1.本发明涉及基于结核分枝杆菌抗原多肽的融合蛋白,用于预防、抑制或治疗由结核复合物物种引起的感染和/或疾病。具体而言,本发明涉及包含不引发针对bcg和/或esat-6重复的免疫应答的抗原的融合蛋白。融合蛋白可包含早期和晚期抗原的组合。此外,本发明涉及包含融合蛋白的疫苗、免疫原性组合物和药物组合物。
背景技术:
2.据世界卫生组织(who)称,由结核分枝杆菌(m.tuberculosis)引起的人类结核病是一个严重的全球健康问题,每年造成数百万人死亡。20世纪60和70年代,新发肺结核病例呈下降趋势,但这一积极趋势已被打破,部分原因是艾滋病(aids)的出现和结核分枝杆菌的耐多药菌株的出现。
3.目前可用于临床的唯一疫苗是bcg(卡介苗),这种疫苗的效力仍有争议。bcg是一种减毒活疫苗,在大多数结核病流行地区广泛用于婴儿。bcg通常会在结核病动物模型中诱导高水平的获得性耐药性,并且在儿童中它可以预防传播形式的结核病,例如脑膜炎和粟粒性结核病。当给幼儿服用时,它可以预防结核病多年,但随后疗效下降。各种对照试验的比较表明,卡介苗对成人的保护功效差异很大,功效范围从无效到80%保护。这使得针对结核分枝杆菌的新型改良疫苗的开发成为当务之急,who已将其列为高度优先事项。
4.目前,有几种新的结核疫苗处于临床试验阶段。由于普遍认为需要强大的细胞免疫应答来预防结核感染和疾病,因此目前大多数临床结核病候选疫苗旨在从cd4 或cd8 t细胞诱导经典的th1细胞因子,如ifn-γ或tnf-α,而没有解决关于提高t细胞质量的问题,t细胞质量在保护性免疫中起着重要作用。目前的结核病候选疫苗包括各种疫苗平台,例如灭活的全细胞或全细胞提取物、基于病毒载体的疫苗、活的重组bcg或减毒bcg疫苗,以及dna疫苗。然而,到目前为止,上述疫苗平台都没有产生令人信服的临床结果。
5.提供有效结核疫苗的替代策略围绕着基于结核分枝杆菌抗原融合蛋白的亚单位疫苗展开。亚单位方法已被认为具有许多优点,例如提高的安全性和稳定性以及增强先前bcg疫苗接种的能力。人们已经努力将多种结核分枝杆菌抗原的融合蛋白与适宜的佐剂结合以诱导强大的细胞免疫应答。
6.wo2010006607a2中证明了当与佐剂ic31一起施用时,仅由两种结核分枝杆菌抗原组成的简单融合物可起到结核疫苗的作用。wo2006136162a2、wo2014063704a2和wo2015161853a1都公开了抗结核感染和疾病的融合蛋白,其中多种结核分枝杆菌抗原被混合以提供能够诱导强免疫应答的亚单位疫苗。然而,上述融合蛋白的共同点是,目前还没有一种相应的亚单位疫苗形成商业上可用的抗结核疫苗。
7.因此,改进的结核疫苗具有产生临床上可接受疫苗的潜力,将是有利的。特别地,能够诱导强的高质量免疫应答从而有增强保护能力的融合蛋白将是有利的。
技术实现要素:
8.本文提供了融合蛋白,其在概念上不同于先前开发的用于预防、抑制或治疗结核感染和/或疾病的融合蛋白。本文所述的融合蛋白是基于不引发针对bcg的免疫应答的抗原(本文称为“bcg-抗原”)和/或“esat-6重复”,其中融合蛋白中存在一个以上esat-6抗原拷贝。融合蛋白可包含早期和晚期抗原的组合。与现有结核疫苗相比,本文提供的融合蛋白产生改善的免疫应答。
9.因此,本发明的一个目的涉及提供包含bcg-抗原和/或esat-6重复的融合蛋白,其诱导强的细胞免疫应答,同时改善t细胞的质量。
10.本发明的另一个目的涉及提供包含早期和晚期结核抗原组合的融合蛋白,其既能加速mtb感染后的免疫应答,又能在感染的晚期慢性阶段诱导具有特异性识别细菌潜力的t细胞。
11.本发明的另一个目的是提供融合蛋白,其可作为独立疫苗或与bcg组合,有效地用在疫苗或免疫原性组合物中以预防、抑制或治疗结核感染和/或疾病。
12.因此,本发明的一个方面涉及一种包含至少五种源自结核分枝杆菌的抗原的融合蛋白。
13.本发明的一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含早期和晚期抗原。
14.本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述抗原从bcg中缺失、不分泌或具有低表达。
15.本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含至少两个esat-6重复,例如至少三个esat-6重复,例如至少四个esat-6重复,例如至少五个esat-6重复。
16.本发明的另一方面涉及包含如本文所述的融合蛋白的疫苗或免疫原性组合物。
17.本发明的另一方面是提供本文所述的融合蛋白或本文所述的疫苗或免疫原性组合物,用于针对由毒性分枝杆菌引起的感染和/或疾病,对受试者进行疫苗接种或免疫。
18.本发明的另一方面是提供一种试剂盒,包括:
19.i)如本文所述的融合蛋白或如本文所述的疫苗或免疫原性组合物,
20.ii)bcg,以及
21.iii)可选地,使用说明。
22.本发明的另一方面是提供一种核酸序列,其包含编码如本文所述的融合蛋白的序列。
23.本发明的另一方面是提供一种重组表达载体,其包含如本文所述的核苷酸序列可操作地连接至一个或多个适于指导在适宜宿主中产生融合蛋白的控制序列。
24.本发明的另一方面是提供一种重组宿主细胞,其包含如本文所述的表达载体。
25.附图简要说明
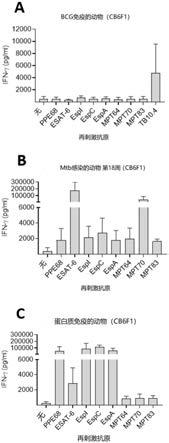
26.图1显示所选的抗原的免疫识别,牛分枝杆菌(m.bovis)bcg danish不诱导针对该抗原的t细胞应答(bcg-抗原)。(a)牛分枝杆菌bcg danish免疫、(b)结核分枝杆菌erdman感染、或(c)用各个抗原免疫后的免疫应答。
27.图2显示所选的含有bcg-抗原的融合蛋白的示意图。
28.图3显示本文所述的另外所选的融合蛋白的示意图。
29.图4显示mtb感染期间esat-6和mpt70的免疫动力学识别。在mtb erdman感染后3、12和20周,在肺中测量(a)抗原特异性干扰素-γ释放或(b)抗原特异性cd4 t细胞的频率的倍数增加。
30.图5显示含有晚期表达抗原的疫苗诱导长期保护。在气溶胶mtb感染后(a)4周和(b)18周,用h104或h105免疫或注射生理盐水的mtb感染动物的肺cfu。h105包括抗原mpt64、mpt70和mpt83。
31.图6显示bcg和h107/caf01的共同施用增加了两种疫苗接种模型中的特异性h107免疫应答。在(a)标准预防模型和(b)bcg再接种模型中,在h107疫苗前12个月施用牛分枝杆菌bcg danish,在h107/caf01免疫后三周,测量脾脏中h107特异性cd4 t细胞的频率。
32.图7显示bcg共同施用影响融合蛋白特异性cd4 t细胞的分化,这取决于融合蛋白是否含有bcg-抗原(h107)或bcg
抗原(h65)。(a)基于单细胞的细胞因子表达,概括显示了t细胞分化,改编自seder等人(2008)。(b)基于对细胞因子ifn-γ、tnf-α和/或il-2的单细胞表达,位于脾脏中的cd4细胞的细胞因子表达谱。(c)表达klrg1的融合蛋白特异性cd4 t细胞的频率。
33.图8显示当含有bcg-抗原的融合蛋白(h107)与bcg共同施用时,针对mtb感染的保护作用增加。mtb erdman感染后4周(a)和18周(b)的肺cfu。
34.图9显示牛分枝杆菌bcg和在caf01中配制的h104-h107融合蛋白的共接种诱导了针对mtb感染的显著保护作用。显示了mtb erdman感染后第4周的肺cfu。
35.图10显示将游离esat-6蛋白添加到疫苗制剂中没有增加疫苗引发的免疫应答,也没有减少mtb感染后的分枝杆菌数量。在(a)疫苗接种后三周和(b)mtb感染后三周测定疫苗接种组中esat-6特异性cd4 t细胞的数量。(c)感染六周后,测定肺部分枝杆菌的数量。
36.图11显示在融合蛋白中重复esat-6导致针对esat-6的免疫反应增加,并且在预防性疫苗接种模型中提供更好的保护作用。(a)免疫后三周,在脾脏中测量h64(一个esat-6拷贝)和h76(五个esat-6拷贝)免疫后的抗原特异性反应。(b)在mtb感染六周后测定肺中的分枝杆菌数量。
37.图12显示在融合分子中重复esat-6改善了esat-6特异性t细胞向感染部位的募集,并改善了疫苗在暴露后的疫苗接种模型中的保护功效。用抗生素治疗感染mtb erdman的动物。在治疗终止前两周,开始用h83/caf01(一个esat-6拷贝)和h84/caf01(四个esat-6拷贝)进行免疫。(a)免疫后两周,肺中esat-6特异性cd4 t细胞的数量。(b)感染后22和35周(最后一次免疫后6和19周)肺中分枝杆菌的数量。
38.图13显示在融合蛋白中重复esat-6增加了免疫应答并改善了疫苗的保护功效。从(a)接种129sc的小鼠和(b)cb6f1小鼠分离的脾脏中的特异性esat-6反应。(c)来自bcg和亚单位共免疫小鼠脾脏中的esat-6特异性免疫应答以及感染4周后肺部的保护功效。
39.图14显示在caf01佐剂中配制的h107融合蛋白是比在caf01中配制的h56更好的疫苗。在mtb感染后4至12周测定肺中的保护功效(δlog
10
cfu
生理盐水-δlog
10
cfu
免疫
)。
40.图15显示在第一次免疫后三周(第二剂h107后一周),与单独bcg相比,在bcg h107组中观察到tb10.4应答增加,这意味着h107 bcg共同施用增加了bcg特异性免疫应答,因此h107发挥作为bcg佐剂的作用。
41.图16显示h107和h107e融合蛋白的组合物的示意图。图16a显示来自大肠杆菌的h107和h107e融合蛋白表达的sds-page和蛋白质印迹(western blots)。与h107相比,h107e增加了蛋白质表达。
42.图17
43.与bcg共接种后,通过表达细胞因子的cd4 t细胞(图17a,左)和经elisa检测的ifnγ释放(图17a,右)二者来测量免疫原性。h107和h107e诱导相似程度的免疫应答。图17b显示h107e与h107一样,诱导对同样的各个抗原的免疫应答。图17c显示,感染后,h107e赋予与bcg相似或更好的保护作用,与bcg和h107e单独相比,bcg h107e共接种导致保护作用显著增加。图17d显示,在bcg记忆小鼠中,与h65(bcg )相比,h107e(bcg-)疫苗接种导致cd4 t细胞分化较少(质量更好),这是通过功能分化评分、fds以及产生il-17的cd4 t细胞比例来衡量。
44.下面将更详细地描述本发明。
具体实施方式
45.定义
46.在进一步详细讨论本发明之前,首先定义以下术语和约定:
47.抗原
48.在本上下文中,术语“抗原”指能够诱导免疫应答的分子,例如免疫原性多肽。抗原产生的免疫应答可以是b细胞驱动的(抗体介导的免疫应答)和/或t细胞驱动的(细胞免疫应答)。
49.本文所述的抗原源自结核分枝杆菌(m.tuberculosis或mtb)。
50.早期和晚期抗原
51.结核分枝杆菌感染的过程基本上经历3个阶段;(i)急性期,(ii)潜伏期/慢性期和可能的(iii)再激活期。结核分枝杆菌的基因表达模式在这些阶段中发生变化。
52.因此,在本文中,术语“早期抗原”指主要在急性期表达的抗原,而“晚期抗原”指主要在潜伏/慢性期表达的抗原。
53.融合蛋白
54.在本文中,术语“融合蛋白”指包含随机顺序的来自结核分枝杆菌或其类似物的两个或更多个抗原的多肽。这些抗原可融合在一起,不管有或没有不同长度和序列的氨基酸接头。
55.融合蛋白可通过可操作地连接两个或更多个编码感兴趣抗原的氨基酸序列的异源核酸序列来产生。为了避免下游生产中的蛋白质聚集,融合蛋白中的所有半胱氨酸可用任何氨基酸代替,但丝氨酸是优选的替代物,因为它与半胱氨酸的结构高度相似。
56.融合蛋白或抗原可包含适当的纯化标签(或亲和标签)以允许从粗生物源(例如重组表达系统)中纯化。纯化标签包括但不限于his标签、几丁质结合蛋白(cbp)、麦芽糖结合蛋白(mbp)和谷胱甘肽-s-转移酶(gst)。
57.在本文中,术语“融合蛋白”与术语“亚单位疫苗”可互换使用。
58.多肽
59.在本文中,术语“多肽”指由通过肽键连接的氨基酸残基、相关的天然存在的结构
变体和/或其合成的非天然存在的类似物组成的聚合物。本文使用常规符号来描述多肽序列:多肽序列的左手端是氨基端(n-端);多肽序列的右手端是羧基端(c-端)
60.多肽可以通过糖基化、脂化、辅基或通过包含另外的氨基酸,例如纯化标签(例如his-标签)或信号肽来进行化学修饰。纯化标签用于获得高纯度的蛋白质制剂。如果用在n-端,his-标签可包含甲硫氨酸作为第一个氨基酸,随后是6-8个组氨酸,如果用在c端,则包含6-8个组氨酸随后是终止密码子。当用在n端时,可删除编码该多肽融合物基因中的甲硫氨酸起始密码子,以避免错误的翻译起始位点。
61.每个多肽由特定的核酸序列编码。应当理解,此类序列包括其类似物和变体,其中此类核酸序列已通过替换、插入、添加或删除一种或多种核酸进行修饰。替换优选地是密码子使用上的保守替换,其不会导致氨基酸序列的任何变化,但可被引入以增强蛋白质的表达。
