一种联合治疗癌症的药物及s100a9-cxcl12信号抑制剂的应用
技术领域
1.本发明涉及癌症治疗技术领域,具体而言,涉及一种联合治疗癌症的药物及s100a9-cxcl12信号抑制剂的应用。
背景技术:
2.乳腺癌是女性最常见的癌症,是全世界女性恶性肿瘤中死亡率最高的原因之一。大约10%的乳腺癌病例是可遗传的,与多个基因的胚系突变有关,包括乳腺癌相关基因1(brca1)、brca2、p53、atm等。其中brca1是重要的相关基因,其原因在于:1)胚系brca1突变约占家族性乳腺癌病例的20%~25%, 2)散发性乳腺癌病例中约1/3brca1表达显著降低,3)brca1缺乏的癌多为三阴性乳腺癌(tnbc; er-、pr-和her2-),由于缺乏有效的治疗手段,在乳腺癌亚型中预后最差。大量研究表明,brca1在许多生物学过程中发挥重要作用,包括转录调控、中心体复制、多个细胞周期检查点的调控、同源重组介导的dna双链断裂(dsb)修复、dna复制、蛋白质稳定和线粒体自噬。
3.由于全身敲除brca1将导致胚胎致死,发明人先前已经构建了携带mmtv-cre介导在乳腺组织中条件特异性的敲除brca1(brca1co/co;mmtv-cre)的突变小鼠模型,发现该突变小鼠模型在长时间后将发生乳腺肿瘤,且p53杂合状态(brca1co/co;p53 /-;mmtv-cre)将加快乳腺癌发生。由于brca1 在hr介导的dsb修复中的重要作用,brca1突变小鼠和人类brca1突变携带者的肿瘤突变负担(tmb) 显著高于brca1高表达的乳腺癌,包括更高的基因突变频率和广泛的基因组改变,包括严重的染色体畸变和非整倍体。研究还表明,brca1缺乏的癌症与肿瘤浸润淋巴细胞(til)的数量增多相关,伴随免疫调节基因的表达增加,包括pdcd1(pd-1)、pd-l1和ctla4。hr缺陷和/或参与免疫检查点的基因表达增加表明brca1缺陷的癌症对icb应具有更高的敏感性,因为这些因素与治疗的反应增加有关。尽管已有上述研究结果,然而,brca1缺陷型乳腺癌对icb的反应与brca1野生型乳腺癌相比并没有显著改善。这些现象表明,决定brca1缺陷型癌症对icb反应性的因素有待进一步阐明。
4.鉴于此,特提出本发明。
技术实现要素:
5.本发明的目的在于提供一种联合治疗癌症的药物及s100a9-cxcl12信号抑制剂的应用从而为 s100a9和/或cxcl12蛋白高表达的癌症提供有效免疫治疗策略。
6.尽管只有在少数癌症患者观察到免疫检查点封锁(icb)具有良好的效果,然而免疫检查点封锁(icb) 仍是癌症治疗的有力手段。基于乳腺癌相关基因1(brca1)缺失导致的乳腺癌对该治疗手段没有显著反应,发明人研究了其潜在机制,分析了brca1突变小鼠模型和人异种移植模型的乳腺和肿瘤组织,从而找出与肿瘤进展和icb反应相关的内在决定因素。
7.发明人发现,brca1缺失激活了s100a9-cxcl12信号通路从而促进了癌症进展,并
触发髓系衍生抑制性细胞(mdscs)的增殖和积累,形成益于肿瘤生长的微环境,从而使肿瘤对icb不敏感。s100a9
‑ꢀ
cxcl12信号抑制剂与pd1抑制剂联合治疗可有效抑制这些致瘤作用。本发明为s100a9和/或cxcl12 蛋白水平升高患者的有效免疫治疗提供了一种新的选择性策略。
8.本发明是这样实现的:
9.本发明提供了一种s100a9-cxcl12信号抑制剂和pd1抑制剂在制备联合治疗癌症药物中的应用。
10.发明人通过cytof(质谱流式细胞术)和数据非依赖质谱技术(dia-ms)对brca1缺失的乳腺组织和肿瘤进行研究。采用这两种方法进一步分析表明,在上皮细胞中brca1负向调控s100a9,brca1缺失时将激活s100a9-cxcl12信号通路,且该信号通路将诱导建立肿瘤微环境,使肿瘤细胞对icb耐受。基于此,发明人发现,可通过s100a9-cxcl12通路抑制剂与pd1抑制剂联合用药来解决,为s100a9和/ 或cxcl12蛋白高表达的乳腺癌提供有效免疫治疗策略。
11.在本发明应用较佳的实施方式中,s100a9-cxcl12信号抑制剂包括不限于:tasquinimod(他喹莫德, cas no.:254964-60-8)、it1t、it1t dihydrochloride、amd 3465、amd 3465 hexahydrobromide和paquinimod (帕奎莫德,cas no.:248282-01-1)。
12.在一种可选的实施方式中,上述s100a9-cxcl12信号抑制剂是通过抑制s100a9表达或抑制cxcl12 表达,从而抑制髓系衍生抑制性细胞(mdscs)的积累以及抑制肿瘤耐受微环境的形成。
13.cxcl12(也称为sdf-1或基质衍生因子-1(stromal derived factor-1))是在许多肿瘤中过表达的趋化因子,其活化位于癌干细胞以及许多免疫细胞的表面上的cxcr4受体。
14.在一种可选的实施方式中,s100a9-cxcl12信号抑制剂包括不限于:tasquinimod和amd 3465。
15.在本发明应用较佳的实施方式中,pd1抑制剂是抗体。
16.在本发明应用较佳的实施方式中,上述抗体选自抗pd-1抗体。
17.在本发明应用较佳的实施方式中,上述抗体包括不限于如下组中的至少一种:纳武单抗(nivolumab)、派姆单抗(pembrolizumab)、皮地利珠单抗(pidilizumab)、兰伯丽珠单抗(lambrolizumab)、bms-936559、阿特珠单抗(atezolizumab)、amp-224、amp224、aunp12、bgb108、mcla134、medi0680、pdrool、 regn2810、shr1210、stial lox、stial llo、tsr042,bms-936558、bgb-a317、bcd-100和js001。
18.在本发明应用较佳的实施方式中,上述癌症为实体肿瘤。
19.在本发明应用较佳的实施方式中,上述实体肿瘤是选自如下的癌症:乳腺癌、胰腺癌、肺癌或肝癌。
20.在其他实施方式中,本领域的技术人员可以根据本发明的构思用于其他实体肿瘤的治疗也均属于本发明的保护范围。
21.其他实体肿瘤包括不限于:
22.鳞状细胞癌、非小细胞肺癌、鳞状非小细胞肺癌(nsclc)、非鳞状nsclc、神经胶质瘤、胃肠癌、肾癌、卵巢癌、结肠直肠癌、子宫内膜癌、肾癌、前列腺癌、甲状腺癌、神经母细胞瘤、成胶质细胞瘤、胃癌、膀胱癌、肝癌、结肠癌、头颈癌、胃癌、生殖细胞瘤、小儿肉瘤、鼻窦
自然杀伤癌、黑色素瘤、皮肤癌、骨癌、宫颈癌、子宫癌、输卵管癌、子宫内膜癌、宫颈癌、阴道癌、外阴癌、肛门区癌、睾丸癌、食管癌、小肠癌、内分泌系统癌、甲状旁腺癌、肾上腺癌、软组织肉瘤、尿道癌、输尿管癌、阴茎癌、肾盂癌、中枢神经系统(cns)肿瘤、原发性cns淋巴瘤、肿瘤血管生成、脊柱肿瘤、脑癌、脑干胶质瘤、垂体腺瘤、卡波西肉瘤、表皮样癌、鳞状细胞癌、儿童期的实体肿瘤、环境诱导的癌症、病毒相关的癌症和病毒起源的癌症。
23.在本发明应用较佳的实施方式中,上述乳腺癌为乳腺癌相关基因1(brca1)缺失所致。
24.在本发明应用较佳的实施方式中,上述癌症为对免疫检查点封锁不敏感的癌症;
25.在一种可选的实施方式中,上述癌症为s100a9和/或cxcl12的转录水平和/或蛋白水平高表达的癌症。上述s100a9和/或cxcl12的转录水平和/或蛋白水平高表达是指,与癌变前的细胞、癌变前的组织或发生乳腺癌相关基因1(brca1)缺失之前相比,细胞、组织中的s100a9和/或cxcl12的转录水平和/或蛋白水平呈现略微、明显或显著地提高。
