一种管理玉米螟对bt毒素抗性的方法
(一)技术领域
1.本发明涉及一种管理鳞翅目害虫玉米螟,包括欧洲玉米螟(ostrinia nubilalis)和亚洲玉米螟(ostrinia furnacalis)对苏云金芽孢杆菌bt毒素发生抗性的方法。
(二)
背景技术:
2.害虫给全球农业生产带来每年数十亿美元的损失。害虫的防治目前主要依靠使用化学农药和推广转基因作物。化学农药的残留会对人体健康和环境带来不良影响。利用生物技术方法培育含有抗虫基因的转基因抗虫农作物,可以大幅度降低化学杀虫剂的使用,有效保护农作物免受害虫为害。杀虫蛋白质有多种,目前使用最多的是来自苏云金芽孢杆菌(bacillus thuringiensis,bt)的杀虫晶体蛋白,如cry1a、cry1f等;以及bt营养期杀虫蛋白,如vip3等(estruch,warren et al.1996,adang,crickmore et al.2014)。bt杀虫蛋白质在转基因植物上的应用已经超过二十年,2017年全球抗虫转基因作物的应用面积超过7000万公顷(isaaa 2018)。
3.杀虫晶体蛋白是发现最早、使用最广泛的bt杀虫蛋白质。目前商业化应用的抗虫转基因玉米中使用较多的杀虫晶体蛋白是cry1ab,cry1fa和cry2ab等,这些杀虫蛋白质对部分鳞翅目昆虫如玉米螟、棉铃虫、小菜蛾、二化螟、三化螟、大螟、稻纵卷叶螟等具有较高的杀虫活性。其中,转cry1ab基因玉米、转cry1ac基因棉花已经被广泛应用。frankenhuyzen和jakka等总结多项研究的生物活性测定数据后指出,cry1ab\1ac蛋白对鳞翅目螟蛾科害虫的杀虫活性较高,但是对多种鳞翅目夜蛾科害虫(如草地贪夜蛾、甜菜夜蛾、小地老虎等)的杀虫活性较低(frankenhuyzen2009,jakka,ferr
é
et al.2015)。cry1d与cry1f与cry1a类蛋白在氨基酸序列上同源性不高,对鳞翅目昆虫的杀虫谱也与cry1a类蛋白有差异。cry1fa蛋白对草地贪夜蛾、棉铃虫、粉纹夜蛾、小菜蛾、甜菜夜蛾等具有高杀虫生物活性,转cry1fa基因玉米在美洲地区被大量应用于防治草地贪夜蛾等鳞翅目害虫(frankenhuyzen 2009)。
4.长期大量地使用单一的杀虫蛋白质,可以导致害虫对bt杀虫蛋白质抗性的产生和发展(carriere,crickmore et al.2015)。自2005年发现第一例bt抗性害虫以来,全球抗性害虫发生量逐年升高,并呈现快速增长的趋势(tabashnik and carri
è
re 2017)。田间检测到部分鳞翅目害虫已经产生了抗性,如bagla等报道了中国新疆、印度等地种植的表达cry1ac蛋白的转基因棉花上发现了抗性棉铃虫和抗性棉红铃虫(bagla 2010);monnerat等报道了巴西种植的表达cry1fa蛋白的转基因玉米产品上发现了抗性草地贪夜蛾(monnerat,martins et al.2015)。
5.对于治理害虫抗性的发生,国际上通行的方式有两种,即基因堆叠策略和高剂量/庇护所策略。基因堆叠策略的主要途径是将两种表达单一抗虫蛋白的转化体进行杂交,使得后代植株能够同时表达两种抗虫蛋白,例如拜耳(原孟山都)公司的power core
tm
玉米是将表达cry1a.105、cry2ab蛋白的mon89034与表达cry1f蛋白的tc1507进行杂交、育种后开发的产品。
6.中国专利cn102843903b中提供了一种利用cry1da和cry1fa蛋白管理草地贪夜蛾
对bt毒素的抗性的方法;中国专利cn102753012b提供了一种将cry1i、cry1e等与cry1fa配合用于控制草地贪夜蛾和欧洲玉米螟的方法。上述cry蛋白与cry1fa蛋白都是bt杀虫晶体蛋白cry1家族的成员,其氨基酸序列相似度较高,杀虫机理比较相似(frankenhuyzen,2017)。
7.vip3家族蛋白与cry1或cry2家族蛋白在进化上没有同源性,在杀虫机理上完全不同(chakroun,2016)。vip3蛋白与杀虫晶体蛋白cry在昆虫中肠上具有不同的结合受体,它们对昆虫的成毒机制不同,不存在交互抗性。chakroun等总结文献研究指出,cry1a、cry1fa等蛋白质在昆虫中肠上典型的几种结合受体均无法与vip3蛋白发生相互作用,杀虫晶体蛋白与vip3蛋白在昆虫中肠的作用机理不同(chakroun,banyuls et al.2016)。wang等通过研究草地贪夜蛾发现,cry1f蛋白在草地贪夜蛾中肠上的结合受体蛋白质为abcc2,而vip3a蛋白的受体则是sr
