非靶向和靶向性il
‑
10 fc融合蛋白
背景技术:
1.本技术要求2019年2月21日提交的美国临时申请号62/808,749和2019年2月21日提交的美国临时申请号62/808,751的权益,其内容通过引用整体并入本文。
背景技术
[0002][0003]
为了让免疫系统产生有效的抗肿瘤应答,必须发生两件事。肿瘤环境中的t细胞必须首先接合由肿瘤细胞上的主要组织相容性复合物(mhc)呈递的抗原性肿瘤肽。接下来,t细胞必须由细胞因子诸如il
‑
15和il
‑
2诱导产生共刺激细胞因子诸如ifnγ。在不存在细胞因子诱导的情况下,识别仅肿瘤肽会导致t细胞变得无反应性,由此导致具有耐受性。因此,在癌症免疫治疗中非常有前景的方法是基于细胞因子的治疗。实际上,il
‑
2已被批准用于患有转移性肾细胞癌和恶性黑色素瘤的患者。
[0004]
除了t细胞无反应性之外,肿瘤细胞还通过mhc分子的下调来逃避免疫应答(seliger等人,2002)。虽然白介素10(il
‑
10)是传统上认可其免疫抑制特性的细胞因子(taga,k.和tosato, g.,1992),但已发现它在肿瘤环境中具有免疫刺激功能。与此一致,mumm等人(2011)报道, il
‑
10治疗显著增加肿瘤微环境中cd8
t细胞的数量、所述肿瘤内cd8
t细胞的ifnγ和颗粒酶表达,以及肿瘤内抗原呈递分子诸如mhc
‑
i和mhc
‑
ii的表达,所有这些都是效应子抗肿瘤应答的基本机制。
[0005]
生物功能il
‑
10是通过两个il
‑
10单体之间的非共价相互作用形成的结构域交换的同源二聚体。生物功能il
‑
10二聚体结合il
‑
10受体,所述受体由两个α和两个β亚基(或r1和 r2亚基)组成。值得注意的是,il
‑
10二聚体在各个il
‑
10单体之间的相互作用被破坏时变得没有生物活性。此外,与其他细胞因子一样,il
‑
10具有短的血浆半衰期。rachmawati等人 (2004)报道,注射到大鼠体内的人il
‑
10经历快速初始清除,得到1.7分钟的半衰期。因此,在第一方面,本发明通过提供il
‑
10融合蛋白以及被工程化为具有降低效力的新型il
‑
10变体来解决生物功能il
‑
10二聚体的潜在破坏及其短循环半衰期问题。
[0006]
虽然il
‑
10可以增强免疫应答,例如在癌症中,但是通过刺激肿瘤内cd8
t细胞,il
‑
10 也可增强通过cd4
调节性t细胞对免疫应答的抑制。与此一致,chan等人(2015)报道称il
‑ꢀ
10刺激cd8
t细胞的ifnγ分泌,将il
‑
10与大量pbmc一起孵育实际上导致ifnγ分泌的抑制,表明il
‑
10的免疫抑制作用通过非cd8
淋巴细胞增强。此外,til中的cd8/cd4 t细胞比率高通常被认为是肿瘤疗法的良好预后指标。因此,在另一方面,本发明提供了靶向cd8 的il
‑
10融合蛋白。
[0007]
t细胞活化后,免疫检查点蛋白诸如pd
‑
1被上调,以通过在与免疫检查点配体诸如pd
‑ꢀ
l1结合后耗尽活化的t细胞来排除自身免疫。然而,免疫检查点蛋白在肿瘤浸润的淋巴细胞 (til)中也上调,并且免疫检查点配体在肿瘤细胞上过表达,作为肿瘤细胞的另一免疫逃逸机制。已经证明,通过诸如(纳武单抗)和(派姆单抗)等药物阻断免疫检查点相互作用来消除对til的抑制作用对癌症治疗高度有效。尽管诸如纳武单抗和
派姆单抗的检查点阻断疗法有前景,但许多患者仍然无法实现对单独的检查点阻断的足够的反应。因此,需要鉴定与检查点阻断相叠加的治疗方式,其能够提高患者的反应率。这可能特别复杂,因为另外的治疗方式不应与检查点阻断竞争。因此,在另一方面,本发明提供了靶向pd
‑
1的il
‑ꢀ
10融合蛋白,其对表达pd
‑
1的til具有选择性,并且不与可与其组合的检查点阻断抗体竞争。
技术实现要素:
[0008]
在一个方面,本发明提供一种异源二聚体fc融合蛋白,其包含:(a)融合蛋白,其包含第一蛋白结构域、第二蛋白结构域和第一fc结构域,其中第一蛋白结构域与第二蛋白结构域共价连接,并且其中第二蛋白结构域与第一fc结构域共价连接;以及(b)第二fc结构域;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一蛋白结构域包含第一il
‑
10单体结构域并且第二蛋白结构域包含第二il
‑
10单体结构域。在一些情况下,异源二聚体fc融合蛋白具有单链il
‑
10异源fc融合体或scil10
‑
异源fc形式。
[0009]
在一些实施方案中,第二蛋白结构域与第一fc结构域的n端共价连接。在某些实施方案中,第二蛋白结构域与第一fc结构域的c端共价连接。在一些实施方案中,第二蛋白结构域通过第一结构域接头与第一fc结构域共价连接。在一些实施方案中,第一蛋白结构域通过第二结构域接头与第二蛋白结构域连接。
[0010]
在一些实施方案中,促进第一和第二fc结构域的异源二聚化的修饰为选自由以下组成的组的一组氨基酸取代:根据eu编号的l368d/k370s和s364k;l368d/k370s和s364k/e357l; l368d/k370s和s364k/e357q;t411e/k360e/q362e和d401k;l368e/k370s和s364k; k370s和s364k/e357q以及t366s/l368a/y407v:t366w(任选地包括二硫桥键, t366s/l368a/y407v/y349c:t366w/s354c)。在一些实施方案中,第一和/或第二fc结构域具有另外一组包括根据eu编号的q295e/n384d/q418e/n421d的氨基酸取代。在一些实施方案中,第一和/或第二fc结构域具有由以下组成的另外一组氨基酸取代:根据eu编号的 g236r/l328r、e233p/l234v/l235a/g236_/s239k、 e233p/l234v/l235a/g236_/s239k/a327g、e233p/l234v/l235a/g236_/s267k/a327g、 e233p/l234v/l235a/g236_以及e233p/l234v/l235a/g236_/s267k。
[0011]
在一些实施方案中,异源二聚体fc融合蛋白在第一和/或第二fc结构域中包含氨基酸修饰g446del/k4447del。
[0012]
在一些实施方案中,第二fc结构域和/或第二fc结构域包含选自由根据eu编号的 m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。
[0013]
在一些实施方案中,第一和/或第二il
‑
10单体结构域具有选自由以下组成的组的多肽序列:seq id no:1(人il
‑
10(109h)前体序列)、seq id no:2(人il
‑
10(109l)前体序列)、seqid no:3(人il
‑
10(109h)成熟形式序列)、seq id no:4(人il
‑
10(109l)成熟形式序列),如图1所描绘。
[0014]
在一些实施方案中,第一和/或第二il
‑
10单体结构域是变体il
‑
10单体结构域。在一些实施方案中,变体il
‑
10单体结构域包括具有一个或多个氨基酸取代的il
‑
10单体结构域,所述氨基酸取代导致对il
‑
10受体的亲和力改变、效力改变、脱酰胺作用的潜力改变、天
冬氨酸异构化的潜力改变、降解相关ptm的潜力改变、潜在降解位点改变、二硫桥改变和/或潜在 n
‑
糖基化位点改变。
[0015]
在一些实施方案中,变体il
‑
10单体结构域在选自由n21、d28、q38、m39、d41、q42、 l43、d44、n45、i87、e142、d144、e151和n160组成的组的氨基酸残基处包含一个或多个氨基酸修饰。在一些实施方案中,变体il
‑
10单体结构域包含选自n21d、d28n、q38e、 m39t、d41n、q42e、l43v、d44n、n45d、i87a、e142q、d144n、e151q和n160del的一个或多个氨基酸修饰。
[0016]
在一些实施方案中,与第二il
‑
10单体结构域共价连接的第一il
‑
10单体结构域形成单链 il
‑
10,其包含seq id no:23或具有氨基酸修饰109l和269l的seq id no:23。在一些实施方案中,单链il
‑
10包含选自由n21、q38、d41、n45、d144、e151、n181、q198、n205、 d304、e311和n320组成的组的一个或多个氨基酸修饰。
[0017]
在一些实施方案中,单链il
‑
10包含选自由n21d、q38e、d41n、n45d、d144n、e151q、 n181d、q198e、n205d、d304n、e311q和n320del组成的组的一个或多个氨基酸修饰。在一些实施方案中,单链il
‑
10包含选自n21d/n181d/n320del、n45d/n205d/n320del、 q38e/n45d/n205d/n320del、d41n/n45d/n205d/n320del、n45d/d144n/n205d/n320del、n45d/e151q/n205d/n320del、n45d/n181d/n205d/n320del、n45d/n205d/d304n/n320del、 n21d/n45d/n181d/n205d/n320del、q38e/n45d/q198e/n205d/n320del、 q42e/n45d/q202e/n205d/n320del和n45d/e151q/n205d/e311q/n320del的氨基酸修饰。
[0018]
在一些实施方案中,fc融合蛋白(例如,scil10
‑
异源fc(scil10
‑
heterofc)融合蛋白)是 xenp30005(seq id no:25和26)、xenp30008(seq id no:27和28)或xenp30013(seqid no:29和30),如图78a
‑
78g中所描绘。
[0019]
在一些实施方案中,fc融合蛋白是选自xenp25239、xenp25240和xenp25241的成员,如图21a
‑
21c中所描绘。
[0020]
本文提供了一种包含本文概述的异源二聚体fc融合蛋白(例如,scil10
‑
异源fc融合蛋白)的组合物,其用于治疗受试者的癌症。此外,本文提供编码本文概述的异源二聚体fc融合蛋白的一种或多种核酸。此外,本文提供一种宿主细胞,其包含编码本文概述的异源二聚体fc融合蛋白的一种或多种核酸。
[0021]
在另一方面,本文提供一种制备异源二聚体fc融合蛋白(例如,scil10
‑
异源fc融合蛋白) 的方法,其包括在借由产生异源二聚体fc融合蛋白的条件下培养所描述的任何宿主细胞;以及回收异源二聚体fc融合蛋白。
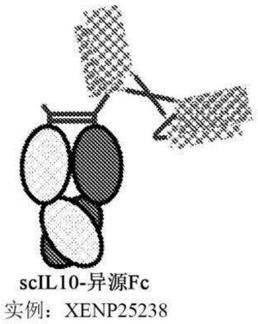
[0022]
在另一方面,本文提供一种纯化本文所述的异源二聚体fc融合蛋白的方法。所述方法包括:(a)提供包含异源二聚体fc融合蛋白的组合物;(b)将组合物上样到离子交换柱上;以及 (c)收集含有异源二聚体fc融合蛋白的级分。
[0023]
在另一方面,本发明提供一种异源二聚体fc融合蛋白,其包含:(a)第一融合蛋白,其包含抗原结合结构域(abd)和第一fc结构域,其中抗原结合结构域与第一fc结构域的n端连接;以及(b)第二融合蛋白,其包含第一蛋白结构域、第二蛋白结构域和第二fc结构域,其中第一蛋白结构域与第二蛋白结构域共价连接,并且其中第二蛋白结构域与第二fc结构域共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一蛋白结构域包含第一il
‑
10单体结构域并且第二蛋白结构域包含第二il
‑
10单体结构域。在一些情况下,异源二聚体fc融合蛋白具有抗x x单链il
‑
10异源fc融合体或
抗x xscil10
‑
异源fc形式。在一些实施方案中,scil10的il
‑
10单体与fc结构域的n端或c端连接。
[0024]
在一些实施方案中,第二蛋白结构域(例如,第二il
‑
10单体结构域)与第二fc结构域的 n端共价连接。在一些实施方案中,第二蛋白结构域与第二fc结构域的c端共价连接。在一些实施方案中,使用第一结构域接头将第一蛋白结构域(例如,第一il
‑
10单体结构域)与第二蛋白结构域(例如,第二il
‑
10单体)连接。在一些实施方案中,使用第二结构域接头将抗原结合结构域与第一fc结构域连接。
[0025]
在一些实施方案中,促进第一和第二fc结构域的异源二聚化的修饰是选自由以下组成的组的一组氨基酸取代:根据eu编号的l368d/k370s和s364k;l368d/k370s和s364k/e357l; l368d/k370s和s364k/e357q;t411e/k360e/q362e和d401k;l368e/k370s和s364k; k370s和s364k/e357q以及t366s/l368a/y407v:t366w(任选地包括二硫桥键, t366s/l368a/y407v/y349c:t366w/s354c)。
[0026]
在一些实施方案中,第一和/或第二fc结构域具有另外一组包括根据eu编号的 q295e/n384d/q418e/n421d的氨基酸取代。
[0027]
在一些实施方案中,第一和/或第二fc结构域具有由以下组成的另外一组氨基酸取代:根据eu编号的g236r/l328r、e233p/l234v/l235a/g236_/s239k、 e233p/l234v/l235a/g236_/s239k/a327g、e233p/l234v/l235a/g236_/s267k/a327g、 e233p/l234v/l235a/g236_以及e233p/l234v/l235a/g236_/s267k。
[0028]
在一些实施方案中,第一fc结构域和/或第二fc结构域包含选自由根据eu编号的 m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。
[0029]
在一些实施方案中,第一和/或第二il
‑
10单体结构域包含选自由以下组成的组的多肽序列:seq id no:1(人il
‑
10(109h)前体序列)、seq id no:2(人il
‑
10(109l)前体序列)、seqid no:3(人il
‑
10(109h)成熟形式序列)、seq id no:4(人il
‑
10(109l)成熟形式序列),如图1中所描绘。
[0030]
在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域是变体il
‑
10单体结构域。
[0031]
在一些实施方案中,变体il
‑
10单体结构域包括具有一个或多个氨基酸取代的il
‑
10单体结构域,所述氨基酸取代导致对il
‑
10受体的亲和力改变、效力改变、脱酰胺作用的潜力改变、天冬氨酸异构化的潜力改变、降解相关ptm的潜力改变、潜在降解位点改变、二硫桥改变和/或潜在n
‑
糖基化位点改变。
[0032]
在一些实施方案中,变体il
‑
10单体结构域在选自由n21、d28、q38、m39、d41、q42、 l43、d44、n45、i87、e142、d144、e151和n160组成的组的氨基酸残基处包含一个或多个氨基酸修饰。在一些实施方案中,变体il
‑
10单体结构域包含选自n21d、d28n、q38e、 m39t、d41n、q42e、l43v、d44n、n45d、i87a、e142q、d144n、e151q和n160del的一个或多个氨基酸修饰。
[0033]
在一些实施方案中,与第二il
‑
10单体结构域共价连接的第一il
‑
10单体结构域形成单链 il
‑
10,其包含seq id no:23或具有氨基酸修饰109l和269l的seq id no:23。在一些实施方案中,单链il
‑
10还包含选自由n21、q38、d41、n45、d144、e151、n181、q198、 n205、d304、e311和n320组成的组的一个或多个氨基酸修饰。在一些实施方案中,单链il
‑ꢀ
10还包含选自由n21d、q38e、d41n、n45d、d144n、e151q、n181d、q198e、n205d、 d304n、e311q和
mab c[pd
‑
1]_h1.48_l1.45、mab c[pd
‑
1]_h1.48_l1.117、mab c[pd
‑
1]_h1.48_l1.129、mabc[pd
‑
1]_h1.48_l1.135、mab c[pd
‑
1]_h1.48_l1.136、mab c[pd
‑
1]_h1.48_l1.140、mab c[pd
‑ꢀ
1]_h1.48_l2、mab c[pd
‑
1]_h1.125_l1、mab c[pd
‑
1]_h1.125_l1.1、mab c[pd
‑ꢀ
1]_h1.125_l1.3、mab c[pd
‑
1]_h1.125_l1.45、mab c[pd
‑
1]_h1.125_l1.117、mab c[pd
‑ꢀ
1]_h1.125_l1.129、mab c[pd
‑
1]_h1.125_l1.135、mab c[pd
‑
1]_h1.125_l1.136、mab c[pd
‑ꢀ
1]_h1.125_l1.140、mab c[pd
‑
1]_h1.125_l2、mab c[pd
‑
1]_h1.130_l1、mab c[pd
‑ꢀ
1]_h1.130_l1.1、mab c[pd
‑
1]_h1.130_l1.3、mab c[pd
‑
1]_h1.130_l1.45、mab c[pd
‑ꢀ
1]_h1.130_l1.117、mab c[pd
‑
1]_h1.130_l1.129、mab c[pd
‑
1]_h1.130_l1.135、mab c[pd
‑ꢀ
1]_h1.130_l1.136、mab c[pd
‑
1]_h1.130_l1.140、mab c[pd
‑
1]_h1.130_l2、mab c[pd
‑ꢀ
1]_h1.132_l1、mab c[pd
‑
1]_h1.132_l1.1、mab c[pd
‑
1]_h1.132_l1.3、mab c[pd
‑ꢀ
1]_h1.132_l1.45、mab c[pd
‑
1]_h1.132_l1.117、mab c[pd
‑
1]_h1.132_l1.129、mab c[pd
‑ꢀ
1]_h1.132_l1.135、mab c[pd
‑
1]_h1.132_l1.136、mab c[pd
‑
1]_h1.132_l1.140、mab c[pd
‑ꢀ
1]_h1.132_l2、mab c[pd
‑
1]_h1.169_l1、mab c[pd
‑
1]_h1.169_l1.1、mab c[pd
‑ꢀ
1]_h1.169_l1.3、mab c[pd
‑
1]_h1.169_l1.45、mab c[pd
‑
1]_h1.169_l1.117、mab c[pd
‑ꢀ
1]_h1.169_l1.129、mab c[pd
‑
1]_h1.169_l1.135、mab c[pd
‑
1]_h1.169_l1.136、mab c[pd
‑ꢀ
1]_h1.169_l1.140、mab c[pd
‑
1]_h1.169_l2、mab c[pd
‑
1]_h1.175_l1、mab c[pd
‑ꢀ
1]_h1.175_l1.1、mab c[pd
‑
1]_h1.175_l1.3、mab c[pd
‑
1]_h1.175_l1.45、mab c[pd
‑ꢀ
1]_h1.175_l1.117、mab c[pd
‑
1]_h1.175_l1.129、mab c[pd
‑
1]_h1.175_l1.135、mab c[pd
‑ꢀ
1]_h1.175_l1.136、mab c[pd
‑
1]_h1.175_l1.140、mab c[pd
‑
1]_h1.175_l2、mab c[pd
‑ꢀ
1]_h2_l1、mab c[pd
‑
1]_h2_l1.1、mab c[pd
‑
1]_h2_l1.3、mab c[pd
‑
1]_h2_l1.45、mabc[pd
‑
1]_h2_l1.117、mab c[pd
‑
1]_h2_l1.129、mab c[pd
‑
1]_h2_l1.135、mab c[pd
‑ꢀ
1]_h2_l1.136、mab c[pd
‑
1]_h2_l1.140和mab c[pd
‑
1]_h2_l2,如图104a
‑
104b中所描绘。
[0038]
在一些实施方案中,cd8结合结构域是人源化的cd8 abd。在一些实施方案中,人源化的cd8 abd包含选自由如图92中所描绘的okt8_h2l1和1c11b3_h1l1组成的组的可变重链和可变轻链对。
[0039]
在一些实施方案中,nkg2d结合结构域是人源化的nkg2d abd。在一些实施方案中,人源化的nkg2d abd包含选自由以下组成的组的可变重链和可变轻链对:ms[nkg2d] h0_l0、1d7b4[nkg2d]_h1_l1、kyk
‑
1.0[nkg2d]_h1_l1、kyk
‑
2.0[nkg2d]_h0_l0、 11b2d10[nkg2d]_h0_l0、6e5a7[nkg2d]_h0_l0、6h7e7[nkg2d]_h0_l0、mabe[nkg2d]_h1_l1、16f31[nkg2d]_h1_l1、mab d[nkg2d]_h1_l1、 1d7b4[nkg2d]_h1_l1、mab a[nkg2d]_h1_l1、mab a[nkg2d]_h2_l1、maba[nkg2d]_h1_l2、mab a[nkg2d]_h2_l2、mab b[nkg2d]_h1_l1、mabb[nkg2d]_h1_l1.1、mab b[nkg2d]_h1_l2、mab b[nkg2d]_h2_l1、mabb[nkg2d]_h2_l1.1、mab b[nkg2d]_h2_l2、mab b[nkg2d]_h3_l1、mabb[nkg2d]_h3_l1.1、mab b[nkg2d]_h3_l2、mab c[nkg2d]_h1_l1、mabc[nkg2d]_h2_l1、mab c[nkg2d]_h1_l2和mab c[nkg2d]_h2_l2,如图93a
‑
93c中所描绘。
[0040]
本文提供一种包含所描述的异源二聚体fc融合蛋白(例如,抗x x scil10
‑
异源fc融合蛋白)的组合物,其用于治疗受试者的癌症。此外,提供了一种或多种编码本文所述的任何异源二聚体fc融合蛋白的核酸。此外,提供了一种宿主细胞,其包含一种或多种编码本文所述的任何异源二聚体fc融合蛋白的核酸。
[0041]
在一个方面,提供了一种制备异源二聚体fc融合蛋白(例如,抗x x scil10
‑
异源fc融合蛋白)的方法,其包括在借由产生异源二聚体fc融合蛋白的条件下培养本文所述的宿主细胞;以及回收蛋白质。
[0042]
在另一方面,提供了一种纯化本文所述的异源二聚体fc融合蛋白(例如,抗x x scil10
‑ꢀ
异源fc融合蛋白)的方法。所述方法包括:(a)提供包含异源二聚体fc融合蛋白的组合物;(b) 将组合物上样到离子交换柱上;以及(c)收集含有异源二聚体fc融合蛋白的级分。
[0043]
在另一方面,本发明提供一种异源二聚体fc融合蛋白,其包含:(a)第一融合蛋白,其包含第一抗原结合结构域和第一fc结构域,其中第一抗原结合结构域(abd)与第一fc结构域的n端共价连接;以及(b)第二融合蛋白,其包含第二抗原结合结构域(abd)、第二fc结构域、第一蛋白结构域和第二蛋白结构域,其中第二抗原结合结构域与第二fc结构域的n端共价连接,其中第一蛋白结构域与第二蛋白结构域共价连接,并且其中第二蛋白结构域与第二fc结构域的c端共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一蛋白结构域包含第一il
‑
10单体结构域并且第二蛋白结构域包含第二il
‑
10单体结构域。在一些情况下,异源二聚体fc融合蛋白具有(抗x)2x异源 fc
‑
单链il
‑
10融合体或(抗x)2
‑
异源fc
‑
scil10融合体形式。
[0044]
在一些实施方案中,促进第一和第二fc结构域的异源二聚化的修饰是选自由以下组成的组的一组氨基酸取代:根据eu编号的l368d/k370s和s364k;l368d/k370s和s364k/e357l; l368d/k370s和s364k/e357q;t411e/k360e/q362e和d401k;l368e/k370s和s364k; k370s和s364k/e357q以及t366s/l368a/y407v:t366w(任选地包括二硫桥键, t366s/l368a/y407v/y349c:t366w/s354c)。
[0045]
在一些实施方案中,第一和/或第二fc结构域具有另外一组包括根据eu编号的 q295e/n384d/q418e/n421d的氨基酸取代。
[0046]
在一些实施方案中,第一和/或第二fc结构域具有由以下组成的另外一组氨基酸取代:根据eu编号的g236r/l328r、e233p/l234v/l235a/g236_/s239k、 e233p/l234v/l235a/g236_/s239k/a327g、e233p/l234v/l235a/g236_/s267k/a327g、 e233p/l234v/l235a/g236_以及e233p/l234v/l235a/g236_/s267k。
[0047]
在一些实施方案中,第一fc结构域和/或第二fc结构域包含选自由根据eu编号的 m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。
[0048]
在一些实施方案中,第一和/或第二il
‑
10单体结构域包含选自由以下组成的组的多肽序列:seq id no:1(人il
‑
10(109h)前体序列)、seq id no:2(人il
‑
10(109l)前体序列)、seqid no:3(人il
‑
10(109h)成熟形式序列)、seq id no:4(人il
‑
10(109l)成熟形式序列),如图1中所描绘。
[0049]
在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域是变体il
‑
10单体结构域。
[0050]
在一些实施方案中,变体il
‑
10单体结构域包括具有一个或多个氨基酸取代的il
‑
10单体结构域,所述氨基酸取代导致对il
‑
10受体的亲和力改变、效力改变、脱酰胺作用的潜力改变、天冬氨酸异构化的潜力改变、降解相关ptm的潜力改变、潜在降解位点改变、二硫桥改变和/或潜在n
‑
糖基化位点改变。
[0051]
在一些实施方案中,变体il
‑
10单体结构域在选自由n21、d28、q38、m39、d41、q42、 l43、d44、n45、i87、e142、d144、e151和n160组成的组的氨基酸残基处包含一个或多个氨基酸修饰。在一些实施方案中,变体il
‑
10单体结构域包含选自n21d、d28n、q38e、 m39t、d41n、q42e、l43v、d44n、n45d、i87a、e142q、d144n、e151q和n160del的一个或多个氨基酸修饰。
[0052]
在一些实施方案中,与第二il
‑
10单体结构域共价连接的第一il
‑
10单体结构域形成单链 il
‑
10,其包含seq id no:23或具有氨基酸修饰109l和269l的seq id no:23。
[0053]
在一些实施方案中,单链il
‑
10还包含选自由n21、q38、d41、n45、d144、e151、n181、 q198、n205、d304、e311和n320组成的组的一个或多个氨基酸修饰。在一些实施方案中,单链il
‑
10还包含选自由n21d、q38e、d41n、n45d、d144n、e151q、n181d、q198e、 n205d、d304n、e311q和n320del组成的组的一个或多个氨基酸修饰。在一些实施方案中,单链il
‑
10包含选自n21d/n181d/n320del、n45d/n205d/n320del、 q38e/n45d/n205d/n320del、d41n/n45d/n205d/n320del、n45d/d144n/n205d/n320del、 n45d/e151q/n205d/n320del、n45d/n181d/n205d/n320del、n45d/n205d/d304n/n320del、 n21d/n45d/n181d/n205d/n320del、q38e/n45d/q198e/n205d/n320del、 q42e/n45d/q202e/n205d/n320del和n45d/e151q/n205d/e311q/n320del的氨基酸修饰。
[0054]
在一些实施方案中,第一abd和/或第二abd选自由pd
‑
1结合结构域、非竞争性pd
‑ꢀ
1结合结构域、tigit结合结构域、cd8结合结构域和nkg2d结合结构域组成的组。
[0055]
在一些实施方案中,pd
‑
1结合结构域是人源化的pd
‑
1abd。在一些实施方案中,人源化的pd
‑
1abd包含选自由以下组成的组的可变重链和可变轻链对:1c11[pd
‑
1]_h0l0、 1c11[p,d
‑
1]_h3l3、1c11[pd
‑
1]_h3.240_l3.148、1c11[pd
‑
1]_h3.241_l3.148、1c11[pd
‑ꢀ
1]_h3.234_l3.144、1c11[pd
‑
1]_h3.241_l3.92、1c11[pd
‑
1]_h3.303_l3.152、 1c11_h3.329_l3.220、1c11_h3.328_l3.152、派姆单抗可变重链和可变轻链、纳武单抗可变重链和可变轻链、皮地利珠单抗可变重链和可变轻链、mk
‑
3475可变重链和可变轻链、bap049 克隆e可变重链和可变轻链、bap049克隆b可变重链和可变轻链、h7798n[pd
‑
1]可变重链和可变轻链、h1h3 var 6[pd
‑
1]可变重链和可变轻链、ape2058[pd
‑
1]可变重链和可变轻链、h005
‑
1[pd
‑
1]可变重链和可变轻链、317
‑
4b6[pd
‑
1]可变重链和可变轻链、326
‑
4a3[pd
‑
1]可变重链和可变轻链、hpd
‑
1mab 7(1.2)[pd
‑
1]可变重链和可变轻链、克隆38[pd
‑
1]可变重链和可变轻链、克隆39[pd
‑
1]可变重链和可变轻链、克隆41[pd
‑
1]可变重链和可变轻链、克隆48[pd
‑ꢀ
1]可变重链和可变轻链、pd1
‑
17[pd
‑
1]可变重链和可变轻链、pd1
‑
28[pd
‑
1]可变重链和可变轻链、pd1
‑
33[pd
‑
1]可变重链和可变轻链、pd1
‑
35[pd
‑
1]可变重链和可变轻链、lopd180可变重链和可变轻链、ab948可变重链和可变轻链、人源化eh
‑
12.2h7[pd
‑
1]可变重链和可变轻链、rg1h10可变重链和可变轻链、rg1h10
‑
h2a
‑
22
‑
1s可变重链和可变轻链、rg1h10
‑
h2a
‑ꢀ
27
‑
2s可变重链和可变轻链、rg1h10
‑
3c可变重链和可变轻链、rg1h10
‑
16c可变重链和可变轻链、rg1h10
‑
17c可变重链和可变轻链、rg1h10
‑
19c可变重链和可变轻链、rg1h10
‑ꢀ
21c可变重链和可变轻链、rg1h10
‑
23c2可变重链和可变轻链、mab7[pd
‑
1]以及pd1ab
‑ꢀ
6[pd
‑
1]可变重链和可变轻链,如图100a
‑
100g中所描绘。
[0056]
在一些实施方案中,非竞争性pd
‑
1结合结构域是不与纳武单抗和/或派姆单抗结合相同表位的人源化的非竞争性pd1 abd。在一些实施方案中,人源化的非竞争性pd
‑
1abd包含选自由以下组成的组的可变重链和可变轻链对:mab a[pd
‑
1]_h1_l1、mab b[pd
‑
1]_
h1_l1、 mab c[pd
‑
1]_h1_l1、mab c[pd
‑
1]_h1_l1.1、mab c[pd
‑
1]_h1_l1.3、mab c[pd
‑ꢀ
1]_h1_l1.45、mab c[pd
‑
1]_h1_l1.117、mab c[pd
‑
1]_h1_l1.129、mab c[pd
‑
1]_h1_l1.135、 mab c[pd
‑
1]_h1_l1.136、mab c[pd
‑
1]_h1_l1.140、mab c[pd
‑
1]_h1_l2、mab c[pd
‑ꢀ
1]_h1.19_l1、mab c[pd
‑
1]_h1.19_l1.1、mab c[pd
‑
1]_h1.19_l1.3、mab c[pd
‑ꢀ
1]_h1.19_l1.45、mab c[pd
‑
1]_h1.19_l1.117、mab c[pd
‑
1]_h1.19_l1.129、mab c[pd
‑ꢀ
1]_h1.19_l1.135、mab c[pd
‑
1]_h1.19_l1.136、mab c[pd
‑
1]_h1.19_l1.140、mab c[pd
‑ꢀ
1]_h1.19_l2、mab c[pd
‑
1]_h1.48_l1、mab c[pd
‑
1]_h1.48_l1.1、mab c[pd
‑
1]_h1.48_l1.3、 mab c[pd
‑
1]_h1.48_l1.45、mab c[pd
‑
1]_h1.48_l1.117、mab c[pd
‑
1]_h1.48_l1.129、mabc[pd
‑
1]_h1.48_l1.135、mab c[pd
‑
1]_h1.48_l1.136、mab c[pd
‑
1]_h1.48_l1.140、mab c[pd
‑ꢀ
1]_h1.48_l2、mab c[pd
‑
1]_h1.125_l1、mab c[pd
‑
1]_h1.125_l1.1、mab c[pd
‑ꢀ
1]_h1.125_l1.3、mab c[pd
‑
1]_h1.125_l1.45、mab c[pd
‑
1]_h1.125_l1.117、mab c[pd
‑ꢀ
1]_h1.125_l1.129、mab c[pd
‑
1]_h1.125_l1.135、mab c[pd
‑
1]_h1.125_l1.136、mab c[pd
‑ꢀ
1]_h1.125_l1.140、mab c[pd
‑
1]_h1.125_l2、mab c[pd
‑
1]_h1.130_l1、mab c[pd
‑ꢀ
1]_h1.130_l1.1、mab c[pd
‑
1]_h1.130_l1.3、mab c[pd
‑
1]_h1.130_l1.45、mab c[pd
‑ꢀ
1]_h1.130_l1.117、mab c[pd
‑
1]_h1.130_l1.129、mab c[pd
‑
1]_h1.130_l1.135、mab c[pd
‑ꢀ
1]_h1.130_l1.136、mab c[pd
‑
1]_h1.130_l1.140、mab c[pd
‑
1]_h1.130_l2、mab c[pd
‑ꢀ
1]_h1.132_l1、mab c[pd
‑
1]_h1.132_l1.1、mab c[pd
‑
1]_h1.132_l1.3、mab c[pd
‑ꢀ
1]_h1.132_l1.45、mab c[pd
‑
1]_h1.132_l1.117、mab c[pd
‑
1]_h1.132_l1.129、mab c[pd
‑ꢀ
1]_h1.132_l1.135、mab c[pd
‑
1]_h1.132_l1.136、mab c[pd
‑
1]_h1.132_l1.140、mab c[pd
‑ꢀ
1]_h1.132_l2、mab c[pd
‑
1]_h1.169_l1、mab c[pd
‑
1]_h1.169_l1.1、mab c[pd
‑ꢀ
1]_h1.169_l1.3、mab c[pd
‑
1]_h1.169_l1.45、mab c[pd
‑
1]_h1.169_l1.117、mab c[pd
‑ꢀ
1]_h1.169_l1.129、mab c[pd
‑
1]_h1.169_l1.135、mab c[pd
‑
1]_h1.169_l1.136、mab c[pd
‑ꢀ
1]_h1.169_l1.140、mab c[pd
‑
1]_h1.169_l2、mab c[pd
‑
1]_h1.175_l1、mab c[pd
‑ꢀ
1]_h1.175_l1.1、mab c[pd
‑
1]_h1.175_l1.3、mab c[pd
‑
1]_h1.175_l1.45、mab c[pd
‑ꢀ
1]_h1.175_l1.117、mab c[pd
‑
1]_h1.175_l1.129、mab c[pd
‑
1]_h1.175_l1.135、mab c[pd
‑ꢀ
1]_h1.175_l1.136、mab c[pd
‑
1]_h1.175_l1.140、mab c[pd
‑
1]_h1.175_l2、mab c[pd
‑ꢀ
1]_h2_l1、mab c[pd
‑
1]_h2_l1.1、mab c[pd
‑
1]_h2_l1.3、mab c[pd
‑
1]_h2_l1.45、mabc[pd
‑
1]_h2_l1.117、mab c[pd
‑
1]_h2_l1.129、mab c[pd
‑
1]_h2_l1.135、mab c[pd
‑ꢀ
1]_h2_l1.136、mab c[pd
‑
1]_h2_l1.140和mab c[pd
‑
1]_h2_l2,如图104a
‑
104b中所描绘。
[0057]
在一些实施方案中,cd8结合结构域是人源化的cd8 abd。在一些实施方案中,人源化的cd8 abd包含选自由如图92中所描绘的okt8_h2l1和1c11b3_h1l1组成的组的可变重链和可变轻链对。
[0058]
在一些实施方案中,nkg2d结合结构域是人源化的nkg2d abd。在一些实施方案中,人源化的nkg2d abd包含选自由以下组成的组的可变重链和可变轻链对:ms[nkg2d] h0_l0、1d7b4[nkg2d]_h1_l1、kyk
‑
1.0[nkg2d]_h1_l1、kyk
‑
2.0[nkg2d]_h0_l0、 11b2d10[nkg2d]_h0_l0、6e5a7[nkg2d]_h0_l0、6h7e7[nkg2d]_h0_l0、mabe[nkg2d]_h1_l1、16f31[nkg2d]_h1_l1、mab d[nkg2d]_h1_l1、 1d7b4[nkg2d]_h1_l1、mab a[nkg2d]_h1_l1、mab a[nkg2d]_h2_l1、maba[nkg2d]_h1_l2、mab a[nkg2d]_h2_l2、mab b[nkg2d]_h1_l1、mabb[nkg2d]_h1_l1.1、mab b[nkg2d]_h1_l2、mab b[nkg2d]_h2_l1、mabb[nkg2d]_h2_l1.1、
mab b[nkg2d]_h2_l2、mab b[nkg2d]_h3_l1、mabb[nkg2d]_h3_l1.1、mab b[nkg2d]_h3_l2、mab c[nkg2d]_h1_l1、mabc[nkg2d]_h2_l1、mab c[nkg2d]_h1_l2和mab c[nkg2d]_h2_l2,如图93a
‑
93c中所描绘。
[0059]
提供了一种包含本文概述的任何异源二聚体fc融合蛋白(例如,抗x)2
‑
异源fc
‑
scil10融合体)的组合物,其用于治疗受试者的癌症。还提供了一种或多种编码本文概述的任何异源二聚体fc融合蛋白的核酸。还提供了一种宿主细胞,其包含一种或多种编码本文概述的任何异源二聚体fc融合蛋白的核酸。
[0060]
在一个方面,提供了一种制备本文概述的任何异源二聚体fc融合蛋白(例如,抗x)2
‑
异源fc
‑
scil10融合体)的方法,其包括在借由产生异源二聚体fc融合蛋白的条件下培养所概述的宿主细胞;以及回收蛋白质。
[0061]
在另一方面,本发明提供了一种二聚体fc融合蛋白,其包含:(a)第一融合蛋白,其包含第一il
‑
10单体结构域和第一fc结构域,其中il
‑
10单体结构域与第一fc结构域共价连接;以及(b)第二融合蛋白,其包含第二il
‑
10单体结构域和第二fc结构域,其中第二il
‑
10单体结构域与第二fc结构域共价连接。在一些实施方案中,二聚体fc融合体具有(il
‑
10)2
‑
fc融合体形式。
[0062]
在一些实施方案中,第一和第二融合蛋白是相同的。在一些实施方案中,第一il
‑
10单体结构域和第二il
‑
10单体结构域是相同的。
[0063]
在一些实施方案中,第一和/或第二fc结构域包含选自由以下组成的组的一组氨基酸取代:根据eu编号的c219s、c220s、s228p、g236r/l328r、e233p/l234v/l235a/g236_/s239k、 e233p/l234v/l235a/g236_/s239k/a327g、e233p/l234v/l235a/g236_/s267k/a327g、 e233p/l234v/l235a/g236_和e233p/l234v/l235a/g236_/s267k。
[0064]
在一些实施方案中,第一和/或第二fc结构域具有另外一组包括根据eu编号的 q295e/n384d/q418e/n421d的氨基酸取代。
[0065]
在一些实施方案中,第一和/或第二fc结构域具有由以下组成的另外一组氨基酸取代:根据eu编号的g236r/l328r、e233p/l234v/l235a/g236_/s239k、 e233p/l234v/l235a/g236_/s239k/a327g、e233p/l234v/l235a/g236_/s267k/a327g、 e233p/l234v/l235a/g236_以及e233p/l234v/l235a/g236_/s267k。
[0066]
在一些实施方案中,第一和/或第二fc结构域包含选自由根据eu编号的m428l、n434s 和m428l/n434s组成的组的另一氨基酸取代。
[0067]
在一些实施方案中,第一il
‑
10单体结构域与第一fc结构域的n端共价连接并且第二 il
‑
10单体结构域与第二fc结构域的n端共价连接。在某些实施方案中,第一il
‑
10单体结构域与第一fc结构域的c端共价连接并且第二il
‑
10单体结构域与第二fc结构域的c端共价连接。
[0068]
在一些实施方案中,使用第一结构域接头将第一il
‑
10单体结构域与第一fc结构域连接,和/或使用第二结构域接头将第二il
‑
10单体结构域与第二fc结构域连接。在特定实施方案中,使用第一结构域接头将第一il
‑
10单体结构域与第一fc结构域连接,并且使用第二结构域接头将第二il
‑
10单体结构域与第二fc结构域连接。在另外的实施方案中,使用第一结构域接头将第一il
‑
10单体结构域与第一fc结构域连接。在另外的实施方案中,使用第二结构域接头将第二il
‑
10单体结构域与第二fc结构域连接。
[0069]
在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域在位置109处具有亮氨酸而不是组氨酸。换句话说,在一些实施方案中,第一il
‑
10单体结构域在位置109处具有亮氨酸而不是组氨酸。在一些实施方案中,第二il
‑
10单体结构域在位置109处具有亮氨酸而不是组氨酸。在一些实施方案中,第一il
‑
10单体结构域和第二il
‑
10单体结构域在位置109处具有亮氨酸而不是组氨酸。
[0070]
在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域在位置109处具有组氨酸而不是亮氨酸。换句话说,在一些实施方案中,第一il
‑
10单体结构域和第二il
‑
10 单体结构域在位置109处具有组氨酸而不是亮氨酸。在某些实施方案中,第一il
‑
10单体结构域在位置109处具有组氨酸而不是亮氨酸。在其他实施方案中,第二il
‑
10单体结构域在位置109处具有组氨酸而不是亮氨酸。
[0071]
在一些实施方案中,第一和/或第二il
‑
10单体结构域包含选自由以下组成的组的多肽序列:seq id no:1(人il
‑
10(109h)前体序列)、seq id no:2(人il
‑
10(109l)前体序列)、seqid no:3(人il
‑
10(109h)成熟形式序列)、seq id no:4(人il
‑
10(109l)成熟形式序列)。在一些实施方案中,第一il
‑
10单体结构域包含选自由以下组成的组的多肽序列:seq id no: 1(人il
‑
10(109h)前体序列)、seq id no:2(人il
‑
10(109l)前体序列)、seq id no:3(人il
‑ꢀ
10(109h)成熟形式序列)、seq id no:4(人il
‑
10(109l)成熟形式序列)。在某些实施方案中,第二il
‑
10单体结构域包含选自由以下组成的组的多肽序列:seq id no:1(人il
‑
10(109h) 前体序列)、seq id no:2(人il
‑
10(109l)前体序列)、seq id no:3(人il
‑
10(109h)成熟形式序列)、seq id no:4(人il
‑
10(109l)成熟形式序列)。
[0072]
在一些实施方案中,第一和/或第二il
‑
10单体结构域是变体il
‑
10单体结构域。在一些实施方案中,第一il
‑
10单体结构域是变体il
‑
10单体结构域。在一些实施方案中,第二il
‑ꢀ
10单体结构域是变体il
‑
10单体结构域。
[0073]
在一些实施方案中,变体il
‑
10单体结构域在选自由n21、d28、q38、m39、d41、q42、 l43、d44、n45、i87、e142、d144、e151和n160组成的组的氨基酸残基处包含一个或多个氨基酸修饰。在一些实施方案中,变体il
‑
10单体结构域包含选自由n21d、d28n、q38e、m39t、d41n、q42e、l43v、d44n、n45d、i87a、e142q、d144n、e151q和n160del组成的组的一个或多个氨基酸修饰。
[0074]
在一些实施方案中,(il
‑
10)2
‑
fc融合体形式的第一和第二融合蛋白各自包含选自由以下组成的组的多肽序列:xenp24628、xenp24629、xenp24630、xenp24631、xenp24632、 xenp24633和xenp24634,如图17
‑
18中所描绘。
[0075]
在一个方面,本发明提供了一种异源二聚体fc融合蛋白,其包含:a)第一融合蛋白,其包含第一il
‑
10单体结构域和第一fc结构域,其中第一il
‑
10单体结构域与第一fc结构域共价连接;以及(b)第二fc结构域;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰。在一些实施方案中,异源二聚体fc融合蛋白具有(il10
‑
nc
‑
il10)
‑
异源fc融合体形式。
[0076]
在一些实施方案中,异源二聚体fc融合蛋白还包含与第一il
‑
10单体结构域非共价连接的第二il
‑
10单体结构域。在一些实施方案中,第一il
‑
10单体结构域与第一fc结构域的n 端连接。在一些实施方案中,第一il
‑
10单体结构域与第一fc结构域的c端连接。在一些实施方案中,使用第一结构域接头将第一il
‑
10单体结构域与第一fc结构域连接。
m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。
[0087]
在一些实施方案中,插入肽包含工程化在il
‑
10单体结构域的螺旋d与e之间的结构域接头。
[0088]
在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域包含选自由以下组成的组的多肽序列:如图15d中所描绘的seq id no:24(il10m1)和如图75a
‑
75b中所描绘的huil10m1变体。
[0089]
在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域包括变体il
‑
10结构域,所述变体il
‑
10结构域包含选自由以下组成的组的一个或多个氨基酸修饰:n21d、d28n、q38e、m39t、d41n、q42e、l43v、d44n、n45d、i87a、e142q、d144n、e151q、 c12a、c108a、q38e/d41n、q38e/q42e、q38e/n45d、q38e/e142q、q38e/d144n、 d41n/q42e、d41n/n45d、d41n/e142q、d41n/d144n、q42e/n45d、q42e/e142q、 q42e/d144n、n45d/e142q、n45d/d144n、e142q/d144n、n21d/q42e、n21d/n45d、 n21d/e151q、n21d/n45e、q42e/e151q、n45d/e151q、n21d/q42e/n45d、 n21d/q42e/e151q、q42e/n45d/e151q、n21d/q42e/n45e、n21d/q42e/n45d/e151q、 f37c/m140c、q38c/s141c、d41c/k138c、l47c/k138c、l48c/e142c、s51c/a120c、 d55c/a120c、f56c/y153c、c62a/c114a、a64c/s118c、m68c/v121c、v76c/a139c、l47q、 s118a和a139q。
[0090]
在一些实施方案中,第一融合蛋白和第二融合蛋白是相同的。在一些实施方案中, (il10m1)2
‑
fc融合体的第一融合蛋白和第二融合蛋白各自包含xenp25236的多肽序列,如图26中所描绘。在一些实施方案中,(il10m1)2
‑
fc融合体的第一融合蛋白和第二融合蛋白各自包含xenp25237的多肽序列,如图27中所描绘。
[0091]
在另一方面,本发明提供一种异源二聚体fc融合蛋白,其包含:(a)融合蛋白,其包含第一蛋白结构域、第二蛋白结构域和第一fc结构域,其中第一蛋白结构域与第一fc结构域连接,并且其中第二蛋白结构域与第一蛋白结构域共价连接;以及(b)第二fc结构域;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一蛋白结构域包含第一il10单体并且第二蛋白结构域包含第二il10单体,并且其中第一和第二il10 单体中的每一个包含插入肽。异源二聚体fc融合蛋白具有(il10m1)2
‑
异源fc融合体形式。
[0092]
在一些实施方案中,第一蛋白结构域与第一fc结构域的n端连接。在一些实施方案中,第一蛋白结构域与第一fc结构域的c端连接。在一些实施方案中,使用第一结构域接头将第一蛋白结构域与第二蛋白结构域连接,和/或使用第二结构域接头将第一蛋白结构域与第一 fc结构域连接。
[0093]
在一些实施方案中,促进第一和第二fc结构域的异源二聚化的修饰是选自由以下组成的组的一组氨基酸取代:根据eu编号的l368d/k370s和s364k;l368d/k370s和s364k/e357l; l368d/k370s和s364k/e357q;t411e/k360e/q362e和d401k;l368e/k370s和s364k; k370s和s364k/e357q以及t366s/l368a/y407v:t366w(任选地包括二硫桥键, t366s/l368a/y407v/y349c:t366w/s354c)。
[0094]
在一些实施方案中,第一和/或第二fc结构域具有另外一组包括根据eu编号的 q295e/n384d/q418e/n421d的氨基酸取代。
[0095]
在一些实施方案中,第一和/或第二fc结构域具有由以下组成的另外一组氨基酸
取代:根据eu编号的g236r/l328r、e233p/l234v/l235a/g236_/s239k、 e233p/l234v/l235a/g236_/s239k/a327g、e233p/l234v/l235a/g236_/s267k/a327g、 e233p/l234v/l235a/g236_以及e233p/l234v/l235a/g236_/s267k。
[0096]
在一些实施方案中,第一第二fc结构域和/或第二fc结构域包含选自由根据eu编号的 m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。
[0097]
在一些实施方案中,插入肽包含工程化在il
‑
10单体结构域的螺旋d与e之间的结构域接头。
[0098]
在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域包含选自由以下组成的组的多肽序列:如图15d中所描绘的seq id no:24(il10m1)和如图75a
‑
75b中所描绘的huil10m1变体。
[0099]
在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域包括变体il
‑
10结构域。在一些实施方案中,变体il
‑
10结构域包含选自由以下组成的组的一个或多个氨基酸修饰:n21d、d28n、q38e、m39t、d41n、q42e、l43v、d44n、n45d、i87a、e142q、 d144n、e151q、c12a、c108a、q38e/d41n、q38e/q42e、q38e/n45d、q38e/e142q、 q38e/d144n、d41n/q42e、d41n/n45d、d41n/e142q、d41n/d144n、q42e/n45d、 q42e/e142q、q42e/d144n、n45d/e142q、n45d/d144n、e142q/d144n、n21d/q42e、 n21d/n45d、n21d/e151q、n21d/n45e、q42e/e151q、n45d/e151q、n21d/q42e/n45d、 n21d/q42e/e151q、q42e/n45d/e151q、n21d/q42e/n45e、n21d/q42e/n45d/e151q、 f37c/m140c、q38c/s141c、d41c/k138c、l47c/k138c、l48c/e142c、s51c/a120c、 d55c/a120c、f56c/y153c、c62a/c114a、a64c/s118c、m68c/v121c、v76c/a139c、l47q、 s118a和a139q。
[0100]
在另一方面,本发明提供一种异源二聚体fc融合蛋白,其包含:(a)融合蛋白,其包含第一蛋白结构域和第一fc结构域,其中第一蛋白结构域与第一fc结构域共价连接;以及(b)第二fc结构域;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一蛋白结构域包含含有插入肽的il
‑
10单体。异源二聚体fc融合体具有 (il10m1)1
‑
异源fc融合体形式。
[0101]
在一些实施方案中,第一蛋白结构域(例如,包含插入肽的il
‑
10单体)与第一fc结构域的n端连接。在一些实施方案中,第一蛋白结构域与第一fc结构域的c端连接。在一些实施方案中,使用结构域接头将第一蛋白结构域与第一fc结构域连接。
[0102]
在一些实施方案中,促进第一和第二fc结构域的异源二聚化的修饰是选自由以下组成的组的一组氨基酸取代:根据eu编号的l368d/k370s和s364k;l368d/k370s和s364k/e357l; l368d/k370s和s364k/e357q;t411e/k360e/q362e和d401k;l368e/k370s和s364k; k370s和s364k/e357q以及t366s/l368a/y407v:t366w(任选地包括二硫桥键, t366s/l368a/y407v/y349c:t366w/s354c)。
[0103]
在一些实施方案中,第一和/或第二fc结构域具有另外一组包括根据eu编号的 q295e/n384d/q418e/n421d的氨基酸取代。
[0104]
在一些实施方案中,第一和/或第二fc结构域具有由以下组成的另外一组氨基酸取代:根据eu编号的g236r/l328r、e233p/l234v/l235a/g236_/s239k、 e233p/l234v/l235a/g236_/s239k/a327g、e233p/l234v/l235a/g236_/s267k/a327g、 e233p/l234v/l235a/g236_以及e233p/l234v/l235a/g236_/s267k。
[0105]
在一些实施方案中,第一fc结构域和/或第二fc结构域包含选自由根据eu编号的 m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。