26.本发明还提供了一种联合治疗癌症的药物组合物,其包括s100a9-cxcl12信号抑制剂和pd1抑制剂。
27.在一种可选的实施方式中,上述癌症选自如下的癌症:乳腺癌、胰腺癌、肺癌或肝癌;
28.在一种可选的实施方式中,上述s100a9-cxcl12信号抑制剂选自如下组中的至少一种:tasquinimod、 it1t、it1t dihydrochloride、amd 3465、amd 3465hexahydrobromide和paquinimod。
29.本发明具有以下有益效果:
30.发明人发现:s100a9-cxcl12信号抑制剂与pd1抑制剂联合治疗可使得癌症对于免疫检查点封锁敏感,这样有助于提高免疫检查点封锁对于癌症治疗的有效性。本发明为s100a9和/或cxcl12蛋白水平升高患者的有效免疫治疗提供了一种新的选择性策略。
附图说明
31.为了更清楚地说明本发明实施例的技术方案,下面将对实施例中所需要使用的附图作简单地介绍,应当理解,以下附图仅示出了本发明的某些实施例,因此不应被看作是对范围的限定,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他相关的附图。
32.图1为brca1缺乏会导致乳腺和肿瘤的免疫抑制的实验验证结果图;
33.图2为s100a9的致癌作用在brca1-mt小鼠和人类乳腺癌患者中的验证结果图;
34.图3为s100a9基因受brca1和s100a9的调控实验结果图;
35.图4为brca1 mt乳腺中的肿瘤免疫耐受微环境实验结果图;
36.图5为s100a9和cxcl12之间的正反馈调控增强brca1-mt上皮细胞中的致癌信号实验结果图;
37.图6为在体内和体外实验验证在mdscs在brca1 mt中的扩增和累积结果图;
38.图7为乳腺癌联合治疗的效果验证结果图;
39.图8为cytof门控策略。
具体实施方式
40.现将详细地提供本发明实施方式的参考,其一个或多个实例描述于下文。提供每一实例作为解释而非限制本发明。实际上,对本领域技术人员而言,显而易见的是,可以对本发明进行多种修改和变化而不背离本发明的范围或精神。例如,作为一个实施方式的部分而说明或描述的特征可以用于另一实施方式中,来产生更进一步的实施方式。
41.除非另有定义,否则本文使用的所有技术和科学术语具有与本公开内容所属领域的普通技术人员通常理解的含义相同的含义。尽管与本文描述的那些方法和材料类似或等同的任何方法和材料都可用于本文的制剂或单位剂量的实践或测试,但现在描述一些方法和材料。除非另有说明,否则本文采用或考虑的技术是标准方法。材料、方法和实例仅是说明性而非限制性的。
42.除非另外指明,否则实践本发明将采用细胞生物学、分子生物学(包含重组技术)、微生物学、生物化学和免疫学的常规技术,所述常规技术在本领域技术人员的能力范围内。文献中充分解释了这种技术,如《分子克隆:实验室手册(molecular cloning:a laboratory manual)》,第二版(sambrook等人,1989);《寡核苷酸合成(oligonucleotide synthesis)》(m.j.gait编,1984);《动物细胞培养(animal cell culture)》(r.i.freshney 编,1987);《酶学方法(methods in enzymology)》(学术出版社有限公司(academic press,inc.);《实验免疫学手册(handbook of experimental immunology)》(d.m.weir和c.c.blackwell编);《哺乳动物细胞用基因转移载体(gene transfer vectors for mammalian cells)》(j.m.miller和m.p.calos编,1987);《当代分子生物学方法(current protocols in molecular biology)》(f.m.ausubel等人编,1987);《pcr:聚合酶链反应(pcr:thepolymerase chain reaction)》(mullis等人编,1994);以及《当代免疫学方法(current protocols in immunology)》 (j.e.coligan等人编,1991),所述文献中的每个文献均通过引用明确并入本文中。
43.为使本发明实施例的目的、技术方案和优点更加清楚,下面将对本发明实施例中的技术方案进行清楚、完整地描述。实施例中未注明具体条件者,按照常规条件或制造商建议的条件进行。所用试剂或仪器未注明生产厂商者,均为可以通过市售购买获得的常规产品。
44.以下结合实施例对本发明的特征和性能作进一步的详细描述。
45.如下实施例中所采用的实验方法如下:
46.为了找出决定brca1缺陷型肿瘤对icb的反应性的内在因素,发明人采用了两种方法,即cytof (质谱流式细胞术),和数据非依赖型质谱(dia-ms),以分析brca1缺陷的乳腺组织和来自brca1突变小鼠模型和人类异种移植模型的肿瘤。潜在的候选基因通过功能性方法进行验证,包括基因过表达和 crispr-cas9介导的敲除,然后是化学抑制剂与pd-1抗体相结合的癌症治疗。
47.(1)肿瘤模型和治疗方法
48.本研究中进行的所有动物工作均经澳门大学动物伦理委员会批准。动物在特定的无病原体(spf)动物设施中保持在12小时光照/12小时黑暗循环下,可免费获得水和标准小鼠饮食。室温保持在23
±
3℃范围内。湿度水平保持在40-70%之间。所有动物研究均在雌性小鼠身上进行,处理在戊巴比妥钠麻醉下进行。 brca1条件性敲除小鼠模型(brca1co/co;
mmtv-cre)在发明人的实验室中建立并已在之前进行了描述(x. xu et al.,conditional mutation of brca1 in mammary epithelial cells results in blunted ductal morphogenesis andtumour formation.nat genet 22,37-43(1999))。为了建立肿瘤同种异体移植物,构建了brca1 wt(b477)乳腺上皮细胞系和brca1-mt乳腺将上皮(g600)细胞(2x105细胞)植入裸鼠的右侧乳腺脂肪垫。将 emt6、sgs100a9-emt6和sgcxcl12-emt6细胞(1x106细胞)植入balb/c小鼠的右侧乳腺脂肪垫中。将lmbg(545)细胞(4x106细胞)植入fvb小鼠的右侧乳腺脂肪垫中。从第7天开始,每隔一天测量一次肿瘤(长度和宽度,单位为毫米)。根据澳门大学批准的动物方案,在实验结束时收集肿瘤样本。根据以下公式计算肿瘤体积(以mm3为单位):v=0.5
×
h2×
l,其中h是较短的直径,l是较长的直径。每周测量肿瘤3次,在肿瘤直径小于或约2厘米之前采集样本,这是澳门大学动物伦理委员会允许的最大肿瘤大小/负荷。为了分析血液和乳腺组织中的mdsc,从6个月大的wt和brca1co/co; mmtv-cre小鼠或移植肿瘤大小达到0.5厘米的荷瘤裸鼠中收集两种组织的免疫细胞.对于 brca1co/co;mmtv-cre小鼠中肿瘤生长的药物治疗,发明人首先在肿瘤达到1厘米大小时切除原发性肿瘤,然后用药物治疗小鼠。对于lmbg-fvb小鼠,药物在注射lmbg细胞系后一天给药。在切除原发性肿瘤或注射lmbg细胞后,将小鼠随机分为对照组和药物治疗组。对照组小鼠腹腔注射pbs。对于apd1 单药治疗组,apd1在术后第二天和第一次抗体注射后三周腹腔注射两次(0.2mg/小鼠)。amd3465单药治疗组隔日皮下注射(0.2mg/小鼠)。对于tas单药治疗组,每天口服tas(一种s100a9抑制剂)(0.2mg/ 小鼠)。对于联合治疗,pd1抗体从术后第二天开始每21天腹腔注射一次,每隔一天注射一次amd3465 (0.2mg/小鼠)和/或每天口服tas(0.2mg/小鼠)。
