性别/组[0390]给药体积:5μl[0391]参数分析:将使用kaplan-meier方法197对小鼠在第4、8、16和20周的存活率进行检测,然后进行死后组织病理学检查,以评估肿瘤大小和侵袭性(h&e染色)、细胞死亡(tunel)和增殖(抗人细胞核)(表11)。[0392][0393][0394]环境条件[0395]居住条件:2只动物/笼,光/暗循环12小时。[0396]饮食:随意进食进水。[0397]下表19示出了seqidno:1至324的序列:[0398][0399][0400][0401][0402][0403][0404][0405][0406][0407][0408][0409][0410][0411][0412][0413][0414][0415][0416][0417][0418]尽管本发明的某些当前优选实施例已在本文中具体描述,但对于本发明所属领域的技术人员来说,显而易见的是,可以在不脱离本发明的精神和范围的情况下对本文所示和描述的各种实施方案进行变更和修改。[0419]参考文献(说明书,实施例除外)sci5,573-587,doi:10.1002/wcs.1304(2014).[0445]26samuels,i.s.etal.deletionoferk2mitogen-activatedproteinkinaseidentifiesitskeyrolesincorticalneurogenesisandcognitivefunction.jneurosci28,6983-6995,doi:28/27/6983[pii]10.1523/jneurosci.0679-08.2008(2008).[0446]27shors,t.j.etal.neurogenesisintheadultisinvolvedintheformationoftracememories.nature410,372-376,doi:10.1038/35066584(2001).[0447]28villeda,s.a.etal.theageingsystemicmilieunegativelyregulatesneurogenesisandcognitivefunction.nature477,90-94,doi:10.1038/nature10357(2011).[0448]29bird,c.m.&burgess,n.thehippocampussupportsrecognitionmemoryforfamiliarwordsbutnotunfamiliarfaces.currbiol18,1932-1936,doi:10.1016/j.cub.2008.10.046(2008).[0449]30t.j.shors,g.m.,a.beylin,m.zhao,t.rydel,e.gould.neurogenesisintheadultisinvolvedintheformationoftracememories.nature410,372-376(2001).[0450]31vanelzakker,m.,fevurly,r.d.,breindel,t.&spencer,r.l.environmentalnoveltyisassociatedwithaselectiveincreaseinfosexpressionintheoutputelementsofthehippocampalformationandtheperirhinalcortex.learnmem15,899-908,doi:10.1101/lm.1196508(2008).[0451]32thornton,g.k.&woods,c.g.primarymicrocephaly:doallroadsleadtorome?trendsgenet25,501-510,doi:10.1016/j.tig.2009.09.011(2009).[0452]33ghaziuddin,m.,zaccagnini,j.,tsai,l.&elardo,s.ismegalencephalyspecifictoautism?jintellectdisabilres43(pt4),279-282(1999).[0453]34kuhn,h.g.,cooper-kuhn,c.m.,boekhoorn,k.&lucassen,p.j.changesinneurogenesisindementiaandalzheimermousemodels:aretheyfunctionallyrelevant?eurarchpsychiatryclinneurosci257,281-289,doi:10.1007/s00406-007-0732-4(2007).[0454]35baker,m.tumourssparkstem-cellreview.nature457,941,doi:10.1038/457941a(2009).[0455]36brederlau,a.etal.transplantationofhumanembryonicstemcell-derivedcellstoaratmodelofparkinson'sdisease:effectofinvitrodifferentiationongraftsurvivalandteratomaformation.stemcells24,1433-1440,doi:2005-0393[pii]10.1634/stemcells.2005-0393(2006).[0456]37burns,t.c.,verfaillie,c.m.&low,w.c.stemcellsforischemicbraininjury:acriticalreview.jcompneurol515,125-144,doi:10.1002/cne.22038(2009).[0457]38carlson,a.p.,schermer,c.r.&lu,s.w.retrospectiveevaluationofanemiaandtransfusionintraumaticbraininjury.jtrauma61,567-571,doi:theregulationofneuronalexcitability.jneurosci30,13235-13245,doi:10.1523/jneurosci.1981-10.2010(2010).[0484]65escayg,a.etal.mutationsofscn1a,encodinganeuronalsodiumchannel,intwofamilieswithgefs 2.natgenet24,343-345,doi:10.1038/74159(2000).[0485]66martin,d.etal.restrepressesasubsetofthepancreaticendocrinedifferentiationprogram.devbiol405,316-327,doi:10.1016/j.ydbio.2015.07.002(2015).[0486]67abderrahmani,a.etal.neuronaltraitsarerequiredforglucose-inducedinsulinsecretion.febslett565,133-138,doi:10.1016/j.febslet.2004.04.002(2004).