62.多肽可以重组地或合成地产生,例如,使用自动化多肽合成仪。
63.结核分枝杆菌
64.在本文中,术语“结核分枝杆菌”指分枝杆菌科的病原菌种结核分枝杆菌,其可以是结核感染和疾病的病原体。结核分枝杆菌在本文中可缩写为mtb。
65.如本文所述,结核分枝杆菌感染经历三个阶段。在本文中,术语“感染”指这三个阶段中的任何一个,即针对由毒性分枝杆菌引起的疾病的疫苗接种或免疫可包括急性期、潜伏/慢性期和再激活期。
66.卡介苗(bcg)
67.在本文中,术语“卡介苗(bcg)”指源自牛分枝杆菌的活的、减毒的结核病细菌的菌株。bcg是目前唯一商业上可获得的针对结核感染和疾病的疫苗。
68.bcg是约100年前巴斯德研究所通过对牛分枝杆菌进行减毒开发的,在此过程中,毒株失去了一个重要的基因片段,该基因片段编码毒力相关抗原,如esat-6。这个原始突变被称为rd1。bcg疫苗在随后的30-40年间分发至全球各种实验室和生产设备。这产生了各种疫苗亚株,通常以其生产实验室的位置命名(bcg danish,prague,tokyo等)。由于这些菌株中的许多最初是通过连续培养繁殖的,因此已观察到原始基因组的大量缺失,并且在不同的亚菌株中具有不同的分布。在已分析的bcg疫苗株中,总共报告了至少12个主要的改变/缺失(rd1-11和sigk突变)。其中有些缺失包含免疫学上重要的具有疫苗潜力的抗原,而有些缺失在免疫学上是沉默的。
69.最初的rd1缺失是包含对疫苗非常重要的免疫显性抗原的区域的一个实例。本文公开了在自然感染期间被强烈识别的抗原:rd1、rd2以及表达受sigk调节的抗原。来自所有三个区域的抗原的涵盖范围已经提供了足够多的抗原以构建强大的基于多蛋白的疫苗,还提供了在感染不同阶段表达的抗原。
70.在本文中,bcg菌株可分为两大类:早期bcg菌株和晚期bcg菌株。不要将早期和晚期bcg菌株与早期和晚期抗原混淆。
71.早期bcg菌株缺少rd1区域。早期bcg菌株包括bcg russia、bcg japan、bcg moreau、bcg sweden和bcg birkhaug。
72.晚期bcg菌株缺少rd1和rd2区域,并且在sigk中有突变。晚期bcg菌株包括bcg tice、bcg frappier、bcg pasteur、bcg denmark、bcg glaxo、bcg prague、bcg china以及
基因改造的bcg菌株vpm1002。
73.晚期和早期bcg菌株不具有相同的基因型和表型。因此,在晚期bcg菌株中,rd2区编码的一些抗原已被删除,而其他抗原表达不佳或不分泌。因此,对于这些缺失的、表达不佳或不分泌的抗原,用这些晚期bcg菌株进行疫苗接种不会诱导免疫应答。
74.bcg
抗原
75.在本文中,术语“bcg
抗原”指由给定bcg菌株表达并在用给定bcg菌株疫苗接种后引发免疫应答(通过受刺激细胞的培养物中的ifn-γ释放来测量)的抗原。因此,包含bcg
抗原的融合蛋白,当在用已供给的bcg菌株进行初始疫苗接种之后施用时,可用于增强先前由所述已供给的bcg菌株诱导的免疫应答。
76.在本文中,bcg
抗原可根据相关bcg菌株来定义,选自bcg danish,bcg pasteur和bcg prague和/或这些菌株的任何衍生物(例如vpm1002)。因此,bcg
抗原包括但不限于:rv1886c(ag85b)、rv3804c(ag85a)、rv0288(tb10.4)、rv0287(esxg)、rv3478(ppe60)、rv0475(hbha)、rv3890c(esxc)、rv3891c(esxd)、rv1284(cana)、rv3019c(esxr)、rv3020c(esxs)、rv3017c(esxq)、rv2031c(hspx)、rv0983(pepd)、rv1196(ppe18)、rv2608(ppe42)、rv3619(esxv)和rv3620(esxw),及其变体。
77.bcg-抗原
78.在本文中,术语“bcg-抗原”指在用已供给的bcg菌株接种后不引发免疫应答(通过ifn-γ释放来测量)的抗原,因为它或者被删除、不分泌或者具有低表达水平。因此,包含bcg-抗原的融合蛋白,当在用已供给的bcg菌株进行初始疫苗接种后施用时,不会增强先前由所述已供给的bcg菌株诱导的免疫应答。因此,bcg-抗原可被定义为功能性阴性。
79.在本文中,bcg-抗原可根据相关bcg菌株来定义,选自bcg danish、bcg pasteur和bcg prague和/或这些菌株的任何衍生物(例如vpm1002)。因此,bcg抗原可定义为与这些bcg菌株相关的功能性阴性。
80.因此,bcg-抗原包括但不限于:rv3875(esat-6)、rv3873(ppe68)、rv3876(espi)、rv3615c(espc)、rv3616c(espa)、rv1980c(mpt64)、rv2875(mpt70)、rv2873(mpt83),及其变体。
81.bcg-抗原可用于引发互补免疫应答,即免疫应答针对的mtb抗原与初始bcg疫苗接种的抗原不同。
82.免疫原性表位
83.多肽的免疫原性表位是多肽的一部分,其在动物或人类中和/或通过本文所述的任何生物测定法测定的生物样品中引发免疫应答。多肽的免疫原性表位可以是t细胞表位或b细胞表位。免疫原性表位可以与多肽的一个或几个相对较小的部分相关,它们可以分散在整个多肽序列中或位于多肽的特定部分。对于一些多肽,甚至已经证明表位分散在整个多肽中,覆盖了完整的序列(ravn,1999)。
84.为了鉴定在免疫应答过程中被识别的相关t细胞表位,可以使用“蛮力”法:因为t细胞表位是线性的,多肽的缺失突变体,如果被系统地构建,将会揭示多肽的哪些区域在免疫识别中是必不可少的,例如,通过对这些缺失突变体进行例如本文所述的ifn-γ测定。另一种方法利用重叠肽检测mhc ii类表位,优选合成的,长度为来自多肽的例如20个氨基酸残基。这些肽可以在生物测定中进行测试(例如本文所述的ifn-γ测定),其中一些将产生
阳性反应(因此具有免疫原性),作为该肽中存在t细胞表位的证据。为了检测mhc i类表位,可以预测那些会结合的肽(stryhn,1996),然后合成这些肽并在相关的生物测定中测试它们,例如本文所述的ifn-γ测定。所述肽的长度优选为来自多肽的例如8至11个氨基酸残基。
85.尽管已显示t细胞表位的最小长度至少为6个氨基酸,但此类表位由更长的氨基酸片段构成也是正常的。因此,优选本发明的多肽片段的长度具有至少7个氨基酸残基,例如至少8、至少9、至少10、至少12、至少14、至少16,至少18、至少20、至少22、至少24和至少30个氨基酸残基。因此,在本发明的重要实施方式中,优选的是多肽片段的长度为至多50个氨基酸残基,例如至多40、35、30、25和20个氨基酸残基。预期长度介于10到30个氨基酸残基的肽将被证明是最有效的mhc ii类表位,因此本发明方法中使用的多肽片段的特别优选的长度是18,例如15、14、13、12以及甚至11个氨基酸残基。预期长度在7到12个氨基酸残基之间的肽将被证明是最有效的mhc i类表位,因此用于本发明方法中的多肽片段的特别优选的长度是11,例如10、9、8以及甚至7个氨基酸残基。
86.多肽的免疫原性部分(包含免疫原性表位的片段),包括该免疫原性表位,可被基因异源人群的大部分(高频)或小部分(低频)识别。此外,一些免疫原性部分诱导高免疫应答(显性),而其他免疫原性部分诱导较低但仍然显著的应答(次显性)。高频》《低频可能与广泛分布的mhc分子(hla型)甚至多个mhc分子结合的免疫原性部分有关(sinigaglia,1988;kilgus,1991)。包含来自所述多肽的免疫原性表位的片段可以作为至少10个氨基酸长度的重叠肽存在,从而跨越几个表位。
87.变体
88.在本文中,术语“变体”指本文所述的抗原、多肽或融合蛋白,它们“基本上同源”和/或保留所参照的抗原、多肽或融合蛋白的至少大量的免疫原性。
89.在本文中,术语“基本上同源”指一种抗原、多肽或融合蛋白的氨基酸序列,与其所参照的抗原、多肽或融合蛋白具有至少80%的序列同一性,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。在本文中,术语“大量的免疫原性”指一种抗原、多肽或融合蛋白,与其所参照的抗原、多肽或融合蛋白相比,保留至少80%的免疫原性,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的免疫原性。
90.因此,如本文所述的抗原、多肽或融合蛋白的变体可具有与其所参照的抗原、多肽或融合蛋白相同的免疫原性或至少相同的免疫原性。类似地,与本文所述的一种抗原、多肽或融合蛋白具有确定的序列同一性的抗原、多肽或融合蛋白可具有与其所参照的抗原、多肽或融合蛋白相同的免疫原性或至少相同的免疫原性。
91.抗原重复
92.在本文中,术语“抗原重复”指融合蛋白中存在多于一个抗原拷贝的抗原。抗原重复的不同抗原拷贝在融合蛋白中可定位为(i)连续地,(ii)与不同于该抗原重复的抗原交替地,或(iii)被不同于该抗原重复的抗原隔开。抗原重复可包括至少两个抗原重复,例如至少三个抗原重复,例如至少四个抗原重复,例如至少五个抗原重复。因此,具有例如至少三个抗原重复的融合蛋白包括所述抗原的至少三个拷贝。
93.在本文中,优选的是包含esat-6的抗原重复(即esat-6重复)的融合蛋白。
94.接头分子
95.在本文中,术语“接头分子”或“接头”指融合蛋白中的抗原之间出现的肽序列。接头通常由甘氨酸和丝氨酸等柔性残基组成,因此融合蛋白的相邻结构域相对于彼此自由移动,并在分泌/制造过程中独立正确折叠。如有必要,可使用更长的接头以确保融合蛋白的两个相邻结构域在空间上不会相互干扰。
96.应当理解,遗传水平的接头是由核酸组成。因此,编码本文所述的融合蛋白的核酸可包括由核酸序列表示的接头。
97.共接种
98.在本文中,术语“共接种”指同时施用两种不同的疫苗和/或免疫原性组合物,例如同时施用如本文所述的亚单位疫苗(或融合蛋白)和bcg疫苗,或者用bcg疫苗接种,然后在预期bcg可以在宿主中持续存在的时间段内进行亚单位疫苗接种。
99.术语“共接种”、“共免疫”和“共同施用”在本文中可互换使用。
100.疫苗和免疫原性组合物
101.在本文中,术语“疫苗”和“免疫原性组合物”指包含至少一种抗原的组合物,所述抗原能够提供对结核感染或疾病的主动获得性免疫。“疫苗”或“免疫原性组合物”可优选地包括如本文所述的融合蛋白,其能够提供对结核感染或疾病的主动获得性免疫。因此,与未接种疫苗的动物相比,在用毒性分枝杆菌攻击后,本文所述的疫苗或免疫原性组合物能够减少靶器官中的细菌负荷、延长存活时间和/或减少动物的体重减轻。
102.疫苗或免疫原性组合物可包括免疫学和药学上可接受的载体或媒剂。适宜的载体包括但不限于:多肽通过疏水性非共价相互作用相结合的聚合物,例如塑料,例如聚苯乙烯;或多肽共价结合的聚合物,例如多糖;或多肽,例如牛血清白蛋白、卵清蛋白或锁孔血蓝蛋白。适宜的媒剂包括但不限于稀释剂和悬浮剂。
103.此外,疫苗或免疫原性组合物优选地包括一种或多种佐剂。
104.佐剂
105.在本文中,术语“佐剂”指增强对抗原的免疫应答的化合物或混合物。佐剂可作为缓慢释放抗原的组织贮库以及作为淋巴系统激活剂,其非特异性地增强免疫应答。通常,在没有佐剂的情况下,单独用抗原进行初次攻击将无法引发体液或细胞的免疫应答。
106.佐剂包括但不限于:中性佐剂制剂、阴离子佐剂制剂、阳离子佐剂制剂(例如“caf01”、“caf04”、“caf09”和“caf10”)、阳离子脂质体(例如二甲基二(十八烷基)溴化铵(dda))、quil a、qs21、聚i:c、氢氧化铝、弗氏不完全佐剂、ifn-γ、il-2、il-12、单磷酰脂质a(mpl)、海藻糖二霉菌酸酯(tdm)、海藻糖二山萮酸酯(tdb)、胞壁酰二肽(mdp)、单分枝酰基甘油(mmg)、cpg和“ic31”,或其组合。
107.表达载体
108.在本文中,术语“表达载体”指将重组遗传材料转移到宿主细胞中的用作媒剂的dna分子。表达载体的四种主要类型是质粒、噬菌体和其他病毒、粘粒和人工染色体。表达载体本身通常是一种dna序列,其由插入物(异质核酸序列,转基因)和作为表达载体“骨架”的更大序列组成。表达载体的目的是将遗传信息传递给宿主,通常是在靶细胞中分离、增殖或表达插入物。表达载体特别适用于在靶细胞中表达异源序列,并且通常具有驱动异源序列表达的启动子序列。
109.可操作地连接
110.在本文中,术语“可操作地连接”指元件连接成为功能单元(例如基因或开放阅读框)的一部分。因此,通过将启动子可操作地连接至编码多肽的核酸序列,这两个元件成为功能单元——基因的一部分。表达控制序列(启动子)与核酸序列的连接使得启动子指导核酸序列转录成为可能。通过可操作地连接两个编码多肽的异源核酸序列,这些序列成为功能单元——开放阅读框的一部分;该开放阅读框编码融合蛋白,包含由所述异源核酸序列编码的氨基酸序列通过可操作地连接两个氨基酸序列,这些序列成为相同功能单元——多肽的一部分。可操作地连接两个异源氨基酸序列产生杂交(融合)多肽。
