无热量甜味剂及合成方法
1.本技术为分案申请,原申请的申请日为2015年10月2日,申请号为201580053379.5(pct/us2015/053777),发明名称为“无热量甜味剂及合成方法”。
2.相关申请案的交叉引用
3.本技术要求保护2014年10月3日提交的,标题为“无热量甜味剂及合成方法”的美国临时专利申请第62/059,498号及2014年12月31日提交的,标题为“无热量甜味剂及合成方法”的美国临时专利申请第62/098,929号的优先权,所述专利申请的公开内容据此通过引用整体并入。
4.支持提交序列表的声明
5.本文提供了序列表的纸质复印件和含有命名为“19452382_1.txt”,大小为60,601字节(正如在microsoftexplorer中所测量)的文件的序列表的计算机可读形式并且通过引用并入本文。该序列表由seq id no:1
‑
12组成。
6.公开背景
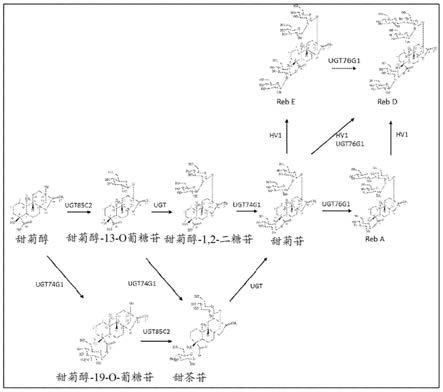
7.本公开总体上涉及天然甜味剂。更具体地,本公开涉及无热量甜味剂及合成无热量甜味剂的方法。
8.甜菊醇糖苷是从甜叶菊(stevia rebaudiana)叶片分离的天然产物。甜菊醇糖苷广泛地用作高强度、低热量的甜味剂并且明显比蔗糖更甜。作为天然甜味剂,不同的甜菊醇葡糖苷具有不同的甜度和后味。甜菊醇糖苷的甜度明显高于蔗糖的甜度。例如,甜菊苷比蔗糖甜100
‑
150倍,具有苦的后味。莱鲍迪苷c(rebaudioside c)比蔗糖甜40
‑
60倍。杜尔可苷a(dulcosidea)比蔗糖甜约30倍。
9.天然存在的甜菊醇糖苷共有相同的基本甜菊醇结构,但是在c13和c19位置的碳水化合物残基(例如,葡萄糖、鼠李糖和木糖残基)的含量上不同。具有已知结构的甜菊醇糖苷包括,甜菊醇、甜菊苷、莱鲍迪苷a、莱鲍迪苷b、莱鲍迪苷c、莱鲍迪苷d、莱鲍迪苷e、莱鲍迪苷f和杜尔可苷a(参见例如,表1)。其它甜菊醇糖苷为莱鲍迪苷m、莱鲍迪苷n和莱鲍迪苷o。
10.表1.甜菊醇糖苷。
11.12.13.[0014][0015]
按干重计,甜菊苷、莱鲍迪苷a、莱鲍迪苷c和杜尔可苷a分别占叶中甜菊醇糖苷总重量的9.1、3.8、0.6和0.3%,而其它甜菊醇糖苷以低得多的量存在。来自甜叶菊植株的提取物可商购,通常含有作为主要化合物的甜菊苷和莱鲍迪苷a。其它甜菊醇糖苷在甜叶菊提取物中作为微量组分存在。例如,商用制剂中莱鲍迪苷a的量可以在总甜菊醇糖苷含量的约20%至大于90%变化,而莱鲍迪苷b的量可为总甜菊醇糖苷的约1
‑
2%,莱鲍迪苷c的量可为
总甜菊醇糖苷的约7
‑
15%,且莱鲍迪苷d的量可为总甜菊醇糖苷的约2%。
[0016]
大多数甜菊醇糖苷是使用尿苷5
’‑
二磷酸葡萄糖(udp
‑
葡萄糖)作为糖部分的供体,通过通常由udp
‑
糖基转移酶(ugt)催化的甜菊醇的几个糖基化反应形成的。植物中的ugt构成了酶的非常不同的类,所述酶将葡萄糖残基从udp
‑
葡萄糖转移到甜菊醇。例如,甜菊苷的c
‑
13
‑
o
‑
葡萄糖的c
‑3’
的糖基化产生莱鲍迪苷a;且甜菊苷的19
‑
o
‑
葡萄糖的c
‑2’
的糖基化产生莱鲍迪苷e。进一步地莱鲍迪苷a(在c
‑2’‑
19
‑
o
‑
葡萄糖处)或莱鲍迪苷e(在c
‑3’‑
13
‑
o
‑
葡萄糖处)的糖基化生成莱鲍迪苷d。(图1)。
[0017]
替代性甜味剂正受到越来越多的关注,因为意识到许多疾病与高糖食品和饮料的消耗相关。虽然人造甜味剂可用,但许多人造甜味剂如甘素(dulcin)、环己氨基磺酸钠和糖精由于对其安全性的考虑已被一些国家禁止或限制。因此,天然来源的无热量甜味剂变得越来越受欢迎。广泛使用甜叶菊甜味剂的主要障碍之一是其不良的口味属性。因此,需要开发替代性甜味剂及其生产方法以提供甜味潜能和风味性质的最佳组合。
[0018]
发明概述
[0019]
本公开总体上涉及天然甜味剂。更具体地,本公开涉及无热量甜味剂和合成无热量甜味剂的方法。
[0020]
合成莱鲍迪苷v。一方面,本公开涉及由以下化学结构组成的合成莱鲍迪苷(莱鲍迪苷v):
[0021][0022]
合成莱鲍迪苷w。一方面,本公开涉及由以下化学结构组成的合成莱鲍迪苷(莱鲍
迪苷w):
[0023][0024]