49.质谱流式细胞术(cytof)
50.将来自小鼠的wtmg、mtmg、adj.mg和肿瘤组织进行消化得到单细胞,具体步骤为,将乳腺和肿瘤组织切成小块,用含有5%fbs、5μg/ml胰岛素(i-1882;sigma-aldrich)、500ng/ml氢化可的松(h0888; sigma-aldrich)的消化培养基消化,10ng/ml egf(13247-051,invitrogen),iii 300u/ml胶原酶(s4m7602s, worthington,lakewood,nj)和100u/ml透明质酸酶(h3506;sigma-aldrich)在37℃下2小时.然后在用hpss缓冲液洗涤一次后,在含有5mg/ml分散酶ii(10295825001;roche diagnostics,indianapolis, in)和脱氧核糖核酸酶(58c10349;worthington,lakewood,nj)的培养基中继续消化样品5分钟。 (14170-112;bio-tech)。然后,在用hbss缓冲液洗涤后,组织消化物通过40μm过滤器进入预热的无血清培养基中。所有单细胞悬液在室温(rt)下通过低渗裂解2分钟去除红细胞。将细胞在预热的无血清培养基中重悬至2x107个细胞/ml的密度,并向细胞悬液中加入等体积的10μm顺铂工作溶液(顺铂的最终浓度:5mm)。将单细胞重新悬浮并在室温下孵育5分钟。然后,加入5倍体积的含染色细胞血清的培养基以淬灭顺铂染色,并将细胞以300x g离心5分钟。然后用1ml maxpar cell staining buffer 洗涤细胞一次,将300万个细胞悬浮在80μl maxpar cell staining buffer中。将悬浮的细胞加入到20μl 的fc阻断溶液中。将细胞在4℃下孵育10分钟,然后重悬于50μl maxpar细胞染色缓冲液中。向每个试管中加入50微升抗体混合物,轻轻涡旋后,将细胞在室温下孵育30分钟。然后用1ml maxpar cellstaining buffer洗涤细胞两次,并加入1ml细胞嵌入溶液(用maxpar fix和perm buffer稀释的cell-idintercalator-ir)。将样品充分混合并在4℃下放置过夜。第二天,细胞用1ml maxpar细胞
5分钟内5-8%b,5-20分钟内8-18%b,20-70分钟内18-40%b,70-72分钟内40-95%b。馏分收集:从洗脱后的第一分钟到72分钟结束,将不同时间收集的馏分合并成12个组分,45℃真空干燥。
61.2.2数据依赖性采集
62.将上述12种馏分溶解在样品溶液(0.1%fa,2%acn)中并加载到连接到电喷雾电离质谱仪/esi质谱仪(orbitrap eclipse mass spectrometer-thermo fisher scientific-usa)的nano lc上,以采用dda模型。具体步骤和参数如下:流动相流速为600nl/min,a相(0.1%fa)和b相(0.1%fa,80%acn)。将样品加载到柱子上(150μm i.d.
×
150mm,填充有acclaim pepmap rplc c18,1.9μm,),每个馏分分析120分钟。分离得到的多肽直接进入质谱仪进行在线检测。ms1扫描是在350-1500m/z的范围内以70,000的分辨率采集的,然后自动增益控制(agc)设置为4.0e5,最大进样时间为50ms。对于ms2扫描,采集范围为200-2000m/z,分辨率为17,500,随后agc设置为3.0e5,最大进样时间为72 ms。归一化碰撞能量(nce)为27%,激活类型-hcd(碰撞能量35%)。将15个样品(每个5μg)的肽溶解在样品溶液(0.1%fa,2%acn)中,并在dia模型中采集。每个样品以一式两份的技术重复上样。具体参数如下:ms1扫描是在400-1210m/z的范围内以120,000的分辨率采集的,然后是设置为 4.0e5的agc,最大进样时间为50ms。对于ms2扫描,采集范围为200-2000m/z,分辨率为17,500,随后agc设置为3.0e5,最大进样时间为72ms。nce为27%,激活类型-hcd(碰撞能量35%)。
63.4.特定数据库的建立
64.将上述12个dda ms生成的文件导入proteome discoverer(2.4版thermo scientific)进行检索和鉴定。检索参数如下:固定修饰:氨基甲酰(c);可变修饰:氧化(m);酶:胰蛋白酶;最大丢失数:2;肽质量分辨率:20ppm;碎片质量:0.6da;质量值:单同位素;显着性阈值:0.05。
65.5.dia分析
66.使用spectronaut软件对dia原始数据进行分析,详细参数如下:肽段长度:7-30;酶/裂解规则:胰蛋白酶/p;消化类型:特异;丢失数:2;碎片离子m/z:400-1210;最大变量修饰:5;固定修饰:氨基甲酰(c);可变修饰:乙酰基(蛋白质n端),氧化(m)psm-fdr:0.01;肽fdr:0.01;蛋白质组fdr: 0.01;前体qvalue截止值:0.01;蛋白质qvalue截止值(运行):0.05。
67.细胞系,培养条件和病毒感染条件如下:
68.wt(b477)乳腺上皮细胞系来源于brca1-wt小鼠的乳腺,而brca1-mt(g600)乳腺上皮细胞系来源于brca1-mt小鼠(11).mda-mb-231、hek293t、emt6、raw264.7细胞系来自atcc。本研究中的所有细胞系均在补充有10%胎牛血清(fbs)(sigma,st.louis,mo)、10μg/ml胰岛素(invitrogen, carlsbad,ca)、1%l-谷氨酰胺(life technologies)和0.6%pen-strep(life technologies)。emt6细胞用补充有10%fbs、10μg/ml胰岛素、1%l-谷氨酰胺和0.6%pen-strep的rpmi 1640(life technologies, carlsbad,ca)一起培养。对于慢病毒转导,用pspax2(3μg)或pmd2.g(9μg)和转移质粒(12μg) 转染293t细胞。加入包装好的病毒72小时,用嘌呤霉素筛选转染细胞。用lipofectamine 3000转染试剂将brca1 cdna和shrna质粒转染到细胞中。本文中使用的sgrna 序列见表1。
prjna719246[https://www.ncbi.nlm.nih.gov/bioproject/prjna719246/]和prjna719077 [https://www.ncbi.nlm.nih.gov/bioproject/prjna719077/]。dia-ms蛋白质组学数据已保存在proteomicsidentifications数据库(pride)中,访问号如下:pxd030328(dia原始数据) [http://www.ebi.ac.uk/pride/archive/projects/pxd030328]和pxd030355(dda原始数据) [http://www.ebi.ac.uk/pride/archive/projects/pxd030355]。使用了来自癌症基因组图谱 (tcga-brca.samplemap/hiseqv2)数据库和gse19783-gpl6480数据集的乳腺癌数据。本发明提供了源数据。其余数据可在文章、补充信息或源数据文件中找到。