111.哺乳动物
112.在本文中,术语“哺乳动物”指属于哺乳动物纲的任何动物,包括但不限于:啮齿动物、灵长类动物、有蹄类动物和食肉动物。有蹄类动物包括但不限于:牛、马、猪、绵羊、山羊和骆驼。
113.在本文中,哺乳动物优选地是人或家畜。
114.序列同一性
115.在本文中,术语“序列同一性”指基因或蛋白质之间分别在核苷酸、碱基或氨基酸水平上的序列同一性。具体而言,如果dna序列的转录物可以转录为相同的rna序列,则认为dna和rna序列是相同的。
116.因此,在本文中,“序列同一性”是氨基酸水平上蛋白质之间同一性的度量和核苷酸水平上核酸之间同一性的度量。当序列比对时,可通过比较每个序列中给定位置的氨基酸序列来确定蛋白质序列同一性。类似地,当序列比对时,可通过比较每个序列中给定位置的核苷酸序列来确定核酸序列同一性。
117.为确定两个氨基酸序列或两个核酸的同一性百分比,将序列进行比对以达到最佳比较目的(例如,可在第一个氨基酸序列或核酸序列中引入空位(gap)以与第二个氨基酸或核酸序列进行最佳比对)。然后比较相应氨基酸位置或核苷酸位置处的氨基酸残基或核苷酸。当第一个序列中的一个位置被与第二个序列中相应位置的相同氨基酸残基或核苷酸占据时,则该位置的分子是相同的。两个序列之间的同一性百分比是这些序列共享的相同位置数量的函数(即,%同一性=#相同位置数/#总位置数(例如,重叠位置)
×
100)。在一种实施方式中,两个序列的长度相同。
118.在另一种实施方式中,两个序列具有不同的长度并且空位被视为不同的位置。人们可以手动比对序列并计数相同氨基酸的数量。或者,比对两个序列以确定同一性百分比可用数学算法来实现。这种算法被并入(altschul等人,1990)的nblast和xblast程序中。blast核苷酸搜索可用nblast程序进行,得分=100,字长=12,以获得与本发明的核酸分子同源的核苷酸序列。blast蛋白质搜索可用xblast程序进行,得分=50,字长=3,以获得与本发明的蛋白质分子同源的氨基酸序列。
119.为了获得用于比较目的而引入空位的序列比对,可使用gapped blast。或者,可用psi-blast进行迭代搜索,以检测分子间的远近关系。当使用blast、xblast和gapped blast程序时,可使用各程序的缺省参数。见http://www.ncbi.nlm.nih.gov。或者,可在序列比对后,例如通过embl数据库中的blast程序(www.ncbi.nlm.gov/cgi-bin/blast),计算序列同一性。通常,可使用关于例如“评分矩阵”和“空位罚分”的缺省设置来进行比对。在本发明的
上下文中,blastn和psi blast缺省设置可能是有利的。
120.两个序列之间的同一性百分比可使用与上述类似的技术来确定,允许或不允许有空位。在计算同一性百分比时,只计数完全匹配处。因此,本发明的一种实施方式涉及的本发明的序列具有一定程度的序列变异。
121.用于结核疫苗的融合蛋白
122.据估计,世界上约有四分之一的人口感染了结核病,这种疾病导致每年超过100万人死亡。这使得结核病成为全球最致命的传染病。尽管如此,仍然没有开发出有效的疫苗来替代和增强一百年前开发的有争议且不可靠的卡介苗bcg疫苗。基于不同平台的策略,例如灭活的全细胞或全细胞提取物、基于病毒载体的疫苗、活的重组bcg、减毒bcg疫苗或dna疫苗,在很大程度上都未能提供令人鼓舞的结果,作出增强型结核疫苗的承诺。
123.另一种方法是设计融合蛋白,包括选择用于生产结核亚单位疫苗的抗原。这种类型的结核疫苗结核疫苗被认为具有一系列超越其他疫苗策略的优势,包括改善疫苗的安全性和稳定性。通过精心设计融合蛋白,可以诱导高质量t细胞的强的细胞免疫应答。因此,本文提供了可有效用在疫苗或免疫原性组合物中的融合蛋白,用于预防、抑制或治疗结核感染和/或疾病。
124.因此,本发明的一个方面涉及一种融合蛋白,其包含至少两种源自结核分枝杆菌的抗原。
125.结核分枝杆菌的基因组包含大约4000个基因,其中许多基因编码的蛋白质不适合作为抗结核感染或疾病的亚单位疫苗中的抗原,因为它们不能诱导足够强度或质量的免疫应答。因此,为了免疫原性,本文所述的融合蛋白的抗原被仔细地选择,包括但不限于rv3875(esat-6)、rv3873(ppe68)、rv3876(espi)、rv3615c(espc)、rv3616c(espa)、rv1980c(mpt64)、rv2875(mpt70)、rv2873(mpt83)、rv1886c(ag85b)、rv3804c(ag85a)、rv0288(tb10.4)、rv0287(esxg)、rv3478(ppe60)、rv0475(hbha)、rv3890c(esxc)、rv3891c(esxd)、rv1284(cana)、rv3019c(esxr)、rv3020c(esxs)、rv3017c(esxq)、rv2031c(hspx)、rv0983(pepd)、rv1196(ppe18)、rv2608(ppe42)、rv3619(esxv)、rv3620(esxw)、rv2660c、rv3614(espd)、rv3865(espf)、rv3849(espr)、rv3872(pe35)和rv3881(espb)。
126.因此,本发明的一种实施方式涉及如本文所述的融合蛋白,其中所述抗原选自:seq id no:1(esat-6)、seq id no:2(ppe68)、seq id no:3(espi)、seq id no:4(espc)、seq id no:5(espa)、seq id no:6(mpt64)、seq id no:7(mpt70)、seq id no:8(mpt83)、seq id no:10(ag85b)、seq id no:11(ag85a)、seq id no:12(tb10.4)、seq id no:13(esxg)、seq id no:14(ppe60)、seq id no:15(hbha)、seq id no:16(esxc)、seq id no:17(esxd)、seq id no:18(cana)、seq id no:19(esxr)、seq id no:20(esxs)、seq id no:21(esxq)、seq id no:22(hspx)、seq id no:23(pepd)、seq id no:24(ppe18)、seq id no:25(ppe42)、seq id no:26(esxv)、seq id no:27(esxw)、seq id no:28(rv2660c)、seq id no:29(rv3614)、seq id no:30(rv3865)、seq id no:31(rv3849)、seq id no:32(rv3872)和seq id no:33(rv3881),及其变体。
127.本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含至少三种抗原,例如至少四种抗原,例如至少五种抗原,例如至少六种抗原,例如至少七种抗原,例如至少八种抗原。
128.结核分枝杆菌感染的过程基本上经历了3个阶段。在急性期,细菌在器官中增殖,直到免疫应答增强。特异性致敏的cd4 t淋巴细胞介导对感染的控制,最重要的介质分子似乎是干扰素γ(ifn-γ)。细菌负荷开始下降,并建立了一个潜伏期/慢性期,其中细菌负荷在低水平保持稳定。在这个阶段,结核分枝杆菌从活跃增殖进入休眠,基本上变成非复制的并留在肉芽肿内。在一些情况下,感染进入再激活阶段,休眠细菌再次开始复制。
129.结核分枝杆菌从最初感染到潜伏期的过渡伴随着基因表达的变化。免疫应答的抗原特异性也可能发生变化,因为细菌在从活跃复制到休眠的过渡期间,细菌调节着基因表达。控制潜伏/慢性感染的免疫应答的全部性质以及导致再激活的因素尚未完全阐明。然而,有些证据表明负责的主要细胞类型发生了转变。虽然在急性期cd4 t细胞是必不可少的且足以控制感染,但研究表明在潜伏期/慢性期cd8 t细胞应答更为重要。
130.目前有几种新的tb疫苗正在临床试验中。然而,它们主要是经典的预防性疫苗,是基于在感染早期表达的有限数量的抗原。然而,作为上述表达动态的直接结果,呈现给t细胞的表位模式会随着时间发生根本性变化。因此,不受理论束缚,本文预期一种融合蛋白,包含分别在感染早期和晚期高表达的两种抗原,可用在一种强效疫苗中,该疫苗可(i)加速mtb感染后的免疫应答和(ii)诱导t细胞有可能在晚期慢性期特异性识别细菌。这种融合蛋白设计的好处还包括改善的表位覆盖以及靶向急性和潜伏/慢性感染的能力。
131.因此,本发明的一种实施方式涉及如本文所述的融合蛋白,其中该融合蛋白包含早期和晚期抗原。
132.本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述早期抗原选自:rv3875(esat-6)、rv3873(ppe68)、rv3876(espi)、rv3615c(espc)、rv3616c(espa)、rv1886c(ag85b)、rv3804c(ag85a)、rv0288(tb10.4)、rv0287(esxg)、rv3478(ppe60)、rv0475(hbha)、rv3890c(esxc)、rv3891c(esxd)、rv1284(cana)、rv3019c(esxr)、rv3020c(esxs)、rv3017c(esxq)、rv0983(pepd)、rv1196(ppe18)、rv2608(ppe42)、rv3619(esxv)、rv3620(esxw)、rv3614(espd)、rv3865(espf)、rv3849(espr)、rv3872(pe35)和rv3881(espb),以及它们的变体。
133.本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述晚期抗原选自rv2875(mpt70)、rv2873(mpt83)、rv2031c(hspx)和rv2660c,以及它们的变体。
134.在受结核病困扰的国家,大多数人口都接种了bcg——一种来自牛分枝杆菌的活的、减毒结核菌菌株。针对结核病增强保护作用的一种普遍策略是增强作为对初始bcg疫苗接种的反应而产生的免疫应答。这可以通过施用结核抗原来寻求实现,这些抗原被bcg表达并且在初始bcg疫苗接种后就已经引发了免疫应答。此类抗原在本文中称为bcg
抗原并且可作为亚单位疫苗的一部分施用,目的是增强由初始bcg疫苗接种诱导的免疫应答。然而,分枝杆菌感染,包括bcg疫苗接种和mtb感染,诱导t细胞成为质量差的t细胞,并且已经证实它们在用亚单位疫苗加强后也极难程序重调(reprogram),结果令人失望。
135.本文公开了在概念上不同于上述bcg
加强疫苗的融合蛋白变体。令人惊奇的是,已发现对bcg呈功能性阴性的结核抗原(即在bcg疫苗接种后不引发免疫应答的抗原)可用在融合蛋白中以提供显著增强的抗结核分枝杆菌的保护作用。这类抗原在本文中被称为bcg-抗原并且可用在融合蛋白中作为独立疫苗的一部分或与bcg组合以引发一种互补免疫应答,该互补免疫应答针对的bcg抗原不同于最初bcg疫苗接种的抗原。
136.不受理论束缚,可以预期,至少部分增加的保护作用是由于本文公开的融合蛋白所诱导的免疫应答的质量。bcg疫苗接种主要是一种分枝杆菌感染,由于慢性抗原刺激,它会诱导短寿命的t细胞,这些t细胞只能在很差的程度上穿透肺组织并对抗感染。bcg疫苗接种的最初免疫应答留下的这种足迹或免疫学遗产随后很难改变。本文公开的融合蛋白是基于bcg-抗原的变体形式,其不受这种免疫学遗产的限制,因此可以绕过t细胞质量障碍;基于bcg
抗原的传统亚单位疫苗通常受到该障碍的限制。
137.因此,本文所述的融合蛋白的变体是基于不引发针对bcg的免疫应答的抗原,即bcg-抗原。与现有的结核疫苗相比,本文提供的融合蛋白产生改善的免疫应答。
138.因此,本发明的一种实施方式涉及如本文所述的融合蛋白,其中至少一种抗原不引发针对bcg的免疫应答。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中至少一种抗原对于bcg是功能阴性的。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中至少一种抗原是bcg-抗原。
139.需要注意的是,存在多种不同的bcg菌株,不同菌株之间的基因型和表型不同。因此,为了确定抗原是bcg
或bcg-抗原,本文参考晚期bcg菌株,优选bcg danish、bcg pasteur、bcg prague或vpm1002。因此,本发明的一种实施方式涉及如本文所述的融合蛋白,其中bcg选自晚期bcg菌株,优选bcg danish、bcg pasteur或bcg prague。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中至少一种抗原不引发针对晚期bcg菌株的免疫应答。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中至少一种抗原不引发针对bcg菌株的免疫应答,所述bcg菌株选自bcg danish、bcg pasteur和bcg prague。