由莱鲍迪苷g生成莱鲍迪苷v的方法。另一方面,本公开涉及一种由莱鲍迪苷g合成莱鲍迪苷v的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷g,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,和hv1 udp
‑
糖基转移酶,有或无蔗糖合酶(sus);以及温育所述反应混合物足够时间以生成莱鲍迪苷v,其中葡萄糖共价偶联至莱鲍迪苷g以生成莱鲍迪苷v。
[0025]
由莱鲍迪苷g生成莱鲍迪苷v的方法。另一方面,本公开涉及一种由莱鲍迪苷g合成莱鲍迪苷v的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷g,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自尿苷二磷酸糖基转移酶(eugt11)、udp
‑
糖基转移酶
‑
蔗糖合酶(sus)融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶),有或无蔗糖合酶(sus);以及温育所述反应混合物足够时间以生成莱鲍迪苷v,其中葡萄糖共价偶联至莱鲍迪苷g以生成莱鲍迪苷v。
[0026]
由莱鲍迪苷ka生成莱鲍迪苷v的方法。另一方面,本公开涉及一种由莱鲍迪苷ka合成莱鲍迪苷v的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷ka,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自udp
‑
糖基转移酶(ugt76g1;seq id no:1)和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶),有或无蔗糖合酶(sus);以及温育所述反应混合物足够时间以生成莱鲍迪苷v,其中葡萄糖共价偶联至莱鲍迪苷ka以生成莱鲍迪苷v。
[0027]
由甜茶苷生成莱鲍迪苷v的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪
苷v的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自udp
‑
糖基转移酶(ugt76g1)、hv1和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶),有或无蔗糖合酶(sus);以及温育所述反应混合物足够时间以生成莱鲍迪苷v,其中葡萄糖共价偶联至甜茶苷以生成莱鲍迪苷ka。连续地,葡萄糖共价偶联至莱鲍迪苷ka以生成莱鲍迪苷v。葡萄糖共价偶联至甜茶苷以生成莱鲍迪苷g。连续地,葡萄糖共价偶联至莱鲍迪苷g以生成莱鲍迪苷v。
[0028]
由甜茶苷生成莱鲍迪苷v的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷v的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自udp
‑
糖基转移酶(ugt76g1)、eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶),有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷v,其中葡萄糖共价偶联至甜茶苷以生成莱鲍迪苷ka并且葡萄糖共价偶联至莱鲍迪苷ka以生成莱鲍迪苷v。葡萄糖共价偶联至甜茶苷以生成莱鲍迪苷g并且葡萄糖共价偶联至莱鲍迪苷g以生成莱鲍迪苷v。
[0029]
由莱鲍迪苷v生成莱鲍迪苷w的方法。另一方面,本公开涉及一种由莱鲍迪苷v合成莱鲍迪苷w的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷v,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自udp
‑
糖基转移酶(ugt76g1)和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶),有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷w,其中葡萄糖共价偶联至莱鲍迪苷v以生成莱鲍迪苷w。
