1.本发明涉及一种基因工程亚单位疫苗的制备方法及其应用,更具体一点说,涉及一种基于反向疫苗学技术的猪肺炎支原体多表位基因工程亚单位疫苗的制备方法及其应用,属于生物技术中的基因工程领域。
背景技术:
2.猪支原体肺炎(mycoplasmal pneumoniae of swine,mps)是由于感染猪肺炎支原体(mycoplasma hyopneumonia,mhp)而引发的慢性呼吸道传染病,又称猪地方性肺炎(porcine enzootic pneumonia,pep)。仔猪感染偏多,临床上以哮喘、阵发性痉挛、咳嗽、食欲减退和肺部水肿为主要特征,最终导致窒息死亡,该病具有感染率高、传染性高的特点。尽管mps的死亡率比较低,但mhp的感染能够降低宿主的免疫力,较易导致继发感染,从而增加死亡率并带来更大的经济损失,常见引发继发感染的病原体包括多杀性巴氏杆菌(pasteurella multocida)、猪繁殖呼吸综合症病毒(porcine reproductive and respiratory syndrome virus, prrsv)等。mhp的体外培养需要使用血清等昂贵试剂,因此限制了商业灭活疫苗或减毒活疫苗的工业化生产及大规模使用,同时商业疫苗只提供部分保护而无法完全清除该病原体。另一方面,mhp的基因组具有多变性并且蛋白质组表现出复杂的翻译后修饰,因此常规基于毒力因子的亚单位疫苗可能具有有限的适用性,对一些变异毒株无法发挥保护作用,需要新的疫苗开发策略。
3.基于反向疫苗学技术的多表位疫苗已经在登革热、丝状虫、假单胞菌、hiv 以及疟疾等疾病和病原体的疫苗及药物开发中得到应用,多表位疫苗具有安全性好、成本低、使用简便及易区分感染动物和免疫动物的优势而具有良好的应用前景。一般反向疫苗学在疫苗开发流程中首先从泛基因组分析开始,不需要培养病原微生物,并且病原在不同时期和环境表达的蛋白都会被分析作为候选抗原,因此可以提高疫苗的普遍适用性。
技术实现要素:
4.为了解决上述现有技术问题,本发明提供具有普遍适用性,对一些变异毒株也能够发挥保护作用等技术特点的一种基于反向疫苗学技术的猪肺炎支原体多表位基因工程亚单位疫苗的制备方法及其应用。
5.为了实现上述目的,本发明是通过以下技术方案实现的:
6.本发明一种基于反向疫苗学技术的猪肺炎支原体多表位基因工程亚单位疫苗的制备方法,该制备方法为:
7.步骤1):通过反向疫苗学的方法筛选了mhp基因组中合适的候选抗原基因;
8.步骤2):从筛选出的候选抗原基因中筛选出可以结合mhc-i类和mhc-ii 类分子的抗原表位;
9.步骤3):将筛选得到的抗原表位序列通过接头融合表达制备重组蛋白 mhpmev;
10.步骤4):将重组蛋白mhpmev按照一定的浓度与佐剂混合,获得猪肺炎支原体多表位基因工程亚单位疫苗。
11.作为优选的,筛选能够结合mhc-i类和mhc-ii类分子的抗原表位为11条短肽,并且11条短肽间均能够自由组合,所述的11条短肽为khytynfdfdy、 qvdgyfigl、kifdnnlnf、kltflisfy、ladlllafy、wkeittaee、 fklvipdnk、nkkdknpkd、kltnkhspe、qniklkplkfnte、 ekifeirfetnd。
12.作为优选的,筛选能够结合mhc-i类分子的抗原表位为6条短肽,并且6 条短肽间均能够自由组合,所述的6条短肽为khytynfdf、ytynfdfdy、 qvdgyfigl、kifdnnlnf、kltflisfy、ladlllafy。
13.作为优选的,筛选能够结合mhc-ii类分子的抗原表位为10条短肽,并且 10条短肽间均能够自由组合,所述的10条短肽为kltnkhspe、wkeittaee、 fklvipdnk、feirfetnd、kifeirfet、ekifeirfe、niklkplkf、 qniklkplk、lkplkfnte、nkkdknpkd。
14.作为优选的,用“aayaay”接头或其他蛋白接头连接抗原表位以构成一段完整的蛋白质序列,所述蛋白质序列seq id no.2所示。