[0093]
统计分析
[0094]
为了量化肿瘤细胞或免疫细胞密度,使用imagej软件捕获并分析了if染色的肿瘤切片图像。使用 graphpad prism 8.0进行统计分析。未配对的双尾学生t检验用于比较两组之间的差异。对于多组之间的比较,使用单向方差分析或双向方差分析。用双尾皮尔逊检验分析相关性。统计信息,包括平均值和统计显着性值,在文本或图形图例中显示。在图表上,条形代表范围或标准偏差(sd)。通过kaplan-meier分析确定生存预后。通过在线patternhunter分析(https://www.metaboanalyst.ca/faces/home.xhtml)dia-ms 数据,并通过pearson r》0.5过滤基因,调整p值《0.05。rna测序数据的上调和下调基因的过滤阈值分别为》1.2或《0.83的倍数变化,p值《0.05。
[0095]
研究批准:根据机构指南,所有小鼠品系均保存在澳门大学健康科学学院的动物实验中心。所有实验均经澳门大学健康科学学院动物伦理委员会批准(协议id:umare-amend-100)。
[0096]
实施例1
[0097]
本实施例论证了brca1缺失将导致mdscs增加,而产生的肿瘤耐受微环境促进癌症的形成。
[0098]
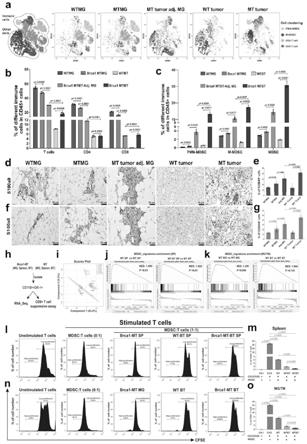
为了探讨brca1缺失是否能诱导免疫应答缺陷,发明人利用brca1co/co;mmtv-cre小鼠模型 (gemm)(x.xu et al.,conditional mutation of brca1 in mammary epithelial cells results in blunted ductalmorphogenesis and tumour formation.nat genet 22,37-43(1999)),对野生型(wt)乳腺组织(wtmg)、brca1 突变(mt)乳腺组织(mtmg)、肿瘤邻近乳腺组织(tu-adj.mg)、野生型肿瘤(wt tumor)和brca1-mt 肿瘤进行免疫分析(结果参照附图1中a-e)。利用cytof抗体:cd45、cd3e、cd4、cd8a、cd11b、 ly6g和ly6c(附图1中f),基于cytobank-tsne分析揭示了基于多个单细胞数据集的复杂云图(图1中a 所示)。
[0099]
图1中a所示,tsne分析来自野生型乳腺组织(wtmg,n=6只小鼠)、brca1突变乳腺组织(mtmg, n=6只小鼠)、癌旁组织(mt肿瘤adj.mg,n=3只小鼠)、野生型肿瘤和brca1突变型肿瘤组织的免疫细胞种类。
[0100]
研究发现,与wtmg组织相比,t细胞数量在mtmg中(包括cd4 t和cd8 t细胞)有所减少,但在tu-adj和肿瘤组织中均显著减少(图1中b)。相反,相对于wtmg组织,在mtmg组织中mdsc 细胞群有一定程度的增加,mdsc细胞群包括多核mdscs(pmn-mdscs)和单核mdscs(m-mdscs);而mdsc细胞群在brca1co/co;mmtv-cre小鼠的tu-adj mg和肿瘤组织中大幅度增多,但在野生型肿瘤组织中却增加不多(参照图1中c所示)。
mt荷瘤小鼠的乳腺肿瘤组织的 mdsc(brca1-mtbt)。mdsc与t细胞的比例为1:1(n=3只小鼠/组)。
[0108]
实施例2
[0109]
本实施例提出并验证了s100a9是brca1突变型乳腺组织中的关键潜在诱导因子。
[0110]
为了鉴定诱导肿瘤耐受微环境的潜在因子,发明人采用数据非依赖型质谱法(dia-ms)。
[0111]
对8-10个月野生型(wtmg)、brca1突变型乳腺组织(mtmg)、癌旁乳腺组织(tumor-associatedmg)、野生型肿瘤(wtbt)和brca1突变型乳腺肿瘤组织(图2中a)进行质谱检测pca分析表明,来源相同的组织可以聚类,且可区分来源不同的组织(图2中b)。
[0112]
图2中a为dia-ms分析肿瘤发生过程中乳腺组织的工作流程,包括来自8-10个月野生型(wtmg)和 brca1-mt(mtmg)小鼠的乳腺组织、癌旁乳腺组织(tu-adj.mg),和来自野生型(wtbt)和brca1-mt(mtbt) 小鼠的肿瘤(n=3只小鼠/每个基因型)。
[0113]
为了验证数据的重现性,发明人考察了dia-ms数据采集中重复样品的变异系数(cv)%。发现重复蛋白的变异系数小于20%,cv%的中位数在0.8-1.21之间(附图2中a),同时pearson相关系数在0.90以上(图 2中c),表明dia-ms在dia蛋白质组数据的定量方面具有很高的重现性。为了找到brca1突变诱导的潜在关键因子,发明人采用patternhunter分析方法(皮尔逊相关系数r》0.5,校正p值《0.05)对dia-ms数据进行了分析(图2中d,附图2中b),从而鉴定到了632显著差异蛋白符合在brca1突变诱导的肿瘤形成过程中逐渐升高的特性。同时,分析brca1野生型和突变型肿瘤组织,发现725个蛋白显著上调(fc》2,pvalue《0.05) (附图2中b)。
[0114]
图2中d为肿瘤发生过程中s100a9蛋白水平增加的模式(a)来自spectronaut的dia数据分析和pearson 相关性的统计分析(fdr《1,r=0.8343,调整p值=0.024。原始数据由log10转换,箱线图和须线图总结了归一化值,线显示sd)。数据来自n=3个生物学独立的重复。
[0115]
然后,发明人用以上得来的632个和725个蛋白与45个来自人数据库的brca1负调控的蛋白进行venny 分析(图2e-g和附图2b),得到s100a9(1,2),s100a8,pglyrp1和colgalt1四个共有基因。
[0116]
其中,图2中e为来自癌症基因组图谱(tcga)数据库(http://www.linkedomics.org/admin.php)的临床蛋白质组学肿瘤分析联盟(cptac)火山图的brca1和s100a9在蛋白质水平上的pearson相关性。f为e中与brca1在蛋白质水平上呈负相关的前50个基因的热图。g为来自dia分析的小鼠候选基因列表与受 brca1基因负调控的前45个基因重叠结果图。
[0117]
这里,发明人选择s100a8和s100a9作为首选研究基因,因为先前的cytof数据揭示了他们在肿瘤微环境(tme)中的作用。由于s100a8基因敲除导致小鼠胚胎早期死亡,并且s100a9-/-小鼠在侵袭性肺炎球菌肺炎(32只)中的中性粒细胞募集过程中发挥重要作用,发明人推测s100a9蛋白可能在介导肿瘤细胞与tme之间的动态相互作用中发挥重要作用。与上述分析一致,所有5组均鉴定出s100a9肽(附图2 中c),且鉴定出的s100a9肽段数量随brca1突变诱导的肿瘤发育阶段而增加(附图2中d)。在对人类数据库的分析中,发明人发现s100a9表达水平与人类乳腺癌的发育阶段呈正相关(附图2中e),pearson相关分析揭示乳
腺癌患者brca1与s100a9呈负相关(附图2中f)。
[0118]
然后发明人用两对brca1突变和野生型的人源肿瘤异种移植模型进行了体内验证,发现s100a9阳性细胞在brca1突变模型中显著高于野生型(图2中h-2i)。相一致的是,s100a9的高水平表达对乳腺癌患者的预后较差(图2中j)。图2中h为s100a9抗体对brca1-wt和brca1-mt乳腺癌组织染色的代表性图像(来自pdx模型的每组n=2)从jacksonlaboratory获得。i为(h)中每个视野s100a9阳性细胞的量化。