[0487]68martin,d.etal.functionalsignificanceofrepressorelement1silencingtranscriptionfactor(rest)targetgenesinpancreaticbetacells.diabetologia51,1429-1439,doi:10.1007/s00125-008-0984-1(2008).[0488]69rigamonti,d.etal.lossofhuntingtinfunctioncomplementedbysmallmoleculesactingasrepressorelement1/neuronrestrictivesilencerelementsilencermodulators.jbiolchem282,24554-24562,doi:10.1074/jbc.m609885200(2007).[0489]70sipione,s.etal.earlytranscriptionalprofilesinhuntingtin-induciblestriatalcellsbymicroarrayanalyses.hummolgenet25,210,doi:10.1093/hmg/ddv416(2016).[0490]71zuccato,c.etal.huntingtininteractswithrest/nrsftomodulatethetranscriptionofnrse-controlledneuronalgenes.natgenet35,76-83,doi:10.1038/ng1219(2003).[0491]72bao,s.etal.targetingcancerstemcellsthroughl1camsuppressesgliomagrowth.cancerres68,6043-6048,doi:10.1158/0008-5472.can-08-1079(2008).[0492]73jackson,m.,hassiotou,f.&nowak,a.glioblastomastem-likecells:attherootoftumorrecurrenceandatherapeutictarget.carcinogenesis36,177-185,doi:10.1093/carcin/bgu243(2015).[0493]74liu,g.etal.analysisofgeneexpressionandchemoresistanceofcd133 cancerstemcellsinglioblastoma.molcancer5,67,doi:10.1186/1476-4598-5-67(2006).[0494]75persano,l.,rampazzo,e.,basso,g.&viola,g.glioblastomacancerstemcells:roleofthemicroenvironmentandtherapeutictargeting.biochempharmacol85,612-622,doi:10.1016/j.bcp.2012.10.001(2013).[0495]76conti,l.etal.restcontrolsself-renewalandtumorigeniccompetenceofhumanglioblastomacells.plosone7,e38486,doi:10.1371/journal.pone.0038486(2012).[0496]77kamal,m.m.etal.restregulatesoncogenicpropertiesofglioblastomastemcells.stemcells30,405-414,doi:10.1002/stem.1020(2012).[0497]78wagoner,m.p.&roopra,a.arestderivedgenesignaturestratifiesglioblastomasintochemotherapyresistantandresponsivedisease.bmcgenomics13,686,doi:10.1186/1471-2164-13-686(2012).[0498]79taylor,p.etal.restisanovelprognosticfactorandtherapeutictargetformedulloblastoma.molcancerther11,1713-1723,doi:10.1158/1535-7163.mct-11-0990(2012).[0499]80jin,h.etal.identificationofre1-silencingtranscriptionfactorasapromoterofmetastasisinpancreaticcancer.frontoncol9,291,doi:10.3389/fonc.2019.00291(2019).[0500]参考文献(仅针对实施例)[0501]1bruce,a.w.etal.genome-wideanalysisofrepressorelement1silencingtranscriptionfactor/neuron-restrictivesilencingfactor(rest/nrsf)targetgenes.procnatlacadsciusa101,10458-10463,doi:10.1073/pnas.04018271010401827101[pii](2004).[0502]2nesti,e.,corson,g.m.,mccleskey,m.,oyer,j.a.&mandel,g.c-terminaldomainsmallphosphatase1andmapkinasereciprocallycontrolreststabilityandneuronaldifferentiation.procnatlacadsciusa111,e3929-3936,doi:1414770111[pii]10.1073/pnas.1414770111(2014).[0503]3nesti,e.methodsandcompositionsusefulinmanipulatingthestabilityofre1silencingtranscriptionfactor.us,ptcpatent(2014,2015).[0504]4nesti,e.harnessingthemastertranscriptionalrepressorresttoreciprocallyregulateneurogenesis..neurogenesis2,doi:10.1080/23262133.2015.1055419(2015).[0505]5arranz-gibert,p.etal.immunosilencingpeptidesbystereochemicalinversionandsequencereversal:retro-d-peptides.scirep8,6446,doi:10.1038/s41598-018-24517-6(2018).