1.本发明涉及生物技术领域,具体地涉及一种神经元发育调控多肽及其应用。
背景技术:
2.l型电压门控钙通道又称为cav1通道,共有cav1.1至cav1.4四种亚型。其中cav1.2和cav1.3在神经系统广泛表达,介导神经元兴奋-转录耦联过程,进而调控神经元发育。cav1通道碳末端(distal c-terminus,dct)被报道具有抑制自身钙内流水平的作用,称为碳末端介导的抑制作用(c-terminus mediated inhibition,cmi)。与此同时,cav1.2的一种dct片段ccatc(calcium channel associated transcriptional regulator)被报道可由独立于通道的外显子表达,并作为转录因子进核,进而促进神经元发育过程。除ccatc外,神经元中还存在水解酶直接截切通道而产生的dct片段,分别为cav1.3通道的dct剪切片段cctd及cav1.2的dct剪切片段cctc。这两种剪切片段一方面可能具有通道抑制作用进而抑制神经元发育,另一方面可能具有转录因子作用促进神经元发育,合理优化通道剪切片段可实现同一基因编码多肽基于不同条件对神经元发育进行双向调控。
技术实现要素:
3.本发明旨在设计一类基因编码的神经元发育调控多肽,该多肽具有同一核心片段基于不同细胞定位序列引起不同神经元发育方向的效果。
4.为达上述目的,本发明提供一种基因编码的神经元发育调控多肽,该基因编码的神经元发育调控多肽组成包括:
5.第一核酸序列以及第二核酸序列。
6.其中,所述第一核酸序列为l型电压门控钙通道碳末端片段,所述第二核酸序列为核输出序列或核定位序列。
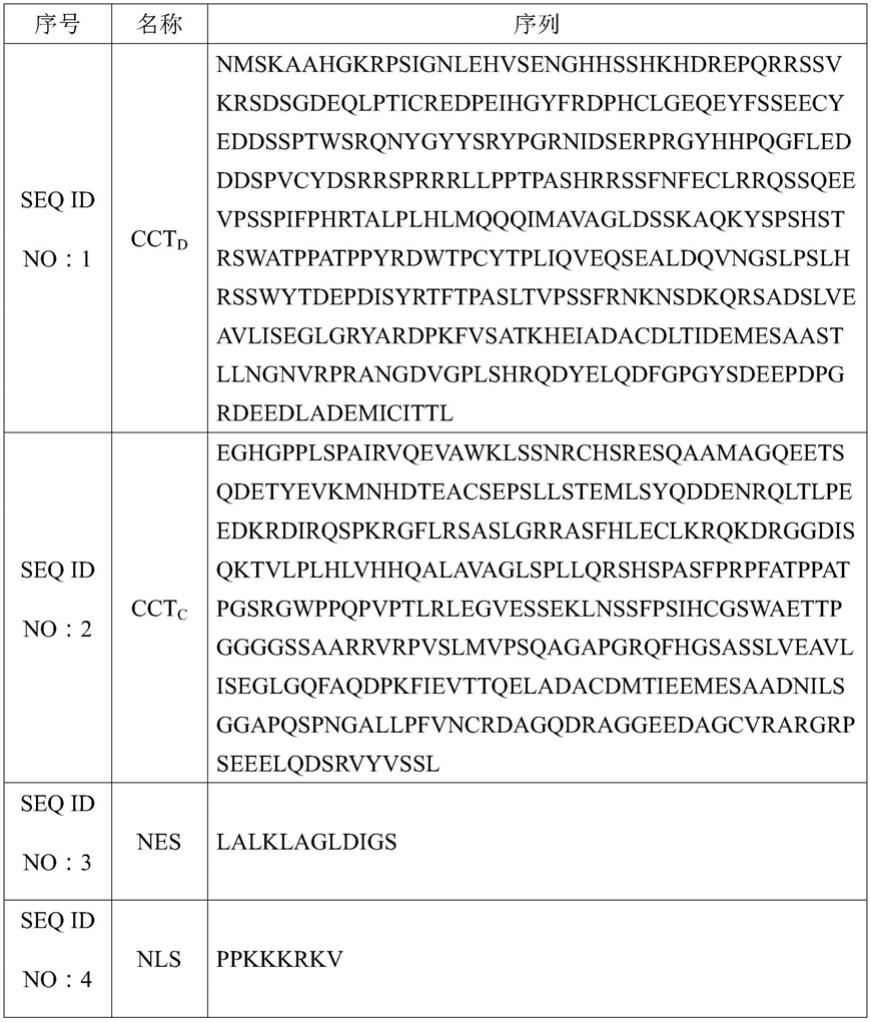
7.本发明的第一核酸序列为cctd或cctc。
8.本发明的cctd的氨基酸序列如下所示:
9.nmskaahgkrpsignlehvsenghhsshkhdrepqrrssvkrsdsgdeqlpticredpeihgyfrdphclgeqeyfsseecyeddssptwsrqnygyysrypgrnidserprgyhhpqgfledddspvcydsrrsprrrllpptpashrrssfnfeclrrqssqeevpsspifphrtalplhlmqqqimavagldsskaqkyspshstrswatppatppyrdwtpcytpliqveqsealdqvngslpslhrsswytdepdisyrtftpasltvpssfrnknsdkqrsadslveavliseglgryardpkfvsatkheiadacdltidemesaastllngnvrprangdvgplshrqdyelqdfgpgysdeepdpgrdeedlademicittl(seq id no:1);
10.本发明的cctc的氨基酸序列如下所示:
11.eghgpplspairvqevawklssnrchsresqaamagqeetsqdetyevkmnhdteacsepsllstemlsyqddenrqltlpeedkrdirqspkrgflrsaslgrrasfhleclkrqkdrggdisqktvlplhlvhhqalavaglspllqrshspasfprpfatppatpgsrgwppqpvptlrlegvesseklnssfpsihcgswaettpggggssaarrvrpvslmvpsqagapgrqfhgsasslveavliseglgqfaqdpkfievttqeladacdmtieemesaadnilsggap
qspngallpfvncrdagqdraggeedagcvrargrpseeelqdsrvyvssl(seq id no:2)。
12.本发明的核输出序列为nes(nuclear export signal),其氨基酸序列为lalklagldigs(seq id no:3)。第一核酸序列和第二核酸序列构成的细胞质定位调控多肽nes-cctc或nes-cctd能够用于神经元发育负向调控。
13.本发明的核定位序列为nls(nuclear localizationsignal),其氨基酸序列为ppkkkrkv(seq id no:4)。第一核酸序列和第二核酸序列构成的细胞核调控多肽nls-cctc或nls-cctd能够用于神经元发育正向调控。
14.本发明还提供一种神经元发育负向调控的方法,使用上述基因编码的神经元发育调控多肽,且该第二核酸序列为核输出序列。
15.本发明的核输出序列为nes。
16.本发明又提供一种神经元发育正向调控的方法,使用上述基因编码的神经元发育调控多肽,且该第二核酸序列为核定位序列。
17.本发明的核定位序列为nls。
18.表1氨基酸序列
[0019][0020]
本发明的基因编码的神经元发育调控多肽具有至少以下优点之一:
[0021]
(1)发明人首次发现并验证了cctd和cctc对于cav1通道的抑制能力。
[0022]
(2)本发明的第一核酸序列l型电压门控钙通道碳末端片段具有基于不同细胞定位进而双向调控神经元发育的能力。
附图说明
[0023]
图1为实施例1的cctc和cctd对cav1通道作用的研究实验结果。
[0024]
图2为实施例2的cctc和cctd在离体培养皮层神经元中对突起发育水平的影响的实验结果。
[0025]
图3为实施例3的利用nes和nls改造后的cctd对神经元突起发育的双向调控的研究的实验结果。
[0026]
图4为实施例4的利用nes和nls定位序列改造后的cctc对神经元突起发育的双向调控的研究的实验结果。
具体实施方式
[0027]
下面将结合实施例对本发明的方案进行解释。本领域技术人员将会理解,下面的实施例仅用于说明本发明,而不应视为限定本发明的范围。实施例中未注明具体技术或条件的,按照本领域内的文献所描述的技术或条件或者按照产品说明书进行。所用试剂或仪器未注明生产厂商者,均为可以通过市购获得的常规产品。
[0028]
实施例1:cctd和cctc对于cav1通道作用的研究
[0029]
在该实施例中,验证了cctd和cctc对于cav1通道的抑制能力。在人配肾细胞hek293中表达cav1.3通道,并同时表达cctd或cctc片段,利用膜片钳全细胞记录方法检测cav1.3通道钙电流,结果见图1。
[0030]