[0119]
接下来,发明人用brca1co/co;mmtv-cre模型鼠进行了进一步验证,发现s100a9的mrna和蛋白表达水平均在brca1突变组织中显著增加(图2中k-n)。同时免疫荧光显示,s100a9蛋白在上皮细胞和m2巨噬细胞中均高表达(附图2中g-2h)。
[0120]
具体地,图2中j为基于kaplan-meierplotter网站 (https://kmplot.com/analysis/index.php?p=service&cancer=breast)的乳腺癌患者(n=4934)的s100a9表达的生存结果。k-l为通过qpcr(n=3只小鼠/组)揭示的4个月brca1-wt和mt小鼠(k)和brca1mt和wt乳腺肿瘤(l)中乳腺组织s100a9和s100a8的相对表达)。m-n为在4个月大的乳腺组织(m)和brca1mt和wt肿瘤(n)中s100a9和s100a8的蛋白质水平通过蛋白质印迹测定结果图(n=3只小鼠/组)。
[0121]
以上数据表明,s100a9是形成肿瘤微环境从而诱导乳腺癌进展的关键潜在因素。
[0122]
实施例3
[0123]
本实施例论证了s100a9的表达受brca1的负向调控和自身的正向调控。
[0124]
由于s100a9的mrna和蛋白水平在brca1突变型小鼠的癌前乳腺和肿瘤组织中均高表达,因此发明人考察s100a9的表达是否受到brca1的负向调控。
[0125]
研究发现,s100a9和s100a8的mrna水平在brca1突变细胞系(g600)中比野生型乳腺上皮细胞(b477) 分别高4倍和5倍(图3中a),s100a9/s100a8的表达剂量依赖性brca1表达的降低(图3中b)。相反,当过表达brca1时,s100a9/s100a8mrna表达降低到原有水平的20%和30%(图3中c)。同时在mda-mb-231(231) 人基底型乳腺癌细胞系中也得到同样的结果(图3中d),即brca1负调控s100a9。图3中d为s100a9 和s100a8在mda-mb-231(231)对照和231细胞中表达shbrca1(敲低brca1)的表达量统计图。
[0126]
为了进一步研究s100a9是否在转录水平受brca1调控,发明人对前期(x.xu et al.,brca1 repressesdnareplication initiation through antagonizing estrogen signaling and maintains genome stability in parallelwith wee1-mcm2 signaling during pregnancy.hum mol genet 28,842-857(2019))构建的brca1-mt(g600) 和wt(b477)乳腺上皮细胞系进行了荧光素酶(luc)报告基因检测(附图3中e)。然后在不同条件下测定启动子活性。数据显示,仅用s100a9启动子-luc转染brca1-mt细胞72h后,luc活性分别提高3.8倍和1.5倍(图3中e)。将brca1在野生型和突变型细胞中过表达时,s100a9-luc活性分别下调为40%和60%(图 3中e),表明brca1对s100a9启动子具有抑制作用。由于s100a9蛋白能够激活和招募mdscs,而活化的 mdscs能够分泌s100a9,所以s100a9可通过正反馈调节环来调控自身的表达。
[0127]
同时发现,过表达s100a9,luc活性在brca1-mt细胞中进一步增加5倍,wt中3倍(图3中e)。通过对2kbs100a9启动子的系列连续缩短,确定该启动子的功能域在-500~-700碱基对范围内(附图3中f)。同时在蛋白水平上,通过在b477和231细胞系上敲低brca1,通过免
疫荧光(if)和westernblot分析发现 brca1可负调控s100a9(图3中f)。
[0128]
综上所述,这些数据表明s100a9在转录水平和蛋白水平的表达均受到brca1的负调控,而s100a9 可通过自身的启动子区进行自身正调控。
[0129]
实施例4
[0130]
本实施例论证了s100a9增加诱导的肿瘤免疫微环境可在brca1-mt早期乳腺组织中形成。
[0131]
据报道,升高的s100a9水平对骨髓细胞具有多效性,并可阻止这些细胞的正常分化(p.cheng et al., inhibition of dendritic cell differentiation and accumulation of myeloid-derived suppressor cells in cancer isregulated by s100a9 protein.j exp med 205,2235-2249(2008))。因此,发明人假设brca1-mt乳腺上皮组织中s100a9水平升高可能会逐渐诱导免疫抑制环境的形成并促进乳腺组织中肿瘤的发生和进展。
[0132]
为了验证这一假设,发明人首先研究了s100a9及其伴侣蛋白s100a8在乳腺不同细胞亚群、不同发育阶段和肿瘤组织中的表达。发明人发现s100a8/s100a9的mrna水平在多个发育阶段的mtmg组织和肿瘤组织中增加(附图4中a-4c),同时s100a9mrna在管腔和基质亚群中(图4中a和附图4中d),以及 brca1-mt乳腺上皮细胞系(g600)中显著增加(图3中a)。与mrna表达增加一致(附图4中a-4c和图3a), s100a8/s100a9蛋白水平在mt乳腺上皮细胞系(g600)、乳腺组织和肿瘤组织中也显著升高(图4中b和图2m-2n)。通过对多个发育阶段的乳腺组织进行ck18和s100a9的免疫共染色发现,s100a9蛋白水平在癌旁组织和肿瘤组织也显著增加(图4中c和补充图4中e),表明brca1的缺失将诱导s100a9在mrna 和蛋白质水平的表达。
[0133]
图4中a为s100a9/s100a8的mrna在wt4个月大的乳腺(wtv4mg)和mt4个月大的乳腺(mtv4mg) 的腔和基质细胞亚群中的表达(n=3只小鼠)。b为通过蛋白质印迹法检测wt(b477)和mt(g600)乳腺上皮细胞系和肿瘤组织中s100a9的蛋白质水平(n=3个体实验向上和n=3小鼠向下)。c为s100a9(红色)和 ck18(绿色)与wtv4mg、mtv4mg、wtv6mg和mtv6mg组织上的抗体共染色(每组n=3对)。d 为分别在4个月和6个月时对wt和mt小鼠的血液和乳腺组织进行facs分析的s100a9和arg1阳性细胞群(facs门控策略见补充图8c,n=3只小鼠/组)。e为通过if(40x共聚焦显微镜)与s100a9(红色) 和cd206(绿色)抗体(左图)共染色以及与s100a9(红色)和ck18(绿色)抗体(右图)在肿瘤邻近组织上共染色,比例尺:20μm。)(n=3只小鼠和3个单独的实验)。
[0134]
s100a9是一种趋化因子。因此,为了研究s100a9阳性细胞在mtmg组织中数量是否增加,发明人对血液和乳腺组织进行了流式细胞术(facs)分析。数据显示,与wt小鼠相比,4个月大的mt小鼠血液和乳腺中的s100a9阳性细胞分别略有增加。值得注意的是,与wt小鼠相比,6个月大的mt小鼠血液中 s100a9阳性细胞群的增加达到30%,乳腺中达到46%(图4中d)。与wt小鼠相比,在4/6个月大的mt 小鼠的血液和乳腺组织中,也观察到arg1(mdsc的另一种标志物)的类似增加的表达模式(图4中d)。这些发现表明,乳腺上皮细胞中s100a9蛋白水平升高可促使额外的mdsc向mt乳腺组织募集,从而抑制细胞毒性t细胞的增殖和功能。
