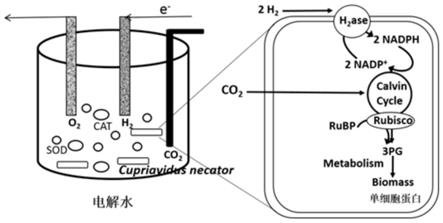
1.本公开属于微生物和基因工程技术领域,具体来说,本公开涉及一种包含重组氢氧化细菌的碳固定系统,以及合成目标产物的方法。
背景技术:
2.单细胞蛋白(single cell protein,scp)又称微生物蛋白、菌体蛋白,是利用各种基质大规模培养细菌、酵母菌、霉菌、微型藻等而获得的微生物蛋白。单细胞蛋白的化学组成中一般以蛋白质、脂肪为主,含粗蛋白50%~85%,并且氨基酸组分齐全,还含维生素、无机盐、脂肪和糖类等,其营养价值优于鱼粉和大豆粉。单细胞蛋白不仅能制成“人造肉”,供人们直接食用,还常作为食品添加剂,用以补充蛋白质或维生素、矿物质等。目前,已发现藻类、细菌和高等真菌均可生产scp产品。其中,通过微生物大量表达合成蛋白质,或者将整个菌体作为单细胞蛋白,有望突破传统的农业和养殖业限制,实现一种高效、快速、节能、减废的绿色饲料蛋白生产方式。
3.氢氧化细菌(hydrogen
‑
oxidizing bacteria)是能够利用h2作为电子供体、o2作为电子受体并同化co2的一类兼性化能自养型细菌。近年来随着可再生能源技术和生物技术的发展,以及全球气候变化、国际形势动荡和自然病灾突发等带来的挑战,以二氧化碳为碳源,以可再生能源产生的氢气为能量,培养氢氧化细菌生产单细胞蛋白的技术逐渐受到关注,具有重要的应用潜力。
4.专利文献1公开了一种使用化能自养微生物来从工业废物俘获碳的系统和方法。系统包括工业源;以及包括微生物的生物反应器。生物反应器被供给有来自源的废物流,该废物流向微生物提供碳;并且也被供给有氢原料。微生物从氢原料获取能量,通过化能自养将提供的碳转换为化学产物。该方法虽然能够将工业废物中的碳氧化物转化为诸如羟基链烷酸酯类、聚羟基链烷酸酯类的有机化合物,然而,其化能自养微生物存在对氢原料的利用率低,转化有机化合物的效率需提高的问题。并且,该微生物反应器未用于生产单细胞蛋白,无法实现对单细胞蛋白的高效生产。
5.专利文献2公开了将如可再生h2和废弃co2发生炉煤气或合成气等气态底物转化为高蛋白生物质的微生物和生物过程,所述高蛋白生物质可以直接用于人类营养,或用作植物、真菌或其它微生物的营养素或用作土壤碳、氮和其它矿物质营养素的来源。然而,该方法同样存在对h2、co2的利用率不高的问题,限制了转化生产生物质、氨基酸、蛋白质等的效率。并且,该方法需要先电解水收集h2,再进行h2、co2和o2的气体发酵生产,h2的收集过程会导致生产成本的增加,并且存在潜在的安全风险。
6.引用文献:
7.专利文献1,cn103958687a
8.专利文献2,cn110678539a
技术实现要素:
9.发明要解决的问题
10.鉴于现有技术存在的问题,例如,微生物以h2为能源固定co2的过程存在对h2的利用率低,固定co2的效率低,导致生产单细胞蛋白的速率和产量低;以及h2的收集过程存在成本高、并具有一定安全隐患的问题。为此,本公开提供了一种碳固定系统,通过电解水产生氢气,原位为重组氢氧化细菌提供能源,无需氢气的收集再利用,避免了收集氢气导致的成本提高和安全性降低的问题;另一方面,碳固定系统使用氢元件和固碳元件选择性加强的重组氢氧化细菌,重组氢氧化细菌对氢气的利用效率及固碳反应的效率提高,能够实现对目标产物的高效生产,得到高产量的目标产物(例如,单细胞蛋白)。
11.用于解决问题的方案
12.第一方面,本公开提供了一种碳固定系统,其中,所述碳固定系统:
13.反应容器,具有容置反应溶液的反应内腔,所述反应溶液中包含重组氢氧化细菌:
14.电解单元,所述电解单元部分伸入所述反应溶液中,用于电解所述反应溶液产生能源气体;
15.通气单元,所述通气单元用于向所述反应溶液内通入碳源气体;
16.其中,所述重组氢氧化细菌包括通过氢氧化反应产生能量的氢元件,和通过固碳反应合成目标产物的固碳元件;并且,与野生型氢氧化细菌相比,所述重组微生物具有如下(i)
‑
(ii)中至少一项所示的特性:
17.(i)氢元件中一个或多个与氢氧化反应相关的蛋白活性,或蛋白编码基因的表达水平被增强;
18.(ii)固碳元件中一个或多个与固碳反应相关的蛋白活性,或蛋白编码基因的表达水平被增强。
19.在一些实施方式中,根据本公开所述的碳固定系统,其中,所述能源气体包含氢气,所述碳源气体包含二氧化碳;优选地,每分钟向所述反应溶液内通入碳源气体的体积与所述反应内腔的体积比为(2
‑
5):10。
20.在一些实施方式中,根据本公开所述的碳固定系统,其中,所述反应溶液中包含超氧化物歧化酶、过氧化氢酶的至少一种;
21.可选地,所述超氧化物歧化酶来源于热栖热菌、嗜热毛壳菌、嗜热子囊菌、大肠杆菌、枯草芽孢杆菌、酿酒酵母的至少一种;
22.可选地,所述过氧化氢酶来源于大肠杆菌、流产布鲁氏菌、黑曲霉菌、枯草芽孢杆菌、布氏栖热菌、粗糙脉孢菌、粘质沙雷氏菌的至少一种;
23.可选地,所述反应溶液中,所述超氧化物歧化酶的含量不小于7u/ml,优选地,所述超氧化物歧化酶的含量不小于30u/ml,更优选地,所述超氧化物歧化酶的含量不小于60u/ml;
24.可选地,所述反应溶液中,所述超氧化物歧化酶的含量不小于20u/ml,优选地,所述超氧化物歧化酶的含量不小于80u/ml,更优选地,所述超氧化物歧化酶的含量不小于160u/ml.。
25.在一些实施方式中,根据本公开所述的碳固定系统,其中,所述反应溶液是包含磷酸盐的发酵培养基,所述磷酸盐的浓度为25
‑
35mm;
26.可选地,所述磷酸盐选自磷酸二氢钾、磷酸氢二钾、磷酸二氢钠、磷酸氢二钠中的一种或两种以上的组合;
27.优选地,所述发酵培养基中还包含亚铁盐;更优选地,所述亚铁盐为硫酸亚铁,所述硫酸亚铁的浓度为30
‑
60mg/l。
28.在一些实施方式中,根据本公开所述的碳固定系统,其中,所述电解单元包括阳极电极和阴极电极;可选地,所述阳极电极为钛片,所述阴极电极为镍片;
29.优选地,所述重组氢氧化细菌靠近所述阴极电极伸入所述反应溶液的一端。
30.在一些实施方式中,根据本公开所述的碳固定系统,其中,所述与固碳反应相关的蛋白包括核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶;优选地,所述与固碳反应相关的蛋白还包括核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的分子伴侣;更优选地,所述分子伴侣为groes和groel。
31.在一些实施方式中,根据本公开所述的碳固定系统,其中,所述核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶包括如(a1)
‑
(a2)任一项所示的第一亚基,如(a3)
‑
(a4)任一项所示的伴侣蛋白,以及如(a5)
‑
(a6)任一项所示的第二亚基:
32.(a1)如seq id no:1所示氨基酸序列的多肽,或者如seq id no:1所示氨基酸序列经过取代、重复、缺失或添加一个或多个氨基酸,且具有seq id no:1所示序列的多肽活性的多肽;