15.作为优选的,通过反向疫苗学的方法筛选mhp基因组中合适的毒力因子的步骤为:
16.a)获取若干猪肺炎支原体mhp菌株的基因组数据,对其进行泛基因组分析,将核心基因翻译后得到核心蛋白,以其中一株中的保守核心蛋白作为标准进行进一步的筛选;
17.b)将核心蛋白比对到猪的蛋白质数据库中,设定同源蛋白的标准,根据标准从核心蛋白中去除宿主的同源蛋白;
18.c)将去除了宿主同源蛋白的mhp核心蛋白进行蛋白亚细胞定位预测,在预测结果中选择mhp的外膜蛋白作为候选蛋白进行进一步筛选;
19.d)获取毒力因子数据库的全部蛋白序列,将外膜蛋白比对到毒力因子数据库中并筛选具有显著比对结果的毒力因子,最终得到有效的抗原蛋白。
20.作为优选的,从筛选出的毒力因子中进一步筛选出可以结合mhc-i类和 mhc-ii类分子的抗原表位步骤为:
21.a)进行毒力因子的信号肽预测,根据预测结果去除毒力因子的信号肽,对去除了信号肽的毒力因子进行免疫原性预测;
22.b)对去除信号肽的毒力因子进行b细胞抗原表位预测;
23.c)从b细胞抗原表位中预测可以结合mhc-ii类分子的抗原表位并预测其免疫原性,然后对免疫原性得分大于0.5的抗原表位进行细胞毒性预测,并且预测其诱导产生ifn-γ的能力,选择可以诱导产生ifn-γ的抗原表位,将其作为能够结合mhc-ii类分子的抗原表位;
24.d)从毒力因子中直接预测可以结合mhc-i类分子的抗原表位,并计算其免疫原性,在结果中选择阳性短肽作为结合mhc-i类分子的抗原表位。
25.作为优选的,将抗原表位通过蛋白接头连接制备重组蛋白mhpmev的步骤为:
26.合并带有重叠序列的可结合mhc-i类和mhc-ii类分子的抗原表位,然后用“aayaay”接头连接以形成一段完整的蛋白质序列。
27.本发明为一种基于反向疫苗学技术的猪肺炎支原体多表位基因工程亚单位疫苗,包括重组蛋白mhpmev,所述重组蛋白mhpmev的基因序列为seq idno.1,所述重组蛋白
qniklkplkfnte的反应结果统计图。
51.图21是本发明elisa法测定免疫小鼠血清抗体对化学合成抗原表位 ekifeirfetnd的反应结果统计图。
52.图22是本发明mhpmev免疫小鼠后脾淋巴细胞增殖实验结果统计图。
53.图23是本发明elisa法测定免疫小鼠血清中ifn-γ水平结果统计图。
54.图24是本发明elisa法测定免疫小鼠血清中il-4水平结果统计图。
55.图25是本发明elisa法测定免疫仔猪血清抗体对重组蛋白mhpmev的反应结果统计图。
56.图26是本发明elisa法测定免疫仔猪血清抗体对重组蛋白mhp597的反应结果统计图。
57.其中,图1中用abcpred预测到的抗原表位数目。
58.图2中用bcepred预测到的抗原表位数目。
59.图3中a:重组蛋白mhpmev的表达框;b:最终表达的重组蛋白mhpmev 的蛋白序列及其物理属性预测结果。
60.图4中,a图:m:蛋白分子量标准;1:未诱导阴性对照;b图:m:蛋白分子量标准;1、2:0.1mm iptg,28℃诱导表达上清和沉淀;3、4:0.2mmiptg,28℃诱导表达上清和沉淀;5、6:0.1mm iptg,18℃诱导表达上清和沉淀;7、8:0.2mm iptg,18℃诱导表达上清和沉淀。
61.图5中,m:蛋白分子量标准;1-8m urea:菌体破碎沉淀物使用不同浓度尿素的溶解上清液。
62.图6中,m:蛋白分子量标准;1:菌体破碎后上清液;2:洗涤液i洗出液; 3:洗涤液ii洗出液;4:洗涤液iii洗出液;5:8m urea溶解上清液;6:复性后上清;7:复性后沉淀。
63.图7中,m:蛋白分子量标准;1:mhpmevc6his复性上清液;2:流穿液; 3:25mm咪唑洗涤液;4:50mm咪唑洗涤液;5:250mm咪唑洗脱液;6: 500mm咪唑洗脱液。
64.图8为血清中总igg1水平测定,其中,*p《0.05,**p《0.01,***p《0.001, ****p《0.0001,统计学显著性分析使用two-way anova。