膜片钳全细胞记录方法具体包括以下步骤:在hek293细胞系中利用磷酸钙转染方法过表达cav1.3通道所需的几个亚基,包括成孔亚基α1dl(区别不同通道的主要亚基),辅助亚基α2δ和β2a。在此基础上,共表达cctc或cctd以验证其对通道的作用。利用膜片钳全细胞记录方法,将细胞钳制在-70mv,并给予不同电压刺激,在电压钳模式下记录细胞电流大小。
[0031]
如图1所示,a为:通道电流示意图,以-10mv激活电压作为代表,其中红色代表钙离子电流,左侧红色指示柱指示钙电流大小,灰色为钡离子电流,细胞浴液为10mm钡离子,钡离子电流峰值归一化到钙离子峰值大小,以表针通道钙依然失活水平大小。从左至右依次为,cav1.3通道对照组,通道与cctc共表达组,通道与cctd共表达组。b为:通道钙依赖失活曲线,其中钙依然失活以s
ca
表征,s
ca
=1-r
50
,而r
50
=i
50
/i
peak
,其中i
50
和i
peak
分别为通道在刺激50ms电流大小以及电流峰值大小,r
50
越大,s
ca
越小,钙依赖失活越强;以-10mv的s
ca
作为通道失活特征指标,图中红色和灰色分别为钙电流和钡电流随着激活电压变化的失活统计曲线。绿色区域指代由cctc或cctd造成的钙依赖失活曲线减小范围。c为:通道峰值钙电流曲线(激活曲线),i
ca
等于峰值电流pa与细胞电容pf的比值,单位为pa/pf,以-10mv峰值电流作为通道特征钙电流大小,记为i
ca
。绿色区域指代由cctc或cctd造成的激活曲线减少范围。
[0032]
由图1可知,在表达cctd或cctc之后,cav1.3通道钙依赖失活水平(s
ca
)和峰值电流(i
ca
)均有显著下降,呈现出典型的碳末端介导的抑制作用(cmi),cav1通道整体钙电流呈现抑制状态,说明cctd和cctc具有抑制cav1通道进而抑制神经元发育的潜力。
[0033]
实施例2:cctc和cctd在离体培养皮层神经元中对突起发育水平影响的研究
[0034]
将黄色荧光蛋白yfp标记于cctd及cctc以便于观察在细胞中的表达情况。将黄色荧光蛋白yfp标记后的cctd及cctc离体培养五天的小鼠皮层神经元中过表达2天时间,同时过表达青色荧光蛋白cfp用于追踪神经元突起,以cfp yfp作为对照组,结果见图2。
[0035]
如图2所示,a为:新生icr小鼠皮层神经元在体外培养5天,转染质粒两天后在共聚焦显微镜下观察神经元形态,上图为cfp通道与yfp通道混合后的示意图,下图为追踪的神经元凸起示意图。b为:神经元凸起总长度总结图。c为:cctc或cctd核质比与神经元凸起总长度关系,灰色指示线提示正相关关系。
[0036]
由图2的结果可知,综合统计所有神经元突起总长度后,发现cctd及cctc与对照组
无区别(如图2中a、b),间接提示cctd及cctc可能具有转录因子作用促进神经元发育进而平衡通道抑制带来的负面作用。考虑到转录因子作用在细胞核中起效而通道抑制作用在细胞质中起效,进一步刻画了两种片段核浆比(细胞核荧光值与细胞质荧光值的比值,nucleus/cytosol ratio,n/cratio)与突起总长度的关系,其结果如图2中c所示。图2中c的结果显示,转染了两种碳末端片段的神经元突起总长度均与片段核浆比正相关,即碳末端片段越趋近于细胞质(核浆比越低)时,神经元发育越呈现受损状态,碳末端片段越趋近于细胞核(核浆比越高)时,神经元发育越呈现促进状态。