33.(a2)由与seq id no:6所示的核苷酸序列具有至少90%的序列同一性的序列编码的,具有seq id no:1所示序列的多肽活性的多肽;
34.(a3)如seq id no:2所示氨基酸序列的多肽,或者如seq id no:2所示氨基酸序列经过取代、重复、缺失或添加一个或多个氨基酸,且具有seq id no:2所示序列的多肽活性的多肽;
35.(a4)由与seq id no:7所示的核苷酸序列具有至少90%的序列同一性的序列编码的,具有seq id no:2所示序列的多肽活性的多肽;
36.(a5)如seq id no:3所示氨基酸序列的多肽,或者如seq id no:3所示氨基酸序列经过取代、重复、缺失或添加一个或多个氨基酸,且具有seq id no:3所示序列的多肽活性的多肽;
37.(a6)由与seq id no:8所示的核苷酸序列具有至少90%的序列同一性的序列编码的,具有seq id no:3所示序列的多肽活性的多肽;
38.可选地,所述重组氢氧化细菌包含如下(b1)
‑
(b3)任一项所示的groes:
39.(b1)包含如seq id no:4所示氨基酸序列的,并且具有groes蛋白活性的多肽;
40.(b2)如seq id no:4所示氨基酸序列经过取代、重复、缺失或添加一个或多个氨基酸,且具有groes蛋白活性的多肽;
41.(b3)由与seq id no:9所示的核苷酸序列具有至少90%的序列同一性的序列编码的,具有groes蛋白活性的多肽;
42.可选地,所述重组氢氧化细菌包含如下(c1)
‑
(c3)任一项所示的groel:
43.(c1)包含如seq id no:5所示氨基酸序列的,并且具有groel蛋白活性的多肽;
44.(c2)如seq id no:5所示氨基酸序列经过取代、重复、缺失或添加一个或多个氨基酸,且具有groel蛋白活性的多肽;
45.(c3)由与seq id no:10所示的核苷酸序列具有至少90%的序列同一性的序列编
码的,具有groel蛋白活性的多肽。
46.在一些实施方式中,根据本公开所述的碳固定系统,其中,所述重组氢氧化细菌包含表达核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的重组表达载体;优选地,所述重组氢氧化细菌包含表达核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶、groes和groel的重组表达载体;
47.优选地,所述重组表达载体的核苷酸序列如seq id no:11或seq id no:12所示。
48.在一些实施方式中,根据本公开所述的碳固定系统,其中,所述与氢氧化反应相关的蛋白包括氢化酶;优选地,所述重组氢氧化细菌中氢化酶的编码基因与强启动子相连接。
49.在一些实施方式中,根据本公开所述的碳固定系统,其中,所述重组氢氧化细菌为贪铜菌属细菌;优选地,所述重组氢氧化细菌为钩虫贪铜菌或其衍生菌株。
50.第二方面,本公开提供了一种合成目标产物的方法,其中,所述方法包括如下步骤:
51.使重组氢氧化细菌与能源气体和碳源气体接触,对所述重组氢氧化细菌进行发酵培养,得到目标产物;优选地,所述目标产物为单细胞蛋白;
52.其中,所述重组氢氧化细菌包括通过氢氧化反应产生能量的氢元件,和通过固碳反应合成目标产物的固碳元件;
53.并且,与野生型氢氧化细菌相比,所述重组氢氧化细菌具有如下(i)
‑
(ii)中至少一项所示的特性:
54.(i)氢元件中一个或多个与氢氧化反应相关的蛋白活性,或蛋白编码基因的表达水平被增强;
55.(ii)固碳元件中一个或多个与固碳反应相关的蛋白活性,或蛋白编码基因的表达水平被增强;
56.优选地,所述能源气体包含氢气,所述碳源气体包含二氧化碳。
57.在一些实施方式中,根据本公开所述的方法,其中,所述发酵培养的温度为25
‑
35℃,所述发酵培养的ph为5.5
‑
8.0。
58.在一些实施方式中,根据本公开所述的方法,其中,使所述重组氢氧化细菌与能源气体接触的步骤包括:对包含重组氢氧化细菌的反应溶液进行电解,得到包含氢气的能源气体,重组氢氧化细菌与电解产生的氢气相接触;优选地,所述电解水的电压为
‑
1.5~
‑
3v,优选
‑
2.5v。
59.发明的效果
60.在一些实施方式中,本公开提供的碳固定系统,使用氢元件和固碳元件选择性加强的重组氢氧化细菌,对氢原料的利用高,且固碳反应的合成效率高,能够显著提高对单细胞蛋白等目标产物的产率。碳固定系统通过电解反应溶液产生能源气体,具体为电解水产生氢气,能够原位为氢氧化细菌提供能源,电解反应与固碳反应同步发生,无需气体收集再利用过程,具有成本降低、技术简单、容易控制、安全性高等优点。
61.在一些实施方式中,碳固定系统使用的重组氢氧化细菌,具有提高的核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的酶活性或蛋白编码基因的表达水平。通过提高重组氢氧化细菌内的固碳反应效率,进而提高目标产物的产率。
62.在一些实施方式中,碳固定系统使用的重组氢氧化细菌,其氢化酶的编码基因与强启动子相连接,通过增强氢化酶的表达,能够提高重组氢氧化细菌代谢h2产生能量的效
率,以提高重组氢氧化细菌的固碳反应效率。
63.在一些实施方式中,本公开提供的合成目标产物的方法,具有目标产物的产量高,且操作简单,合成过程容易控制,安全性高等优势,适合对单细胞蛋白的大规模工业化生产。
附图说明
64.图1示出了碳固定系统中电解水与固碳反应同步发酵产单细胞蛋白的过程示意图。
65.图2示出了不同电解电压下突变株m2的生长曲线(od
600
)。
66.图3示出了电解过程中超氧离子及过氧化氢浓度的浓度变化曲线。
67.图4示出了超氧化物歧化酶及过氧化氢酶在电驱动作用下的稳定性。
68.图5示出了不同酶浓度下突变株m2的生长曲线(od
600
)。
69.图6示出了大肠杆菌mg1655、氢氧细菌钩虫贪铜菌h16与突变株m2的生长曲线(od
600
)。
70.图7示出了突变株m2及m3的生长曲线(od
600
)。
具体实施方式
71.当在权利要求和/或说明书中与术语“包含”联用时,词语“一(a)”或“一(an)”可以指“一个”,但也可以指“一个或多个”、“至少一个”以及“一个或多于一个”。
72.如在权利要求和说明书中所使用的,词语“包含”、“具有”、“包括”或“含有”是指包括在内的或开放式的,并不排除额外的、未引述的元件或方法步骤。
73.在整个申请文件中,术语“约”表示:一个值包括测定该值所使用的装置或方法的误差的标准偏差。
74.虽然所公开的内容支持术语“或”的定义仅为替代物以及“和/或”,但除非明确表示仅为替代物或替代物之间相互排斥外,权利要求中的术语“或”是指“和/或”。
75.当用于权利要求书或说明书时,选择/可选/优选的“数值范围”既包括范围两端的数值端点,也包括相对于前述数值端点而言,所述数值端点中间所覆盖的所有自然数。
76.如本公开所使用的,术语“野生型”指在自然界中可以找到的对象。例如,一种可以从自然界的一个来源中分离出来并且在实验室中没有被人类有意修改的多肽、多核苷酸序列或微生物是天然存在的。如本公开所用的,“天然存在”和“野生型”是同义词。