140.晚期bcg菌株还可具有以下特征:(i)缺失一些由已删除的rd2区编码的抗原;(ii)其他抗原表达差或不分泌。因此,对于这些缺失、表达差或不分泌的抗原,在接种晚期bcg菌株后不会诱导免疫应答。因此,本发明的一种实施方式涉及如本文所述的融合蛋白,其中抗原从bcg中缺失、不分泌或具有低表达。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中抗原在晚期bcg菌株中缺失、不分泌或具有低表达,优选地所述晚期bcg菌株选自bcg danish、bcg pasteur和bcg prague。
141.本发明的另一种实施方式涉及如本文所述的融合蛋白,其中抗原选自位于rd1内的抗原、位于rd2内的抗原和其表达受sigk调节的抗原,以及它们的组合。
142.虽然结核分枝杆菌的基因组包含大约4000个基因,但与bcg的基因组有约98%的大量重叠。因此,潜在的bcg-抗原的数量被限制在大约100种抗原,bcg不引发针对这些抗原的t细胞反应。本文鉴定了适于包含在融合蛋白中用于抗结核感染和/或疾病的疫苗中的bcg-抗原。因此,本发明的一种实施方式涉及如本文所述的融合蛋白,其中抗原选自:rv3875(esat-6)、rv3873(ppe68)、rv3876(espi)、rv3615c(espc)、rv3616c(espa)、rv1980c(mpt64)、rv2875(mpt70)和rv2873(mpt83),及其变体。应当理解,本文所述的抗原、多肽或融合蛋白的变体可具有相同的免疫原性,即它们在诱导免疫应答方面同样有效,或至少有与其参照的抗原、多肽或融合蛋白相同的免疫原性。本发明的又一种实施方式涉及如本文所述的融合蛋白,其中抗原选自:
143.a)氨基酸序列,选自seq id no:1(esat-6)、seq id no:2(ppe68)、seq id no:3(espi)、seq id no:4(espc)、seq id no:5(espa)、seq id no:6(mpt64)、seq id no:7(mpt70)和seq id no:8(mpt83),及其变体或免疫原性表位,或者
144.b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。
145.已经发现一些抗原片段是有利的。因此,本发明的一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含选自以下的氨基酸序列:
146.a)seq id no:34(h107b)、seq id no:35(h107c)或seq id no:91(h107e),或其变体或免疫原性表位,或
147.b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。
148.本发明的一种优选实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含:
149.a)seq id no:1(esat-6)、seq id no:2(ppe68)、seq id no:3(espi)、seq id no:4(espc)和seq id no:5(espa),或其变体或免疫原性表位,或者
150.b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。
151.本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含选自以下的氨基酸序列:
152.a)seq id no:36(h106)或seq id no:38(h104),或其变体或免疫原性表位,或者
153.b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。
154.本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含:
155.a)seq id no:1(esat-6)、seq id no:2(ppe68)、seq id no:3(espi)、seq id no:4(espc)、seq id no:5(espa)、seq id no:6(mpt64)、seq id no:7(mpt70)和seq id no:8(mpt83),或其变体或免疫原性表位,或者
156.b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。
157.本发明的另一种实施方式涉及一种如本文所述的融合蛋白,其中所述融合蛋白包含选自以下的氨基酸序列:
158.a)seq id no:37(h105)或其变体或免疫原性表位,或者
159.b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。
160.本发明的另一种实施方式涉及如本文所述的融合蛋白,其中b)的氨基酸序列与a)的任一氨基酸序列具有至少90%的序列同一性,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。应当理解,与本文所述的一种抗原、多肽
或融合蛋白具有确定的序列同一性的抗原、多肽或融合蛋白可具有相同的免疫原性,即它们在诱导免疫应答方面同样有效,或至少与它所参照的抗原、多肽或融合蛋白具有相同的免疫原性。
161.如本文所述,可以预期用早期和晚期抗原进行疫苗接种产生改善的免疫应答。本文从有限数量的bcg-抗原中鉴定出早期和晚期抗原。因此,本发明的一种优选实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含早期抗原和晚期抗原,它们都不引发针对bcg的免疫应答(即bcg-抗原)。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中不引发针对bcg的免疫应答的晚期抗原是rv2875(mpt70)和/或rv2873(mpt83),及其变体。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中不引发针对bcg的免疫应答的早期抗原选自rv3875(esat-6)、rv3873(ppe68)、rv3876(espi)、rv3615c(espc)和rv3616c(espa),及其变体。
162.本文公开的融合蛋白的一些变体限于仅包含bcg-抗原。因此,本发明的一个实施方案涉及如本文所述的融合蛋白,其中所述融合蛋白仅包含不引发针对bcg的免疫反应的抗原(即bcg-抗原)。
163.本文所述的融合蛋白也可通过改变6kda早期分泌抗原靶点(esat-6)的含量来改进。esat-6是结核分枝杆菌重要的分泌蛋白和强效t细胞抗原。esat-6蛋白是由该细菌组成性表达并大量分泌,在人和动物中感染后均可被识别。因此,esat-6被认为是一种重要的抗原,可被有利地包含在针对结核感染和疾病的亚单位疫苗中。然而,由于天然esat-6分子是一种只有95个氨基酸的小蛋白质,因此在人和动物中针对esat-6产生的免疫应答程度相对较低。
164.本文公开了包含多个esat-6重复的融合蛋白的变体,因此esat-6相对地代表了融合蛋白的较大部分。不受理论束缚,本文预期,增加融合蛋白中esat-6抗原的相对含量减轻了具有低免疫原性的天然esat-6的挑战。对于包含一个以上esat-6拷贝的融合蛋白,每次出现的抗原称为esat-6重复,例如包含四个esat-6抗原拷贝的融合蛋白可以说包含四个esat-6重复。因此,本发明的一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含至少两个esat-6重复,例如至少三个esat-6重复,例如至少四个esat-6重复,例如至少五个esat-6重复。本发明的一种优选实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含至少四个esat-6重复。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含四个esat-6重复。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中每个esat-6重复由seq id no:1或与seq id no:1具有至少80%的序列同一性(例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性)的氨基酸序列表示。
165.esat-6重复原则上可以任何顺序分布在融合蛋白中,例如在c端、n端、连续地、交替地或隔开的。本发明的一种实施方式涉及如本文所述的融合蛋白,其中esat-6重复被至少一种不同于esat-6的抗原分开。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中esat-6重复与不同于esat-6的抗原交替定位。
166.含有esat-6重复的融合蛋白中适用的抗原优选具有高度免疫原性并且在结核感染后在人和动物中均被识别。因此,适于与esat-6重复一起用于融合蛋白中的抗原包括但不限于:rv3873(ppe68)、rv3876(espi)、rv3615c(espc)、rv3616c(espa)、rv1980c(mpt64)、
rv2875(mpt70)、rv2873(mpt83)、rv1886c(ag85b)、rv3804c(ag85a)、rv0288(tb10.4)、rv0287(esxg)、rv3478(ppe60)、rv0475(hbha)、rv3890c(esxc)、rv3891c(esxd)、rv1284(cana)、rv3019c(esxr)、rv3020c(esxs)、rv3017c(esxq)、rv2031c(hspx)、rv0983(pepd)、rv1196(ppe18)、rv2608(ppe42)、rv3619(esxv)、rv3620(esxw)、rv2660c、rv3614(espd)、rv3865(espf)、rv3849(espr)、rv3872(pe35)和rv3881(espb)。
167.因此,本发明的一种实施方式涉及如本文所述的融合蛋白,其中抗原选自seq id no:2(ppe68)、seq id no:3(espi)、seq id no:4(espc)、seq id no:5(espa)、seq id no:6(mpt64)、seq id no:7(mpt70)、seq id no:8(mpt83)、seq id no:10(ag85b)、seq id no:11(ag85a)、seq id no:12(tb10.4)、seq id no:13(esxg)、seq id no:14(ppe60)、seq id no:15(hbha)、seq id no:16(esxc)、seq id no:17(esxd)、seq id no:18(cana)、seq id no:19(esxr)、seq id no:20(esxs)、seq id no:21(esxq)、seq id no:22(hspx)、seq id no:23(pepd)、seq id no:24(ppe18)、seq id no:25(ppe42)、seq id no:26(esxv)、seq id no:27(esxw)、seq id no:28(rv2660c)、seq id no:29(rv3614)、seq id no:30(rv3865)、seq id no:31(rv3849)、seq id no:32(rv3872)和seq id no:33(rv3881),及其变体,并且其中所述融合蛋白包含至少两个esat-6重复,例如至少三个esat-6重复,例如至少四个esat-6重复,例如至少五个esat-6重复。
168.如本文所述的融合蛋白还包括含有早期和晚期两种抗原或bcg-抗原与esat-6重复相结合的融合蛋白。因此,本发明的一种优选实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含早期和晚期两种抗原,以及至少两个esat-6重复,例如至少三个esat-6重复,例如至少四个esat-6重复,例如至少五个esat-6重复。本发明的另一种优选实施方式涉及如本文所述的融合蛋白,其中至少一种抗原不引发针对bcg的免疫应答,并且其中所述融合蛋白包含至少两个esat-6重复,例如至少三个esat-6重复,例如至少四个esat-6重复,例如至少五个esat-6重复。
169.本发明的一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含:
170.a)seq id no:1(esat-6)、seq id no:2(ppe68)、seq id no:3(espi)、seq id no:4(espc)、seq id no:5(espa)、seq id no:6(mpt64)、seq id no:7(mpt70)和seq id no:8(mpt83),或其变体或免疫原性表位,或者
171.b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性,并且
172.其中所述融合蛋白包含与不同于esat-6的抗原交替定位的四个esat-6重复。
173.本发明的另一种特别优选的实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含早期和晚期两种bcg-抗原与esat-6重复相结合。这种融合蛋白的一个变体称为h107(seq id no:9)。因此,本发明的一种优选实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包括由seq id no:9表示的氨基酸序列。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含:
174.a)seq id no:9(h107)或其变体或免疫原性表位,或
175.b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同
一性,并且
176.其中b)的氨基酸序列具有与seq id no:9相同的免疫原性或至少相同的免疫原性。
177.h107融合蛋白的变体称为h107e(seq id no:91)。因此,本发明的一种优选实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含由seq id no:91表示的氨基酸序列。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含:
178.a)seq id no:91(h107e)或其变体或免疫原性表位,或
179.b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性,并且
180.其中b)的氨基酸序列具有与seq id no:91相同的免疫原性或至少相同的免疫原性。
181.本文所述的融合蛋白的抗原可通过接头连接,例如肽接头。因此,本发明的一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白的抗原与接头分子连接。
182.为了能够产生融合蛋白或抗原,它们可包含适当的纯化标签以允许从例如重组表达系统中纯化。因此,本发明的一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含纯化标签。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述纯化标签选自his-标签、几丁质结合蛋白(cbp)、麦芽糖结合蛋白(mbp)和谷胱甘肽-s-转移酶(gst)。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述纯化标签是his-标签。
183.融合蛋白的变体可包括bcg
抗原。因此,本发明的一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含至少一种引发针对bcg的免疫应答的抗原。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述融合蛋白包含至少一种bcg
抗原。本发明的又一种实施方式涉及如本文所述的融合蛋白,其中所述至少一种引发针对bcg的免疫应答的抗原选自rv1886c(ag85b)、rv3804c(ag85a)、rv0288(tb10.4)、rv0287(esxg)、rv3478(ppe60)、rv0475(hbha)、rv3890c(esxc)、rv3891c(esxd)、rv1284(cana)、rv3019c(esxr)、rv3020c(esxs)、rv3017c(esxq)、rv2031c(hspx)、rv0983(pepd)、rv1196(ppe18)、rv2608(ppe42)、rv3619(esxv)和rv3620(esxw),及其变体。本发明的另一种实施方式涉及如本文所述的融合蛋白,其中所述至少一种引发针对bcg的免疫应答的抗原选自:
184.a)氨基酸,选自seq id no:10(ag85b)、seq id no:11(ag85a)、seq id no:12(tb10.4)、seq id no:13(esxg)、seq id no:14(ppe60)、seq id no:15(hbha)、seq id no:16(esxc)、seq id no:17(esxd)、seq id no:18(cana)、seq id no:19(esxr)、seq id no:20(esxs)、seq id no:21(esxq)、seq id no:22(hspx)、seq id no:23(pepd)、seq id no:24(ppe18)、seq id no:25(ppe42)、seq id no:26(esxv)和seq id no:27(esxw),及其变体或免疫原性表位,或者
185.b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。
186.本文公开的融合蛋白是有免疫原性的并且可有效地用于疫苗或免疫原性组合物
中以预防、抑制或治疗结核感染和/或疾病。这种疫苗或免疫原性组合物可以是预防性或治疗性地使用。因此,本发明的一个方面涉及包含如本文所述的融合蛋白的疫苗或免疫原性组合物。
187.为了确保疫苗或免疫原性组合物的最佳性能,优选的是它包含免疫学和药学上可接受的载体、媒剂或佐剂。因此,本发明的一种实施方式涉及如本文所述的疫苗或免疫原性组合物,其中所述疫苗或免疫原性组合物还包含免疫学和药学上可接受的载体、媒剂或佐剂。本发明的另一种实施方式涉及如本文所述的疫苗或免疫原性组合物,其中所述疫苗或免疫原性组合物还包含一种或多种佐剂。本发明的另一种实施方式涉及如本文所述的疫苗或免疫原性组合物,其中所述佐剂选自中性佐剂制剂、阴离子佐剂制剂、阳离子佐剂制剂、阳离子脂质体(例如二甲基二(十八烷基)溴化铵(dda)、quil a、qs21、聚i:c、氢氧化铝、弗氏不完全佐剂、ifn-γ、il-2、il-12、单磷酰脂a(mpl)、海藻糖二霉酸酯(tdm)、海藻糖二山嵛酸酯(tdb)、胞壁酰二肽(mdp)、单分枝酰基甘油(mmg)、cpc和“ic31”,或其组合。本发明的一种实施方式涉及如本文所述的疫苗或免疫原性组合物,其中所述佐剂为阳离子佐剂制剂1(caf01)。在另一种优选实施方式中,所述佐剂为包含dda、mmg和cpg的阳离子佐剂制剂10(caf10)。可在本发明的疫苗或免疫原性组合物中用作佐剂的其他阳离子佐剂制剂如下表1中所列:
188.表1caf佐剂命名法
[0189][0190]
缩写:dda:n,n-二甲基-n,n-二(十八烷基)铵(溴化物盐)。dspc:1,2-二硬脂酰-sn-甘油-3-磷酸胆碱。dspe-peg:1,2-二硬脂酰-sn-甘油-3-磷酸乙醇胺-n-[羧基(聚乙二醇)-2000](钠盐)。tdb:a,a-海藻糖6,6
’‑
二山嵛酸酯。mmg:合成的单分枝酰基甘油。聚-ic:聚肌苷酸-聚胞苷酸(钠盐)。cpg:5
’‑
c-磷酸-g-3’寡核苷酸。mpl:单磷酰脂质a。
[0191]
本文公开的融合蛋白可用于独立疫苗或与bcg疫苗组合使用。因此,本发明的一种实施方式涉及如本文所述的疫苗或免疫原性组合物,其中所述疫苗或免疫原性组合物还包含bcg。
[0192]
本发明的另一个方面涉及如本文所述的融合蛋白或如本文所述的疫苗或免疫原性组合物,用于针对由毒性分枝杆菌引起的感染和/或疾病,对受试者进行疫苗接种或免
疫。
[0193]
毒性分枝杆菌可感染多种多样的动物,并且在某些情况下会对例如农场动物造成挑战。因此,本发明的一种实施方式涉及如本文所述使用的融合蛋白、疫苗或免疫原性组合物,其中受试者是哺乳动物。然而,提供本文公开的融合蛋白的主要目的是改善当前对抗人类结核感染和/或疾病的全球挑战的可能性。因此,本发明的一种优选实施方式涉及如本文所述使用的融合蛋白、疫苗或免疫原性组合物,其中哺乳动物是人。
[0194]
结核可由不同的毒性分枝杆菌引起。因此,本发明的一种实施方式涉及如本文所述使用的融合蛋白、疫苗或免疫原性组合物,其中毒性分枝杆菌选自结核分枝杆菌(m.tuberculosis)、牛分枝杆菌(m.bovis)、非洲分枝杆菌(m.africanum)、卡内蒂分枝杆菌(m.canetti)、和田鼠分枝杆菌(m.microti),优选结核分枝杆菌(m.tuberculosis)。本发明的一种优选实施方式涉及如本文所述使用的融合蛋白、疫苗或免疫原性组合物,其中毒性分枝杆菌是结核分枝杆菌。
[0195]
应用的方式可以多种多样。任何用于疫苗施用的常规方法都是适用的,包括但不限于:口服制剂、栓剂、肠胃外,和通过注射,例如皮下或肌肉内。本发明的一种实施方式涉及如本文所述使用的融合蛋白、疫苗或免疫原性组合物,其中所述融合蛋白、疫苗或免疫原性组合物通过选自以下的途径施用:口服、肠胃外、皮下和肌肉内。疫苗的剂量将取决于施用途径,并根据待接种疫苗的人的年龄、以及在较小程度上根据待接种疫苗的人的体型有所不同。
[0196]
如本文所述,融合蛋白、疫苗或免疫原性组合物可有利地与bcg疫苗组合施用。因此,本发明的一种实施方式涉及如本文所述使用的融合蛋白、疫苗或免疫原性组合物,其中在施用融合蛋白、疫苗或免疫原性组合物之前、同时或之后施用bcg。令人惊讶地发现,当与本文公开的融合蛋白、疫苗或免疫原性组合物同时施用时,bcg可提供强佐剂作用,产生高质量的强免疫应答。因此,本发明的一种优选实施方式涉及如本文所述使用的融合蛋白、疫苗或免疫原性组合物,其中bcg的施用与融合蛋白、疫苗或免疫原性组合物的施用同时进行。本发明的另一种实施方式涉及如本文所述使用的融合蛋白、疫苗或免疫原性组合物,其中受试者先前已接种bcg疫苗。
[0197]
由于本文公开的融合蛋白、疫苗或免疫原性组合物可与bcg疫苗协同作用,因此它们可方便地一起提供以方便应用。因此,本发明的一个方面涉及一种试剂盒,包括:
[0198]
i)本文所述的融合蛋白或本文所述的疫苗或免疫原性组合物,
[0199]
ii)bcg,和
[0200]
iii)可选地,使用说明。
[0201]
本发明的一种实施方式涉及如本文所述的试剂盒,其中i)和ii)用于同时、分开或顺序施用。
[0202]
本文公开的融合蛋白可通过设计编码融合蛋白的表达载体构建体并将表达载体引入适宜的重组表达系统来重组生产。因此,本发明的一个方面涉及一种核酸序列,其包含编码如本文所述的融合蛋白的序列。
[0203]
本发明的另一个方面涉及一种重组表达载体,其包含如本文所述的核苷酸序列,所述核苷酸序列可操作地连接至一个或多个适于指导在适宜宿主中产生融合蛋白的控制序列。
[0204]
本发明的另一方面涉及一种重组宿主细胞,其包含如本文所述的表达载体。
[0205]
适用于生产本文公开的融合蛋白的表达载体可包括选自以下组的核酸:seq id no:46(esat-6)、seq id no:47(ppe68)、seq id no:48(espi)、seq id no:49(espc)、seq id no:50(espa)、seq id no:51(mpt64)、seq id no:52(mpt70)、seq id no:53(mpt83)、seq id no:55(ag85b)、seq id no:56(ag85a)、seq id no:57(tb10.4)、seq id no:58(esxg)、seq id no:59(ppe60)、seq id no:60(hbha)、seq id no:61(esxc)、seq id no:62(esxd)、seq id no:63(cana)、seq id no:64(esxr)、seq id no:65(esxs)、seq id no:66(esxq)、seq id no:67(hspx)、seq id no:68(pepd)、seq id no:69(ppe18)、seq id no:70(ppe42)、seq id no:71(esxv)、seq id no:72(esxw)、seq id no:73(rv2660c)、seq id no:74(rv3614)、seq id no:75(rv3865)、seq id no:76(rv3849)、seq id no:77(rv3872)和seq id no:78(rv3881),及其变体。
[0206]
此外,选择的融合蛋白可由选自以下的核酸编码:seq id no:54(h107)、seq id no:79(h107b)、seq id no:80(h107c)、seq id no:92(h107e)、seq id no:81(h106)、seq id no:82(h105)和seq id no:83(h104),及其变体。