65.图9为血清抗体对重组蛋白mhpmev的elisa检测结果,其中,*p《0.05, **p《0.01,***p《0.001,****p《0.0001,统计学显著性分析使用two-wayanova。
66.图10为血清抗体对重组蛋白mhp597的elisa检测结果,其中,*p《0.05, **p《0.01,***p《0.001,****p《0.0001,统计学显著性分析使用two-wayanova。
67.图11-21分别是对11条合成的抗原短肽进行间接elisa测定血清抗体的抗原抗体反应,短肽序列标记在测定结果的左上角,其中*p《0.05,**p《0.01, ***p《0.001,****p《0.0001,统计学显著性分析使用two-way anova。
68.图22为免疫后第35和42d,相同浓度的重组蛋白mhpmev对mhpmev-25 和mhpmev-50两个实验组的脾淋巴细胞增殖刺激实验结果,其中,*p《0.05,**p《0.01,***p《0.001,****p《0.0001,统计学显著性分析使用two-wayanova。
69.图23为小鼠血清中细胞因子ifn-γ的水平检测结果,其中,*p《0.05,** p《0.01,***p《0.001,****p《0.0001,统计学显著性分析使用two-wayanova。
70.图24为小鼠血清中细胞因子il-4的水平检测结果,其中,*p《0.05,**p 《0.01,***p《0.001,****p《0.0001,统计学显著性分析使用two-wayanova。
71.图25为仔猪免疫血清抗体对重组蛋白mhpmev的elisa检测结果,其中 *p《0.05,**p《0.01,***p《0.001,****p《0.0001,统计学显著性分析使用two-way anova。
72.图26为仔猪免疫血清抗体对重组蛋白mhp597的检测结果,其中,*p《 0.05,**p《0.01,***p《0.001,****p《0.0001,统计学显著性分析使用 two-way anova。
具体实施方式
73.以下结合说明书附图1-26,对本发明作进一步说明,但本发明并不局限于以下实施例。
74.本发明创造性的通过反向疫苗学的方法筛选了mhp基因组中合适的毒力因子,从这些毒力因子中继续筛选出可以结合mhc-i类和mhc-ii类分子的抗原表位,然后将抗原表位进一步整合为重组蛋白,探究了开发mhp多表位疫苗的可行性,并通过小鼠和仔猪免疫对该多表位疫苗的免疫效果进行评价,为新型猪支原体肺炎疫苗的开发提供了新思路。
75.一种基于反向疫苗学技术的猪肺炎支原体多表位基因工程亚单位疫苗的制备方法及步骤:
76.1.1对猪肺炎支原体的泛基因组分析以及毒力因子的筛选
77.从ncbi数据库中下载21株mhp菌株的基因组数据,这些菌株的基因组信息详见表1,使用bioperl(https://bioperl.org/)脚本bp_genbank2gff3.pl将所有基因组数据中的gbff格式文件转化成gff格式文件,然后使用软件roary (http://sanger-pathogens.github.io/roary/)对其进行泛基因组分析。在roary的分析结果中,核心基因在99%~100%菌株中存在并具有高度同源性,将核心基因翻译后得到核心蛋白,以mhp232株中的核心蛋白作为标准进行进一步的筛选,泛基因组的分析结果详见表2。
78.为了保证最终得到的蛋白全部为宿主的异源蛋白并能够引发免疫反应,通过在线blastp(https://blast.ncbi.nlm.nih.gov/blast.cgi)程序将核心蛋白比对到猪(sus scrofa,taxid:9823)的蛋白质数据库中,将同源蛋白的标准设定为 percentage identity》35%,bit score》100且e-value《1e-5
,然后根据以上标准从核心蛋白中去除宿主的同源蛋白。