[0106]
在一些实施方案中,插入肽包含工程化在il
‑
10单体结构域的螺旋d与e之间的结构域接头。
[0107]
在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域包含选自由以下组成的组的多肽序列:如图15d中所描绘的seq id no:24(il10m1)和如图75a
‑
75b中所描绘的huil10m1变体。
[0108]
在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域包括变体il
‑
10结构域,所述变体il
‑
10结构域包含选自由以下组成的组的一个或多个氨基酸修饰:n21d、 d28n、q38e、m39t、d41n、q42e、l43v、d44n、n45d、i87a、e142q、d144n、e151q、 c12a、c108a、q38e/d41n、q38e/q42e、q38e/n45d、q38e/e142q、q38e/d144n、 d41n/q42e、d41n/n45d、d41n/e142q、d41n/d144n、q42e/n45d、q42e/e142q、 q42e/d144n、n45d/e142q、n45d/d144n、e142q/d144n、n21d/q42e、n21d/n45d、 n21d/e151q、n21d/n45e、q42e/e151q、n45d/e151q、n21d/q42e/n45d、 n21d/q42e/e151q、q42e/n45d/e151q、n21d/q42e/n45e、n21d/q42e/n45d/e151q、 f37c/m140c、q38c/s141c、d41c/k138c、l47c/k138c、l48c/e142c、s51c/a120c、 d55c/a120c、f56c/y153c、c62a/c114a、a64c/s118c、m68c/v121c、v76c/a139c、l47q、 s118a和a139q。
[0109]
在另一方面,本发明提供一种异源二聚体fc融合蛋白,其包含:(a)第一融合蛋白,其包含第一蛋白结构域和第一fc结构域,其中第一蛋白结构域与第一fc结构域共价连接;以及 (b)第二融合蛋白,其包含第二蛋白结构域和第二fc结构域,其中第二蛋白结构域与第二fc 结构域共价连接;其中第一蛋白结构域包含il
‑
10单体结构域的螺旋a
‑
d并且第二蛋白结构域包含il
‑
10单体结构域的螺旋e
‑
f。异源二聚体fc融合蛋白具有(分裂il10)1
‑
异源fc形式。
[0110]
在一些实施方案中,第一蛋白结构域与第一fc结构域的n端共价连接,和/或第二蛋白结构域与第二fc结构域的n端共价连接。
[0111]
在一些实施方案中,第一蛋白结构域与第一fc结构域的c端共价连接,和/或第二蛋白结构域与第二fc结构域的c端共价连接。
[0112]
在一些实施方案中,第一蛋白结构域通过第一结构域接头与第一fc结构域共价连接,和 /或第二蛋白结构域通过第二结构域接头与第二fc结构域共价连接。
[0113]
在一些实施方案中,第一fc结构域和/或第二fc结构域包含选自由根据eu编号的 m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。
[0114]
在一些实施方案中,第一蛋白结构域包含如图15a中所描绘的seq id no:21(hl
‑
10(a
‑ꢀ
d))的多肽序列或与seq id no:21具有至少90%(例如,90%、91%、92%、93%、94%、95%、 96%、97%、98%、99%或更高)序列同一性的多肽序列。在一些实施方案中,第二蛋白结构域包含如图15b中所描绘的seq id no:22(hl
‑
10(e
‑
f))的多肽序列或与seq id no:22具有至少90%(例如,90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更高)序列同一性的多肽序列。
[0115]
在一些实施方案中,(分裂il10)1
‑
异源fc形式的异源二聚体fc融合蛋白包含:(a)第一融合蛋白,其具有如图30中所描绘的xenp25242链1的多肽序列,以及(b)第二融合蛋
白,其具有如图30中所描绘的xenp25242链2的多肽序列。
[0116]
在一些实施方案中,(分裂il10)1
‑
异源fc形式的异源二聚体fc融合蛋白包含:(a)第一融合蛋白,其具有如图30中所描绘的xenp25243链1的多肽序列,以及(b)第二融合蛋白,其具有如图30中所描绘的xenp25243链2的多肽序列。
[0117]
在一些实施方案中,(分裂il10)1
‑
异源fc形式的异源二聚体fc融合蛋白包含:(a)第一融合蛋白,其具有如图30中所描绘的xenp25244链1的多肽序列,以及(b)第二融合蛋白,其具有如图30中所描绘的xenp25244链2的多肽序列。
[0118]
提供了一种核酸,其编码本文所述的任何异源二聚体融合蛋白。提供了一种宿主细胞,其包含编码本文所述的任何异源二聚体融合蛋白的核酸。
[0119]
在一些实施方案中,提供一种制备异源二聚体fc融合蛋白(例如,(分裂il10)1
‑
异源fc 融合蛋白)的方法,其包括在借由产生异源二聚体fc融合蛋白的条件下培养所描述的宿主细胞;以及回收异源二聚体fc融合蛋白。
[0120]
在另一方面,本发明提供一种异源二聚体fc融合蛋白,其包含:(a)第一融合蛋白,其包含第一蛋白结构域、第二蛋白结构域和第一fc结构域,其中第一蛋白结构域与第二蛋白结构域共价连接,并且第二蛋白结构域与第一fc结构域共价连接;以及(b)第二融合蛋白,其包含第三蛋白结构域、第四蛋白结构域和第二fc结构域,其中第三蛋白结构域与第四蛋白结构域共价连接,并且第四蛋白结构域与第二fc结构域共价连接;其中第一蛋白结构域和第二蛋白结构域各自包含il
‑
10单体结构域的螺旋a
‑
d,并且其中第三蛋白结构域和第四蛋白结构域各自包含il
‑
10单体结构域的螺旋e
‑
f。异源二聚体fc融合蛋白具有(分裂il10)2
‑
异源fc融合体形式。
[0121]
在一些实施方案中,第二蛋白与第一fc结构域的n端共价连接,和/或第四蛋白结构域与第二fc结构域的n端共价连接。
[0122]
在一些实施方案中,第二蛋白与第一fc结构域的c端共价连接,和/或第四蛋白结构域与第二fc结构域的c端共价连接。
[0123]
在一些实施方案中,第一蛋白结构域通过第一结构域接头与第二蛋白结构域共价连接,和/或第三蛋白结构域通过第二结构域接头与第三蛋白结构域共价连接。
[0124]
在一些实施方案中,第二蛋白结构域通过第三结构域接头与第一fc结构域共价连接,和 /或第四蛋白结构域通过第四结构域接头与第二fc结构域共价连接。
[0125]
在一些实施方案中,第一和/或第二fc结构域包含选自由根据eu编号的m428l、n434s 和m428l/n434s组成的组的另一氨基酸取代。在一个实施方案中,第一fc结构域包含选自由m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。在一个实施方案中,第二 fc结构域包含选自由m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。在一些实施方案中,第一和第二fc结构域各自包含选自由m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。
[0126]
在一些实施方案中,第一蛋白结构域和/或第二蛋白结构域包含如图15a中所描绘的seqid no:21(hil
‑
10(a
‑
d))的多肽序列或与seq id no:21具有至少90%(例如,90%、91%、 92%、93%、94%、95%、96%、97%、98%、99%或更高)序列同一性的多肽序列。在一些实施方案中,第三蛋白结构域和/或第四蛋白结构域包含如图15b中所描绘的seq id no:22(hil
‑ꢀ
10(e
‑
f))的多肽序列或与seq id no:22具有至少90%(例如,90%、91%、92%、
93%、94%、95%、96%、97%、98%、99%或更高)序列同一性的多肽序列。
[0127]
提供了一种核酸,其编码本文所述的任何异源二聚体融合蛋白(例如,(分裂il10)2
‑
异源 fc融合体)。提供了一种宿主细胞,其包含编码本文所述的任何异源二聚体融合蛋白的核酸。
[0128]
在一些实施方案中,提供了一种制备异源二聚体fc融合蛋白(例如,(分裂il10)2
‑
异源fc 融合蛋白)的方法,其包括在借由产生异源二聚体fc融合蛋白的条件下培养所描述的宿主细胞;以及回收异源二聚体fc融合蛋白。
[0129]
在另一方面,本发明提供一种异源二聚体fc融合蛋白,其包含:(a)第一融合蛋白,其包含抗原结合结构域(abd)和第一fc结构域,其中抗原结合结构域与第一fc结构域的n端共价连接;以及(b)第二融合蛋白,其包含蛋白结构域和第二fc结构域,其中蛋白结构域与第二 fc结构域的n端共价连接,并且其中蛋白结构域包含第一il
‑
10单体结构域。异源二聚体fc 融合蛋白具有抗x x il10
‑
异源fc融合体形式。
[0130]
在一些实施方案中,异源二聚体fc融合蛋白还包含与第一il
‑
10单体结构域非共价连接的第二il
‑
10单体结构域。
[0131]
在一些实施方案中,使用第一结构域接头将抗原结合结构域与第一fc结构域连接。在一些实施方案中,使用第二结构域接头将第一il
‑
10单体结构域与第二fc结构域连接。
[0132]
在一些实施方案中,促进第一和第二fc结构域的异源二聚化的修饰是选自由以下组成的组的一组氨基酸取代:根据eu编号的l368d/k370s和s364k;l368d/k370s和s364k/e357l; l368d/k370s和s364k/e357q;t411e/k360e/q362e和d401k;l368e/k370s和s364k; k370s和s364k/e357q以及t366s/l368a/y407v:t366w(任选地包括二硫桥键, t366s/l368a/y407v/y349c:t366w/s354c)。
[0133]
在一些实施方案中,第一和/或第二fc结构域具有另外一组包括根据eu编号的 q295e/n384d/q418e/n421d的氨基酸取代。
[0134]
在一些实施方案中,第一和/或第二fc结构域具有由以下组成的另外一组氨基酸取代:根据eu编号的g236r/l328r、e233p/l234v/l235a/g236_/s239k、 e233p/l234v/l235a/g236_/s239k/a327g、e233p/l234v/l235a/g236_/s267k/a327g、 e233p/l234v/l235a/g236_以及e233p/l234v/l235a/g236_/s267k。
[0135]
在一些实施方案中,第一fc结构域和/或第二fc结构域包含选自由根据eu编号的 m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。
[0136]
在一些实施方案中,第一和/或第二il
‑
10单体结构域包含选自由以下组成的组的多肽序列:seq id no:1(人il
‑
10(109h)前体序列)、seq id no:2(人il
‑
10(109l)前体序列)、seq id no:3(人il
‑
10(109h)成熟形式序列)、seq id no:4(人il
‑
10(109l)成熟形式序列),如图1中所描绘。
[0137]
在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域是变体il
‑
10单体结构域。
[0138]
在一些实施方案中,变体il
‑
10单体结构域包括具有一个或多个氨基酸取代的il
‑
10单体结构域,所述氨基酸取代导致对il
‑
10受体的亲和力改变、效力改变、脱酰胺作用的潜力改变、天冬氨酸异构化的潜力改变、降解相关ptm的潜力改变、潜在降解位点改变、二硫
桥改变和/或潜在n
‑
糖基化位点改变。在一些实施方案中,变体il
‑
10单体结构域在选自由n21、 d28、q38、m39、d41、q42、l43、d44、n45、i87、e142、d144、e151和n160组成的组的氨基酸残基处包含一个或多个氨基酸修饰。在一些实施方案中,变体il
‑
10单体结构域包含选自由n21d、d28n、q38e、m39t、d41n、q42e、l43v、d44n、n45d、i87a、e142q、 d144n、e151q和n160del组成的组的一个或多个氨基酸修饰。
[0139]
在一些实施方案中,抗原结合结构域(abd)选自由pd
‑
1结合结构域、非竞争性pd
‑
1结合结构域、tigit结合结构域、cd8结合结构域和nkg2d结合结构域组成的组。
[0140]
在一些实施方案中,pd
‑
1结合结构域是人源化的pd
‑
1abd。在一些实施方案中,人源化的pd
‑
1abd包含选自由以下组成的组的可变重链和可变轻链对:1c11[pd
‑
1]_h0l0、 1c11[p,d
‑
1]_h3l3、1c11[pd
‑
1]_h3.240_l3.148、1c11[pd
‑
1]_h3.241_l3.148、1c11[pd
‑ꢀ
1]_h3.234_l3.144、1c11[pd
‑
1]_h3.241_l3.92、1c11[pd
‑
1]_h3.303_l3.152、 1c11_h3.329_l3.220、1c11_h3.328_l3.152、派姆单抗可变重链和可变轻链、纳武单抗可变重链和可变轻链、皮地利珠单抗可变重链和可变轻链、mk
‑
3475可变重链和可变轻链、bap049 克隆e可变重链和可变轻链、bap049克隆b可变重链和可变轻链、h7798n[pd
‑
1]可变重链和可变轻链、h1h3 var 6[pd
‑
1]可变重链和可变轻链、ape2058[pd
‑
1]可变重链和可变轻链、 h005
‑
1[pd
‑
1]可变重链和可变轻链、317
‑
4b6[pd
‑
1]可变重链和可变轻链、326
‑
4a3[pd
‑
1]可变重链和可变轻链、hpd
‑
1mab 7(1.2)[pd
‑
1]可变重链和可变轻链、克隆38[pd
‑
1]可变重链和可变轻链、克隆39[pd
‑
1]可变重链和可变轻链、克隆41[pd
‑
1]可变重链和可变轻链、克隆48[pd
‑ꢀ
1]可变重链和可变轻链、pd1
‑
17[pd
‑
1]可变重链和可变轻链、pd1
‑
28[pd
‑
1]可变重链和可变轻链、pd1
‑
33[pd
‑
1]可变重链和可变轻链、pd1
‑
35[pd
‑
1]可变重链和可变轻链、lopd180可变重链和可变轻链、ab948可变重链和可变轻链、人源化eh
‑
12.2h7[pd
‑
1]可变重链和可变轻链、rg1h10可变重链和可变轻链、rg1h10
‑
h2a
‑
22
‑
1s可变重链和可变轻链、rg1h10
‑
h2a
‑ꢀ
27
‑
2s可变重链和可变轻链、rg1h10
‑
3c可变重链和可变轻链、rg1h10
‑
16c可变重链和可变轻链、rg1h10
‑
17c可变重链和可变轻链、rg1h10
‑
19c可变重链和可变轻链、rg1h10
‑ꢀ
21c可变重链和可变轻链、rg1h10
‑
23c2可变重链和可变轻链、mab7[pd
‑
1]以及pd1ab
‑ꢀ
6[pd
‑
1]可变重链和可变轻链,如图100a
‑
100g中所描绘。
[0141]
在一些实施方案中,非竞争性pd
‑
1结合结构域是不与纳武单抗和/或派姆单抗结合相同表位的人源化的非竞争性pd1 abd。在一些实施方案中,人源化的非竞争性pd
‑
1abd包含选自由以下组成的组的可变重链和可变轻链对:mab a[pd
‑
1]_h1_l1、mab b[pd
‑
1]_h1_l1、 mab c[pd
‑
1]_h1_l1、mab c[pd
‑
1]_h1_l1.1、mab c[pd
‑
1]_h1_l1.3、mab c[pd
‑ꢀ
1]_h1_l1.45、mab c[pd
‑
1]_h1_l1.117、mab c[pd
‑
1]_h1_l1.129、mab c[pd
‑
1]_h1_l1.135、 mab c[pd
‑
1]_h1_l1.136、mab c[pd
‑
1]_h1_l1.140、mab c[pd
‑
1]_h1_l2、mab c[pd
‑ꢀ
1]_h1.19_l1、mab c[pd
‑
1]_h1.19_l1.1、mab c[pd
‑
1]_h1.19_l1.3、mab c[pd
‑ꢀ
1]_h1.19_l1.45、mab c[pd
‑
1]_h1.19_l1.117、mab c[pd
‑
1]_h1.19_l1.129、mab c[pd
‑ꢀ
1]_h1.19_l1.135、mab c[pd
‑
1]_h1.19_l1.136、mab c[pd
‑
1]_h1.19_l1.140、mab c[pd
‑ꢀ
1]_h1.19_l2、mab c[pd
‑
1]_h1.48_l1、mab c[pd
‑
1]_h1.48_l1.1、mab c[pd
‑
1]_h1.48_l1.3、 mab c[pd
‑
1]_h1.48_l1.45、mab c[pd
‑
1]_h1.48_l1.117、mab c[pd
‑
1]_h1.48_l1.129、mabc[pd
‑
1]_h1.48_l1.135、mab c[pd
‑
1]_h1.48_l1.136、mab c[pd
‑
1]_h1.48_l1.140、mab c[pd
‑ꢀ
1]_h1.48_l2、mab c[pd
‑
1]_h1.125_l1、mab c[pd
‑
1]_h1.125_l1.1、mab c[pd
‑ꢀ
1]_
h1.125_l1.3、mab c[pd
‑
1]_h1.125_l1.45、mab c[pd
‑
1]_h1.125_l1.117、mab c[pd
‑ꢀ
1]_h1.125_l1.129、mab c[pd
‑
1]_h1.125_l1.135、mab c[pd
‑
1]_h1.125_l1.136、mab c[pd
‑ꢀ
1]_h1.125_l1.140、mab c[pd
‑
1]_h1.125_l2、mab c[pd
‑
1]_h1.130_l1、mab c[pd
‑ꢀ
1]_h1.130_l1.1、mab c[pd
‑
1]_h1.130_l1.3、mab c[pd
‑
1]_h1.130_l1.45、mab c[pd
‑ꢀ
1]_h1.130_l1.117、mab c[pd
‑
1]_h1.130_l1.129、mab c[pd
‑
1]_h1.130_l1.135、mab c[pd
‑ꢀ
1]_h1.130_l1.136、mab c[pd
‑
1]_h1.130_l1.140、mab c[pd
‑
1]_h1.130_l2、mab c[pd
‑ꢀ
1]_h1.132_l1、mab c[pd
‑
1]_h1.132_l1.1、mab c[pd
‑
1]_h1.132_l1.3、mab c[pd
‑ꢀ
1]_h1.132_l1.45、mab c[pd
‑
1]_h1.132_l1.117、mab c[pd
‑
1]_h1.132_l1.129、mab c[pd
‑ꢀ
1]_h1.132_l1.135、mab c[pd
‑
1]_h1.132_l1.136、mab c[pd
‑
1]_h1.132_l1.140、mab c[pd
‑ꢀ
1]_h1.132_l2、mab c[pd
‑
1]_h1.169_l1、mab c[pd
‑
1]_h1.169_l1.1、mab c[pd
‑ꢀ
1]_h1.169_l1.3、mab c[pd
‑
1]_h1.169_l1.45、mab c[pd
‑
1]_h1.169_l1.117、mab c[pd
‑ꢀ
1]_h1.169_l1.129、mab c[pd
‑
1]_h1.169_l1.135、mab c[pd
‑
1]_h1.169_l1.136、mab c[pd
‑ꢀ
1]_h1.169_l1.140、mab c[pd
‑
1]_h1.169_l2、mab c[pd
‑
1]_h1.175_l1、mab c[pd
‑ꢀ
1]_h1.175_l1.1、mab c[pd
‑
1]_h1.175_l1.3、mab c[pd
‑
1]_h1.175_l1.45、mab c[pd
‑ꢀ
1]_h1.175_l1.117、mab c[pd
‑
1]_h1.175_l1.129、mab c[pd
‑
1]_h1.175_l1.135、mab c[pd
‑ꢀ
1]_h1.175_l1.136、mab c[pd
‑
1]_h1.175_l1.140、mab c[pd
‑
1]_h1.175_l2、mab c[pd
‑ꢀ
1]_h2_l1、mab c[pd
‑
1]_h2_l1.1、mab c[pd
‑
1]_h2_l1.3、mab c[pd
‑
1]_h2_l1.45、mabc[pd
‑
1]_h2_l1.117、mab c[pd
‑
1]_h2_l1.129、mab c[pd
‑
1]_h2_l1.135、mab c[pd
‑ꢀ
1]_h2_l1.136、mab c[pd
‑
1]_h2_l1.140和mab c[pd
‑
1]_h2_l2,如图104a
‑
104b中所描绘。
[0142]
在一些实施方案中,cd8结合结构域是人源化的cd8 abd。在一些实施方案中,人源化的cd8 abd包含选自由如图92中所描绘的okt8_h2l1和1c11b3_h1l1组成的组的可变重链和可变轻链对。
[0143]
在一些实施方案中,nkg2d结合结构域是人源化的nkg2d abd。在一些实施方案中,人源化的nkg2d abd包含选自由以下组成的组的可变重链和可变轻链对:ms[nkg2d] h0_l0、1d7b4[nkg2d]_h1_l1、kyk
‑
1.0[nkg2d]_h1_l1、kyk
‑
2.0[nkg2d]_h0_l0、 11b2d10[nkg2d]_h0_l0、6e5a7[nkg2d]_h0_l0、6h7e7[nkg2d]_h0_l0、mabe[nkg2d]_h1_l1、16f31[nkg2d]_h1_l1、mab d[nkg2d]_h1_l1、 1d7b4[nkg2d]_h1_l1、mab a[nkg2d]_h1_l1、mab a[nkg2d]_h2_l1、maba[nkg2d]_h1_l2、mab a[nkg2d]_h2_l2、mab b[nkg2d]_h1_l1、mabb[nkg2d]_h1_l1.1、mab b[nkg2d]_h1_l2、mab b[nkg2d]_h2_l1、mabb[nkg2d]_h2_l1.1、mab b[nkg2d]_h2_l2、mab b[nkg2d]_h3_l1、mabb[nkg2d]_h3_l1.1、mab b[nkg2d]_h3_l2、mab c[nkg2d]_h1_l1、mabc[nkg2d]_h2_l1、mab c[nkg2d]_h1_l2和mab c[nkg2d]_h2_l2,如图93a
‑
93c中所描绘。
[0144]
提供了一种核酸,其编码抗x x il
‑
10
‑
异源fc融合体形式的本文所述的任何异源二聚体融合蛋白。提供了一种宿主细胞,其包含编码本文所述的任何异源二聚体融合蛋白的核酸。
[0145]
在一些实施方案中,提供了一种制备异源二聚体fc融合蛋白(例如,抗x x il
‑
10
‑
异源fc 融合蛋白)的方法,其包括在借由产生异源二聚体fc融合蛋白的条件下培养所描述的宿主细胞;以及回收异源二聚体fc融合蛋白。
[0146]
在一些实施方案中,本文提供一种纯化抗x x il
‑
10
‑
异源fc融合体形式的本文所
述的异源二聚体fc融合蛋白的方法。所述方法包括:(a)提供包含异源二聚体fc融合蛋白的组合物; (b)将组合物上样到离子交换柱上;以及(c)收集含有异源二聚体fc融合蛋白的级分。
附图说明
[0147]
图1描绘人il
‑
10(il
‑
10(109h)和il
‑
10(109l)序列)及其受体的序列。
[0148]
图2描绘小鼠il
‑
10及其受体的序列,以有利于在临床前研究中研究本发明的il
‑
10融合蛋白。
[0149]
图3描绘食蟹猴il
‑
10及其受体的序列,以有利于在临床前研究中研究本发明的il
‑
10融合蛋白。
[0150]
图4描绘病毒il
‑
10同源物的序列。
[0151]
图5描绘fc异源二聚化变体组的有用对(包括偏斜和pi变体)。没有对应的“单体2”的变体是可以在任一单体上单独使用的pi变体。
[0152]
图6描绘等构变体抗体恒定区和它们各自的取代的列表。pi_(
‑
)表示较低的pi变体,而 pi_( )表示较高的pi变体。这些可以任选地且独立地与本发明的其它异源二聚化变体(以及其它变体类型,如本文所概述的)组合。
[0153]
图7描绘消融了fcγr结合的有用消融变体(有时称为“敲除”或“ko”变体)。通常,在两种单体上都发现了消融变体,尽管在一些情况下它们可能仅在一种单体上。
[0154]
图8示出本发明的il
‑
10融合体的“非细胞因子”组分的特别有用的实施方案。
[0155]
图9示出本发明的“非细胞因子”/“非fv”组分的特别有用的实施方案。
[0156]
图10示出基于人igg1的几个有用的同源二聚体il
‑
10融合体骨架的序列,而无细胞因子序列。同源二聚体fc骨架1基于人igg1(356e/358m同种异型),并且包括 e233p/l234v/l235a/g236del/s267k消融变体和c220s。同源二聚体fc骨架2基于人igg1 (356d/358l同种异型),并且包括e233p/l234v/l235a/g236del/s267k消融变体和c220s。同源二聚体fc骨架3基于人igg4和消融了fab臂交换(如本领域已知的)的s228p(根据eu编号;在kabat中为s241p)变体。同源二聚体fc骨架4基于人igg2,并且包括s267k消融变体。同源二聚体骨架4的替代形式可以包括c219s和/或c220s。这些序列可以与利用同源二聚体fc区的本发明的任何il
‑
10融合体一起使用。这些序列还可以与利用同源二聚体fc区的本发明的靶向性il
‑
10融合体的形式一起使用。在包括与fc共价连接的可变重结构域的靶向性il
‑
10融合体形式中,可变重结构域可通过相应的ch1结构域和部分铰链区与fc结构域共价连接,其例示性序列描绘于图12中,并且可排除c220s和/或c219s取代。
[0157]
图11a
‑
11b示出基于人igg1的几种有用的异源二聚体il
‑
10融合体骨架的序列,而无细胞因子序列。虽然这些骨架可用于本发明的il
‑
10融合体,但它们也可用于本发明的靶向性 il
‑
10融合体的形式。异源二聚体fc骨架1基于人igg1(356e/358m同种异型),并包括处于第一异源二聚体fc链上的l368d/k370s偏斜变体和q295e/n384d/q418e/n421d pi变体、处于第二异源二聚体fc链上的s364k/e357q偏斜变体,以及处于两条链上的 e233p/l234v/l235a/g236del/s267k消融变体和c220s。异源二聚体fc骨架2基于人igg1 (356e/358m同种异型),并包括处于第一异源二聚体fc链上的l368d/k370s偏斜变体和 q295e/n384d/q418e/n421d pi变体、处于第二异源二聚体fc链上的s364k偏斜变体,以及处于两条
链上的e233p/l234v/l235a/g236del/s267k消融变体和c220s。异源二聚体fc骨架3基于人igg1(356e/358m同种异型),并包括处于第一异源二聚体fc链上的l368e/k370s 偏斜变体和q295e/n384d/q418e/n421d pi变体、处于第二异源二聚体fc链上的s364k偏斜变体,以及处于两条链上的e233p/l234v/l235a/g236del/s267k消融变体和c220s。异源二聚体fc骨架4基于人igg1(356e/358m同种异型),并包括处于第一异源二聚体fc链上的 k360e/q362e/t411e偏斜变体和q295e/n384d/q418e/n421d pi变体、处于第二异源二聚体 fc链上的d401k偏斜变体,以及处于两条链上的e233p/l234v/l235a/g236del/s267k消融变体和c220s。异源二聚体fc骨架5基于人igg1(356d/358l同种异型),并包括处于第一异源二聚体fc链上的l368d/k370s偏斜变体和q295e/n384d/q418e/n421d pi变体、处于第二异源二聚体fc链上的s364k/e357q偏斜变体,以及处于两条链上的 e233p/l234v/l235a/g236del/s267k消融变体和c220s。异源二聚体fc骨架6基于人igg1 (356e/358m同种异型),并包括处于第一异源二聚体fc链上的l368d/k370s偏斜变体和 q295e/n384d/q418e/n421d pi变体、处于第二异源二聚体fc链上的s364k/e357q偏斜变体,以及处于两条链上的e233p/l234v/l235a/g236del/s267k消融变体、去除糖基化的n297a 变体以及c220s。异源二聚体fc骨架7基于人igg1(356e/358m同种异型),并包括处于第一异源二聚体fc链上的l368d/k370s偏斜变体和q295e/n384d/q418e/n421d pi变体、处于第二异源二聚体fc链上的s364k/e357q偏斜变体,以及处于两条链上的 e233p/l234v/l235a/g236del/s267k消融变体、去除糖基化的n297s变体以及c220s。异源二聚体fc骨架8基于人igg4,并包括处于第一异源二聚体fc链上的l368d/k370s偏斜变体和q295e/n384d/q418e/n421d pi变体、处于第二异源二聚体fc链上的s364k/e357q偏斜变体,以及处于两条链上的消融了fab臂交换(如本领域所已知的)的s228p(根据eu编号,在kabat中为s241p)变体。异源二聚体fc骨架9基于人igg2,并包括处于第一异源二聚体 fc链上的l368d/k370s偏斜变体和q295e/n384d/q418e/n421d pi变体、处于第二异源二聚体fc链上的s364k/e357q偏斜变体。异源二聚体fc骨架10基于人igg2,并包括处于第一异源二聚体fc链上的l368d/k370s偏斜变体和q295e/n384d/q418e/n421d pi变体、处于第二异源二聚体fc链上的s364k/e357q偏斜变体,以及处于两条链上的s267k消融变体。异源二聚体fc骨架9和10的替代形式可以包括c220s和/或c219s(在基于igg2的骨架的情况下)。异源二聚体fc骨架11基于人igg1(356e/358m同种异型),并包括处于第一异源二聚体fc链上的l368d/k370s偏斜变体和q295e/n384d/q418e/n421d pi变体、处于第二异源二聚体fc链上的s364k/e357q偏斜变体,以及处于两条链上的 e233p/l234v/l235a/g236del/s267k消融变体、m428l/n434s xtend变体以及c220s。异源二聚体fc骨架12基于人igg1(356e/358m同种异型),并包括处于第一异源二聚体fc链上的l368d/k370s偏斜变体、处于第二异源二聚体fc链上的s364k/e357q偏斜变体和 p217r/p229r/n276k pi变体,以及处于两条链上的e233p/l234v/l235a/g236del/s267k消融变体和c220s。
[0158]
如本领域技术人员将认识到并在下文概述的,这些序列可以与需要异源二聚体fc区的任何il
‑
10融合体形式一起使用。应当注意,对于c端fc融合体形式(例如异源fc
‑
scil10),骨架还可在一条或两条链上包含缺失k447。
[0159]
这些序列也可以与需要异源二聚体fc区的本发明的任何靶向性il
‑
10融合体一起使用。在包括与fc共价连接的可变重结构域的靶向性il
‑
10融合体形式中,可变重结构域可通过相应的ch1结构域和部分铰链区与fc结构域共价连接,其例示性序列描绘于图12中,并
且可排除c220s和/或c219s取代。
[0160]
在这些骨架的每一个骨架内包括与所列举的序列90%、95%、98%和99%相同(如本文所定义)和/或含有1、2、3、4、5、6、7、8、9或10个额外的氨基酸取代的序列(与附图的“亲本”相比,如本领域技术人员将理解,与亲本人igg1(或igg2或igg4,这取决于骨架)相比,已经含有多个氨基酸修饰)。也就是说,除了此图的骨架中所含的偏斜变体、pi变体和消融变体之外或作为其替代方案,所列举的骨架可含有另外的氨基酸修饰(通常是氨基酸取代)。
[0161]
图12描绘例示性的ch1和部分铰链区,在靶向性il
‑
10融合体的特定形式的背景下,它们可用于将可变结构域与骨架共价连接,如图10和11中所描绘。
[0162]
图13描绘可用于本发明的开瓶器(bottle
‑
opener)和中心
‑
scfv双特异性抗体的同源轻链(即恒定轻链)的“非fv”骨架。
[0163]
图14描绘可用于本发明的il
‑
10融合蛋白的il
‑
10组分和配置的卡通示意图。图14a描绘包含成熟形式il
‑
10的残基1
‑
116的hil
‑
10(a
‑
d)结构域。图14b描绘包含成熟形式il
‑
10 的残基117
‑
160的hil
‑
10(e
‑
f)结构域。图14c描绘通过在第一il
‑
10单体的hil
‑
10(a
‑
d)与第二il
‑
10单体的hil
‑
10(e
‑
f)之间进行结构域交换而形成的生物活性il
‑
10同源二聚体。图14d描绘单链il
‑
10复合物或“scil
‑
10”,其包含任选地通过接头与第二il
‑
10单体共价连接的第一il
‑
10单体。图14e描绘josephson等人(2000)描述的单体il
‑
10或“il10m1”,其通过在il
‑
10单体中的hil
‑
10(a
‑
d)与hil
‑
10(e
‑
f)结构域之间工程化结构域接头而生成。图 14f描绘分裂il
‑
10,其包含与hil
‑
10(e
‑
f)结构域非共价连接的hil
‑
10(a
‑
d)结构域。
[0164]
图15描绘可用于本发明的il
‑
10融合蛋白的il
‑
10组分的序列和配置。图15a描绘包含成熟形式il
‑
10的残基1
‑
116的hil
‑
10(a
‑
d)结构域。图15b描绘包含成熟形式il
‑
10的残基 117
‑
160的hil
‑
10(e
‑
f)结构域。图15c描绘单链il
‑
10复合物或“scil
‑
10”,其包含与第二 il
‑
10单体共价连接的第一il
‑
10单体,但是应注意,scil
‑
10可以任选地在第一与第二il
‑
10 单体之间包括如本领域已知的接头。图15d描绘josephson等人(2000)描述的单体il
‑
10或“il10m1”,其通过在il
‑
10单体中的hil
‑
10(a
‑
d)与hil
‑
10(e
‑
f)结构域之间工程化结构域接头而生成,但是本领域技术人员应认识到,可以通过本领域已知的其他接头替代所述接头。应注意,虽然本文描绘的il
‑
10序列包含109h,但所述序列还可以包含109l,以及90%、 95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0165]
图16描绘il10
‑
fc类别的il
‑
10融合体的例示性形式。n端il
‑
10fc融合体或“(il10)2ꢀ‑
fc”(图16a)形式包括两个相同的单体,每个单体包含任选地通过结构域接头与同源二聚体 fc链的n端共价连接的il
‑
10单体(例如,xenp24628)。c端il
‑
10fc融合体或“fc
‑ꢀ
(il10)
2”(图16b)形式包括两个相同的单体,每个单体包含任选地通过结构域接头与同源二聚体fc链的c端共价连接的il
‑
10单体(例如,xenp24632)。“(il10
‑
nc
‑
il10)
‑
异源fc”(图 16c)形式包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接(任选地通过结构域接头)的第一il
‑
10单体;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链,同时第二il
‑
10单体单独转染,以形成非共价il
‑
10二聚体(例如,xenp25955)。“异源fc
‑
(il10
‑
nc
‑
il10)”(图16d)形式包含:第一单体,其包含与第一异源二
聚体fc链的 c端共价连接(任选地通过结构域接头)的第一il
‑
10单体;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链,同时第二il
‑
10单体单独转染,以形成结构域交换的il
‑
10二聚体。
[0166]
图17描绘(il10)2‑
fc形式的il10
‑
fc类别的例示性il
‑
10融合体的序列(例如,xenp24628、 xenp246289、xenp24630和xenp24631)。il
‑
10序列为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l,但il
‑
10融合体可以利用包含109h的il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0167]
图18描绘fc
‑
(il10)2形式的il10
‑
fc类别的例示性il
‑
10融合体的序列(例如,xenp24632、 xenp24633和xenp24634)。il
‑
10序列为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l,但il
‑
10融合体可以利用包含109h 的il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、 5、6、7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0168]
图19描绘(il10
‑
nc
‑
il10)
‑
异源fc形式的il10
‑
fc类别的例示性il
‑
10融合体的序列(例如,xenp25955)。il
‑
10序列为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l,但il
‑
10融合体可以利用包含109h的il
‑
10 序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、 8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/ 去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0169]
图20a
‑
图20b描绘scil10
‑
fc类别的il
‑
10融合体的例示性形式,其利用如图15中描绘的单链il
‑
10(“scil10”)复合物。n端单链il
‑
10fc融合体或“scil10
‑
异源fc”(图20a)形式包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接(任选地通过结构域接头) 的scil
‑
10;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链(例如,xenp25238)。c端单链il
‑
10fc融合体或“异源fc
‑
scil10”(图20b)形式包含:第一单体,其包含与第一异源二聚体fc链的c端共价连接(任选地通过结构域接头)的scil
‑
10;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链(例如, xenp28740)。以此类别的形式使用的scil
‑
10复合物包含任选地通过结构域接头与第二il
‑
10 单体共价连接的第一il
‑
10单体。
[0170]
图21a
‑
图21c描绘scil10
‑
异源fc形式的scil10
‑
fc类别的例示性il
‑
10融合体的序列 (例如,xenp25238、xenp25239、xenp25240、xenp25241、xenp25880和xenp28295)。il
‑
10序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文
描绘的il
‑
10序列包含109l(和269l,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109h(和269h,在scil
‑
10的背景中)的il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0171]
图22描绘异源fc
‑
scil10形式的scil10
‑
fc类别的例示性il
‑
10融合体的序列(例如, xenp28740)。il
‑
10序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l(和269l,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109h(和269h,在scil
‑
10的背景中)的il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0172]
图23a
‑
图23f描绘il10m1
‑
fc类别的il
‑
10融合体的例示性形式,其利用josephson等人(2000)描述的单体il
‑
10(il10m1),如图14和15中所描绘。n端二价il10m1 fc融合体或“(il10m1)2‑
fc”(图23a)形式包括两个相同的单体,每个单体包含任选地通过结构域接头与同源二聚体fc链的n端共价连接的il10m1(例如,xenp25326)。c端二价il10m1 fc融合体或“fc
‑
(il10m1)
2”(图23b)形式包括两个相同的单体,每个单体包含任选地通过结构域接头与同源二聚体fc链的c端共价连接的il10m1(例如,xenp25327)。单臂n端二价il10m1 fc融合体或“(il10m1)2‑
异源fc”(图23c)形式包含:第一单体,其包含与进一步共价连接至第一异源二聚体fc链的n端(任选地通过接头)的第二il10m1共价连接(任选地通过接头)的第一il10m1;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc 链。单臂c端二价il10m1 fc融合体或“异源fc
‑
(il10m1)
2”(图23d)形式包含:第一单体,其包含与进一步共价连接至第一异源二聚体fc链的c端(任选地通过接头)的第二il10m1共价连接(任选地通过接头)的第一il10m1;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链。n端一价il10m1融合体或“(il10m1)1‑
异源fc”(图23e)形式包含与第一异源二聚体fc链的n端共价连接(任选地通过接头)的il10m1和包含为“仅 fc”或“空fc”的互补的第二异源二聚体fc链的第二单体。c端一价il10m1融合体或“异源fc
‑
(il10m1)
1”(图23f)形式包含与第一异源二聚体fc链的c端共价连接(任选地通过接头)的il10m1和包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链的第二单体。在此类别的形式中使用的il10m1通过在il
‑
10单体的螺旋d与e之间工程化结构域接头而生成。
[0173]
图24描绘(il10m1)1‑
异源fc形式的il10m1
‑
fc类别的例示性il
‑
10融合体的序列(例如, xenp14246)。il
‑
10m1序列为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il10m1、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il10m1序列包含109h(根据不具有内部接头的成熟形式il
‑
10单体编号),但il
‑
10融合体可以利用包含109l的il10m1序列,以及90%、95%、98%和99%相同 (如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的il10m1 序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0174]
图25描绘异源fc
‑
(il10m1)1形式的il10m1
‑
fc类别的例示性il
‑
10融合体的序列(例如, xenp14247)。il
‑
10m1序列为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il10m1、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il10m1序列包含109h(根据不具有内部接头的成熟形式il
‑
10单体编号),但il
‑
10融合体可以利用包含109l的il10m1序列,以及90%、95%、98%和99%相同 (如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的il10m1 序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0175]
图26描绘(il10m1)2‑
fc形式的il10m1
‑
fc类别的例示性il
‑
10融合体的序列(例如, xenp25236)。il
‑
10m1序列为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il10m1、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il10m1序列包含109h(根据不具有内部接头的成熟形式il
‑
10单体编号),但il
‑
10融合体可以利用包含109l的il10m1序列,以及90%、95%、98%和99%相同 (如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的il10m1 序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0176]
图27描绘fc
‑
(il10m1)2形式的il10m1
‑
fc类别的例示性il
‑
10融合体的序列(例如, xenp25237)。il
‑
10m1序列为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il10m1、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il10m1序列包含109h(根据不具有内部接头的成熟形式il
‑
10单体编号),但il
‑
10融合体可以利用包含109l的il10m1序列,以及90%、95%、98%和99%相同 (如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的il10m1 序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0177]
图28描绘(il10m1)2‑
异源fc形式的il10m1
‑
fc类别的例示性il
‑
10融合体的序列(例如, xenp26887)。il
‑
10m1序列为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il10m1、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il10m1序列包含109h(根据不具有内部接头的成熟形式il
‑
10单体编号),但il
‑
10融合体可以利用包含109l的il10m1序列,以及90%、95%、98%和99%相同 (如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的il10m1 序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0178]
图29描绘分裂il10
‑
fc类别的il
‑
10融合体的例示性形式,其利用如图14a
‑
14f和15a
‑ꢀ
15d中所描绘的hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f)结构域。n端一价分裂il
‑
10fc融合体或“(分裂 il10)1‑
异源fc”(图29a)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接 (任选地通过接头)的hil
‑
10(a
‑
d)结构域;以及第二单体,其包含与互补的第二异源二聚体fc 链的n端共价连接(任选地通过接头)的hil
‑
10(e
‑
f)结构域(例如,xenp25242)。c端一价分裂il
‑
10fc融合体或“异源fc
‑
(分裂il10)
1”(图29b)包含:第一单体,其包含与第一异源二聚体fc链的c端共价连接(任选地通过接头)的hil
‑
10(a
‑
d)结构域;以及第二单
体,其包含与互补的第二异源二聚体fc链的c端共价连接(任选地通过接头)的hil
‑
10(e
‑
f)结构域。n 端二价分裂il
‑
10fc融合体或“(分裂il10)2‑
异源fc”(图29c)包含:第一单体,其包含与进一步共价连接(任选地通过接头)至第一异源二聚体fc链的n端的第二hil
‑
10(a
‑
d)结构域共价连接(任选地通过接头)的第一hil
‑
10(a
‑
d)结构域;以及第二单体,其包含与进一步共价连接(任选地通过接头)至互补的第二异源二聚体fc链的n端的第二hil
‑
10(e
‑
f)结构域共价连接(任选地通过接头)的第一hil
‑
10(e
‑
f)结构域。c端二价分裂il
‑
10fc融合体或“异源fc
‑ꢀ
(分裂il10)
2”(图29d)包含:第一单体,其包含与进一步共价连接(任选地通过接头)至第一异源二聚体fc链的c端的第二hil
‑
10(a
‑
d)结构域共价连接(任选地通过接头)的第一hil
‑
10 (a
‑
d)结构域;以及第二单体,其包含与进一步共价连接(任选地通过接头)至互补的第二异源二聚体fc链的c端的第二hil
‑
10(e
‑
f)结构域共价连接(任选地通过接头)的第一hil
‑
10(e
‑
f) 结构域的。
[0179]
图30描绘分裂il10
‑
异源fc形式的分裂il10
‑
fc类别的例示性il
‑
10融合体的序列(例如,xenp25242、xenp25243和xenp25244)。il
‑
10组分的序列为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10组分、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的hil
‑
10(a
‑
d)序列包含109l(根据成熟形式il
‑
10单体编号),但il
‑
10融合体可以利用包含109h的hil
‑
10(a
‑
d)序列。此外,il
‑
10融合体可以利用90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、 4、5、6、7、8、9或10个氨基酸取代的hil
‑
10(a
‑
d)和/或hil
‑
10(e
‑
f)序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0180]
图31描绘异源fc
‑
分裂il10形式的分裂il10
‑
fc类别的例示性il
‑
10融合体的序列(例如,xenp25879)。il
‑
10组分的序列为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10组分、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的hil
‑
10(a
‑
d)序列包含109l(根据成熟形式il
‑
10单体编号),但il
‑
10融合体可以利用包含109h的hil
‑
10(a
‑
d)序列。此外,il
‑
10融合体可以利用90%、 95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个氨基酸取代的hil
‑
10(a
‑
d)和/或hil
‑
10(e
‑
f)序列,所述氨基酸取代包括调节亲和力/效力、引入/ 去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0181]
图32a
‑
图32c描绘a)色谱图,其示出xenp25880的纯化部分2(在蛋白a色谱法之后的阳离子交换色谱法)以及如图32a中所描绘的从阳离子交换分离中分离出的峰b相对于峰 bc的纯度和同质性,如通过b)利用多角度光散射的分析型尺寸排阻色谱法(asec
‑
mals)和 c)分析型阳离子交换色谱法(aciex)所确定的。图32b还描绘了如通过多角度光散射确定的峰中的蛋白质物类的分子量。
[0182]
图33a
‑
图33e描绘在a)cd4
cd25
cd45ra
‑
t细胞、b)cd4
cd25
中
cd45ra
t细胞、c)cd8
cd25
cd45ra
‑
t细胞、d)cd8
cd25
中
cd45ra
t细胞以及e)tregs上通过 il10
‑
fc形式(xenp24628)、scil10
‑
fc形式(xenp25880)、il10m1
‑
fc形式(xenp25236)和分裂il10
‑
fc形式(xenp25242)的il
‑
10融合蛋白以及重组il
‑
10对照实现的stat3磷酸化的诱导。数据显示,scil0
‑
fc形式的il
‑
10融合体(xenp25880)在各种淋巴细胞群上诱导stat3磷酸化。
[0183]
图34a
‑
图34e描绘在a)cd4
cd45ra
‑
t细胞、b)cd4
cd45ra
t细胞、c) cd8
cd45ra
‑
t
细胞、d)cd8
cd45ra
t细胞以及e)tregs上通过具有n端il
‑
10 (xenp25981)和c端il
‑
10(xenp28741)的il
‑
10融合蛋白以及重组il
‑
10对照实现的stat3 磷酸化的诱导。数据显示,c端fc融合体xenp28741比n端fc融合体xenp25981更有效地诱导stat3磷酸化。
[0184]
图35a
‑
图35d描绘例示性il
‑
10单体变体的序列,所述变体经过设计以降低其对il
‑
10 受体复合物的亲和力和/或降低其效力。应注意,这些il
‑
10变体序列是基于il
‑
10(109l)序列,但是它们可以基于il
‑
10(109h)序列。本文中的每个取代可以单独使用或与本文描绘的任何其他取代组合使用。另外,如本领域技术人员将清楚的那样,虽然本文中的取代是在il
‑
10 单体的背景下描述的,但每个取代都可以在il
‑
10结构域(例如hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f))、 scil
‑
10和il10m1的背景下单独使用或组合使用。此外,此图中描绘的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0185]
图36a
‑
图36d描绘例示性il10m1变体的序列,所述变体经过设计以降低其对il
‑
10受体复合物的亲和力和/或降低其效力。应注意,这些il
‑
10变体序列是基于il
‑
10(109h)序列,但是它们可以基于il
‑