[0135]
为了进一步评估brca1-mt小鼠乳腺组织中的tme,发明人进行了if染色和蛋白质印迹分析。发明人发现1)s100a9阳性mdsc侵入肿瘤邻近的乳腺组织(图4中e);2)s100a9蛋
白由mdscs分泌到brca1-mt 小鼠的肿瘤邻近乳腺组织和体外培养细胞的上清液中(图4中f);3)tgf-β和il-10的蛋白质水平在4个月的乳腺组织中均增加,这可以触发s100a9迁移至细胞膜和细胞表面以促进s100a9蛋白分泌(图4中 g-h)。
[0136]
图4中f为从邻近肿瘤的乳腺(n=3只小鼠)中的肿瘤细胞和mdsc细胞分泌s100a9蛋白(左)并存在于培养的癌细胞的上清液中(右)(n=3)。g为wt和brca1mt小鼠4个月时(n=3只小鼠)乳腺组织中s100a9、tgf-β和il-10的蛋白质水平。h为wt和brca1mt小鼠6个月时(n=3只小鼠)乳腺组织中 s100a9、tgf-β和il-10的蛋白质水平。
[0137]
这些数据表明,brca1-mt乳腺上皮细胞中升高的s100a9信号传导可以作为导致mdsc积累的内在因素,并在肿瘤形成之前很久就在brca1-mt乳腺组织中建立有利于肿瘤发生的肿瘤允许微环境。
[0138]
实施例5
[0139]
本实施例论证了s100a9和cxcl12之间的正反馈调节能放大brca1-mt上皮细胞中的致癌信号。
[0140]
为了了解s100a9在brca1-mt乳腺上皮细胞中的致癌作用,发明人首先对四种具有s100a9差异表达的乳腺上皮细胞系进行了rna测序,包括wt(b477-ctr)、过表达s100a’9(b477-oe-s100a9)、brca1-mt (g600-ctr)和敲除s100a9(sgs100a9)-brca1-mt细胞系(g600-sgs100a9)(附图s5b-s5d)。通过 b477-oe-s100a9与b477-ctr细胞相比,以及g600-sgs100a9与g600-ctr相比,453个共有基因上调在 b477-oe-s100a9,下调在g600-sgs100a9(图5中a)。
[0141]
图5中a为g600中上调基因、b477细胞中过表达-s100a9(b477-oe-s100a9)和g600细胞中表达 sgs100a9(g600-sgs100a9)中下调基因的维恩图分析。
[0142]
对这453个基因功能的kegg通路分析表明,细胞因子-细胞因子受体相互作用通路是前10种激活的致癌通路之一(图5中b)。
[0143]
值得注意的是,cxcl12,也称为基质细胞衍生因子1(sdf1),与这些途径相关,在453个基因中排名第一,就其相对于对照水平的倍数变化而言(图5中c和附图5中e)。
[0144]
图5中c为通过比较四种不同组细胞的基因表达谱,获得前20个差异表达基因,包括b477-ctr(b1-3)、 b477-oes100a9(p1-3)、g600-ctr(g1-3)和g600-sgs100a9(g10-1-3)。
[0145]
接下来,发明人想探索cxcl12是否可以在brca1-mt上皮细胞中介导s100a9信号。发明人首先检查了b477和g600细胞中10种细胞因子的表达,发现只有s100a9和cxcl12在brca1-mt细胞中上调(附图5中a)。该数据表明cxcl12可能是brca1-s100a9调节的主要细胞因子。然后,发明人进一步研究了b477 中过表达s100a9(b477-oe-s100a9)中的cxcl12表达和g600细胞系中敲除s100a9(g600-sgs100a9)(附图5 中b-d),数据显示cxcl12的mrna和蛋白质水平在b477-oe-s100a9细胞中增加,但在g600-sgs100a9 中降低(图5中d-e),证实s100a9的表达可以影响brca1-mt乳腺上皮细胞中cxcl12的表达。
[0146]
由于显示cxcl12可以诱导stat3的磷酸化,因此,发明人探究s100a9是否通过cxcl12调节pstat3的表达。发现当s100a9在emt6细胞中过表达时,cxcl12和pstat3的蛋白质水平增加,并且这种增加可以被sgs100a9或sgcxcl12抑制在emt6-oe-s100a9和b477-oe-s100a9中(图5中f-h),在231和545细胞系中也观察到了同样的结果(附图5中f-h)。
[0147]
用已知的s100a9抑制剂tasquinimod(tas)进一步验证发现,545和231细胞中抑制
s100a9,cxcl12的 mrna水平降低(附图5中i-j),在g600细胞系中s100a9、cxcl12和pstat3的蛋白质水平也降低(图5 中g和5m),这证实了cxcl12可介导s100a9对pstat3的作用。
[0148]
图5中g为s100a9抑制剂tasquinimod(tas-50μm)对s100a9、cxcl12和pstat3的蛋白质水平的作用结果图。h为蛋白质印迹检测oe-s100a9、oe-s100a9/sgcxcl12或oe-cxcl12的wt(b477)细胞中s100a9、 cxcl12和pstat3的蛋白水平结果图。i,k分别为不同时间过程(0-48小时)和不同剂量下(0ug-4ug)在 b477(i)细胞系中oe-s100a9和sgs100a9在g600(k)细胞中cxcl12的蛋白质水平变化结果图。j,l分别为不同时间进程(0-48小时)和不同剂量(0ug-4ug)的b477(j)和sgcxcl12g600(l)细胞中s100a9 蛋白水平的变化结果图。
[0149]
据报道,s100a9通过其晚期糖基化终产物(rage)的同源受体发挥作用(k.narumi et al., proinflammatory proteins s100a8/s100a9 activate nk cells via interaction with rage.j immunol 194, 5539-5548(2015))。为了检查s100a9诱导的增加的pstat3是否由rage受体介导,发明人用rage抑制剂fps-zm1处理brca1-mt细胞,并检测到s100a9、cxcl12和pstat3的蛋白质水平没有明显变化(图5 中m),表明s100a9对cxcl12/pstat3的调节与rage无关。
[0150]
已有研究表明,除了rage之外,s100a9的受体还包括tlr4、cd36和emmpirin。
[0151]
值得注意的是,发明人的数据还显示,与b477对照组相比,cxc12的敲除组中s100a9也降低了,而 s100a9的蛋白质水平在cxcl12的过表达中有所增加(图5中h;附图5中f),且在b477和231细胞中均观察到cxcl12过表达可诱导s100a9水平的增加(图5中j;附图5中l),表明s100a9和cxcl12之间可能存在正反馈调节。为了进一步证明此正反馈调节回路,发明人进行了时间过程和剂量依赖性实验,以检测b477、g600和mda-mb-231细胞系中cxcl12和s100a9的动态变化。数据表明b477和mda-mb-231 细胞中s100a9或cxcl12的过表达(oe)可以增加cxcl12和s100a9的蛋白质水平(图5中i、5j和附图5 中k、5l)。相反,在g600中sgs100a9和sgcxcl12的表达分别降低了cxcl12和s100a9的蛋白质水平(图 5中k,5l),表明升高的s100a9诱导的致癌信号可以通过s100a9和cxcl12之间的正反馈调节环进一步放大肿瘤发生的概率。为了进一步证明s100a9-cxcl12相互作用与brca1的缺失有关,发明人在wt细胞系中敲低brca1并在brca1-mt细胞系中过表达brca1,发现s100a9、cxcl12和pstat3的蛋白质水平在 shbrca1-b477细胞中增加,在oe-mbrca1-g600细胞中降低(图5中n)。