‑
c(wang et al.,2019)。
8.但是目前广泛使用的杀虫蛋白vip3aa对玉米螟没有杀虫活性。因此,使用vip3aa对玉米螟的抗性管理没有作用。本发明首次披露通过在转基因玉米中同时表达bt晶体杀虫蛋白质和对玉米螟具有杀虫活性的vip3基因,如vip3af,vip3ag和vip3ab,以控制和减缓玉米螟的抗性发生和发展。
(三)
技术实现要素:
9.本发明提供一种管理玉米螟对bt毒素发生抗性的方法,将多种bt营养期杀虫蛋白vip3用于降低抗bt杀虫晶体蛋白cry的玉米螟抗性。本发明将两种具有完全不同作用机理的bt抗虫蛋白在同一植物转化体中高剂量表达,从而达到延缓玉米螟对bt杀虫蛋白抗性发生的目的。本发明构建了包含所述的bt抗虫蛋白编码基因的t
‑
dna载体,并将其导入植物基因组中,获得的转基因植物对普通玉米螟和抗性玉米螟都具有高效杀虫效果,实现延缓玉米螟对bt毒素抗性发生的目标。
10.本发明采用的技术方案是:
11.本发明提供一种管理玉米螟对bt杀虫蛋白抗性的方法,所述的方法通过在同一植物细胞中同时表达至少一种bt杀虫晶体蛋白cry和至少一种bt营养期杀虫蛋白vip3实现。所述bt杀虫晶体蛋白包括cry1和cry2蛋白质家族中的一种或多种,优选包括cry1ab、cry1f、cry2ab;所述bt营养期杀虫蛋白是对玉米螟具有杀虫活性的vip3蛋白,包括vip3af、vip3ag、vip3ab中的任意一种或多种。
12.进一步,所述cry1ab核苷酸序列如seq id no:4所示,氨基酸序列如seq id no:1所示;cry1fa核苷酸序列如seq id no:5所示,氨基酸序列如seq id no:2所示;cry2ab核苷酸序列如seq id no:6所示,氨基酸序列如seq id no:3所示。
13.进一步,vip3af核苷酸序列如seq id no:10所示,氨基酸序列如seq id no:7所示;vip3ag核苷酸序列如seq id no:11所示,氨基酸序列如seq id no:8所示;vip3ab核苷酸序列如seq id no:12所示,氨基酸序列如seq id no:9所示。
14.需要指出的是,bt杀虫晶体蛋白cry和bt营养期杀虫蛋白vip3可以由不同的核苷酸序列编码。特别是在不同植物中,不同密码子的使用频率可以是不同的,可以根据在不同植物中的应用使用不同的核苷酸序列。例如玉米等单子叶植物往往使用gc含量高的密码子,而双子叶植物中gc含量往往低一些。因此,只要是使用了本发明披露的编码了bt杀虫晶
体蛋白或bt营养期杀虫蛋白vip3的氨基酸序列的基因,均应视为本发明的内容。
15.进一步,本发明管理玉米螟对bt杀虫蛋白抗性的方法包括:(1)构建同时含bt杀虫晶体蛋白cry编码基因表达框和bt营养期杀虫蛋白vip3编码基因表达框的转化载体,再将转化载体的t
‑
dna导入到植物细胞中,获得能够表达bt杀虫蛋白的转基因植物细胞,进而延缓玉米螟对bt毒素抗性发生;(2)分别构建含bt杀虫晶体蛋白cry编码基因表达框的转化载体和含bt营养期杀虫蛋白vip3编码基因表达框的转化载体,并分别将各个转化载体的t
‑
dna导入到植物细胞中,分别获得表达bt杀虫晶体蛋白cry的转基因植物细胞和表达bt营养期杀虫蛋白vip3的转基因植物细胞,然后将两种转基因植物细胞通过植物杂交产生同时表达bt杀虫晶体蛋白cry和bt营养期杀虫蛋白vip3的转基因植物细胞,进而延缓玉米螟对bt毒素抗性发生。
16.进一步,bt杀虫晶体蛋白cry编码基因的表达框由启动子、编码基因和终止子构成。所述的启动子包括但不限于玉米泛素启动子pzmubi、花椰菜花叶病毒35s启动子pcamv 35s,优选启动子为pzmubi(核苷酸序列如seq id no:13所示);所述的终止子包括但不限于农杆菌烟脂碱合成酶终止子tnos,花椰菜花叶病毒35s rna的终止子tcamv 35s,优选终止子为tnos(核苷酸序列如seq id no:14所示)。