10(109l)序列。此图中描绘的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0186]
图37a
‑
图37e描绘(il10m1)1‑
异源fc形式的il10m1
‑
fc类别的例示性il
‑
10融合体的序列,其包含被工程化以降低亲和力和效力的il10m1变体。il
‑
10m1序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示 il10m1、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il10m1序列包含 109h(根据不具有内部接头的成熟形式il
‑
10单体编号),但il
‑
10融合体可以利用包含109l 的il10m1序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、 5、6、7、8、9或10个另外的氨基酸取代的il10m1序列,所述氨基酸取代包括调节亲和力 /效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0187]
图38描绘包含与人il
‑
10r1结合的亲和工程化的il10m1变体的(il10m1)1‑
异源fc融合体的解离常数(k
d
)、缔合速率(k
a
)和解离速率(k
d
),如通过octet(一种基于bli的方法)所确定的。将二价人il
‑
10
‑
fc捕获在ar2g生物传感器上并浸入多种浓度的指定测试制品中。*表示来自双相传感图的拟合不佳。数据显示,我们工程化了对il
‑
10r1具有一定范围的亲和力的il
‑
10变体。
[0188]
图39a
‑
图39g描绘例示性scil
‑
10变体的序列,所述变体经过设计以降低其对il
‑
10受体复合物的亲和力和/或降低其效力。应注意,这些il
‑
10变体序列是基于il
‑
10(109l)序列,但是它们也可以基于il
‑
10(109h)序列。此图中描绘的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或增强稳定性的取代。还应注意,这些scil
‑
10变体可以任选地在每个il
‑
10序列之间包括接头,但是取代的编号将基于如图15c所描绘的全长scil
‑
10序列。
[0189]
图40a
‑
图40n描绘scil10
‑
异源fc形式的scil10
‑
fc类别的例示性il
‑
10融合体的序列,其包含被工程化以降低亲和力和效力的scil
‑
10变体。il
‑
10序列加下划线,接头加双下划线 (但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示
il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l(和269l,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109h(和269h,在scil
‑
10的背景中)的 il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、 6、7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0190]
图41a
‑
图41f描绘异源fc
‑
scil10形式的scil10
‑
fc类别的例示性il
‑
10融合体的序列,其包含被工程化以降低亲和力和效力的scil
‑
10变体。il
‑
10序列加下划线,接头加双下划线 (但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l(和269l,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109h(和269h,在scil
‑
10的背景中)的 il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、 6、7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0191]
图42a
‑
图42e描绘在a)cd4
cd45ra
‑
t细胞、b)cd4
cd45ra
t细胞、c) cd8
cd45ra
‑
t细胞、d)cd8
cd45ra
t细胞以及e)tregs上通过包含亲和工程化的scil
‑ꢀ
10变体的scil10
‑
异源fc融合体实现的stat3磷酸化的诱导。数据说明我们工程化了具有广泛效力的il
‑
10融合体(如stat3磷酸化的诱导所指示)。
[0192]
图43a
‑
图43c描绘包含亲和工程化的scil
‑
10变体的scil10
‑
异源fc融合体对纯化的cd8
t细胞的a)ifnγ分泌,b)增殖(如通过表达ki67的细胞的百分比所指示)以及c)活化(如通过cd25表达所指示)的增强。数据显示,本发明的il
‑
10融合蛋白增强cd8
t细胞的 ifnγ分泌、cd8
t细胞的活化以及cd8
t细胞的增殖。
[0193]
图44a
‑
图44c描绘包含另外的亲和工程化的scil
‑
10变体的scil10
‑
异源fc融合体对纯化的cd8
t细胞的a)ifnγ分泌,b)增殖(如通过表达ki67的细胞的百分比所指示)以及c) 活化(如通过cd25表达所指示)的增强。数据显示,本发明的il
‑
10融合蛋白增强cd8
t细胞的ifnγ分泌、cd8
t细胞的活化以及cd8
t细胞的增殖。
[0194]
图45a
‑
图45c描绘cd8
t细胞的a)ifnγ分泌、b)增殖和c)活化的ec50与通过包含另外的亲和工程化的scil
‑
10变体的scil10
‑
异源fc融合体诱导stat3磷酸化的ec50之间的相关性。数据显示,stat3磷酸化的诱导的ec50与cd8
t细胞的ifnγ分泌、活化以及 cd8
t细胞的增殖的增强的ec50正相关。
[0195]
图46a
‑
图46c描绘a)色谱图,其示出xenp25238的纯化部分2(在蛋白a色谱法之后的阳离子交换色谱法),以及如图46a中所描绘的从阳离子交换分离中分离出的峰b相对于峰bc的纯度和同质性,如通过b)利用多角度光散射的分析型尺寸排阻色谱(asec
‑
mals)和 c)分析型阳离子交换色谱(aciex)所确定的。图46b还描绘了如通过多角度光散射确定的峰中的蛋白质物类的分子量。
[0196]
图47描绘在a)cd4
cd25
cd45ra
‑
t细胞、b)cd4
cd25
中
cd45ra
t细胞、c) cd8
cd25
cd45ra
‑
t细胞、d)cd8
cd25
中
cd45ra
t细胞,以及e)tregs上通过具有 scil
‑
10复合物(在异源二聚体fc链上具有较低pi(xenp25238)和在异源二聚体链上具有较高 pi(xenp25880))
的scil10
‑
异源fc融合蛋白实现的stat3磷酸化的诱导。
[0197]
图48描绘单链il
‑
10复合物(scil
‑
10)的例示性序列,其在第一il
‑
10单体与第二il
‑
10 单体之间包含结构域接头。il
‑
10序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体与接头之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l(和269l,在全长scil
‑
10的背景中),但 il
‑
10融合体可以利用包含109h(和269h,在scil
‑
10的背景中)的il
‑
10序列,以及90%、 95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0198]
图49a
‑
图49b描绘scil10
‑
异源fc形式的scil10
‑
fc类别的例示性il
‑
10融合体的序列,其包含在第一与第二il
‑
10单体之间具有结构域接头的scil
‑
10复合物。il
‑
10序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l(和269l,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109h(和269h,在 scil
‑
10的背景中)的il
‑
10序列。另外,虽然本文描绘的il
‑
10序列是野生型,但il
‑
10融合体可以利用90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、 8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/ 去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0199]
图50a
‑
图50e描绘在a)cd4
cd25
cd45ra
‑
t细胞、b)cd4
cd25
中
cd45ra
t细胞、c)cd8
cd25
cd45ra
‑
t细胞、d)cd8
cd25
中
cd45ra
t细胞,以及e)tregs上通过不具有接头(xenp25238)、具有工程化在scil
‑
10复合物内的g4s接头(xenp25239)、(g4s)2接头(xenp25240)和(g4s)3接头(xenp25241)的scil10
‑
异源fc融合蛋白实现的stat3磷酸化的诱导。
[0200]
图51a
‑
图51b描绘来自生产批次20180213的a)xenp25880和b)xenp25981的蛋白a纯化(纯化部分1)样品的分析型阳离子交换分离曲线。
[0201]
图52描绘各种生产批次的xenp25880和xenp25981的总蛋白质收率(在纯化部分1中通过蛋白a纯化的所有蛋白质;mg/l)、同源二聚体百分比和异源二聚体百分比(通过蛋白a 纯化材料的分析型阳离子交换分离确定),以及同源二聚体和异源二聚体收率(基于总蛋白质收率和同源二聚体或异源二聚体百分比计算)。数据显示xenp25981的产生通常导致scil10
‑ꢀ
异源fc融合体的收率高于空fc同源二聚体,表明n45d取代提高了异源二聚体收率。
[0202]
图53描绘xenp25986和xenp25993(具有n45d和n205d修饰的例示性变体scil10
‑ꢀ
异源fc融合体)的序列。il
‑
10序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l(和269l,在scil
‑
10的背景中),但 il
‑
10融合体可以利用包含109h(和269h,在scil
‑
10的背景中)的il
‑
10序列。另外,il
‑
10 融合体可以利用90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、 7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0203]
图54描绘来自生产批次20180213的xenp25880、xenp25981、xenp25986和 xenp25993的总蛋白质收率(在纯化部分1中通过蛋白a纯化的所有蛋白质;mg/l)、同源二聚体百分比和异源二聚体百分比(通过蛋白a纯化材料的分析型阳离子交换分离确定),以及同源二聚体和异源二聚体收率(基于总蛋白质收率和同源二聚体或异源二聚体百分比计算)。数据显示,包含具有n45d(和n205d)取代的il
‑
10变体的scil10
‑
异源fc融合体具有提高的异源二聚体收率。
[0204]
图55描绘例示性il
‑
10单体变体的序列,所述变体经过设计以去除脱酰胺的潜力。应注意,这些il
‑
10变体序列是基于il
‑
10(109l)序列,但是它们可以基于il
‑
10(109h)序列。本文的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。另外,如本领域技术人员将清楚的那样,虽然本文中的取代是在il
‑
10单体的背景下描述的,但每个取代都可以在il
‑
10结构域(例如hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f))、scil
‑
10和il10m1的背景下单独使用或组合使用。
[0205]
图56a
‑
图56c描绘例示性scil
‑
10变体的序列,所述变体经过设计以去除脱酰胺的潜力。应注意,这些il
‑
10变体序列是基于il
‑
10(109l)序列,但是它们也可以基于il
‑
10(109h)序列。还应注意,虽然这些il
‑
10变体序列包括n45d取代以提高收率,但它们可以在没有n45d 取代的情况下使用。此图中描绘的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。还应注意,这些scil
‑
10变体可以任选地在每个il
‑
10序列之间包括接头,但是取代的编号将基于如图15a
‑
15d所描绘的全长scil
‑
10序列。
[0206]
图57描绘例示性il
‑
10单体变体的序列,所述变体经过设计以去除天冬氨酸异构化的潜力。应注意,这些il
‑
10变体序列是基于il
‑
10(109l)序列,但是它们可以基于il
‑
10(109h) 序列。本文的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。另外,如本领域技术人员将清楚的那样,虽然本文中的取代是在il
‑
10单体的背景下描绘的,但每个取代都可以在il
‑
10结构域(例如hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f))、scil
‑
10和il10m1 的背景下单独使用或组合使用。
[0207]
图58描绘例示性scil
‑
10变体的序列,所述变体经过设计以去除天冬氨酸异构化的潜力。应注意,这些il
‑
10变体序列是基于il
‑
10(109l)序列,但是它们也可以基于il
‑
10(109h)序列。还应注意,虽然这些il
‑
10变体序列包括n45d取代以提高收率,但它们可以在没有n45d 取代的情况下使用。此图中描绘的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。还应注意,这些scil
‑
10变体可以任选地在每个il
‑
10序列之间包括接头,但是取代的编号将基于如图15a
‑
图15d所描绘的全长scil
‑
10序列。
[0208]
图59a
‑
图59h描绘scil10
‑
异源fc形式的scil10
‑
fc类别的例示性il
‑
10融合体的序列,其包含被工程化以去除降解相关ptm的潜力的scil
‑
10变体。il
‑
10序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑ꢀ
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l (和269l,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109h(和269h,在
scil
‑
10的背景中)的il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、 3、4、5、6、7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0209]
图60描绘异源fc
‑
scil10形式的scil10
‑
fc类别的例示性il
‑
10融合体的序列,其包含被工程化以去除降解相关ptm的潜力的scil
‑
10变体。il
‑
10序列加下划线,接头加双下划线 (但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l(和269l,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109h(和269h,在scil
‑
10的背景中)的 il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、 6、7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0210]
图61a
‑
图61c描绘a)xenp25981、b)xenp28641和c)xenp28635的蛋白a纯化(纯化部分1)样品的分析型阳离子交换分离曲线,其示出带电荷变体的去除。
[0211]
图62a
‑
图62e描绘在a)cd4
cd45ra
‑
t细胞、b)cd4
cd45ra
t细胞、c) cd8
cd45ra
‑
t细胞、d)cd8
cd45ra
t细胞以及e)tregs上通过包含被工程化以去除潜在降解位点的scil
‑
10变体的scil10
‑
异源fc融合体实现的stat3磷酸化的诱导。数据显示,被工程化以去除潜在降解位点的scil
‑
10变体不丧失其活性。
[0212]
图63a
‑
图63c描绘包含被工程化以去除潜在降解位点的scil
‑
10变体的scil10
‑
异源fc 融合体对纯化的cd8
t细胞的a)ifnγ分泌,b)增殖(如表达ki67的细胞的百分比所指示) 以及c)活化(如cd25表达所指示)的增强。数据显示,被工程化以去除潜在降解位点的scil
‑ꢀ
10变体不丧失其活性。
[0213]
图64描绘例示性il
‑
10单体变体的序列,所述变体经过设计以去除潜在的n
‑
糖基化位点。应注意,这些il
‑
10变体序列是基于il
‑
10(109l)序列,但是它们可以基于il
‑
10(109h) 序列。本文的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。另外,如本领域技术人员将清楚的那样,虽然本文中的取代是在il
‑
10单体的背景下描述的,但每个取代都可以在il
‑
10结构域(例如hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f))、scil
‑
10和il10m1 的背景下单独使用或组合使用。
[0214]
图65a
‑
图65b描绘例示性scil
‑
10变体的序列,所述变体经过设计以去除潜在的n
‑
糖基化位点。应注意,这些il
‑
10变体序列是基于il
‑
10(109l)序列,但是它们也可以基于il
‑
10 (109h)序列。还应注意,虽然这些il
‑
10变体序列包括n45d取代以提高收率,但它们可以在没有n45d取代的情况下使用。此图中描绘的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。还应注意,这些scil
‑
10变体可以任选地在每个il
‑
10序列之间包括接头,但是取代的编号将基于如图15c所描绘的全长scil
‑
10序列。
[0215]
图66a
‑
图66d描绘scil10
‑
异源fc形式的scil10
‑
fc类别的例示性il
‑
10融合体的序列,其包含被工程化以去除潜在n
‑
糖基化位点的scil
‑
10变体。il
‑
10序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指
示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l(和 269l,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109h(和269h,在scil
‑
10的背景中)的il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、 4、5、6、7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力 /效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0216]
图67描绘例示性il
‑
10单体变体的序列,所述变体经过设计以去除二硫桥。应注意,这些il
‑
10变体序列是基于il
‑
10(109l)序列,但是它们可以基于il
‑
10(109h)序列。本文的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。另外,如本领域技术人员将清楚的那样,虽然本文中的取代是在il
‑
10单体的背景下描述的,但每个取代都可以在il
‑
10结构域(例如hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f))、scil
‑
10和il10m1的背景下单独使用或组合使用。
[0217]
图68描绘例示性scil
‑
10变体的序列,所述变体经过设计以去除二硫桥。应注意,这些 il
‑
10变体序列是基于il
‑
10(109l)序列,但是它们也可以基于il
‑
10(109h)序列。还应注意,虽然这些il
‑
10变体序列包括n45d取代以提高收率,但它们可以在没有n45d取代的情况下使用。还应注意,虽然这些il
‑
10变体序列包括n320a取代以降低脱酰胺作用的潜力,但它们可以在没有n320a取代的情况下使用。此图中描绘的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。还应注意,这些scil
‑
10变体可以任选地在每个il
‑
10序列之间包括接头,但是取代的编号将基于如图15c所描绘的全长scil
‑
10序列。
[0218]
图69描绘scil10
‑
异源fc形式的scil10
‑
fc类别的例示性il
‑
10融合体的序列,其包含被工程化以去除二硫桥的scil
‑
10变体。il
‑
10序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l(和269l,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109h(和269h,在scil
‑
10的背景中)的il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或 10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0219]
图70a
‑
图70b描绘例示性il10m1变体的序列,所述变体经过设计以引入一个或多个另外的二硫桥或去除一个或多个二硫桥。应注意,这些il
‑
10变体序列是基于il
‑
10(109h)序列,但是它们也可以基于il
‑
10(109l)序列。此图中描绘的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、通过收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。另外,如本领域技术人员将清楚的那样,虽然本文中的取代是在il10m1的背景下设计的,但每个取代都可以在il
‑
10单体、il
‑
10结构域(例如hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f))以及scil
‑
10的背景下使用。
[0220]
图71a
‑
图71e描绘(il10m1)1
‑
异源fc形式的il10m1
‑
fc类别的例示性il
‑
10融合体的序列,其包含经过设计以引入一个或多个另外的二硫桥或去除一个或多个二硫桥的
il10m1 变体。il
‑
10序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109h(和269l,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109l(和269h,在scil
‑
10的背景中)的il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的il
‑ꢀ
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0221]
图72a
‑
图72aq描绘il10m1核心残基处的计算机模拟点突变对于其对il10m1的亲和力及其稳定性的影响。wt表示野生型残基,pos表示在il
‑
10中的位置(根据人il
‑
10成熟形式序列编号),并且mut表示取代。此图中描绘的预测为工程化具有改善稳定性的il10m1变体提供背景。
[0222]
图73描绘例示性il10m1变体的序列,所述变体经过设计以改善稳定性。应注意,这些 il
‑
10变体序列是基于il
‑
10(109h)序列,但是它们也可以基于il
‑
10(109l)序列。此图中描绘的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。另外,如本领域技术人员将清楚的那样,虽然本文中的取代是在il10m1的背景下设计的,但每个取代都可以在il
‑
10单体、il
‑
10结构域(例如hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f))以及scil
‑
10的背景下使用。
[0223]
图74a
‑
图74b描绘(il10m1)1‑
异源fc形式的il10m1
‑
fc类别的例示性il
‑
10融合体的序列,其包含被工程化以便改善稳定性的il10m1变体。il
‑
10m1序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il10m1、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il10m1序列包含109h(根据不具有内部接头的成熟形式il
‑
10单体编号),但il
‑
10融合体可以利用包含109l的il10m1 序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、 8、9或10个另外的氨基酸取代的il10m1序列。
[0224]
图75a
‑
图75b描绘例示性il10m1变体的序列,所述变体经过设计在螺旋d与e之间具有替代接头,以增加刚性或增加柔性。应注意,这些il
‑
10变体序列是基于il
‑
10(109h)序列,但是它们可以基于il
‑
10(109l)序列。此图中描绘的il10m1序列中的每一个可以包括本文描绘的一个或多个取代,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0225]
图76a
‑
图76d描绘(il10m1)1‑
异源fc形式的il10m1
‑
fc类别的例示性il
‑
10融合体的序列,其包含被工程化为在螺旋d与e之间具有替代接头,以增加刚性或增加柔性的il10m1 变体。il
‑
10m1序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il10m1、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il10m1序列包含109h(根据不具有内部接头的成熟形式il
‑
10单体编号),但il
‑
10融合体可以利用包含109l的il10m1序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的il10m1序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0226]
图77a
‑
图77b描绘在il
‑
10(109h)序列的背景中工程化的优选il
‑
10变体的序列。此图中描绘的每个取代可以单独使用或与本文描绘的任何其他取代组合使用,包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0227]
图78a
‑
图78g描绘scil10
‑
异源fc形式的scil10
‑
fc类别的il
‑
10融合体的例示性序列,其包含在il
‑
10il
‑
10(109h)序列的背景中工程化的优选il
‑
10变体。il
‑
10序列为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑ꢀ
10组分、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109h (和269h,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109l(和269l,在scil
‑
10的背景中)的il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、 3、4、5、6、7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0228]
图79a
‑
图79g描绘异源fc
‑
scil10形式的scil10
‑
fc类别的il
‑
10融合体的例示性序列,其包含在il
‑
10il
‑
10(109h)序列的背景中工程化的优选il
‑
10变体。il
‑
10序列为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑ꢀ
10组分、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109h (和269h,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109l(和269l,在scil
‑
10的背景中)的il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、 3、4、5、6、7、8、9或10个另外的氨基酸取代的il
‑
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0229]
图80a
‑
图80j描绘在cd4
cd45ra
t细胞上通过a)基于109h变体的例示性il
‑
10融合体;b)xenp27220相对于xenp30004;c)xenp27226相对于xenp30008;d)xenp27991 相对于xenp30009;e)xenp27990相对于xenp30010;f)xenp27993相对于xenp30011; g)xenp27222相对于xenp30012;h)xenp25986相对于xenp30013;i)xenp25993相对于xenp30014;以及j)xenp25981相对于xenp30005实现的stat3磷酸化的诱导。
[0230]
图81a
‑
图81j描绘在cd4
cd45ra
‑
t细胞上通过a)基于109h变体的例示性il
‑
10融合体;b)xenp27220相对于xenp30004;c)xenp27226相对于xenp30008;d)xenp27991 相对于xenp30009;e)xenp27990相对于xenp30010;f)xenp27993相对于xenp30011;g)xenp27222相对于xenp30012;h)xenp25986相对于xenp30013;i)xenp25993相对于xenp30014;以及j)xenp25981相对于xenp30005实现的stat3磷酸化的诱导。
[0231]
图82a
‑
图82j描绘在cd8
cd45ra
t细胞上通过a)基于109h变体的例示性il
‑
10融合体;b)xenp27220相对于xenp30004;c)xenp27226相对于xenp30008;d)xenp27991 相对于xenp30009;e)xenp27990相对于xenp30010;f)xenp27993相对于xenp30011; g)xenp27222相对于xenp30012;h)xenp25986相对于xenp30013;i)xenp25993相对于xenp30014;以及j)xenp25981相对于xenp30005实现的stat3磷酸化的诱导。
[0232]
图83a
‑
图83j描绘在cd8
cd45ra
‑
t细胞上通过a)基于109h变体的例示性il
‑
10融合体;b)xenp27220相对于xenp30004;c)xenp27226相对于xenp30008;d)xenp27991 相对于
xenp30009;e)xenp27990相对于xenp30010;f)xenp27993相对于xenp30011; g)xenp27222相对于xenp30012;h)xenp25986相对于xenp30013;i)xenp25993相对于xenp30014;以及j)xenp25981相对于xenp30005实现的stat3磷酸化的诱导。
[0233]
图84
‑
图84j描绘在tregs上通过a)基于109h变体的例示性il
‑
10融合体;b)xenp27220 相对于xenp30004;c)xenp27226相对于xenp30008;d)xenp27991相对于xenp30009; e)xenp27990相对于xenp30010;f)xenp27993相对于xenp30011;g)xenp27222相对于xenp30012;h)xenp25986相对于xenp30013;i)xenp25993相对于xenp30014;以及j)xenp25981相对于xenp30005实现的stat3磷酸化的诱导。
[0234]
图85a
‑
图85j描绘在单核细胞上通过a)基于109h变体的例示性il
‑
10融合体;b) xenp27220相对于xenp30004;c)xenp27226相对于xenp30008;d)xenp27991相对于 xenp30009;e)xenp27990相对于xenp30010;f)xenp27993相对于xenp30011;g) xenp27222相对于xenp30012;h)xenp25986相对于xenp30013;i)xenp25993相对于 xenp30014;以及j)xenp25981相对于xenp30005实现的stat3磷酸化的诱导。
[0235]
图86a
‑
图86d描绘被工程化具有xtend fc(m428l/n434s)的例示性il
‑
10融合蛋白的序列。il
‑
10序列加下划线,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10单体、接头和fc区之间的一个或多个边界。应注意,虽然本文描绘的il
‑
10序列包含109l(和269l,在scil
‑
10的背景中),但il
‑
10融合体可以利用包含109h(和269h,在scil
‑
10的背景中)的il
‑
10序列,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的il
‑ꢀ
10序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0236]
图87描绘基于il10
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性 il10
‑
fc”类别。“mab
‑
il10”形式(图87a)包括两个相同的单体,每个单体包含与同源二聚体fc链的n端共价连接的抗原结合结构域,所述同源二聚体fc链通过c端任选地通过接头与il
‑
10单体共价连接。“il10
‑
mab”形式(图87b)包括两个相同的单体,每个单体包含任选地通过接头与抗原结合结构域共价连接的il
‑
10单体,所述抗原结合结构域与同源二聚体fc 链的n端共价连接。“mab
‑
中心
‑
il10”形式(图87c)包括两个相同的单体,每个单体包含与 il
‑
10单体共价连接的抗原结合结构域,所述il
‑
10单体任选地通过接头与同源二聚体fc链共价连接。应注意,虽然抗原结合结构域被描述为fab,但抗原结合结构域可以是如本文所定义的任何抗原结合分子,诸如scfv。“抗x x il10
‑
异源fc”形式(图87d)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头)的第一il
‑
10单体,并且第二il
‑
10单体单独转染,以与第一il
‑
10单体形成非共价相互作用。
[0237]
图88a
‑
图88e描绘基于scil10
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性scil10
‑
fc”类别。“抗x x scil10
‑
异源fc”形式(图88a)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头)的scil
‑
10。在一个方面,“抗x x 异源fc
‑
scil10”形式(图88b)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含通过c端与scil
‑
10共价连接(任选地通过结构
域接头)的第二异源二聚体fc链。在另一方面,“抗x x异源fc
‑
scil10”形式包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域,所述第一异源二聚体fc链通过c端与scil
‑
10共价连接(任选地通过结构域接头);以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链。“(抗x)2‑
异源fc
‑
scil10”形式(图 88c)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的抗原结合结构域,所述第二异源二聚体fc链通过c端与scil
‑
10共价连接(任选地通过结构域接头)。“(抗x)2‑
中心
‑ꢀ
scil10”形式(图88d)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与scil
‑
10共价连接的抗原结合结构域,所述scil
‑
10 与第二异源二聚体fc链的n端共价连接(任选地通过接头)。“scil10
‑
(抗x)2‑
异源fc”形式 (图88e)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含任选地通过接头与抗原结合结构域共价连接的scil
‑
10,所述抗原结合结构域与第二异源二聚体fc链的n端共价连接(任选地通过接头)。应注意,虽然抗原结合结构域被描绘为fab,但抗原结合结构域可以是如本文所定义的任何抗原结合分子,诸如scfv。
[0238]
图89a
‑
图89m描绘基于il10m1
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性il10m1
‑
fc”类别。“抗x x(il10m1)1‑
异源fc”形式(图89a)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头)的il10m1。在一个方面,“抗x x异源fc
‑
(il10m1)
1”形式(图89b)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含通过c端与il10m1共价连接(任选地通过结构域接头)的第二异源二聚体fc链。在另一方面,“抗x x异源fc
‑
(il10m1)
1”形式包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域,所述第一异源二聚体fc链通过c端与il10m1共价连接(任选地通过结构域接头);以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链。“(抗x)2‑
异源fc
‑ꢀ
(il10m1)
1”形式(图89c)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的抗原结合结构域,所述第二异源二聚体fc链通过c端与il10m1共价连接(任选地通过结构域接头)。“(il10m1)1‑
(抗x)2‑
异源fc”形式(图89d)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含任选地通过接头与抗原结合结构域共价连接的il10m1,所述抗原结合结构域与第二异源二聚体fc链的n端共价连接(任选地通过接头)。“(抗x)2‑
中心
‑
(il10m1)
1”形式(图89e)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与il10m1共价连接的抗原结合结构域,所述il10m1与第二异源二聚体fc链的n端共价连接(任选地通过接头)。“抗x x(il10m1)2‑
异源fc”形式(图89f)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含任选地通过结构域接头与第二 il10m1共价连接的第一il10m1,所述第二il10m1与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头)。在一个方面,“抗x x异源fc
‑
(il10m1)
2”形式(图89g)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含通过c端与第一il10m1共价连接(任选地通过结构域接头)的第二异源二聚体fc 链,所述第一il10m1任选地通过结构域接头与
第二il10m1共价连接。在另一方面,“抗xx异源fc
‑
(il10m1)
2”形式包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域,所述第一异源二聚体fc链通过c端与第一il10m1共价连接(任选地通过结构域接头),所述第一il10m1任选地通过结构域接头与第二il10m1共价连接;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链。“(抗x)2‑
异源fc
‑ꢀ
(il10m1)
2”形式(图89h)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的抗原结合结构域,所述第二异源二聚体fc链通过c端与第一il10m1共价连接(任选地通过结构域接头),所述第一il10m1任选地通过结构域接头与第二il10m1共价连接。“(il10m1)2‑
(抗 x)2‑
异源fc”形式(图89i)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含任选地通过接头与第二il10m1共价连接的第一 il10m1,所述第二il10m1与抗原结合结构域共价连接(任选地通过结构域接头),所述抗原结合结构域与第二异源二聚体fc链的n端共价连接(任选地通过接头)。“(抗x)2‑
中心
‑ꢀ
(il10m1)
2”形式(图89j)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第一il10m1共价连接的抗原结合结构域,所述第一il10m1任选地通过结构域接头与第二il10m1共价连接,所述第二il10m1与第二异源二聚体fc链的n端共价连接(任选地通过接头)。“mab
‑
(il10m1)
2”形式(图89k)包含两个相同的单体,每个单体包含与同源二聚体fc链的n端共价连接的抗原结合结构域,所述同源二聚体fc链通过c端任选地通过接头与il10m1共价连接。“(il10m1)2‑
mab”形式(图 89l)包含两个相同的单体,每个单体包含任选地通过接头与抗原结合结构域共价连接的 il10m1,所述抗原结合结构域与同源二聚体fc链的n端共价连接。“mab
‑
中心
‑
(il10m1)
2”形式(图89m)包含两个相同的单体,每个单体包含与il10m1共价连接的抗原结合结构域,所述il10m1任选地通过接头与同源二聚体fc链共价连接。应注意,虽然抗原结合结构域被描绘为fab,但抗原结合结构域可以是如本文所定义的任何抗原结合分子,诸如scfv。
[0239]
图90描绘基于分裂il10
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性分裂il10
‑
fc”类别。在一个方面,“抗x x(分裂il10)1‑
异源fc”形式(图90a)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头)的hil
‑
10(a
‑
d)结构域,以及单独转染以使其与hil
‑
10(a
‑
d)结构域非共价相互作用的hil
‑
10(e
‑
f)结构域。在另一方面,“抗x x(分裂il10)1‑
异源fc”形式包含:第一单体,其包含与第一异源二聚体fc 链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n 端共价连接(任选地通过结构域接头)的hil
‑
10(e
‑
f)结构域,以及单独转染以使其与hil
‑
10(e
‑ꢀ
f)结构域非共价相互作用的hil
‑
10(a
‑
d)结构域。“(抗x)2x异源fc
‑
(分裂il10)
1”形式(图90b)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域,所述第一异源二聚体fc链通过c端与hil
‑
10(a
‑
d)结构域共价连接;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的抗原结合结构域,所述第二异源二聚体fc链通过c 端与hil
‑
10(e
‑
f)结构域共价连接。“(分裂il10)1‑
(抗x)2‑
异源fc”形式(图90c)包含:第一单体,其包含与抗原结合结构域共价连接的hil
‑
10(a
‑
d)结构域,所述抗原结合结构域与第一异源二聚体fc链的n端共价连接;以及第二单体,其包含与抗原结合结构域共价连接的
cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人员将理解的,v
h
和v
l
结构域可以被格式化为fab或scfv,以用于本发明的靶向cd8的il
‑
10融合蛋白。
[0242]
图93a
‑
图93c描绘例示性的抗nkg2d抗体的v
h
和v
l
结构域序列。cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人员将理解的,v
h
和 v
l
结构域可以被格式化为fab或scfv,以用于本发明的靶向nkg2d的il
‑
10融合蛋白。
[0243]
图94描绘“抗x x(il10m1)1‑
异源fc”形式的例示性的靶向cd8的il
‑
10融合体的序列。il
‑
10组分为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10组分、可变区和恒定/fc区之间的一个或多个边界。cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法 (如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr 且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人员将理解的,v
h
和v
l
结构域可以被格式化为fab或scfv,以用于本发明的靶向cd8的il
‑
10融合蛋白。
[0244]
图95描绘“mab
‑
(il10m1)
2”形式的例示性的靶向cd8的il
‑
10融合体的序列。il
‑
10 组分为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10组分、可变区和恒定/fc区之间的一个或多个边界。cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人员将理解的,v
h
和 v
l
结构域可以被格式化为fab或scfv,以用于本发明的靶向cd8的il
‑
10融合蛋白。
[0245]
图96a
‑
图96c描绘“抗x x scil10
‑
异源fc”形式的例示性的靶向cd8的il
‑
10融合体的序列。il
‑
10组分为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10组分、可变区和恒定/fc区之间的一个或多个边界。 cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人员将理解的,v
h
和v
l
结构域可以被格式化为fab或scfv,以用于本发明的靶向cd8的il
‑ꢀ
10融合蛋白。应注意,虽然本文描绘的il
‑
10序列包含109l,但所述序列还可以包含109h,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或 10个另外的氨基酸取代的序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0246]
图97a
‑
图97c描绘“抗x x scil10
‑
异源fc”形式的例示性的靶向nkg2d的il
‑
10融合体的序列。il
‑
10组分为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10组分、可变区和恒定/fc区之间的一个或多个边界。 cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人
员将理解的,v
h
和v
l
结构域可以被格式化为fab或scfv,以用于本发明的靶向nkg2d的 il
‑
10融合蛋白。应注意,虽然本文描绘的il
‑
10序列包含109l,但所述序列还可以包含109h,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0247]
图98描绘包含例示性的il
‑
10效力变体的“抗x x scil10
‑
异源fc”形式的例示性的靶向cd8的il
‑
10融合体的序列。il
‑
10组分为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10组分、可变区和恒定/fc区之间的一个或多个边界。cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人员将理解的,v
h
和v
l
结构域可以被格式化为fab或scfv,以用于本发明的靶向cd8的il
‑
10融合蛋白。应注意,虽然本文描绘的il
‑
10序列包含109l,但所述序列还可以包含109h,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、 5、6、7、8、9或10个另外的氨基酸取代的序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0248]
图99a
‑
图99e描绘在a)cd4
cd45ra
‑
t细胞、b)cd4
cd45ra
t细胞、c) cd8
cd45ra
‑
t细胞、d)cd8
cd45ra
t细胞以及e)tregs上通过靶向cd8和nkg2d的 il
‑
10融合体实现的stat3磷酸化的诱导。数据显示,靶向cd8的il
‑
10融合体xenp25794 比非靶向性il
‑
10融合体(xenp25880)远远更有效地在cd8
t细胞上诱导stat3磷酸化。此外,与cd4
t细胞相比,靶向nkg2d的il
‑
10融合体(xenp25952)还显示出针对cd8
t细胞的选择性。
[0249]
图100a
‑
图100g描绘例示性的人源化抗pd
‑
1抗体的v
h
和v
l
结构域序列。cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr 且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人员将理解的,v
h
和v
l
结构域可以被格式化为fab或scfv,以用于本发明的靶向pd
‑
1的il
‑
10融合蛋白。
[0250]
图101描绘“抗x x scil10
‑
异源fc”形式的例示性的靶向pd
‑
1的il
‑
10融合体的序列。 il
‑
10组分为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10组分、可变区和恒定/fc区之间的一个或多个边界。cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr 且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人员将理解的,v
h
和v
l
结构域可以被格式化为fab或scfv,以用于本发明的靶向pd
‑
1的il
‑
10融合蛋白。应注意,虽然本文描绘的il
‑
10序列包含109l,但所述序列还可以包含109h,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0251]
图102a
‑
图102c描绘包含例示性的il
‑
10效力变体的“抗x x scil10
‑
异源fc”形式的例示性的靶向pd
‑
1的il
‑
10融合体的序列。il
‑
10组分为斜体,接头加双下划线(但是如本
领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10组分、可变区和恒定/fc区之间的一个或多个边界。cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人员将理解的,v
h
和v
l
结构域可以被格式化为fab或scfv,以用于本发明的靶向pd
‑
1的il
‑
10融合蛋白。应注意,虽然本文描绘的il
‑
10序列包含109l,但所述序列还可以包含109h,以及90%、95%、98%和99%相同(如本文所定义),和/或含有 1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0252]
图103描绘包含例示性的il
‑
10效力变体的“抗x x异源fc
‑
scil10”形式的例示性的靶向pd
‑
1的il
‑
10融合体的序列。il
‑
10组分为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10组分、可变区和恒定/fc区之间的一个或多个边界。cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人员将理解的,v
h
和v
l
结构域可以被格式化为fab或scfv,以用于本发明的靶向pd
‑
1的il
‑
10融合蛋白。应注意,虽然本文描绘的il
‑
10序列包含109l,但所述序列还可以包含109h,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代的序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0253]
图104a
‑
图104b描绘非竞争性的抗pd
‑
1mab a、mab b和mab c的例示性人源化可变区的序列。cdr为粗体,并且斜线表示可变结构域的边界。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人员将理解的,v
h
和v
l
结构域可以被格式化为fab 或scfv,以用于本发明的靶向pd
‑
1的il
‑
10融合蛋白。
[0254]
图105描绘基于纳武单抗的二价抗pd
‑
1mab、内部制备的派姆单抗、嵌合mab a、嵌合mab b和嵌合mab c的表位分箱,如通过归一化的bli响应octet所指示。大于0.5的归一化bli响应表示抗体对不会分箱到同一表位。
[0255]
图106a
‑