[0506]6hall,e.d.,bryant,y.d.,cho,w.&sullivan,p.g.evolutionofpost-traumaticneurodegenerationaftercontrolledcorticalimpacttraumaticbraininjuryinmiceandratsasassessedbythedeolmossilverandfluorojadestainingmethods.jneurotrauma25,235-247,doi:10.1089/neu.2007.0383(2008).[0507]7calderone,a.etal.ischemicinsultsderepressthegenesilencerrestinneuronsdestinedtodie.jneurosci23,2112-2121,doi:23/6/2112[pii](2003).[0508]8kaneko,n.,hwang,j.y.,gertner,m.,pontarelli,f.&zukin,r.s.caseinkinase1suppressesactivationofrestininsultedhippocampalneuronsandhaltsischemia-inducedneuronaldeath.jneurosci34,6030-6039,doi:34/17/6030[pii]10.1523/jneurosci.4045-13.2014(2014).[0509]9noh,k.m.etal.repressorelement-1silencingtranscriptionfactor(rest)-dependentepigeneticremodelingiscriticaltoischemia-inducedneuronaldeath.procnatlacadsciusa109,e962-971,doi:1121568109[pii]10.1073/pnas.1121568109(2012).[0510]10abe,k.therapeuticpotentialofneurotrophicfactorsandneuralstemcellsagainstischemicbraininjury.jcerebbloodflowmetab20,1393-1408,doi:10.1097/00004647-200010000-00001(2000).[0511]11kanding,p.k.g.,andramondiaz-arrastia.intranslationalresearchintraumaticbraininjury(edgrantglaskowitzd)ch.14,(taylorandfrancis,2016).[0512]12pasquina,p.,kirtley,r.&ling,g.moderate-to-severetraumaticbraininjury.seminneurol34,572-583,doi:10.1055/s-0034-1396010(2014).[0513]13hwang,j.y.,kaneko,n.,noh,k.m.,pontarelli,f.&zukin,r.s.thegenesilencingtranscriptionfactorrestrepressesmir-132expressioninhippocampalneuronsdestinedtodie.jmolbiol426,3454-3466,doi:10.1016/j.jmb.2014.07.032(2014).[0514]14mcclelland,s.etal.thetranscriptionfactornrsfcontributestoepileptogenesisbyselectiverepressionofasubsetoftargetgenes.elife3,e01267(2014).[0515]15lu,t.etal.restandstressresistanceinageingandalzheimer'sdisease.nature507,448-454,doi:nature13163[pii]10.1038/nature13163(2014).[0516]16orta-salazar,e.etal.rest/nrsf-inducedchangesofchatproteinexpressionintheneocortexandhippocampusofthe3xtg-admousemodelforalzheimer'sdisease.lifesciences116,83-89,doi:10.1016/j.lfs.2014.09.013(2014).[0517]17sharp,d.j.,scott,g.&leech,r.networkdysfunctionaftertraumaticbraininjury.natrevneurol10,156-166,doi:10.1038/nrneurol.2014.15(2014).[0518]18roopra,a.,dingledine,r.&hsieh,j.epigeneticsandepilepsy.epilepsia53suppl9,2-10,doi:10.1111/epi.12030(2012).[0519]19bergsland,m.,covacu,r.,perezestrada,c.,svensson,m.&brundin,l.nitricoxide-inducedneuronaltogliallineagefate-changedependsonnrsf/restfunctioninneuralprogenitorcells.stemcells32,2539-2549,doi:10.1002/stem.1749(2014).[0520]20conaco,c.,otto,s.,han,j.j.&mandel,g.reciprocalactionsofrestandamicrornapromoteneuronalidentity.procnatlacadsciusa103,2422-2427,doi:0511041103[pii]10.1073/pnas.0511041103(2006).[0521]21covey,m.v.,streb,j.w.,spektor,r.&ballas,n.restregulatesthepoolsizeofthedifferentneurallineagesbyrestrictingthegenerationofneuronsandoligodendrocytesfromneuralstem/progenitorcells.development139,2878-2890,doi:dev.074765[pii]10.1242/dev.074765(2012).[0522]22gao,z.etal.themasternegativeregulatorrest/nrsfcontrolsadultneurogenesisbyrestrainingtheneurogenicprograminquiescentstemcells.jneurosci31,9772-9786,doi:31/26/9772[pii]10.1523/jneurosci.1604-11.2011(2011).