17.进一步,bt营养期杀虫蛋白vip3编码基因的表达框由启动子、编码基因和终止子构成。所述的启动子包括但不限于玉米泛素启动子pzmubi、花椰菜花叶病毒35s启动子pcamv 35s,优选启动子为pzmubi(核苷酸序列如seq id no:13所示);所述的终止子包括但不限于农杆菌烟脂碱合成酶终止子tnos,花椰菜花叶病毒35s rna的终止子tcamv 35s,优选终止子为tnos(核苷酸序列如seq id no:14所示)。
18.进一步,所述的bt杀虫晶体蛋白cry编码基因表达框和bt营养期杀虫蛋白vip3编码基因表达框的相对顺序以及表达框在转化载体中的插入位置没有限制,只要包含有所述的bt杀虫晶体蛋白cry编码基因表达框和所述的bt营养期杀虫蛋白vip3编码基因表达框的转化载体均应视为本发明的内容。所述转化载体的基础载体为常规的植物遗传转化载体,包括但不限于pcambia1300、pcambia1301、pcambia2301等。
19.本发明还提供一种用于玉米螟对bt杀虫蛋白抗性管理的转化载体,所述转化载体含bt杀虫晶体蛋白cry编码基因表达框和bt营养期杀虫蛋白vip3编码基因表达框,优选所述转化载体包括同时含bt杀虫晶体蛋白cry编码基因表达框和bt营养期杀虫蛋白vip3编码基因表达框的转化载体,或者分别包含bt杀虫晶体蛋白cry编码基因表达框的转化载体和bt营养期杀虫蛋白vip3编码基因表达框的转化载体;所述bt杀虫晶体蛋白包括cry1和cry2蛋白质家族中的一种或多种,优选包括cry1ab、cry1f、cry2ab;所述bt营养期杀虫蛋白是对玉米螟具有杀虫活性的vip3蛋白,包括vip3af、vip3ag、vip3ab中的任意一种或多种。
20.本发明还提一种用于玉米螟对bt杀虫蛋白抗性管理的抗虫植物细胞,所述的植物细胞同时表达至少一种所述的bt杀虫晶体蛋白cry和至少一种所述的bt营养期杀虫蛋白vip3。只要将能够同时表达本发明所述的bt杀虫晶体蛋白cry和bt营养期杀虫蛋白vip3的植物细胞应用于制备转基因农作物,均应视为本发明的内容。
21.进一步,本发明所述的抗虫植物细胞可以通过植物遗传转化方法获得。将所述的同时包含一种所述cry基因和vip3基因的转化载体的t
‑
dna通过遗传转化方法整合到植物基因组中,或者将含所述bt杀虫晶体蛋白cry基因转化载体的植物细胞和bt营养期杀虫蛋
no:6所示;vip3af蛋白的氨基酸序列如seq id no:7所示,编码基因vip3af的核苷酸序列如seq id no:10所示;vip3ag蛋白的氨基酸序列如seq id no:8所示,编码基因vip3ag的核苷酸序列如seq id no:11所示;vip3ab蛋白的氨基酸序列如seq id no:9所示,编码基因vip3ab的核苷酸序列如seq id no:12所示。
32.利用大肠杆菌(escherichia coli)pet表达系统进行重组蛋白质的表达和制备。将目的基因导入pet表达载体中,再将表达载体转入合适的表达菌株,在一定条件下诱导目的基因的表达,制备重组蛋白质。本实施例中重组cry蛋白和重组vip3蛋白的制备过程完全一致,以下以cry1ab蛋白为例进行说明。
33.cry1ab蛋白的制备过程如下:将cry1ab基因克隆到表达载体pet28a(novagen,69015
‑
3,美国)的bamhi和saci这二个限制性内切酶酶切位点之间,获得重组质粒pet28a
‑
cry1ab;重组质粒通过热击转化法导入大肠杆菌bl21(de3)细胞系,在含50mg/l卡那霉素的lb固体培养基上37℃过夜培养,筛选得到单克隆阳性菌落;单克隆阳性菌落接种至含50mg/l卡那霉素的lb液体培养基,37℃过夜培养后,取0.2ml菌液继代接种至装有200毫升含50mg/l卡那霉素的lb液体培养液的锥形瓶中,37℃振荡培养至od
600
=0.6,然后加入iptg(isopropylβ
‑
d
‑1‑
thiogalactopyranoside)至终浓度为0.5mm,并继续在同样的条件下振荡培养4小时,诱导重组蛋白质的表达;表达完成后,将培养液5000rpm离心15分钟,弃掉上清培养液,收集细胞沉淀,并以50毫升20mm磷酸缓冲液(pbs,ph=7.4)重悬细胞;随后将重悬液置于冰水中充分冷却后,用超声波破碎仪(宁波新芝生物科技公司,型号jy92
‑
iin)粉碎细胞(探头直径10mm,以650w、50%机显功率破碎15min,工作3s间歇6s),12000rpm离心10min后,分别收集上清和沉淀,并采用聚丙烯酰胺凝胶电泳(sds
‑
page)验证目标蛋白的条带大小,将含有正确目的条带的蛋白质样品储存于