[0030]
由莱鲍迪苷g生成莱鲍迪苷w的方法。另一方面,本公开涉及一种由莱鲍迪苷g合成莱鲍迪苷w的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷g,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自udp
‑
糖基转移酶(ugt76g1)、udp
‑
糖基转移酶
‑
蔗糖合酶融合酶和hv1的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶);有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷w,其中葡萄糖通过hv1共价偶联至莱鲍迪苷g以生成莱鲍迪苷v。连续地,葡萄糖通过ugt76g1共价偶联至莱鲍迪苷v以生成莱鲍迪苷w。
[0031]
由莱鲍迪苷g生成莱鲍迪苷w的方法。另一方面,本公开涉及一种由莱鲍迪苷g合成莱鲍迪苷w的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷g,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自ugt76g1、eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶);以及温育所述反应混合物足够时间以生成莱鲍迪苷w,其中葡萄糖通过eugt11共价偶联至莱鲍迪苷g以生成莱鲍迪苷v。连续地,葡萄糖通过ugt76g1共价偶联至莱鲍迪苷v以生成莱鲍迪苷w。
[0032]
由莱鲍迪苷ka生成莱鲍迪苷w的方法。另一方面,本公开涉及一种由莱鲍迪苷ka合成莱鲍迪苷w的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷ka;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自udp
‑
糖基转移酶(ugt76g1)和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶),有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷w,其中葡萄糖共
价偶联至莱鲍迪苷ka以生成莱鲍迪苷v。连续地,葡萄糖共价偶联至莱鲍迪苷v以生成莱鲍迪苷w。
[0033]
由甜茶苷生成莱鲍迪苷w的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷d的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自ugt76g1、hv1和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶),有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷w。
[0034]
由甜茶苷生成莱鲍迪苷w的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷w的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自ugt76g1、eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶),有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷w。
[0035]
由甜茶苷生成甜茶苷和莱鲍迪苷ka的混合物的方法。另一方面,本公开涉及一种由甜茶苷合成甜菊苷和莱鲍迪苷ka的混合物的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成甜菊苷和莱鲍迪苷ka的混合物,其中葡萄糖共价偶联至甜茶苷的c2
’‑
19
‑
o
‑
葡萄糖以生成莱鲍迪苷ka;葡萄糖共价偶联至甜茶苷的c2
’‑
13
‑
o
‑
葡萄糖以生成甜菊苷。
[0036]