77.如本公开所使用的,术语“微生物”是肉眼难以观测的微小生物的总称,包括细菌、真菌等等。由于微生物的表面积与体积比很大,能够快速与外界环境进行物质交换,产生代谢产物。本公开中的微生物特指能够发酵培养,产生糖类、脂质、氨基酸、核苷酸等代谢产物的发酵微生物。
78.如本公开所使用的,术语“氢氧化细菌”(hydrogen
‑
oxidizing bacteria)利用氢气作为电子供体,将二氧化碳还原为有机物。氢氧化细菌为兼性化能自养细菌,其包括通过氢氧化反应产生能量的氢元件,和通过固碳反应合成目标产物的固碳元件。
79.示例性的,氢氧化细菌包括但不限于假单胞菌属(pseudomonas)、副球菌属(paracoccus)、黄杆菌属(flavobacterium)、分支杆菌属(mycobacterium)、产碱菌属
(alcaligenes)、诺卡氏菌属(nocardia)、醋酸杆菌属(acetobacter)、棒杆菌属(corynebacterium)、贪铜菌属(cupriavidus)等。在一些实施方式中,氢氧化细菌为贪铜菌属(cupriavidus)细菌。在一些具体的实施方式中,氢氧化细菌为钩虫贪铜菌(cupriavidus necator)。
80.如本公开所使用的,术语“能源气体”是指能够被氢氧化细菌利用,通过代谢反应产生能量的气体。在一些实施方式中,能源气体为包含氢气的气体。氢氧化细菌以氢气为电子供体,通过氢氧化反应产生其固碳反应所需的能量。
81.如本公开所使用的,术语“重组微生物”是以基因工程方法对野生型微生物或亲本微生物进行改造,得到的不同于野生型微生物或亲本微生物的改造的微生物。实施方案包括但不限于引入微生物的外源基因、可操作地连接到异源启动子的内源蛋白质编码序列和具有经修改的蛋白质编码序列的基因。重组基因保存在微生物的基因组、微生物中的质粒或微生物中的噬菌体上。
82.进一步的,本公开中的“重组微生物”为“重组氢氧化细菌”。
83.如本公开所使用的,术语“衍生菌株”包括其中一个或多个内源基因或序列被缺失或修饰和/或一个或多个外源基因或序列被引入的修饰的生物体。
84.如本公开所使用的,术语“异养”指代不能合成生物从二氧化碳生存和生长所需的所有有机化合物,并且必须利用有机化合物进行生长的生物。
85.如本公开所使用的,术语“自养”指代能合成生物从二氧化碳生存和生长所需的所有有机化合物,并且可利用其进行生长的生物。
86.如本公开所使用的,术语“核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶”(ribulose
‑
1,5
‑
bisphosphate carboxylase/oxygenase)是光合作用固定co2的关键酶,它催化核酮糖
‑
1,5
‑
二磷酸(rubp)的羧化反应,此反应是光合碳还原(pcr)循环,也即calvin
‑
benson
‑
bassham循环的限速步骤。除此之外,rubp还催化rubp与o2的反应,将氧加在rubp上。
87.如本公开所使用的,术语“分子伴侣”(chaperone)又称为侣伴蛋白(molecular chaperone),是一类协助细胞内分子组装和协助蛋白质折叠的蛋白质。groes、groel是来源于大肠杆菌的分子伴侣,在肽链的折叠过程中发挥重要作用,能够促进蛋白的正确折叠、组装。
88.如本公开所使用的,术语“氢化酶”以h2为电子供体,通过催化h2的氧化代谢产生能量。示例性的,氢化酶包括膜结合的氢化酶和可溶性的氢化酶。其中,膜结合的氢化酶使得细菌可以将h2氧化成h
并释放能量,生成的电子经电子传递链传递给o2。而可溶性氢化酶则直接催化h2作为还原剂使nad
还原成nadh。
89.如本公开所使用的,术语“超氧化物歧化酶”(superoxide dismutase,sod)是生物体内存在的一种抗氧化金属酶,它能够催化超氧阴离子自由基歧化生成氧和过氧化氢。
90.如本公开所使用的,术语“过氧化氢酶”是一类广泛存在于生物体中的抗氧化剂,其作用是催化过氧化氢转化为水和氧气的反应。
91.如本公开所使用的,术语“编码基因”是指能够通过一定的规则指导蛋白的合成dna分子,蛋白编码基因指导蛋白合成的过程一般包括以双链dna为模板的转录过程和以mrna为模板的翻译过程。编码基因含有cds序列(coding sequence),能够指导编码蛋白质的mrna的产生。
92.如本公开所使用的,术语“表达”包括涉及rna产生及蛋白产生的任何步骤,包括但不限于:转录、转录后修饰、翻译、翻译后修饰和分泌。
93.如本公开所使用的,术语“多肽”、“酶”、“多肽或酶”、“多肽/酶”具有相同的含义,其在本公开中可以互换。前述术语是指一种由和很多氨基酸通过肽键组成的聚合物,其可能含有或可能不含有如磷酸基和甲酰基的修饰。
94.如本公开所使用的,术语“酶活性”“蛋白活性”也被表述为“比活力”或“比活度”,其在本公开中具有相同的含义,可以互换使用。其是指每毫克多肽(酶、蛋白)的酶活力(u/mg)、蛋白活力(u/mg)。
95.如本公开所使用的,术语“可操作地连接”是指如下的构造:调控序列相对于多核苷酸的编码序列安置在适当位置,从而使得该调控序列指导该编码序列的表达。示例性的,所述调控序列可以选自启动子和/或增强子编码的序列。
96.如本公开所使用的,术语“基因工程改造”是指任何对野生型菌株或亲本菌株进行的遗传操作,包括但不限于各种分子生物学手段。本公开中的术语“修饰”、“基因编辑”、“基因改造”、“基因工程改造”可相互替换。
97.如本公开所使用的,术语“内源的”指在生物体或细胞内自然表达或产生的多核苷酸、多肽或其他化合物。也就是说,内源性多核苷酸、多肽或其他化合物不是外源的。例如,当细胞最初从自然界分离时,细胞中存在一种“内源性”多核苷酸或多肽。
98.如本公开所使用的,术语“外源的”指在需要表达的特定细胞或有机体中天然发现或表达的任何多核苷酸或多肽。外源多核苷酸、多肽或其他化合物不是内源性的。
99.如本公开所使用的,术语“氨基酸突变”或“核苷酸突变”,包括“取代、重复、缺失或添加一个或多个氨基酸或核苷酸”。在本公开中,术语“突变”是指核苷酸序列或者氨基酸序列的改变。在一个具体的实施方式中,术语“突变”是指“取代”。
100.在一个实施方式中,本公开的“突变”可以选自“保守突变”。在本公开中,术语“保守突变”是指可正常维持蛋白质的功能的突变。保守突变的代表性例子为保守置换。
101.如本公开所使用的,术语“保守置换”涉及用具有类似侧链的氨基酸残基替换氨基酸残基。本领域已经定义了具有类似侧链的氨基酸残基家族,并且包括具有碱性侧链(例如赖氨酸、精氨酸和组氨酸)、酸性侧链(例如天冬氨酸和谷氨酸)、不带电极性侧链(例如甘氨酸、天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、酪氨酸、和半胱氨酸)、非极性侧链(例如丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、蛋氨酸和色氨酸)、β
‑