[0207]
表2概述显示抗原和融合蛋白以及序列号和表达模式:
[0208]
[0209]
[0210][0211]
本技术中引用的所有专利和非专利参考文献均将其全部内容引入本文作为参考。
[0212]
应当注意,在本发明的一个方面的上下文中描述的实施方式和特征也适用于本发明的其他方面。本发明的实施方式和特征在以下项目中也进行了概述。
[0213]
项目
[0214]
1、一种融合蛋白,包含至少两种源自结核分枝杆菌的抗原。
[0215]
2、根据项目1所述的融合蛋白,其中所述融合蛋白包含至少三种抗原,例如至少四种抗原,例如至少五种抗原,例如至少六种抗原,例如至少七种抗原,例如至少八种抗原。
[0216]
3、根据项目1或2任一项所述的融合蛋白,其中所述融合蛋白包含早期抗原和晚期抗原。
[0217]
4、根据前述项目任一项所述的融合蛋白,其中至少一种抗原不引发针对bcg的免疫应答。
[0218]
5、根据前述项目任一项所述的融合蛋白,其中所述抗原从bcg中缺失、不分泌或在bcg中具有低表达。
[0219]
6、根据前述项目任一项所述的融合蛋白,其中所述抗原选自:
[0220]
rv3875(esat-6)、rv3873(ppe68)、rv3876(espi)、rv3615c(espc)、rv3616c(espa)、rv1980c(mpt64)、rv2875(mpt70)和rv2873(mpt83),及其变体。
[0221]
7、根据前述项目任一项所述的融合蛋白,其中所述抗原选自:
[0222]
a)氨基酸序列,选自seq id no:1(esat-6)、seq id no:2(ppe68)、seq id no:3(espi)、seq id no:4(espc)、seq id no:5(espa)、seq id no:6(mpt64)、seq id no:7(mpt70)和seq id no:8(mpt83),及其变体或免疫原性表位,或者
[0223]
b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同
一性。
[0224]
8、根据前述项目任一项所述的融合蛋白,其中所述融合蛋白包含:
[0225]
a)seq id no:1(esat-6)、seq id no:2(ppe68)、seq id no:3(espi)、seq id no:4(espc)和seq id no:5(espa),或其变体或其免疫原性表位,或者
[0226]
b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。
[0227]
9、根据前述项目任一项所述的融合蛋白,其中所述融合蛋白包含:
[0228]
a)seq id no:1(esat-6)、seq id no:2(ppe68)、seq id no:3(espi)、seq id no:4(espc)、seq id no:5(espa)、seq id no:6(mpt64)、seq id no:7(mpt70)和seq id no:8(mpt83),或其变体或免疫原性表位,或者
[0229]
b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。
[0230]
10、根据项目7-9任一项所述的融合蛋白,其中b)的氨基酸序列与a)的任一氨基酸序列具有至少90%的序列同一性,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。
[0231]
11、根据前述项目任一项所述的融合蛋白,其中所述融合蛋白包含至少两个esat-6重复,例如至少三个esat-6重复,例如至少四个esat-6重复,例如至少五个esat-6重复。
[0232]
12、根据前述项目任一项所述的融合蛋白,其中所述融合蛋白包含至少四个esat-6重复。
[0233]
13、根据项目11或12任一项所述的融合蛋白,其中所述esat-6重复被至少一种不同于esat-6的抗原隔开。
[0234]
14、根据项目11-13任一项所述的融合蛋白,其中所述esat-6重复与不同于esat-6的抗原交替定位。
[0235]
15、根据前述项目任一项所述的融合蛋白,其中所述融合蛋白包含:
[0236]
a)seq id no:1(esat-6)、seq id no:2(ppe68)、seq id no:3(espi)、seq id no:4(espc)、seq id no:5(espa)、seq id no:6(mpt64)、seq id no:7(mpt70)和seq id no:8(mpt83),或其变体或免疫原性表位,或者
[0237]
b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性;并且
[0238]
其中所述融合蛋白包含与不同于esat-6的抗原交替定位的四个esat-6重复。
[0239]
19、根据前述权利要求任一项所述的融合蛋白,其中所述融合蛋白包含选自以下的任一项所表示的氨基酸序列:
[0240]
a)seq id no:9、seq id no:34、seq id no:35、seq id no:36、seq id no:37、seq id no:38或seq id no:91,或其变体或免疫原性表位;或者
[0241]
b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同
一性。
[0242]
20、根据前述项目任一项所述的融合蛋白,其中所述融合蛋白包含由seq id no:9或seq id no:91表示的氨基酸序列。
[0243]
21、根据权利要求任一项所述的融合蛋白,其中所述融合蛋白由选自以下的核酸序列编码:
[0244]
a)seq id no:54(h107)、seq id no:79(h107b)、seq id no:80(h107c)、seq id no:92(h107e)、seq id no:81(h106)、seq id no:82(h105)和seq id no:83(h104),或其变体或片段;或者
[0245]
b)与a)的任一氨基酸序列具有至少80%序列同一性的核酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。
[0246]
22、根据前述项目任一项所述的融合蛋白,其中所述融合蛋白的抗原与接头分子连接。
[0247]
23、根据前述项目任一项所述的融合蛋白,其中所述融合蛋白包含至少一种引发针对bcg的免疫应答的抗原。
[0248]
24、根据项目23所述的融合蛋白,其中所述至少一种引发针对bcg的免疫应答的抗原选自rv1886c(ag85b)、rv3804c(ag85a)、rv0288(tb10.4)、rv0287(esxg)、rv3478(ppe60)、rv0475(hbha)、rv3890c(esxc)、rv3891c(esxd)、rv1284(cana)、rv3019c(esxr)、rv3020c(esxs)、rv3017c(esxq)、rv2031c(hspx)、rv0983(pepd)、rv1196(ppe18)、rv2608(ppe42)、rv3619(esxv)和rv3620(esxw),及其变体。
[0249]
25、根据项目23或24任一项所述的融合蛋白,其中所述至少一种引发针对bcg的免疫应答的抗原选自:
[0250]
a)氨基酸,选自seq id no:10(ag85b)、seq id no:11(ag85a)、seq id no:12(tb10.4)、seq id no:13(esxg)、seq id no:14(ppe60)、seq id no:15(hbha)、seq id no:16(esxc)、seq id no:17(esxd)、seq id no:18(cana)、seq id no:19(esxr)、seq id no:20(esxs)、seq id no:21(esxq)、seq id no:22(hspx)、seq id no:23(pepd)、seq id no:24(ppe18)、seq id no:25(ppe42)、seq id no:26(esxv)和seq id no:27(esxw),及其变体或免疫原性表位,或者
[0251]
b)与a)的任一氨基酸序列具有至少80%序列同一性的氨基酸序列,例如至少90%,例如至少95%,例如至少96%,例如至少97%,例如至少98%,例如至少99%的序列同一性。
[0252]
26、一种疫苗或免疫原性组合物,包含前述项目任一项所述的融合蛋白。
[0253]
27、根据项目26所述的疫苗或免疫原性组合物,其中所述疫苗或免疫原性组合物还包含一种或多种佐剂。
[0254]
28、根据项目27所述的疫苗或免疫原性组合物,其中所述佐剂选自中性佐剂制剂、阴离子佐剂制剂、阳离子佐剂制剂、阳离子脂质体(例如二甲基二(十八烷基)溴化铵(dda)、quil a、qs21、聚i:c、氢氧化铝、弗氏不完全佐剂、ifn-γ、il-2、il-12、单磷酰脂a(mpl)、海藻糖二霉酸酯(tdm)、海藻糖二山嵛酸酯(tdb)、胞壁酰二肽(mdp)、单分支酰基甘油(mmg)、cpg和“ic31”,或其组合。
[0255]
29、根据项目26-28任一项所述的疫苗或免疫原性组合物,其中所述疫苗或免疫原
性组合物还包含bcg。
[0256]
30、根据项目1-25任一项所述的融合蛋白或根据项目26-29任一项所述的疫苗或免疫原性组合物,用于针对由毒性分枝杆菌引起的感染和/或疾病,对受试者进行疫苗接种或免疫。
[0257]
31、根据项目30所述使用的融合蛋白、疫苗或免疫原性组合物,其中所述受试者是哺乳动物。
[0258]
32、根据项目第31所述使用的融合蛋白、疫苗或免疫原性组合物,其中所述哺乳动物是人。
[0259]
33、根据项目30-32任一项所述使用的融合蛋白、疫苗或免疫原性组合物,其中所述毒性分枝杆菌选自结核分枝杆菌、牛分枝杆菌、非洲分枝杆菌、卡内蒂分枝杆菌和田鼠分枝杆菌,优选结核分枝杆菌。
[0260]
34、根据项目30-33任一项所述使用的融合蛋白、疫苗或免疫原性组合物,其中在施用所述融合蛋白、疫苗或免疫原性组合物之前、同时或之后施用bcg。
[0261]
35、根据项目34所述使用的融合蛋白、疫苗或免疫原性组合物,其中在施用所述融合蛋白、疫苗或免疫原性组合物的同时施用bcg。
[0262]
36、根据项目30-35任一项所述使用的融合蛋白、疫苗或免疫原性组合物,其中所述受试者先前已接种bcg。
[0263]
37、一种试剂盒,包括:
[0264]
i)根据项目1-25任一项所述的融合蛋白或根据项目26-29任一项所述的疫苗或免疫原性组合物,
[0265]
ii)bcg,和
[0266]
iii)可选地,使用说明。
[0267]
38、根据项目37所述的试剂盒,其中i)和ii)用于同时、分开或顺序施用。
[0268]
39、一种核酸序列,包含编码项目1-25任一项所述的融合蛋白的序列。
[0269]
40、一种重组表达载体,包含项目39所述的核苷酸序列,该核苷酸序列可操作地连接至一个或多个适于指导在适宜宿主中产生融合蛋白的控制序列。
[0270]
41、一种重组宿主细胞,包含项目40所述的表达载体。
[0271]
现在将在以下非限制性实施例中更详细地描述本发明。
[0272]
实施例
[0273]
实施例1:bcg疫苗接种不引发针对h104-h107融合蛋白的结核分枝杆菌抗原的免疫应答
[0274]
材料和方法
[0275]
六至十周龄雌性cb6f1小鼠用牛分枝杆菌bcg danish免疫,或通过气雾剂途径感染25-50cfu毒性mtb erdman,或用2μg单蛋白质免疫3次,单蛋白质配制在总体积为200μl的阳离子佐剂制剂1(caf01)中。每种蛋白质疫苗间隔两周。bcg免疫后10周或mtb感染后18周或最后一次单蛋白质免疫后2周,通过使脾脏穿过100μm细胞过滤器,从各个动物获得脾细胞的单细胞悬液。将细胞在rpmi中洗涤两次,然后测定从每个脾脏中获得的细胞数并调整为2
×
106细胞/ml。为了刺激细胞,将100μl(2
×
105个细胞)等分到96孔板的各个孔中,并加入纯化的重组单mtb抗原(2μg/ml)。使用rpmi培养基将每孔的总体积调整到200μl,然后将
细胞与各个抗原在37℃于co2培养箱中孵育3-5天。在该时间点,通过夹心elisa测量细胞上清液中细胞因子ifn-γ的浓度。
[0276]
结果
[0277]
选择一组潜在的候选抗原用于设计具有非-bcg(bcg-)抗原的亚单位疫苗,并研究它们是否在bcg疫苗接种后引起免疫应答。用纯化为重组蛋白的指定单抗原(2μg/ml)刺激从小鼠分离的脾细胞(也参见实施例2)。刺激后,测量细胞释放到培养基中的细胞因子ifn-γ。在无抗原孵育的细胞(“无”)培养基中测量的细胞因子ifn-γ浓度建立了非刺激(non-stimulated)细胞的基线。在bcg免疫的动物中,没有一种候选抗原引起免疫应答(图1a)。相反,由bcg表达的tb10.4抗原引起显著的免疫应答(图1a)。在mtb感染的动物中,所有抗原都引起免疫应答,证实这些抗原是有免疫原性的并在mtb感染期间被表达(图1b)。类似地,在接种了单蛋白质的动物中,所有抗原都引起免疫应答,表明这些抗原在cb6f1小鼠品系中有免疫原性(图1c)。