79.将去除了宿主同源蛋白的mhp核心蛋白提交到psortb version 3.0 (http://www.psort.org/psortb/)进行蛋白亚细胞定位预测,参数中的“choose anorganism type”设置为“bacteria”,“choose gram strain”设置为“negative”,在预测结果中选择分类为“outer membrane”的蛋白作为mhp的外膜蛋白。其它分类为“unknown”的蛋白则继续提交到cello2go (http://cello.life.nctu.edu.tw/cello2go/)网站上进行蛋白亚细胞定位预测,在分析结果中选择分类为“outermembrane”的蛋白,最后合并psortb version 3.0和 cello2go预测的外膜蛋白做进一步筛选。
80.为了得到有效的毒力因子,下载毒力因子数据库vfdb(virulence factordatabase,http://www.mgc.ac.cn/vfs/main.htm)的全部蛋白序列,用本地blastp 程序将上述筛选到的外膜蛋白比对到该数据库中进一步筛选具有显著比对结果的抗原蛋白,最终得到4个有效的毒力因子。
81.1.2毒力因子的免疫原性预测
82.使用signalip-5.0server(http://www.cbs.dtu.dk/services/signalp/)进行毒
(de3)宿主中,经过抗性筛选及重组质粒鉴定后做菌种保藏,将其作为表达重组蛋白的工程菌,命名为e.coli bl21(de3)-pet22b-mhpmev。
91.1.6重组蛋白mhpmev的表达条件探究及鉴定
92.取上述工程菌于5ml lb液体培养基中培养过夜,添加amp抗生素储液使其终浓度为20μg/ml。次日经过扩大培养之后,在50ml培养体积下探究重组蛋白mhpmev在以下4个条件中的表达情况
①
培养温度:28℃,iptg终浓度: 0.1mm,培养时间:16h;
②
培养温度:28℃,iptg终浓度:0.2mm,培养时间:16h;
③
培养温度:18℃,iptg终浓度:0.1mm,培养时间:24h;
④
培养温度:18℃,iptg终浓度:0.2mm,培养时间:24h。
93.诱导表达结束后离心收集菌体,经过超声破碎后取上清160μl于1.5ml ep 管中,用枪头挑取沉淀重悬于160μl pbs缓冲液中,然后加入40μl 5
×
loadingbuffer,加热煮沸10min后做12%sds-page,电泳结果如图4所示。
94.将破碎后的菌体沉淀分别重悬在1~8m共8中不同浓度的尿素溶液中,室温震荡混匀30min后离心分离上清液进行12%sds-page,转膜后用小鼠6
×
histag单抗为一抗鉴定重组蛋白,检测结果如图5所示。
95.1.7mhpmev的大量表达及包涵体的变复性
96.1)对上述工程菌在5ml lb液体培养基中培养过夜,然后转接到1l培养基中扩大培养4~5h,添加iptg储液至终浓度为0.2mm,然后培养24h。诱导表达结束后,超声破碎菌体,12000rpm离心30min分离上清和沉淀。
97.2)用洗涤液i(50mm tris-hcl,10mm edta,5%甘油,1%triton x-100,100 mmβ-巯基乙醇,ph 8.0),洗涤液ii(洗涤液i 0.3m urea),洗涤液iii (洗涤液i 0.6m urea)依次洗涤包涵体沉淀,每次洗涤后12000rpm离心 15min收集沉淀。
98.3)洗涤结束后对沉淀用20~30ml溶解液(50mm tris-hcl,5%甘油,8m urea) 均匀重悬包涵体沉淀,然后用超声促溶,溶解结束后12000rpm离心20min 取上清保存于4℃冰箱。
99.4)将8m urea溶解上清液装入3.5kda透析袋中,依次在复性液i(复性液iv 6m urea),复性液ii(复性液iv 4m urea),复性液iii(复性液iv 2 m urea)和复性液iv(50mm tris-hcl,5%甘油,ph 8.0)中进行透析复性,每种复性液的透析时间均为3h。复性结束后,取出透析袋中液体,12000rpm 离心20min取上清。
100.5)取破碎后上清,破碎后沉淀,洗涤液i洗出液,洗涤液ii洗出液,洗涤液iii 洗出液,8m urea溶解沉淀,8m urea溶解上清和复性后上清液做12% sds-page电泳,电泳结果如图6所示。