支链(例如苏氨酸、缬氨酸和异亮氨酸)和芳香侧链(例如酪氨酸、苯丙氨酸、色氨酸和组氨酸)。如本公开所使用的,“保守置换”通常在蛋白质的一个或多个位点上交换一种氨基酸。这种取代可以是保守的。作为被视作保守置换的置换,此外,保守突变还包括起因于基因所来源的个体差异、株、种的差异等天然产生的突变。
102.如本公开所使用的,术语“多核苷酸”指由核苷酸组成的聚合物。多核苷酸可以是单独片段的形式,也可以是更大的核苷酸序列结构的一个组成部分,其是从至少在数量或浓度上分离一次的核苷酸序列衍生而来的,能够通过标准分子生物学方法(例如,使用克隆载体)识别、操纵以及恢复序列及其组分核苷酸序列。当一个核苷酸序列通过一个dna序列(即a、t、g、c)表示时,这也包括一个rna序列(即a、u、g、c),其中“u”取代“t”。换句话说,“多核苷酸”指从其他核苷酸(单独的片段或整个片段)中去除的核苷酸聚合物,或者可以是一
个较大核苷酸结构的组成部分或成分,如表达载体或多顺反子序列。多核苷酸包括dna、rna和cdna序列。“重组多核苷酸”属于“多核苷酸”中的一种。
103.如本公开所使用的,术语“载体”指的是dna构建体,其含有与合适的控制序列可操作地连接的dna序列,从而在合适的宿主中表达目的基因。“重组表达载体”指用于表达例如编码所需多肽的多核苷酸的dna结构。重组表达载体可包括,例如包含i)对基因表达具有调控作用的遗传元素的集合,例如启动子和增强子;ii)转录成mrna并翻译成蛋白质的结构或编码序列;以及iii)适当的转录和翻译起始和终止序列的转录亚单位。重组表达载体以任何合适的方式构建。载体的性质并不重要,并可以使用任何载体,包括质粒、病毒、噬菌体和转座子。用于本公开的可能载体包括但不限于染色体、非染色体和合成dna序列,例如细菌质粒、噬菌体dna、酵母质粒以及从质粒和噬菌体dna的组合中衍生的载体,来自如牛痘、腺病毒、鸡痘、杆状病毒、sv40和伪狂犬病等病毒的dna。
104.如本公开所使用的,术语“引入”包括“转染”、“转化”或“转导”并且指代将核酸序列并入至真核或原核细胞中,在所述真核或原核细胞,核酸序列可并入至所述细胞的基因组(例如,染色体、质粒、质体或线粒体dna)中,转化成能独立复制的复制子,或独立于细胞的基因组表达。“转染”、“转化”或“转导”的方法包括但不限于电穿孔法、磷酸钙(capo4)沉淀法、氯化钙(cacl2)沉淀法、微注射法、聚乙二醇(peg)法、deae
‑
葡聚糖法、阳离子脂质体法以及乙酸锂
‑
dmso法。
105.如本公开所使用的,术语“培养”指代使细胞(例如微生物细胞)群体在合适的生长条件下、在液态或固态培养基中生长。
106.如本公开所使用的,术语“产率”具有本领域技术人员普遍理解的意思,即生成目标产物所消耗的底物占总的底物的百分比。在本公开中,“目标产物产率”和“底物转化率”可以相互替代使用。
107.如本公开所使用的,术语“产量”指代由进料产生的产物相对于如果所有进料物质转化为产物将产生的物质总量的量。举例来说,氨基酸产率可以表示为相对于如果100%的进料转化为氨基酸的理论产率,所产生的氨基酸的%。
108.本公开的重组微生物的培养可以根据本领域的常规方法进行,包括但不限于孔板培养、摇瓶培养、批次培养、连续培养和分批补料培养等,并可以根据实际情况适当地调整各种培养条件如温度、时间和培养基的ph值等。
109.除非另外定义或由背景清楚指示,否则在本公开中的全部技术与科学术语具有如本公开所属领域的普通技术人员通常理解的相同含义。
110.重组氢氧化细菌
111.本公开中的重组氢氧化细菌,包括通过氢氧化反应产生能量的氢元件,和通过固碳反应合成目标产物的固碳元件;并且,与野生型氢氧化细菌相比,所述重组微生物具有如下(i)
‑
(ii)中至少一项所示的特性:
112.(i)氢元件中一个或多个与氢氧化反应相关的蛋白活性,或蛋白编码基因的表达水平被增强;
113.(ii)固碳元件中一个或多个与固碳反应相关的蛋白活性,或蛋白编码基因的表达水平被增强。
114.重组氢氧化细菌具有加强的氢元件,以及固碳元件,其利用氢气代谢生产能量的
利用率提高,可为固碳元件提供丰富的能量来源,进一步加强固碳元件合成目标产物的反应效率,使重组氢氧化细菌合成目标产物的产量和速率显著提升。
115.在一些实施方式中,与固碳反应相关的蛋白包括核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶,核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶是卡尔文循环固定co2的关键酶,通过加强核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的酶活性或蛋白编码基因的表达水平,能够提高固碳元件固定co2的效率,进而提高重组氢氧化细菌合成单细胞蛋白等目标产物的产率。
116.在一些可选地实施方式中,核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶包括第一亚基,伴侣蛋白,以及第二亚基。在一些优选的实施方式中,核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶由第一亚基,伴侣蛋白,以及第二亚基组成。
117.在一些可选地实施方式中,第一亚基为氨基酸序列如seq id no:1所示的多肽,或者如seq id no:1所示氨基酸序列经过取代、重复、缺失或添加一个或多个氨基酸,且具有第一亚基活性的多肽。
118.在一些可选地实施方式中,第一亚基是由于与seq id no:6所示的核苷酸序列具有至少90%的序列同一性的序列编码的,具有第一亚基活性的多肽。示例性的,编码第一亚基的核苷酸序列与seq id no:6所示序列具有至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%的序列同一性。
119.在一些可选地实施方式中,伴侣蛋白为氨基酸序列如seq id no:2所示的多肽,或者如seq id no:2所示氨基酸序列经过取代、重复、缺失或添加一个或多个氨基酸,且具有伴侣蛋白活性的多肽。
120.在一些可选地实施方式中,伴侣蛋白是由于与seq id no:7所示的核苷酸序列具有至少90%的序列同一性的序列编码的,具有伴侣蛋白活性的多肽。示例性的,编码伴侣蛋白的核苷酸序列与seq id no:7所示序列具有至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%的序列同一性。
121.在一些可选地实施方式中,第二亚基为氨基酸序列如seq id no:3所示的多肽,或者如seq id no:3所示氨基酸序列经过取代、重复、缺失或添加一个或多个氨基酸,且具有第二亚基活性的多肽。
122.在一些可选地实施方式中,第二亚基是由于与seq id no:8所示的核苷酸序列具有至少90%的序列同一性的序列编码的,具有第二亚基活性的多肽。示例性的,编码第二亚基的核苷酸序列与seq id no:8所示序列具有至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%的序列同一性。