[0278]
结论
[0279]
针对八种候选疫苗抗原的任何一种,牛分枝杆菌bcg danish免疫均未诱导细胞免疫应答。相反,在mtb感染或单蛋白质免疫后,所有候选抗原都引起免疫应答。因此,h104-h107的抗原不与牛分枝杆菌bcg danish共享抗原,并且对mtb有特异性。在本文中这样的抗原被称为bcg-抗原。相反,那些bcg免疫引起t细胞应答的抗原在本文中被称为bcg
抗原,例如tb10.4。
[0280]
实施例2:融合蛋白的构建
[0281]
材料与方法
[0282]
基于从已发表的结核分枝杆菌菌株h37rv的基因组序列翻译的蛋白质序列,合成了编码单一和融合蛋白的基因。合成的dna序列经过密码子优化以在大肠杆菌中表达,并在诱导型启动子t7启动子的控制下克隆到pj411表达载体中并转化到大肠杆菌中。pj411载体在框内编码n端his标签,框内带有编码序列用于插入的重组蛋白。对于每种蛋白质,当体外培养达到~0.5od
600
的密度时,会诱导带有his标签的重组蛋白的表达。第二天,将细菌收获、裂解,将重组蛋白用三步纯化过程进行纯化。这些过程包括从沉淀的重组蛋白(包涵体)中去除可溶性大肠杆菌蛋白、固定化金属亲和层析以及根据蛋白质电荷进行阴离子或阳离子交换。纯化后,通过质谱确认纯化的蛋白质的身份,并通过扫描考马斯染色的sds-凝胶确认纯度。最后,测定蛋白质浓度。
[0283]
结果
[0284]
基于实施例1的结果,设计了带有bcg-抗原的融合蛋白。重组融合蛋白的选择如(图2和3)中所示。从6l起始培养物中纯化的融合蛋白的产量在0.6mg到20mg之间不等。纯化的蛋白质的浓度在0.2mg/ml到0.7mg/ml之间不等。纯度为95%或更高。
[0285]
结论
[0286]
所有单一和融合蛋白都以足够的数量和质量产生,以成为适用于疫苗的抗原并执行以下实施例中描述的所有需要的免疫和细胞刺激。
[0287]
实施例3:mtp70是一种在晚期mtb感染期间表达上调的抗原(即“晚期抗原”)
[0288]
材料与方法
[0289]
用25-50cfu毒性mtb erdman气溶胶感染雌性cb6f1小鼠。在三、十二和二十周后处
死小鼠进行免疫分析。将肺使用t型管gentlemacs(miltenyi biotec)轻度匀浆,并使用胶原酶iv(sigma-aldrich)在37℃消化30-60分钟。通过使组织穿过100μm细胞过滤器获得单细胞悬液。将细胞在rpmi中洗涤两次,然后进行抗原刺激用于细胞内细胞因子染色(ics)和elisa。
[0290]
ics:共1-2
×
106个肺细胞在含有1μg/ml抗-cd28和抗-cd49d、加或不加5μg esat-6或mpt70 pepmix的rpmi 10%fcs中体外刺激1小时,然后在10μg/ml布雷非德菌素a(sigma-aldrich)存在下,在37℃的自动加热器中孵育5小时,孵育后将细胞冷却至4℃。第二天,细胞在facs缓冲液(含1%fcs的1
×
pbs)中洗涤,并在4℃使用抗-cd3、抗-cd4和抗-cd44对表面标志物进行染色。表面染色15-30分钟后,细胞在facs缓冲液中洗涤,使用cytofix/cytoperm试剂盒(bd)根据制造商的说明进行透化,然后用抗-ifn-γ、抗-tnf-α、抗-il-2、抗-il-17a进行细胞内染色。随后将细胞洗涤,重悬于facs缓冲液中并用lsr facs fortessa(bd)分析。所有流式细胞仪数据用flowjo软件v.10(treestar,ashland,or,usa)进行分析。
[0291]
elisa:用esat-6或mpt70 pepmix在37℃和5%co2下刺激肺细胞。三天后,通过培养上清液的夹心elisa测量分泌的ifn-γ。
[0292]
结果
[0293]
mtb感染后三周,通过ifn-γelisa(图4a)和分泌细胞因子的cd4 t细胞(图4b)二者测量,有明显的esat-6应答。然而,在该时间点,没有针对mpt70的免疫应答。在感染后12周,针对mpt70的免疫应答很小,但可检测到,在第20周时增强。这支持esat-6作为“早期抗原”和mpt70作为“晚期抗原”,其特征在于感染的晚期/慢性阶段的免疫识别。
[0294]
结论
[0295]
针对esat-6和mpt70的免疫应答动力学支持:在bcg-抗原中,esat-6是“早期抗原”;而mpt70是“晚期抗原”,因为其表达在晚期感染期间上调。
[0296]
实施例4:晚期抗原改善长期保护作用
[0297]
材料与方法
[0298]
各组小鼠用2μg融合蛋白免疫3次,该融合蛋白配制在总体积为200μl的阳离子佐剂制剂1(caf01)中。每轮疫苗接种相隔2周。第三次免疫后六周,所有动物用结核分枝杆菌erdman菌株进行气溶胶攻击(20-50cfu)。四或十八周后,通过铺板连续稀释的肺匀浆来测定各个肺中的分枝杆菌数量。
[0299]
结果
[0300]
为了检验“晚期抗原”对长期保护作用的重要性,将用含有早期和晚期抗原(h105)(seq id no:37)的融合蛋白免疫的感染动物中的细菌负荷,与用只含有早期表达抗原(h104)(seq id no:38)的融合蛋白免疫的感染动物进行比较。在感染的早期(感染4周),所有四种融合蛋白均诱导显著保护作用,保护水平无差异(图5a)。在感染的慢性阶段(18周),晚期抗原的添加产生了影响,因为h105比缺乏晚期抗原的h104提供了更好的保护作用(p=0.044)(图5b)。
[0301]
结论
[0302]
在疫苗中加入晚期抗原改善了长期保护/遏制(图5)。
[0303]
实施例5:bcg-抗原很高效,并且与bcg
抗原相比,能与bcg“协同”作用
[0304]
材料与方法
[0305]
各组cb6f1小鼠(n=4)用h107和bcg(bcg h107)共免疫,先在尾巴根部的左侧通过皮下注射(s.c.)施予bcg,然后在第二天在同一注射部位施予存在于caf01中的1μg h107(seq id no:9)。两周(右侧)和四周(左侧)后再次接种h107。最后一次免疫后两周,对动物实施安乐死,并通过使脾脏穿过100μm细胞过滤器获得单细胞悬液。如实施例3中所述,将细胞在rpmi中洗涤两次,然后进行抗原刺激用于ics,不同之处在于这次,表面染色中包括抗-klrg1。在第二个实验中,除小的生理盐水对照组(n=4)外,所有动物均接受牛分枝杆菌bcg danish免疫并休息12个月。在该时间点,将bcg免疫的动物分为四组(每组n=4),用牛分枝杆菌bcg再免疫一次或用h107/caf01或生理盐水再免疫3次。第三次免疫后三周,对动物实施安乐死,并从脾脏获得单细胞悬液,并使用h107作为抗原进行刺激用于ics(如实施例3中所述)。
[0306]
在分开的实验中,用bcg以及从h104-h107(bcg-抗原)、h74(bcg-抗原,seq id no:42)或h65(bcg
抗原,seq id no:43)选一种共免疫各组的小鼠(如上所述))。对照组用h107/caf01或bcg免疫或注射生理盐水。在mtb菌株erdman感染后4周和18周,测量各个肺中的细菌负荷。
[0307]
结果
[0308]
在生理盐水和bcg免疫的动物中,正如预期的那样,没有针对h107的免疫应答,因为h107不与bcg共享抗原。用h107免疫导致形成强烈的cd4t细胞应答,并且重要的是,通过bcg的共同施用(bcg h107,图6a)这种应答被显著增强。这表明bcg共同施用能够增强h107(bcg-抗原)的免疫应答,即使这两种疫苗之间不共享抗原。
[0309]
世界上大多数人口在新生儿时就进行了bcg免疫,由于最近的临床试验取得了可喜的结果,bcg再接种越来越受到关注。为了研究bcg的辅佐剂(co-adjuvant)作用是否也出现在已接种bcg的动物中,采用了bcg再接种模型,其中两次bcg免疫间隔一年(图6b)。正如预期的那样,在生理盐水、bcg或bcg再接种组中都没有针对h107的应答。相反,在最初接种过bcg的动物中,用h107疫苗接种导致明显的cd4 t细胞应答;当h107与二次bcg疫苗接种共同施用时,该应答显著增加。这表明在已接种过bcg疫苗的动物中bcg也可发挥辅佐剂的作用。
[0310]
用活的bcg进行疫苗接种诱导cd4 t细胞产生高度分化的表型,已知晓这种细胞具有较差的保护能力。相比之下,亚单位疫苗引起cd4 t细胞分化较少。与分化程度较低的t细胞(例如中央记忆细胞)相比,高度分化的cd4 t细胞(效应记忆和效应细胞)表达ifn-γ,但tnf-γ和il-2的共表达逐渐减弱(图7a)。为了表征t细胞分化,功能分化评分(fds)可定义为ifn-γ 细胞除以ifn-γ-细胞。bcg疫苗接种主要导致产生ifn-γ的效应子和效应记忆cd4 t细胞(以及非常少的ifn-γ细胞),产生的分化指数为13.3(图7b,上)。bcg和bcg
疫苗(h65)的共免疫使细胞因子表达谱有所改善,有更多的ifn-γ细胞,产生的fds为1.9(图7b,中)。然而,bcg与h107(bcg-抗原)的共接种使细胞因子表达谱有更大的改善,其中主要是ifn-γ中央型记忆cd4 t细胞,产生的fds是0.5。这说明当疫苗与bcg一起施用时,与bcg
抗原相比,使用bcg-抗原时可以获得改善的t细胞质量。高度分化的t细胞也以klrg1的表达为特征,并且证实了与bcg和bcg h65二者相比,bcg h107产生最低频率的表达klrg1的cd4 t细胞(图7c)。
[0311]
为了比较bcg与bcg-抗原(h107)或bcg
抗原(h65)共免疫的保护效果,用mtb erdman气溶胶感染免疫的小鼠。h74包含在内作为先前描述的融合蛋白的参考(wo2015161853a1)。在感染四周后,用h107和h74二者共免疫增加了牛分枝杆菌bcg疫苗的保护效力,而h65则没有。对于h107,这种增加很显著,不仅与h107或bcg单独相比(p《0.0001),还有与bcg与h74(p=0.0489)和h65(p《0.0001)的共免疫相比(图8a)。值得注意的是,h107作为独立疫苗施用比bcg和bcg h65更有效,与bcg h74相当。在晚期时间点(图8b),单独bcg和bcg h65共接种的保护功效都丧失了,但h107/caf01仍显著防御mtb(p《0.0001),并且用bcg和h107融合蛋白共接种仍然是最好的保护组。其他bcg-融合体h104-h106,当这些疫苗在第4周与bcg共同施用时,也诱导了相似水平的保护(图9)。
[0312]
结论
[0313]
将牛分枝杆菌bcg与由bcg-抗原组成的亚单位疫苗共同施用,在数量(图6)和质量(图7)两个方面均产生了增强的针对亚单位疫苗的免疫应答,并改善了气溶胶mtb感染后的保护作用(图8和图9)。h107作为独立疫苗比bcg和bcg h65更有效,与bcg h74相当,表明当与bcg共同施用以及作为独立疫苗时,本文公开的bcg-抗原组合是有强效的。
[0314]
实施例6:融合蛋白中的esat-6重复增加了免疫原性
[0315]
材料与方法
[0316]
在第一个实验中,各组的雌性cb6f1小鼠用caf01中的5μg h56(seq id no:45)或caf01中的5μg h56 5μg esat-6免疫3次。第三次免疫后三周,从每组两只动物中分离脾细胞,将2
×
105个细胞/孔用esat-6蛋白在37℃体外刺激6小时。通过ics测定cd4 t细胞的数量,该细胞产生细胞因子ifn-γ、tnf-α或il-2作为对刺激的应答。免疫后六周,用mtb菌株erdman感染小鼠,三周后从每组四只动物中分离肺细胞,并按照上述脾细胞那样处理,以测定产生细胞因子的cd4 t细胞的数量。感染的六周后,通过将各小鼠的肺匀浆铺在7h11琼脂上测定细菌数量(每组n=8)(图10)。
[0317]
在第二个实验中(图11),各组小鼠分别用含有一个(h64,seq id no:44)或五个(h76,seq id no:41)esat-6分子拷贝的h64/caf01或h76/caf01进行免疫。第三次免疫后三周,用h64和h76两种融合蛋白中存在的单抗原刺激脾细胞,并通过ics测定产生ifn-γ、tnf-α或il-2的cd4t细胞的频率。用rv0287蛋白进行刺激被包括在内作为阴性对照。在mtb菌株erdman感染六周后,测定肺中的细菌数量。
[0318]
在第三个实验中(图12),使用mtb复发模型研究esat-6重复在另一个分子骨架中的影响。在mtb erdman气溶胶感染六周后,在饮水中随意给予小鼠异烟肼(0.1g/l)和利福布汀(0.1g/l)六周。感染后第10周开始用0.5μg h83(一个esat-6拷贝,seq id no:40)和h84(四个esat-6拷贝,seq id no:39)或生理盐水(对照)免疫3次,每次间隔3周。在最后一次免疫(感染的第18周)后两周,取出肺,通过ics进行免疫分析,并且在感染后第22周和第35周,通过将匀浆铺在7h11琼脂上来评估肺的细菌负荷。