‑
80℃冰箱中备用。
34.cry重组蛋白存在于超声破碎后的沉淀中,将含cry1ab蛋白的沉淀以carbonate buffer(50mm na2co3,ph=10)重悬2h后即可溶解,获得重组蛋白cry1ab溶液,采用莫纳生物科技公司eva3100超微量核酸蛋白检测仪(货号gd30101)测定溶液中a280的吸光度,即可得到重组蛋白质中目的蛋白的浓度。用同样的carbonate buffer缓冲液调整蛋白浓度均为1.5mg/ml,得到重组cry1ab蛋白溶液,同样条件下制备蛋白浓度为1.5mg/ml的重组蛋白cry1fa溶液,蛋白浓度为1.5mg/ml的重组蛋白cry2ab溶液。
35.vip3a重组蛋白存在于超声破碎后的上清中,将携带有vip3af、vip3ag或vip3ab蛋白的上清进行his
‑
tag纯化(qiagen,ni
‑
nta superflow,lot no.160049084),纯化步骤参照该试剂的说明书进行,具体如下:(1)向柱中填入1ml ni
‑
nta树脂,静置至分层;(2)流出保存液,用8~10倍柱体积的lysis buffer清洗树脂至完全平衡;(3)将样品加入到柱中,封口,轻轻摇晃(或将柱平躺),在室温(或4℃,视蛋白情况而定)下静置30min;(4)将柱子放置在铁架台上,以1ml/min的流速释放上清液;(5)用5倍柱体积wash buffer清洗,流速1ml/min,洗掉杂蛋白;(6)用5倍柱体积elution buffer洗脱,流速1ml/min,保存洗脱液。最后采用莫纳eva3100检测仪测定洗脱液中a280的吸光度,即可得到重组蛋白质中目的蛋白的浓度,用同样的carbonate buffer缓冲液调整蛋白浓度为2.5mg/ml,获得vip3a重组蛋白溶液。同样条件下,制备蛋白浓度为2.5mg/ml含重组蛋白vip3ag溶液,蛋白浓度为2.5mg/ml的含重组蛋白vip3ab蛋白溶液。
36.所述蛋白纯化过程中lysis buffer的配方为:20mm nah2po4、300mm nacl、10mm咪
唑,用naoh调整ph至8.0;wash buffer的配方为:20mm nah2po4、300mm nacl、20mm咪唑,用naoh调整ph至8.0;elution buffer的配方为20mm nah2po4、300mm nacl、250mm咪唑,用naoh调整ph至8.0。
37.所述lb液体培养基组成为:蛋白胨10g/l、酵母提取物5g/l、氯化钠10g/l,溶剂为双蒸水,120℃高温高压灭菌15分钟;lb固体培养基是在lb液体培养基中再加入15g/l琼脂,高温高压灭菌后倒置于已灭菌的培养皿中冷却即可。
38.通过相同的方法分别制备得到cry1fa、cry2ab、vip3af、vip3ag和vip3ab蛋白,储存在
‑
80℃冰箱中备用。此外,用相同方法制备vip3aa20蛋白(ncbi accession no.abg20429)作为实验对照,该蛋白是商业化转基因产品mir162(usda petition for determination of nonregulated status for insect
‑
resistant mir162 maize,07
‑
253
‑
01p)中表达的bt杀虫蛋白质。
39.实施例2、重组蛋白质对玉米螟的生物活性测定
40.实验对象:选取新生2h以内的普通玉米螟(采集自海南三亚试验基地田间)、抗cry1ab玉米螟和抗cry1fa玉米螟的初孵幼虫作为被试昆虫。抗性玉米螟(抗cry1ab玉米螟和抗cry1fa玉米螟)通过人工饲养筛选获得,具体方法参考文献资料(kain,wendy c.,et al."inheritance of resistance to bacillus thuringiensis cry1ac toxin in a greenhouse
‑
derived strain of cabbage looper(lepidoptera:noctuidae)."journal of economic entomology 97.6(2004):2073
‑