图106c描绘包含例示性的il
‑
10效力变体的例示性的非竞争性的靶向pd
‑
1的 il
‑
10融合体的序列。il
‑
10组分为斜体,接头加双下划线(但是如本领域技术人员将了解的,接头可以被其它接头替代),并且斜线(/)指示il
‑
10组分、可变区和恒定/fc区之间的一个或多个边界。cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。如本领域技术人员将理解的,v
h
和v
l
结构域可以被格式化为fab或scfv,以用于本发明的靶向pd
‑ꢀ
1的il
‑
10融合蛋白。应注意,虽然本文描绘的il
‑
10序列包含109l,但所述序列还可以包含 109h,以及90%、95%、98%和99%相同(如本文所定义),和/或含有1、2、3、4、5、6、7、 8、9或10个另外的氨基酸取代的序列,所述氨基酸取代包括调节亲和力/效力、引入/去除二硫桥、
提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的取代。
[0256]
图107a
‑
图107e描绘在a)cd4
cd45ra
‑
cd25
t细胞、b)cd4
cd45ra
cd25
t细胞、c)cd8
cd45ra
低
cd25
高
t细胞、d)cd8
cd45ra
高
cd25
中
t细胞以及e)tregs上通过靶向pd
‑
1的il
‑
10融合体实现的stat3磷酸化的诱导。数据显示,靶向pd
‑
1的il
‑
10融合体比非靶向性il
‑
10融合体(xenp25880)远远更有效地在各种t细胞群上诱导stat3磷酸化。
[0257]
图108a
‑
图108c描绘靶向pd
‑
1的il
‑
10融合体对纯化的cd8
t细胞的a)ifnγ分泌, b)增殖(如表达ki67的细胞的百分比所指示)以及c)活化(如cd25表达所指示)的增强。
[0258]
图109描绘非靶向性il
‑
10融合蛋白(xenp30007、xenp30008和xenp30010)和具有不同scil
‑
10效力变体的靶向pd
‑
1的il
‑
10融合蛋白(xenp30520、xenp30521和xenp30522) 对cd8
t细胞增殖的诱导(如总cd8
t细胞群中cd8
ki67 的百分比所指示)。数据显示,对于各效力降低的il
‑
10
‑
fc融合体,通过pd
‑
1靶向恢复了诱导cd8
t细胞的效力。
[0259]
图110描绘在cd8
pd
‑1
t细胞上通过具有基于mab c_h1_l1.1的pd
‑
1靶向臂 (xenp30521、xenp31266、xenp31268和xenp31267)、基于1c11的pd
‑
1靶向臂 (xenp31270、xenp31269、xenp31272和xenp31271)或基于numax/莫维珠单抗(motavizumab)的rsv靶向臂(xenp31274、xenp31273、xenp31276和xenp31275)的靶向性il
‑
10融合蛋白实现的stat3磷酸化的诱导。数据显示,靶向pd
‑
1的il
‑
10融合蛋白比相应的靶向rsv的il
‑
10融合蛋白(具有相同的il
‑
10变体和相同的末端连接)更有效。对于效力较弱的il
‑
10融合蛋白(具有il10.105),scil
‑
10部分的c端连接导致效力降低;对于效力较强的il
‑
10融合蛋白,scil
‑
10部分的c端连接导致效力增强。
[0260]
图111描绘人和食蟹猴的tigit的序列,以促进与两者结合的抗原结合结构域的开发,以便于临床开发。
[0261]
图112描绘具有消融变体(e233p/l234v/l235a/g236del/s267k,“igg1_pva_/s267k”) 的噬菌体来源的抗tigit抗体的序列。cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。
[0262]
图113描绘xenp19351的序列,其为基于10a7的二价抗tigit mab,所述10a7如 wo 2015/009856中所描述,是一种已知的tigit/tigit配体相互作用阻断剂。cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表 1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。
[0263]
图114描绘xenp19352的序列,其为基于1f4的二价抗tigit mab,所述1f4如wo 2015/009856中所描述,是一种已知的tigit/tigit配体相互作用的非阻断剂。cdr加有下划线。如本文所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表 1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包括带下划线的cdr且还包括使用其它编号系统的v
h
和v
l
结构域内所包括的cdr。
[0264]
图115描绘含有克隆2a5b4的例示性板的每个孔中的单细胞门(single cell gate)内的平均 mfi(指示噬菌体克隆与jurkat
‑
tigit细胞的结合)。
[0265]
图116。单细胞门内的jurkat
‑
tigit细胞以单参数直方图表示(与含有克隆2a5b4
对cd8
t细胞增殖的诱导(如总cd8
t细胞群中cd8
ki67 的百分比所指示)。数据显示,对于各效力降低的il
‑
10
‑
fc融合体,通过tigit靶向恢复了诱导cd8
t细胞的效力。
具体实施方式
[0274]
i.定义
[0275]
为了可以更全面地理解本技术,下面阐述几个定义。此类定义意在涵盖语法等同物。
[0276]
本文中的“消融”意指结合和/或活性的降低或去除。因此,举例来说,“消融fcγr结合”意指初始结合比不含有特定变异的fc区小50%的fc区氨基酸变异,其中优选小于70
‑ꢀ
80
‑
90
‑
95
‑
98%的结合损失,并且一般来说,结合低于在biacore分析中可检测到的结合水平。在fcγr结合的消融中特别有用的是图7所示的那些。然而,除非另外指出,否则本发明的fc 单体保持与fcrn的结合。
[0277]
如本文所使用的,“adcc”或“抗体依赖性细胞介导的细胞毒性”意指其中表达fcγr 的非特异性细胞毒性细胞识别靶细胞上的结合抗体并且随后引起靶细胞溶解的细胞介导反应。adcc与结合fcγriiia相关;与fcγriiia结合的增加引起adcc活性的增加。如本文所讨论的,本发明的许多实施方案完全消融adcc活性。
[0278]
本文中的“修饰”意指多肽序列中的氨基酸取代、插入和/或缺失或对与蛋白质化学连接的部分的更改。例如,修饰可以是与蛋白质连接的改变的碳水化合物或peg结构。本文中通过“氨基酸修饰”意指多肽序列中的氨基酸取代、插入和/或缺失。为清楚起见,除非另外说明,否则氨基酸修饰总是针对由dna编码的氨基酸,例如具有dna和rna形式的密码子的20种氨基酸。
[0279]
本文中通过“氨基酸取代”或“取代”意指用不同的氨基酸替换亲本多肽序列中特定位置的氨基酸。特别地,在一些实施方案中,取代是针对并非天然存在于特定位置、并非天然存在于生物体内或任何生物体内的氨基酸。举例来说,取代e272y或272y是指在位置272 处的谷氨酸被酪氨酸替换的变体多肽,在这种情况下是fc变体。为清楚起见,已经被工程化以改变核酸编码序列但不改变起始氨基酸(例如,将cgg(编码精氨酸)换成cga(仍然编码精氨酸)以增加宿主生物体表达水平)的蛋白质不是“氨基酸取代”;即,尽管产生了编码相同蛋白质的新基因,但是如果蛋白质在其开始的特定位置处具有相同氨基酸,则所述蛋白质不是氨基酸取代。
[0280]
如本文所使用的,“氨基酸插入”或“插入”意指在亲本多肽序列中的特定位置处添加氨基酸残基或序列。举例来说,
‑
233e表示在位置233之后和位置234之前插入谷氨酸。另外,
‑
233ade或a233ade表示在位置233之后和位置234之前插入alaaspglu。
[0281]
如本文所使用,“氨基酸缺失”或“缺失”意指在亲本多肽序列的特定位置处去除氨基酸残基或序列。例如,e233
‑
、e233#、e233()或e233del指代位置233处的谷氨酸的缺失。另外,eda233
‑
或eda233#指代开始于位置233的序列gluaspala的缺失。
[0282]
如本文所使用的,“变体蛋白质”、“蛋白质变体”或“变体”意指由于至少一个修饰而与亲本蛋白质不同的蛋白质。蛋白质变体可以指代蛋白质本身、包含所述蛋白质的组合物、对所述蛋白质进行编码的氨基酸序列或对所述蛋白质进行编码的dna序列。优选地,蛋白质变体与亲本蛋白相比具有至少一种氨基酸修饰,例如,与亲本相比,约一种到约七十种
氨基酸修饰,并且优选地约一种到约五种氨基酸修饰。修饰可以是添加、缺失或取代。如下文所描述的,在一些实施方案中,亲本多肽,例如,fc亲本多肽,是人野生型序列,如来自人igg1、 igg2、igg3或igg4的fc区。本文的蛋白质变体序列将优选与亲本蛋白序列具有至少约80%的同一性,并且最优选至少约90%的同一性,更优选至少约95
‑
98
‑
99%的同一性。
[0283]
如本文所使用的,“蛋白质”意指至少两个共价连接的氨基酸,包括蛋白质、多肽、寡肽和肽。当生物功能分子包含两种或更多种蛋白质时,每种蛋白质可称为“单体”或称为“亚基”或称为“结构域”;并且生物功能分子可称为“复合物”。
[0284]
如本文所用的“残基”意指蛋白质中的位置及其相关的氨基酸实体。例如,天冬酰胺 297(也称为asn297或n297)是人抗体igg1中位置297处的残基。
[0285]
如本文所使用的,“igg亚类修饰”或“同种型修饰”意指将一个igg同种型的一个氨基酸转换为不同的比对igg同种型中的对应氨基酸的氨基酸修饰。例如,因为在eu位置296 处igg1包含酪氨酸并且igg2包含苯丙氨酸,所以igg2中的f296y取代被认为是igg亚类修饰。
[0286]
如本文所用的“非天然存在的修饰”意指不是同型的氨基酸修饰。例如,因为igg均不包含位置434处的丝氨酸,所以igg1、igg2、igg3或igg4(或其杂交体)中的取代434s被视为非天然存在的修饰。
[0287]
如本文所用的“氨基酸”和“氨基酸实体”意指由dna和rna编码的20种天然存在的氨基酸之一。
[0288]
如本文所用的“效应子功能”意指由抗体fc区与fc受体或配体的相互作用产生的生物化学事件。效应子功能包括但不限于adcc、adcp和cdc。
[0289]
如本文所使用的,“igg fc配体”或“fc配体”意指来自与igg抗体的fc区结合以形成fc/fc配体复合物的任何生物体的分子,优选地多肽。fc配体包括但不限于fcγri、fcγrii、 fcγriii、fcrn、c1q、c3、甘露聚糖结合凝集素、甘露糖受体、葡萄球菌蛋白a、链球菌蛋白g和病毒性fcγr。fc配体还包括fc受体同源物(fcrh),其为与fcγr同源的fc受体家族 (davis等人,2002,《免疫评论(immunological reviews)》190:123
‑
136,其以引用的方式整体并入本文中)。fc配体可包括结合fc的未发现的分子。特别的igg fc配体是fcrn和fcγ受体。
[0290]
如本文所用的“fcγ受体”、“fcγr”或“fcγr(fcgammar)”意指结合igg抗体fc区并且由fcγr基因编码的蛋白质家族的任何成员。在人类中,这个家族包括但不限于 fcγri(cd64),包括同种型fcγria、fcγrib和fcγric;fcγrii(cd32),包括同种型fcγriia(包括同种异型h131和r131)、fcγriib(包括fcγriib
‑
1和fcγriib
‑
2)和fcγriic;和fcγriii(cd16),包括同种型fcγriiia(包括同种异型v158和f158)和fcγriiib(包括同种异型fcγriiib
‑
na1和 fcγriiib
‑
na2)(jefferis等人,2002,《免疫学快报(immunol lett)》82:57
‑
65,其以引用的方式整体并入本文中),以及任何未发现的人类fcγr或fcγr同种型或同种异型。fcγr可来自任何生物体,包括但不限于人类、小鼠、大鼠、兔和猴。小鼠fcγr包括但不限于fcγri(cd64)、 fcγrii(cd32)、fcγriii(cd16)和fcγriii
‑
2(cd16
‑
2)以及任何未发现的小鼠fcγr或fcγr同种型或同种异型。
[0291]
如本文所用的“fcrn”或“新生儿fc受体”意指结合igg抗体fc区,并且至少部分地由fcrn基因编码的蛋白质。fcγr可来自任何生物体,包括但不限于人类、小鼠、大鼠、兔和
猴。如本领域已知的,功能性fcrn蛋白包括两种多肽,常常被称为重链和轻链。轻链是β
‑ꢀ2‑
微球蛋白(β
‑2‑
微球蛋白),并且重链由fcrn基因编码。除非本文另外指出,否则fcrn或 fcrn蛋白是指fcrn重链与β2
‑
微球蛋白的复合物。各种fc变体可以用于增加与fcrn的结合并且在一些情况下用于增加血清半衰期。通常,除非另有说明,否则本发明的fc单体保持与fcrn的结合(并且,如下所述,可以包括氨基酸变体以增加与fcrn的结合)。
[0292]
如本文所用,“亲本多肽”意指随后修饰以生成变体的起始多肽。亲本多肽可以是天然存在的多肽或者天然存在的多肽的变体或工程化型式。亲本多肽可指代多肽本身、包含所述亲本多肽的组合物,或编码所述亲本多肽的氨基酸序列。
[0293]
如本文所用,“fc”或“fc区”或“fc结构域”意指包含抗体的恒定区,在一些情况下不包含所有第一恒定区免疫球蛋白结构域(例如,ch1)或其一部分,并且在一些情况下,任选地包括铰链的全部或一部分的多肽。对于igg,fc结构域包括免疫球蛋白结构域ch2和ch3 (cγ2和cγ3),以及任选地ch1(cγ1)与ch2(cγ2)之间的铰链区的全部或一部分。因此,在一些情况下,fc结构域从n端到c端包括ch2
‑
ch3和铰链
‑
ch2
‑
ch3。在一些实施方案中, fc结构域是来自igg1、igg2、igg3或igg4的fc结构域,其中igg1铰链
‑
ch2
‑
ch3和igg4 铰链
‑
ch2
‑
ch3特别用于许多实施方案中。另外,在人igg1 fc结构域的情况下,铰链时常包括c220s氨基酸取代。另外,在人igg4 fc结构域的情况下,铰链时常包括s228p氨基酸取代。尽管fc区的边界可变化,但是人类igg重链fc区通常被定义为包括其羧基端的残基 e216、c226或a231,其中编号是根据如kabat中的eu索引。在一些实施方案中,如下文更全面地描述,对fc区进行氨基酸修饰,例如以改变与一种或多种fcγr或fcrn的结合。
[0294]
如本领域的技术人员将理解的,在不同的编号系统之间,重恒定区结构域的精确编号和布置可以有所不同。根据eu和kabat的重恒定区编号的有用比较如下,参见edelman等人, 1969,《美国国家科学院院刊(proc natl acad sci usa)》63:78
‑
85,以及kabat等人,1991,《免疫学关注的蛋白质的序列(sequences of proteins of immunological interest)》,第5版,美国公共卫生署(united states public health service),国立卫生研究院(national institute of health),贝塞斯达(bethesda),所述文献通过引用整体并入。
[0295]
表1
[0296] eu编号kabat编号ch1118
‑
215114
‑
223铰链216
‑
230226
‑
243ch2231
‑
340244
‑
360ch3341
‑
447361
‑
478
[0297]
如本文所用,“fc变体”或“变体fc”意指包括fc结构域中的氨基酸修饰的蛋白质。修饰可以是添加、缺失或取代。本发明的fc变体根据构成它们的氨基酸修饰来定义。因此,例如,n434s或434s是相对于亲本fc多肽在位置434处具有丝氨酸的取代的fc变体,其中编号是根据eu索引。同样地,m428l/n434s定义了相对于亲本fc多肽具有取代m428l和 n434s的fc变体。wt氨基酸的身份标识可以是非特定的,在此情况下前述变体称为 428l/434s。应注意,提供取代的顺序是任意的,也就是说,例如,428l/434s是与434s/428l 相同的fc变体等。对于本发明所讨论的涉及抗体或其衍生物和片段的所有位置,除非另外指出,否则根据
eu索引进行氨基酸位置编号。eu索引或如kabat或eu编号方案中的eu索引是指eu抗体的编号(edelman等人,1969,《美国国家科学院院刊》63:78
‑
85,所述文献特此通过引用整体并入)。修饰可以是添加、缺失或取代。
[0298]
如本文所用,“融合蛋白”是指至少两个蛋白质或蛋白结构域的共价连接。融合蛋白可包括人工序列,例如,结构域接头、变体fc结构域、变体il
‑
10结构域等,如本文所描述的。本文中的“fc融合蛋白”或“免疫粘附素”是指包括fc区的蛋白质,所述fc区通常(任选地通过结构域接头,如本文所述)与一个或多个不同的蛋白结构域连接。因此,“il
‑
10fc融合体”包含与il
‑
10、il
‑
10(a
‑
d)结构域、il10(e
‑
f)结构域、单体il
‑
10和/或单链il
‑
10复合物(scil
‑
10)连接的fc区(任选地但通常通过结构域接头),如本文所述。在一些情况下,两种 fc融合蛋白可以形成同源二聚体fc融合蛋白或异源二聚体fc融合蛋白,后者是优选的。在一些情况下,异源二聚体fc融合蛋白的一个单体包含单独的fc结构域(例如,空fc结构域),并且另一个单体是fc融合体,包含变体fc结构域和il
‑
10结构域,如本文所概述的。
[0299]
如本文所用的“位置”意指蛋白质序列中的位置。位置可顺序地或根据已建立格式例如抗体编号的eu索引进行编号。
[0300]
在本文中,在本发明的异源二聚体蛋白的单体的上下文中,“链型(strandedness)”意指,类似于dna的“相匹配”的两个链,异源二聚化变体并入到每个单体中以便保持、产生和/ 或增强“相匹配”以形成异源二聚体的能力。例如,如果一些pi变体被工程化为单体a(例如,使pi更高),则作为也可以利用的“电荷对”的空间变体不干扰pi变体,例如使pi更高的电荷变体被放在同一个“链”或“单体”上以保存两种功能性。类似地,对于如下文更充分地概述的以成对的组出现的“偏斜”变体,技术人员将认为pi决定将进入并入所述对中的一组的链或单体,从而使得还使用偏斜变体的pi使pi分离最大化。
[0301]
在本文中,“野生型或wt”意指在自然界发现的氨基酸序列或核苷酸序列,包含等位变异。wt蛋白质具有尚未经有意修饰的氨基酸序列或核苷酸序列。
[0302]
本发明的异源二聚体蛋白通常是分离的或重组的。在用于描述本文所公开的各种多肽时,“分离的”意指已经从表达多肽的细胞或细胞培养物中鉴定并且分离和/或回收的多肽。通常,将通过至少一个纯化步骤来制备分离的多肽。“分离的蛋白质”是指基本上不含来自细胞培养物的如宿主细胞蛋白等其它蛋白质的蛋白质。“重组”意指在外源性宿主细胞中使用重组核酸技术生成的蛋白质。
[0303]
关于蛋白质序列的“氨基酸序列同一性百分比(%)”被定义为,在比对序列并引入缺口 (如果必要的话)以实现最大序列同一性百分比之后并且在不将任何保守取代视为序列同一性的一部分的情况下,候选序列中的与特定(亲本)序列中的氨基酸残基相同的氨基酸残基的百分比。出于确定氨基酸序列同一性百分比的目的的比对可以以本领域的能力范围内的各种方式实现,例如使用可公开获得的计算机软件,如blast、blast
‑
2、align或megalign (dnastar)软件实现。本领域技术人员可以确定用于测量比对的适当参数,包括在进行比较的序列的全长内实现最大比对所需的任何算法。一种特定程序是美国公开号20160244525的 [0279]到[0280]段落中所概述的align
‑
2程序,所述美国专利特此通过引用并入。
[0304]
本发明的氨基酸序列(“发明序列”)与亲本氨基酸序列之间的同一性的程度被计算为在两个序列的比对中精确匹配的数除以“发明序列”的长度或亲本序列的长度,以最短
者为准。结果以同一性百分比表示。
[0305]
在一些实施方案中,两个或更多个氨基酸序列具有至少50%、60%、70%、80%或90%同一性。在一些实施方案中,两个或更多个氨基酸序列具有至少95%、97%、98%、99%或甚至 100%同一性。
[0306]“il
‑
10单体”或“il
‑
10单体结构域”在本文是指由6个螺旋组成的单体,所述螺旋在本文称为螺旋a
‑
f。如本文所讨论的,il10单体结构域可以是野生型人序列(例如来自图1的 seq id no:1)或其变体(例如,参见例如图35a
‑
35d)。注意,在本文所述的形式和序列的背景中,残基109(基于成熟形式il10序列编号)可以是h或l。
[0307]
本发明的il
‑
10单体,当以本文所述的形式一起存在时,结合由两个α和两个β亚基(或 r1和r2亚基)组成的il
‑
10受体。可以在相互作用的解离常数(k
d
)方面表达特异性结合的强度或亲和力,其中更小的k
d
表示更大的亲和力并且更大的k
d
表示更小的亲和力。结合特性可以通过本领域众所周知的方法,如生物层干涉测量法和基于表面等离振子共振的方法来确定。一种此类方法需要测量抗原结合位点/抗原或受体/配体复合物缔合和解离的速率,其中速率取决于复合物配偶体的浓度、相互作用的亲和力以及在两个方向上同等地影响速率的几何参数。因此,可以确定缔合速率(k
a
)和解离速率(k
d
),并且k
d
/k
a
的比率等于解离常数k
d
(参见《自然(nature)》361:186
‑
187(1993)和davies等人(1990)《生物化学年鉴(annual revbiochem)》59:439
‑
473)。
[0308]
特定分子的特异性结合可以例如通过针对配体(在这种情况下通常为受体)的k
d
为以下的分子来表现:至少约10
‑4m、至少约10
‑5m、至少约10
‑6m、至少约10
‑7m、至少约10
‑8m、至少约10
‑9m、可替代地至少约10
‑
10
m、至少约10
‑
11
m、至少约10
‑
12
m或更大。典型地,与其受体特异性地结合的分子的k
d
将是对照分子相对于受体的k
d
的20倍、50倍、100倍、 500倍、1000倍、5,000倍、10,000倍或更多倍。
[0309]
而且,特定分子的特异性结合可以例如通过针对配体或受体的k
a
或缔合速率为以下的分子来表现:针对相对于对照物的受体大至少20倍、50倍、100倍、500倍、1000倍、5,000 倍、10,000倍或更多倍。
[0310]
本文中“融合”或“共价连接”是指组分(例如,il
‑
10单体结构域和fc结构域)直接或间接地通过本文所概述的结构域接头通过肽键连接。
[0311]
如本文所用,术语“单链”是指包括通过肽键线性连接的氨基酸结构域的分子。在某些实施方案中,生物功能il10是单链il
‑
10复合物或“scil10”,即两个il
‑
10单体结构域融合以形成单个肽链。在一个特定的此类实施方案中,第一il
‑
10单体的c端连接至第二il
‑
10 单体的n端,在本文中命名为“scil10”。应注意的是,其中两个il
‑
10单体位于同一氨基酸链上的这些单链构建体仍然是含有两个氨基酸链的异源二聚体分子的一部分(例如,scil10 组分和“空臂”fc组分)。
[0312]
ii.二聚体fc融合蛋白
[0313]
在一些方面,本发明涉及包括fc区的二聚体fc融合蛋白,所述fc区通常(任选地通过结构域接头)与一个或多个不同的蛋白结构域连接。
[0314]
在一个方面,二聚体fc融合蛋白是同源二聚体fc融合蛋白。在另一方面,二聚体fc融合蛋白是异源二聚体fc融合蛋白。如将认识到的,除非另外指明,否则本文对本公开所涵盖的融合蛋白的组分的讨论在适当时适用于同源二聚体和异源二聚体fc融合蛋白两者。
[0315]
在一个方面,二聚体fc融合蛋白是il
‑
10二聚体fc融合蛋白,其包括处于不同取向的 il
‑
10单体结构域,使得其一起存在以与il10受体结合。fc结构域可以来源于igg fc结构域,例如igg1、igg2、igg3或igg4 fc结构域,其中igg1 fc结构域在本发明中特别有用。如本文所述,igg1 fc结构域可通常但不总是与消融变体结合使用以消融效应子功能。类似地,当期望低效应子功能时,可使用igg4 fc结构域。
[0316]
对于本文所述的任何fc融合蛋白,每条链的羧基端部分限定主要负责效应子功能的恒定区。kabat等人收集了重链和轻链的可变区的多个一级序列。基于序列的保守度,其将单个一级序列分类为cdr和框架并且列出其列表(参见《免疫学相关序列》,第5版,nih公布案第 91
‑
3242号,e.a.kabat等人,其以引用的方式整体并入本文中)。在整个本发明说明书中,当提及可变结构域中的残基(大致是轻链可变区的残基1
‑
107和重链可变区的残基1
‑
113)时通常使用kabat编号系统并且fc区使用eu编号系统(例如kabat等人,同上(1991))。
[0317]
在免疫球蛋白的igg亚类中,重链中存在几种免疫球蛋白结构域。本文中通过“免疫球蛋白(ig)结构域”意指具有不同三级结构的免疫球蛋白的区域。本发明感兴趣的是重链结构域,包括恒定重链(ch)结构域和铰链结构域。在igg抗体的上下文中,igg同种型各自具有三个ch区。因此,在igg的上下文中,“ch”结构域如下:“ch1”是指根据如kabat中的eu索引的位置118
‑
215。“铰链”是指根据如kabat中的eu索引的位置216
‑
230。“ch2”是指根据如kabat中的eu索引的位置231
‑
340,并且“ch3”是指根据如kabat中的eu索引的位置341
‑
447。如表1所示,在不同的编号系统之间,重链结构域的精确编号和布置可以有所不同。如本文所示和下文所述的,pi变体可以处于一个或多个ch区以及铰链区中,如下文讨论的。
[0318]
在免疫球蛋白的igg亚类中,重链中存在几种免疫球蛋白结构域。本文中通过“免疫球蛋白(ig)结构域”意指具有不同三级结构的免疫球蛋白的区域。本发明感兴趣的是重链结构域,包括恒定重链(ch)结构域和铰链结构域。在igg抗体的上下文中,igg同种型各自具有三个ch区。因此,在igg的上下文中,“ch”结构域如下:“ch1”是指根据如kabat中的eu索引的位置118
‑
215。“铰链”是指根据如kabat中的eu索引的位置216
‑
230。“ch2”是指根据如kabat中的eu索引的位置231
‑
340,并且“ch3”是指根据如kabat中的eu索引的位置341
‑
447。如表1所示,在不同的编号系统之间,重链结构域的精确编号和布置可以有所不同。如本文所示和下文所述的,pi变体可以处于一个或多个ch区以及铰链区中,如下文讨论的。
[0319]
重链的另一种类型的ig结构域是铰链区。“铰链”或“铰链区”或“抗体铰链区”或“免疫球蛋白铰链区”在本文中意指包含抗体的第一与第二重链恒定结构域之间的氨基酸的柔性多肽。在结构上,igg ch1结构域在eu位置215处终止,并且igg ch2结构域在残基 eu位置231处开始。因此,对于igg,抗体铰链在本文中定义为包括位置216(igg1中的e216) 到230(igg1中的p230),其中编号是根据如kabat中的eu索引。在一些实施方案中,例如在fc区的上下文中,包括铰链,通常是指位置216
‑
230。如本文所指出,pi变体还可以在铰链区中制成。
[0320]
如本文所述和本领域已知的,本发明的二聚体蛋白包含不同的结构域,所述结构域也可以重叠。这些结构域包括但不限于fc结构域、ch2结构域、ch3结构域和铰链结构域以及 il
‑
10单体结构域。如本文所述,这些结构域以不同的形式连接在一起,如图16、20、23和 29中总体概述的。
[0321]
在本文所述的一些实施方案中,当蛋白结构域例如il
‑
10单体与fc结构域连接时,
是蛋白结构域的c端连接于fc结构域的n端。在本文所述的其他实施方案中,当蛋白结构域例如il
‑
10单体与fc结构域连接时,是蛋白质片段的n端连接于fc结构域的c端。
[0322]
在fc结构域蛋白的本文概述的一些构建体和序列中,il
‑
10单体的c端与结构域接头的 n端连接,其c端与恒定fc结构域(n
‑
il
‑
10单体结构域
‑
接头
‑
fc结构域
‑
c)的n端连接,尽管如将认识到的,可以将组分的顺序切换为:(n
‑
fc结构域
‑
接头
‑
il
‑
10单体结构域
‑
c)。在本文概述的其它构建体和序列中,第一蛋白结构域的c端与第二蛋白结构域的n端连接,任选地通过结构域接头,第二蛋白结构域的c端与恒定fc结构域的n端连接,任选地通过结构域接头。在另一个构建体中,第一蛋白结构域的n端与第二蛋白结构域的c端连接,任选地通过结构域接头,第二蛋白质片段的n端与恒定fc结构域的c端连接,任选地通过结构域接头。在本文概述的其它构建体和序列中,提供了未附接至第一蛋白质片段或第二蛋白质片段的恒定fc结构域。二聚体fc融合蛋白可以含有两个或更多个本文所述的示例性单体fc结构域蛋白
‑
如将认识到的,同源二聚体fc融合蛋白将含有两个相同的单体fc结构域蛋白,而异源二聚体fc融合蛋白将含有两个或更多个不同的单体fc结构域蛋白。
[0323]
在一些实施方案中,接头是“结构域接头”,其用于将如本文概述的任何两个结构域连接在一起,其中的一些在附图中描绘。虽然可以使用任何合适的接头,但许多实施方案使用甘氨酸
‑
丝氨酸聚合物,包括例如(gs)n、(gsggs)n、(ggggs)n和(gggs)n,其中n是至少一的整数(并且通常为0到1到2到3到4到5)以及允许具有足够的长度和柔性的两个结构域重组连接的任何肽序列以允许每个结构域保留其生物学功能。在一些情况下,并且注意“链型”,如下文所概述,接头是带电荷的结构域接头。
[0324]
如上文所指出,本公开涵盖异源二聚体和非异源二聚体(本文也称为同源二聚体)形式。如本领域技术人员将认识到的,此类同源二聚体形式可以是单特异性抗体(例如“传统单克隆抗体”和/或结合单一靶标的fc融合蛋白)或非异源二聚体双特异性形式。因此,本发明提供单克隆(单特异性)fc融合蛋白或非异源二聚体双特异性蛋白。非异源二聚体双特异性形式是本领域已知的,并且包括如spiess等人,《分子免疫学(molecular immunology)》(67):95
‑
106 (2015)和kontermann,mabs 4:2,182
‑
197(2012)中总体上描绘的许多不同形式,所述两个文献均通过引用明确并入,特别是针对附图、图例以及对其中形式的引用而明确并入。
[0325]
在一些实施方案中,本发明提供异源二聚体fc融合蛋白,其依赖于使用两个不同的重链变体fc序列,其将自组装以形成异源二聚体fc结构域融合多肽。在一个实施方案中,异源二聚体fc融合蛋白含有至少两个恒定结构域,所述至少两个恒定结构域可以被工程化以产生异源二聚体,如pi工程化。可以使用的其它fc结构域包括含有已经过pi工程化的本发明的 ch1、ch2、ch3和铰链结构域中的一个或多个的片段。具体地,图16c
‑
d、20a
‑
b、23c
‑
f 和29a
‑
d中描绘的形式是异源二聚体fc融合蛋白,意指所述蛋白质具有自组装成异源二聚体fc结构域的两个缔合的fc序列和至少一个蛋白质片段(例如,1、2或更多个蛋白质片段)。在一些情况下,第一蛋白质片段连接至第一fc序列,并且第二蛋白质片段连接至第二fc序列。在一些情况下,异源二聚体fc融合蛋白含有第一蛋白质片段,所述第一蛋白质片段连接至第二蛋白质片段,所述第二蛋白质片段接至第一fc序列;以及第二fc序列,所述第二fc 序列不连接至第一或第二蛋白质片段。
[0326]
本发明涉及用于提供允许与一种或多种结合配偶体、配体或受体结合的二聚体fc
融合蛋白的新型构建体。fc融合构建体基于抗体的重链的两个fc结构域(例如,组装成“二聚体”的两个“单体”)的自组装性质。通过改变如下面更充分讨论的每个单体的氨基酸序列来制备异源二聚体fc融合体形式。因此,本发明总体涉及非异源二聚体和异源二聚体fc融合蛋白的产生,其可以以若干方式共接合一种或多种结合配偶体或配体或受体。在另外的实施方案中,异源二聚体fc融合蛋白依赖于恒定区中的在每条链上不同的氨基酸变体以促进异源二聚体形成和/或允许相比于同源二聚体更容易地纯化异源二聚体。存在多种机制可以用于生成本发明的异二聚体。另外,如本领域技术人员将了解的,这些机制可以组合以确保高异源二聚化。因此,导致异源二聚体产生的氨基酸变体被称为“异源二聚化变体”。如下文所讨论的,异源二聚化变体可以包括空间变体(例如,下文所描述的“旋钮与孔(knobs and holes)”或“偏斜”变体和下文所描述的“电荷对”)以及“pi变体”,这允许纯化同源二聚体远离异源二聚体。如在特此通过引用以其全文并入的wo2014/145806中通常描述的,并且具体如下文所讨论的“异源二聚化变体”,异源二聚化的有用机制包括“旋钮与孔”(“kih”;有时在本文描述为“偏斜”变体(参见wo2014/145806中的讨论))、如wo2014/145806中所描述的“静电转向”或“电荷对”、如wo2014/145806中所描述的pi变体以及如wo2014/145806以及下文中所概述的一般另外的fc变体。
[0327]
在本发明中,存在可以使纯化异源二聚体蛋白和抗体变得容易的若干种基本机制;一种机制依赖于使用pi变体,使得每个单体以及随后每个二聚体物类具有不同pi,从而允许等电纯化a
‑
a、a
‑
b和b
‑
b二聚体蛋白。可替代地,一些形式也允许基于大小的分离。如下文中进一步概述的,还可能相比于同源二聚体“偏斜”异源二聚体的形成。因此,空间异源二聚化变体和pi或电荷对变体的组合特别用于本发明。
[0328]
通常,特别用于本发明的实施方案依赖于包括偏斜变体的变体组,所述偏斜变体相比于同源二聚化形成鼓励异源二聚化形成、与pi变体偶联,所述pi变体增加两个单体与每个二聚体物类之间的pi差。
[0329]
另外,如下文更全面地概述的,取决于异源二聚体fc融合蛋白的形式,pi变体可以包含在单体的恒定结构域和/或fc结构域内,或可以使用结构域接头。也就是说,本发明提供了在单体中的一个或两个单体和/或带电结构域接头上的pi变体。另外,用于替代性功能的另外的氨基酸工程化还可赋予pi变化,如fc、fcrn和ko变体。
[0330]
在将pi用作分离机制以允许纯化异源二聚体蛋白的本发明中,可以将氨基酸变体引入到单体多肽中的一个或两个中;也就是说,可以使单体之一(为了简单起见,在本文称为“单体 a”)的pi工程化远离单体b,或者单体a和单体b两者均可以随着单体a的pi增加和单体 b的pi降低而改变。如所讨论的,任一个单体或两个单体的pi变化可以通过以下进行:去除或添加带电荷残基(例如,由带正电荷的氨基酸残基或带负电荷的氨基酸残基替代中性氨基酸,例如,谷氨酰胺到谷氨酸)、将带电荷残基从带正电或带负电改变为相反电荷(例如,天冬氨酸到赖氨酸)或将带电荷残基改变为中性残基(例如,电荷损失;赖氨酸到丝氨酸)。多种这些变体在附图中示出。
[0331]
因此,本发明的这个实施方案提供了产生单体中的至少一个单体的充分pi变化,从而使得异源二聚体可以与同源二聚体分离。如本领域技术人员将了解的并且如下文中进一步讨论的,这可以通过以下来完成:使用“野生型”重链恒定区和已经工程化以增大或减小其pi的变体区(wt a:b 或wt a:b
‑
)或增大一个区并减小另一个区(a :b
‑
或a
‑
:b )。
[0332]
因此,通常,本发明的一些实施方案的组分是恒定区中的氨基酸变体,所述氨基酸变体涉及通过将氨基酸取代(“pi变体”或“pi取代”)并入到单体中的一个或两个单体中来更改二聚体蛋白的单体中的至少一个单体(如果不是两个单体的话)的等电点(pi)。异源二聚体与两个同源二聚体的分离可以在两个单体的pi相差少至0.1个ph单位时完成,其中0.2、0.3、0.4 和0.5或更多个ph单位均可用于本发明。
[0333]
如本领域技术人员将了解的,每一个或两个单体上要包括的用于得到良好分离的pi变体数量将部分地取决于组分的起始pi。也就是说,为了确定对哪个单体进行工程化或处于哪个“方向”(例如,更具阳性或更具阴性),计算fc结构域的序列,并且在一些情况下,计算与 fc结构域连接的一个或多个蛋白结构域,并据此做出决定。如本领域已知的,不同的fc结构域和/或蛋白结构域将具有用于本发明中的不同起始pi。通常,如本文所概述的,pi被工程化以使各个单体的总pi差为至少约0.1log,如本文所概述的优选的是0.2到0.5。
[0334]
此外,如本领域技术人员将了解的并且如本文所概述的,在一些实施方案中,可以基于大小来使异源二聚体与同源二聚体分离。如图所示,例如,若干种形式允许基于大小来分离异源二聚体和同源二聚体。
[0335]
在pi变体用于实现异源二聚化的情况下,通过使用一个或多个fc结构域的一个或多个恒定区,提供了用于设计和纯化异源二聚体fc融合蛋白的更模块化的方法。因此,在一些实施方案中,异源二聚化变体(包括偏斜和纯化异源二聚化变体)必须被工程化。另外,在一些实施方案中,通过从不同的igg同种型输入pi变体,由pi变体造成免疫原性的可能性显著降低,使得在未引入显著免疫原性的情况下改变pi。因此,要解决的另一个问题是阐明具有高人序列含量的低pi恒定结构域,例如最小化或避免任何特定位置处的非人残基。
[0336]
可与此pi工程化一起发生的附加益处同样是血清半衰期的延长和fcrn结合的增加。也就是说,如ussn 13/194,904(以引用方式以其全文并入)中所描述的,降低抗体恒定结构域(包括在抗体和fc融合体中找到的那些)的pi可能导致血清保留在体内的时间更长。具有增加的血清半衰期的这些pi变体也促进pi变化以进行纯化。
[0337]
另外,应注意,异源二聚化变体的pi变体给予了fc融合蛋白的分析和质量控制过程另外的益处,因为在同源二聚体存在时消除、最小化和进行区分的能力是显著的。类似地,能够可靠地测试异源二聚体fc融合蛋白产生的可重现性很重要。
[0338]
a.异源二聚化变体
[0339]
本发明提供了包括呈各种形式的异源二聚体fc融合蛋白的异源二聚体蛋白,所述异源二聚体蛋白利用异源二聚体变体以允许异源二聚体形成和/或远离同源二聚体而纯化。异源二聚体融合构建体基于两个fc结构域的自组装性质,例如,组装成“二聚体”的两个“单体”。
[0340]
存在适合的多对异源二聚化偏斜变体组。这些变体以成“对”的“组”出现。也就是说,所述对中的一组并入第一单体中并且所述对中的另一组并入第二单体中。应注意,这些组不一定表现为“孔中旋钮”变体,一个单体上的残基与另一个单体上的残基一一对应;也就是说,这些对组形成了两个单体之间的界面,所述界面鼓励异源二聚体形成并且不鼓励同源二聚体形成,从而允许在生物学条件下自发形成的异源二聚体的百分比超过90%,而非预期的 50%(25%同源二聚体a/a:50%异源二聚体a/b:25%同源二聚体b/b)。
[0341]
b.空间变体
[0342]
在一些实施方案中,可以通过添加空间变体来促进异源二聚体形成。也就是说,通过改变每条重链中的氨基酸,相比于形成具有相同fc氨基酸序列的同源二聚体,不同的重链更可能缔合以形成异源二聚体结构。合适的空间变体包括在ussn 15/141,350的图29中以及图5 中,所有所述文献特此通过引用以其全文并入。
[0343]
一种机制在本领域通常称为“旋钮和孔”,是指形成空间影响以利于异源二聚体形成和不利于同源二聚体形成的氨基酸工程化,如在ussn 61/596,846,ridgway等人,proteinengineering 9(7):617(1996);atwell等人,j.mol.biol.1997 270:26;美国专利第8,216,805号中所述,所有这些都以引用方式全文并入本文中。附图标识了许多依赖于“旋钮和孔”的“单体a
‑
单体b”对。另外,如merchant等人,nature biotech.16:677(1998)中所述,这些“旋钮和孔”突变可以与二硫键组合以偏斜形成异源二聚化。
[0344]
用于产生异源二聚体的另一种机制有时称为“静电转向”,如gunasekaran等人,j.biol. chem.285(25):19637(2010)中所述,所述文献以引用方式全文并入本文中。这在本文有时被称为“电荷对”。在此实施方案中,静电用于使形成朝异源二聚化偏斜。如本领域的技术人员将理解的,这些电荷对还可能影响pi并因此影响纯化,并且因此在一些情况下还可以被视为 pi变体。然而,由于这些是为了强制发生异源二聚化而产生并且不是作为纯化工具使用,因此其被分类为“空间变体”。这些包括但不限于d221e/p228e/l368e与d221r/p228r/k409r 配对(例如,这些是“单体”对应组)和c220e/p228e/368e与c220r/e224r/p228r/k409r配对。
[0345]
其它单体a和单体b变体可以任选且独立地以任何量与其它变体(如本文中概述的pi变体或us 2012/0149876的图37中所示的其它空间变体,其全部以引用方式明确并入本文中) 组合。
[0346]
在一些实施方案中,本文所概述的空间变体可以任选且独立地与任何pi变体(或其它变体,如fc变体、fcrn变体等)并入到一个或两个单体中,并且可以独立且任选地包含在本发明的蛋白质中或并不包含在内。
[0347]
合适的偏斜变体的列表在图5中找到。在多个实施方案中,特别适用的是以下组中的对,包括但不限于s364k/e357q:l368d/k370s;l368d/k370s:s364k;l368e/k370s:s364k; t411t/e360e/q362e:d401k;l368d/k370s:s364k/e357l;k370s:s364k/e357q;以及 t366s/l368a/y407v:t366w(任选地包括二硫桥键,t366s/l368a/y407v/y349c: t366w/s354c)。就命名法而言,对“s364k/e357q:l368d/k370s”意指单体中的一者具有双变体组s364k/e357q并且另一者具有双变体组l368d/k370s;如上文所述,这些对的“链型”取决于起始pi。
[0348]
c.异源二聚体的pi(等电点)变体
[0349]
通常,如本领域技术人员将了解的,存在两种一般类别的pi变体:增大蛋白质的pi(碱性变化)的变体和减小蛋白质的pi(酸性变化)的变体。如本文所述,可以使用这些变体的所有组合:一个单体可以是野生型或是不表现出与野生型显著不同的pi的变体,并且另一个可以更具碱性或更具酸性。可替代地,可改变每个单体,一个更具碱性且一个更具酸性。
[0350]
pi变体的优选组合在ussn 15/141,350的图30中示出,所有这些文献都以引用的方式全文并入本文中。如本文所概述和附图中所示,这些变化相对于igg1示出,但所有同种型可以这种方式改变,以及同种型杂合体。在重链恒定结构域来自igg2
‑
4的情况下,也可以
使用 r133e和r133q。
[0351]
在一个实施方案中,如果fc单体中的一个包括ch1结构域,那么pi变体的优选组合具有包含208d/295e/384d/418e/421d变体(当相对于人igg1时为 n208d/q295e/n384d/q418e/n421d)的一个单体。在一些例子中,第二单体包含带正电荷的结构域接头,包括(gkpgs)4。在一些情况下,第一单体包括ch1结构域,包括位置208。因此,在不包括ch1结构域的构建体中(例如对于在其中一个结构域上不利用ch1结构域的异源二聚体fc融合蛋白),优选的负pi变体fc组包括295e/384d/418e/421d变异(当相对于人 igg1时为q295e/n384d/q418e/n421d)。
[0352]
在一些实施方案中,在fc结构域的铰链中进行突变,包括位置216、217、218、219、 220、221、222、223、224、225、226、227、228、229和230。因此,pi突变以及特别地取代可以在位置216
‑
230中的一个或多个位置中进行,其中1、2、3、4或5个突变可用于本发明中。同样,设想了单独或与其它结构域中的其它pi变体一起的所有可能的组合。
[0353]
可用于降低铰链结构域的pi的具体取代包括但不限于221位处的缺失、222位处的非天然缬氨酸或苏氨酸、223位处的缺失、224位处的非天然谷氨酸、225位处的缺失、235位处的缺失,以及236位处的缺失或非天然丙氨酸。在一些情况下,仅在铰链结构域中进行pi取代,并且在其它情况下,这些取代以任何组合添加至其它结构域中的其它pi变体。
[0354]
在一些实施方案中,可以在ch2区域中进行突变,包括位置233、234、235、236、274、 296、300、309、320、322、326、327、334和339。应该注意,可以进行233
‑
236的改变以增加igg2骨架中的效应子功能(以及327a)。同样,可以对这14个位置进行所有可能的组合;例如,pi抗体可具有1、2、3、4、5、6、7、8、9或10个ch2 pi取代。
[0355]
可用于降低ch2结构域的pi的具体取代包括但不限于:位置274处的非天然谷氨酰胺或谷氨酸、位置296处的非天然苯丙氨酸、位置300处的非天然苯丙氨酸、位置309处的非天然缬氨酸、位置320处的非天然谷氨酸、位置322处的非天然谷氨酸、位置326处的非天然谷氨酸、位置327处的非天然甘氨酸、位置334处的非天然谷氨酸、位置339处的非天然苏氨酸,以及ch2内的和与其它结构域的所有可能组合。
[0356]
在这个实施方案中,突变可以独立且任选地选自位置355、359、362、384、389、392、 397、418、419、444和447。可用于降低ch3结构域的pi的具体取代包括但不限于:位置 355处的非天然谷氨酰胺或谷氨酸、位置384处的非天然丝氨酸、位置392处的非天然天冬酰胺或谷氨酸、位置397处的非天然甲硫氨酸、位置419处的非天然谷氨酸、位置359处的非天然谷氨酸、位置362处的非天然谷氨酸、位置389处的非天然谷氨酸、位置418处的非天然谷氨酸、位置444处的非天然谷氨酸,以及位置447处的缺失或非天然天冬氨酸。
[0357]
d.同种型变体
[0358]
另外,本发明的许多实施方案依赖于特定位置处的pi氨基酸从一个igg同种型到另一个的“输入”,由此降低或消除将不想要的免疫原性引入变体中的可能性。许多这些在美国公开申请第2014/0370013号的图21中示出,该美国公开申请以引用方式并入本文中。也就是说,出于多种原因,包括高效应子功能,igg1是治疗性抗体的常见同种型。然而,igg1的重链恒定区具有比igg2的重链恒定区更高的pi(8.10相对于7.31)。通过将特定位置的igg2残基引入igg1骨架中,使得所得单体的pi降低(或提高)且另外表现出较长的血清半衰期。例如,igg1在位置137处具有甘氨酸(pi 5.97),并且igg2具有谷氨酸(pi 3.22);输入谷
氨酸将影响所得蛋白质的pi。如下文所述,通常需要许多氨基酸取代来显著影响变体fc融合蛋白的 pi。然而,应该注意,如下文所论述,即使igg2分子的变化也允许增加血清半衰期。
[0359]
在其它实施方案中,进行非同种型氨基酸变化,以减少所得蛋白质的总电荷状态(例如通过将更高的pi氨基酸变为更低的pi氨基酸),或允许在结构中调节稳定性等等,如下文更进一步描述。
[0360]
另外,通过pi工程化重恒定结构域和轻恒定结构域两者,可以看到异源二聚体的每个单体的显著变化。如本文所讨论的,使两个单体的pi相差至少0.5可以通过离子交换色谱法或等电聚焦或对等电点敏感的其它方法进行分离。
[0361]
e.计算pi
[0362]
每个单体的pi可以取决于变体重链恒定结构域的pi和总单体的pi,所述总单体包括变体重链恒定结构域和融合配偶体。因此,在一些实施方案中,使用美国公开申请第 2014/0370013号中的图19的图表,基于变体重链恒定结构域来计算pi变化。如本文所讨论的,要工程化哪个单体通常由每个单体的固有pi来决定。
[0363]
f.还赋予了更好的fcrn体内结合的pi变体
[0364]
在pi变体减小单体的pi的情况下,其可以具有提高体内血清固位的附加益处。
[0365]
虽然仍在审查中,但是据信,fc区在体内具有较长的半衰期,因为在核内体中以ph 6与 fcrn的结合螯合了fc(ghetie和ward,1997《今日免疫学(immunol today.)》18(12):592
‑
598,所述文献特此通过引用整体并入)。核内体区室然后使fc循环到细胞表面。一旦区室向胞外空间打开,则约7.4的较高ph诱导将fc释放回到血液中。在小鼠中,dall'acqua等人示出了,在ph为6和ph为7.4时具有增加的fcrn结合的fc突变体实际上具有降低的血清浓度和与野生型fc相同的半衰期(dall'acqua等人,2002,《免疫学杂志(j.immunol.)》169:5171
‑ꢀ
5180,所述文献特此通过引用整体并入)。据认为,在ph为7.4时fcrn的fc的增加的亲和力阻止将fc释放回到血液中。因此,将增加fc体内半衰期的fc突变理想地将增强较低ph 下的fcrn结合,同时仍允许fc在较高ph下释放。在6.0至7.4的ph范围内,氨基酸组胺酸改变了其电荷状态。因此,在fc/fcrn复合物中的重要位置找到his残基并不出乎意料。
[0366]
g.用于额外功能的额外fc变体
[0367]
除了pi氨基酸变体之外,存在出于各种原因可以进行的许多有用的fc氨基酸修饰,包括但不限于更改与一个或多个fcγr的结合、与fcrn的更改结合等。
[0368]
因此,本发明的蛋白质可以包括氨基酸修饰,包括本文所概述的异源二聚化变体,所述异源二聚化变体包括pi变体和空间变体。每组变体可以独立且任选地包括在任何特定异源二聚体蛋白中或不包括在内。
[0369]
h.fcγr变体
[0370]
因此,存在可以更改与fcγ受体中的一个或多个的结合而进行的多个有用fc取代。导致增加的结合以及减少的结合的取代可以是有用的。例如,已知与fcγriiia的结合增加引起adcc增加(抗体依赖性细胞介导的细胞毒性;细胞介导的反应,其中表达fcγr的非特异性细胞毒性细胞识别靶细胞上所结合的抗体且随后造成靶细胞溶解)。类似地,在一些情况下,与fcγriib(一种抑制性受体)的结合减少也可能是有利的。可以用于本发明中的氨基酸取代包括ussn 11/124,620(特别是图41)、ussn 11/174,287、ussn 11/396,495、
ussn 11/538,406 中列出的那些,其均明确地以引用方式整体并入本文中并且特别是针对其中公开的变体。可以使用的特定变体包括但不限于236a、239d、239e、332e、332d、239d/332e、267d、267e、 328f、267e/328f、236a/332e、239d/332e/330y、239d、332e/330l、243a、243l、264a、 264v和299t。
[0371]
另外,增加对fcγriic的亲和力的氨基酸取代也可包括在本文概述的fc结构域变体中。举例来说,ussn 11/124,620和14/578,305中描述的取代是有用的。
[0372]
另外,存在额外的fc取代,其可用于增加与fcrn的结合和增加血清半衰期,如具体公开于ussn 12/341,769中,其以引用的方式全文并入本文中,包括但不限于434s、434a、 428l、308f、259i、428l/434s、259i/308f、436i/428l、436i or v/434s、436v/428l和 259i/308f/428l。
[0373]
i.消融变体
[0374]
类似地,另一类功能变体是“fcγr消融变体”或“fc敲除(fcko或ko)”变体。在这些实施方案中,对于一些治疗应用,期望减少或去除fc结构域与一种或多种或所有fcγ受体(例如fcγr1、fcγriia、fcγriib、fcγriiia等)的正常结合,以避免其它作用机制。即,例如,在许多实施方案中,特别是在使用免疫调节蛋白时,期望消融fcγriiia结合以消除或显著降低 adcc活性,使得一个fc结构域包含一个或多个fcγ受体消融变体。这些消融变体在ussn 15/141,350的图31中描绘,其均通过引用方式整体并入本文中,并且每个都可以独立地并且任选地被包括或排除,其中优选的方面利用选自由以下项组成的组的消融变体:
[0375]
根据eu索引,g236r/l328r、e233p/l234v/l235a/g236del/s239k、 e233p/l234v/l235a/g236del/s267k、e233p/l234v/l235a/g236del/s239k/a327g、 e233p/l234v/l235a/g236del/s267k/a327g和e233p/l234v/l235a/g236del。此外,图7中还描绘了用于本发明中的消融变体。应该注意的是,本文提及的消融变体消融fcγr结合,但通常不消融fcrn结合。
[0376]
j.异源二聚体变体和fc变体的组合
[0377]
如本领域技术人员将了解的,所有所列举的异源二聚化变体(包括偏斜变体和/或pi变体) 可以以任何方式任选且独立地组合,只要其保持其“链型”或“单体分隔”即可。另外,所有这些变体可以组合成异源二聚化形式中的任一种异源二聚化形式。
[0378]
在pi变体的情况下,虽然附图中示出了特别有用的实施方案,但是可以遵循改变两个单体之间的pi差以促进纯化的基本规则,以生成其它组合。
[0379]
另外,任何异源二聚化变体、偏斜和pi也可独立且任选地与如本文一般概述的fc消融变体、fc变体、fcrn变体组合。
[0380]
另外,单体fc结构域可包含一组氨基酸取代,该一组氨基酸取代包括 c220s/s267k/l368d/k370s或c220s/s267k/s364k/e357q。
[0381]
另外,异源二聚体fc融合蛋白可以包含偏斜变体(例如,如ussn 15/141,350的图1a
‑
1c 中所示的一组氨基酸取代,所有文献以引用的方式全文并入本文中),其中特别有用的偏斜变体选自由以下组成的组:s364k/e357q:l368d/k370s;l368d/k370s:s364k;l368e/k370s: s364k;t411t/e360e/q362e:d401k;l368d/k370s:s364k/e357l;k370s:s364k/e357q; t366s/l368a/y407v:t366w;以及t366s/l368a/y407v:t366w(任选地包括二硫桥键, t366s/l368a/y407v/y349c:t366w/s354c)、任选的消融变体、任选的带电荷的结构域接头;以及任选的pi变体。
[0382]
在一些实施方案中,fc结构域包含选自由以下组成的组的一个或多个氨基酸取代:根据eu索引的236r、s239d、s239e、f243l、m252y、v259i、s267d、s267e、s67k、s298a、 v308f、l328f、l328r、330l、i332d、i332e、m428l、n434a、n434s、236r/l328r、 s239d/i332e、236r/l328f、v259i/v308f、s267e/l328f、m428l/n43s、y436i/m428l、 n436v/m428l、v436i/n434s、y436v/n434s、s239d/i332e/330l、m252y/s54t/t256e、 v259i/v308f/m428l、e233p/l234v/l235a/g236_/s239k、 e233p/l234v/l235a/g236_/s239k/a327g、e233p/l234v/l235a/g236_/s267k/a327g、 e233p/l234v/l235a/g236_以及e233p/l234v/l235a/g236_/s267k。
[0383]
在一个实施方案中,可用于本发明的偏斜和pi变体的特定组合是t366s/l368a/y407v: t366w(任选地包括二硫桥键,t366s/l368a/y407v/y349c:t366w/s354c),其中一种单体包含q295e/n384d/q418e/n481d并且另一单体包含带正电荷的结构域接头。如本领域将理解的,“孔包旋钮(knobs in holes)”变体不改变pi,并且因此可用于任一单体上。