由甜茶苷生成莱鲍迪苷ka的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷ka的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物和hv1,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷ka,其中葡萄糖共价偶联至甜茶苷的c2
’‑
19
‑
o
‑
葡萄糖以生成莱鲍迪苷ka。
[0037]
由甜茶苷生成莱鲍迪苷g的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷g的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自ugt76g1和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷g,其中葡萄糖共价偶联至甜茶苷的c3
’‑
13
‑
o
‑
葡萄糖以生成莱鲍迪苷g。
[0038]
由莱鲍迪苷ka生成莱鲍迪苷e的方法。另一方面,本公开涉及一种由莱鲍迪苷ka合成莱鲍迪苷e的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷ka,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物和hv1 udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷e,其中葡萄糖共价偶联至莱鲍迪苷ka的c2’13
‑
o
‑
葡萄糖以生成莱鲍迪苷e。
[0039]
由莱鲍迪苷ka生成莱鲍迪苷e的方法。另一方面,本公开涉及一种由莱鲍迪苷ka合成莱鲍迪苷e的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷ka,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷e,其中葡萄糖共价偶联至莱鲍迪苷ka的c2’13
‑
o
‑
葡萄糖以生成
莱鲍迪苷e。
[0040]
由甜茶苷生成莱鲍迪苷e的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷e的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,和选自eugt11和udp
‑
糖基转移酶
‑
蔗糖合成融合酶的udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷e,其中葡萄糖共价偶联至甜茶苷以生成莱鲍迪苷ka和甜菊苷的混合物。连续地,葡萄糖共价偶联至莱鲍迪苷ka和甜菊苷以生成莱鲍迪苷e。
[0041]
由甜茶苷生成莱鲍迪苷e的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷e的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物和hv1 udp
‑
糖基转移酶,有或无蔗糖合酶;温育所述反应混合物足够时间以生成莱鲍迪苷e,其中葡萄糖共价偶联至甜茶苷以生成莱鲍迪苷ka;以及与hv1一起进一步温育莱鲍迪苷ka以生成莱鲍迪苷e。
[0042]
由甜茶苷生成莱鲍迪苷d3的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷d3的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的udp
‑
糖基转移酶,有或无蔗糖合酶;温育所述反应混合物足够时间以生成甜菊苷和莱鲍迪苷d3的混合物,其中葡萄糖共价偶联至甜茶苷以生成甜菊苷和莱鲍迪苷ka的混合物;与eugt11一起进一步温育甜菊苷和莱鲍迪苷ka的混合物以生成莱鲍迪苷e,其中葡萄糖共价偶联至甜菊苷和莱鲍迪苷ka以生成莱鲍迪苷e;以及与eugt11一起进一步温育莱鲍迪苷e以生成莱鲍迪苷d3,其中葡萄糖共价偶联至莱鲍迪苷e以生成莱鲍迪苷d3。
[0043]
由莱鲍迪苷ka生成莱鲍迪苷d3的方法。另一方面,本公开涉及一种由莱鲍迪苷ka合成莱鲍迪苷d3的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷ka,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的udp