2078.;huang,fangneng,et al."baseline susceptibility and changes in susceptibility to bacillus thuringiensis subsp.kurstaki under selection pressure in european corn borer(lepidoptera:pyralidae)."journal of economic entomology 90.5(1997):1137
‑
1143.)提供的方法进行筛选。抗性玉米螟对bt杀虫晶体蛋白(cry1ab或cry1fa)不敏感,普通玉米螟对bt杀虫晶体蛋白质敏感。
41.实验组:实施例1方法制备的重组蛋白质(重组蛋白cry1ab溶液,重组蛋白cry1fa溶液,重组蛋白cry2ab溶液,重组蛋白vip3af溶液,重组蛋白vip3ag溶液,重组蛋白vip3ab蛋白溶液)进行生物活性的预实验(分别设置1ng/cm2、10ng/cm2、100ng/cm2、1000ng/cm2、10000ng/cm2五组浓度);根据预实验结果,将被试昆虫全部死亡的最低浓度设为上限浓度,设置8组浓度梯度(浓度范围如下表1所示,以ph=7.4的pbs缓冲液稀释),测定其对昆虫具有50%致死率的浓度区间。每组浓度测试虫数为80头。
42.阴性对照组:阴性对照的准备方法与实施例1相同,唯一的区别是大肠杆菌表达菌株中转入的质粒为不含任何插入dna序列的pet28a空载体。
43.测定方法:以表面平铺法测定重组蛋白质对昆虫的致死中浓度lc
50
。
44.昆虫人工饲料煮熟后平铺到24孔培养板(每孔直径为1.5cm)中,等待饲料凝固。每块24孔板中随机设置20个孔为实验组,4个为阴性对照组。实验组每孔的饲料表面铺上重组蛋白200微升,阴性对照组每孔的饲料表面铺上阴性对照蛋白200微升,待整块板铺完后将其放置在120rpm的摇床上,在室温(25
‑
30℃)下晃动至液体晾干。每孔接入2头被试昆虫,并用透气封口膜封闭,在28℃培养箱中遮光饲养7天,统计昆虫死亡率,得出对应的致死中浓度lc
50
,结果如下表2所示。
45.表1对玉米螟生物活性测定的重组蛋白浓度梯度表(ng/cm2)
46.重组蛋白质浓度1浓度2浓度3浓度4浓度5浓度6浓度7浓度8cry1ab10203040506075100cry1fa10203040506075100cry2ab50100250500750100015002000vip3af10255075100125150200vip3ag10255075100125150200vip3ab10255075100125150200
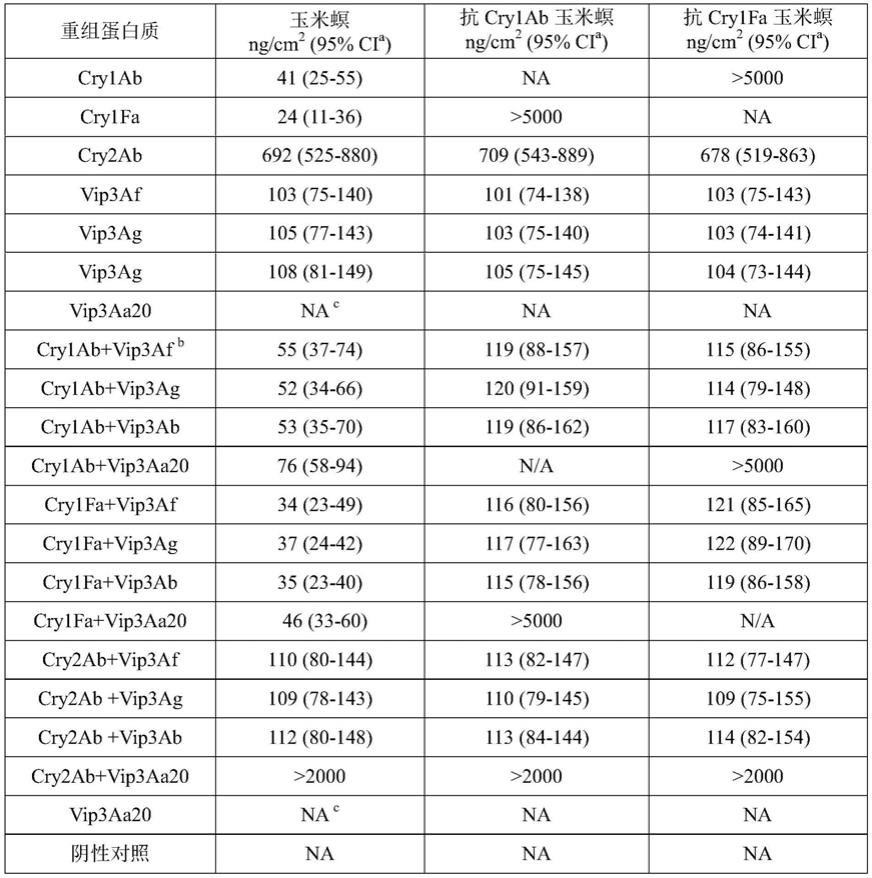
47.表2重组蛋白质对玉米螟和抗性玉米螟的致死中浓度lc
50
[0048][0049]
注:a、95%ci,95%置信区间。b、cry1ab vip3af,cry1ab、vip3af蛋白以摩尔比1:1的混合物,以下类同。c、na,not active。
[0050]
从上述对玉米螟的生物活性测定结果可以得出结论:(1)vip3af、vip3ag和vip3ab蛋白对玉米螟具有极高的杀虫活性,而vip3aa20蛋白对玉米螟没有活性;(2)vip3af、vip3ag、vip3ab蛋白对抗性玉米螟和普通玉米螟的lc
50
没有显著差异,都具有极高活性,表明vip3af、vip3ag、vip3ab蛋白与cry1ab或cry2ab蛋白之间均不存在对玉米螟的交互抗性;(3)将cry蛋白与vip3蛋白按摩尔比1:1比例进行物理混合后,混合蛋白对普通玉米螟和抗
性玉米螟都具有高杀虫活性。
[0051]
实施例3、转化载体的构建
[0052]
在本实施例中,转化载体是基于pcambia1300(ncbi序列编号af234296)载体而构建的,本发明的读者可以根据本实施例提供的方法,在其他植物遗传转化载体上实现相同的效果,例如pcambia1301、pcambia2301等。本实施例以构建含有cry1fa和vip3af两个基因的表达框为例阐述转化载体的构建方法,将cry1fa基因替换为任意本发明中所述的cry蛋白、将vip3af基因替换为任意本发明中所述的vip3蛋白,均能实现转化载体的构建。此外,抗虫基因的表达框中使用的启动子可以是任意组成型启动子,如花椰菜花叶病毒35s启动子pcamv35s、玉米泛素启动子pzmubi等;终止子的选择没有限制,例如常用的农杆菌nos终止子tnos、花椰菜花叶病毒35s终止子t35s等等都可以作为抗虫基因的终止子。