123.在一些具体的实施方式中,通过向氢氧化细菌中导入外源的核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的编码基因,以上调氢氧化细菌中核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的表达水平。进一步的,是通过向氢氧化细菌中导入由第一亚基,伴侣蛋白,以及第二亚基组成的核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的编码基因。
124.在一些优选的实施方式中,与固碳反应相关的蛋白还包括groes、groel,groes、groel是促进核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的蛋白折叠、组装的分子伴侣,可促进核酮
糖
‑
1,5
‑
二磷酸羧化酶/加氧酶形成正确的空间构象,从而使其具有高的酶活性。
125.在一些可选地实施方式中,groes为包含如seq id no:4所示氨基酸序列的,并且具有groes蛋白活性的多肽。作为优选,groes的氨基酸序列如seq id no:4所示。groel为包含如seq id no:5所示氨基酸序列的,并且具有groel蛋白活性的多肽。作为优选,groel的氨基酸序列如seq id no:5所示。
126.在一些可选地实施方式中,groes为如seq id no:4所示氨基酸序列经过取代、重复、缺失或添加一个或多个氨基酸,且具有groes蛋白活性的多肽。groel为如seq id no:5所示氨基酸序列经过取代、重复、缺失或添加一个或多个氨基酸,且具有groel蛋白活性的多肽。
127.在一些可选地实施方式中,groes是由与seq id no:9所示的核苷酸序列具有至少90%的序列同一性的序列编码的,具有groes蛋白活性的多肽。示例性的,编码groes的核苷酸序列与seq id no:9所示序列具有至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%的序列同一性。groel是由与seq id no:10所示的核苷酸序列具有至少90%的序列同一性的序列编码的,具有groel蛋白活性的多肽。示例性的,编码groel的核苷酸序列与seq id no:10所示序列具有至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%的序列同一性。
128.在一些具体的实施方式中,通过向氢氧化细菌中导入外源的groes、groel的编码基因,实现在氢氧化细菌中表达groes、groel,从而促进核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的蛋白折叠、组装。进一步的,是通过向氢氧化细菌中导入核苷酸序列如seq id no:9所示的编码基因,以及核苷酸序列如seq id no:10所示的编码基因。
129.在一些实施方式中,所述重组氢氧化细菌包含表达核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的重组表达载体;利用重组表达载体提高重组氢氧化氢细菌内核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的表达水平。在一些优选地实施方式中,所述重组氢氧化细菌包含表达核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶、groes和groel的重组表达载体。利用包含核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶、groes和groel的重组表达载体,能够在重组氢氧化氢细菌内高水平表达具有正确空间结构的核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶。
130.在一些具体的实施方式中,所述重组表达载体的核苷酸序列如seq id no:11或seq id no:12所示。
131.在一些实施方式中,与氢氧化反应相关的蛋白包括氢化酶。氢化酶能够以氢气为电子供体,通过氧化氢气代谢产生能量,用于卡尔文循环等固碳反应。重组氢氧化细菌中氢化酶的酶活性提高,或氢化酶的编码基因的表达水平提高,能够提高氢氧化细菌对氢气的利用率,进而提高重组氢氧化细菌内代谢氢产生的能量,促进固碳反应的发生。
132.在一些实施方式中,氢氧化细菌内的氢化酶包括但不限于:膜连接氢化酶(mbh)、可溶性氢化酶(sh)、调节性氢化酶(rh)、假设性氢化酶(hy4)。此处的氢元件为膜连接氢化酶(mbh)及可溶性氢化酶(sh)。在一些实施方式中,重组氢氧化细菌中与氢氧化反应相关的蛋白活性,或蛋白编码基因的表达水平被增强,是指膜连接氢化酶(mbh)、可溶性氢化酶(sh)中的一种或两种酶的活性增强或表达水平增强。
133.优选地,所述重组氢氧化细菌中氢化酶的编码基因与强启动子相连接。示例性的,
将氢化酶的编码基因的启动子优化为bba_j23119,以提高重组氢氧化细菌中氢化酶的表达水平。
134.在一些实施方式中,所述重组氢氧化细菌为贪铜菌属细菌。在一些优选地实施方式中,所述重组氢氧化细菌为钩虫贪铜菌或其衍生菌株。在另外一些实施方式中,重组氢氧化细菌还可以是其他种类的氢氧化细菌,例如,假单胞菌属细菌、棒杆菌属细菌等等。
135.碳固定系统
136.本公开的碳固定系统包括:
137.反应容器,具有容置反应溶液的反应内腔,所述反应溶液中包含根据本公开所述的重组氢氧化细菌;
138.电解单元,所述电解单元部分伸入所述反应溶液中,用于电解反应溶液产生能源气体;
139.通气单元,所述通气单元用于向所述反应溶液内通入碳源气体。
140.碳固定系统通过电解水产生包含氢气的能源气体,为重组氢氧化细菌原位提供能源,避免了氢气收集再利用的过程。能够降低碳固定系统合成目标产物的成本、提高使用安全性。重组氢氧化细菌利用能源气体和碳源气体,合成单细胞蛋白等目标产物。本公开中的重组氢氧化细菌对氢能源的利用效率高,固碳反应的效率高,包含重组氢氧化细菌的碳固定系统具有显著提高的目标产物产量和生产速率。碳固定系统用于生产单细胞蛋白等目标产物,具有产量高、技术简单、容易控制、安全性高的优势,适合规模化生产。
141.在一些实施方式中,能源气体包含氢气,碳源气体包含二氧化碳;重组氢氧化细菌以氢气为能源,以二氧化碳为碳源,高效合成单细胞蛋白等目标产物。在一些优选的实施方式中,每分钟向所述反应溶液内通入碳源气体的体积与所述反应内腔的体积比为(2
‑
5):10。示例性的,当反应内腔的体积为100ml时,则向反应溶液内通入碳源气体的速率为20
‑
50ml/min。进一步的,碳源气体通入的流速为20ml/min、25ml/min、30ml/min、35ml/min、40ml/min、50ml/min等等。通过控制气体通入反应溶液中的流速,对于提高重组氢氧化细菌的生长速率,进而提高目标单细胞蛋白的产量和生产速率具有重要意义。