‑
糖基转移酶,有或无蔗糖合酶;温育所述反应混合物足够时间以生成莱鲍迪苷d3,其中葡萄糖共价偶联至莱鲍迪苷ka以生成莱鲍迪苷e;以及进一步温育莱鲍迪苷e与eugt11的混合物以生成莱鲍迪苷d3,其中葡萄糖共价偶联至莱鲍迪苷e以生成莱鲍迪苷d3。
[0044]
由莱鲍迪苷e生成莱鲍迪苷z的方法。另一方面,本公开涉及一种由莱鲍迪苷e合成莱鲍迪苷z的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷e,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,hv1和蔗糖合酶;温育所述反应混合物足够时间以生成莱鲍迪苷z,其中葡萄糖共价偶联至莱鲍迪苷e的c2
’‑
13
‑
o
‑
葡萄糖以生成莱鲍迪苷z1。葡萄糖共价偶联至莱鲍迪苷e的c2
’‑
19
‑
o
‑
葡萄糖以生成莱鲍迪苷z2。
[0045]
由莱鲍迪苷d生成莱鲍迪苷m的方法。另一方面,本公开涉及一种由莱鲍迪苷d合成莱鲍迪苷m的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷d,选自蔗糖、尿苷二磷酸(udp)、尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)及其组合的底物,及选自ugt76g1、udp
‑
糖基转移酶
‑
蔗糖合酶融合酶及其组合的udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷m,其中葡萄糖共价偶联至莱鲍迪苷d以生成莱鲍迪苷m。
[0046]
由甜菊苷生成莱鲍迪苷d和莱鲍迪苷m的方法。另一方面,本公开涉及一种由甜菊苷合成莱鲍迪苷d和莱鲍迪苷m的方法。该方法包括制备反应混合物,所述反应混合物包含甜菊苷,选自蔗糖、尿苷二磷酸(udp)、尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)及其组合的底物,及选自hv1、ugt76g1、udp
‑
糖基转移酶
‑
蔗糖合酶融合酶及其组合的udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷m。在某些实施方案中,葡萄糖共价偶联至甜菊苷以生成莱鲍迪苷a和/或莱鲍迪苷e。连续地,葡萄糖共价偶联至莱鲍迪苷a和/或莱鲍迪苷e以生成莱鲍迪苷d,并且葡萄糖共价偶联至莱鲍迪苷d以生成莱鲍迪苷m。
[0047]
由莱鲍迪苷a生成莱鲍迪苷d和莱鲍迪苷m的方法。另一方面,本公开涉及一种由莱鲍迪苷a合成莱鲍迪苷d和莱鲍迪苷m的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷a,选自蔗糖、尿苷二磷酸(udp)、尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)及其组合的底物,及选自hv1、ugt76g1、udp
‑
糖基转移酶
‑
蔗糖合酶融合酶及其组合的udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷m,其中葡萄糖共价偶联至莱鲍迪苷a以生成莱鲍迪苷d,并且葡萄糖共价偶联至莱鲍迪苷d以生成莱鲍迪苷m。
[0048]
由莱鲍迪苷e生成莱鲍迪苷d和莱鲍迪苷m的方法。另一方面,本公开涉及一种由莱鲍迪苷e合成莱鲍迪苷d和莱鲍迪苷m的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷e,选自蔗糖、尿苷二磷酸(udp)、尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)及其组合的底物,及选自ugt76g1、udp
‑
糖基转移酶
‑
蔗糖合酶融合酶及其组合的udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷m,其中葡萄糖共价偶联至莱鲍迪苷e以生成莱鲍迪苷d,并且其中葡萄糖共价偶联至莱鲍迪苷d以生成莱鲍迪苷m。
[0049]