[0053]
本发明涉及的编码基因均由上海生工公司进行人工合成。编码基因cry1ab的核苷酸序列如seq id no:4所示,编码基因cry1fa的核苷酸序列如seq id no:5所示,编码基因cry2ab的核苷酸序列如seq id no:6所示;编码基因vip3af的核苷酸序列如seq id no:10所示,编码基因vip3ag的核苷酸序列如seq id no:11所示,编码基因vip3ab的核苷酸序列如seq id no:12所示。
[0054]
启动子均选用玉米泛素启动子pzmubi(核苷酸序列如seq id no:13所示),终止子均为农杆菌烟脂碱合成酶终止子tnos(核苷酸序列如seq id no:14所示)。将pcambia1300载体用限制性内切酶hindiii和kpni进行双酶切,经琼脂糖凝胶电泳、割胶回收后,已被线性化的载体片段储存在
‑
20℃冰箱中备用。
[0055]
设计合成表3所示pcr引物,扩增获得用于快速克隆的元件片段:
[0056]
启动子pzmubi(genebank no:s94464)从玉米的基因组中通过pcr获得,使用的引物分别是:zmubi
‑
f(5
’‑
gcatgcctacagtgcagcgtgacccggtcgtgc
‑3’
);zmubi
‑
r(5
’‑
tctagagtcgacctgcagaagtaacaccaaacaacag)。选用快速无缝克隆试剂盒(诺唯赞公司产品clonexpress multis one step cloning kit,货号c113
‑
01),实验步骤参照说明书,获得玉米泛素启动子pzmubi,核苷酸序列如seq id no.13所示。
[0057]
农杆菌烟脂碱合成酶终止子tnos由上海生工公司人工合成,核苷酸序列如seq id no.14所示。
[0058]
表3各片段根据无缝克隆(clonexpress)方法设计的引物序列
[0059]
[0060]
pcr反应选用高保真酶2
×
phanta max master mix(诺唯赞,货号p515
‑
01),反应体系按说明书进行,按如下表4进行反应:
[0061]
表4
[0062][0063]
pcr产物经琼脂糖凝胶电泳、割胶回收后,测定回收产物的浓度。
[0064]
快速无缝克隆反应参考试剂盒(诺唯赞公司货号c113
‑
01)说明书,按如下表5体系配制:
[0065]
表5无缝克隆反应液
[0066][0067]
将表5无缝克隆反应液混匀后,37℃孵育30min,结束后快速放置到冰上冷却2min以上。随后进行大肠杆菌dh5α感受态细胞(上海唯地生物公司,cat#:dl1001)的转化:取出感受态细胞后冰上融化,加入上述反应完成后的无缝克隆反应液,冰浴25min,随后42℃水浴热激45s,再在冰上快速冷却2min,加入1ml lb培养液,37℃下200rpm振荡培养1h,随后在含有50mg/l卡那霉素的lb固体培养基上涂板,37℃过夜培养后挑单个克隆进行测序鉴定。最终构建完成的载体如图1所示。获得转化载体pcambia1300
‑
pzmubi
‑
cry
‑
tnos
‑
pzmubi
‑
vip3
‑
tnos,其中cry为cry1ab、cry1fa、cry2ab;vip3为vip3af、vip3ag、vip3ab。
[0068]
实施例4、制备用于玉米遗传转化的根癌农杆菌
[0069]
由实施例3制备得到的包含转化载体的大肠杆菌经测序鉴定正确后保存在
‑
80℃冰箱。将保存的大肠杆菌菌株在含有50mg/l卡那霉素的lb固体培养基上划线,37℃过夜培养后挑取单克隆至含有50mg/l卡那霉素的3毫升lb液体培养基中,37℃过夜培养后提取质粒。将上述质粒通过电击转化法转入农杆菌eha105的感受态细胞中,在同时含有50mg/l卡那霉素、25mg/l利福平和100mg/l链霉素的yep固体培养基上均匀涂板,置于28℃下培养48~72h,待长出单克隆后挑取单菌落至同时含有50mg/l卡那霉素、25mg/l利福平和100mg/l链霉素的yep液体培养基中,28℃培养48~72h后,获得农杆菌培养液,提取质粒进行测序鉴定,鉴定正确的阳性克隆菌保存于
‑
80℃冰箱中备用。
[0070]
lb培养基组成:蛋白胨10g/l、酵母提取物5g/l、nacl 10g/l,溶剂为水,高温高压灭菌;若配制固体培养基,则加入15g/l琼脂。
[0071]