142.在一些实施方式中,反应容器具有容置反应溶液的反应内腔,以及将反应容器的反应内腔与外界连通的第一连通口。电解单元和通气单元可以通过连通口与反应内腔中的反应溶液接触,提供重组氢氧化细菌所需要的能源气体和碳源气体。本公开中的反应容器可以是能够实现重组氢氧化细菌的发酵培养的任意类型的生物反应器。
143.进一步的,反应容器上还开设有用于向反应内腔中通入反应溶液的第二连通口。第一连通口和第二连通口可以是同一开口或是不同开口。在一些可选地实施方式中,通过第二连通口,可以向反应内腔中周期性地提供反应溶液。
144.在一些实施方式中,所述反应溶液是包含磷酸盐的发酵培养基。其中,发酵培养基中磷酸盐的浓度为25
‑
35mm,示例性的,发酵培养基中磷酸盐的浓度为26mm、28mm、29mm、30mm、32mm、34mm等等。在一些优选的实施方式中,发酵培养基中磷酸盐的浓度为30mm。在一些具体的实施方式中,所述磷酸盐选自磷酸二氢钾、磷酸氢二钾、磷酸二氢钠、磷酸氢二钠中的一种或两种以上的组合。在一些优选的实施方式中,磷酸盐为磷酸二氢钠、磷酸氢二钠中的一种或两种。
145.进一步的,发酵培养基中还包含镍、亚铁、铁、钙、镁等离子。在一些优选的实施方
式中,发酵培养基中包含亚铁盐。示例性的,亚铁盐为氯化亚铁、硫酸亚铁等。在一些优选的实施方式中,亚铁盐为硫酸亚铁,发酵培养基中硫酸亚铁的浓度为30
‑
60mg/l,示例性的,发酵培养基中硫酸亚铁的浓度为35mg/l、38mg/l、40mg/l、42mg/l、45mg/l、48mg/l、50mg/l、52mg/l、54mg/l、56mg/l、57mg/l、59mg/l等等。在一些优选的实施方式中,发酵培养基中硫酸亚铁的浓度为50mg/l。
146.本公开中的发酵培养基适于重组氢氧化细菌的自养培养,能够维持重组氢氧化细菌的快速生长、代谢。
147.在另外一些实施方式中,发酵培养基中还可以包含其他种类的磷酸盐,或其他种类的亚铁盐,只要其能够形式适于重组氢氧化细菌的生长、代谢的营养环境即可。
148.在一些实施方式中,反应溶液中包含超氧化物歧化酶、过氧化氢酶的至少一种。通过添加超氧化物歧化酶、过氧化氢酶,有利于减轻对重组氢氧化细菌的生长抑制因素,维持重组氢氧化细菌的快速生长、增殖活性。进一步的,反应溶液中超氧化物歧化酶的含量为≥7u/ml,过氧化氢酶的含量为≥20u/ml;示例性的,过氧化氢酶的含量为23u/ml、超氧化物歧化酶的含量为7.5u/ml,过氧化氢酶的含量为46u/ml、超氧化物歧化酶的含量为15u/ml,过氧化氢酶的含量为90u/ml、超氧化物歧化酶的含量为30u/ml,过氧化氢酶的含量为180u/ml、超氧化物歧化酶的含量为60u/ml,过氧化氢酶的含量为230u/ml、超氧化物歧化酶的含量为75u/ml等等。当过氧化氢酶的含量为至少180u/ml、超氧化物歧化酶的含量为至少60u/ml时,重组氢氧化细菌具有高的生长、繁殖速度,在生长7天后od
600
可达8.9以上。而当23u/ml≤过氧化氢酶<180u/ml,7.5u/ml≤超氧化物歧化酶<60u/ml时,od
600
可达到2
‑
8的范围。
149.在一些优选的实施方式中,超氧化物歧化酶来源于热栖热菌(thermus thermophilus,uniprot编号p61503)、嗜热毛壳菌(chaetomium thermophilum,uniprot编号q1heq0)、嗜热子囊菌(thermoascus aurantiacus,uniprot编号q1hdv5)、大肠杆菌(escherichia coli,uniprot编号p00448、p0agd1、p0agd3等)、枯草芽孢杆菌(bacillus subtilis,uniprot编号p54375)、酿酒酵母(saccharomyces cerevisiae,uniprot编号p00447)等。
150.在一些优选的实施方式中,过氧化氢酶来源于大肠杆菌(escherichia coli,uniprot编号p13029)、流产布鲁氏菌(brucella abortus,uniprot编号p0a327)、黑曲霉菌(aspergillus niger,uniprot编号a2q7t1)、枯草芽孢杆菌(bacillus subtilis,uniprot编号q9aqq9)、布氏栖热菌(thermus brockianus,uniprot编号q596k8)、粗糙脉孢菌(neurospora crassa,uniprot编号q8x182)、粘质沙雷氏菌(serratia marcescens,uniprot编号a0a221fs36、a0a5c7ble8等)等。
151.电解单元用于电解反应溶液产生包含氢气的能源气体,具体地,电解单元用于电解水产生氢气。在一些实施方式中,电解单元包括阳极电极和阴极电极,阳极电极和阴极电极分别部分地伸入反应溶液中。其中,阳极电极和阴极电极伸入反应溶液的一端为电解端;阳极电极和阴极电极中远离所述电解端的另一端与电源相连,为电连接端。在通电后,电解单元的阳极处发生析氧反应,阴极处发生析氢反应,从而将水电解为氢气和氧气。
152.在一些可选地实施方式中,所述阳极电极为钛片,所述阴极电极为镍片。利用两者作为电解单元的电极,能够完成电解水、产生氢气和氧气的过程。在另外一些实施方式中,
电解单元也可以由其他种类的阳极材料、阴极材料形成阳极电极和阴极电极。
153.进一步的,对应阳极电极和阴极电极,可以将反应容器的反应内腔分为阳极室和阴极室。阳极电极的电解端位于阳极室中,阴极电极的电解端位于阴极室中。并且,重组氢氧化细菌位于电解单元的阴极室中,靠近阴极电极伸入所述反应溶液的电解端,为重组氢氧化细菌充分利用由阴极电极产生的氢气提供便利。
154.在一些优选的实施方式中,阴极室和阳极室之间还可设置分隔层,避免重组氢氧化细菌流向阳极室内。例如,分隔层可以是位于阴极室和阳极室之间的质子交换膜。
155.通气单元用于向所述反应溶液内通入包含二氧化碳的碳源气体,在一些实施方式中,通气单元包括伸入反应溶液中的管路,以及与所述管路相连接的储气瓶。在一定压力作用下,将储气瓶内的二氧化碳气体经管路输入反应溶液中,为重组氢氧化细菌提供碳原料。二氧化碳气体可以是二氧化碳的含量超过99%的纯净气体,或是含有一定量其他种类气体的混合气体。在一些优选的实施方式中,
156.显而易见的是,具有电解水与重组氢氧化细菌处于同一空间碳固定系统落入本公开的范围内,并不排除能够通过其他电极或电解方式或非钩虫贪铜菌的氢氧化细菌,只要它具有对应的功能并表现出对应功效即可。
157.本公开中的碳固定系统,通过优化反应溶液的物质含量,能够有效解除重组氢氧化细菌的生长抑制条件,实现生物量的积累;重组氢氧化细菌在优选条件下生长7天后od
600
可到达8.9,显著提高在电驱动条件下碳固定系统生产单细胞蛋白的产量。并且,通过使用氢元件和固碳元件优化的重组氢氧化细菌,能够显著提高碳固定系统生产单细胞蛋白的产率。与未优化的氢氧化细菌相比,本公开中使用重组氢氧化细菌的碳固定系统生产单细胞蛋白的速率可提高1.7倍,具有规模化工业应用的价值。
158.合成目标产物的方法