[0523]23kohyama,j.etal.bmp-inducedrestregulatestheestablishmentandmaintenanceofastrocyticidentity.jcellbiol189,159-170,doi:jcb.200908048[pii]10.1083/jcb.200908048(2010).[0524]24mandel,g.etal.repressorelement1silencingtranscriptionfactor(rest)controlsradialmigrationandtemporalneuronalspecificationduringneocorticaldevelopment.procnatlacadsciusa108,16789-16794,doi:1113486108[pii]10.1073/pnas.1113486108(2011).[0525]25vincent,a.s.,roebuck-spencer,t.m.&cernich,a.cognitivechangesanddementiariskaftertraumaticbraininjury:implicationsforagingmilitarypersonnel.alzheimersdement10,s174-187,doi:10.1016/j.jalz.2014.04.006(2014).[0526]26kirn,j.r.therelationshipofneurogenesisandgrowthofbrainregionstosonglearning.brainlang115,29-44,doi:s0093-934x(09)00135-7[pii]10.1016/j.bandl.2009.09.006(2010).[0527]27oomen,c.a.,bekinschtein,p.,kent,b.a.,saksida,l.m.&bussey,t.j.adulthippocampalneurogenesisanditsroleincognition.wileyinterdisciprevcognsci5,573-587,doi:10.1002/wcs.1304(2014).[0528]28samuels,i.s.etal.deletionoferk2mitogen-activatedproteinkinaseidentifiesitskeyrolesincorticalneurogenesisandcognitivefunction.jneurosci28,6983-6995,doi:28/27/6983[pii]10.1523/jneurosci.0679-08.2008(2008).[0529]29shors,t.j.etal.neurogenesisintheadultisinvolvedintheformationoftracememories.nature410,372-376,doi:10.1038/35066584(2001).[0530]30villeda,s.a.etal.theageingsystemicmilieunegativelyregulatesneurogenesisandcognitivefunction.nature477,90-94,doi:10.1038/nature10357(2011).[0531]31bird,c.m.&burgess,n.thehippocampussupportsrecognitionmemoryforfamiliarwordsbutnotunfamiliarfaces.currbiol18,1932-1936,doi:10.1016/j.cub.2008.10.046(2008).[0532]32t.j.shors,g.m.,a.beylin,m.zhao,t.rydel,e.gould.neurogenesisintheadultisinvolvedintheformationoftracememories.nature410,372-376(2001).[0533]33vanelzakker,m.,fevurly,r.d.,breindel,t.&spencer,r.l.environmentalnoveltyisassociatedwithaselectiveincreaseinfosexpressionintheoutputelementsofthehippocampalformationandtheperirhinalcortex.learnmem15,899-908,doi:10.1101/lm.1196508(2008).[0534]34thornton,g.k.&woods,c.g.primarymicrocephaly:doallroadsleadtorome?trendsgenet25,501-510,doi:10.1016/j.tig.2009.09.011(2009).[0535]35ghaziuddin,m.,zaccagnini,j.,tsai,l.&elardo,s.ismegalencephalyspecifictoautism?jintellectdisabilres43(pt4),279-282(1999).[0536]36kuhn,h.g.,cooper-kuhn,c.m.,boekhoorn,k.&lucassen,p.j.changesinneurogenesisindementiaandalzheimermousemodels:aretheyfunctionallyrelevant?eurarchpsychiatryclinneurosci257,281-289,doi:10.1007/s00406-007-0732-4(2007).[0537]37baker,m.tumourssparkstem-cellreview.nature457,941,doi:10.1038/457941a(2009).[0538]38brederlau,a.etal.transplantationofhumanembryonicstemcell-derivedcellstoaratmodelofparkinson'sdisease:effectofinvitrodifferentiationongraftsurvivalandteratomaformation.stemcells24,1433-1440,doi:2005-0393[pii]10.1634/stemcells.2005-0393(2006).[0539]39burns,t.c.,verfaillie,c.m.&low,w.c.stemcellsforischemicbraininjury:acriticalreview.jcompneurol515,125-144,doi:10.1002/cne.22038(2009).[0540]40carlson,a.p.,schermer,c.r.&lu,s.w.retrospectiveevaluationofanemiaandtransfusionintraumaticbraininjury.jtrauma61,567-571,doi:10.1097/01.ta.0000231768.44727.a200005373-200609000-00007[pii](2006).