yep培养基组成:蛋白胨10g/l、酵母提取物10g/l、nacl 5g/l,溶剂为水,高温高压灭菌;若配制固体培养基,则加入15g/l琼脂。
[0072]
实施例5、转基因玉米的获得
[0073]
转基因玉米的获得方法是根据现有技术,采用根癌农杆菌介导法转化玉米幼胚(ishida,saito et al.1996),具体步骤如下:取授粉后8~10天的玉米穗,收集所有的未成熟胚(大小为1.0~1.5mm)。取实施例4中阳性克隆菌的农杆菌细胞与未成熟胚共培育2~3天(22℃)。随后,转移未成熟胚到愈伤诱导培养基(含200mg/l特美汀)上,28℃暗培养10~14天。接下来将所有的愈伤组织转移到带有50ng/ml潮霉素的筛选培养基上进行筛选,28℃暗培养2~3周。转移所有的组织到新鲜潮霉素的筛选培养基上进行第二次筛选,28℃暗培养2~3周。随后转移所有筛选后仍然成活的胚性组织到再生培养基上,28℃暗培养10~14天,每皿一个株系。转移胚性组织到新鲜的再生培养基上,26℃光照培养10~14天。转移所有发育完全的植株到生根培养基上,26℃光照培养直到根系发育完全。将cry为cry1fa,vip3为vip3af的转基因玉米t0代转化体记为fz。
[0074]
实施例6、转基因农作物转化体中抗虫蛋白表达量的测定
[0075]
由于转化体表现型与非转基因玉米差异较大,从实施例5获得的t0代转化体fz中,取植株表现型与非转基因无明显差异的10个株系,使用新鲜组织测定每个转化体中抗虫蛋白的含量,具体为:取每个株系在v6叶期的新鲜叶片组织50mg,每个叶片组织取3份样品作为重复。对于新鲜组织,经液氮研磨后加入500微升pbs缓冲液,充分混合后12000rpm离心5min,将上清液用pbs缓冲液稀释500倍后进行酶联免疫法(elisa)测定。使用的试剂盒为上海佑隆生物公司的cry1f elisa定量试剂盒(产品编号aa0641),参照产品使用说明书进行操作;测定vip3a蛋白的试剂盒为vip3a elisa定量试剂盒(产品编号aa1641),实验操作方法与上述cry1f elisa试剂盒一致。测得的t0代转基因玉米转化体中cry1fa蛋白和vip3af蛋白的结果表6所示。实验结果表明,t0代转基因玉米转化体中cry1fa蛋白表达量在2.9
‑
15.5μg/g
‑
fwt之间,vip3af表达量在2.9
‑
13.6μg/g
‑
fwt之间,转化体个体之间不同蛋白的表达量有所差异。
[0076]
同样方法检测,实施例5获得的cry为cry1ab、cry2ab;vip3为vip3ag、vip3ab的转基因玉米t0代转化体中cry蛋白含量在3.3~10.6μg/g
‑
fwt之间,vip3蛋白含量在2.9~12.8μg/g
‑
fwt之间。
[0077]
表6 t0代玉米转化体中抗虫蛋白表达量的测定
[0078][0079]
注:*μg/g
‑
fwt:微克每克新鲜组织
[0080]
实施例7、转基因抗虫玉米转化体的抗虫性鉴定
[0081]
通过实施例5、6所述方法获得的转基因植物转化体,移栽至温室,待植物能够稳定生长(玉米进入v6
‑
8叶期)后进行生物活性测定。选取实施例6中测定了抗虫蛋白表达量的
转化体进行生测,选用玉米螟和抗性玉米螟(抗cry1ab玉米螟和抗cry1fa玉米螟)进行转基因玉米转化体的生物活性测定。取5
‑
6cm新鲜的植物叶片组织,放置在直径7.5cm的培养皿中,垫上一片湿润的滤纸,接入10头初孵2h内的幼虫,盖上盖子后用封口膜封闭培养皿,防止幼虫逃逸。每个转化体设置5个重复的处理组,每次生物测定实验以单独表达vip3aa20蛋白的转基因玉米转化体az
‑
54(vip3aa20蛋白表达量为12.5μg/g
‑
fwt)作为实验对照,以相同生长期的非转基因植物作为阴性对照。5天后,统计每个实验组中幼虫的存活头数,计算出幼虫的死亡率。根据统计得到的死亡率数据。表达cry1fa和vip3af的转基因玉米抗虫性,结果见表7所示。
[0082]
生测结果表明,相比于非转基因玉米和表达单独vip3aa20蛋白(表达量为12.5μg/g
‑
fwt)的转基因玉米az
‑
54,同时表达cry蛋白和vip3a蛋白的玉米转化体fz能在72h内杀灭玉米螟和抗性玉米螟幼虫,显示出对普通玉米螟和抗性玉米螟均具有极高的杀虫活性,这一结论与实施例2中bt蛋白的生测数据一致。此外,从本实施例的生测数据中可以看到,bt蛋白的表达量与杀虫效率存在一定的相关性。例如,转化体fz72中cry1fa和vip3af蛋白的表达量相对较低,分别为2.9、7.5μg/g