159.本公开提供的合成目标产物的方法,包括如下步骤:使根据本公开所述的重组氢氧化细菌与氢气和二氧化碳接触,对所述重组氢氧化细菌进行发酵培养,得到目标产物。
160.在一些实施方式中,使重组氢氧化细菌与氢气接触的步骤包括:对包含重组氢氧化细菌的反应溶液进行电解,使重组氢氧化细菌与电解水产生的氢气相接触。在一些优选的实施方式中,电解水的电压为
‑
1.5~
‑
3v,优选
‑
2.5v。示例性的,电解水的电压为
‑
1.5v、、
‑
1.8v、
‑
2v、
‑
2.3v、
‑
2.5v、
‑
2.8v、
‑
3v等等。
161.在一些实施方式中,发酵培养的温度为25
‑
35℃,示例性的,发酵培养的温度为26℃、27℃、28℃、29℃、30℃、31℃、32℃、33℃等等。在一些优选的实施方式中,发酵培养的温度为30℃。在一些实施方式中,发酵培养的ph为5.5
‑
8.0,示例性的,发酵培养的ph为5.5
‑
6,6
‑
6.5,6.5
‑
7,7
‑
7.5,7.5
‑
8等等。在一些优选地实施方式中,发酵培养的ph为6.0
‑
7.0。
162.在一些具体的实施方式中,本公开中合成目标产物的方法,包括使用本公开所述的碳固定元件合成目标产物的步骤。
163.在一些优选的实施方式中,目标产物为单细胞蛋白。本公开提供的方法中,进一步对发酵条件和水解电压进行优化,从而为重组氢氧化细菌适合其生长、代谢产生单细胞蛋白的环境条件,提高发酵生产单细胞蛋白的产量及速率。
164.在本领域,用于操纵微生物的方法是已知的,如《分子生物学现代方法》(online isbn:9780471142720,john wiley and sons,inc.)、《微生物代谢工程:方法和规程》
(qiong cheng ed.,springer)和《系统代谢工程:方法和规程》(hal s.alper ed.,springer)等出版物中被解释。
165.在本公开中,核苷酸或氨基酸的编号的含义如下:
166.seq id no:1所示的序列是来源于synechococcus sp.pcc 7002的核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的第一亚基的氨基酸序列;
167.seq id no:2所示的序列是来源于synechococcus sp.pcc 7002的核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的伴侣蛋白的氨基酸序列;
168.seq id no:3所示的序列是来源于synechococcus sp.pcc 7002的核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的第二亚基的氨基酸序列;
169.seq id no:4所示的序列是来源于c.necator h16的groes的氨基酸序列;
170.seq id no:5所示的序列是来源于c.necator h16的groel的氨基酸序列;
171.seq id no:6所示的序列是来源于synechococcus sp.pcc 7002的核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的第一亚基的核苷酸序列;
172.seq id no:7所示的序列是来源于synechococcus sp.pcc 7002的核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的伴侣蛋白的核苷酸序列;
173.seq id no:8所示的序列是来源于synechococcus sp.pcc 7002的核酮糖
‑
1,5
‑
二磷酸羧化酶/加氧酶的第二亚基的核苷酸序列;
174.seq id no:9所示的序列是来源于c.necator h16的groes的核苷酸序列;
175.seq id no:10所示的序列是来源于c.necator h16的groel的核苷酸序列;
176.seq id no:11所示的序列重组质粒pbbr
‑
sprubisco的核苷酸序列;
177.seq id no:12所示的序列重组质粒pbbr
‑
syprubisco
‑
cngroesl的核苷酸序列。
178.实施例
179.本公开的其他目的、特征和优点将从以下详细描述中变得明显。但是,应当理解的是,详细描述和具体实施例(虽然表示本公开的具体实施方式)仅为解释性目的而给出,因为在阅读该详细说明后,在本公开的精神和范围内所作出的各种改变和修饰,对于本领域技术人员来说将变得显而易见。
180.本实施例中所用到的实验技术与实验方法,如无特殊说明均为常规技术方法,例如下列实施例中未注明具体条件的实验方法,通常按照常规条件如sambrook等人,分子克隆:实验室手册(new york:cold spring harbor laboratory press,1989)中所述的条件,或按照制造厂商所建议的条件。
181.除非另有说明,以下实施例中使用的原料和试剂均为市售商品,或者可以通过已知方法制备。
182.本发明实施例中使用的部分材料信息如下:
183.pet20b、pet28a载体,novagen,madison,wi;
184.大肠杆菌表达菌bl21(de3),invitrogen,carlsbad,ca;
185.实施例1:超氧化物歧化酶的表达及酶活测定
186.来源于嗜热栖热菌(thermus thermophilus,uniprot编号p61503)的超氧化物歧化酶基因的dna序列可从uniprot的官方网站(https://www.uniprot.org/)上获得。通过simple cloning
3.的方法,将上述基因克隆至pet20b载体(novagen,madison,wi)中,获得
相应的表达载体pet20b
‑
ttcsod。
187.重组蛋白在大肠杆菌bl21(de3)中表达,经70℃热处理30分钟对其破碎粗酶液进行纯化,并通过检测抑制50%连苯三酚自氧化确定其活性。实验结果显示,嗜热栖热菌来源的超氧化物歧化酶在25℃下的比酶活为120u/mg。
188.实施例2:过氧化氢酶的表达及酶活测定
189.来源于大肠杆菌(escherichia coli,uniprot编号p13029)的过氧化氢酶基因的dna序列可从uniprot的官方网站(https://www.uniprot.org/)上获得。通过simple cloning的方法,将上述基因克隆至pet20b载体(novagen,madison,wi)中,获得相应的表达载体pet20b
‑
eccat。
190.重组蛋白在大肠杆菌bl21(de3)中表达,经镍柱纯化对其破碎粗酶液进行纯化,并通过检测过氧化氢在240nm下吸光值降低计算其活性。结果显示,大肠杆菌来源的过氧化氢酶在25℃下的比酶活为3000u/mg。
191.实施例3:重组氢氧化细菌的构建
192.(1)在野生型cupriavidus necator h16菌株的基础上,采用pk18mobsacb质粒敲除了两个限制性内切酶基因h16_a0006和h16_a0008
‑