[0541]41carmeliet,p.&storkebaum,e.vascularandneuronaleffectsofvegfinthenervoussystem:implicationsforneurologicaldisorders.semincelldevbiol13,39-53,doi:10.1006/scdb.2001.0290s1084952101902903[pii](2002).[0542]42choe,y.,kozlova,a.,graf,d.&pleasure,s.j.bonemorphogenicproteinsignalingisamajordeterminantofdentatedevelopment.jneurosci33,6766-6775,doi:33/16/6766[pii]10.1523/jneurosci.0128-13.2013(2013).[0543]43erdo,f.etal.host-dependenttumorigenesisofembryonicstemcelltransplantationinexperimentalstroke.jcerebbloodflowmetab23,780-785,doi:10.1097/01.wcb.0000071886.63724.fb(2003).[0544]44greig,n.h.etal.incretinmimeticsaspharmacologictoolstoelucidateandasanewdrugstrategytotreattraumaticbraininjury.alzheimersdement10,s62-75,doi:s1552-5260(13)02925-7[pii]10.1016/j.jalz.2013.12.011(2014).[0545]45kaplan,g.b.,vasterling,j.j.&vedak,p.c.brain-derivedneurotrophicfactorintraumaticbraininjury,post-traumaticstressdisorder,andtheircomorbidconditions:roleinpathogenesisandtreatment.behavpharmacol21,427-437,doi:10.1097/fbp.0b013e32833d8bc9(2010).[0546]46knoepfler,p.s.deconstructingstemcelltumorigenicity:aroadmaptosaferegenerativemedicine.stemcells27,1050-1056,doi:10.1002/stem.37(2009).dysfunctionsinneuropathicpain.journalofneuroscience30,4806-4814,doi:10.1523/jneurosci.5541-09.2010(2010).[0561]61alvarado,s.etal.peripheralnerveinjuryisaccompaniedbychronictranscriptome-widechangesinthemouseprefrontalcortex.molpain9,21,doi:10.1186/1744-8069-9-21(2013).[0562]62hammer,p.etal.mrna-seqwithagnosticsplicesitediscoveryfornervoussystemtranscriptomicstestedinchronicpain.genomeres20,847-860,doi:10.1101/gr.101204.109(2010).[0563]63willis,d.e.,wang,m.,brown,e.,fones,l.&cave,j.w.selectiverepressionofgeneexpressioninneuropathicpainbytheneuron-restrictivesilencingfactor/repressorelement-1silencingtranscription(nrsf/rest).neuroscienceletters625,20-25,doi:10.1016/j.neulet.2015.12.003(2016).[0564]64ballas,n.,grunseich,c.,lu,d.d.,speh,j.c.&mandel,g.restanditscorepressorsmediateplasticityofneuronalgenechromatinthroughoutneurogenesis.cell121,645-657,doi:s0092-8674(05)00285-0[pii]10.1016/j.cell.2005.03.013(2005).[0565]65chong,j.a.etal.rest:amammaliansilencerproteinthatrestrictssodiumchannelgeneexpressiontoneurons.cell80,949-957,doi:0092-8674(95)90298-8[pii](1995).[0566]66mortazavi,a.,leeperthompson,e.c.,garcia,s.t.,myers,r.m.&wold,b.comparativegenomicsmodelingofthenrsf/restrepressornetwork:fromsingleconservedsitestogenome-widerepertoire.genomeres16,1208-1221,doi:gr.4997306[pii]10.1101/gr.4997306(2006).[0567]67otto,s.j.etal.anewbindingmotifforthetranscriptionalrepressorrestuncoverslargegenenetworksdevotedtoneuronalfunctions.jneurosci27,6729-6739,doi:27/25/6729[pii]10.1523/jneurosci.0091-07.2007(2007).[0568]68schoenherr,c.j.&anderson,d.j.silencingisgolden:negativeregulationinthecontrolofneuronalgenetranscription.curropinneurobiol5,566-571,doi:0959-4388(95)80060-3[pii](1995).[0569]69schoenherr,c.j.,paquette,a.j.&anderson,d.j.identificationofpotentialtargetgenesfortheneuron-restrictivesilencerfactor.procnatlacadsciusa93,9881-9886(1996).