‑
fwt,表现在生测试验中即为24h玉米螟的死亡率为80%,该死亡率数据在所有转化体中是最低的。总的来说,表达cry1fa和vip3af的转化体fz能够在72h内高效杀灭玉米螟、抗cry1ab玉米螟和抗cry1fa玉米螟,而vip3aa20蛋白的转化体az
‑
54对玉米螟没有杀虫效果,因此本实施例所述的fz转化体可以为玉米螟对bt蛋白的抗性管理提供了一种新的研究思路和产品解决方案。
[0083]
表7玉米螟和抗性玉米螟饲喂转基因玉米转化体后的死亡率统计
[0084][0085]
注:
*
ck:对照组非转基因玉米。
[0086]
同样条件下,测试实施例3中其他转化载体转化植物获得的转基因玉米(如同时表达cry1ab vip3ag蛋白的转化体、同时表达cry1fa vip3ab蛋白的转化体),结果显示所有筛选得到的转化体都能够在72h内杀灭普通玉米螟和抗性玉米螟。这一结果与实施例2中原核表达的蛋白质对玉米螟生物活性测定的结论一致。因此,在同一转基因玉米中同时表达所述cry蛋白与所述vip3蛋白,在防治玉米螟的同时,可以有效降低抗性玉米螟的发生,是一种在玉米上全新的管理玉米螟对bt毒素抗性的有效手段。
[0087]
实施例8、通过植物杂交方法获得的转基因玉米及其对玉米螟的生物活性
[0088]
通过本发明所述的转化载体构建方法(实施例3)和植物遗传转化方法(实施例4、5),可以获得表达一种或多种cry蛋白的转基因玉米,也可以获得表达一种或多种vip3蛋白
的转基因玉米。在实施例3中描述了转化载体的构建方法,可以制备得到含有一个或多个cry基因的植物转化载体,也可以制备得到含有一个或多个vip3基因的植物转化载体。在实施例4中详细描述了通过农杆菌制备转基因玉米的具体步骤。通过上述实施例,将只有cry基因的转化载体导入植物细胞,可以获得只表达cry蛋白的转基因玉米;或将只有vip3基因的转化载体导入植物细胞,可以获得只表达vip3蛋白的转基因玉米。
[0089]
例如,通过本发明实施例3、4、5中提供的方法,获得了一种表达cry1fa蛋白的转基因玉米转化体tf43和一种表达vip3af蛋白的转基因玉米转化体tm19。通过elisa方法测定tf43中cry1fa蛋白的表达量达到10.4μg/g
‑
fwt,tm19中vip3af蛋白的表达量达到20.5μg/g
‑
fwt。将tf43和tm19植株同时种植在温室中,在生育期通过常规的有性生殖方式进行植物杂交,在玉米开花期将tf43的花粉人工授粉至tm19的雌穗上,获得了同时表达cry1fa蛋白和vip3af蛋白的杂交玉米后代tmf4319;或是将tm19的花粉人工授粉至tf43的雌穗上,获得了同时表达cry1fa蛋白和vip3af蛋白的杂交玉米后代tmf1943。将tmf1943和tmf4319种子种植于试验田中,用转基因快速检测试纸(上海佑隆生物公司,货号aa0831
–
cp4)对每棵植株进行检测,只留下检测阳性的植株,剔除阴性植株;再对阳性植株进行外源蛋白表达量的测定,使用实施例6中所述的elisa方法,测定tmf1943中cry1fa蛋白的表达量平均为10.1μg/g
‑
fwt,vip3af蛋白的表达量平均为19.7μg/g
‑
fwt;tmf4319中cry1fa蛋白的表达量平均为10.3μg/g
‑
fwt,vip3af蛋白的表达量平均为19.6μg/g
‑
fwt。
[0090]
对tmf4319和tmf1943进行玉米螟的生物活性测定(测定方法如实施例7中所述的方法一致),结果显示,玉米螟、抗cry1ab玉米螟和抗cry1fa玉米螟均在24h内全部死亡,表明tmf4319和tmf1943对普通玉米螟和抗性玉米螟都具有极高的杀虫活性。因此,通过植物杂交方法在同一植株中同时表达所述的cry蛋白和所述的vip3蛋白,也可以实现玉米螟抗性管理的目的,可以有效减缓抗性害虫的发生。
[0091]
最后,以上列举的仅是本发明的若干实施例。显然,本发明不限于以上实施例,还可以有许多延伸和拓展。本领域的普通技术人员能从本发明公开的内容直接到导出或联想到的所有延伸,均应认为是本发明的保护范围。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。