101.1.8重组蛋白mhpmev的镍离子亲和层析纯化
102.将复性之后的重组mhpmev蛋白用镍离子层析进行纯化,纯化方法如下:
103.1)将镍离子亲和层析柱中的液体流下,用10倍柱体积的ddh2o清洗,然后用 3倍柱体积的纯化洗涤液his buffer a(50mm tris-hcl,200mm nacl,25mm 咪唑,10%甘油,ph 8.0)洗涤3次进行柱平衡。
104.2)加入0.45μm滤膜过滤的重组蛋白mhpmev复性后上清液,在摇床震荡并冰浴结合2小时以上,然后流出柱内液体作为流穿液。
105.3)用5倍柱体积的纯化洗涤液his buffer a洗涤2次,每次在摇床上冰浴震荡洗涤
5min以除去杂蛋白。
106.4)2倍柱体积的纯化洗脱液his buffer b(50mm tris-hcl,200mm nacl,250 mm咪唑,10%甘油,ph 8.0)洗涤2次,在摇床上冰浴震荡10min以洗脱目的蛋白。
107.5)洗脱后的目的蛋白在透析液(50mm tris-hcl,5%甘油,ph8.0)中4℃透析过夜,然后bca法进行蛋白定量并将蛋白浓度调整到0.1mg/ml,最后分装保存在-80℃作为动物免疫实验的免疫原。重组蛋白mhpmev的纯化结果如图7所示。
108.1.9动物免疫实验分组及免疫原制备
109.将32只6-8周龄的雌性balb/c小鼠随机分成以下4组:
①
pbs组;
②
m hpm ev-25免疫实验组;
③
m hpm ev-25免疫实验组;
④
商业疫苗组(支必宁,猪支原体肺炎活疫苗(168株),兆丰华生物科技(南京)有限公司)。各组的免疫原制备方法如下:
110.①
无菌pbs缓冲液,每次每只小鼠免疫注射200μl;
111.②
将分装的重组蛋白mhpmev储液稀释到合适的浓度,和弗式佐剂等体积乳化后每次免疫每只小鼠注射25μg mhpmev重组蛋白,注射体积200μl;
112.③
将分装的重组蛋白mhpmev储液稀释到合适的浓度,和弗式佐剂等体积乳化后每次免疫每只小鼠注射50μg mhpmev重组蛋白,注射体积200μl;
113.④
用支必宁专用稀释液重悬冻干疫苗,每次免疫每只小鼠注射200μl。
114.重组蛋白溶液和弗式佐剂混合乳化的方法如下:
115.重组蛋白和弗式佐剂的混合乳化方法参照文献(koh,y.t.,s.a.higgins,j.s. weber,et al.immunological consequences of using three different clinical/laboratorytechniques of emulsifying peptide-based vaccines in incomplete freund's adjuvant[j]. j transl med,2006.4:p.42)进行;简述如下:用5ml注射器取适当稀释的重组 mhpmev蛋白,然后另一个5ml注射器取等体积的弗式佐剂,两个注射器之间用乳胶管连接,然后推注乳化持续约30min,然后将液体滴加到冷水面上,液滴不扩散则说明乳化完成。将乳化后的样品冰上保存,当天制备并且当天使用。
[0116]
1.10小鼠免疫及样品收集
[0117]
在第0,14和28d时进行小鼠免疫,第一次免疫使用弗式完全佐剂(初次免疫),后两次免疫使用弗式不完全佐剂(加强免疫)。在第0d时收集血清作为阴性对照,免疫后每隔7d收集一次血清,整个免疫时间为42d,共收集7次血清样品。在免疫后第35和42d进行血清中细胞因子ifn-γ和il-4的检测,同时做脾淋巴细胞增殖实验以确定重组mhpmev蛋白刺激的细胞免疫水平。
[0118]
1.11免疫小鼠血清检测
[0119]
1)参照试剂盒的使用说明(mouse igg1 elisa kit,索莱宝)对免疫后第35和 42d血清中的总igg1抗体水平进行检测,检测结果如图8所示。
[0120]
2)elisa抗原工作液的制备
[0121]