[0570]70rose,k.etal.transcriptionalrepressionofthemchannelsubunitkv7.2inchronicnerveinjury.pain152,742-754,doi:10.1016/j.pain.2010.12.028(2011).[0571]71costigan,m.etal.multiplechronicpainstatesareassociatedwithacommonaminoacid-changingalleleinkcns1.brain133,2519-2527,doi:10.1093/brain/awq195(2010).[0572]72zhang,f.etal.repressorelement1-silencingtranscriptionfactordrivesthedevelopmentofchronicpainstates.pain,doi:10.1097/j.pain.0000000000001633(2019).[0573]73zhang,j.,chen,s.r.,chen,h.&pan,h.l.re1-silencingtranscriptionfactorcontrolstheacute-to-chronicneuropathicpaintransitionandchrm2receptorgeneexpressioninprimarysensoryneurons.jbiolchem293,19078-19091,doi:10.1074/jbc.ra118.005846(2018).[0574]74ueda,h.etal.amimeticofthemsin3-bindinghelixofnrsf/restamelioratesabnormalpainbehaviorinchronicpainmodels.bioorgmedchemlett27,4705-4709,doi:10.1016/j.bmcl.2017.09.006(2017).[0575]75uchida,h.,sasaki,k.,ma,l.&ueda,h.neuron-restrictivesilencerfactorcausesepigeneticsilencingofkv4.3geneafterperipheralnerveinjury.neuroscience166,1-4,doi:10.1016/j.neuroscience.2009.12.021(2010).[0576]76gervasinm.,d.a.,clarkdm.,dinglem,,pisarchikav.,nestilj.c-terminaldomainsmallphosphatase1(ctdsp1)regulatesgrowthfactorexpressionandaxonalregeneration.journaloftranslationalmedicinemanuscriptedsubmittedforpublication.(2020).[0577]77nesti,e.,pisarchik,a.compositionsandmethodsforderepressingre1silencingtranscriptionfactortargetgenes.usapatent(2019).[0578]78cole,l.a.,kurscheid,s.,nekrasov,m.,domaschenz,r.,dennis,j.h.,tremethick,d.j.redefiningthenucleosomalarchitectureofactiveandinactivepromotersinthecontextofcellularplasticityandcancer.naturecommunicationsrevisionreturned(2020).[0579]79ren,y.,vera,d.l.,hughes,k.a.&dennis,j.h.stimulationofthedrosophilaimmunesystemaltersgenome-widenucleosomeoccupancy.genomdata3,146-147,doi:10.1016/j.gdata.2015.01.001(2015).[0580]80sexton,b.s.etal.thespring-loadedgenome:nucleosomeredistributionsarewidespread,transient,anddna-directed.genomeres24,251-259,doi:10.1101/gr.160150.113(2014).[0581]81sexton,b.s.etal.hierarchicalregulationofthegenome:globalchangesinnucleosomeorganizationpotentiategenomeresponse.oncotarget7,6460-6475,doi:10.18632/oncotarget.6841(2016).[0582]82lunyak,v.v.&rosenfeld,m.g.norestforrest:rest/nrsfregulationofneurogenesis.cell121,499-501,doi:10.1016/j.cell.2005.05.003(2005).[0583]83dirks,p.b.makingacommitment:neuronsrefusecancer'sadvances.natneurosci22,507-508,doi:10.1038/s41593-019-0373-8(2019).[0584]84kaspar,a.a.&reichert,j.m.futuredirectionsforpeptidetherapeuticsdevelopment.drugdiscovtoday18,807-817,doi:10.1016/j.drudis.2013.05.011(2013).[0585]85hamzeh-mivehroud,m.,alizadeh,a.a.,morris,m.b.,bretchurch,w.&dastmalchi,s.phagedisplayasatechnologydeliveringonthepromiseofpeptidedrugdiscovery.drugdiscoverytoday18,1144-1157,doi:10.1016/j.drudis.2013.09.001(2013).[0586]86hoekstra,e.,peppelenbosch,m.p.&fuhler,g.m.meetingreporteurophosphatase2015:phosphatasesasdrugtargetsincancer.cancerresearch76,193-196,doi:10.1158/0008-5472.can-15-2091(2016).[0587]87shi,y.serine/threoninephosphatases:mechanismthroughstructure.cell139,468-484,doi:10.1016/j.cell.2009.10.006(2009).