另一方面,本公开涉及一种可口服消耗产品,其包含增甜量的选自莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷g、莱鲍迪苷ka、莱鲍迪苷m及其组合的莱鲍迪苷,其中可口服消耗产品选自饮料产品和可消耗产品。
[0050]
另一方面,本公开涉及一种饮料产品,其包含增甜量的选自莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷g、莱鲍迪苷ka、莱鲍迪苷m及其组合的莱鲍迪苷。莱鲍迪苷以约5ppm至约100ppm的浓度存在于饮料产品中。在一些实施方案中,低浓度的莱鲍迪苷,例如低于100ppm,具有与浓度介于10,000和30,000ppm之间的蔗糖溶液相当的甜度。
[0051]
另一方面,本公开涉及一种可消耗产品,其包含增甜量的选自莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷g、莱鲍迪苷ka、莱鲍迪苷m及其组合的莱鲍迪苷。莱鲍迪苷以约5ppm至约100ppm的浓度存在于可消耗产品中。在一些实施方案中,低浓度的莱鲍迪苷,例如低于100ppm,具有与浓度介于10,000和30,000ppm之间的蔗糖溶液相当的甜度。
[0052]
另一方面,本公开涉及一种由以下化学结构组成的甜味剂:
[0053][0054]
另一方面,本公开涉及一种由以下化学结构组成的甜味剂:
[0055][0056]
在可以与任何前述实施方案组合的某些实施方案中,莱鲍迪苷v或莱鲍迪苷w或莱鲍迪苷g或莱鲍迪苷ka或莱鲍迪苷m可以是唯一的甜味剂,并且所述产品具有相当于约1%至约4%(w/v
‑
%)蔗糖溶液的甜味强度。在可以与任何前述实施方案组合的某些实施方案中,可口服消耗产品还可包括附加甜味剂,其中所述产品具有相当于约1%至约10%(w/v
‑
%)蔗糖溶液的甜味强度。在可以与任何前述实施方案组合的某些实施方案中,产品中的每种增甜成分均可为高强度甜味剂。在可以与任何前述实施方案组合的某些实施方案中,产品中的每种增甜成分均可为天然高强度甜味剂。在可以与任何前述实施方案组合的某些实施方案中,附加甜味剂可为选自以下的一种或多种甜味剂:甜叶菊提取物、甜菊醇糖苷、甜菊苷、莱鲍迪苷a、莱鲍迪苷b、莱鲍迪苷c、莱鲍迪苷d、莱鲍迪苷d3、莱鲍迪苷e、莱鲍迪苷f、莱鲍迪苷g、莱鲍迪苷ka、莱鲍迪苷m、杜尔可苷a、甜茶苷、甜菊醇二糖苷、蔗糖、高果糖玉米糖浆、果糖、葡萄糖、木糖、阿拉伯糖、鼠李糖、赤藓糖醇、木糖醇、甘露糖醇、山梨糖醇、肌醇、acek、阿斯巴甜(aspartame)、纽甜(neotame)、三氯蔗糖(sucralose)、糖精、柚皮苷二氢查耳酮(nardhc)、新橙皮苷二氢查耳酮(ndhc)、甜茶苷、罗汉果苷iv(mogroside iv)、赛门苷i(siamenoside i)、罗汉果苷v、莫那甜(monatin)、索马甜(thaumatin)、莫奈林(monellin)、布拉奇因(brazzein)、l
‑
丙氨酸、甘氨酸、罗汉果、赫南德辛(hernandulcin)、叶甜素(phyllodulcin)、三叶苷(trilobtain)及其组合。在可以与任何前述实施方案组合的某些实施方案中,饮料产品和可消耗产品还可包括一种或多种选自以下的添加剂:碳水化合物、多元醇、氨基酸或其盐、聚氨基酸或其盐、糖酸或其盐、核苷酸、有机酸、无机酸、有机盐、有机酸式盐、有机碱式盐、无机盐、苦味化合物、食用香料、调味成分、涩味化合物、蛋白质、蛋白质水解物、表面活性剂、乳化剂、类黄酮、醇、聚合物及其组合。在可以与任何前述
实施方案组合的某些实施方案中,莱鲍迪苷v在将其添加到产品中之前具有按重量计约50%至约100%的纯度。在可以与任何前述实施方案组合的某些实施方案中,w在将其添加到产品中之前具有按重量计约50%至约100%的纯度。在可以与任何前述实施方案组合的某些实施方案中,产品中的莱鲍迪苷v为莱鲍迪苷v多晶型物或非晶形莱鲍迪苷v。在可以与任何前述实施方案组合的某些实施方案中,产品中的莱鲍迪苷v为莱鲍迪苷v立体异构体。在可以与任何前述实施方案组合的某些实施方案中,产品中的莱鲍迪苷w为莱鲍迪苷w多晶型物或非晶形莱鲍迪苷w。在可以与任何前述实施方案组合的某些实施方案中,产品中的莱鲍迪苷w为莱鲍迪苷w立体异构体。
[0057]