[0384]
在另一个实施方案中,本发明的异源二聚体包括异源二聚体变体和fc结构域变体的组合,诸如附图中描绘的那些。
[0385]
在一些实施方案中,示例性ch1和部分铰链区用于将可变结构域共价连接至本发明的靶向性il
‑
10融合蛋白的fc骨架。此类序列在图12和相应seq id no的序列表中提供。
[0386]
在一些实施方案中,本文提供的任何非靶向和靶向性il
‑
10融合蛋白的fc结构域包含促进fc结构域的异源二聚化的修饰。在一些实施方案中,促进异源二聚化的修饰是选自由以下组成的组的一组氨基酸取代:根据eu编号的l368d/k370s和s364k;l368d/k370s和 s364k/e357l;l368d/k370s和s364k/e357q;t411e/k360e/q362e和d401k;l368e/k370s 和s364k;k370s和s364k/e357q以及t366s/l368a/y407v:t366w(任选地包括二硫桥键, t366s/l368a/y407v/y349c:t366w/s354c)。在一些实施方案中,fc结构域具有另外一组包括根据eu编号的q295e/n384d/q418e/n421d的氨基酸取代。在一些实施方案中,fc结构域具有由以下组成的另外一组氨基酸取代:根据eu编号的g236r/l328r、 e233p/l234v/l235a/g236_/s239k、e233p/l234v/l235a/g236_/s239k/a327g、 e233p/l234v/l235a/g236_/s267k/a327g、e233p/l234v/l235a/g236_以及 e233p/l234v/l235a/g236_/s267k。在一些实施方案中,fc结构域包含选自由根据eu编号的m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。另外的修饰在附图中提供,包括图5a
‑
11c。
[0387]
iii.白介素10
[0388]
本发明涉及白介素10的生物学功能形式。如上所述,白介素10(或“il
‑
10”或“il10”) 的生物学功能形式是通过两个il
‑
10单体(在本文也称为“il
‑
10单体结构域”)之间的非共价相互作用形成的结构域交换的同源二聚体。生物功能il
‑
10二聚体结合il
‑
10受体,所述受体由两个α和两个β亚基(或r1和r2亚基)组成。值得注意的是,il
‑
10二聚体在各个il
‑
10单体之间的相互作用被破坏时变得没有生物活性。根据本文进一步详细描述的fc融合蛋白,可以使用il
‑
10单体结构域的不同形式。一些示例性形式在图14中示出。每个单体可包含两个结构域:(i)(a
‑
d)结构域(图14a),其包含人il
‑
10蛋白的成熟形式的残基1
‑
116,以及(ii)(e
‑ꢀ
f)结构域(图14b),其包含人il
‑
10蛋白的成熟形式的残基117
‑
160
‑
。如上文所论述,生物活性il
‑
10包含结构域交换的同源二聚体,其在图14c中示意性地示出。在本发明中使用的另一形式是单链il
‑
10复合物或“scil
‑
10”,其包含任选地通过接头与第二
il
‑
10单体共价连接的第一il
‑
10单体(参见图14d)。在本发明中使用的另一il
‑
10形式在图14e中示出,其描绘了例如josephson等人(2000)中描述的单体il
‑
10或“il10m1”,所述文献特此通过引用整体并入,特别是与il10m1相关的所有教导,其通过在hil
‑
10(a
‑
d)(例如,il
‑
10单体的a
‑ꢀ
d结构域)与hil
‑
10(e
‑
f)(例如,il
‑
10单体的e
‑
f结构域)之间工程化结构域接头而生成。在本发明中使用的另一il
‑
10形式是分裂il
‑
10形式,其包含与hil
‑
10(e
‑
f)结构域非共价连接的hil
‑
10(a
‑
d)结构域,如图14f所描绘。
[0389]
在一些实施方案中,人il
‑
10蛋白具有在ncbi参考seq.no.np_000563.1、seq id no: 1(人il
‑
10(109h)前体序列)、seq id no:2(人il
‑
10(109l)前体序列)、seq id no:3(人il
‑ꢀ
10(109h)成熟形式序列)、seq id no:4(人il
‑
10(109l)成熟形式序列)中列出的氨基酸序列,全部如图1中所描绘。应当理解,在本发明中使用的il
‑
10蛋白可在位置109处包括组氨酸 (109h)或在其他实施方案中在位置109处包括亮氨酸(109l)。在一些实施方案中,il
‑
10单体结构域序列与全部如图1所示的seq id no:1(人il
‑
10(109h)前体序列)、seq id no:2(人 il
‑
10(109l)前体序列)、seq id no:3(人il
‑
10(109h)成熟形式序列)、seq id no:4(人il
‑ꢀ
10(109l)成熟形式序列)具有至少90%,例如90%、91%、92%、93%、94%、95%、96%、97%、 98%、99%或更大的序列同一性。fc融合蛋白的il
‑
10单体结构域在图1描绘的氨基酸序列中还可具有1、2、3、4、5、6、7、8或9个氨基酸突变。在本发明中使用的il
‑
10组分的另外的示例性序列和配置在包括图15a
‑
15d、21a
‑
21c、22、35a
‑
35d、36a
‑
36e、39a
‑
39g、 40a
‑
40m、41a
‑
41f、48、49a
‑
49b、53、55、56a
‑
56c、57、58、59a
‑
59h、60、64、65a
‑
65b、 66a
‑
66d、67、68、69、70a
‑
70b、71a
‑
71e、73、4a
‑
74b、75a
‑
75b、76a
‑
76d、78a
‑
78g、 79a
‑
79g和86a
‑
86d的附图以及相应序列标识符的序列表中提供。
[0390]
本发明还提供了变体il
‑
10单体结构域。这些变体可用作本文所述的生物功能il
‑
10复合物以及il
‑
10
‑
fc融合体中的任何融合体的一部分。
[0391]
a.表达变体
[0392]
在一些实施方案中,本发明的il
‑
10单体结构域亚基可以包括改善/增加表达的变体,特别是对于异源二聚体形式。此类变体可包括降低或去除脱酰胺作用的潜力、降低或去除天冬氨酸异构化的潜力、降低或去除降解相关ptm的潜力、减少或去除潜在降解位点、减少/去除二硫桥和/或减少或去除潜在n
‑
糖基化位点的变体。在本发明中使用的il
‑
10组分和表达变体的示例性序列在包括图15a
‑
15d、21a
‑
21c、22、35a
‑
35d、36a
‑
36e、39a
‑
39g、40a
‑
40m、 41a
‑
41f、48、49a
‑
49b、53、55、56a
‑
56c、57、58、59a
‑
59h、60、64、65a
‑
65b、66a
‑
66d、 67、68、69、70a
‑
70b、71a
‑
71e、73、4a
‑
74b、75a
‑
75b、76a
‑
76d、78a
‑
78g、79a
‑
79g和 86a
‑
86d的附图以及相应序列标识符的序列表中提供。
[0393]
应当理解,本发明的il
‑
10fc融合蛋白的不同形式将含有不同的氨基酸修饰和氨基酸修饰的组合,以实现上述功能,并从而改善/增加本发明的蛋白质的表达。本发明的fc融合蛋白的不同形式的不同变体序列的示例性实施方案描绘于图15a
‑
15d、21a
‑
21c、22、35a
‑
35d、 36a
‑
36e、39a
‑
39g、40a
‑
40m、41a
‑
41f、48、49a
‑
49b、53、55、56a
‑
56c、57、58、59a
‑ꢀ
59h、60、64、65a
‑
65b、66a
‑
66d、67、68、69、70a
‑
70b、71a
‑
71e、73、4a
‑
74b、75a
‑
75b、 76a
‑
76d、78a
‑
78g、79a
‑
79g和86a
‑
86d以及相应序列标识符的序列表中。
[0394]
b.亲和力变体
[0395]
本发明提供了il
‑
10变体,其形成对il
‑
10受体的结合亲和力发生改变(即,降低或
增加) 的生物功能il
‑
10。此类变体也可或替代地显示出改变的(降低的或增加的)效力。
[0396]
il
‑
10上可以引入氨基酸修饰的合适位点包括n21、d28、q38、m39、d41、q42、l43、d44、n45、i87、e142、d144和e151(根据人il
‑
10(il
‑
10成熟形式序列,如图1所描绘,具有109h或109l序列)编号)。这些位点处的例示性修饰包括n21d、d28n、q38e、m39t、 d41n、q42e、l43v、d44n、n45d、i87a、e142q、d144n和e151q。在一些实施方案中, il
‑
10变体包含选自由以下组成的组的一个或多个取代:n21、d28、q38、d41、d44、q42、 n45、e142、d144和e151(根据如图1所描绘的人il
‑
10成熟形式序列编号)。在一些实施方案中,il
‑
10变体包含选自由以下组成的组的一个或多个取代:n21d、d28n、q38e、d41n、 q42e、d44n、n45d、e142q、d144n和e151q(根据如图1所描绘的人il
‑
10成熟形式序列编号)。在一些实施方案中,il
‑
10变体还包含选自由m39、l43和i87组成的组的一个或多个取代。在一些情况下,取代包括m39t、l43v、i87a,以及其任何组合。il
‑
10亲和力/效力变体可以在这些位点中的一个或多个处包括修饰。在另外的实施方案中,用于改变亲和力和/ 或效力的一种或多种氨基酸修饰可以与一种或多种另外的修饰(包括表达变体)组合。这些修饰可以单独使用或与任何其他il
‑
10变体组合使用,诸如调节亲和力/效力、引入/去除二硫桥、提高收率、去除电荷变体、去除糖基化位点和/或改善稳定性的变体。
[0397]
在本发明中使用的亲和力/效力变体的示例性组合包括q38e/d41n、q38e/q42e、 q38e/n45d、q38e/e142q、q38e/d144n、d41n/q42e、d41n/n45d、d41n/e142q、 d41n/d144n、q42e/n45d、q42e/e142q、q42e/d144n、n45d/e142q、n45d/d144n、 e142q/d144n、n21d/q42e、n21d/n45d、n21d/e151q、n21d/n45e、q42e/e151q、 n45d/e151q、n21d/q42e/n45d、n21d/q42e/e151q、q42e/n45d/e151q、n21d/q42e/n45e 和n21d/q42e/n45d/e151q。
[0398]
在一些实施方案中,本发明的任何形式的il
‑
10变体包含任何选自由以下组成的组的以下取代:q38e/q198e、d41n/d201n、q42e/q202e、n45d/n205d、e142q/e302q、 e142q/e302q、d144n/d304n、q38e/d41n/q198e/d201n、q38e/q42e/q198e/q202e、 q38e/n45d/q198e/n205d、q38e/e142q/q198e/e302q、q38e/d144n/q198e/d304n、 d41n/q42e/d201n/q202e、d41n/n45d/d201n/n205d、d41n/e142q/d201n/e302q、 d41n/d144n/d201n/d304n、q42e/n45d/q202e/n205d、q42e/e142q/q202e/e302q、 q42e/d144n/q202e/d304n、n45d/e142q/n205d/e302q、n45d/d144n/n205d/d304n、 e142q/d144n/e302q/d304n、n21d/n181d、e151q/e311q、n21d/q42e/n181d/q202e、 n21d/n45d/n181d/n205e、n21d/n45d/n181d/n205d、n21d/e151q/n181d/e311q、 q42e/e151q/e311q、n45d/e151q/n205d/e311q、n21d/q42e/n45d/n181d/q202e/n205e、 n21d/q42e/e151q/n181d/q202e/e311q、q42e/n45d/e151q/q202e/n205d/e311q、 n21d/q42e/n45d/e151q/n181d/q202e/n205d/e311q、q38e/n45d/n205d、 d41n/n45d/n205d、n45d/d144n/n205d、n205d、n45d、n21d/n45d/n205d、 n45d/n181d/n205d、n45d/e151q/n205d、n45d/n205d/e311q以及n45d/n205d/d304n。
[0399]
在一些实施方案中,本发明的任何形式的il
‑
10变体包含任何选自由以下组成的组的以下取代:n10a、n10q、n92a、n126a、n160a、n10a/n45d/n170a/n205d、 n45d/n92a/n205d/n252a、n45d/n126a/n205d/n286a、n45d/n160a/n205d、 n45d/n205d/n320a、n45d/n160a/n205d/n320a、n10a/n45d/n170a/n205d/n320a、 n10a/n45d/n170a/n205d/n320_、n10a/n45d/n170a/n205d/r319_/n320_、 n10a/n45d/n170a/n205d/i318_/r319_/n320_、 n10a/n45d/n170a/n205d/k317_/i318_/r319_/n320_、n10a/n45d/n205d/n320a、n45d/
n170a/n205d/n320a、n10q/n45d/n170q/n205d/n320a、d28a以及 d28a/n45d/d188a/n205d。在一些实施方案中,scil10变体包含任何选自由以下组成的组的以下取代:n10a、n92a、n126a、n160a、n170a、n170q、n252a、n286a、n320a、k317_、 i318_、r319_、n320_,以及其任何组合。在一些实施方案中,一个或多个取代降低融合蛋白脱酰胺作用的潜力。
[0400]
在一些实施方案中,本发明的任何形式的il
‑
10变体包含任何选自由以下组成的组的以下取代:d28a和d28a/n45d/d188a/n205d。在一些实施方案中,一个或多个取代降低天冬氨酸异构化的潜力。
[0401]
在一些实施方案中,本发明的任何形式的il
‑
10变体包含任何选自由以下组成的组的以下取代:n116d、n116q、k117p、s118a、n45d/n116d/n205d、n45d/n116d/n205d、 n45d/k117p/n205d、n45d/s118a/n205d、n45d/n205d/n276d、n45d/n205d/n276q、 n45d/n205d/k277p、n45d/n205d/s178a以及n45d/n116d/n205d/n276d。在一些实施方案中,取代减少n
‑
糖基化位点。
[0402]
在一些实施方案中,本发明的任何形式的il
‑
10变体包含任何选自由以下组成的组的以下取代:c12a/c108a和c12a/n45d/c108a/c172a/n205d/c268a/n320a。在一些实施方案中,一个或多个取代降低形成二硫桥的潜力。
[0403]
在一些实施方案中,本发明的任何形式的il
‑
10变体包含任何选自由以下组成的组的以下取代:f37c/m140c、q38c/s141c、d41c/k138c、l47c/k138c、l48c/e142c、s51c/a120c、 d55c/a120c、f56c/y153c、c62a/c114a、a64c/s118c、m68c/v121c以及v76c/a139c。在一些实施方案中,il10m1变体在以下残基处包含一个或多个半胱氨酸取代:螺旋a
‑
d中的f37、q38、d41、l47、l48、s51、d55、f56、a64、m68和v76;以及螺旋e
‑
f中的s118、 a120、v121、k138、a139、m140、s141、e142和y153。在一些情况下,在il10m1变体中发现以下氨基酸对之间的二硫桥:f37c:m140c、q38c:s141c、d41c:k138c、l47c: k138c、l48c:e142c、s51c:a120c、d55c:a120c、f56c:y153c、a64c:s118c、m68c: v121c以及v76c:a139c促进螺旋a
‑
d与螺旋e
‑
f之间的相互作用的稳定。在一些实施方案中,一个或多个取代允许形成二硫桥。
[0404]
在一些实施方案中,本发明的任何形式的il
‑
10变体包含任何选自由以下组成的组的以下取代:l47q、s118a和a139q。在一些实施方案中,任何形式的il
‑
10变体包含任何选自由以下组成的组的以下取代:l19、l23、l26、a29、f30、v33、f37、l47、l48、l52、f56、 a64、l65、s66、m68、i69、f71、y72、v76、m77、a80、e81、i87、v91、l94、g95、l98、l101、l105、l112、s118、v121、v124、f128、k138、a139、m140、s141、f143、f146、 i147和i150。
[0405]
在本发明中使用的il
‑
10组分和亲和力变体的示例性序列提供于包括图15a
‑
15d、21a
‑ꢀ
21c、22、35a
‑
35d、36a
‑
36e、39a
‑
39g、40a
‑
40m、41a
‑
41f、48、49a
‑
49b、53、55、56a
‑ꢀ
56c、57、58、59a
‑
59h、60、64、65a
‑
65b、66a
‑
66d、67、68、69、70a
‑
70b、71a
‑
71e、 73、4a
‑
74b、75a
‑
75b、76a
‑
76d、78a
‑
78g、79a
‑
79g和86a
‑
86d的附图以及相应序列标识符的序列表中。
[0406]
生物功能il
‑
10二聚体可以包含两个野生型il
‑
10单体或变体il
‑
10单体和野生型il
‑
10 单体。
[0407]
iv.抗原结合结构域
[0408]
如下文所论述的,通常使用术语“抗体”或“抗原结合结构域”。可用于本发明的抗原结合结构域可呈现如本文所述的多种形式,包括本文所述和附图中所描绘的传统抗体以及抗体衍生物、片段和模拟物。在一些实施方案中,本发明提供含有检查点抗原结合结构域
和fc 结构域的抗体融合蛋白。在一些实施方案中,本发明提供含有免疫细胞抗原结合结构域和fc 结构域的抗体融合蛋白。
[0409]
传统抗体结构单位通常包括四聚体。每个四聚体通常由相同的两对多肽链构成,每对具有一条“轻链”(通常具有约25kda的分子量)和一条“重链”(通常具有约50
‑
70kda的分子量)。人轻链被分类为κ轻链和λ轻链。本发明涉及通常基于igg类的抗体或抗体片段(抗体单体),其具有几个亚类,包括但不限于igg1、igg2、igg3和igg4。通常,igg1、igg2和 igg4比igg3更频繁地使用。应注意,igg1具有在356(d或e)和358(l或m)处具有多型性的不同同种异型。本文描述的序列使用356d/358m同种异型,然而本文包括其它同种异型。也就是说,本文包括的包含igg1 fc结构域的任何序列可以具有替代356d/358m同种异型的 356e/358l。
[0410]
另外,本文中的许多序列的至少一个在位置220处的半胱氨酸被丝氨酸替代;通常,这是在本文中所描绘的大多数序列的“scfv单体”侧,尽管它也可以在“fab单体”侧或两者上,以减少二硫化物的形成。本文序列中特别包括这些半胱氨酸中的一个或两个被替代 (c220s)。
[0411]
靶向性il
‑
10fc融合蛋白的pd
‑
1抗原结合结构域(abd)(例如,抗pd
‑
1组分)通常是一组6个cdr和/或可变重结构域和可变轻结构域,其形成可结合人pd
‑
1的fv结构域。如本文所示,抗pd
‑
1abd可以是scfv的形式,其中vh和vl结构域使用scfv接头连接,所述scfv接头可以任选地是带电荷的scfv接头。如本领域技术人员所理解的,scfv可以从n端到c端组装为n
‑
vh
‑
scfv接头
‑
vl
‑
c或为n
‑
vl
‑
scfv接头
‑
vh
‑
c,其中scfv结构域的c端通常与铰链
‑
ch2
‑
ch3 fc结构域连接。可以以scfv形式或fab形式使用合适的fv(包括cdr组和可变重/可变轻结构域),在附图中示出以及在wo2017/218707和在2018年11月8日提交的pct/us2018/059887中公开,其特此明确地整体并入本文,特别是用于描绘抗pd
‑
1序列的附图、图例和seq标识符。
[0412]
在一些实施方案中,pd
‑
1abd基于1c11克隆,如图所示。在一些实施方案中,本发明的pd
‑
1abd基于:基于图100a
‑
100b中所示的1c11克隆的重链的变体重链和基于图100a
‑ꢀ
100b中所示的1c11克隆的轻链的变体轻链。在一些实施方案中,pd
‑
1abd包含变体重链和轻链对,其选自由图100a
‑
100g和相应的seq id no中描绘的重链和轻链对组成的组,包括1c11[pd
‑
1]_h0l0、1c11[pd
‑
1]_h3l3、1c11[pd
‑
1]_h3.240_l3.148、1c11[pd
‑ꢀ
1]_h3.241_l3.148、1c11[pd
‑
1]_h3.234_l3.144、1c11[pd
‑
1]_h3.241_l3.92、1c11[pd
‑ꢀ
1]_h3.303_l3.152、1c11_h3.329_l3.220、1c11_h3.328_l3.152、派姆单抗vh和vl链、纳武单抗vh和vl链、皮地利珠单抗vh和vl链、mk
‑
3475vh和vl链、bap049克隆evh和vl链、bap049克隆b vh和vl链、h7798n vh和vl链、h1h3 var6 vh和vl链、 ape2058 vh和vl链、h005
‑
1 vh和vl链、317
‑
4b6、326
‑
4a3 vh和vl链、hpd
‑
1mab7 (1.2)vh和vl链、克隆38[pd
‑
1]vh和vl链、克隆39[pd
‑
1]vh和vl链、克隆41[pd
‑
1] vh和vl链、克隆48[pd
‑
1]vh和vl链、pd1
‑
17[pd
‑
1]vh和vl链、pd1
‑
28[pd
‑
1]vh和 vl链、pd1
‑
33[pd
‑
1]vh和vl链、pd1
‑
35[pd
‑
1]vh和vl链、lopd180[pd
‑
1]vh和vl 链、ab948[pd
‑
1]vh和vl链、人源化的eh
‑
12.2h7 vh和vl链、rg1h10 vh和vl链、 rg1h10
‑
h2a
‑
22
‑
1s vh和vl链、rg1h10
‑
h2a
‑
27
‑
2s vh和vl链、rg1h10
‑
3c vh和vl 链、rg1h10
‑
16c vh和vl链、rg1h10
‑
17c vh和vl链、rg1h10
‑
19c vh和vl链、 rg1h10
‑
21c vh和vl链、rg1h10
‑
23c2 vh和vl链、mab7[pd
‑
1]vh和vl链以及 pd1ab
‑
6[pd
‑
1]vh和vl链。如本领
域技术人员将理解的,vh和vl结构域可以被格式化为fab或scfv,以用于本发明的靶向pd
‑
1的il
‑
10融合蛋白。
[0413]
在一些实施方案中,非竞争性pd
‑
1abd基于mab a、mab b和mab c克隆,如图所示。在一些实施方案中,本发明的非竞争性pd
‑
1abd基于:基于图104a
‑
104b中所示的maba、mab b或mab c克隆的重链的变体重链和基于图104a
‑
104b中所示的mab a、mab b 或mab c克隆的轻链的变体轻链。在一些实施方案中,非竞争性pd
‑
1abd包含变体重链和轻链对,其选自由图104a
‑
104b和相应的seq id no中描绘的重链和轻链对组成的组,包括mab a[pd
‑
1]_h1_l1和mab b[pd
‑
1]_h1_l1。
[0414]
在一些实施方案中,非竞争性pd
‑
1abd包含选自由mab c[pd
‑
1]_h1、mab c[pd
‑ꢀ
1]_h1.19、mab c[pd
‑
1]_h1.48、mab c[pd
‑
1]_h1.125、mab c[pd
‑
1]_h1.130、mab c[pd
‑ꢀ
1]_h1.132、mab c[pd
‑
1]_h1.169、mab c[pd
‑
1]_h1.175和mab c[pd
‑
1]_h2组成的组的变体重链,以及选自由mab c[pd
‑
1]_l1、mab c[pd
‑
1]_l1.1、mab c[pd
‑
1]_l1.3、mab c[pd
‑ꢀ
1]_l1.45、mab c[pd
‑
1]_l1.117、mab c[pd
‑
1]_l1.129、mab c[pd
‑
1]_l1.135、mab c[pd
‑ꢀ
1]_l1.136、mab c[pd
‑
1]_l1.140和mab c[pd
‑
1]_l2组成的组的变体轻链。如本领域技术人员将理解的,vh和vl结构域可以被格式化为fab或scfv,以用于本发明的靶向性il
‑
10融合蛋白。
[0415]
在一些实施方案中,tigit abd包含选自由2a5b4[tigit]_h1_l1、4.1d3.q1e(替拉格鲁单抗(tiragolumab))[tigit]_h0_l0、10a7[tigit]vh和vl链、1f4[tigit]以及4.1d3[tigit] vh和vl链组成的组的变体重链。如本领域技术人员将理解的,vh和vl结构域可以被格式化为fab或scfv,以用于本发明的靶向性il
‑
10融合蛋白。
[0416]
在一些实施方案中,tigit abd包含选自由hu14d7 vh1、hu14d7 vh2和hu14d7 vh3 组成的组的变体重链,以及选自由hu14d7 vl1、hu14d7 vl2和hu14d7 vl3组成的组的变体轻链。如本领域技术人员将理解的,vh和vl结构域可以被格式化为fab或scfv,以用于本发明的靶向性il
‑
10融合蛋白。
[0417]
在一些实施方案中,tigit abd包含选自由hu26b10 vh1、hu26b10 vh2和hu26b10 vh3组成的组的变体重链,以及选自由hu26b10 vl1、hu26b10 vl2和hu26b10 vl3组成的组的变体轻链。如本领域技术人员将理解的,vh和vl结构域可以被格式化为fab或scfv,以用于本发明的靶向性il
‑
10融合蛋白。
[0418]
在一些实施方案中,tigit abd包含选自由以下组成的组的变体重链和轻链对: meb125.31c6.a1.205 vh4/vl1、meb125.31c6.a1.205 vh5/vl4、meb125.31c6.a1.205 vh5/vl3、15a6 vh和vl链、22g2a6 vh和vl链、11g11 vh和vl链、10d7 vh和vl 链、10d7 vh和vl链、313r19 vh和vl链以及艾替吉利单抗(etigilimab)vh和vl链。如本领域技术人员将理解的,vh和vl结构域可以被格式化为fab或scfv,以用于本发明的靶向性il
‑
10融合蛋白。
[0419]
在一些实施方案中,nkg2d abd包含选自由以下组成的组的变体重链和轻链对: ms[nkg2d]h0_l0、1d7b4[nkg2d]_h1_l1、kyk
‑
1.0[nkg2d]_h1_l1、kyk
‑ꢀ
2.0[nkg2d]_h0_l0、11b2d10[nkg2d]_h0_l0、6e5a7[nkg2d]_h0_l0、6h7e7[nkg2d]_h0_l0、mab e[nkg2d]_h1_l1、16f31[nkg2d]_h1_l1、mabd[nkg2d]_h1_l1、1d7b4[nkg2d]_h1_l1、mab a[nkg2d]_h1_l1、maba[nkg2d]_h1_l2、mab a[nkg2d]_h2_l1、mab a[nkg2d]_h2_l2、mabb[nkg2d]_h1_l1、mab b[nkg2d]_h1_l1.1、mab b[nkg2d]_h1_l2、mabb[nkg2d]_h2_l1、mab b[nkg2d]_
h2_l1.1、mab b[nkg2d]_h2_l2、mabb[nkg2d]_h3_l1、mab b[nkg2d]_h3_l1.1、mab b[nkg2d]_h3_l2、mabc[nkg2d]_h1_l1、mab c[nkg2d]_h1_l2、mab c[nkg2d]_h2_l1以及mabc[nkg2d]_h2_l2。如本领域技术人员将理解的,vh和vl结构域可以被格式化为fab或 scfv,以用于本发明的靶向性il
‑
10融合蛋白。
[0420]
在一些实施方案中,cd8 abd包含选自由okt8_h2_l1和1c11b3[cd8]_h1_l1组成的组的变体重链和轻链对。如本领域技术人员将理解的,vh和vl结构域可以被格式化为fab 或scfv,以用于本发明的靶向性il
‑
10融合蛋白。
[0421]
v.结构域接头
[0422]
在一些实施方案中,il
‑
10单体和/或il
‑
10单体螺旋结构域(例如,螺旋(a
‑
d)和(e
‑
f)通过接头连接在一起。任选地,单体和/或单体螺旋结构域不通过接头连接。在其他实施方案中, il
‑
10单体或螺旋结构域非共价连接。在一些实施方案中,il
‑
10单体和/或il
‑
10单体螺旋结构域通过接头连接至fc结构域。在某些实施方案中,il
‑
10单体和/或螺旋结构域直接连接至 fc结构域,诸如不使用接头。在一些情况下,不使用接头将il
‑
10单体或il
‑
10单体螺旋结构域连接至fc结构域。
[0423]
在一些实施方案中,接头是“结构域接头”,其用于将如本文概述的任何两个结构域连接在一起。虽然可以使用任何适合的接头,但是许多实施方案利用甘氨酸
‑
丝氨酸聚合物以及允许以足够允许每个结构域保持其生物功能的长度和柔性来重组连接这两个结构域的任何肽序列,所述甘氨酸
‑
丝氨酸聚合物包含例如(gs)
n
、(gsggs)
n
、(ggggs)
n
和(gggs)
n
,其中n 是至少为0(并且通常为0到1到2到3到4到5)的整数。在某些情况下,有用的接头包括 (ggggs)0或(ggggs)1或(ggggs)2。图103中描绘了例示性结构域接头。在一些情况下并且在注意“链型”时,如下文所概述的,如本文所讨论的可以使用带电荷的结构域接头。
[0424]
vi.本发明的有用非靶向形式
[0425]
如图16、20、23和29所示,存在许多本发明的非靶向性il
‑
10融合蛋白的有用形式。通常,本发明的融合蛋白具有两种功能组分:il
‑
10二聚体组分和fc组分,两者都可以采取与本文概述的形式不同的形式,并且其中的每一者都可以以任何配置与其它组分组合。如图87、88、89和90所示,存在许多本发明的靶向性il
‑
10融合蛋白的有用形式。通常,本发明的融合蛋白具有两种功能组分:il
‑
10组分、fc组分和抗原结合结构域,它们全部可以采用与本文概述的形式不同的形式,并且其中的每一者都可以以任何配置与其它组分组合。应当理解,本文所述的任何形式中的il
‑
10组分可包括在位置109处具有组氨酸或在位置109处具有亮氨酸的il
‑
10。il
‑
10组分还可包括本文描述的和本领域已知的任何il
‑
10单体变体。
[0426]
在一些实施方案中,il
‑
10单体任选地用结构域接头共价连接,并且在本文称为单链il
‑ꢀ
10复合物或“scil
‑
10”。
[0427]
在一些实施方案中,il
‑
10单体不共价连接,而是与作为二聚体组装的第一和第二fc结构域分别共价连接。
[0428]
在一些实施方案中,il
‑
10单体的不同螺旋彼此连接或与第一和第二fc结构域连接。在另外的实施方案中,il
‑
10单体的螺旋(a
‑
d)在单链中与螺旋(e
‑
f)共价连接,任选地通过结构域接头。在其他实施方案中,螺旋(a
‑
d)共价连接至第一fc结构域(任选地通过结构域接头)并且螺旋(e
‑
f)共价连接至第二fc结构域(任选地通过结构域接头),使得当第一
和第二fc结构域缔合时,(a
‑
d)螺旋与(e
‑
f)螺旋非共价相互作用。应当理解,在一些实施方案中,第一和第二fc结构域是相同的,并且在一些实施方案中,第一和第二fc结构域是不相同的。
[0429]
第一和/或第二fc结构域可在fc区具有缺失以帮助增强可制造性。在一些实施方案中,此类缺失可包括但不限于g446del/k447del。
[0430]
第一和第二fc结构域可以具有选自由以下组成的组的一组氨基酸取代:根据eu编号的 a)l368d/k370s和s364k;b)l368d/k370s和s364k/e357l;c)l368d/k370s和 s364k/e357q;d)t411e/k360e/q362e和d401k;e)l368e/k370s和s364k;f)k370s和 s364k/e357q;以及g)t366s/l368a/y407v:t366w(任选地包括二硫桥键, t366s/l368a/y407v/y349c:t366w/s354c)。
[0431]
在一些实施方案中,第一和/或第二fc结构域具有另外一组包括根据eu编号的 q295e/n384d/q418e/n421d的氨基酸取代。
[0432]
任选地,第一和/或第二fc结构域具有由以下组成的另外一组氨基酸取代:根据eu编号的g236r/l328r、e233p/l234v/l235a/g236_/s239k、 e233p/l234v/l235a/g236_/s239k/a327g、e233p/l234v/l235a/g236_/s267k/a327g、 e233p/l234v/l235a/g236_以及e233p/l234v/l235a/g236_/s267k。
[0433]
任选地,第一和/或第二fc结构域具有用于半衰期延长的428l/434s变体。
[0434]
应当理解,本文论述的任何氨基酸取代,包括任何前述取代,可与g445del/k447del修饰以任何组合使用。
[0435]
在另外的优选实施方案中,本文所述的形式含有抗原结合结构域,其中那些抗原结合结构域靶向il
‑
15。在另外的实施方案中,抗原结合结构域是非竞争性pd
‑
1结合结构域。
[0436]
a.il10
‑
fc融合体形式
[0437]
为了工程化其中预复合了il
‑
10同源二聚体的il
‑
10融合蛋白,一个实施方案包括il10
‑ꢀ
fc融合体形式(图16中描绘的卡通示意图)。这种类别的一种这样的形式是(il10)2‑
fc形式(图 16a中描绘的卡通示意图),其包含两个相同的单体,每个单体包含与同源二聚体fc链的n 端共价连接的il
‑
10单体。(il10)2
‑
fc形式的例示性蛋白质包括xenp24628、xenp24629、 xenp24630和xenp24631,其序列在图17中描绘。
[0438]
另一个实施方案是fc
‑
(il10)2形式(图16b中描绘的卡通示意图),其包含两个相同的单体,每个单体包含与同源二聚体fc链的c端共价连接的il
‑
10单体。fc
‑
(il10)2形式的例示性蛋白质包括xenp24632、xenp24633和xenp246334,其序列在图18中描绘。
[0439]
另一个实施方案是(il10
‑
nc
‑
il10)
‑
异源fc形式(图16c中描绘的卡通示意图),其包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接(任选地通过结构域接头)的第一 il
‑
10单体;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链,同时第二il
‑
10单体单独转染,以形成非共价il
‑
10二聚体。(il10
‑
nc
‑
il10)
‑
异源fc形式的例示性蛋白质包括xenp25955,其序列在图19中描绘。
[0440]
在一些实施方案中,这种形式的il
‑
10单体包含本文所述的任何l
‑
10单体结构域(例如,变体il
‑
10单体结构域)(例如,图55、57、64、67、70a
‑
70b和73、相应的seq id no和序列表中所示的那些)。
[0441]
b.scil10
‑
fc融合体形式
[0442]
在另一方面,本公开提供了一种将il
‑
10同源二聚体预复合为单链il
‑
10复合物(或“scil
‑ꢀ
10”)的形式,其中第一il
‑
10单体与第二il
‑
10单体共价连接。一种类别的利用scil
‑
10的 il
‑
10融合体在本文中称为scil10
‑
fc融合体(图20中描绘的卡通示意图)。
[0443]
在一些实施方案中,这种形式的scil
‑
10包含氨基酸序列,诸如但不限于seq id no:23 和本文所述的任何scil10变体(例如,图21a
‑
21c、22、35a
‑
35d、39a
‑
39g、40a
‑
40m、41a
‑ꢀ
41f、48、49a
‑
49b、53、55、56a
‑
56c、58、59a
‑
59h、60、64、65a
‑
65b、66a
‑
66d、67、 68、69、70a
‑
70b、73、78a
‑
78g、79a
‑
79g和86a
‑
86d、相应的seq id no以及序列表中所示的那些)。在一些实施方案中,scil
‑
10组分包含与seq id no:23具有至少90%(例如,至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更大)序列同一性的氨基酸序列。
[0444]
一个实施方案是scil10
‑
异源fc形式(图20a中描绘的卡通示意图),其包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的scil
‑
10;以及第二单体,其包含为“仅 fc”或“空fc”的互补的第二异源二聚体fc链。scil10
‑
异源fc形式的例示性蛋白质包括 xenp25238、xenp25239、xenp25240、xenp25241、xenp25880和xenp28295,其序列在图21a
‑
21c和序列表中的相应seq id no中描绘。
[0445]
在一些实施方案中,scil10
‑
异源fc融合蛋白包含图78a
‑
78g和序列表中针对 xenp30003、xenp30004、xenp30005、xenp30006、xenp30007、xenp30008、xenp30009、 xenp30010、xenp30011、xenp30012、xenp30013、xenp30014和xenp30015列出的序列。xenp30007也称为scil10.104
‑
fc。xenp30008也称为scil10.105
‑
fc。xenp30010也称为scil10.107
‑
fc。
[0446]
在一些实施方案中,异源fc
‑
scil10融合蛋白包含图79a
‑
79g和序列表中针对 xenp30016、xenp30017、xenp30018、xenp30019、xenp30020、xenp30021、xenp30022、 xenp30023、xenp30024、xenp30025、xenp30026、xenp30027以及xenp30028列出的序列。
[0447]
在一些实施方案中,在fc结构域中含有m428l/n434s取代的scil10融合蛋白包含图 86a
‑
86d和序列表中针对xenp31091、xenp31092、xenp31093、xenp31094、xenp31095、 xenp31096、xenp31830和xenp31831列出的序列。
[0448]
另一个实施方案是异源fc
‑
scil10形式(图20b中描绘的卡通示意图),其包含:第一单体,其包含与第一异源二聚体fc链的c端共价连接(任选地通过结构域接头)的scil
‑
10;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链。异源fc
‑
scil10 的例示性蛋白质是xenp28740,其序列在图22和序列表中的相应seq id no中描绘。
[0449]
在一些实施方案中,此示例性形式的fc融合蛋白包含图20a所示的形式,其具有il
‑
10 变体和/或在fc结构域中具有氨基酸取代。在一些实施方案中,此示例性形式的fc融合蛋白具有图21a和相应序列标识符的序列表中列出的序列。在一些实施方案中,fc融合蛋白具有图78a
‑
78g和序列表中针对xenp30003、xenp30004、xenp30005、xenp30006、 xenp30007、xenp30008、xenp30009、xenp30010、xenp30011、xenp30012、xenp30013、 xenp30014和xenp30015列出的序列。在另外的实施方案中,此示例性形式的fc融合蛋白具有图78a
‑
78g中针对xenp30005(seq id no:25和seq id no:26)、xenp30008(seqid no:27和seq id no:28)以及xenp30013(seq id no:29和seq id no:30)列出的序列。在另外的实施方案中,此示例性形式的fc融合蛋白包含图78a
‑
78g中针对xenp30005、 xenp30008和
xenp30013列出的序列,其具有选自下组的另外的氨基酸修饰(单独或以任何组合):根据eu编号的g445del、k447del、g445del/k447del、m428l、n434s和m428l/n434s。
[0450]
c.il10m1
‑
fc融合体
[0451]
在另一方面,本公开提供了一种被工程化以规避对两个il
‑
10单体之间的结构域交换的需要的形式。如上文所论述,形成生物活性il
‑
10同源二聚体的非共价相互作用的破坏产生生物学上无活性的il
‑
10单体。然而,josephson等人(2000)报道了“il
‑
10m1”(或“il10m1”;图15d中描绘的序列),其为通过在il
‑
10单体的螺旋d与e之间工程化gly
‑ꢀ
ser接头(gggsgg;seq id no:xx)而生成的生物活性il
‑
10单体。josephson等人报道,il
‑ꢀ
10m1能够结合il
‑
10r1并募集il
‑
10r2,以诱导il
‑
10细胞反应。因此,在此方面,fc融合蛋白利用il10m1,并且这种形式在本文中称为il10m1
‑
fc融合体(图23a
‑
23f中描绘的卡通示意图)。
[0452]
在一些实施方案中,这种形式的il10m1包含氨基酸序列,诸如但不限于seq id no:24 和本文所述的任何il10m1变体(例如,图36a
‑
36e、55、57、64、67、70a
‑
70b、71a
‑
71e、 73、74a
‑
74b、75a
‑
75b和76a
‑
76d、相应的seq id no以及序列表中所示的那些)。在一些实施方案中,il10m1变体包含选自由以下组成的组的一个或多个取代:n21d、d28n、q38e、 m39t、d41n、q42e、l43v、d44n、n45d、i87a、e142q、d144n、e151q、q38e/d41n、 q38e/q42e、q38e/n45d、q38e/e142q、q38e/d144n、d41n/q42e、d41n/q42e、d41n/n45d、 d41n/e142q、d41n/d144n、q42e/n45d、q42e/e142q、q42e/d144n、n45d/e142q、 n45d/d144n、e142q/d144n、n21d/q42e、n21d/n45d、n21d/e151q、n21d/n45e、 q42e/e151q、n45d/e151q、n21d/q42e/n45d、n21d/q42e/e151q、q42e/n45d/e151q、 n21d/q42e/n45e和n21d/q42e/n45d/e151q。在一些实施方案中,il10m1组分包含与seqid no:24具有至少90%(例如,至少90%、91%、92%、93%、94%、95%、96%、97%、98%、 99%或更大)序列同一性的氨基酸序列。
[0453]
在一些情况下,所述形式的il10m1组分包含选自由图75a
‑
75b和相应序列标识符的序列表中描绘的序列组成的组的氨基酸序列。在一些实施方案中,il10m1组分包含选自由以下组成的组的接头(例如,插入肽):pggsgg(seq id no:xx)、gpgsgg(seq id no:xx)、 ggpsgg(seq id no:xx)、gggpgg(seq id no:xx)、gggspg(seq id no:xx)、 gggsgp(seq id no:xx)、ggggg(seq id no:xx)、gggg(seq id no:xx)、ggg、 gg、g以及ggggsggggs(seq id no:xx)。
[0454]
在一个实施方案中,(il10m1)1‑
异源fc(图23e中描绘的卡通示意图)包含与第一异源二聚体fc链的n端共价连接(任选地通过接头)的il10m1,以及包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链的第二单体。(il10m1)1
‑
异源fc形式的例示性蛋白质是 xenp14246,其序列在图24中描绘。在另一个实施方案中,异源fc
‑
(il10m1)1形式(图23f 中描绘的卡通示意图)包含与第一异源二聚体fc链的c端共价连接(任选地通过接头)的 il10m1,以及包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链的第二单体。异源 fc
‑
(il10m1)1形式的例示性蛋白质是xenp14247,其序列在图25和序列表中的相应seq idno中描绘。在一些实施方案中,(il10m1)1‑
异源fc融合蛋白包含图37a
‑
37e和序列表中针对xenp25753、xenp25754、xenp25755、xenp25756、xenp25757、xenp25758、 xenp25759、xenp25760、xenp25761、xenp25763、xenp25766、xenp25767以及 xenp25768和相应序列标识符列出的序列。
[0455]
在另一个实施方案中,(il10m1)2‑
fc形式(图23a中描绘的卡通示意图)包含两个
相同的单体,其中每个单体包含与同源二聚体fc链的n端共价连接的il10m1。(il10m1)2‑
fc形式的例示性蛋白质是xenp25236,其序列在图26中描绘。在另一个实施方案中,fc
‑
(il10m1)2形式(图23b中描绘的卡通示意图)包含两个相同的单体,其中每个单体包含与同源二聚体fc 链的c端共价连接的il10m1。fc
‑
(il10m1)2形式的例示性蛋白质是xenp25237,其序列在图27和序列表中的相应seq id no中描绘。
[0456]
在另一个实施方案中,(il10m1)2‑
异源fc形式(图23c中描绘的卡通示意图)包含:第一单体,其包含与进一步共价连接(任选地通过接头)至第一异源二聚体fc链的n端的第二 il10m1共价连接(任选地通过接头)的第一il10m1;以及第二单体,其包含为“仅fc”或“空 fc”的互补的第二异源二聚体fc链。(il10m1)2
‑
异源fc形式的例示性蛋白质是xenp26887,其序列在图28和序列表中的相应seq id no中描绘。
[0457]
d.分裂il10
‑
fc融合体
[0458]
在另一方面,代替像在il10m1中那样在il10(a
‑
d)与il10(e
‑
f)结构域之间使用gly
‑ꢀ
ser接头,通过将il
‑
10单体“分裂”成其两个结构域(即hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f),其序列分别描绘于图15a
‑
15b以及seq id no:21和seq id no:22中)生成生物活性il
‑
10单体,并且il
‑
10结构域的二聚化将提供在本文称为“分裂il10”的生物活性分子。因此,本公开提供在本文中称为分裂il10
‑
fc融合体的形式(图29a
‑
29d中描绘的卡通示意图)。
[0459]
在一些实施方案中,这种形式的hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f)结构域包含氨基酸序列,分别诸如但不限于seq id no:21和seq id no:22以及本文所述的任何hil
‑
10(a
‑
d)和hil
‑ꢀ
10(e
‑
f)结构域变体(例如,图55、57、64、67、70a
‑
70b和73、相应的seq id no以及序列表中所示的那些)。
[0460]
一个实施方案是分裂il10
‑
异源fc形式(图29a中描绘的卡通示意图),其包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的hil
‑
10(a
‑
d);以及第二单体,其包含与互补的第二异源二聚体fc链的n端共价连接的hil
‑
10(e
‑
f),使得异源二聚体fc链的二聚化迫使il
‑
10结构域二聚化。分裂il10
‑
异源fc形式的例示性蛋白质包括xenp25242、 xenp25243和xenp25244,其序列在图30和序列表中的相应seq id no中描绘。
[0461]
在另一个实施方案中,异源fc
‑
分裂il10形式(图29b中描绘的卡通示意图)包含:第一单体,其包含与第一异源二聚体fc链的c端共价连接(任选地通过接头)的hil
‑
10(a
‑
d)结构域;以及第二单体,其包含与互补的第二异源二聚体fc链的c端共价连接(任选地通过接头) 的hil
‑
10(e
‑
f)结构域。异源fc
‑
分裂il10形式的例示性蛋白质包括xenp25879,其序列在图31和序列表中的相应seq id no中描绘。
[0462]
vii.本发明的有用靶向形式
[0463]
如图87、88、89和90中所示,存在许多本发明的靶向性il
‑
10融合蛋白的有用形式。通常,本发明的融合蛋白具有若干组分:il
‑
10组分、fc组分和抗原结合结构域,它们全部可以采用与本文所概述的形式不同的形式,并且其中的每一者都可以以任何配置与其它组分组合。应当理解,本文所述的任何形式的il
‑
10组分可包括在位置109处具有组氨酸或在位置 109处具有亮氨酸的il
‑
10。il
‑
10组分还可包括本文描述的和本领域已知的任何il
‑
10单体、 il10m1、scil10和分裂il10变体。如上所述,此类靶向性il
‑
10融合蛋白的fc结构域包括促进fc结构域的异源二聚化的修饰。
[0464]
a.靶向性il10
‑
fc融合体形式
[0465]
图87a
‑
图87d描绘了基于il10
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性il10
‑
fc”类别。图88a
‑
图88e描绘了基于scil10
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性scil10
‑
fc”类别。图89a
‑
图89m描绘了基于il10m1
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性il10m1
‑
fc”类别。图90a
‑
图90h 描绘了基于分裂il10
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性分裂 il10
‑
fc”类别。
[0466]
在一些实施方案中,本发明的靶向性il
‑
10fc融合蛋白包含抗原结合结构域,诸如fab 或scfv。在一些实施方案中,“mab
‑
il10”形式(参见,例如,图87a)包含两个相同的单体,每个单体包含与同源二聚体fc链的n端共价连接的抗原结合结构域,所述同源二聚体fc链通过c端任选地通过接头与il
‑
10单体共价连接。在一些实施方案中,“il10
‑
mab”形式(参见,例如,图87b)包含两个相同的单体,每个单体包含任选地通过接头与抗原结合结构域共价连接的il
‑
10单体,所述抗原结合结构域与同源二聚体fc链的n端共价连接。在一些实施方案中,“mab
‑
中心
‑
il10”形式(参见,例如,图87c)包含两个相同的单体,每个单体包含与il
‑
10单体共价连接的抗原结合结构域,所述il
‑
10单体任选地通过接头与同源二聚体fc 链共价连接。应注意,虽然抗原结合结构域被描绘为fab,但抗原结合结构域可以是如本文所定义的任何抗原结合分子,诸如scfv。在一些实施方案中,“抗x x il10
‑
异源fc”形式(参见,例如,图87d)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头)的第一il
‑
10单体,并且第二il
‑
10单体单独转染,以与第一il
‑
10单体形成非共价相互作用。应当理解,本文所述的任何形式的抗原结合结构域,包括图87a
‑
87d中所示的那些,可针对任何合适的抗原,包括但不限于cd8、nkg2d、pd
‑
1和tigit。在一些实施方案中,本文所述的任何il
‑
10单体或其变体可以用于靶向性il10
‑
fc形式。
[0467]
在一些实施方案中,本文所述形式的cd8抗原结合结构域包含图92中描绘的可变重链和可变轻链对以及序列表中的相应seq id no的序列。cd8 abd的有用的可变重链和可变轻链对的非限制性实例包括okt8_h2_l1和1c11b3_h1_l1。
[0468]
在一些实施方案中,本文所述形式的nkg2d抗原结合结构域包含图93a
‑
93c中描绘的可变重链和可变轻链对以及序列表中的相应seq id no的序列。nkg2d abd的有用的可变重链和可变轻链对的非限制性实例包括ms[nkg2d]h0_l0、1d7b4[nkg2d]_h1_l1、kyk
‑ꢀ
1.0[nkg2d]_h1_l1、kyk
‑
2.0[nkg2d]_h0_l0、11b2d10[nkg2d]_h0_l0、 6e5a7[nkg2d]_h0_l0、6h7e7[nkg2d]_h0_l0、mab e[nkg2d]_h1_l1、 16f31[nkg2d]_h1_l1、mab d[nkg2d]_h1_l1、1d7b4[nkg2d]_h1_l1、maba[nkg2d]_h1_l1、mab a[nkg2d]_h2_l1、mab a[nkg2d]_h1_l2、maba[nkg2d]_h2_l2、mab b[nkg2d]_h1_l1、mab b[nkg2d]_h1_l1.1、mabb[nkg2d]_h1_l2、mab b[nkg2d]_h2_l1、mab b[nkg2d]_h2_l1.1、mabb[nkg2d]_h2_l2、mab b[nkg2d]_h3_l1、mab b[nkg2d]_h3_l1.1、mabb[nkg2d]_h3_l2、mab c[nkg2d]_h1_l1、mab c[nkg2d]_h2_l1、mabc[nkg2d]_h1_l2以及mab c[nkg2d]_h2_l2。
[0469]
在一些实施方案中,本文所述形式的pd
‑
1抗原结合结构域包含图100a
‑
100g中描绘的可变重链和可变轻链对以及序列表中的相应seq id no的序列。pd
‑
1abd的有用的可变重链和可变轻链对的非限制性实例包括1c11[pd
‑
1]_h0l0、1c11[pd
‑
1]_h3l3、1c11[pd
‑ꢀ
1]_h3.240_l3.148、1c11[pd
‑
1]_h3.241_l3.148、1c11[pd
‑
1]_h3.234_l3.144、1c11[pd
‑ꢀ
1]_h3.241_l3.92、1c11[pd
‑