elisa实验中进行的检测的抗原包括:重组mhpmev蛋白,重组蛋白 mhp597核酸酶(登录号为aav27984.1,依据参考文献(li,p.,y.zhang,x.li,etal.mycoplasma hyopneumoniae mhp597 is a cytotoxicity,inflammation andimmunosuppression associated nuclease[j].vet microbiol,2019.235:p.53-62.)进行制备,分装保存前进行核酸酶的活性鉴定)以及化学合成的11条短肽(由生工生物工程(上海)股份有限公司合
成,纯度为75%)。包板时重组蛋白的工作浓度为1μg/ml,使用常规的elisa抗原包被缓冲液(na2co3/nahco3缓冲液)进行稀释;化学合成短肽的工作浓度为100μg/ml,需要使用高纯度的pbs缓冲液(北京索莱宝科技有限公司)进行稀释。
[0122]
3)免疫血清的elisa检测
[0123]
为了评估小鼠对于重组mhpmev蛋白所产生的体液免疫效应,采用elisa 法检测抗体水平。分别将制成工作液的重组mhpmev、重组mhp597和11条化学合成的抗原表位包被96孔板(corning,stripwell
tm microplate),elisa 封闭液(5%脱脂奶粉,5%bsa溶于pbs中)37℃封闭2h,孵育抗体稀释液(2%脱脂奶粉,1%bsa溶于pbs中)1:100稀释的血清样品2h,然后再孵育二抗,加显色液显色。其中上述每两步间均用elisa洗涤液洗板4次,每次1min。最后加入elisa终止液终止显色反应,检测450nm波长下的光密度(optical density, od)值,检测结果如图9-21所示。结果显示,实验组mhpmev-25和mhpmev-50 的免疫血清中均可检测到与重组蛋白mhpmev、mhp597以及化学合成的抗原表位的特异性抗体。
[0124]
1.12脾淋巴细胞增殖实验
[0125]
为了评估小鼠对于重组mhpmev蛋白抗原所产生的细胞免疫效应,使用小鼠淋巴细胞分离液(北京达科为生物技术有限公司)对免疫后35和42d的小鼠进行脾淋巴细胞分离,具体操作流程参见产品说明书。对分离后的各组脾淋巴细胞进行细胞计数,96孔板每孔加入100μl稀释至4
×
106个/ml的细胞悬液。 37℃,5%co2培养箱中培养至细胞完全贴壁后,分别在每孔加入100μl稀释后的重组蛋白mhpmev(实验组,工作浓度为10μg/ml),1640培养基(阴性对照组)与刀豆蛋白a(cona,阳性对照组,工作浓度为10μg/ml)。将细胞放入co2培养箱继续培养42h后每孔加入20μl mtt(5mg/ml),继续培养4 h,弃尽细胞培养上清,每孔加入100μl dmso,于振荡器上震荡混匀5min,最后检测490nm波长下的od值,按照以下公式计算刺激指数(si),si=刺激孔od-阴性对照孔od平均值/cona阳性对照组-阴性对照孔od平均值,结果如图22所示。
[0126]
1.13细胞因子检测
[0127]
参照试剂盒的使用说明(mouse ifn-γelisa kit和mouse il-4elisa kit,索莱宝)对免疫后第35和42d血清中ifn-γ和il-4的含量进行检测,结果如图 23-24所示。
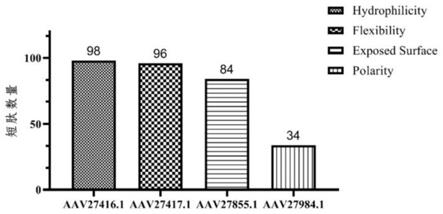
[0128]
综合体液免疫和细胞免疫的检测结果,在小鼠免疫实验中,50μg注射剂量和25μg注射剂量诱导产生的血清抗体对重组蛋白mhpmev和mhp597的检测结果没有明显的差异,但是在细胞免疫检测中,50μg注射剂量诱导的脾淋巴细胞增殖水平和细胞因子水平均高于25μg注射剂量,因此相较于25μg,在小鼠免疫中,50μg为每次免疫单只小鼠的最佳注射剂量。
[0129]
1.14仔猪免疫及血清抗体检测
[0130]
为了在本体动物仔猪中确定mhpmev的免疫原性,15只4周龄雌性仔猪 (7-8公斤,猪肺炎支原体抗原和抗体阴性)被随机分为3组,每组5只,组名以及注射免疫原如下:
[0131]
①