[0152]
总之,发明人的数据表明,由brca1缺陷诱导的s100a9的致癌激活可以上调cxcl12并激活pstat3,上调的cxcl12蛋白可以进一步放大brca1-mt乳腺上皮细胞中的s100a9致癌信号,进而可通过旁分泌方式产生了肿瘤免疫抑制微环境(图5o),然而,这种机制需要进一步验证和阐明。
[0153]
实施例6
[0154]
本实施例论证了s100a9/cxcl12信号轴从上皮细胞到免疫细胞。
[0155]
为了检查来自brca1-mt乳腺上皮的s100a9/cxcl12信号是否会影响mdsc,发明人首先检查了来自 wt和brca1-mt小鼠的mdsc细胞在不同条件培养基中的作用(图6中a-c)。
[0156]
图6中a-c为条件培养基(cm)中来自wt(a)和brca1-mt(b)小鼠脾脏的mdscs迁移测定,包括来自 b477细胞(ctr-b477)的cms、b477细胞中的oe-s100a9(oe-s100a9-b477)、g600细胞(ctr-g600)和表达 s100a9的g600细胞(sg-s100a9-g600)。
[0157]
数据显示:
[0158]
1)来自具有oe-s100a9的b477细胞的条件培养基中的mdsc比仅来自b477细胞的对照培养基中的 mdsc通过膜迁移多,但来自brca1-mt的mdsc比来自wt小鼠的mdsc迁移更多;
[0159]
2)尽管来自g600细胞的条件培养基中在wt和brca1-mt小鼠中观察到与上述相同的迁移模式,但是更多的招募来自从brca1-mt小鼠分离的mdscs;
[0160]
3)如果条件培养基来自g600细胞表达sgs100a9,迁移细胞的数量显著减少,表明不同基因型细胞分泌的s100a9蛋白不仅对mdscs的迁移有影响,而且对brca1-mt小鼠的mdscs有更强的影响。
[0161]
为了研究s100a9/cxcl12信号如何影响髓系细胞,发明人用各种类型的条件培养基(cm)培养raw264.7 细胞系(一种鼠白血病单核细胞/巨噬细胞系)的细胞48小时并监测增殖,迁移和对巨噬细胞分子的作用。虽然来自wt乳腺上皮细胞(b477)的cm几乎没有影响,但来自mt乳腺上皮细胞(g600)的cm显着增加了raw264.7细胞的集落大小和迁移。这种效应主要是由s100a9引起的,因为敲除s100a9在g600细胞系中,cm诱导的集落大小和迁移明显减少,而在b477细胞中过表达s100a9而增加(图6中d-g)。在用敲低brca1的b477的培养基处理raw264.7细胞,也观察到raw264.7细胞的集落大小和迁移增加,而过表达brca1却减少(附图6中a-c)。
[0162]
与这些观察结果一致,在b477过表达的cm和g600的cm处理raw264.7细胞,负责巨噬细胞增殖(cyclind1)、扩增(pstat3)和免疫抑制(arg1)的分子和cxcl12的蛋白水平均有增加。但在来自 g600-sgs100a9细胞的cm培养的raw264.7细胞中减少(图6中h)。同时用添加了s100a9重组蛋白的 b477-cm处理raw264.7,以上这些分子均有增加,而添加s100a9抑制剂-tas可抑制这些分子的表达(图 6中i)。使用emt6细胞的cm观察到类似的结果(图6中j)。
[0163]
图6中i-j为raw264.7细胞中s100a9、cxcl12、pstat3和arg1的蛋白水平结果图,用来自b477(i) 或emt6(j)细胞系的cms与s100a9蛋白(0.1mg/ml)或s100a9蛋白与tas抑制剂一起处理的cm。
[0164]
这些研究表明,s100a9可以作为从brca1-mt乳腺上皮细胞和wt乳腺癌细胞到免疫细胞的信号分子。为了支持cxcl12也可以介导s100a9在免疫细胞中的作用,发明人研究了来自brca1-mt乳腺上皮(g600) 细胞和oe-s100a9-emt6细胞的cm如何控制raw264.7细胞中分子的作用。
[0165]
发明人发现tas和amd3465(一种cxcl12受体抑制剂)可抑制s100a9介导的s100a9、cxcl12、pstat3 和arg1的蛋白水平的增加(图6中k)。在与来自oe-s100a9-emt6细胞的cm一起培养的raw264.7细胞中观察到类似的效果(图6中l)。这些结果表明cxcl12介导s100a9信号从上皮细胞到免疫细胞。
[0166]
值得注意的是,cxcl12的抑制也减少了s100a9,表明免疫细胞中也存在同样的s100a9-cxcl12轴的正反馈调节回路。为了阐明癌症中升高的s100a9信号传导是否会增加小鼠中mdsc的累积,发明人构建了强力霉素(dox)诱导s100a9表达的b477细胞系统(brca1-wt-dox)和g600细胞系统(brca1-mt-dox)(附图 6中d,e)。然后发明人将这些细胞系统植入裸鼠的乳腺脂肪垫中,并在肿瘤直径达到1厘米时诱导s100a9 表达。发明人分别在wt-dox和mt-dox小鼠中诱导s100a9表达48小时后,在血液和乳腺肿瘤组织中检测到s100a9阳性群
体和cd11b/gr1双阳性mdsc亚群增加;然而,在mt-dox小鼠模型中观察到更高水平的增加(图6中m-n,(n=4只小鼠/组))。
[0167]
此外,发明人还检查了sgs100a9在同种异体移植小鼠肿瘤模型中的作用,数据表明cd11b/gr1双阳性mdsc在b477-sgs100a9(wt-sgs100a9)组和g600-sgs100a9(mt-sgs100a9)组的血液中的数量分别从36%大幅减少至12%和62%至22%(图6中o-p,(n=4只小鼠/组))。brca1-mt小鼠脾脏中增加的tgf-β、arg1和cxcl12蛋白水平进一步支持了这一发现(附图6中f)。最后,发明人证明sgcxcl12可以阻断s100a9 过表达所诱导的mdsc在肿瘤生长的作用(图6中q-r和附图6中g)。图6中q-r为mdsc来自balb/c 小鼠的乳腺肿瘤组织(q)和血液样本(r)细胞通过cytof分析(每组n=6-10只小鼠,并将它们随机混合成3 个样本进行cytof分析)。
[0168]
总之,这些数据揭示了s100a9-cxcl12在诱导mdsc积累以建立免疫抑制/肿瘤允许环境中的重要作用。
[0169]
实施例7
[0170]
本实施例论证了抑制s100a9-cxcl12信号传导使乳腺癌对免疫检查点治疗(icb)敏感。
[0171]
以上数据显示,在乳腺上皮细胞中,brca1突变可通过s100a9-cxcl12信号轴激活肿瘤相关通路,并通过招募相关免疫细胞而形成肿瘤微环境。为了研究抑制s100a9-cxcl12轴在肿瘤发生过程中的精准治疗的作用,发明人用emt6细胞系,敲除s100a9(sgs100a9)和cxcl12(sgcxcl12)并原位植入balb/c 同系小鼠中,发现敲除这两个基因显著抑制了肿瘤的发生和生长(图7中a-7b)(n=15只小鼠)。而在体外敲除这两个基因并不影响细胞的生长(图7中c),表明这些基因是通过肿瘤免疫微环境起作用的。
[0172]