1]_h3.303_l3.152、1c11_h3.329_l3.220、1c11_h3.328_l3.152、派姆单抗可变重链和可变轻链、纳武单抗可变重链和可变轻链、皮地利珠单抗可变重链和可变轻链、mk
‑
3475可变重链和可变轻链、bap049克隆e可变重链和可变轻链、bap049克隆 b可变重链和可变轻链、h7798n[pd
‑
1]可变重链和可变轻链、h1h3 var 6[pd
‑
1]可变重链和可变轻链、ape2058[pd
‑
1]可变重链和可变轻链、h005
‑
1[pd
‑
1]可变重链和可变轻链、317
‑ꢀ
4b6[pd
‑
1]可变重链和可变轻链、326
‑
4a3[pd
‑
1]可变重链和可变轻链、hpd
‑
1mab 7(1.2)[pd
‑ꢀ
1]可变重链和可变轻链、克隆38[pd
‑
1]可变重链和可变轻链、克隆39[pd
‑
1]可变重链和可变轻链、克隆41[pd
‑
1]可变重链和可变轻链、克隆48[pd
‑
1]可变重链和可变轻链、pd1
‑
17[pd
‑ꢀ
1]可变重链和可变轻链、pd1
‑
28[pd
‑
1]可变重链和可变轻链、pd1
‑
33[pd
‑
1]可变重链和可变轻链、pd1
‑
35[pd
‑
1]可变重链和可变轻链、lopd180可变重链和可变轻链、ab948可变重链和可变轻链、人源化eh
‑
12.2h7[pd
‑
1]可变重链和可变轻链、rg1h10可变重链和可变轻链、 rg1h10
‑
h2a
‑
22
‑
1s可变重链和可变轻链、rg1h10
‑
h2a
‑
27
‑
2s可变重链和可变轻链、 rg1h10
‑
3c可变重链和可变轻链、rg1h10
‑
16c可变重链和可变轻链、rg1h10
‑
17c可变重链和可变轻链、rg1h10
‑
19c可变重链和可变轻链、rg1h10
‑
21c可变重链和可变轻链、 rg1h10
‑
23c2可变重链和可变轻链、mab7[pd
‑
1]以及pd1ab
‑
6[pd
‑
1]可变重链和可变轻链。
[0470]
在一些实施方案中,本文所述形式的非竞争性pd
‑
1抗原结合结构域包含图104a
‑
104b 中描绘的可变重链和可变轻链对以及序列表中的相应seq id no的序列。非竞争性pd
‑
1abd 的有用的可变重链和可变轻链对的非限制性实例包括mab a[pd
‑
1]_h1_l1、mab b[pd
‑ꢀ
1]_h1_l1、mab c[pd
‑
1]_h1_l1、mab c[pd
‑
1]_h1_l1.1、mab c[pd
‑
1]_h1_l1.3、mab c[pd
‑ꢀ
1]_h1_l1.45、mab c[pd
‑
1]_h1_l1.117、mab c[pd
‑
1]_h1_l1.129、mab c[pd
‑
1]_h1_l1.135、 mab c[pd
‑
1]_h1_l1.136、mab c[pd
‑
1]_h1_l1.140、mab c[pd
‑
1]_h1_l2、mab c[pd
‑ꢀ
1]_h1.19_l1、mab c[pd
‑
1]_h1.19_l1.1、mab c[pd
‑
1]_h1.19_l1.3、mab c[pd
‑ꢀ
1]_h1.19_l1.45、mab c[pd
‑
1]_h1.19_l1.117、mab c[pd
‑
1]_h1.19_l1.129、mab c[pd
‑ꢀ
1]_h1.19_l1.135、mab c[pd
‑
1]_h1.19_l1.136、mab c[pd
‑
1]_h1.19_l1.140、mab c[pd
‑ꢀ
1]_h1.19_l2、mab c[pd
‑
1]_h1.48_l1、mab c[pd
‑
1]_h1.48_l1.1、mab c[pd
‑
1]_h1.48_l1.3、 mab c[pd
‑
1]_h1.48_l1.45、mab c[pd
‑
1]_h1.48_l1.117、mab c[pd
‑
1]_h1.48_l1.129、mabc[pd
‑
1]_h1.48_l1.135、mab c[pd
‑
1]_h1.48_l1.136、mab c[pd
‑
1]_h1.48_l1.140、mab c[pd
‑ꢀ
1]_h1.48_l2、mab c[pd
‑
1]_h1.125_l1、mab c[pd
‑
1]_h1.125_l1.1、mab c[pd
‑ꢀ
1]_h1.125_l1.3、mab c[pd
‑
1]_h1.125_l1.45、mab c[pd
‑
1]_h1.125_l1.117、mab c[pd
‑ꢀ
1]_h1.125_l1.129、mab c[pd
‑
1]_h1.125_l1.135、mab c[pd
‑
1]_h1.125_l1.136、mab c[pd
‑ꢀ
1]_h1.125_l1.140、mab c[pd
‑
1]_h1.125_l2、mab c[pd
‑
1]_h1.130_l1、mab c[pd
‑ꢀ
1]_h1.130_l1.1、mab c[pd
‑
1]_h1.130_l1.3、mab c[pd
‑
1]_h1.130_l1.45、mab c[pd
‑ꢀ
1]_h1.130_l1.117、mab c[pd
‑
1]_h1.130_l1.129、mab c[pd
‑
1]_h1.130_l1.135、mab c[pd
‑ꢀ
1]_h1.130_l1.136、mab c[pd
‑
1]_h1.130_l1.140、mab c[pd
‑
1]_h1.130_l2、mab c[pd
‑ꢀ
1]_h1.132_l1、mab c[pd
‑
1]_h1.132_l1.1、mab c[pd
‑
1]_h1.132_l1.3、mab c[pd
‑ꢀ
1]_h1.132_l1.45、mab c[pd
‑
1]_h1.132_l1.117、mab c[pd
‑
1]_h1.132_l1.129、mab c[pd
‑ꢀ
1]_h1.132_l1.135、mab c[pd
‑
1]_h1.132_l1.136、mab c[pd
‑
1]_h1.132_l1.140、mab c[pd
‑ꢀ
1]_h1.132_l2、mab c[pd
‑
1]_h1.169_l1、mab c[pd
‑
1]_h1.169_l1.1、mab c[pd
‑ꢀ
1]_h1.169_l1.3、mab c[pd
‑
1]_h1.169_l1.45、mab c[pd
‑
1]_h1.169_l1.117、mab c[pd
‑ꢀ
1]_
h1.169_l1.129、mab c[pd
‑
1]_h1.169_l1.135、mab c[pd
‑
1]_h1.169_l1.136、mab c[pd
‑ꢀ
1]_h1.169_l1.140、mab c[pd
‑
1]_h1.169_l2、mab c[pd
‑
1]_h1.175_l1、mab c[pd
‑ꢀ
1]_h1.175_l1.1、mab c[pd
‑
1]_h1.175_l1.3、mab c[pd
‑
1]_h1.175_l1.45、mab c[pd
‑ꢀ
1]_h1.175_l1.117、mab c[pd
‑
1]_h1.175_l1.129、mab c[pd
‑
1]_h1.175_l1.135、mab c[pd
‑ꢀ
1]_h1.175_l1.136、mab c[pd
‑
1]_h1.175_l1.140、mab c[pd
‑
1]_h1.175_l2、mab c[pd
‑ꢀ
1]_h2_l1、mab c[pd
‑
1]_h2_l1.1、mab c[pd
‑
1]_h2_l1.3、mab c[pd
‑
1]_h2_l1.45、mabc[pd
‑
1]_h2_l1.117、mab c[pd
‑
1]_h2_l1.129、mab c[pd
‑
1]_h2_l1.135、mab c[pd
‑ꢀ
1]_h2_l1.136、mab c[pd
‑
1]_h2_l1.140以及mab c[pd
‑
1]_h2_l2。
[0471]
在一些实施方案中,本文所述形式的tigit抗原结合结构域包含图122a
‑
122c中描绘的可变重链和可变轻链对以及序列表中的相应seq id no的序列。tigit abd的有用的可变重链和可变轻链对的非限制性实例包括2a5b4[tigit]_h1_l1、genentech_4.1d3.q1e(替拉格鲁单抗)_h0_l0、10a7[tigit]可变重链和可变轻链、1f4[tigit]可变重链和可变轻链、 4.1d3[tigit]可变重链和可变轻链、hu14d7 vh1_vl1、hu14d7 vh1_vl2、hu14d7 vh1_vl3、hu14d7 vh2_vl1、hu14d7 vh2_vl2、hu14d7 vh2_vl3、hu14d7 vh3_vl1、 hu14d7 vh3_vl2、hu14d7 vh3_vl3、hu26b10 vh1_vl1、hu26b10 vh1_vl2、hu26b10 vh1_l3、hu26b10 vh2_vl1、hu26b10 vh2_vl2、hu26b10 vh2_vl3、hu26b10 vh3_vl1、 hu26b10 vh3_l2、hu26b10 vh3_vl3、meb125.31c6.a1.205 vh4/vl1、 meb125.31c6.a1.205 vh5/vl4、meb125.31c6.a1.205 vh5/vl3、15a6可变重链和可变轻链、22g2可变重链和可变轻链、11g11可变重链和可变轻链、10d7可变重链和可变轻链、313r19可变重链和可变轻链,以及艾替吉利单抗可变重链和可变轻链。
[0472]
b.靶向性scil10
‑
fc融合体形式
[0473]
在另外的实施方案中,本公开提供了诸如图88a
‑
图88e中所示的那些的形式。图88描绘了基于scil10
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性scil10
‑
fc”类别。在一些实施方案中,“抗x x scil10
‑
异源fc”形式(参见,例如,图88a)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头)的scil
‑
10。在一些实施方案中,“抗x x异源fc
‑
scil10”形式(参见,例如,图88b)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含通过c端与scil
‑
10共价连接(任选地通过结构域接头)的第二异源二聚体fc链。在一些实施方案中,“抗x x异源fc
‑
scil10”形式包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域,所述第一异源二聚体fc链通过c端与scil
‑
10共价连接(任选地通过结构域接头);以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc 链。在一些实施方案中,“(抗x)2‑
异源fc
‑
scil10”形式(参见,例如,图88c)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的抗原结合结构域,所述第二异源二聚体fc链通过c端与scil
‑
10共价连接(任选地通过结构域接头)。在一些实施方案中,“(抗x)2‑
中心
‑ꢀ
scil10”形式(参见,例如,图88d)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与scil
‑
10共价连接的抗原结合结构域,所述scil
‑
10与第二异源二聚体fc链的n端共价连接(任选地通过接头)。在一些实施方案中,“scil10
‑
(抗x)2‑
异
源fc”形式(参见,例如,图88e)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含任选地通过接头与抗原结合结构域共价连接的scil
‑
10,所述抗原结合结构域与第二异源二聚体fc链的n端共价连接(任选地通过接头)。应注意,虽然抗原结合结构域被描绘为fab,但抗原结合结构域可以是如本文所定义的任何抗原结合分子,诸如scfv。应当理解,本文所述的任何形式的抗原结合结构域,包括图88a
‑
88e中所示的那些,可针对任何合适的抗原,包括但不限于cd8、 nkg2d、pd
‑
1和tigit。
[0474]
在一些实施方案中,靶向性il
‑
10fc融合蛋白(例如,靶向性抗cd8 abd x scil10异源二聚体fc融合蛋白)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗 cd8抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的scil
‑ꢀ
10。在一些实施方案中,scil
‑
10通过结构域接头与第二fc链共价连接。在一些实施方案中,抗cd8 abd包含1c11b3_h1_l1的vh和vl组合。在一些实施方案中,融合蛋白的scil
‑ꢀ
10包含图15c的序列或本文所述的任何scil10单体变体。在一些实施方案中,scil10的每个单体包含如图39a
‑
39g、55、56a
‑
56c、57、58、64和65a
‑
65b中列出的氨基酸序列。在一些实施方案中,scil选自由以下组成的组:huil10_huil10、huil10.22_huil10.22、 huil10.38_huil10.38、huil10.64_huil10.64、huil10.102_huil10.102、huil10.104_huil10.104、 huil10.105_huil10.105以及huil10.107_huil10.107,如附图所描绘。在一些实施方案中,scil 单体选自由以下组成的组:huil10、huil10.22、huil10.38、huil10.64、huil10.102、huil10.104、 huil10.105以及huil10.107,如附图所描绘。
[0475]
在一些实施方案中,靶向性抗cd8 abd x scil10异源二聚体fc融合蛋白包含:第一单体,其包含scil10、结构域接头和第一fc结构域;第二单体,其包含抗cd8 vh结构域和第二fc结构域;以及抗cd8 vl结构域,使得vh结构域和vl结构域形成抗cd8 abd。在一些情况下,scil10包含两个il
‑
10单体。在一些情况下,scil10包含通过接头共价连接的两个il
‑
10单体。在一些实施方案中,靶向性il
‑
10fc融合蛋白是xenp25415、xenp25791、 xenp25794、xenp26883和xenp26879,如图96a
‑
图96c和图98以及与序列表中的seqid no相对应的序列中所描绘。
[0476]
在一些实施方案中,靶向性il
‑
10fc融合蛋白(例如,靶向性抗nkg2d abd x scil10异源二聚体fc融合蛋白)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗 nkg2d抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的 scil
‑
10。在一些实施方案中,scil
‑
10通过结构域接头与第二fc链共价连接。在一些实施方案中,抗nkg2d abd包含选自由以下组成的组的任一个的vh和vl组合:ms[nkg2d] h0_l0、1d7b4[nkg2d]_h1_l1、kyk
‑
1.0[nkg2d]_h1_l1、kyk
‑
2.0[nkg2d]_h0_l0、 11b2d10[nkg2d]_h0_l0、6e5a7[nkg2d]_h0_l0、6h7e7[nkg2d]_h0_l0、mabe[nkg2d]_h1_l1、16f31[nkg2d]_h1_l1、mab d[nkg2d]_h1_l1、 1d7b4[nkg2d]_h1_l1、mab a[nkg2d]_h1_l1、mab a[nkg2d]_h2_l1、maba[nkg2d]_h1_l2、mab a[nkg2d]_h2_l2、mab b[nkg2d]_h1_l1、mabb[nkg2d]_h1_l1.1、mab b[nkg2d]_h1_l2、mab b[nkg2d]_h2_l1、mabb[nkg2d]_h2_l1.1、mab b[nkg2d]_h2_l2、mab b[nkg2d]_h3_l1、mabb[nkg2d]_h3_l1.1、mab b[nkg2d]_h3_l2、mab c[nkg2d]_h1_l1、mabc[nkg2d]_h2_l1、mab c[nkg2d]_h1_l2以及mab c[nkg2d]_h2_l2。在一些实施方案中,融合蛋白的scil
‑
10包含图15c的序列或本文所述的任何scil10单体变体。在一些实施方案中,scil10的每个单体包含如图39a
‑
39g、55、56a
‑
56c、57、58、64
和65a
‑
65b中列出的氨基酸序列。在一些实施方案中,scil选自由以下组成的组:huil10_huil10、 huil10.22_huil10.22、huil10.38_huil10.38、huil10.64_huil10.64、huil10.102_huil10.102、 huil10.104_huil10.104、huil10.105_huil10.105以及huil10.107_huil10.107,如附图所描绘。在一些实施方案中,scil单体选自由以下组成的组:huil10、huil10.22、huil10.38、huil10.64、 huil10.102、huil10.104、huil10.105以及huil10.107,如附图所描绘。
[0477]
在一些实施方案中,靶向性抗nkg2d abd x scil10异源二聚体fc融合蛋白包含:第一单体,其包含scil10、结构域接头和第一fc结构域;第二单体,其包含抗nkg2d vh结构域和第二fc结构域;以及抗nkg2d vl结构域,使得vh结构域和vl结构域形成抗nkg2dabd。在一些情况下,scil10包含两个il
‑
10单体。在一些情况下,scil10包含通过接头共价连接的两个il
‑
10单体。在一些实施方案中,靶向nkg2d的il
‑
10fc融合蛋白是 xenp25952、xenp30526、xenp30527、xenp30528和xenp31819,如图97a
‑
图97c以及与序列表中的seq id no相对应的序列中所描绘。
[0478]
在一些实施方案中,靶向性il
‑
10fc融合蛋白(例如,靶向性抗pd
‑
1abd x scil10异源二聚体fc融合蛋白)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗 pd
‑
1抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的 scil
‑
10。在某些实施方案中,“抗pd
‑
1x异源fc
‑
scil10”形式的靶向性il
‑
10fc融合蛋白包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗pd
‑
1抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的c端共价连接的scil
‑
10。在一些实施方案中,scil
‑
10通过结构域接头与第二fc链共价连接。在一些实施方案中,抗pd
‑
1abd包含选自由以下组成的组的任一个的vh和vl组合:1c11[pd
‑
1]_h0l0、1c11[p,d
‑
1]_h3l3、 1c11[pd
‑
1]_h3.240_l3.148、1c11[pd
‑
1]_h3.241_l3.148、1c11[pd
‑
1]_h3.234_l3.144、 1c11[pd
‑
1]_h3.241_l3.92、1c11[pd
‑
1]_h3.303_l3.152、1c11_h3.329_l3.220、 1c11_h3.328_l3.152、派姆单抗可变重链和可变轻链、纳武单抗可变重链和可变轻链、皮地利珠单抗可变重链和可变轻链、mk
‑
3475可变重链和可变轻链、bap049克隆e可变重链和可变轻链、bap049克隆b可变重链和可变轻链、h7798n[pd
‑
1]可变重链和可变轻链、h1h3 var 6[pd
‑
1]可变重链和可变轻链、ape2058[pd
‑
1]可变重链和可变轻链、h005
‑
1[pd
‑
1]可变重链和可变轻链、317
‑
4b6[pd
‑
1]可变重链和可变轻链、326
‑
4a3[pd
‑
1]可变重链和可变轻链、 hpd
‑
1mab 7(1.2)[pd
‑
1]可变重链和可变轻链、克隆38[pd
‑
1]可变重链和可变轻链、克隆39[pd
‑
1]可变重链和可变轻链、克隆41[pd
‑
1]可变重链和可变轻链、克隆48[pd
‑
1]可变重链和可变轻链、pd1
‑
17[pd
‑
1]可变重链和可变轻链、pd1
‑
28[pd
‑
1]可变重链和可变轻链、pd1
‑ꢀ
33[pd
‑
1]可变重链和可变轻链、pd1
‑
35[pd
‑
1]可变重链和可变轻链、lopd180可变重链和可变轻链、ab948可变重链和可变轻链、人源化eh
‑
12.2h7[pd
‑
1]可变重链和可变轻链、rg1h10 可变重链和可变轻链、rg1h10
‑
h2a
‑
22
‑
1s可变重链和可变轻链、rg1h10
‑
h2a
‑
27
‑
2s可变重链和可变轻链、rg1h10
‑
3c可变重链和可变轻链、rg1h10
‑
16c可变重链和可变轻链、 rg1h10
‑
17c可变重链和可变轻链、rg1h10
‑
19c可变重链和可变轻链、rg1h10
‑
21c可变重链和可变轻链、rg1h10
‑
23c2可变重链和可变轻链、mab7[pd
‑
1]以及pd1ab
‑
6[pd
‑
1]可变重链和可变轻链。在一些实施方案中,融合蛋白的scil
‑
10包含图15c的序列或本文所述的任何scil10单体变体。在一些实施方案中,scil10的每个单体包含如图39a
‑
39g、55、56a
‑ꢀ
56c、57、58、64和
65a
‑
65b中列出的氨基酸序列。在一些实施方案中,scil选自由以下组成的组:huil10_huil10、huil10.22_huil10.22、huil10.38_huil10.38、huil10.64_huil10.64、 huil10.102_huil10.102、huil10.104_huil10.104、huil10.105_huil10.105以及 huil10.107_huil10.107,如附图所描绘。在一些实施方案中,scil单体选自由以下组成的组: huil10、huil10.22、huil10.38、huil10.64、huil10.102、huil10.104、huil10.105以及huil10.107,如附图所描绘。
[0479]
在一些实施方案中,靶向性抗pd
‑
1abd x scil10异源二聚体fc融合蛋白包含:第一单体,其从n端到c端包含:scil10、结构域接头和第一fc结构域;第二单体,其从n端到 c端包含:抗pd
‑
1vh结构域和第二fc结构域;以及抗pd
‑
1vl结构域,使得vh结构域和vl结构域形成抗pd
‑
1abd。在某些实施方案中,靶向性抗pd
‑
1abd x scil10异源二聚体fc融合蛋白包含:第一单体,其从n端到c端包含:第一fc结构域、结构域接头和scil10;第二单体,其从n端到c端包含:抗pd
‑
1vh结构域和第二fc结构域;以及抗pd
‑
1vl结构域,使得vh结构域和vl结构域形成抗pd
‑
1abd。在一些情况下,scil10包含两个il
‑ꢀ
10单体。在一些情况下,scil10包含通过接头共价连接的两个il
‑
10单体。在一些实施方案中,靶向pd
‑
1的il
‑
10fc融合蛋白是xenp25953、xenp27830、xenp27831、xenp31269、 xenp31270、xenp31271和xenp31272,如图101、图102a
‑
102c和图103以及与序列表中的seq id no相对应的序列中所描绘。
[0480]
在一些实施方案中,靶向性il
‑
10fc融合蛋白(例如,靶向性的非竞争性抗pd
‑
1abd xscil10异源二聚体fc融合蛋白)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的非竞争性抗pd
‑
1抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的scil
‑
10。在某些实施方案中,“抗pd
‑
1x异源fc
‑
scil10”形式的靶向性il
‑ꢀ
10fc融合蛋白包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的非竞争性抗pd
‑
1抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的c端共价连接的 scil
‑
10。在一些实施方案中,scil
‑
10通过结构域接头与第二fc链共价连接。在一些实施方案中,抗pd
‑
1abd包含选自由以下组成的组的任一个的vh和vl组合:mab a[pd
‑ꢀ
1]_h1_l1、mab b[pd
‑
1]_h1_l1、mab c[pd
‑
1]_h1_l1、mab c[pd
‑
1]_h1_l1.1、mab c[pd
‑ꢀ
1]_h1_l1.3、mab c[pd
‑
1]_h1_l1.45、mab c[pd
‑
1]_h1_l1.117、mab c[pd
‑
1]_h1_l1.129、 mab c[pd
‑
1]_h1_l1.135、mab c[pd
‑
1]_h1_l1.136、mab c[pd
‑
1]_h1_l1.140、mab c[pd
‑ꢀ
1]_h1_l2、mab c[pd
‑
1]_h1.19_l1、mab c[pd
‑
1]_h1.19_l1.1、mab c[pd
‑
1]_h1.19_l1.3、 mab c[pd
‑
1]_h1.19_l1.45、mab c[pd
‑
1]_h1.19_l1.117、mab c[pd
‑
1]_h1.19_l1.129、mabc[pd
‑
1]_h1.19_l1.135、mab c[pd
‑
1]_h1.19_l1.136、mab c[pd
‑
1]_h1.19_l1.140、mab c[pd
‑ꢀ
1]_h1.19_l2、mab c[pd
‑
1]_h1.48_l1、mab c[pd
‑
1]_h1.48_l1.1、mab c[pd
‑
1]_h1.48_l1.3、 mab c[pd
‑
1]_h1.48_l1.45、mab c[pd
‑
1]_h1.48_l1.117、mab c[pd
‑
1]_h1.48_l1.129、mabc[pd
‑
1]_h1.48_l1.135、mab c[pd
‑
1]_h1.48_l1.136、mab c[pd
‑
1]_h1.48_l1.140、mab c[pd
‑ꢀ
1]_h1.48_l2、mab c[pd
‑
1]_h1.125_l1、mab c[pd
‑
1]_h1.125_l1.1、mab c[pd
‑ꢀ
1]_h1.125_l1.3、mab c[pd
‑
1]_h1.125_l1.45、mab c[pd
‑
1]_h1.125_l1.117、mab c[pd
‑ꢀ
1]_h1.125_l1.129、mab c[pd
‑
1]_h1.125_l1.135、mab c[pd
‑
1]_h1.125_l1.136、mab c[pd
‑ꢀ
1]_h1.125_l1.140、mab c[pd
‑
1]_h1.125_l2、mab c[pd
‑
1]_h1.130_l1、mab c[pd
‑ꢀ
1]_h1.130_l1.1、mab c[pd
‑
1]_h1.130_l1.3、mab c[pd
‑
1]_h1.130_l1.45、mab c[pd
‑ꢀ
1]_h1.130_l1.117、mab c[pd
‑
1]_h1.130_l1.129、mab c[pd
‑
1]_h1.130_l1.135、mab c[pd
‑ꢀ
1]_h1.130_l1.136、mab c[pd
‑
1]_h1.130_l1.140、mab c[pd
‑
1]_h1.130_l2、mab c[pd
‑ꢀ
1]_h1.132_l1、mab c[pd
‑
1]_h1.132_l1.1、mab c[pd
‑
1]_h1.132_l1.3、mab c[pd
‑ꢀ
1]_h1.132_l1.45、mab c[pd
‑
1]_h1.132_l1.117、mab c[pd
‑
1]_h1.132_l1.129、mab c[pd
‑ꢀ
1]_h1.132_l1.135、mab c[pd
‑
1]_h1.132_l1.136、mab c[pd
‑
1]_h1.132_l1.140、mab c[pd
‑ꢀ
1]_h1.132_l2、mab c[pd
‑
1]_h1.169_l1、mab c[pd
‑
1]_h1.169_l1.1、mab c[pd
‑ꢀ
1]_h1.169_l1.3、mab c[pd
‑
1]_h1.169_l1.45、mab c[pd
‑
1]_h1.169_l1.117、mab c[pd
‑ꢀ
1]_h1.169_l1.129、mab c[pd
‑
1]_h1.169_l1.135、mab c[pd
‑
1]_h1.169_l1.136、mab c[pd
‑ꢀ
1]_h1.169_l1.140、mab c[pd
‑
1]_h1.169_l2、mab c[pd
‑
1]_h1.175_l1、mab c[pd
‑ꢀ
1]_h1.175_l1.1、mab c[pd
‑
1]_h1.175_l1.3、mab c[pd
‑
1]_h1.175_l1.45、mab c[pd
‑ꢀ
1]_h1.175_l1.117、mab c[pd
‑
1]_h1.175_l1.129、mab c[pd
‑
1]_h1.175_l1.135、mab c[pd
‑ꢀ
1]_h1.175_l1.136、mab c[pd
‑
1]_h1.175_l1.140、mab c[pd
‑
1]_h1.175_l2、mab c[pd
‑ꢀ
1]_h2_l1、mab c[pd
‑
1]_h2_l1.1、mab c[pd
‑
1]_h2_l1.3、mab c[pd
‑
1]_h2_l1.45、mabc[pd
‑
1]_h2_l1.117、mab c[pd
‑
1]_h2_l1.129、mab c[pd
‑
1]_h2_l1.135、mab c[pd
‑ꢀ
1]_h2_l1.136、mab c[pd
‑
1]_h2_l1.140和mab c[pd
‑
1]_h2_l2。
[0481]
在一些实施方案中,靶向性的非竞争性抗pd
‑
1abd x scil10异源二聚体fc融合蛋白包含:第一单体,其从n端到c端包含:scil10、结构域接头和第一fc结构域;第二单体,其从n端到c端包含:抗pd
‑
1vh结构域和第二fc结构域;以及抗pd
‑
1vl结构域,使得 vh结构域和vl结构域形成非竞争性抗pd
‑
1abd。在某些实施方案中,靶向性的非竞争性抗pd
‑
1abd x scil10异源二聚体fc融合蛋白包含:第一单体,其从n端到c端包含:第一 fc结构域、结构域接头和scil10;第二单体,其从n端到c端包含:抗pd
‑
1vh结构域和第二fc结构域;以及抗pd
‑
1vl结构域,使得vh结构域和vl结构域形成非竞争性抗pd
‑ꢀ
1abd。在一些情况下,scil10包含两个il
‑
10单体。在一些情况下,scil10包含通过接头共价连接的两个il
‑
10单体。在一些实施方案中,非竞争性的靶向pd
‑
1的il
‑
10fc融合蛋白是 xenp30520、xenp30521、xenp30522、xenp31266、xenp31267和xenp31268,如图106a
‑ꢀ
106c以及与序列表中的seq id no相对应的序列中所描绘。xenp30520也称为αpd
‑
1(mabc_h1_l1.1)x huil10.104。xenp30521也称为αpd
‑
1(mab c_h1_l1.1)x huil10.105。 xenp30522也称为αpd
‑
1(mab c_h1_l1.1)x huil10.107。
[0482]
在一些实施方案中,靶向性il
‑
10fc融合蛋白(例如,靶向性的非竞争性抗tigit abd xscil10异源二聚体fc融合蛋白)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的非竞争性抗tigit抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的 n端共价连接的scil
‑
10。在某些实施方案中,“抗tigit x异源fc
‑
scil10”形式的靶向性 il
‑
10fc融合蛋白包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗tigit 抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的c端共价连接的scil
‑
10。在一些实施方案中,scil
‑
10通过结构域接头与第二fc链共价连接。在一些实施方案中,抗 tigit abd包含选自由以下组成的组的任一个的vh和vl组合:2a5b4[tigit]_h1_l1、 genentech_4.1d3.q1e(替拉格鲁单抗)_h0_l0、10a7[tigit]可变重链和可变轻链、1f4[tigit] 可变重链和可变轻链、4.1d3[tigit]可变重链和可变轻链、hu14d7 vh1_vl1、hu14d7 vh1_vl2、hu14d7 vh1_vl3、hu14d7 vh2_vl1、hu14d7 vh2_vl2、hu14d7 vh2_vl3、 hu14d7 vh3_vl1、hu14d7 vh3_vl2、hu14d7 vh3_vl3、hu26b10 vh1_vl1、hu26b10 vh1_vl2、
hu26b10 vh1_l3、hu26b10 vh2_vl1、hu26b10 vh2_vl2、hu26b10 vh2_vl3、hu26b10 vh3_vl1、hu26b10 vh3_l2、hu26b10 vh3_vl3、meb125.31c6.a1.205 vh4/vl1、 meb125.31c6.a1.205 vh5/vl4、meb125.31c6.a1.205 vh5/vl3、15a6可变重链和可变轻链、22g2可变重链和可变轻链、11g11可变重链和可变轻链、10d7可变重链和可变轻链、 313r19可变重链和可变轻链,以及艾替吉利单抗可变重链和可变轻链。
[0483]
在一些实施方案中,靶向性抗tigit abd x scil10异源二聚体fc融合蛋白包含:第一单体,其从n端到c端包含:scil10、结构域接头和第一fc结构域;第二单体,其从n端到c端包含:抗tigit vh结构域和第二fc结构域;以及抗tigit vl结构域,使得vh结构域和vl结构域形成抗tigit abd。在某些实施方案中,靶向性抗tigit abd x scil10异源二聚体fc融合蛋白包含:第一单体,其从n端到c端包含:第一fc结构域、结构域接头和scil10;第二单体,其从n端到c端包含:抗tigit vh结构域和第二fc结构域;以及抗 tigit vl结构域,使得vh结构域和vl结构域形成非竞争性抗tigit abd。在一些情况下,scil10包含两个il
‑
10单体。在一些情况下,scil10包含通过接头共价连接的两个il
‑
10 单体。在一些实施方案中,抗tigit靶向性il
‑
10fc融合蛋白是xenp30523、xenp30524和 xenp30525,如图123a
‑
123b以及与序列表中的seq id no相对应的序列中所描绘。
[0484]
xenp30523也称为αtigit(2a5b4)x huil10.104。xenp30524也称为αtigit(2a5b4)xhuil10.105。xenp30525也称为αtigit(2a5b4)x huil10.107。
[0485]
c.靶向性il10m1
‑
fc融合体形式
[0486]
图89a
‑
图89m描绘了基于il10m1
‑
fc类别的靶向性il
‑
10融合体的另外的实施方案,本文称为“靶向性il10m1
‑
fc”类别。在一些实施方案中,“抗x x(il10m1)1‑
异源fc”形式(参见,例如,图89a)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头)的il10m1。在一些实施方案中,“抗x x异源fc
‑
(il10m1)
1”形式(参见,例如,图89b)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含通过c端与il10m1共价连接(任选地通过结构域接头)的第二异源二聚体fc链。在一些实施方案中,“抗x x异源fc
‑
(il10m1)
1”形式包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域,所述第一异源二聚体fc 链通过c端与il10m1共价连接(任选地通过结构域接头);以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链。在一些实施方案中,“(抗x)2‑
异源fc
‑ꢀ
(il10m1)
1”形式(参见,例如,图89c)包含:第一单体,其包含与第一异源二聚体fc链的n 端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的抗原结合结构域,所述第二异源二聚体fc链通过c端与il10m1共价连接(任选地通过结构域接头)。在一些实施方案中,“(il10m1)1‑
(抗x)2‑
异源fc”形式(参见,例如,图89d) 包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含任选地通过接头与抗原结合结构域共价连接的il10m1,所述抗原结合结构域与第二异源二聚体fc链的n端共价连接(任选地通过接头)。在一些实施方案中,“(抗x)2‑
中心
‑
(il10m1)
1”形式(参见,例如,图89e)包含:第一单体,其包含与第一异源二聚体fc链的 n端共价连接的抗原结合结构域;以及第二单体,其包含与il10m1共价连接的抗原结合结构域,所述il10m1与第二异源二聚体fc链的n端共价连接(任选地通过接头)。在一些实施方案中,“抗x x(il10m1)2‑
异源
fc”形式(参见,例如,图89f)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含任选地通过结构域接头与第二il10m1共价连接的第一il10m1,所述第二il10m1与第二异源二聚体 fc链的n端共价连接(任选地通过结构域接头)。在一些实施方案中,“抗x x异源fc
‑ꢀ
(il10m1)
2”形式(参见,例如,图89g)包含:第一单体,其包含与第一异源二聚体fc链的n 端共价连接的抗原结合结构域;以及第二单体,其包含通过c端与第一il10m1共价连接(任选地通过结构域接头)的第二异源二聚体fc链,所述第一il10m1任选地通过结构域接头与第二il10m1共价连接。在一些实施方案中,“抗x x异源fc
‑
(il10m1)
2”形式包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域,所述第一异源二聚体fc链通过c端与第一il10m1共价连接(任选地通过结构域接头),所述第一il10m1任选地通过结构域接头与第二il10m1共价连接;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链。在一些实施方案中,“(抗x)2‑
异源fc
‑
(il10m1)
2”形式(参见,例如,图89h)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的抗原结合结构域,所述第二异源二聚体fc链通过c端与第一il10m1共价连接(任选地通过结构域接头),所述第一il10m1任选地通过结构域接头与第二il10m1共价连接。在一些实施方案中,“(il10m1)2‑
(抗x)2‑
异源fc”形式(参见,例如,图89i)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含任选地通过接头与第二il10m1共价连接的第一il10m1,所述第二il10m1与抗原结合结构域共价连接(任选地通过结构域接头),所述抗原结合结构域与第二异源二聚体fc链的n端共价连接(任选地通过接头)。在一些实施方案中,“(抗x)2‑
中心
‑
(il10m1)
2”形式(参见,例如,图89j)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第一il10m1共价连接的抗原结合结构域,所述第一il10m1任选地通过结构域接头与第二il10m1共价连接,所述第二il10m1与第二异源二聚体fc链的n端共价连接(任选地通过接头)。在一些实施方案中,“mab
‑
(il10m1)
2”形式(参见,例如,图89k)包含两个相同的单体,每个单体包含与同源二聚体fc链的n端共价连接的抗原结合结构域,所述同源二聚体fc链通过c端任选地通过接头与il10m1共价连接。在一些实施方案中,“(il10m1)2‑
mab”形式(参见,例如,图89l)包含两个相同的单体,每个单体包含任选地通过接头与抗原结合结构域共价连接的il10m1,所述抗原结合结构域与同源二聚体fc链的n 端共价连接。在一些实施方案中,“mab
‑
中心
‑
(il10m1)
2”形式(参见,例如,图89m)包含两个相同的单体,每个单体包含与il10m1共价连接的抗原结合结构域,所述il10m1任选地通过接头与同源二聚体fc链共价连接。应注意,虽然抗原结合结构域被描绘为fab,但抗原结合结构域可以是如本文所定义的任何抗原结合分子,诸如scfv。
[0487]
在一些实施方案中,靶向性il
‑
10fc融合蛋白(例如,靶向性抗cd8 x il10m1异源二聚体fc融合蛋白)包含:与第一异源二聚体fc链的n端共价连接的抗cd8 abd;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的il10m1。在一些实施方案中,il10m1 通过结构域接头与第二异源二聚体fc链的n端共价连接。在一些实施方案中,抗cd8 abd 包含okt8_h2_l1的vh和vl组合。在一些实施方案中,抗cd8 abd包含1c11b3_h1_l1 的vh和vl组合。在一些实施方案中,靶向性il
‑
10fc融合蛋白是xenp25365或xenp25366,如图94以及与序列表中的seq id no相对应的序列中所描绘。
[0488]
在一些实施方案中,靶向性il
‑
10fc融合蛋白(例如,靶向性抗cd8 mab x(il10m1)2异源二聚体fc融合蛋白)包含两个相同的单体,每个单体包含与同源二聚体fc链的n端共价连接的抗cd8 abd,所述同源二聚体fc链通过c端与il10m1共价连接。在一些实施方案中,fc链通过接头与il10m1连接。在一些实施方案中,每个单体包含两条链,使得第一条链从n端到c端包含:抗cd8 abd的vh结构域、fc结构域、任选的接头和il10m1,并且第二条链包含抗cd9 abd的vl结构域。在一些实施方案中,抗cd8 abd包含 1c11b3_h1_l1的vh和vl组合。在一些实施方案中,靶向性il
‑
10fc融合蛋白是 xenp26172,如图95以及与序列表中的seq id no相对应的序列中所描绘。
[0489]
d.靶向性分裂il10
‑
fc融合体形式
[0490]
图90a
‑
图90h描绘了基于分裂il10
‑
fc类别的靶向性il
‑
10融合体的另外实施方案,本文称为“靶向性分裂il10
‑
fc”类别。在一些实施方案中,“抗x x(分裂il10)1‑
异源fc”形式(参见,例如,图90a)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头)的hil
‑
10(a
‑
d)结构域,以及单独转染以使其与hil
‑
10(a
‑
d)结构域非共价相互作用的hil
‑
10(e
‑
f)结构域。在一些实施方案中,“抗x x(分裂il10)1‑
异源fc”形式包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头)的hil
‑
10(e
‑
f) 结构域,以及单独转染以使其与hil
‑
10(e
‑
f)结构域非共价相互作用的hil
‑
10(a
‑
d)结构域。在一些实施方案中,“(抗x)2x异源fc
‑
(分裂il10)
1”形式(参见,例如,图90b)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域,所述第一异源二聚体fc链通过c端与hil
‑
10(a
‑
d)结构域共价连接;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的抗原结合结构域,所述第二异源二聚体fc链通过c端与hil
‑
10 (e
‑
f)结构域共价连接。在一些实施方案中,“(分裂il10)1‑
(抗x)2‑
异源fc”形式(参见,例如,图90c)包含:第一单体,其包含与抗原结合结构域共价连接的hil
‑
10(a
‑
d)结构域,所述抗原结合结构域与第一异源二聚体fc链的n端共价连接;以及第二单体,其包含与抗原结合结构域共价连接的hil
‑
10(e
‑
f)结构域,所述抗原结合结构域与第二异源二聚体fc链的n端共价连接。在一些实施方案中,“(抗x)2‑
中心
‑
(分裂il10)1‑
异源fc”形式(参见,例如,图 90d)包含:第一单体,其包含与hil
‑
10(a
‑
d)结构域共价连接的抗原结合结构域,所述hil
‑ꢀ
10(a
‑
d)结构域与第一异源二聚体fc链共价连接;以及第二单体,其包含与hil
‑
10(e
‑
f)结构域共价连接的抗原结合结构域,所述hil
‑
10(e
‑
f)结构域与第二异源二聚体fc链共价连接。在一些实施方案中,“抗x x(分裂il10)2‑
异源fc”形式(参见,例如,图90e)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二hil
‑
10(a
‑
d)结构域共价连接的第一hil
‑
10(a
‑
d)结构域,所述第二hil
‑
10(a
‑
d)结构域与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头),以及与第二hil
‑
10 (e
‑
f)结构域共价连接的第一hil
‑
10(e
‑
f)结构域,所述第二hil
‑
10(e
‑
f)结构域单独转染以使其与hil
‑
10(a
‑
d)结构域非共价相互作用。在一些实施方案中,“抗x x(分裂il10)2‑
异源fc”形式包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二hil
‑
10(e
‑
f)结构域共价连接的第一hil
‑
10(e
‑
f)结构域,所述第二hil
‑
10(e
‑
f)结构域与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头),
以及与第二hil
‑
10(a
‑
d)结构域共价连接的第一hil
‑
10(a
‑
d)结构域,所述第二hil
‑
10 (a
‑
d)结构域单独转染以使其与hil
‑
10(e
‑
f)结构域非共价相互作用。在一些实施方案中,“(抗 x)2x异源fc
‑
(分裂il10)
2”形式(参见,例如,图90f)包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域,所述第一异源二聚体fc链通过c端与共价连接至第二hil
‑
10(a
‑
d)结构域的第一hil
‑
10(a
‑
d)结构域共价连接;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接的抗原结合结构域,所述第二异源二聚体fc链通过c 端与共价连接至第二hil
‑
10(e
‑
f)结构域的第一hil
‑
10(e
‑
f)结构域共价连接。在一些实施方案中,“(分裂il10)2‑
(抗x)2‑
异源fc”形式(参见,例如,图90g)包含:第一单体,其包含与第二hil
‑
10(a
‑
d)结构域共价连接的第一hil
‑
10(a
‑
d)结构域,所述第二hil
‑
10(a
‑
d)结构域与共价连接至第一异源二聚体fc链的n端的抗原结合结构域共价连接;以及第二单体,其包含与第二hil
‑
10(e
‑
f)结构域共价连接的第一hil
‑
10(e
‑
f)结构域,所述第二hil
‑
10(e
‑
f)结构域与共价连接至第二异源二聚体fc链的n端的抗原结合结构域共价连接。在一些实施方案中,“(抗x)2‑
中心
‑
(分裂il10)2‑
异源fc”形式(参见,例如,图90h)包含:第一单体,其包含与第一hil
‑
10(a
‑
d)结构域共价连接的抗原结合结构域,所述第一hil
‑
10(a
‑
d)结构域与共价连接至第一异源二聚体fc链的第二hil
‑
10(a
‑
d)结构域共价连接;以及第二单体,其包含与第一hil
‑
10(e
‑
f)结构域共价连接的抗原结合结构域,所述第一hil
‑
10(e
‑
f)结构域与共价连接至第二异源二聚体fc链的第二hil
‑
10(e
‑
f)结构域共价连接。
[0491]
e.其它靶向性的含il10的fc融合体形式
[0492]
在(抗x)2
‑
中心
‑
scil10形式的一些实施方案中,异源二聚体fc融合蛋白包含:a)第一融合蛋白,其包含第一抗原结合结构域和第一fc结构域,其中第一抗原结合结构域与第一fc 结构域的n端共价连接;以及b)第二融合蛋白,其包含第二抗原结合结构域、第二fc结构域、第一蛋白结构域和第二蛋白结构域,其中第一蛋白结构域与第二蛋白结构域共价连接,其中第二蛋白结构域与第二fc结构域的n端共价连接,并且其中第一蛋白结构域与第二抗原结合结构域共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一蛋白结构域包含第一il
‑
10单体并且第二蛋白结构域包含第二il
‑ꢀ
10单体。
[0493]
在scil10
‑
(抗x)2
‑
异源fc形式的一些实施方案中,异源二聚体fc融合蛋白包含:a)第一融合蛋白,其包含第一抗原结合结构域和第一fc结构域,其中第一抗原结合结构域与第一fc 结构域的n端共价连接;以及b)第二融合蛋白,其包含第二抗原结合结构域、第二fc结构域、第一蛋白结构域和第二蛋白结构域,其中第二抗原结合结构域与第二fc结构域的n端共价连接,其中第一蛋白结构域与第二蛋白结构域共价连接,并且其中第二蛋白结构域与第二抗原结合结构域共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一蛋白结构域包含第一il
‑
10单体并且第二蛋白结构域包含第二il
‑
10单体。
[0494]
在scil10
‑
(抗x)2
‑
异源fc抗x x(il10m1)1
‑
异源fc形式的一些实施方案中,异源二聚体fc融合蛋白包含:a)第一融合蛋白,其包含第一抗原结合结构域和第一fc结构域,其中第一抗原结合结构域与第一fc结构域的n端共价连接;以及b)第二融合蛋白,其包含蛋白结构域和第二fc结构域,其中蛋白结构域与第二fc结构域共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中蛋白结构域包含含有插
入肽的il
‑
10单体结构域。在一些实施方案中,蛋白结构域与第二fc结构域的n端共价连接。在一些实施方案中,蛋白结构域与第二fc结构域的c端共价连接。
[0495]
在(抗x)2x异源fc
‑
(il10m1)1形式的一些实施方案中,异源二聚体fc融合蛋白包含: a)第一融合蛋白,其包含第一抗原结合结构域、第一蛋白结构域和第一fc结构域,其中第一抗原结合结构域与第一fc结构域的n端共价连接;以及b)第二融合蛋白,其包含第二抗原结合结构域和第二fc结构域,其中第二抗原结合结构域与第二fc结构域的n端共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中蛋白结构域包含含有插入肽的il
‑
10单体结构域。
[0496]
在一些实施方案中,第一蛋白结构域与第一fc结构域的c端共价连接。在一些实施方案中,第一蛋白结构域与第一抗原结合结构域的n端共价连接。
[0497]
在(抗x)2
‑
中心
‑
(il10m1)1
‑
异源fc形式的一些实施方案中,异源二聚体fc融合蛋白包含:a)第一融合蛋白,其包含第一抗原结合结构域、第一蛋白结构域和第一fc结构域,其中第一抗原结合结构域与第一蛋白结构域共价连接并且其中第一蛋白结构域与第一fc结构域的n端共价连接;以及b)第二融合蛋白,其包含第二抗原结合结构域和第二fc结构域,其中第二抗原结合结构域与第二fc结构域的n端共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中蛋白结构域包含含有插入肽的il
‑
10 单体结构域。
[0498]
在抗x x(il10m1)2
‑
异源fc形式的一些实施方案中,异源二聚体fc融合蛋白包含:a)第一融合蛋白,其包含第一抗原结合结构域和第一fc结构域,其中第一抗原结合结构域与第一 fc结构域的n端共价连接;以及b)第二融合蛋白,其包含第一蛋白结构域、第二蛋白结构域和第二fc结构域,其中第一蛋白结构域与第二蛋白结构域共价连接,并且其中第二蛋白结构域与第二fc结构域的n端共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一和第二蛋白结构域各自包含含有插入肽的il
‑
10单体。
[0499]
在抗x x异源fc
‑
(il10m1)2形式的一些实施方案中,异源二聚体fc融合蛋白包含:a) 第一融合蛋白,其包含第一抗原结合结构域和第一fc结构域,其中第一抗原结合结构域与第一fc结构域的n端共价连接;b)第二fc结构域;以及c)第一蛋白结构域和第二蛋白结构域,其中第一蛋白结构域与第二蛋白结构域共价连接,其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一和第二蛋白结构域各自包含含有插入肽的 il
‑
10单体。在一些实施方案中,第一蛋白结构域与第二蛋白结构域共价连接,并且其中第二蛋白结构域与第一fc结构域的c端共价连接。在一些实施方案中,第一蛋白结构域与第二蛋白结构域共价连接,并且其中第二蛋白结构域与第二fc结构域的c端共价连接。
[0500]
在抗x x异源fc
‑
(il10m1)2形式的一些实施方案中,异源二聚体fc融合蛋白包含:a) 第一融合蛋白,其包含第一抗原结合结构域、第一蛋白结构域、第二蛋白结构域和第一fc结构域,其中第一抗原结合结构域与第一fc结构域的n端共价连接,并且其中第一蛋白结构域与第二蛋白结构域共价连接,并且第二蛋白结构域与第一fc结构域的c端共价连接;以及b)第二融合蛋白,其包含第二抗原结合结构域和第二fc结构域,其中第二抗原结合结构域与第二fc结构域共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一和第二蛋白结构域各自包含含有插入肽的il
‑
10单体结构
域。
[0501]
在抗x x异源fc
‑
(il10m1)2形式的一些实施方案中,异源二聚体fc融合蛋白包含:)第一融合蛋白,其包含第一抗原结合结构域、第一蛋白结构域、第二蛋白结构域和第一fc结构域,其中第一抗原结合结构域与第一fc结构域的n端共价连接,并且其中第一蛋白结构域与第二蛋白结构域共价连接,并且第二蛋白结构域与第一抗原结合结构域的n端共价连接;以及b)第二融合蛋白,其包含第二抗原结合结构域和第二fc结构域,其中第二抗原结合结构域与第二fc结构域共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一和第二蛋白结构域各自包含含有插入肽的il