[0037]
实施例3:利用nes和nls改造后的cctd对神经元突起发育的双向调控的研究
[0038]
在该实施例中,以cctd为例进一步分析核质分布对神经元发育的影响。在cctd片段中分别嵌合核输出序列nes和核定位序列nls能高效的将cctd片段定位于细胞质或细胞核中,并同时融合yfp作为荧光标签,分别得到nes-yfp-cctd和nls-yfp-cctd。以体外培养了15-18天的新生小鼠皮层神经元为考察对象,将yfp作为对照组,nes-yfp-cctd和nls-yfp-cctd转染其中并观察其神经元发育情况,结果见图3。
[0039]
如图3所示,a为:新生icr小鼠皮层神经元体外培养15-18天,转染nes-yfp-cctd或nls-yfp-cctd。左一yfp通道指示cct片段或yfp对照组细胞分别位置,左二cfp通道指示神经元胞体轮廓,左三混合通道指示神经元整体轮廓,最右为神经元凸起追踪图。b为:神经元受到两种不同细胞分布cct片段影响下的凸起总长度变化,及复杂度变化,后者利用sholl analysis统计,其意义为以胞体为中心画10微米同心圆,计算凸起与同心圆相交个数。
[0040]
由图3的结果可知,转染了核输出序列的nes-yfp-cctd的神经元突起总长度显著性低于yfp对照组,而具有核定位序列的nls-yfp-cctd的神经元突起总长度显著性高于yfp对照组。由此可以证明,cctd片段具有基于不同细胞定位进而双向调控神经元发育的能力。
[0041]
实施例4:利用nes和nls改造后的cctc对神经元突起发育的双向调控的研究
[0042]
在该实施例中,以cctc为例进一步分析核质分布对神经元发育的影响。在cctc片段中分别嵌合核输出序列nes和核定位序列nls能高效的将cctc片段定位于细胞质或细胞核中,并同时融合yfp作为荧光标签,分别得到nes-yfp-cctc和nls-yfp-cctc。以体外培养了7天的初生小鼠皮层神经元为考察对象,将yfp作为对照组,nes-yfp-cctc和nls-yfp-cctc转染其中并观察其神经元发育情况,结果见图4。
[0043]
如图4所示,a为:新生icr小鼠皮层神经元在体外培养5天,转染nes-yfp-cctc和nls-yfp-cctc两天后在共聚焦显微镜下观察神经元形态,图中依次为融合通道图、凸起追踪图、胞体放大混合通道图、yfp通道图、以及cfp通道图。b为:神经元凸起总长度总结图。c为:神经元凸起复杂度总结图。
[0044]
由图4的结果可知,细胞质定位片段nes-yfp-cctc显著性抑制了神经元的发育,而细胞核定位片段nls-yfp-cctc显著性促进了神经元的发育。由此可以证明,cctc片段具有基于不同细胞定位进而双向调控神经元发育的能力。
[0045]
虽然本发明内容已以实施方式揭露如上,然其并非用以限定本发明内容,任何熟悉本领域的相关技术人员,在不脱离本发明内容的精神和范围内,当可作各种更动与润饰,因此本发明内容的保护范围当视所附权利要求的保护范围所界定者为准。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。