为了阐明s100a9和cxcl12对肿瘤允许微环境中免疫细胞的影响,发明人对对照组、sgs100a9和 sgcxcl12组进行了cytof分析,发现cd8 t细胞、cd4 t细胞和活性t细胞群增多在sgs100a9和sgcxcl12 组,而mdsc种群减少(图7中d)。表明抑制s100a9或cxcl12信号可以刺激免疫原性并减少乳腺组织中的免疫抑制微环境。图7中d为通过cd3/cd4、cd3/cd8、cd11b/ly6g、cd11b/ly6c和cd3/cd28抗体的cytof分析,在balb/c小鼠中不表达或表达sgs100a9和sgcxcl12的肿瘤组织中不同免疫细胞的百分比(n=3只小鼠/组)。
[0173]
为了评估发明人的发现对人类乳腺癌患者治疗效果的重要性,发明人分析了人的数据库,并发现同样存在brca1负向调控s100a9和cxcl12的关系。来自两个不同数据库(gse19783-gpl6480和tcga) 的数据显示,增加的cxcl12表达与低brca1表达和高s100a9表达相关(图7中e-7f和附图7中a-7g)。
[0174]
值得注意的是,cxcl12的表达在brca1低表达和s100a9高表达的人群中进一步增加(图7中e)。与表达数据一致,s100a9高表达和brca1低表达(p=0.0008748)的人群的生存预后比其他人群差(图7 中f和附图7中f-7g)。与该结果一致,sgs100a9介导的s100a9-231细胞敲低抑制了异种移植小鼠模型中的癌症发生和生长(图7中g-7h)。
[0175]
因为brca1-mt乳腺上皮细胞和免疫细胞表现出高水平的s100a9和pstat3,这可能诱导pd1和/或 pd-l1的过度表达,发明人用pd-1抗体(apd-1)和/或抑制s100a9/cxcl12信号来测试icb的潜在作用。因为大约25%的brca1co/co;mmtv-cre小鼠在大约18个月内随机发生乳腺肿瘤,因此很难在初始阶段检测到原发性肿瘤。因此,发明人决定使用该模型来研
究治疗对肿瘤复发和转移的影响,这在手术切除原发性肿瘤后经常发生。
[0176]
对于该实验,发明人用》100只brca1co/co;mmtv-cre雌性小鼠,将它们交配一到两次(用于激活 mmtv-cre的表达),并监测它们是否出现肿瘤,然后在肿瘤大小约为1.0厘米时切除。发明人将这些小鼠分成具有不同治疗条件的4组(图7中i),并在初始治疗后42天监测复发性肿瘤的生长和肝脏和肺中的转移。对照组(第1组,n=10,仅pbs)中的所有小鼠在原始腺体中都有复发性肿瘤,并且6/10的小鼠还在其他乳腺中出现了额外的肿瘤,肿瘤体积从100mm3到2250mm3不等(图7中j)。8/10小鼠在肝脏或肺中具有转移灶(图7中k)。在第2组(仅apd-1)中,4/6只小鼠出现复发性肿瘤,并且还在其他乳腺中出现了额外的肿瘤,肿瘤体积从150mm3到1800mm3不等,所有6只小鼠都有肺转移,有的转移在肝脏或卵巢(图7中j-k),表明单独icb对这些肿瘤的影响很小。在第3组(s100a9抑制剂tas,n=3)中,2/3的小鼠有复发性肿瘤,但在肝脏或肺中均未观察到转移(图7中j-7k),这表明tas可能会减少肿瘤转移,尽管对复发性肿瘤的生长没有抑制作用。
[0177]
值得注意的是,数据表明抑制剂治疗的效果不如crispr-cas9介导的敲除。发明人认为这主要是因为抑制剂治疗受有效药物浓度的影响,在体内条件下通常无法达到最大值。在第4组(tas/apd-1,n=5)中,所有小鼠均未出现大肿瘤,而在原肿瘤部位出现小残留物/结节(0-230mm3),同时在肝和肺没有转移的发生(图7中j-7k)。
[0178]
对治疗前切除的乳腺肿瘤和治疗后检测到的小结节进行组织学分析,发现s100a9和cxcl12蛋白水平降低(图7中l)。这些数据表明,虽然单独的icb对复发性肿瘤的生长和转移几乎没有影响,这与之前的发现一致,即brca1缺陷小鼠肿瘤对icb不敏感(图2中1),但它可以通过联合抑制s100a9/cxcl12信号来提升icb的治疗效果。
[0179]
接下来,发明人试图利用肿瘤细胞系545评估icb/药物组合对原发性肿瘤的影响,该细胞系来源于 brca1co/co;mmtv-cre乳腺肿瘤,可以在fvb小鼠中形成同种异体肿瘤。crispr-cas9介导的s100a9敲除也可以显着减少肿瘤生长(附图7中h)。
[0180]
发明人首先测试了apd1抗体的单一治疗。与brca1co/co的处理一致;mmtv-cre荷瘤小鼠,对原发性乳腺肿瘤的生长没有抑制作用(图7中m-7n)。发明人还发现tas和amd3465单独处理可增加pdl1和 pd1的表达(附图7中m-p)。然后,发明人用tas apd1和amd apd1联合治疗545-fvb鼠模型。数据显示,所有联合治疗均显示在前10-20天抑制肿瘤生长,并在第21天几乎完全阻断肿瘤生长(图7中o)。治疗在第21天停止,小鼠最多保持29天,当它们被处死并分析肿瘤的大小(图7中p)和体积(附图7 中j)。值得注意的是,所有荷瘤小鼠的脾脏都增大了,在药物治疗后脾脏缩小到大约正常大小(附图7中 l)。同时,第13天和第29天的cytof分析显示,与对照组相比,联合治疗组cd8 t细胞、cd4 t细胞和cd3 /cd28 活性t细胞的数量在13天均显著增加(图7中q)和第29天(图7中r),但在第29天效果更为显著。
[0181]
与上述分析一致,乳腺组织中s100a9、cxcl12和cxcr4的蛋白质水平在apd1-tas和sgs100a9中显著降低(附图7中q)。所有这些数据表明,brca1缺乏通过激活s100a9-cxcl12信号轴在乳腺组织中形成了一个肿瘤耐受微环境,这使癌细胞对icb不敏感,而s100a9-cxcl12信号轴的抑制可以使癌症对icb 敏感。
[0182]
综上,发明人使用多种方法研究了brca1缺乏对tme形成的影响,并取得了几个主要发现:1)乳腺上皮细胞中brca1缺乏增加了s100a9的表达,s100a9被分泌并正向调节其自身和附近的免疫细胞,并逐渐在乳腺组织中形成免疫抑制环境,其特征是s100a9表达升高
和mdsc积累,有利于brca1缺陷乳腺上皮细胞的致癌激活;2)s100a9-cxcl12轴在乳腺上皮与周围免疫环境之间构成正反馈回路,使癌细胞能够抵抗icb;3)抑制s100a9或cxcl12使乳腺癌对icb敏感,这可能提示抑制s100a9-cxcl12高表达是治疗乳腺癌发生和进展的新策略。
[0183]
总之,发明人认为s100a9-cxcl12通过以下作用促进肿瘤发生和肿瘤进展。首先,高水平的 s100a9-cxcl12促进mdsc的积累和扩增,这有助于诱导免疫抑制/肿瘤微环境,从而有利于肿瘤的形成。在这方面,发明人的数据表明crispr-cas9介导的s100a9或cxcl12敲除显著抑制了mdsc活化。而且,用s100a9或cxcl12的抑制剂治疗也显著抑制mdsc积累并减缓肿瘤生长。此外,增多的mdscs 和增加的pstat3可以激活pd-1/pd-l1介导的免疫检查点以支持肿瘤生长。因此,发明人在brca1-mt小鼠中测试了s100a9或cxcl12和pd1抗体抑制剂联合治疗的疗效,发现该治疗显著抑制了乳腺肿瘤的生长、复发和转移。这项研究提供了一种新的策略来抑制s100a9和/或cxcl12表达升高的患者乳腺癌的发生和进展。
[0184]
以上所述仅为本发明的优选实施例而已,并不用于限制本发明,对于本领域的技术人员来说,本发明可以有各种更改和变化。凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。