[0319]
结果
[0320]
esat-6是蛋白质疫苗接种后的低免疫原性抗原,因此需要增加esat-6特异性免疫原性。首先研究了添加更多游离esat-6与含有esat-6的融合蛋白(h56)相结合是否会增加免疫应答。接种后三周,与接受h56 esat-6的小鼠相比,单独接受h56的小鼠中的esat-6特异性免疫应答更高(图10a)。这种差异在气溶胶感染三周后在肺中更为明显。在该时间点,
产生细胞因子的cd4 t细胞的数量差异显著(p=0.0060)(图10b)。两种疫苗接种方案在感染六周后对肺部提供了类似的显著保护作用(相对于未接种疫苗的动物,p=0.0016和p=0.0121)。这表明向疫苗中添加更多的esat-6蛋白未增强免疫原性或保护作用。
[0321]
作为esat6为低免疫原性的解决方案,设计了一些在整个分子构建体中esat-6序列有重复的融合蛋白。比较了由h64(含有一个esat-6分子拷贝)和h76融合蛋白(含有五个esat-6分子拷贝)诱导的免疫应答,显示在h76免疫的动物中,疫苗引发的esat-6特异性cd4 t细胞的数量高三到五倍(图11a)。在气溶胶mtb erdman感染后六周,我们比较了h64和h76免疫动物中的细菌负荷,esat-6特异性免疫应答增强也导致保护作用增强(图11b)。
[0322]
同样,使用不同的分子构建体,将esat-6拷贝数从一个(h83)增加到四个(h84),增强了使用部分抗生素治疗的mtb复发模型中的免疫原性和保护作用。最后一次免疫后两周,与h83组相比,h84组中的esat-6特异性cd4 t细胞应答显著增强(图12a)。部分抗生素治疗结束后,对照组和接种h83动物的细菌负荷从第22周到第35周增加(复发)(图12b)。相反,h84疫苗接种动物的细菌负荷从第22周到第35周下降(图12b),表明esat-6重复模式既增加了esat-6特异性免疫应答,也增加了mtb复发模型中的保护能力。
[0323]
结论
[0324]
通过在疫苗配方中添加游离esat-6蛋白(“游离esat-6”指esat-6蛋白不是融合蛋白的一部分)以及含有esat-6的融合蛋白,不太可能增加疫苗引发的esat-6反应(图10)。相反,在融合蛋白中掺入更多的esat-6拷贝增加了疫苗引发的esat-6特异性cd4 t细胞的数量,并改善了不同动物模型中的保护作用(图11和12)。
[0325]
实施例7:融合蛋白既可以单独使用也可以与bcg一起使用
[0326]
材料与方法
[0327]
在第一个(图13a)和第二个实验(图13b)中,雌性129/sv和cb6f1小鼠分别用caf01中的1μg h74(seq id no:42)、h105(seq id no:37)或h107(seq id no:9)任一种免疫3次。第三次免疫后两周,从每组四只动物中分离脾细胞,1-2
×
106个细胞/孔用esat-6肽在37℃体外刺激6小时,然后通过ics测量产生细胞因子的cd4 t细胞的数量。如前所述,通过elisa测量3-天培养上清液中的ifn-γ分泌的水平。
[0328]
在第三个实验中(图13c),雌性cb6f1小鼠用bcg和在caf01中的1μg h105或h107(bcg h105或bcg h107)共免疫,通过在尾巴根部的左侧皮下注射(s.c.)施予bcg,然后第二天在同一注射部位施予caf01中的h105或h107。随后在两周(右侧)和四周(左侧)后,再次用亚单位疫苗免疫动物。第三次免疫后两周,从每组四只动物中分离脾细胞,1-2
×
106个细胞/孔在37℃用esat-6肽在体外刺激6小时,然后通过ics测量产生细胞因子的cd4 t细胞的数量。如本文所述,通过elisa测量3-天培养上清液中的ifn-γ分泌的水平。最后一次免疫后6周,所有动物均采用mtb erdman菌株气溶胶感染,感染4周后通过铺板器官匀浆测量肺的细菌负荷。
[0329]
结果
[0330]
在129/sv小鼠中,将esat-6拷贝数从一个(h74和h105)增加到四个(h107)导致esat-6应答增强(图13a)。这在cb6f1小鼠中也是如此,通过ics和elisa测量(图13b)。最后测试当这些疫苗与bcg共同施用时是否也是如此。在该方案中,再次,与h105相比,h107显著增加了esat-6特异性免疫应答,这也导致针对mtb气溶胶感染的保护作用增加(图13c)。
[0331]
结论
[0332]
在三个独立的实验中(使用两种不同的小鼠品系),证明了将esat-6拷贝数从一个(h74和h105)增加到四个(h107)增加了esat-6特异性免疫应答。当这些疫苗与bcg共同施用时也是如此。这还减少了细菌负荷,显示在疫苗骨架中重复esat-6是一种强有力的方法来改善免疫原性和疫苗介导的保护作用。此外,数据支持了:融合物不管作为独立的疫苗还是与bcg一起使用都是有效的。
[0333]
实施例8:h107是比现有技术亚单位疫苗h56更好的独立疫苗
[0334]
材料与方法
[0335]
各组小鼠用2μg融合蛋白免疫三次,该融合蛋白是配制在总体积200μl的阳离子佐剂制剂1(caf01)中。每轮疫苗接种相隔2周。第三次免疫后六周,所有动物用结核分枝杆菌菌株erdman进行气溶胶攻击(20-50cfu)。四至十二周后,通过铺板连续稀释的肺匀浆,测定各个肺中的分枝杆菌数量。
[0336]
结果
[0337]
为了比较h107(seq id no:9)相对目前最先进的亚单位疫苗(h56;seq id no:45)的保护效力,在两个(h107)和五个(h56)独立实验中,将各组小鼠用caf01佐剂中的融合蛋白免疫。将动物用mtb菌株erdman进行攻击,并计算各个动物肺中的细菌数量(图14)。根据实验,在mtb菌株erdman攻击后四至十二周测定保护效力。平均而言,h107疫苗提供2.0log10保护,对应细菌负荷减少到1/100。相比之下,h56疫苗使细菌负荷平均减少到约1/7(0.82log10)。
[0338]
结论
[0339]
与最先进的亚单位tb疫苗(h56/caf01)相比,h107/caf01疫苗针对毒性mtb的气溶胶攻击提供了更好的保护作用(图14)。
[0340]
实施例9:bcg h107共同施用增强bcg-特异性免疫应答
[0341]
材料与方法
[0342]
如先前实施例所述,各组空白(naive)cb6f1小鼠用bcg和三次1μg h107(seq id no:9)共免疫(每个时间点n=4)。在免疫后的不同时间点对动物实施安乐死,并通过使脾脏穿过100μm细胞过滤器获得单细胞悬液。如先前实施例中所述,将细胞在rpmi中洗涤两次,然后抗原刺激进行ics。将tb10.4用于刺激,以评估bcg-特异性免疫应答,因为该抗原包含在bcg中,但不包含在h107中。
[0343]
结果
[0344]
在第一次免疫后三周(第二次h107后一周),与单独bcg相比,在bcg h107组中观察到增加的tb10.4应答(图15)。这种差异一直保持到第9周实验终止。这表明h107与bcg的共同施用增加了bcg特异性免疫应答,即使它们不共享抗原。
[0345]
结论
[0346]
先前的实施例表明,bcg h107共同施用导致h107应答增加(bcg发挥h107佐剂的作用)。该实施例表明,h107 bcg的共同施用也增加了bcg-特异性免疫应答,这意味着h107发挥bcg佐剂的作用(图15)。这样,两种疫苗之间就存在真正的协同作用。
[0347]
实施例10:与h107相比,h107e增加了大肠杆菌中的蛋白质表达
[0348]
除其他外,重组蛋白表达的产量取决于蛋白质的氨基酸序列,大规模的疫苗生产
需要最优化的表达过程。为了解决这个问题,开发了h107的高表达版本:h107e。
[0349]
材料与方法
[0350]
为了优化表达,h107(seq id no:9)的rv3876部分中富含脯氨酸的序列aa298-aa517被删除得到h107e(seq id no:91)。通过化学合成制得h107对应的dna序列(seq id no:54)和h107e对应的dna序列(seq id no:92),插入到pj411表达载体(atum,menlo park,ca,us),并转化到在luria-bertani(lb)培养基中生长的大肠杆菌bl21(de3)菌株(agilent,dk)中。在用1mm异丙基β-d-1-硫代吡喃半乳糖苷(iptg)诱导培养后0、1和3小时,通过sds-page或蛋白质印迹(使用识别h107/h107e的mpt70部分的一抗)评估重组蛋白表达水平。在凝胶上的泳道中上样相同数量的细胞(相同的od600)。表达后,从包涵体中纯化重组抗原,并使用5ml histrap hp柱(ge healthcare)进行金属离子亲和层析,然后使用hitrap q hp柱(ge healthcare)进行阴离子交换。通过二辛可宁酸测定法估算最终的蛋白产量。
[0351]
结果
[0352]
从培养诱导实验中明显看出,h107e在大肠杆菌中的表达显著增加。在诱导后1和3小时后,在sds-page凝胶和蛋白质印迹中,对应于h107e的条带均比h107的条带强得多(图16a)。作为比较,从6l发酵物得到的纯化蛋白质的产量:h107为2.4-7.2mg,h107e为15.2mg。
[0353]
结论:
[0354]
与h107相比,h107e序列的改变导致蛋白质表达和产量都显著增加。
[0355]
实施例11:h107e与h107一样有免疫原性,并且还与bcg协同作用
[0356]
材料和方法
[0357]
如前面实施例所述,将各组空白cb6f1小鼠(n=8)用bcg和1μg h107(seq id no:9)或h107e(seq id no:91)共同免疫。最后一次免疫后两周,对动物实施安乐死,通过使脾脏穿过100μm细胞过滤器获得单细胞悬液。如前面实施例所述,将细胞在rpmi中洗涤两次,然后抗原刺激以便进行ics和ifn-γelisa。
[0358]
在第二个实验中,将各组空白cb6f1小鼠(n=6)按上述用bcg和h107e共免疫,对照组用h107e/caf01或bcg免疫或者注射生理盐水。在mtb菌株erdman感染8周后,测量各个肺中的细菌负荷。
[0359]
在第三个实验中,各组bcg记忆cb6f1小鼠(n=6)在尾根处皮下注射2μg h107e或h65免疫3次。最后一次免疫后两周,对动物实施安乐死,通过使腹股沟淋巴结穿过100μm细胞过滤器获得单细胞悬液。将细胞在rpmi中洗涤两次,然后抗原刺激以便进行ics。
[0360]
结果
[0361]
与bcg共接种后,证实h107e和h107具有相同的免疫原性,免疫原性是通过表达细胞因子的cd4 t细胞(图17a,左)和经由elisa测量的ifn-γ释放(图17a,右)来测量。还证实了h107e同h107一样,对相同的各个抗原诱导免疫应答(图17b)。值得注意的是,h107e中rv3876部分的缺失导致rv3876特异性免疫应答略有下降,但针对mpt70和mpt83的免疫应答增加(图17b)。感染后,h107e提供与bcg相似或更好的保护作用,与单独的bcg和h107e相比,bcg h107e共接种导致保护作用显著增加(图17c),正如先前在实施例9中用h107观察到的那样。在bcg记忆小鼠中,与h65(bcg )相比,h107e(bcg-)疫苗接种导致的cd4 t细胞分化更少(质量更好),这是通过功能分化评分fds来测量(图17d)。此外,随着th17细胞在过去十年
中获得了越来越多的关注,th17细胞被认为在mtb感染期间具有保护特性,通过流式细胞术测量表达il-17的cd4 t细胞。与h65相比,h107e诱导的th17细胞比例要高得多,这表明与传统的bcg 疫苗相比,bcg-疫苗在bcg初免(bcg-primed)的个体中产生整体更广泛的免疫应答(图17d)。
[0362]
结论
[0363]
h107e具有与h107相似的免疫原性,并在mtb攻击后诱导显著的保护作用。与h107一样,h107e与bcg协同作用,并且bcg h107e共接种的保护水平显著高于单独的h107e和bcg。在bcg初免的动物中,与h65(bcg )相比,h107e(bcg-)疫苗接种诱导较少的t细胞分化(如同h107)并增加th17细胞的比例,该比例通过表达il-17的cd4 t细胞来测量。
[0364]
参考文献
[0365]
·
wo2010006607a2
[0366]
·
wo2006136162a2
[0367]
·
wo2014063704a2
[0368]
·
wo2015161853a1
[0369]
·
sederetal(2008),nat.rev.immunol.8,247-258
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。