‑
10单体结构域。
[0502]
在抗x)2‑
中心
‑
(il10m1)2‑
异源fc形式的一些实施方案中,异源二聚体fc融合蛋白包含:a)第一融合蛋白,其包含第一抗原结合结构域、第一蛋白结构域、第二蛋白结构域和第一 fc结构域,其中第一蛋白结构域与第二蛋白结构域共价连接,并且第二蛋白结构域与第一fc 结构域的n端共价连接,并且其中第一抗原结合结构域与第一蛋白结构域的n端共价连接;以及b)第二融合蛋白,其包含第二抗原结合结构域和第二fc结构域,其中第二抗原结合结构域与第二fc结构域共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一和第二蛋白结构域各自包含含有插入肽的il
‑
10单体结构域。
[0503]
在mab
‑
(il10m1)2形式的一些实施方案中,异源二聚体fc融合蛋白包含:a)第一融合蛋白,其包含第一抗原结合结构域、第一蛋白结构域和第一fc结构域,其中第一抗原结合结构域与fc结构域的n端共价连接;以及b)第二融合蛋白,其包含第二抗原结合结构域、第二 fc结构域和第二蛋白结构域,其中第二抗原结合结构域与第二fc结构域的n端共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一和第二蛋白结构域各自包含含有插入肽的il
‑
10单体。在一些实施方案中,第一蛋白结构域与第一fc结构域的c端共价连接,并且第二蛋白结构域与第二fc结构域的c端共价连接。在一些实施方案中,第一蛋白结构域与第一抗原结合结构域的n端共价连接,并且第二蛋白结构域与第二抗原结合结构域的n端共价连接。
[0504]
在mab
‑
中心
‑
(il10m1)2形式的一些实施方案中,异源二聚体fc融合蛋白包含:a)第一融合蛋白,其包含第一抗原结合结构域、第一蛋白结构域和第一fc结构域,其中第一抗原结合结构域与第一蛋白结构域的n端共价连接,并且其中第一蛋白结构域与第一fc结构域的 n端共价连接;以及b)第二融合蛋白,其包含第二抗原结合结构域、第二fc结构域和第二蛋白结构域,其中第二抗原结合结构域与第二蛋白结构域的n端共价连接,并且其中第二蛋白结构域与第二fc结构域的n端共价连接;其中第一和第二fc结构域包含促进第一和第二fc 结构域的异源二聚化的修饰,并且其中第一和第二蛋白结构域各自包含含有插入肽的il
‑
10 单体。
[0505]
在mab
‑
中心
‑
(il10m1)2形式的一些实施方案中,异源二聚体fc融合蛋白包含:a)第一融合蛋白,其包含第一抗原结合结构域、第一蛋白结构域和第一fc结构域,其中第一抗原结合结构域与第一蛋白结构域的n端共价连接,并且其中第一蛋白结构域与第一fc结构域的 n端共价连接;以及b)第二融合蛋白,其包含第二抗原结合结构域、第二fc结构域和第二蛋白结构域,其中第二抗原结合结构域与第二蛋白结构域的n端共价连接,并且其中第二
蛋白结构域与第二fc结构域的n端共价连接;其中第一和第二fc结构域包含促进第一和第二fc 结构域的异源二聚化的修饰,并且其中第一和第二蛋白结构域各自包含含有插入肽的il
‑
10 单体。
[0506]
在抗x x(分裂il10)1
‑
异源fc形式的一些实施方案中,异源二聚体fc融合蛋白包含:a) 第一融合蛋白,其包含第一抗原结合结构域和第一fc结构域,其中第一抗原结合结构域与第一fc结构域的n端共价连接;以及b)第二融合蛋白,其包含第二fc结构域和第一蛋白结构域,其中第二fc结构域与第一蛋白结构域共价连接;其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一蛋白结构域包含人il
‑
10的螺旋(a
‑ꢀ
d)。在一些实施方案中,异源二聚体fc融合蛋白还包含与第一蛋白结构域非共价连接的第二蛋白结构域。在一些实施方案中,第二蛋白结构域包含人il
‑
10的螺旋(e
‑
f)。
[0507]
在(抗x)2x异源fc
‑
(分裂il10)1形式的一些实施方案中,异源二聚体fc融合蛋白包含: a)第一融合蛋白,其包含第一蛋白结构域、第一抗原结合结构域和第一fc结构域,其中第一抗原结合结构域与第一fc结构域的n端共价连接;以及b)第二融合蛋白,其包含第二蛋白结构域、第二抗原结合结构域和第二fc结构域,其中第二抗原结合结构域与第二fc结构域的n端共价连接,其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一蛋白结构域是人il
‑
10(a
‑
d)结构域并且第二蛋白结构域是人il
‑
10(e
‑
f) 结构域。在一些实施方案中,第一蛋白结构域与第一fc结构域的c端共价连接,并且第二蛋白结构域与第二fc结构域的c端共价连接。在一些实施方案中,第一蛋白结构域与第一抗原结合结构域的n端共价连接,并且第二蛋白结构域与第二抗原结合结构域的n端共价连接。
[0508]
在(抗x)2‑
中心
‑
(分裂il10)1‑
异源fc形式的一些实施方案中,异源二聚体fc融合蛋白包含:a)第一融合蛋白,其包含第一蛋白结构域、第一抗原结合结构域和第一fc结构域,其中第一抗原结合结构域与第一蛋白结构域共价连接,并且第一蛋白结构域与第一fc结构域的n 端共价连接;以及b)第二融合蛋白,其包含第二蛋白结构域、第二抗原结合结构域和第二fc 结构域,其中第二抗原结合结构域与第二蛋白结构域共价连接,并且第二蛋白结构域与第二 fc结构域的n端共价连接,其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一蛋白结构域是人il
‑
10(a
‑
d)结构域并且第二蛋白结构域是人il
‑ꢀ
10(e
‑
f)结构域。
[0509]
在抗x x(分裂il10)2‑
异源fc形式的一些实施方案中,异源二聚体fc融合蛋白包含: a)第一融合蛋白,其包含第一抗原结合结构域和第一fc结构域,其中第一抗原结合结构域与第一fc结构域的n端共价连接;以及b)第二融合蛋白,其包含第一蛋白结构域和第二蛋白结构域以及第二fc结构域,其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一蛋白结构域包含与第二人il
‑
10(a
‑
d)序列共价连接的第一人il
‑ꢀ
10(a
‑
d)序列,并且第二蛋白结构域包含与第二人il
‑
10(e
‑
f)序列共价连接的第一人il
‑
10(e
‑ꢀ
f)序列。在一些实施方案中,第一蛋白结构域与第二fc结构域共价连接,并且第二蛋白结构域与第一蛋白结构域非共价连接。在一些实施方案中,第二蛋白结构域与第二fc结构域共价连接,并且第一蛋白结构域与第二蛋白结构域非共价连接。
[0510]
在抗x)2x异源fc
‑
(分裂il10)2形式的一些实施方案中,异源二聚体fc融合蛋白包含: a)第一融合蛋白,其包含第一蛋白结构域、第一抗原结合结构域和第一fc结构域;以及
b)第二融合蛋白,其包含第二蛋白结构域、第二抗原结合结构域和第二fc结构域,其中第一和第二fc结构域包含促进第一和第二fc结构域的异源二聚化的修饰,并且其中第一蛋白结构域包含与第二人il
‑
10(a
‑
d)序列共价连接的第一人il
‑
10(a
‑
d)序列,并且第二蛋白结构域包含与第二人il
‑
10(e
‑
f)序列共价连接的第一人il
‑
10(e
‑
f)序列。在一些实施方案中,第一蛋白结构域与第一fc结构域的c端共价连接,并且第二蛋白结构域与第二fc结构域的c端共价连接。在一些实施方案中,第一抗原结合结构域与第一fc结构域的n端共价连接并且第一蛋白结构域与第一抗原结合结构域的n端共价连接,并且其中第二抗原结合结构域与第二fc 结构域的n端共价连接并且第二蛋白结构域与第二抗原结合结构域的n端共价连接。在一些实施方案中,第一蛋白结构域与第一fc结构域的n端共价连接并且第二蛋白结构域与第二 fc结构域的n端共价连接,并且其中第一抗原结合结构域与第一蛋白结构域的n端共价连接并且第二抗原结合结构域与第二蛋白结构域的n端共价连接。
[0511]
在mab
‑
il10形式的一些实施方案中,二聚体fc融合蛋白包含:(a)第一融合蛋白,其包含第一il
‑
10单体结构域、第一fc结构域和第一抗原结合结构域,其中第一il
‑
10单体结构域与第一fc结构域的c端共价连接,并且第一抗原结合结构域与第一fc结构域的n端共价连接;以及(b)第二融合蛋白,其包含第二il
‑
10单体结构域、第二fc结构域和第二抗原结合结构域,其中第二il
‑
10单体结构域与第二fc结构域的c端共价连接,并且第二抗原结合结构域与第二fc结构域的n端共价连接。在一些实施方案中,使用第一结构域接头将第一il
‑ꢀ
10单体结构域与第一fc结构域连接,和/或使用第二结构域接头将第二il
‑
10单体结构域与第二fc结构域连接。在一些实施方案中,使用第三结构域接头将第一抗原结合结构域与第一 fc结构域连接,和/或使用第四结构域接头将第二抗原结合结构域与第二fc结构域连接。在一些实施方案中,第一fc结构域和/或第二fc结构域包含选自由根据eu编号的m428l、 n434s和m428l/n434s组成的组的另一氨基酸取代。在一些实施方案中,第一和/或第二il
‑ꢀ
10单体结构域在位置109处包含亮氨酸。在一些实施方案中,第一和/或第二il
‑
10单体结构域包含选自由以下组成的组的多肽序列:seq id no:1(人il
‑
10(109h)前体序列)、seq idno:2(人il
‑
10(109l)前体序列)、seq id no:3(人il
‑
10(109h)成熟形式序列)、seq id no: 4(人il
‑
10(109l)成熟形式序列)。在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10 单体结构域是变体il
‑
10单体结构域。在一些实施方案中,变体il
‑
10单体结构域包括具有一个或多个氨基酸取代的il
‑
10单体结构域,所述氨基酸取代导致对il
‑
10受体的亲和力改变、效力改变、脱酰胺作用的潜力改变、天冬氨酸异构化的潜力改变、降解相关ptm的潜力改变、潜在降解位点改变、二硫桥改变和/或潜在n
‑
糖基化位点改变。在一些实施方案中,变体il
‑
10单体结构域在选自由n21、d28、q38、m39、d41、q42、l43、d44、n45、i87、e142、 d144和e151组成的组的氨基酸残基处包含一个或多个氨基酸修饰。在一些实施方案中,变体il
‑
10单体结构域包含选自由n21d、d28n、q38e、m39t、d41n、q42e、l43v、d44n、 n45d、i87a、e142q、d144n和e151q组成的组的一个或多个氨基酸修饰。
[0512]
在il10mab形式的一些实施方案中,二聚体fc融合蛋白包含:(a)第一融合蛋白,其包含第一il
‑
10单体结构域、第一fc结构域和第一抗原结合结构域,其中第一il
‑
10单体结构域与第一抗原结合结构域共价连接并且第一抗原结合结构域进一步与第一fc结构域的n端共价连接;以及(b)第二融合蛋白,其包含第二il
‑
10单体结构域、第二fc结构域和第二抗原结合结构域,其中第二il
‑
10单体结构域与第二抗原结合结构域共价连接并且第二抗原
结合结构域与第二fc结构域的n端共价连接。在一些实施方案中,使用第一结构域接头将第一 il
‑
10单体结构域与第一抗原结合结构域连接,和/或使用第二结构域接头将第二il
‑
10单体结构域与第二抗原结合结构域连接。在一些实施方案中,使用第三结构域接头将第一抗原结合结构域与第一fc结构域连接,和/或使用第四结构域接头将第二抗原结合结构域与第二fc 结构域连接。在一些实施方案中,第一和/或第二fc结构域包含选自由根据eu编号的m428l、 n434s和m428l/n434s组成的组的另一氨基酸取代。在一些实施方案中,第一和/或第二il
‑ꢀ
10单体结构域在位置109处包含组氨酸。在一些实施方案中,第一和/或第二il
‑
10单体结构域在位置109处包含亮氨酸。在一些实施方案中,第一和/或第二il
‑
10单体结构域包含选自由以下组成的组的多肽序列:seq id no:1(人il
‑
10(109h)前体序列)、seq id no:2(人il
‑ꢀ
10(109l)前体序列)、seq id no:3(人il
‑
10(109h)成熟形式序列)、seq id no:4(人il
‑
10 (109l)成熟形式序列)。在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域是变体il
‑
10单体结构域。在一些实施方案中,变体il
‑
10单体结构域包括具有一个或多个氨基酸取代的il
‑
10单体结构域,所述氨基酸取代导致对il
‑
10受体的亲和力改变、效力改变、脱酰胺作用的潜力改变、天冬氨酸异构化的潜力改变、降解相关ptm的潜力改变、潜在降解位点改变、二硫桥改变和/或潜在n
‑
糖基化位点改变。在一些实施方案中,变体il
‑
10单体结构域在选自由n21、d28、q38、m39、d41、q42、l43、d44、n45、i87、e142、d144 和e151组成的组的氨基酸残基处包含一个或多个氨基酸修饰。在一些实施方案中,变体il
‑ꢀ
10单体结构域包含选自由n21d、d28n、q38e、m39t、d41n、q42e、l43v、d44n、n45d、 i87a、e142q、d144n和e151q组成的组的一个或多个氨基酸修饰。
[0513]
在mab
‑
中心
‑
il10形式的一些实施方案中,二聚体fc融合蛋白包含:(a)第一融合蛋白,其包含第一il
‑
10单体结构域、第一fc结构域和第一抗原结合结构域,其中第一il
‑
10单体结构域与第一抗原结合结构域共价连接并且第一il
‑
10单体结构域进一步与第一fc结构域的 n端共价连接;以及(b)第二融合蛋白,其包含第二il
‑
10单体结构域、第二fc结构域和第二抗原结合结构域,其中第二il
‑
10单体结构域与第二抗原结合结构域共价连接并且第二il
‑
10 单体结构域与第二fc结构域的n端共价连接。
[0514]
在一些实施方案中,使用第一结构域接头将第一il
‑
10单体结构域与第一抗原结合结构域连接,和/或使用第二结构域接头将第二il
‑
10单体结构域与第二抗原结合结构域连接。在一些实施方案中,使用第三结构域接头将第一il
‑
10单体结构域与第一fc结构域连接,和/或使用第四结构域接头将第二il
‑
10单体结构域与第二fc结构域连接。在一些实施方案中,第一和/或第二fc结构域包含选自由根据eu编号的m428l、n434s和m428l/n434s组成的组的另一氨基酸取代。在一些实施方案中,第一和/或第二il
‑
10单体结构域在位置109处包含组氨酸。在一些实施方案中,第一和/或第二il
‑
10单体结构域在位置109处包含亮氨酸。在一些实施方案中,第一和/或第二il
‑
10单体结构域包含选自由以下组成的组的多肽序列:seqid no:1(人il
‑
10(109h)前体序列)、seq id no:2(人il
‑
10(109l)前体序列)、seq id no: 3(人il
‑
10(109h)成熟形式序列)、seq id no:4(人il
‑
10(109l)成熟形式序列)。在一些实施方案中,第一il
‑
10单体结构域和/或第二il
‑
10单体结构域是变体il
‑
10单体结构域。在一些实施方案中,变体il
‑
10单体结构域包括具有一个或多个氨基酸取代的il
‑
10单体结构域,所述氨基酸取代导致对il
‑
10受体的亲和力改变、效力改变、脱酰胺作用的潜力改变、天冬氨酸异构化的潜力改变、降解相关ptm的潜力改变、潜在降解位点改变、二
硫桥改变和/或潜在 n
‑
糖基化位点改变。变体il
‑
10单体结构域在选自由n21、d28、q38、m39、d41、q42、l43、 d44、n45、i87、e142、d144和e151组成的组的氨基酸残基处包含一个或多个氨基酸修饰。在一些实施方案中,变体il
‑
10单体结构域包含选自由n21d、d28n、q38e、m39t、d41n、 q42e、l43v、d44n、n45d、i87a、e142q、d144n和e151q组成的组的一个或多个氨基酸修饰。
[0515]
应当理解,本文所述的任何形式的抗原结合结构域,包括附图(包括但不限于图87a
‑
87d、图88a
‑
88e、图89a
‑
89m和图90a
‑
90h)中所示的那些,可针对任何合适的抗原,包括但不限于cd8、nkg2d、pd
‑
1和tigit。在一些实施方案中,将结合cd8的本文所述的任何抗原结合结构域(例如,第一和/或第二抗原结合结构域)并入靶向性il
‑
10融合蛋白形式中。示例性cd8 abd的序列提供于图92和序列表中。在一些实施方案中,将结合nkg2d的本文所述的任何抗原结合结构域(例如,第一和/或第二抗原结合结构域)并入靶向形式中。示例性 nkg2d abd的序列提供于图93a
‑
93c和序列表中。在一些实施方案中,将结合pd
‑
1的本文所述的任何抗原结合结构域(例如,第一和/或第二抗原结合结构域)并入靶向形式中。示例性pd
‑
1abd的序列提供于图100a
‑
100g和序列表中。示例性的非竞争性pd
‑
1abd的序列提供于图104a
‑
104b和序列表中。在一些实施方案中,将结合tigit的本文所述的任何抗原结合结构域(例如,第一和/或第二抗原结合结构域)并入靶向形式中。示例性tigit abd的序列提供于图122a
‑
122c和序列表中。
[0516]
在本文所述的靶向形式的另一方面,本文所述的任何il
‑
10单体结构域(例如,变体il
‑
10 单体结构域)(例如,图55、57、64、67、70a
‑
70b和73以及相应的seq id no中所示的那些)可用于适用的靶向性il
‑
10融合蛋白形式。在一些实施方案中,本文所述的hil
‑
10(a
‑
d) 和hil
‑
10(e
‑
f)序列(包括但不限于分裂il10的相应的seq id no:21和22)以及hil
‑
10(a
‑
d) 和hil
‑
10(e
‑
f)变体(例如,图55、57、64、67、70a
‑
70b和73以及相应的seq id no中所示的那些)中的任一个可用于靶向性il
‑
10融合蛋白形式。在某些实施方案中,本文所述的 il10m1序列(包括但不限于seq id no:24)和il10m1变体(例如,图36a
‑
36e、55、57、64、 67、70a
‑
70b、71a
‑
71e、73、74a
‑
74b、75a
‑
75b和76a
‑
76d以及相应的seq id no中所示的那些)中的任一个可用于靶向性il
‑
10融合蛋白形式。在一些实施方案中,il10m1序列中的任一个包含与seq id no:24具有至少90%(例如,至少90%、91%、92%、93%、94%、 95%、96%、97%、98%、99%或更大)序列同一性的氨基酸序列。在一些实施方案中,本文所述的scil
‑
10序列(包括但不限于seq id no:23)和scil10变体(例如,图21a
‑
21c、22、35a
‑ꢀ
35d、39a
‑
39g、40a
‑
40m、41a
‑
41f、48、49a
‑
49b、53、55、56a
‑
56c、58、59a
‑
59h、60、 64、65a
‑
65b、66a
‑
66d、67、68、69、70a
‑
70b、73、78a
‑
78g、79a
‑
79g和86a
‑
86d以及相应的seq id no中所示的那些)中的任一个可用于靶向性il
‑
10融合蛋白形式。在一些实施方案中,scil10序列中的任一个包含与seq id no:23具有至少90%(例如,至少90%、91%、 92%、93%、94%、95%、96%、97%、98%、99%或更大)序列同一性的氨基酸序列。
[0517]
在一个方面,本文提供一种包含本文(包括附图)所概述的异源二聚体fc融合蛋白或二聚体融合蛋白中的任一种的组合物,其用于治疗受试者的癌症。还提供了一种或多种核酸,其编码这种异源二聚体fc融合蛋白或二聚体融合蛋白。还提供了一种宿主细胞,其包含编码本文所述的异源二聚体fc融合蛋白或二聚体融合蛋白的一种或多种此类核酸。
[0518]
在另一方面,本文提供了一种产生所描述的任何异源二聚体fc融合蛋白或二聚体
融合蛋白的方法。在一些实施方案中,所述方法包括在借由产生异源二聚体fc融合蛋白或二聚体融合蛋白的条件下培养本文概述的任何宿主细胞;以及回收蛋白质。
[0519]
viii.本发明的有用实施方案
[0520]
如本领域技术人员将理解的并且如下文更全面地讨论的,本发明的非靶向性il
‑
10融合蛋白可以采取广泛多种配置,如附图中总体上描绘的,所述附图包括但不限于图16a
‑
16d、 20a
‑
20b、23a
‑
23f和29a
‑
29d。本发明的靶向性il
‑
10融合蛋白可以采取广泛多种配置,如附图中总体上描绘的,所述附图包括但不限于图87a
‑
87d、88a
‑
88d、89a
‑
89m和90a
‑
90h。
[0521]
在本发明的一个实施方案中是图20a中描绘的scil10
‑
异源fc形式,其具有il
‑
10变体和/或在fc结构域中具有氨基酸取代。一般来说,这种形式包含:第一单体/融合蛋白,其包含与第一异源二聚体fc链的n端共价连接(任选地通过结构域接头)的scil
‑
10;以及第二单体/融合蛋白,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链。在这样的实施方案中,scil
‑
10包含与第二il
‑
10单体结构域连接的第一il
‑
10单体结构域。
[0522]
在另外的实施方案中,此示例性形式的fc融合蛋白具有分别在图78b、78c和78f中针对xenp30005(seq id no:25和26)、xenp30008(seq id no:27和28)或xenp30013(seqidno:29和30)列出的序列。在另外的实施方案中,此示例性形式的fc融合蛋白包含分别在图78b、78c和78f中针对xenp30005(seq id no:25和26)、xenp30008(seq id no:27 和28)或xenp30013(seq idno:29和30)列出的序列,其具有选自下组的另外的氨基酸修饰 (单独或以任何组合):根据eu编号的g445del、k447del、g445del/k447del、m428l、n434s 和m428l/n434s。
[0523]
在一些实施方案中,本发明的scil10
‑
异源fc形式融合蛋白包含il
‑
10单体,所述单体在图20a中描绘的scil10
‑
异源fc形式内包含图1中描绘的任何人il10前体或成熟序列(109h 或109l)。在另外的实施方案中,il
‑
10单体包含经过设计以降低其对il
‑
10受体复合物的亲和力和/或降低其效力的il
‑
10单体变体。在此类亲和力/效力变体中,变体il
‑
10单体可在选自以下的位置处包含一个或多个取代:n21、d28、q38、m39、d41、q42、l43、d44、n45、 i87、e142、d144和e151,单独或以任何组合。变体il
‑
10单体的示例性序列在图35a
‑
35c 中示出
‑
应当理解,虽然所描绘的序列是在il
‑
10(109l)序列的背景中,但可以在il
‑
10(109h) 序列的背景中作出相同取代。图35a
‑
35c中描绘的任何序列可以在图20a中描绘的scil10
‑ꢀ
异源fc形式的背景中使用。
[0524]
在一些实施方案中,本发明的scil10
‑
异源fc形式融合蛋白在fc结构域中包括改善可制造性的修饰。在另外的实施方案中,本发明的scil10
‑
异源fc形式融合蛋白在一个或两个fc 结构域的fc区中包括g446del/k4447del修饰。在另外的实施方案中,本发明的scil10
‑
异源 fc形式融合蛋白在一个或两个fc结构域中包括g446del/k4447del修饰和m428l/n434s修饰。
[0525]
在另外的实施方案中,本发明的scil10
‑
异源fc形式融合蛋白在第一和第二il
‑
10单体结构域之间和/或在scil10与fc结构域之间包括结构域接头。在另外的实施方案中,所使用的结构域接头选自附图中所示的那些。在另外的实施方案中,在第一il
‑
10单体与第二il
‑
10 单体之间包含结构域接头的scil
‑
10复合物包括如图48中描绘的序列,包括seq id no:xx (scil
‑
10(g4s))、seq id no:xx(scil
‑
10(g4s)2)或seq id no:xx(scil
‑
10(g4s)3)。
abd 包含根据图92和相应seq id no的序列表中列出的任一个的可变重链序列。在另外的实施方案中,人源化的抗cd8 abd包含根据图92和相应seq id no的序列表中列出的任一个的可变轻链序列。在一些实施方案中,人源化的抗cd8 abd包含根据图92和相应seq idno的序列表中列出的任一个的可变重链序列和可变轻链序列。在另外的实施方案中,人源化的抗cd8 abd在抗x x scil10
‑
异源fc形式中与含有m428l/n434s修饰的第一和第二fc结构域中的任一者或两者组合。在另外的实施方案中,本发明的抗x x scil10
‑
异源fc形式融合蛋白是xenp25365、xenp25366和xenp30525。
[0532]
在其中抗x(抗原结合结构域)结合nkg2d的抗x x scil10
‑
异源fc形式的实施方案中,此类形式的例示性序列在图98
‑
100g中提供。在一些实施方案中,抗x x scil10
‑
异源fc形式中使用的nkg2d abd序列是人源化的抗nkg2d abd。在另外的实施方案中,人源化的抗 nkg2d abd包含根据图93a
‑
93c和相应seq id no的序列表中列出的任一个的可变重链序列。在另外的实施方案中,人源化的抗nkg2d abd包含根据图93a
‑
93c和相应seq idno的序列表中列出的任一个的可变轻链序列。在一些实施方案中,人源化的抗nkg2d abd 包含根据图93a
‑
93c和相应seq id no的序列表中列出的任一个的可变重链序列和可变轻链序列。在另外的实施方案中,人源化的抗nkg2d abd在抗x x scil10
‑
异源fc形式中与含有m428l/n434s修饰的第一和第二fc结构域中的任一者或两者组合。在另外的实施方案中,本发明的抗x x scil10
‑
异源fc形式融合蛋白是xenp25953、xenp30526、xenp30527、 xenp30528和xenp31819。
[0533]
ix.本发明的核酸
[0534]
本发明还提供了核酸组合物,所述核酸组合物编码本发明的fc融合蛋白、il
‑
10单体结构域和il
‑
10二聚体复合物(或者,在单体fc结构域蛋白的情况下,还提供编码那些蛋白的核酸)。
[0535]
如本领域技术人员将理解,核酸组合物将取决于融合蛋白的形式。因此,例如,当所述形式需要三个氨基酸序列时,可以将三个核酸序列并入一个或多个表达载体中以用于表达。类似地,仅两种核酸需要一些形式;再次,所述两种核酸可以被放入到一个或两个表达载体中。
[0536]
如本领域已知的,对本发明的组分进行编码的核酸可以并入到如本领域已知并且取决于用于产生本发明的异源二聚体fc融合蛋白的宿主细胞的表达载体中。通常,核酸可操作地连接到任何数量的调控元件(启动子、复制起点、可选择标记、核糖体结合位点、诱导剂等)。表达载体可以是染色体外的或整合的载体。
[0537]
本发明的核酸和/或表达载体然后转化成如本领域众所周知的任何数量的不同类型的宿主细胞,包括哺乳动物细胞、细菌细胞、酵母细胞、昆虫细胞和/或真菌细胞,其中哺乳动物细胞(例如,cho细胞)可用于许多实施方案中。
[0538]
在一些实施方案中,如果根据形式适用的话,编码每个单体的核酸通常在不同或相同的启动子控制下各自包含在单个表达载体内。在本发明的特别有用的实施方案中,这两种或三种核酸中的每种包含在不同表达载体上。
[0539]
通过培养包含如本领域众所周知的一种或多种表达载体的宿主细胞来制备本发明的fc 融合蛋白。一旦产生,就进行传统的融合蛋白或抗体纯化步骤,包括离子交换色谱法步骤。如本文所讨论的,使两个单体的pi相差至少0.5可以通过离子交换色谱法或等电聚
焦或对等电点敏感的其它方法进行分离。也就是说,包含改变每个单体的等电点(pi)的pi取代使得每个单体具有不同的pi并且异源二聚体也具有不同的pi,从而促进异源二聚体的等电纯化(例如,阴离子交换色谱法、阳离子交换色谱法)。这些取代还有助于确定和监测纯化后的任何污染性同源二聚体(例如,ief凝胶、cief和分析iex柱)。
[0540]
x.il
‑
10免疫调节fc融合蛋白的生物和生化功能
[0541]
通常,本发明的fc融合蛋白施用于患有癌症的患者,并且以如本文所描述的多种方式对功效进行评估。因此,在可以进行标准功效测定时,如癌症负载、肿瘤大小、对存在或转移程度的评估等,可以也可以基于免疫状态评估来估计免疫肿瘤学治疗。这可以多种方式进行,包括体外和体内测定。例如,可以与“旧式”测量如肿瘤负荷、大小、浸润性、ln参与、转移等一起进行免疫状态变化(例如,在ipi处理后icos cd4 t细胞的存在)的评估。因此,可以评估以下中的任一个或全部:pvrig对cd4
t细胞活化或增殖、cd8
t(ctl)细胞活化或增殖、cd8
t细胞介导的细胞毒性活性和/或ctl介导的细胞耗竭、nk细胞活性和nk介导的细胞耗竭的抑制作用;pvrig对treg细胞分化和增殖和treg或骨髓来源的抑制细胞 (mdsc)介导的免疫抑制或免疫耐受的增强作用;和/或pvrig对通过免疫细胞产生促炎性细胞因子的影响,例如通过t细胞或其它免疫细胞产生il
‑
2、ifn
‑
γ或tnf
‑
α。
[0542]
在一些实施方案中,通过使用例如cfse稀释方法、免疫效应细胞的ki67细胞内染色和3h
‑
胸腺掺入法评估免疫细胞增殖来进行治疗估计。
[0543]
在一些实施方案中,通过评估基因表达的增加或活化相关标志物的蛋白质含量的增加来评定处理,所述标志物包括以下中的一个或多个:cd25、cd69、cd137、icos、pd1、gitr、 ox40以及通过cd107a的表面表达测量的细胞脱粒。
[0544]
通常,按本领域已知的那样进行基因表达测定。
[0545]
通常,按本领域已知的那样进行基因表达测定。
[0546]
在一些实施方案中,通过评估各种细胞参数如酶活性(包含蛋白酶活性)、细胞膜渗透性、细胞粘附、atp产生、辅酶产生和核苷酸摄取活性来估计通过靶细胞活力检测测量的细胞毒活性进行治疗估计。这些分析的具体实例包括但不限于台盼蓝(trypan blue)或pi染色、
51
cr或
35
s释放法、ldh活性、mtt和/或wst分析、钙黄绿素
‑
am分析、基于发光的分析以及其它分析。
[0547]
在一些实施方案中,通过使用众所周知的技术评估通过细胞因子产生测量的t细胞活性、使用包括但不限于ifnγ、tnfα、gm
‑
csf、il2、il6、il4、il5、il10、il13的细胞因子的培养物上清液中的细胞内度量,来进行处理的评估。
[0548]
因此,可以使用评估以下中的一个或多个的测定来评定治疗:(i)免疫应答的增加;(ii)αβ和/或γδt细胞活化的增加;(iii)细胞毒性t细胞活性的增加;(iv)nk和/或nkt细胞活性的增加;(v)αβ和/或γδt细胞抑制的缓解;(vi)促炎性细胞因子分泌的增加;(vii)il
‑
2分泌的增加;(viii)干扰素γ产生的增加;(ix)th1反应的增加;(x)th2反应的减少;(xi)调节性t细胞 (treg)中的至少一种的细胞数量和/或活性的减少或消除。。
[0549]
a.测量功效和效力的测定
[0550]
在一些实施方案中,使用如本领域已知的混合淋巴细胞反应(mlr)测定来评定t细胞活化。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0551]
在一个实施方案中,信号传导通路测定测量免疫应答的增加或减少,如例如通过
不同因子的磷酸化或去磷酸化或通过测量其它翻译后修饰所测量的。il
‑
12通过stat4的磷酸化介导ifnγ表达和分泌(morinobu等人,2002)。因此,在一个优选实施方案中,信号传导途径测定测量免疫应答的增加或减少,如stat4的磷酸化所指示的。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0552]
在一个实施方案中,信号传导途径分析测量αβ和/或γδt细胞活化的增加或减少,如例如通过细胞因子分泌或通过增殖或通过如例如cd137、cd107a、pd1等活化标志物的表达变化所测量。活性增加指示免疫刺活化性。适当的活性增加概述如下。
[0553]
在一个实施方案中,信号传导通路分析测量细胞毒性t细胞活性的增加或减少,如例如通过直接杀伤靶细胞(例如癌细胞)或通过细胞因子分泌或通过增殖或通过如例如cd137、 cd107a、pd1等活化标志物的表达变化所测量。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0554]
在一个实施方案中,信号传导途径测定测量nk和/或nkt细胞活性的增加或减少,如例如通过直接杀伤靶细胞如例如癌细胞或通过细胞因子分泌或通过如例如cd107a等活化标志物的表达变化所测量的。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0555]
在一个实施方案中,信号传导途径测定测量αβ和/或γδt细胞抑制的增加或减少,如例如通过细胞因子分泌或通过增殖或通过如例如cd137、cd107a、pd1等活化标志物的表达变化所测量的。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0556]
在一个实施方案中,信号传导途径测定测量如例如通过elisa或通过luminex或通过基于多珠方法或通过胞内染色和facs分析或通过alispot等测量的促炎性细胞因子分泌的增加或减少。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0557]
在一个实施方案中,信号传导途径测定测量il
‑
2分泌的增加或减少,如例如通过elisa 或通过luminex或通过基于微珠的多重方法或通过细胞内染色和facs分析或通过alispot等所测量的。活性的增加指示免疫刺激活性。适当的活性增加概述如下。
[0558]
在一个实施方案中,信号传导途径测定测量如例如通过elisa或通过luminex或通过基于多珠方法或通过胞内染色和facs分析或通过alispot等测量的干扰素
‑
γ产生的增加或减少。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0559]
在一个实施方案中,信号传导途径测定测量th1反应的增加或减少,如例如通过细胞因子分泌或通过活化标志物的表达变化所测量的。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0560]
在一个实施方案中,信号传导途径测定测量th2反应的增加或减少,如例如通过细胞因子分泌或通过活化标志物的表达变化所测量的。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0561]
在一个实施方案中,信号传导途径测定测量至少一种调控t细胞(treg)的细胞数量和/或活性的增加或减少,如例如通过流式细胞术或通过ihc所测量的。反应减少指示免疫刺激活性。如下所概述,适当的减少与增加相同。
[0562]
在一个实施方案中,信号传导途径测定测量m2巨噬细胞细胞数量的增加或减少,如例如通过流式细胞术或通过ihc所测量的。反应减少指示免疫刺激活性。如下所概述,适当的减少与增加相同。
[0563]
在一个实施方案中,信号传导途径测定测量m2巨噬细胞促肿瘤发生活性的增加或
减少,如例如通过细胞因子分泌或通过活化标志物的表达变化所测量的。反应减少指示免疫刺激活性。如下所概述,适当的减少与增加相同。
[0564]
在一个实施方案中,信号传导途径测定测量n2嗜中性粒细胞增量的增加或减少,如例如通过流式细胞术或通过ihc所测量的。反应减少指示免疫刺激活性。如下所概述,适当的减少与增加相同。
[0565]
在一个实施方案中,信号传导途径测定测量n2嗜中性粒细胞促肿瘤发生活性的增加或减少,如例如通过细胞因子分泌或通过活化标志物的表达变化所测量的。反应减少指示免疫刺激活性。如下所概述,适当的减少与增加相同。
[0566]
在一个实施方案中,信号传导途径测定测量t细胞活化抑制的增加或减少,如例如通过细胞因子分泌或通过增殖或通过如例如cd137、cd107a、pd1等活化标志物的表达变化所测量的。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0567]
在一个实施方案中,信号传导途径测定测量ctl活化抑制的增加或减少,如例如通过直接杀伤靶细胞如例如癌细胞或通过细胞因子分泌或通过增殖或通过如例如cd137、cd107a、 pd1等活化标志物的表达变化所测量的。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0568]
在一个实施方案中,信号传导途径测定测量αβ和/或γδt细胞耗尽的增加或减少,如例如通过活化标志物的表达变化所测量的。反应减少指示免疫刺激活性。如下所概述,适当的减少与增加相同。
[0569]
在一个实施方案中,信号传导途径测定测量αβ和/或γδt细胞反应的增加或减少,如例如通过细胞因子分泌或通过增殖或通过如例如cd137、cd107a、pd1等活化标志物的表达变化所测量的。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0570]
在一个实施方案中,信号传导途径测定测量抗原特异性记忆反应的刺激的增加或减少,如例如通过细胞因子分泌或通过增殖或通过如例如cd45ra、ccr7等活化标志物的表达变化所测量的。活性增加指示免疫刺激活性。适当的活性增加概述如下。。
[0571]
在一个实施方案中,信号传导途径测定测量如例如通过细胞毒性测定如例如mtt、cr释放、钙黄绿素am或通过基于流式细胞术测定如例如cfse稀释或碘化丙啶染色等测量的癌细胞的凋亡或裂解的增加或减少。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0572]
在一个实施方案中,信号传导途径测定测量如例如通过细胞毒性测定如例如mtt、cr释放、钙黄绿素am或通过基于流式细胞术测定如例如cfse稀释或碘化丙啶染色等测量的刺激对癌细胞的细胞毒性或细胞抑制作用的增加或减少。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0573]
在一个实施方案中,信号传导途径测定测量如例如通过细胞毒性测定如例如mtt、cr释放、钙黄绿素am或通过基于流式细胞术测定如例如cfse稀释或碘化丙啶染色等测量的直接杀伤的癌细胞的增加或减少。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0574]
在一个实施方案中,信号传导途径测定测量th17活性的增加或减小,如例如通过细胞因子分泌或通过增殖或通过活化标志物的表达变化所测量的。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0575]
在一个实施方案中,信号传导途径测定测量补体依赖性细胞毒性和/或抗体依赖
性细胞介导的细胞毒性的诱导的增加或减少,如例如通过细胞毒性测定如例如mtt、cr释放、calcineam或通过基于流式细胞术的测定如例如cfse稀释或碘化丙啶染色等所测量的。活性增加指示免疫刺激活性。适当的活性增加概述如下。
[0576]
在一个实施方案中,例如通过直接杀伤靶细胞(例如癌细胞)或通过细胞因子分泌或通过增殖或通过活化标志物(例如cd137、cd107a、pd1等)的表达变化来测量t细胞活化。对于 t细胞,增殖、细胞表面活化标志物(例如,cd25、cd69、cd137、pd1)、细胞毒性(杀伤靶细胞的能力)和细胞因子产生(例如,il
‑
2、il
‑
4、il
‑
6、ifnγ、tnf
‑
a、il
‑
10、il
‑
17a)的增加将指示免疫调节,这将符合癌细胞的杀伤增强。
[0577]
在一个实施方案中,例如通过直接杀伤靶细胞如例如癌细胞或通过细胞因子分泌或通过增殖或通过改变活化标志物如例如cd107a等的表达测量nk细胞活化。对于nk细胞,增殖增加、细胞毒性(杀伤靶细胞并增加cd107a、颗粒酶和穿孔素表达的能力)、细胞因子产生(例如,ifnγ和tnf)和细胞表面受体表达(例如cd25)将指示免疫调节,其与增强的癌细胞杀伤作用相一致。
[0578]
在一个实施方案中,例如通过细胞因子分泌或通过增殖或通过改变活化标志物的表达来测量γδt细胞活化。
[0579]
在一个实施方案中,例如通过细胞因子分泌或通过活化标志物的表达变化来测量th1细胞活化。
[0580]
活性或应答的适当增加(或减少,如上文所概述的视情况而定)比参考样品或对照样品例如不含有本发明的il
‑
12异源二聚体融合蛋白的测试样品中的信号增加10%、20%、30%、 40%、50%、60%、70%、80%、90%、95%或98%至99%。类似地,与参考或对照样品相比,增加至少一倍、两倍、三倍、四倍或五倍显示功效。
[0581]
xi.治疗
[0582]
一旦制成,在与本发明的异源二聚体fc融合蛋白结合的情况下,通过治疗癌症、通常促进t细胞活化(例如,不再抑制t细胞),本发明的组合物可以用于许多肿瘤学应用。
[0583]
因此,本发明的异源二聚体组合物可以用于治疗这些癌症。
[0584]
a.体内施用的融合蛋白组合物
[0585]
通过将具有期望程度的纯度的融合蛋白与任选的药学上可接受的呈冻干调配物或水性溶液的形式的载体、赋形剂或稳定剂(如《雷明顿药物科学(remington’s pharmaceutical sciences)》, 第16版,osol,a.编辑[1980]中通常概述的),用于以冻干配制物或水溶液形式储存。可接受的载体、缓冲剂、赋形剂或稳定剂在所用剂量和浓度下对接受者无毒,并且包括缓冲剂,例如磷酸盐、柠檬酸盐和其它有机酸;抗氧化剂,包括抗坏血酸和蛋氨酸;防腐剂(例如十八烷基二甲基苯甲基氯化铵;六甲基氯化铵;苯扎氯铵、苄索氯铵;苯酚、丁醇或苄醇;对羟基苯甲酸烷基酯,如对羟基苯甲酸甲酯或对羟基苯甲酸丙酯;儿茶酚;间苯二酚;环己醇;3
‑
戊醇;和间甲酚);低分子量(少于约10个残基)多肽;蛋白质,例如血清白蛋白、明胶或免疫球蛋白;亲水性聚合物,例如聚乙烯吡咯烷酮;氨基酸,例如甘氨酸、谷氨酰胺、天冬酰胺、组氨酸、精氨酸或赖氨酸;单糖、二糖和其它碳水化合物,包括葡萄糖、甘露糖或糊精;螯合剂,例如edta;糖,例如蔗糖、甘露醇、海藻糖或山梨糖醇;成盐抗衡离子,例如钠;金属配合物(例如锌
‑
蛋白质配合物);和/或非离子表面活性剂,例如tween
tm
、pluronics
tm
或聚乙二醇(peg)。
[0586]
b.施用方式
[0587]
本发明的融合蛋白和化疗剂根据已知的方法,如静脉内施用(作为团注物(bolus)或通过在一段时间内连续输注)向受试者施用。
[0588]
c.治疗方式
[0589]
在本发明的方法中,用疗法来提供对疾病或病症的阳性治疗应答。“阳性治疗应答”旨在改善疾病或病症和/或改善与疾病或病症相关联的症状。例如,阳性治疗应答将指对疾病的以下改善中的一种或多种:(1)减少赘生性细胞的数量;(2)增加赘生性细胞死亡;(3)抑制赘生性细胞存活;(5)抑制(即减慢到某种程度并且优选地停止)肿瘤生长;(6)增加患者存活率;以及(7)与疾病或病症相关联的一个或多个症状的一些缓解。
[0590]
可以通过对那个疾病或病症具有特异性的标准化应答标准来确定任何给定疾病或病症的阳性治疗应答。可以使用筛选技术如磁共振成像(mri)扫描、x
‑
射线成像、计算机断层(ct)扫描、骨扫描成像、内窥镜检查和肿瘤活检取样,包含骨髓吸出(bma)和对循环中的肿瘤细胞计数来估计针对肿瘤形态(即总体肿瘤负荷、肿瘤大小等)改变的肿瘤应答。
[0591]
除了这些阳性治疗应答之外,正经受疗法的受试者可能经历与疾病相关联的症状的改善的有益效果。
[0592]
根据本发明的治疗包含所使用的“治疗有效量的”药物。“治疗有效量”是指以剂量计并且持续所需的时间段以实现所希望的治疗结果的有效的量。
[0593]
治疗有效量可以根据因素而变化,如个体的疾病状态、年龄、性别、以及体重、以及药物在个体中引发希望的应答的能力。治疗有效量也是蛋白或蛋白部分的治疗有益效果超过其任何毒性或有害效果的量。
[0594]
肿瘤疗法的“治疗有效量”还可以通过其稳定疾病的进展的能力测量。可以在预示人类肿瘤的功效的动物模型系统中评估化合物抑制癌症的能力。
[0595]
可替代地,通过熟练的从业者已知的体外测定,组合物的这种特性可以通过检验化合物抑制肿瘤生长或诱导凋亡的能力来评估。治疗有效量的治疗化合物可以减小肿瘤大小或以其它方式减轻受试者的症状。本领域的普通技术人员将能够基于如受试者的大小、受试者症状的严重程度和特定的组合物或所选择的施用途径这种因素确定这种量。
[0596]
调整剂量方案以提供最佳的期望应答(例如,治疗应答)。例如,可以施用单个团注物,或者可以随时间施用若干分次剂量,或者可以按治疗状况的紧急情况所表明的成比例地减少或增加剂量。为了方便施用和剂量的均匀性,肠胃外组合物可以按剂量单位调配。如本文使用的剂量单位是指作为针对要治疗受试者的单一剂量适合的物理上离散的单位;每个单位含有与所需药物载体缔合的、经计算产生所期望的治疗效果的预定量的活性化合物。
[0597]
针对本发明的剂量单位形式的规格受制于并直接取决于:(a)活性化合物的独特特性和要实现的特定治疗效果,和(b)个体敏感性对复合用于治疗的此类活性化合物造成的本领域固有限制。
[0598]
用于本发明的异源二聚体蛋白的有效剂量和剂量方案取决于待治疗的疾病或病状并且可以由本领域的技术人员确定。
[0599]
用于本发明的治疗有效量的异源二聚体蛋白的示例性非限制性范围是约0.1
‑
100mg/kg。
[0600]
所有引用的参考文献通过引用以其全部内容明确地并入本文。
[0601]
尽管出于说明的目的已经在上文描述了本发明的具体实施方案,但是本领域技术人员将了解的是,在不脱离如所附权利要求所描述的本发明的情况下可以做出许多细节变化。
[0602]
实施例
[0603]
下文提供的实施例旨在说明本发明。这些实施例不旨在将本发明限制于任何特定的应用或操作理论。根据eu和kabat的重恒定区编号的有用比较如下,参见edelman等人,1969, 《美国国家科学院院刊(proc natl acad sci usa)》63:78
‑
85,以及kabat等人,1991,《免疫学关注的蛋白质的序列(sequences of proteins of immunological interest)》,第5版,美国公共卫生署(united states public health service),国立卫生研究院(national institute of health),贝塞斯达(bethesda),所述文献通过引用整体并入。抗体领域中的技术人员将理解,这个规约由免疫球蛋白序列的特定区域中的非顺序编号组成,所述非顺序编号使得能够对免疫球蛋白家族中的保守位置进行标准化的引用。因此,如通过eu索引定义的任何给定免疫球蛋白的位置不一定对应于其顺序序列。
[0604]
在美国公开2015/0307629、2014/0288275和wo2014/145806中概述了一般和具体的科学技术,其均通过引用明确地整体并入本文中,并且特别是针对其中概述的技术。
[0605]
实施例1:il
‑
10融合蛋白的工程化
[0606]
如上所述,诸如il
‑
10的细胞因子的半衰期短,并且需要高剂量治疗才能在靶标(例如肿瘤部位)处达到足以诱导免疫应答的细胞因子浓度。然而,基于对其他细胞因子的观察,用il
‑ꢀ
10进行高剂量治疗可能会导致全身毒性。此外,天然il
‑
10单体是没有生物活性的,并且生物活性il
‑
10二聚体在两个单体单元之间的相互作用被破坏时变得没有活性。为了解决这两个问题,我们将各种il
‑
10形式工程化为fc融合蛋白(在此统称为il
‑
10融合蛋白或il
‑
10融合体),目的是既促进生物活性il
‑
10,又通过fcrn介导的再循环增强血清半衰期。
[0607]
1a:il
‑
10融合蛋白形式
[0608]
如上所述,生物活性il
‑
10是通过两个il
‑
10单体之间的非共价相互作用形成的结构域交换的同源二聚体。每个il
‑
10单体由6个螺旋组成,本文称为螺旋a
‑
f。先前已有报道,形成生物活性il
‑
10同源二聚体的非共价相互作用是由第一il
‑
10单体的螺旋a
‑
d(或“hil
‑
10(a
‑ꢀ
d)”;成熟形式il
‑
10的残基1
‑
116)与第二il
‑
10单体的螺旋e
‑
f(或“hil
‑
10(e
‑
f)”;成熟形式il
‑
10的残基117
‑
160)之间的结构域交换引起的。在这里,我们描述了四类il
‑
10融合蛋白,其被工程化以保留il
‑
10的二聚体性质或规避对il
‑
10单体之间的结构域交换的需要。
[0609]
1a(a):il10
‑
fc融合体
[0610]
为了工程化其中预复合了il
‑
10同源二聚体的il
‑
10融合蛋白,我们设想的第一il
‑
10融合体类别是il10
‑
fc融合体(图16中描绘的卡通示意图)。我们作为原型工程化的这种类别的一种这样的形式是(il10)2‑
fc形式(图16a中描绘的卡通示意图),其包含两个相同的单体,每个单体包含与同源二聚体fc链的n端共价连接的il
‑
10单体。(il10)2‑
fc形式的例示性蛋白质包括xenp24628,其序列在图17中描绘。我们作为原型工程化的这种类别的另一形式是 fc
‑
(il10)2形式(图16b中描绘的卡通示意图),其包含两个相同的单体,每个
单体包含与同源二聚体fc链的c端共价连接的il
‑
10单体。fc
‑
(il10)2形式的例示性蛋白质包括xenp24632,其序列在图18中描绘。我们作为原型工程化的这种类别的另一形式是(il10
‑
nc
‑
il10)
‑
异源fc 形式(图16c中描绘的卡通示意图),其包含:第一单体,其包含与第一异源二聚体fc链的n 端共价连接(任选地通过结构域接头)的第一il
‑
10单体;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链,同时第二il
‑
10单体单独转染,使得形成非共价 il
‑
10二聚体。(il10
‑
nc
‑
il10)
‑
异源fc形式的例示性蛋白质包括xenp25955,其序列在图19 中描绘。
[0611]
1a(b):scil10
‑
fc融合体
[0612]
接下来,我们试图将il
‑
10同源二聚体预复合为单链il
‑
10复合物(或“scil
‑
10”),其中第一il
‑
10单体与第二il
‑
10单体共价连接。我们设想了一类利用scil
‑
10的il
‑
10融合体,在此称为scil10
‑
fc融合体(图20中描绘的卡通示意图)。我们作为原型工程化的这种类别的一种这样的形式是scil10
‑
异源fc形式(图20a中描绘的卡通示意图),其包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的scil
‑
10;以及第二单体,其包含为“仅 fc”或“空fc”的互补的第二异源二聚体fc链。scil10
‑
异源fc形式的例示性蛋白质包括 xenp25880,其序列在图21中描绘。我们作为原型工程化的这种类别的另一这样的形式是异源fc
‑
scil10形式(图20b中描绘的卡通示意图),其包含:第一单体,其包含与第一异源二聚体fc链的c端共价连接(任选地通过结构域接头)的scil
‑
10;以及第二单体,其包含为“仅 fc”或“空fc”的互补的第二异源二聚体fc链。异源fc
‑
scil10的例示性蛋白质是 xenp28740,其序列在图22中描绘。
[0613]
1a(c):il10m1
‑
fc融合体
[0614]
接下来,我们探索了被工程化以规避对两个il
‑
10单体之间的结构域交换的需要的il
‑
10 融合体形式。如上文所论述,形成生物活性il
‑
10同源二聚体的非共价相互作用的破坏产生生物学上无活性的il
‑
10单体。然而,josephson等人(2000)报道了“il
‑
10m1”(或“il10m1”;图15d中描绘的序列),其为通过在il
‑
10单体的螺旋d与e之间工程化gly
‑ꢀ
ser接头(gggsgg)的而生成的生物活性il
‑
10单体。josephson等人报道,il
‑
10m1能够结合 il
‑
10r1并募集il
‑
10r2,以诱导il
‑
10细胞反应。因此,我们设想了一类利用il10m1的il
‑ꢀ
10融合蛋白,在此称为il10m1
‑
fc融合体(图23中描绘的卡通示意图)。我们作为原型工程化的这种类别的一种这样的形式是(il10m1)1‑
异源fc(图23e中描绘的卡通示意图),其包含:与第一异源二聚体fc链的n端共价连接(任选地通过接头)的il10m1;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链。(il10m1)1‑
异源fc形式的例示性蛋白质是xenp14246,其序列在图24中描绘。我们作为原型工程化的这种类别的另一这样的形式是异源fc
‑
(il10m1)1(图23f中描绘的卡通示意图),其包含:与第一异源二聚体fc链的c端共价连接(任选地通过接头)的il10m1;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体fc链。异源fc
‑
(il10m1)1形式的例示性蛋白质是xenp14247,其序列在图25中描绘。我们作为原型工程化的这种类别的另一形式是(il10m1)2‑
fc形式(图23a 中描绘的卡通示意图),其包含两个相同的单体,其中每个单体包含与同源二聚体fc链的n 端共价连接的il10m1。(il10m1)2‑
fc形式的例示性蛋白质是xenp25236,其序列在图26中描绘。我们作为原型工程化的这种类别的另一形式是fc
‑
(il10m1)2形式(图23b中描绘的卡通示意图),其包含两个相同的单体,其中每个单体包含与同源二聚体fc链的c端共价连接的 il10m1。
fc
‑
(il10m1)2形式的例示性蛋白质是xenp25237,其序列在图27中描绘。我们作为原型工程化的这种类别的另一形式是(il10m1)2‑
异源fc形式(图23c中描绘的卡通示意图),其包含:第一单体,其包含与进一步共价连接至第一异源二聚体fc链的n端(任选地通过接头)的第二il10m1共价连接(任选地通过接头)的第一il10m1;以及第二单体,其包含为“仅 fc”或“空fc”的互补的第二异源二聚体fc链。(il10m1)2‑
异源fc形式的例示性蛋白质是 xenp26887,其序列在图28中描绘。
[0615]
1a(d):分裂il10
‑
fc融合体
[0616]
我们推断,代替像在il10m1中那样在il10(a
‑
d)与il10(e
‑
f)结构域之间使用gly
‑
ser 接头,可通过将il
‑
10单体“分裂”成其两个结构域(即hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f),其序列分别描绘于图15a
‑
15b)来产生生物活性il
‑
10单体,并且il
‑
10结构域的二聚化将提供在本文称为“分裂il10”的生物活性分子。因此,我们设想了利用分裂il10的il
‑
10融合体,在此称为分裂il10
‑
fc融合体(图29中描绘的卡通示意图)。我们作为原型工程化的这种类别的一种这样的形式是分裂il10
‑