9后,获得m1菌株(cupriavidus necator h16δh16_a0006δh16_a0008
‑
9)。
193.(2)在m1菌株的基础上,把氢元件的启动子(具体为膜连接氢化酶(mbh)及可溶性氢化酶(sh)的启动子)优化为bba_j23119,获得m2菌株。
194.(3)选择synechococcus sp.pcc 7002的rubisco,先利用pcr扩增把rubisco大小亚基(genbank:cp000951.1)从基因组上克隆下来,利用双酶切(酶切位点为ndei及saci),连接转化至pbbr
‑
mcs质粒骨架上,构成重组质粒pbbr
‑
sprubisco。
195.把来源于c.necator h16的分子伴侣groes及groel基因利用同样的方式连接至上述质粒上,构建重组质粒pbbr
‑
syprubisco
‑
cngroesl。
196.把pbbr
‑
syprubisco
‑
cngroesl通过接合转移方式转入至m2菌株中,获得m3菌株。
197.实施例4:m2菌株通气发酵产单细胞蛋白
198.此处氢气来源为氢气发生器,二氧化碳及氧气为气瓶,调整通气比例h2:o2:co2为7:1:1,h16/m2/m3异养转接后初始od为0.2左右,在发酵培养基2(medium 2)包括:9.0g/l na2hpo4·
12h2o,1.5g/l kh2po4,1.0g/l(nh4)2so4,80mg/l mgso4·
7h2o,1mg/lcaso4·
2h2o,0.56mg/l niso4·
7h2o,0.4mg/l柠檬酸三铁(ferric citrate),200mg/lnahco3,50mg/l feso4·
7h2o下自养培养24小时后再次进行转接至电解阴极室,用于后续碳固定系统发酵产单细胞蛋白。
199.实施例5:碳固定系统发酵产单细胞蛋白
200.碳固定系统的反应容器设置为阳极室和阴极室的双室:阳极室电极为钛片(3*5cm),阳极室中反应溶液为2mm h2so4,为恢复维持阴极因析出氢气后导致ph升高而不适宜氢氧化细菌生长的ph环境。阴极室电极为镍片(3*5cm),阴极室中反应溶液为包含如下成分的基础培养基:9.0g/l na2hpo4·
12h2o,1.5g/l kh2po4,1.0g/l(nh4)2so4,80mg/l mgso4·
7h2o,1mg/l caso4·
2h2o,0.56mg/l niso4·
7h2o,0.4mg/l ferric citrate,200mg/l nahco3,50mg/l feso4·
7h2o。阳极室与阴极室中间利用nafion 212隔开确保只允许质子通过,参比电极为ag/agcl。
201.用于固碳产单细胞蛋白的钩虫贪铜菌突变菌株m2菌株先经异养培养后,转接经自养培养24小时后再次进行转接至阴极室,初始od
600
为0.15
‑
0.25。添加过氧化氢酶量为23u/ml,超氧化物歧化酶添加量为7.5u/ml,电解电压为
‑
2.5v,向阴极按照流速10ml/min通混合气(co2与o2比例为1:1),并同时补充流速30ml/min的co2气体。此时初始电流为30ma左右。保持发酵反应的温度为30℃,ph为6
‑
7。每天补充同量过氧化氢酶与超氧化物歧化酶,经四天发酵后,od
600
可达2.33
±
0.17。
202.实施例6:电解过程中电压及酶量的优化过程
203.在实施例5条件基础上调整电压为
‑
1.5v、
‑
2v、
‑
2.5v,结果如图2所示,电压为
‑
2.5v时,od为2.32
±
0.17,较
‑
2.0v(od为2.03
±
0.11)增长幅度与
‑
2.0v与
‑
1.5v(od为1.06
±
0.07)的增效相比小很多,考虑能效问题,因此后续电压选取
‑
2.5v。
204.实施例7:电解过程中超氧离子及过氧化氢浓度测定
205.碳固定系统的反应容器设置为阳极室和阴极室的双室:阳极室电极为钛片(3*5cm),阳极室中反应溶液为2mm h2so4。阴极室电极为镍片(3*5cm),阴极室中反应溶液为包含如下成分的基础培养基:9.0g/l na2hpo4·
12h2o,1.5g/l kh2po4,1.0g/l(nh4)2so4,80mg/l mgso4·
7h2o,1mg/l caso4·
2h2o,0.56mg/l niso4·
7h2o,0.4mg/l ferric citrate,200mg/l nahco3,50mg/l feso4·
7h2o。参比电极为ag/agcl,阳极室与阴极室中间利用nafion 212隔开。
206.电解电压为
‑
2.5v,向阴极按照流速10ml/min通混合气(co2与o2比例为1:1),并同时补充流速30ml/min的co2气体。保持发酵反应的温度为30℃,ph为6
‑
7。每隔一个小时抽取样品,利用过氧化氢试剂盒检测过氧化氢浓度,再利用超氧化物歧化酶把氧自由基转换成过氧化氢后,再次测定过氧化氢浓度,利用减差法计算氧自由基浓度。
207.结果如图3所示,电解产生的氧自由基浓度约为0.35mm,过氧化氢浓度约为0.05mm。
208.实施例8:电解过程中超氧化物歧化酶及过氧化氢酶稳定性测定
209.碳固定系统的反应容器设置为阳极室和阴极室的双室:阳极室电极为钛片(3*5cm),阳极室中反应溶液为2mm h2so4,为恢复维持阴极因析出氢气后导致ph升高而不适宜氢氧化细菌生长的ph环境。阴极室电极为镍片(3*5cm),阴极室中反应溶液为包含如下成分的基础培养基:9.0g/l na2hpo4·
12h2o,1.5g/l kh2po4,1.0g/l(nh4)2so4,80mg/l mgso4·
7h2o,1mg/l caso4·
2h2o,0.56mg/l niso4·
7h2o,0.4mg/l ferric citrate,200mg/l nahco3,50mg/l feso4·
7h2o。阳极室与阴极室中间利用nafion 212隔开确保只允许质子通过,参比电极为ag/agcl。
210.用于固碳产单细胞蛋白的钩虫贪铜菌突变菌株(m2菌株)先经异养培养后,转接经自养培养24小时后再次进行转接至阴极室,初始od600为0.15
‑
0.25。添加过氧化氢酶量为180u/ml,超氧化物歧化酶添加量为60u/ml,电解电压为
‑
2.5v,向阴极按照流速10ml/min通混合气(co2与o2比例为1:1),并同时补充流速30ml/min的co2气体。保持发酵反应的温度为30℃,ph为6
‑
7。每隔24h取样测定氧化物歧化酶及过氧化氢酶活性剩余,活性测定方法与实施例1和2中一致。
211.结果如图4所示,氧化物歧化酶及过氧化氢酶活性经1天后剩余80%以下,三天后剩余50%左右。
subtilis.appl.environ.microbiol.78(5):1593
‑
1595.
222.以上,对本公开的实施方式进行了说明。但是,本公开不限定于上述实施方式。凡在本公开的精神和原则之内,所做的任何修改、等同替换、改进等,均应包含在本公开的保护范围之内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。