[0588]88sussman,f.,villaverde,m.c.,l.dominguez,j.&danielson,u.h.ontheactivesiteprotonationstateinasparticproteases:implicationsfordrugdesign.currentpharmaceuticaldesign,volume19,number2319,4257-4275,[0589]doi:10.1002/(sici)1520-667x(1998)10:1《19::aid-mcs3》3.0.co;2-1(2013).[0590]89anobom,c.d.etal.fromstructuretocatalysis:recentdevelopmentsinthebiotechnologicalapplicationsoflipases.biomedresearchinternational2014,doi:10.1155/2014/684506(2014).[0591]90blundell,t.l.,jhoti,h.&abell,c.high-throughputcrystallographyforleaddiscoveryindrugdesign.naturereviewsdrugdiscovery1,45-54,doi:10.1038/nrd706(2002).[0592]91weir,m.r.renininhibitors:novelagentsforrenoprotectionorabetterangiotensinreceptorblockerforbloodpressurelowering?currentopinioninnephrologyandhypertension16,416-421,doi:10.1097/mnh.0b013e328209fe00(2007).[0593]92yeo,m.etal.smallctdphosphatasesfunctioninsilencingneuronalgeneexpression.science307,596-600,doi:307/5709/596[pii]10.1126/science.1100801(2005).[0594]93fulda,s.,wick,w.,weller,m.&debatin,k.m.smacagonistssensitizeforapo2l/trail-oranticancerdrug-inducedapoptosisandinduceregressionofmalignantgliomainvivo.naturemedicine8,808-815,doi:10.1038/nm735(2002).[0595]94noguchi,h.etal.anewcell-permeablepeptideallowssuccessfulallogeneicislettransplantationinmice.naturemedicine10,305-309,doi:10.1038/nm994(2004).[0596]95pooga,m.etal.cellulartranslocationofproteinsbytransportan.thefasebjournal:officialpublicationofthefederationofamericansocietiesforexperimentalbiology15,1451-1453,doi:10.1096/fj.00-0780fje(2001).[0597]96schwarze,s.r.,ho,a.,vocero-akbani,a.&dowdy,s.f.invivoproteintransduction:deliveryofabiologicallyactiveproteinintothemouse.science(newyork,n.y.)285,1569-1572,doi:10.1126/science.285.5433.1569(1999).2398(2015).[0661]160a.etal.perineuraldexmedetomidineeffectsonsciaticnerveinrat.brazilianjournalofanesthesiology(englishedition)67,57-66,doi:10.1016/j.bjane.2015.08.012(2017).[0662]161rigaud,m.etal.speciesandstraindifferencesinrodentsciaticnerveanatomy:implicationsforstudiesofneuropathicpain.pain136,188-201,doi:10.1016/j.pain.2008.01.016(2008).[0663]162sleigh,j.n.,weir,g.a.&schiavo,g.asimple,step-by-stepdissectionprotocolfortherapidisolationofmousedorsalrootganglia.bmcresnotes9,82,doi:10.1186/s13104-016-1915-8(2016).[0664]163ronchi,g.etal.discrepanciesinquantitativeassessmentofnormalandregeneratedperipheralnervefibersbetweenlightandelectronmicroscopy.jperiphernervsyst19,224-233,doi:10.1111/jns.12090(2014).[0665]164varejao,a.s.etal.functionalandmorphologicalassessmentofastandardizedratsciaticnervecrushinjurywithanon-serratedclamp.jneurotrauma21,1652-1670,doi:10.1089/neu.2004.21.1652(2004).[0666]165carriel,v.,garzon,i.,alaminos,m.&cornelissen,m.histologicalassessmentinperipheralnervetissueengineering.neuralregenres9,1657-1660,doi:10.4103/1673-5374.141798(2014).[0667]166hazari,a.,wiberg,m.,johansson-ruden,g.,green,c.&terenghi,g.aresorbablenerveconduitasanalternativetonerveautograftinnervegaprepair.brjplastsurg52,653-657,doi:10.1054/bjps.1999.3184(1999).[0668]167shi,t.j.etal.effectofperipheralnerveinjuryondorsalrootganglionneuronsinthec57bl/6jmouse:markedchangesbothincellnumbersandneuropeptideexpression.neuroscience105,249-263(2001).