异源fc形式(图29a中描绘的卡通示意图),其包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的hil
‑
10(a
‑
d);以及第二单体,其包含与互补的第二异源二聚体fc链的n端共价连接的hil
‑
10(e
‑
f),使得异源二聚体fc链的二聚化迫使il
‑
10结构域二聚化。分裂il10
‑
异源fc形式的例示性蛋白质包括xenp25242,其序列在图30中描绘。我们作为原型工程化的这种类别的另一这样的形式是异源fc
‑
分裂il10形式 (图29b中描绘的卡通示意图),其包含:第一单体,其包含与第一异源二聚体fc链的c端共价连接(任选地通过接头)的hil
‑
10(a
‑
d)结构域;以及第二单体,其包含与互补的第二异源二聚体fc链的c端共价连接(任选地通过接头)的hil
‑
10(e
‑
f)结构域。异源fc
‑
分裂il10形式的例示性蛋白质包括xenp25879,其序列在图31中描绘。
[0617]
1b:原型il
‑
10融合蛋白的产生
[0618]
为了产生xenp24628(il10
‑
fc类别的例示性il
‑
10融合蛋白),通过标准基因合成构建编码il
‑
10单体的质粒,然后将其亚克隆到含有fc融合配偶体(例如同源二聚体fc链,如图 10所描绘)的ptt5表达载体中。
[0619]
为了产生xenp25880(scil10
‑
fc类别的例示性il
‑
10融合蛋白),通过标准基因合成构建编码scil
‑
10复合物的质粒,然后将其亚克隆到含有fc融合配偶体(例如第一异源二聚体fc 链,如图11所描绘)的ptt5表达载体中。还使用了编码空fc(例如,如图11所描绘的相应的第二异源二聚体fc链)的另一ptt5表达载体。
[0620]
为了产生xenp25236(il10m1
‑
fc类别的例示性il
‑
10融合蛋白),通过标准基因合成构建编码il10m1的质粒,然后将其亚克隆到含有fc融合配偶体(例如同源二聚体fc链,如图 10所描绘)的ptt5表达载体中。
[0621]
为了产生xenp25242(分裂il10
‑
fc融合体类别的例示性il
‑
10融合蛋白),通过标准基因合成构建编码hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f)的质粒,然后将其亚克隆到含有fc融合配偶体 (例如异源二聚体fc骨架,如图11所描绘)的ptt5表达载体中。
[0622]
通过在hek293e细胞中进行瞬时转染产生蛋白质,并且通过包括蛋白a色谱法(纯化部分1)然后是离子交换色谱法(纯化部分2)的两步纯化工艺对所述蛋白质进行纯化。
[0623]
1b(a):il
‑
10融合蛋白的纯化
[0624]
为了说明如上所述的il
‑
10融合蛋白的纯化,我们在此描述xenp25880的纯化和表
征。如上所述从hek293e上清液中纯化xenp25880。图32a描绘示出xenp25880的纯化部分2的色谱图(蛋白a色谱法之后的阳离子交换色谱法)。色谱图示出两个峰的分离(峰a和峰 b),通过利用多角度光散射的分析型尺寸排阻色谱法(asec
‑
mals)和分析型阳离子交换色谱法(aciex)来进一步表征所述峰的身份(identity)、纯度和同质性,如下文一般描述的。
[0625]
使用asec
‑
mals分析从xenp25880的纯化部分2中分离出的峰a和b,以推断出其组分蛋白物类。在agilent 1200高效液相色谱(hplc)系统上进行分析。利用280nm的uv检测波长,使用1
×
pbs,ph 7.4作为流动相在4℃下以1.0毫升/分钟的速度将样品注射到 superdex
tm 200 10/300gl柱(通用医疗生命科学公司(ge healthcare life sciences))上,持续25 分钟。在带有t
‑
rex折射率检测器的(加利福尼亚州圣塔芭芭拉的怀雅特技术公司(wyatt technology,santa barbara,cali.))上进行mals。使用agilentopenlab色谱数据系统(cds)chemstation版aic c.01.07版本和astra 6.1.7.15版本进行分析。图32b中描绘了描绘峰a和b的asec分离曲线的色谱图以及通过mals确定的组分物类的mw。曲线示出,峰b包含~92kd的主要物类(dominant species),这与xenp25880的计算分子量89.8kda(基于氨基酸序列)一致。
[0626]
还使用分析型ciex对来自纯化部分2的峰进行分析,以进一步评估峰a和b的纯度和同质性。在agilent 1200高效液相色谱(hplc)系统上进行分析。利用280nm的uv检测波长,在20mm mes、ph 6.0缓冲液中使用0
‑
40%nacl梯度以1.0毫升/分钟将样品注入到 proteomix scx
‑
np5 5μm无孔柱(特拉华州纽瓦克的赛分科技有限公司(sepax technologies, inc.,newark,del.))上。使用agilent openlab cds chemstation版aic c.01.07版本进行分析。图32c中描绘了描绘峰a和b的aciex分离的色谱图。值得注意的是,aciex分离显示,在峰b材料中,除了主峰(dominant peak)之外,还存在电荷变体。这将随后在实施例3d 中解决。
[0627]
1c:il
‑
10融合蛋白的生物活性
[0628]
接下来,我们研究了上述每个类别中的原型il
‑
10融合蛋白是否具有生物活性。在细胞因子与其受体结合后,与细胞因子受体缔合的janus激酶(jak)使stat蛋白磷酸化,然后所述stat蛋白转运到细胞核中以调节进一步的下游过程。具体地,il
‑
10与il
‑
10受体复合物结合并激活jak1和tyk2,它们使stat3磷酸化。因此,我们使用stat3磷酸化作为il10
‑ꢀ
fc形式(xenp24628)、scil10
‑
fc形式(xenp25880)、il10m1
‑
fc形式(xenp25236)和分裂il10
‑ꢀ
fc形式(xenp25242)的原型il
‑
10融合蛋白的生物活性的指标。
[0629]
使用100ng/ml抗cd3抗体(okt3;板结合)刺激人pbmc 2天以诱导il
‑
10受体的表达。刺激后,收获t细胞并用il
‑
10融合体处理15分钟。处理后,将pbmc在室温下用抗cd4
‑ꢀ
bv605(rpa
‑
t4)和抗cd8
‑
alexa700(sk1)抗体染色30
‑
45分钟。将细胞用预冷(
‑
20℃)的90%甲醇洗涤并孵育20
‑
60分钟。用甲醇孵育后,再次洗涤细胞并用抗cd45ra
‑
pe(hi100)、抗 cd25
‑
bv421(m
‑
a251)、抗foxp3
‑
alexa488(259d)和抗pstat3
‑
alexa647(py705)染色以标记各种群体和stat3磷酸化。通过流式细胞术分析pbmc,并且图33中描绘了描绘各种淋巴细胞群上的stat3磷酸化的数据。值得注意的是,数据显示,scil10
‑
fc形式(xenp25880)的原型il
‑
10融合体在各种淋巴细胞群上诱导stat3磷酸化(尽管不如重组il
‑
10有效),而il
‑ꢀ
10
‑
fc、il10m1
‑
fc和分裂il10
‑
fc形式的il
‑
10融合体仅显示基线水平的stat3磷酸化。
[0630]
1d:替代性c端fc融合体形式
[0631]
接下来,我们研究了本发明的融合蛋白中il
‑
10的几何形状和定位是否影响它们的活性。因此,我们生成了il
‑
10融合蛋白,其具有与fc区的c端共价连接的il
‑
10分子。
[0632]
我们作为原型工程化的scil10
‑
fc融合体类别的一种这样的形式是异源fc
‑
scil10形式 (图20b中描绘的卡通示意图),其包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的scil
‑
10;以及第二单体,其包含为“仅fc”或“空fc”的互补的第二异源二聚体 fc链。异源fc
‑
scil10形式的例示性蛋白质是xenp28741,其序列在图22中描绘。
[0633]
在此,我们比较了xenp28741和xenp25981(相应的scil10
‑
异源fc融合体作为比较物;其序列在图21中描绘)对stat3磷酸化的诱导。如实施例1b中总体描述的那样产生和纯化xenp28741和xenp25981。
[0634]
新鲜的pbmc用il
‑
10融合体处理15分钟。处理后,将pbmc在室温下用抗cd3
‑ꢀ
buv395(ucht1)、抗cd4
‑
bv605(rpa
‑
t4)、抗cd8
‑
alexa700(sk1)和抗cd14
‑
bv510(m5e2) 抗体染色30
‑
45分钟。将细胞用预冷(
‑
20℃)的90%甲醇洗涤并孵育20
‑
60分钟。用甲醇孵育后,再次洗涤细胞并用抗cd45ra
‑
pe(hi100)、抗cd25
‑
bv421(m
‑
a251)、抗foxp3
‑
alexa488 (259d)和抗pstat3
‑
alexa647(py705)染色以标记各种群体和stat3磷酸化。通过流式细胞术分析pbmc,并且图34中描绘了描绘各种淋巴细胞群上的stat3磷酸化的数据。数据显示,c端fc融合体xenp28741表现出比n端fc融合体xenp25981更大的诱导stat3磷酸化的效力,表明il
‑
10在本发明的融合蛋白中的位置确实影响它们的生物活性。
[0635]
实施例2:减弱il
‑
10融合蛋白的效力
[0636]
我们推断,降低il
‑
10融合蛋白对il10r1的亲和力(并且进而降低它们的效力)将减少抗原沉默并延长il
‑
10融合蛋白的半衰期。为此,我们设计了il
‑
10变体,目的是降低本发明的 il
‑
10融合蛋白的亲和力和效力,并在scil10
‑
fc融合体的背景下研究它们。
[0637]
2a:工程化il
‑
10变体以降低亲和力和效力
[0638]
使用与il
‑
10r1复合的人il
‑
10的晶体结构(如josephson等人(2001)所报道[pdb代码 1j7v])并且在molecular operating environment(moe;加拿大魁北克省蒙特利尔的化学计算集团公司(chemical computing group,montreal,quebec,canada))软件中建模,我们将以下鉴定为与il
‑
10r1接触的il
‑
10残基:n21、d28、q38、d41、d44、q42、n45、e142、d144和 e151(根据人il
‑
10成熟形式序列编号,如图1所描绘)。
[0639]
基于上文,我们设计了许多在接触残基处具有等构取代的il
‑
10变体(原因是等构取代具有较小的免疫原性潜力),目的是降低生物功能il
‑
10的效力。例示性取代包括n21d、d28n、 q38e、d41n、q42e、d44n、n45d、e142q、d144n和e151q。此外,yoon等人(2012)先前已有报道称,病毒il
‑
10(由eb病毒编码的il
‑
10同源物;其序列在图4中描绘)与人il
‑
10 相比对il
‑
10r1的亲和力要弱得多。我们研究了yoon等人所述的m39、l43和i87处的取代,其相应的病毒il
‑
10残基(即m39t、l43v和i87a)作为对照。可以设想,在il
‑
10单体、 il
‑
10结构域(例如hil
‑
10(a
‑
d)和hil
‑
10(e
‑
f))、il10m1或scil
‑
10的背景下,此处所述的取代可以单独或组合使用。包含如上所述的取代的例示性il
‑
10单体变体的序列在图35中描绘。应该注意的是,il
‑
10单体的取代编号是基于成熟形式il
‑
10序列。
[0640]
2b:变体il
‑
10对il
‑
10受体的亲和力
[0641]
为了研究实施例2a中描述的取代对于il
‑
10对il10r1的亲和力的影响,我们首先在il
‑ꢀ
10m1的背景下对取代进行工程化。例示性il10m1变体的序列在图36中描绘。应该注
culture kit,马里兰州洛克威尔(rockville,md.)的meso scale discovery)评估cd8
t细胞的ifnγ分泌。然后在冰上用抗cd8
‑
pe(rpa
‑
t8)和抗cd25
‑
bv421(m
‑
a251)对细胞染色 45
‑
60分钟。洗涤细胞并且使用ebioscience
tm foxp3/转录因子染色缓冲液套件(transcription factor staining buffer set)(赛默飞世尔科技公司(thermofisher scientific)),使用抗ki67
‑
apc 对细胞染色,并通过流式细胞术进行评估。
[0649]
如图43
‑
44中所描绘的数据显示,il
‑
10融合蛋白确实增强cd8
t细胞的ifnγ分泌,激活cd8
t细胞(如cd25表达所指示),并诱导cd8
t细胞的增殖(如表达ki67的细胞的百分比所指示)。与显示出一定范围的结合亲和力和诱导stat3磷酸化的效力的实施例2b
‑
2c 一致,具有亲和工程化的scil
‑
10变体的融合体在增强cd8
t细胞的ifnγ分泌、增殖和活化方面表现出一定范围的效力。值得注意的是,如图45a
‑
c所描绘,测试制品诱导stat3磷酸化的ec50与cd8
t细胞的ifnγ分泌、活化以及cd8
t细胞增殖的诱导的增强的ec50 正相关。
[0650]
实施例3:工程化scil10
‑
fc形式以优化产生
[0651]
在此,我们描述了改善scil10
‑
fc形式的il
‑
10融合体的产生的工程化尝试。
[0652]
3a:将scil
‑
10从一条fc链交换到另一条
[0653]
如图32a所描绘,峰a与b之间的分离不是最佳的。我们推断这是由于每种物类(异源二聚体相对于同源二聚体)的等电点并且将scil
‑
10复合物从异源二聚体fc区的一个臂(具有较高的pi)交换到异源二聚体fc区的另一个臂(具有较低pi)将在异源二聚体scil10
‑
异源fc物类与同源二聚体(scil10)2‑
fc和空fc物类之间提供更大的等电点差异。因此,我们研究了 xenp25238(图21中描绘的序列)的产生和纯化,其为具有与具有较低pi的异源二聚体fc链共价连接的scil
‑
10复合物的scil10
‑
异源fc融合体形式的il
‑
10融合体。如实施例1b中总体描述的那样产生和纯化xenp25238。图46a描绘示出xenp25238的纯化部分2的色谱图,其示出两个峰的分离(峰b和峰bc),通过asec
‑
mals和aciex进一步表征所述峰的身份、纯度和同质性,如实施例1b(a)总体描述和图46b
‑
c中所描绘的。分析显示峰b包含~88kd的主要物类。值得注意的是,如纯化部分2所指示,包含scil10
‑
异源fc融合体 xenp25238的峰b比包含空fc(pi( ):pi( ))同源二聚体的峰bc占优势。这代表了优于 xenp25880的改进,其中纯化部分2(如图32a中所描绘)产生比包含scil10
‑
异源fc融合体 xenp25880的峰b占优势的包含空fc(pi(
‑
):pi(
‑
))同源二聚体的峰a。与xenp25880一样, aciex分离显示,在包含xenp25238的峰b材料中,除了主峰之外,还存在电荷变体。
[0654]
如实施例1c中所述的那样进行评估stat3磷酸化的实验,以确定xenp25880的生物活性。如图47所描绘的数据显示,将scil
‑
10复合物从异源二聚体fc区的一条链交换到另一条链对活性没有影响。
[0655]
3b:在scil
‑
10构建体内工程化接头
[0656]
作为改善本发明的scil10
‑
fc融合蛋白的产生的另一种方法,我们研究了在scil
‑
10复合物内的第一il
‑
10单体与第二il
‑
10单体之间工程化不同长度的结构域接头。图48描绘了包含结构域接头的例示性scil
‑
10复合物的序列,并且图49描绘了包含scil
‑
10复合物的例示性scil
‑
10
‑
fc融合蛋白的序列,其如实施例1b中总体描述的那样产生和纯化。
[0657]
在scil
‑
10复合物内工程化接头似乎对scil10
‑
异源fc形式的il
‑
10融合体的产生没有实质影响(数据未示出);然而,活性也未受影响,如图50所描绘,其示出具有不同接头长度的 scil10
‑
异源fc融合体对stat3磷酸化的诱导(如实施例2c中描述的那样进行实
验)。
[0658]
3c:通过n45d(和n205d)取代实现的收率提高
[0659]
尽管引入n45d取代的目的是降低il
‑
10对il
‑
10受体的亲和力,但实施例2c中描绘的数据显示,含有n45d和n205d的scil10
‑
异源fc xenp25981表现出与wt scil10
‑
异源fcxenp25880相似的活性。然而,我们惊奇地发现n45d取代提高了scil10
‑
异源fc异源二聚体的收率。
[0660]
图52描绘了来自xenp25880和xenp25981的表达的收率。该图列出总收率(mg/l),其描绘通过蛋白a纯化的含fc蛋白的收率;同源二聚体和异源二聚体的百分比(通过蛋白a纯化的含fc蛋白的aciex分离确定;例示性色谱图描绘于图51中);以及同源二聚体和异源二聚体收率(mg/l),基于总收率和同源二聚体/异源二聚体的百分比计算。虽然收率存在一些批次间的差异,但数据显示xenp25981的产生通常导致scil10
‑
异源fc融合体的收率高于空 fc同源二聚体。
[0661]
为了研究是否是n45d(和n205d)修饰实现了异源二聚体收率提高,我们将n45d(和 n205d)修饰工程化到其他变体(例如xenp25986和xenp25993,其序列在图53中描绘)的背景中。如上所述,图54中描绘了来自xenp25880、xenp25981、xenp25986和xenp25993 的表达的收率。数据显示,相对于wt scil10
‑
异源fc xenp25880,除了q38e(在xenp25986 中)或q42e(在xenp25993中)之外,工程化n45d增强异源二聚体表达。
[0662]
3d:去除电荷变体
[0663]
如实施例1b(a)和3a中所示,scil10
‑
异源fc形式的分子(诸如xenp25880和 xenp25238)的纯化样品除主峰外还含有电荷变体。我们推断这种电荷异质性是由降解相关的翻译后修饰(ptm)(诸如贡献负电荷的脱酰胺作用)引起的。虽然脱酰胺作用通常是缓慢的并且在长期应力后发生(例如在储存期间),但我们假设将生物功能il
‑
10同源二聚体工程化为单链il
‑
10复合物和/或fc融合体扰乱il
‑
10结构,使得特定的天冬酰胺残基(诸如scil
‑
10复合物中的il
‑
10:il
‑
10共价连接界面处;或il
‑
10:fc共价连接界面处的那些)更易发生脱酰胺作用。因此,我们工程化了il
‑
10(和scil
‑
10)变体,其包括用丙氨酸或谷氨酰胺(尽管易发生脱酰胺作用,但远不如天冬酰胺)取代选定的天冬酰胺残基。
[0664]
具体地,先前已经指出,在小残基诸如甘氨酸、丝氨酸或丙氨酸前面的天冬酰胺最易发生脱酰胺作用(stephenson等人(1989))。因此,我们在残基n10、n92、n126和n160(以及 scil
‑
10复合物中的相应残基n170、n252、n286和n320)处工程化修饰。例示性的il
‑
10单体变体包括一个或多个以下取代:n10a、n10q、n92a、n126a、n160a。例示性的scil
‑
10 变体包括一个或多个以下取代:n10a、n92a、n126a、n160a、n170a、n170q、n252a、 n286a、n320a、k317_、i318_、r319_和n320_。例示性il
‑
10单体变体和相应的scil
‑
10变体的序列分别描绘于图55和56中。应该注意的是,il
‑
10变体的取代编号是基于成熟形式 il
‑
10序列,并且scil10变体的取代编号是基于scil
‑
10序列,如图15所描绘。还应注意,对于scil
‑
10变体,单体1和单体2中的取代不必相同。例如,对于命名为huil10.90的scil10 变体,第一单体在n160处不包含修饰,而第二单体在相应残基处包含修饰(即n320a)。
[0665]
可替代地,我们推断scil
‑
10复合物与fc链之间的结构域接头((g4s)2)提供另一脱酰胺位点(即,n
‑
g基序)。因此,我们工程化了xenp28295和xenp28904(不具有结构域接头的 scil10
‑
异源fc融合体)以及xenp28905(另一scil10
‑
异源fc融合体,其不具有结构域接头
并且另外包含具有如上所述的n10a和n170a取代的scil
‑
10变体),其序列在图59中描绘。
[0666]
除了脱酰胺作用之外,天冬氨酸异构化也可促成异质性。如本领域技术人员已知的,存在导致天冬氨酸残基更易发生异构化的特定基序(参见例如,sydow等人(2014),通过引用并入本文)。我们生成了例示性il
‑
10变体,其中残基d28被丙氨酸取代(il
‑
10单体变体和相应的scil
‑
10变体的序列描绘于图57
‑
58中)。
[0667]
scil
‑
10变体作为scil10
‑
异源fc融合蛋白(其序列在图59中描绘)的一部分,以及异源 fc
‑
scil10融合体(其序列在图60中描绘)的一部分工程化,并且如实施例1b中总体描述的那样产生和纯化。如实施例1b(a)中总体描述的那样通过aciex表征xenp28295、xenp28635 和xenp28641的蛋白a纯化样品,其色谱图描绘于图61中。在将图61a中描绘的xenp25981 的色谱图与图61b中描绘的xenp28295的色谱图进行比较时,我们注意到不具有n端接头的融合体的酸性峰消失,这可能是由于n320脱酰胺作用的消除。在将图61a中描绘的 xenp25981的色谱图与图61c中描绘的xenp28295和xenp28641的色谱图进行比较时,我们注意到具有包含n160a和n320a的scil
‑
10变体的融合体的酸性峰消失,这同样可能是由于n320脱酰胺作用的消除。出乎意料的是,如图61d的xenp28635的色谱图所示,n10a 取代(和相应的n170a取代)似乎去除了前延酸性峰(leading acidic peak)。
[0668]
重要的是,被工程化以去除潜在降解位点的scil
‑
10变体不影响il
‑
10融合体的活性,如通过stat3磷酸化的诱导(如实施例2b中描述的那样进行实验;数据描绘于图62中)以及cd8
t细胞的ifnγ分泌、cd8
t细胞增殖和cd8
t细胞活化的增强(如实施例2d中总体描述的那样进行实验;数据描绘于图63中)所指示的。
[0669]
3e:去除潜在n
‑
糖基化位点
[0670]
il
‑
10单体在残基116
‑
118(根据人il
‑
10成熟形式序列)处包括糖基化基序(n
‑
x
‑
s)。虽然已有报道称人il
‑
10在该位点不糖基化(westerhof等人(2012)),但我们设计了il
‑
10变体(和相应的scil
‑
10变体),以消融错误糖基化的潜力。因此,我们在以下残基处工程化修饰:n116、 k117、s118(根据人il
‑
10成熟形式序列),以及scil
‑
10复合物中相应的n276、k277和s278。例示性il
‑
10单体变体包括一个或多个以下取代:n116d、n116q、k117p和s118a。例示性 scil
‑
10变体包括一个或多个以下取代:n116d、n116q、k117p、s118a、n276d、n276q、 k227p和s228a。经过设计以去除潜在n
‑
糖基化位点的例示性il
‑
10单体变体和scil
‑
10变体的序列描绘于图63
‑
63中。scil
‑
10变体作为scil10
‑
异源fc融合蛋白(其序列在图66中描绘)的一部分工程化并且如实施例1b中总体描述的那样产生和纯化。
[0671]
3f:去除二硫桥
[0672]
il
‑
10单体包含两组二硫桥(由c12:c108和c62:c114形成),其稳定螺旋a
‑
d的组装 (walter等人,2014)。我们推断,二硫桥可能会无必要地使本发明的scil10
‑
fc融合蛋白的产生复杂化,并且例如增加异质性。因此,我们对修饰进行工程化以去除二硫桥。具有被引入以消融二硫桥形成的修饰的例示性il
‑
10单体变体(和相应的scil
‑
10变体)的序列描绘于图67
‑ꢀ
68中。scil
‑
10变体作为scil10
‑
异源fc融合蛋白(其序列在图69中描绘)的一部分工程化并且如实施例1b中总体描述的那样产生和纯化。
[0673]
3g:去除c端赖氨酸或甘氨酸
‑
赖氨酸
[0674]
作为改善本发明的il
‑
10融合蛋白的产生/纯化的另一方法,il
‑
10融合蛋白被工程化为去除了fc区c端的赖氨酸(k)或甘氨酸
‑
赖氨酸(gk)。去除了c端k或gk的例示性il
‑
10
融合蛋白的序列散布在整个图中,并具有注释g446del和/或k447del。
[0675]
实施例4:进一步工程化il10m1
‑
fc形式
[0676]
如实施例1c中所述,原型(il10m1)2‑
fc融合体xenp25236在pstat3测定中是没有生物活性的,尽管单独的il10m1分子具有活性,如josephson等人(2000)所述。我们推断,在融合于fc区的背景下保留il10m1的活性需要il10m1分子的进一步工程化。在此,我们描述了il10m1变体的设计,目的是恢复il10m1
‑
fc类别的融合体的活性。应注意,实施例4 的以下小节中的氨基酸编号是根据人il
‑
10成熟形式序列(编号中不包括il10m1接头)。
[0677]
4a:工程化il10m1中的二硫桥
[0678]
在一种方法中,我们推断,鉴于分子内结构域交换(即同一il10m1分子内的螺旋a
‑
d和螺旋e
‑
f)而不是分子间结构域交换(即两个单独il
‑
10单体的螺旋a
‑
d与螺旋e
‑
f之间),稳定螺旋a
‑
d的二硫桥(walter等人,2014)不再是必要的,并且实际上可能对il10m1
‑
fc融合体的活性有害,例如,通过错误折叠。因此,我们设计了il10m1变体,其在c12、c62、c108 和/或c114处包含修饰以去除二硫桥。例示性il10m1变体描绘于图68中,并且包含变体的例示性il10m1
‑
fc融合体描绘于图69中。
[0679]
可替代地,可能鉴于分子内结构域交换(即同一il10m1分子内的螺旋a
‑
d和螺旋e
‑
f)而不是分子间结构域交换(即两个单独il
‑
10单体的螺旋a
‑
d与螺旋e
‑
f之间),需要另外稳定螺旋a
‑
d与螺旋e
‑
f之间的相互作用。基于josephson等人(2000)描述的il10m1的晶体结构 (pdb代码1y6k),我们将以下残基鉴定为适合用半胱氨酸残基取代:螺旋a
‑
d中的f37、 q38、d41、l47、l48、s51、d55、f56、a64、m68和v76;以及s118、a120、v121、k138、 a139、m140、s141、e142和y153。具体地,我们设想在以下残基对之间引入二硫桥:f37c: m140c、q38c:s141c、d41c:k138c、l47c:k138c、l48c:e142c、s51c:a120c、d55c: a120c、f56c:y153c、a64c:s118c、m68c:v121c和v76c:a139c,以促进螺旋a
‑
d与螺旋e
‑
f之间的相互作用的稳定。具有工程化的二硫桥的例示性il10m1变体描绘于图70 中,并且包含变体的例示性il10m1
‑
fc融合体描绘于图71中。
[0680]
4b:稳定性变体
[0681]
与以上稳定il10m1分子的想法一致,我们使用moe软件中的建模来鉴定核心残基中可有助于增加稳定性的单个取代。我们鉴定为适于取代的核心残基包括:l19、l23、l26、a29、 f30、v33、f37、l47、l48、l52、f56、a64、l65、s66、m68、i69、f71、y72、v76、m77、 a80、e81、i87、v91、l94、g95、l98、l101、l105、l112、s118、v121、v124、f128、 k138、a139、m140、s141、f143、f146、i147和i150。使用moe软件中的残基扫描(residuescanning)模块,我们在以上鉴定的残基处进行计算机模拟单点突变,以预测它们对于il10m1 的稳定性及其对il10r1的亲和力的影响,其数据在图72a
‑
aq中示出。
[0682]
此外,我们使用rosetta软件套件(rosettacommons)进行进一步的稳定性建模。il
‑
10二聚体结构是根据rosetta的松弛方案(rosetta’s relax protocols)制备的。接下来,我们运行了各自100个轨迹的多个固定骨架优化方案,以探索在单个起始结构的背景中以及在结构集合 (ensemble)中允许疏水性残基(相对于所有氨基酸残基)作为独立突变或组合的影响。我们还使用了一种灵活骨架方法,其在设计与骨架移动之间进行迭代。我们仅使用了每条链的核心中的如上文所鉴定的位点。
[0683]
出于说明目的,我们设计了il10m1变体并产生具有突变(即l47q、s118a和a139q)
的 il10m1和相应的il10m1
‑
fc融合体,所述突变被一致地预测为改善上述不同方法之间的稳定性,其序列分别描绘于图73
‑
74中。
[0684]
4c:工程化用于il10m1的替代性接头
[0685]
接下来,我们设计了具有不同接头的il10m1变体。在一种方法中,我们旨在通过引入脯氨酸残基、去除丝氨酸残基或缩短接头长度来增加接头的刚性。考虑到这种方法设计的例示性il10m1变体的序列描绘于图75中。可替代地,我们旨在增加接头的柔性,例如,通过延长接头长度。考虑到这种方法设计的例示性il10m1变体的序列还描绘于图75中。具有这些例示性il10m1变体的il10m1
‑
fc融合体的序列描绘于图76中。
[0686]
实施例5:包含具有基于109h的序列的il
‑
10变体的il
‑
10融合体
[0687]
大多数上述il
‑
10融合体及其组分il
‑
10变体是在il
‑
10(109l)序列的背景下生成的。我们在il
‑
10(109h)序列的背景下工程化了例示性il
‑
10变体(其序列描绘于图77中)。接下来,我们工程化了具有il
‑
10(109h)变体的scil10
‑
异源fc和异源fc
‑
scil10形式的il
‑
10融合体,其序列描绘于图78和79中,以确定不同的il
‑
10序列背景是否影响il
‑
10融合体的活性。这些il
‑
10融合体如实施例1b中总体描述的那样产生。
[0688]
具有il
‑
10(109h)变体的il
‑
10融合体的活性在如上文总体描述的pstat3测定中确定,其数据描绘于图80
‑
85中。数据显示,基于il
‑
10(109h)的il
‑
10融合体在各种细胞类型上保持一定范围的效力。令人惊讶的是,数据显示,与基于il
‑
10(109l)序列的相应il
‑
10融合体 (例如相同的变体)相比,基于il
‑
10(109h)序列的各il
‑
10融合体在所研究的各细胞类型上表现出较低功效(即stat3磷酸化的最大诱导)。这表明il
‑
10的位置109处的特定残基可能对于il
‑
10融合体活性的调节很重要。
[0689]
实施例6:进一步工程化il
‑
10融合蛋白以增加半衰期
[0690]
为了进一步增加半衰期,利用xtend fc(m428l/n434s取代)对本发明的il
‑
10融合蛋白进一步工程化。具有xtend的例示性il
‑
10融合蛋白的序列描绘于图86中。
[0691]
实施例7:靶向性il
‑
10融合蛋白的工程化
[0692]
如上所述,我们设想使il
‑
10选择性地靶向表达检查点受体(诸如pd
‑
1)的til,或使il
‑ꢀ
10选择性地靶向cd8
t细胞。因此,我们设想了多种靶向性il
‑
10形式,其卡通示意图描绘于图87
‑
90中。
[0693]
图87描绘基于il10
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性 il10
‑
fc”类别。
[0694]
图88描绘基于scil10
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性 scil10
‑
fc”类别。我们工程化的这种类别的一种这样的形式是“抗x x scil10
‑
异源fc”形式,其包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头)的 scil
‑
10。
[0695]
图89描绘基于il10m1
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性 il10m1
‑
fc”类别。我们工程化的这种类别的一种这样的形式是“抗x x(il10m1)1‑
异源fc”形式,其包含:第一单体,其包含与第一异源二聚体fc链的n端共价连接的抗原结合结构域;以及第二单体,其包含与第二异源二聚体fc链的n端共价连接(任选地通过结构域接头) 的il10m1。我们工程化的这种类别的另一这样的形式是“mab
‑
(il10m1)
2”,其包含两个
相同的单体,每个单体包含与同源二聚体fc链的n端共价连接的抗原结合结构域,所述同源二聚体fc链通过c端任选地通过接头与il10m1共价连接。
[0696]
图90描绘基于分裂il10
‑
fc类别的靶向性il
‑
10融合体的例示性形式,本文称为“靶向性分裂il10
‑
fc”类别。
[0697]
实施例8:靶向cd8的il
‑
10融合蛋白
[0698]
如上所述,il
‑
10主要通过cd8
t细胞增强癌症免疫。实际上,如chan等人(2015)所描述,il
‑
10与大量pbmc一起孵育导致ifnγ分泌的抑制,表明il
‑
10的免疫抑制作用通过非 cd8
细胞类型增强。另外,til中的高cd8/cd4 t细胞比率通常被认为是肿瘤治疗的良好预后标志。因此,我们推断,对靶向cd8的il
‑
10融合蛋白进行工程化,以通过选择性地靶向 cd8
t细胞来增强il
‑
10的免疫刺激潜力将是有用的。
[0699]
8a:原型靶向cd8的il
‑
10融合蛋白的产生
[0700]
可用于本发明的靶向cd8的il
‑
10融合蛋白的例示性抗cd8和抗nkg2d可变区描绘于图92和93中。
[0701]
通过标准基因合成构建编码il
‑
10组分或抗cd8或抗nkg2d可变区的质粒,然后将其亚克隆到含有融合配偶体(例如,骨架和/或ch1/部分铰链,如图10、11和/或12中所描绘)的 ptt5表达载体中。通过在hek293e细胞中进行瞬时转染产生蛋白质,并且通过包括蛋白a 色谱法(ge医疗)和离子交换色谱法的两步纯化工艺对所述蛋白质进行纯化。
[0702]“抗x x(il10m1)1‑
异源fc”的例示性的靶向cd8的il
‑
10融合体的序列描绘于图94 中。“mab
‑
(il10m1)
2”形式的例示性的靶向cd8的il
‑
10融合体的序列描绘于图95中。“抗x x scil10
‑
异源fc”形式的例示性的靶向cd8和靶向nkg2d的il
‑
10融合体的序列分别描绘于图96和98中。包含il
‑
10变体的“抗x x scil10
‑
异源fc”的例示性的靶向cd8的 il
‑
10融合体的序列描绘于图97中。
[0703]
8b:靶向cd8的il
‑
10融合蛋白的生物活性
[0704]
我们在如实施例1d中总体描述的那样进行的实验中研究了原型靶向cd8的il
‑
10融合体在各种淋巴细胞群上对stat3磷酸化的诱导,其数据描绘于图99中。数据显示,靶向cd8 的il
‑
10融合体xenp25794比非靶向性il
‑
10融合体(xenp25880)更有效地在cd8
t细胞上诱导stat3磷酸化。值得注意的是,对于所研究的测试制品浓度范围,靶向cd8的il
‑
10 融合体在cd4
t细胞上诱导stat3磷酸化的效力似乎低于xenp25880。此外,与cd4
t 细胞相比,靶向nkg2d的il
‑
10融合体(xenp25952)还显示出对cd8
t细胞的选择性,表明可使用除cd8之外的其它靶标来使il
‑
10融合蛋白靶向cd8
t细胞。
[0705]
实施例9:靶向pd
‑
1的il
‑
10融合蛋白
[0706]
如上所述,免疫检查点蛋白诸如pd
‑
1在肿瘤浸润淋巴细胞中上调。因此,靶向pd
‑
1的 il
‑
10融合蛋白被设想为在肿瘤环境中选择性地靶向t细胞。
[0707]
9a:原型靶向pd
‑
1的il
‑
10融合蛋白的产生
[0708]
可用于本发明的靶向pd
‑
1的il
‑
10融合蛋白的例示性抗pd
‑
1可变区描绘于图100中。
[0709]
通过标准基因合成构建编码il
‑
10组分或抗pd
‑
1可变区的质粒,然后将其亚克隆到含有融合配偶体(例如,骨架和/或ch1/部分铰链,如图10、11和/或12中所描绘)的ptt5表达载体中。通过在hek293e细胞中进行瞬时转染产生蛋白质,并且通过包括蛋白a色谱法(ge
医疗)和离子交换色谱法的两步纯化工艺对所述蛋白质进行纯化。
[0710]“抗x x scil10
‑
异源fc”形式的例示性的靶向pd
‑
1的il
‑
10融合体的序列描绘于图101 中。此外,包含il
‑
10变体的“抗x x scil10
‑
异源fc”形式的例示性的靶向pd
‑
1的il
‑
10融合体的序列描绘于图102中。“抗x x异源fc
‑
scil10”形式的靶向pd
‑
1的il
‑
10融合蛋白的序列描绘于图103中。
[0711]
9b:非竞争性pd
‑
1结合结构域
[0712]
另外,因为将本发明的靶向性il
‑
10融合蛋白与pd
‑
1阻断抗体组合,或在用pd
‑
1阻断抗体处理后施用本发明的靶向性il
‑
10融合蛋白将是有用的,因此重要的是,靶向性il
‑
10融合蛋白的pd
‑
1靶向臂不与pd
‑
1阻断抗体结合相同或相似的表位。本文考虑的pd
‑
1阻断抗体包括但不限于纳武单抗和派姆单抗。
[0713]
预期用于本发明的靶向pd
‑
1的il
‑
10融合蛋白的例示性的非竞争性抗pd
‑
1结合结构域被称为mab a、mab b和mab c(它们的人源化可变区的序列描绘于图104中,人源化使用串含量优化(string content optimization)进行(参见,例如,2010年2月2日发布的美国专利号 7,657,380))。
[0714]
进行串联表位分箱以证明mab不与纳武单抗和派姆单抗竞争。使用octet htx仪器进行表位分箱。首先使用amc(抗小鼠fc)生物传感器捕获人pd
‑
1的鼠fc融合体,将其浸入100 nm的第一抗体(如图105左侧所指示)中,并且接着浸入100nm的第二抗体(如图105顶部所指示)中。针对将生物传感器浸入hbs
‑
ep缓冲液中,然后浸入抗pd
‑
1抗体中的bli响应对 bli响应进行归一化。如果抗体对提供的归一化bli响应小于0.5,则认为该对竞争或部分竞争且位于同一表位箱中,即识别非常相似或高度重叠的表位。如果抗体对提供的归一化bli 响应大于0.5,则认为该对是非竞争性的,并被分箱至不同的表位。所测试的抗体是基于纳武单抗的二价抗pd
‑
1mab、内部制备的派姆单抗、嵌合mab a、嵌合mab b和嵌合mab c。还包括pd
‑
l1
‑
fc以研究抗体对pd
‑
1:pd
‑
l1相互作用的阻断。分箱显示抗pd
‑
1mab a、mabb和mab c不与纳武单抗或派姆单抗竞争。另外,mab a似乎不阻断pd
‑
1:pd
‑
l1相互作用,而mab b和mab c是pd
‑
1:pd
‑
l1相互作用的部分阻断剂。利用基于人源化mab c的pd
‑
1 靶向臂的例示性的靶向pd
‑
1的il
‑
10融合体的序列描绘于图106中。
[0715]
9c:靶向pd
‑
1的il
‑
10融合蛋白的生物活性
[0716]
我们在如实施例1c中总体描述的那样进行的实验中研究了原型靶向pd1的il
‑
10融合体在各种淋巴细胞群上对stat3磷酸化的诱导,其数据描绘于图107中。应注意,对于该实验,首先使用100ng/ml抗cd3抗体(okt3;板结合)激活pbmc 2天,以诱导pd
‑
1的表达并模拟肿瘤环境。数据显示,靶向pd
‑
1的il
‑
10融合体比非靶向性il
‑
10融合体(xenp25880) 更有效地在各种t细胞群上诱导stat3磷酸化。
[0717]
我们还研究了原型靶向pd
‑
1的il
‑
10融合体对cd8
t细胞群的ifnγ分泌和增殖的增强。对于这些研究,我们使用利用easysep
tm
人cd8
t细胞富集试剂盒(加拿大温哥华的干细胞技术公司)从pbmc中纯化的cd8
t细胞。首先将cd8
t细胞与10μg/ml的固定化抗 cd3抗体(okt3)和2μg/ml的固定化抗cd28抗体(28.2)一起孵育3天,以诱导cd8
t细胞上的il
‑
10受体表达。然后用指定的测试制品激发cd8
t细胞3天,然后用1μg/ml可溶性抗cd3抗体刺激6小时。收获上清液并使用人ifn
‑
γ组织培养试剂盒(马里兰州洛克威尔的 meso scale discovery)评估cd8
t细胞的ifnγ分泌。然后在冰上用抗cd8
‑
pe(rpa
‑
t8)和抗 cd25
‑
bv421(m
‑
a251)对细胞染色45
‑
60分钟。洗涤细胞并且使用ebioscience
tm foxp3/转录因子染色缓冲液套件(赛默飞世尔科技公司),使用抗ki67
‑
apc对细胞染色,并通过流式细胞术进行评估。示出cd8
t细胞的ifnγ分泌、cd8
t细胞增殖和cd8
t细胞活化的数据描绘于图108中。
[0718]
9d:pd
‑
1靶向恢复效力降低的il
‑
10
‑
fc融合体在诱导cd8 t细胞增殖上的效力
[0719]
接下来,研究了pd
‑
1靶向对效力降低的il
‑
10
‑
fc融合体的影响。首先将cd8
t细胞与 10μg/ml的固定化抗cd3抗体(okt3)和2μg/ml的固定化抗cd28抗体(28.2)一起孵育3 天,以诱导cd8
t细胞上的il
‑
10受体表达。然后用指定的测试制品激发cd8
t细胞3天,然后用1μg/ml可溶性抗cd3抗体刺激6小时。然后在冰上用包括抗cd8
‑
bv421(sk1)的染色抗体对细胞染色45
‑
60分钟。接下来,洗涤细胞并且使用ebioscience
tm foxp3/转录因子染色缓冲液套件(赛默飞世尔科技公司),使用抗ki67
‑
apc对细胞染色,并通过流式细胞术进行评估。所研究的测试制品是xenp30007(scil10.104
‑
fc)、xenp30008(scil10.105
‑
fc)、xenp30010(scil10.107
‑
fc)、xenp30520(αpd
‑
1(mab c_h1_l1.1)x huil10.104))、xenp30521 (αpd
‑
1(mab c_h1_l1.1)x huil10.105))和xenp30522(αpd
‑
1(mab c_h1_l1.1)xhuil10.107))。
[0720]
示出cd8
t细胞增殖(如通过为ki67 的cd8
t细胞的百分比所指示)的数据描绘于图 109中。对于每种效力降低的il
‑
10
‑
fc融合体,通过pd
‑
1靶向恢复诱导cd8
t细胞增殖的效力。据推测,被靶向而增殖的cd8
t细胞是表达pd
‑
1的cd8
t细胞,类似于将在肿瘤环境中发现的那些。
[0721]
9e:进一步调整靶向pd
‑
1的il
‑
10
‑
fc融合体的效力
[0722]
接下来,在如上文总体描述的stat3磷酸化测定中研究不同的pd
‑
1靶向臂以及靶向pd
‑ꢀ
1的il
‑
10
‑
fc融合体的形式的影响。所研究的测试制品使用基于mab c_h1_l1.1的pd
‑
1靶向臂、基于1c11的pd
‑
1靶向臂,或基于numax/莫维珠单抗的rsv靶向臂;较弱的il
‑
10 变体(il10.105)或较强的il
‑
10变体(il10.102);以及n端scil
‑
10相对于c端scil
‑
10。
[0723]
示出stat3的磷酸化的数据描绘于图110中。在每种情况下,靶向pd
‑
1的il
‑
10
‑
fc融合体比相应的靶向rsv的il
‑
10
‑
fc融合体更有效。在大多数但并非所有情况下,基于1c11 的pd
‑
1靶向臂比基于mab c_h1_l1.1的pd
‑
1靶向臂在更大程度上增强效力。值得注意的是,对于效力较弱的il
‑
10
‑
fc融合体(具有il10.105),无论特定靶向臂如何,scil
‑
10的c端连接都导致较低的效力;而对于效力较强的il
‑
10
‑
fc融合体(具有il10.102),无论特定靶向臂如何,scil
‑
10的c端连接都导致较大的效力。
[0724]
实施例10:靶向tigit的il
‑
10融合蛋白
[0725]
如上所述,免疫检查点蛋白诸如tigit在肿瘤浸润淋巴细胞中上调。因此,靶向tigit 的il
‑
10融合蛋白被设想为在肿瘤环境中选择性地靶向t细胞。
[0726]
10a:生成tigit结合结构域
[0727]
重组人tigit和食蟹猴tigit(图111中描绘的序列)用于噬菌体淘选。建立在噬菌体外壳蛋白piii上展示fab和scfv变体的内部从头噬菌体文库(in
‑
house de novo phage libraries) (在此分别称为“fab文库”和“scfv文库”)。fab文库和scfv文库均如下淘选五轮:1)人 tigit,2)食蟹猴tigit,3)人tigit,4)食蟹猴tigit,以及5)人tigit,严格程度不断增加 (就抗原浓度以及洗涤严格性而言)。在每轮之后,将洗脱的噬菌体添加至对数期xl1
‑
blue细胞(特拉华州威尔明顿的安捷伦(agilent,wilmington,del.))并在37℃、
250rpm下扩增过夜。
[0728]
对来自fab文库和scfv文库两者的淘选第3轮、第4轮和第5轮中的每一者的192个克隆进行测序,从而产生1,152个克隆。我们研究了这些克隆在tigit转染的jurkat细胞(此处称为jurkat
‑
tigit)上的细胞表面结合。在384孔板上每孔使用100k jurkat
‑
tigit细胞,将其与噬菌体上清液或对照抗tigit
‑
apc(加利福尼亚州圣地亚哥(san diego,calif.)的biolegend) 一起在4℃下孵育1小时并洗涤。然后用二抗对细胞染色并通过ique screener(新墨西哥州阿尔伯克基(albuquerque,n.mex.)的intellicyt)分析噬菌体结合。图115
‑
116描绘了例示性板读出结果和相关直方图(380个克隆 4个对照),其中对照(对照抗tigit mab、亲本k07噬菌体 染色抗体、仅染色抗体以及仅细胞)和我们最喜欢的克隆(即2a5b4;其序列在图112中作为二价mab描绘)突出显示。基于对指示噬菌体克隆与jurkat
‑
tigit细胞的结合的mfi和直方图的分析,从1,152个克隆中选择92个唯一克隆。
[0729]
通过gibson组装构建含有92个克隆的可变重和可变轻结构域的质粒,并将所述质粒亚克隆到含有igg1恒定区的编码序列的ptt5表达载体中(使用 e233p/l234v/l235a/g236del/s67k消融变体)。在hek293e中转染dna以进行表达,并且使用蛋白a色谱法从上清液中纯化所得的二价mab。
[0730]
接下来,我们研究了92种二价mab与jurkat
‑
tigit的结合。将jurkat
‑
tigit细胞与指定浓度的xenp27507和噬菌体来源的抗tigit mab以及对照抗体xenp19351(基于10a7 的二价抗tigit mab,如wo 2015/009856中所述,具有e233p/l234v/l235a/g236del/s67k 消融变体;其序列描绘于图113中)一起在4℃下孵育1小时。然后将细胞用alexa647 affinipure f(ab')2片段山羊抗人igg、fcγ片段特异性二抗(jackson immunoresearch,west grove, penn.)在4℃染色1小时,并通过流式细胞术进行分析。说明xenp27507与5种其它噬菌体来源的mab的结合的数据描绘于图117中,并且示出每种噬菌体来源的mab都能够结合至 jurkat
‑
tigit细胞。
[0731]
最后,我们使用基于biolayer干涉技术(bli)的方法octet针对tigit对92种二价mab 进行亲和筛选。octet的实验步骤通常包括以下内容:固定化(将配体捕获到生物传感器上);缔合(将配体涂布的生物传感器浸入含有分析物的孔中);以及解离(将生物传感器返回到含有缓冲液的孔中)以确定测试制品的亲和力。在数据处理期间用于背景校正的方法中还包括仅含有缓冲液的参考孔。具体地,使用his1k生物传感器捕获带有his标签的人tigit或带有 his标签的食蟹猴tigit,并浸入100nm的每种二价mab中。通过用1:1langmuir结合模型对结合数据进行全局拟合来进行动力学分析。xenp27507、选定的5种噬菌体来源的mab和对照mab xenp19351的所得表观解离常数(k
dapp
)、缔合速率(k
a
)和解离速率(k
d
)描绘于图118 中。基于人和食蟹猴tigit的k
dapp
(k
d
表观,由于亲合力问题)值对克隆进行排序。数据显示,每种噬菌体来源的mab以一定范围的表观k
d
与人和食蟹猴tigit结合。
[0732]
基于以上研究细胞表面tigit结合和k
dapp
的实验,我们鉴定了~20个合适的tigit abd (从1,152个初始序列中),其具有细胞表面抗原结合和亲和力的良好平衡。
[0733]
10a(a):tigit阻断的表征
[0734]
检查点受体/配体相互作用的阻断对于t细胞无反应性的抑制是必要的。因此,靶向性il
‑ꢀ
10融合蛋白的tigit靶向臂能够阻断cd112和pvr与tigit的结合可能是有用的。
[0735]
在第一实验中,我们使用octet进行串联表位分箱,以研究噬菌体来源的mab是否
与 xenp19351(tigit配体的经确认阻断剂)或xenp19352(tigit配体的经确认非阻断剂;其序列描绘于图114中)结合相同的表位,推断与xenp19351分箱至相同表位的噬菌体来源的 mab是tigit/tigit配体相互作用的阻断剂。首先将his1k生物传感器浸入xenp23423中,然后浸入100nm的xenp19351或xenp19352中,并且接着浸入100nm的指定测试制品中。针对将生物传感器浸入hbs
‑
ep缓冲液中,然后浸入抗体中的bli响应对bli响应进行归一化。如果抗体对提供的bli响应小于0.5,则认为该对竞争且位于同一表位箱中,即识别非常相似或高度重叠的表位。如果抗体对提供的归一化bli响应大于0.5,则认为该对是非竞争性的,并被分箱至不同的表位。图119中概括了每个抗体对的归一化的bli响应。数据显示,每种噬菌体来源的mab都被分箱至与xenp19351相同的表位(如低于0.5的bli响应所指示)和与xenp19352不同的表位(如大于0.5的bli响应所指示),表明噬菌体来源的mab 是tigit:tigit配体相互作用的阻断剂。
[0736]
为了证实上文,我们在基于细胞的测定中研究了tigit与细胞表面cd112和pvr的结合的阻断。将指定浓度的噬菌体来源的mab、阳性对照抗体xenp19351(tigit:tigit配体相互作用的经确认阻断剂)或阴性对照抗rsv mab xenp15074与1μg/ml的人tigit的鼠 fc融合体(tigit
‑
mfc)在室温下混合30分钟。然后添加cd112转染或pvr转染的cho细胞,并在4℃下孵育30分钟。洗涤细胞并用抗鼠fc
‑
a647抗体染色以检测tigit
‑
mfc与cho 细胞上的cd112或pvr的结合,并通过流式细胞术进行分析。描绘tigit
‑
mfc与cd112转染的cho细胞或pvr转染的cho细胞的结合的数据分别描绘于图120a
‑
b中,并证实噬菌体来源的mab(包括基于2a5b4的mab)能够阻断tigit与pvr和cd112的结合。在类似地进行的另一个实验中,研究了pvr和cd112的阻断百分比,其数据描绘于图121a
‑
b中。
[0737]
10b:原型靶向tigit的il
‑
10融合蛋白的产生
[0738]
使用如上所述的抗tigit可变区构建本文所述的原型靶向tigit的il
‑
10融合蛋白。可用于本发明的靶向tigit的il
‑
10融合蛋白的另外的抗tigit可变区的序列描绘于图122中。
[0739]
通过标准基因合成构建编码il
‑
10组分或抗tigit可变区的质粒,然后将其亚克隆到含有融合配偶体(例如,骨架和/或ch1/部分铰链,如图10、11和/或12中所描绘)的ptt5表达载体中。通过在hek293e细胞中进行瞬时转染产生蛋白质,并且通过包括蛋白a色谱法(ge 医疗)和离子交换色谱法的两步纯化工艺对所述蛋白质进行纯化。
[0740]“抗x x scil10
‑
异源fc”形式的例示性的靶向tigit的il
‑
10融合体的序列描绘于图 123中。
[0741]
10c:tigit靶向恢复效力降低的il
‑
10
‑
fc融合体在诱导cd8 t细胞增殖上的效力
[0742]
接下来,研究了tigit靶向对效力降低的il
‑
10
‑
fc融合体的影响。首先将cd8
t细胞与10μg/ml的固定化抗cd3抗体(okt3)和2μg/ml的固定化抗cd28抗体(28.2)一起孵育3 天,以诱导cd8
t细胞上的il
‑
10受体表达。然后用指定的测试制品激发cd8
t细胞3天,然后用1μg/ml可溶性抗cd3抗体刺激6小时。然后在冰上用包括抗cd8
‑
bv421(sk1)的染色抗体对细胞染色45
‑
60分钟。接下来,洗涤细胞并且使用ebioscience
tm foxp3/转录因子染色缓冲液套件(赛默飞世尔科技公司),使用抗ki67
‑
apc对细胞染色,并通过流式细胞术进行评估。所研究的测试制品是xenp30007(scil10.104
‑
fc)、xenp30008(scil10.105
‑
fc)、 xenp30010(scil10.107
‑
fc)、xenp30523(αtigit(2a5b4)x huil10.104))、xenp30524 (α
tigit(2a5b4)x huil10.105))和xenp30525(αtigit(2a5b4)x huil10.107))。
[0743]
示出cd8
t细胞增殖(如通过为ki67 的cd8
t细胞的百分比所指示)的数据描绘于图 124中。对于每种效力降低的il
‑
10
‑
fc融合体,通过tigit靶向恢复诱导cd8
t细胞增殖的效力。据推测,被靶向而增殖的cd8
t细胞是表达tigit的cd8
t细胞,类似于将在肿瘤环境中发现的那些。
[0744]
提供上述实施例以向本领域中的技术人员给出如何实施和使用本发明的组合物、系统和方法的实施方案的完整公开和说明,并非意图限制发明人对于他们的发明的主张范围。用于实施本发明的上述模式的修改为熟练的技术人员所显见,并且意图属于以下权利要求范围之内。本说明书中所提及的所有专利和出版物指示本发明所属领域的技术人员的水平。本公开中引用的所有参考文献通过引用并入,其程度如同每个参考文献已经以全文引用的方式单独地并入。
[0745]
使用所有标题和章节命名仅仅是出于清楚和参考的目的并且不应当被认为是以任何方式进行限制。举例而言,本领域中的技术人员应了解根据本文所述的本发明的精神和范围按需要组合来自不同标题和章节的各种方面的有用性。
[0746]
本文所引证的所有参考文献都以引用的方式且出于所有目的完整并入本文中,犹如个别公布或专利或专利申请特别且个别明示出于所有目的以引用的方式整体并入本文。如本领域中的技术人员显而易见,在不背离本技术的精神和范围的情况下可对其进行许多修改和变化。本文所述的特定实施方案和实施例仅以实施例方式提供,并且本技术仅由随附权利要求的术语以及权利要求有权主张的等效物的全部范围限制。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。