生理盐水组(阴性对照组,每次免疫在颈部肌肉注射2ml无菌生理盐水);
[0132]
②
m hpm ev组(实验组,在第0和14天免疫两次,每次免疫取1mg/ml-80℃冻存的mhpmev重组蛋白1ml,和等体积的montanideim isa 206vg佐剂(seppic公司)充分乳化后进行仔猪颈部肌肉注射;
[0133]
③
m hp组(商业疫苗组,每次免疫在颈部肌肉注射1ml,制备方法和实验步骤1.9相同);
[0134]
第一次免疫时即收集血清,然后每隔一周收集一次血清样品,整个动物实验持续4周,共计收集5次血清样品。免疫结束后,参照实验步骤1.10进行血清抗体检测,待测抗原为分装冻存的重组蛋白mhpmev和mhp597,检测结果如图25-26所示。检测结果显示,在免疫后第21和28d,血清抗体对重组蛋白 mhpmev和mhp597的检测结果显著高于生理盐水阴性对照组(p《0.0001)。
[0135]
2.附录
[0136]
表1:用于泛基因组分析的猪肺炎支原体菌株及其基因组信息
[0137][0138]
表2:roary对21株猪肺炎支原体的泛基因组分析结果
[0139][0140]
表3:最终得到的抗原蛋白的信息及其免疫原性预测
[0141][0142]
注:
[0143]
a aav27855.1的蛋白全长为957aa,signalip-5.0server预测其信号肽所在位置为1-31aa。
[0144]
b aav27984.1的蛋白全长为377aa,signalip-5.0server预测其信号肽所在位置为1-22aa
[0145]
c aav27416.1和aav27417.1蛋白没有预测到信号肽序列
[0146]
表4:预测得到的b细胞抗原表位信息
[0147]
[0148][0149]
表5:可诱导产生ifn-γ的抗原表位(结合mhc-ii类分子的抗原表位)
[0150][0151]
表6:可结合mhc-i类分子的抗原表位的预测结果
[0152][0153]
seq id no.1
[0154]
aaacactatacatacaattttgacttcgattacgccgcttacgcggcgtaccaagttgat ggttatttcatcggcctggccgcgtatgctgcttataagatcttcgacaacaacctgaatt ttgcagcgtacgcggcgtacaagttgacgttcctgatttccttttatgcagcttacgcgg cgtatctggcagatctgctcctggcattttatgcgg
cgtatgcggcgtactggaaagaaa tcaccaccgcagaagaggccgcgtacgctgcgtacttcaaattagtgattccggataat aaagcagcttacgcggcgtataacaagaaagacaaaaacccgaaggacgccgcctacg ctgcgtacaagttgaccaataagcacagcccagaggcagcttacgccgcctaccagaat attaaattgaagccgctgaagttcaacaccgaagcagcgtatgcggcgtatgaaaaaat tttcgagatccgttttgagactaacgac
[0155]
seq id no.2
[0156]
khytynfdfdyaayaayqvdgyfiglaayaaykifdnnlnfaayaaykltflisfyaayaay ladlllafyaayaaywkeittaeeaayaayfklvipdnkaayaaynkkdknpkdaayaay kltnkhspeaayaayqniklkplkfnteaayaayekifeirfetnd
[0157]
最后,需要注意的是,本发明不限于以上实施例,还可以有很多变形,如包括利用本方法筛选获得的猪肺炎支原体其他抗原及表位制备的疫苗等。本领域的普通技术人员能从本发明公开的内容中直接导出或联想到的所有变形,均应认为是本发明的保护范围。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。