[0669]168jackson,w.m.etal.mesenchymalprogenitorcellsderivedfromtraumatizedmuscleenhanceneuritegrowth.jtissueengregenmed7,443-451,doi:10.1002/term.539(2013).[0670]169kobielaketza,b.k.,grunbergne,kasperce,osbornel,pryorb,tosininl,wux,andersjj.characterizationofmacrophage/microglialactivationandeffectofphotobiomodulationinthesparednerveinjurymodelofneuropathicpain..painmedicine5,932-946,doi:10.1093/pm/pnw144(2016).[0671]170whiteside,g.t.,adedoyina,leventhall.predictivevalidityofanimalpainmodels?acomparisonofthepharmacokinetic-pharmacodynamicrelationshipforpaindrugsinratsandhumans.neuropharm,767-775,doi:10.1016/j.neuropharm.2008.01.001(2008).[0672]171labuda,c.j.&fuchs,p.n.abehavioraltestparadigmtomeasuretheaversivequalityofinflammatoryandneuropathicpaininrats.expneurol163,490-494,doi:10.1006/exnr.2000.7395(2000).behaviourbydopamineinhibition.nature248,257-258(1974).[0687]186spyraki,c.,fibiger,h.c.&phillips,a.g.attenuationofheroinrewardinratsbydisruptionofthemesolimbicdopaminesystem.psychopharmacology79,278-283(1983).[0688]187phillips,a.g.&lepiane,f.g.reinforcingeffectsofmorphinemicroinjectionintotheventraltegmentalarea.pharmacology,biochemistry,andbehavior12,965-968(1980).[0689]188phillips,a.g.,lepiane,f.g.&fibiger,h.c.dopaminergicmediationofrewardproducedbydirectinjectionofenkephalinintotheventraltegmentalareaoftherat.lifesciences33,2505-2511(1983).[0690]189prusaj,j.j.,rosecransja.inmethodsofbehavioranalysisinneuroscience(edbuccafuscojj)ch.4,(crcpress/taylor&francis,2009).[0691]190carr,k.d.,bak,t.h.,simon,e.j.&portoghese,p.s.effectsoftheselectivekappaopioidantagonist,nor-binaltorphimine,onelectrically-elicitedfeedingintherat.lifesciences45,1787-1792(1989).[0692]191galli,r.etal.isolationandcharacterizationoftumorigenic,stem-likeneuralprecursorsfromhumanglioblastoma.cancerres64,7011-7021,doi:10.1158/0008-5472.can-04-1364(2004).[0693]192garzon-muvdi,t.etal.regulationofbraintumordispersalbynkcc1throughanovelroleinfocaladhesionregulation.plosbiol10,e1001320,doi:10.1371/journal.pbio.1001320(2012).[0694]193guerrero-cazares,h.,chaichana,k.l.&quinones-hinojosa,a.neurospherecultureandhumanorganotypicmodeltoevaluatebraintumorstemcells.methodsmolbiol568,73-83,doi:10.1007/978-1-59745-280-9_6(2009).[0695]194tilghman,j.etal.regulationofglioblastomatumor-propagatingcellsbytheintegrinpartnertetraspanincd151.neoplasia18,185-198,doi:10.1016/j.neo.2016.02.003(2016).[0696]195lal,s.etal.animplantableguide-screwsystemforbraintumorstudiesinsmallanimals.jneurosurg92,326-333,doi:10.3171/jns.2000.92.2.0326(2000).[0697]196manton,c.a.etal.inductionofcelldeathbythenovelproteasomeinhibitormarizomibinglioblastomainvitroandinvivo.scirep6,18953,doi:10.1038/srep18953(2016).[0698]197kaplan,e.l.m.,p.nonparametricestimationfromincompleteobservations.journaloftheamericanstatisticalassociation53,457-481,doi:10.2307/2281868(1958).[0699]198marubinie,v.m.c.estimationofsurvivalprobabilities.analysingsurvivaldatafromclinicaltrialsandobservationalstudies.41–81(johnwileyandsons,1995).当前第1页12当前第1页12
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。