本公开的其它方面涉及一种制备饮料产品和可消耗产品的方法,所述方法通过将选自莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m和莱鲍迪苷g的合成莱鲍迪苷包括到产品或用于制备饮料产品和可消耗产品的成分中进行,其中选自莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m和莱鲍迪苷g的莱鲍迪苷以约5ppm至约100ppm的浓度存在于产品中。本公开的其它方面涉及一种提高饮料产品和可消耗产品甜度的方法,所述方法通过将约5ppm至约100ppm的选自莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m和莱鲍迪苷g的合成莱鲍迪苷添加到饮料产品和可消耗产品中进行,其中与缺少选自莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m和莱鲍迪苷g的合成莱鲍迪苷的相应饮料产品和可消耗产品相比,添加的选自莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m和莱鲍迪苷g的合成莱鲍迪苷提高了饮料产品和可消耗产品的甜度。
[0058]
在可以与任何前述实施方案组合的某些实施方案中,莱鲍迪苷v是唯一的甜味剂,并且产品具有相当于约1%至约4%(w/v
‑
%)蔗糖溶液的甜味强度。在可以与任何前述实施方案组合的某些实施方案中,莱鲍迪苷ka是唯一的甜味剂,并且产品具有相当于约1%至约4%(w/v
‑
%)蔗糖溶液的甜味强度。在可以与任何前述实施方案组合的某些实施方案中,莱鲍迪苷g是唯一的甜味剂,并且产品具有相当于约1%至约4%(w/v
‑
%)蔗糖溶液的甜味强度。在可以与任何前述实施方案组合的某些实施方案中,莱鲍迪苷w是唯一的甜味剂,并且产品具有相当于约1%至约4%(w/v
‑
%)蔗糖溶液的甜味强度。在可以与任何前述实施方案组合的某些实施方案中,莱鲍迪苷m是唯一的甜味剂,并且产品具有相当于约1%至约4%(w/v
‑
%)蔗糖溶液的甜味强度。在可以与任何前述实施方案组合的某些实施方案中,所述方法还包括添加附加甜味剂,其中产品具有相当于约1%至约10%(w/v
‑
%)蔗糖溶液的甜味强度。
[0059]
本公开的其它方面涉及一种制备增甜饮料产品或增甜可消耗产品的方法,所述方法通过以下步骤进行:a)提供含有一种或多种甜味剂的饮料产品或可消耗产品;以及b)向饮料产品或可消耗产品中添加约5ppm至约100ppm的选自莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m和莱鲍迪苷g及其组合的合成莱鲍迪苷。
[0060]
在可以与任何前述实施方案组合的某些实施方案中,所述方法还包括向饮料产品或可消耗产品中添加一种或多种添加剂。在可以与任何前述实施方案组合的某些实施方案中,可口服消耗产品还含有一种或多种添加剂。在可以与任何前述实施方案组合的某些实施方案中,所述一种或多种添加剂选自碳水化合物、多元醇、氨基酸或其盐、聚氨基酸或其盐、糖酸或其盐、核苷酸、有机酸、无机酸、有机盐、有机酸式盐、有机碱式盐、无机盐、苦味化合物、食用香料、调味成分、涩味化合物、蛋白质、蛋白质水解物、表面活性剂、乳化剂、类黄
酮、醇、聚合物及其组合。在可以与任何前述实施方案组合的某些实施方案中,产品中的每种增甜成分均可为高强度甜味剂。在可以与任何前述实施方案组合的某些实施方案中,产品中的每种增甜成分均可为天然高强度甜味剂。在可以与任何前述实施方案组合的某些实施方案中,甜味剂选自甜叶菊提取物、甜菊醇糖苷、甜菊苷、莱鲍迪苷a、莱鲍迪苷b、莱鲍迪苷c、莱鲍迪苷d、莱鲍迪苷d3、莱鲍迪苷e、莱鲍迪苷f、莱鲍迪苷g、莱鲍迪苷ka、莱鲍迪苷m、杜尔可苷a、甜茶苷、甜菊醇二糖苷、蔗糖、高果糖玉米糖浆、果糖、葡萄糖、木糖、阿拉伯糖、鼠李糖、赤藓糖醇、木糖醇、甘露糖醇、山梨糖醇、肌醇、acek、阿斯巴甜、纽甜、三氯蔗糖、糖精、柚皮苷二氢查耳酮(nardhc)、新橙皮苷二氢查耳酮(ndhc)、甜茶苷、罗汉果苷iv、赛门苷i、罗汉果苷v、莫那甜、索马甜、莫奈林、布拉奇因、l
‑
丙氨酸、甘氨酸、罗汉果、赫南德辛、叶甜素、三叶苷及其组合。在可以与任何前述实施方案组合的某些实施方案中,莱鲍迪苷v在将其添加到产品中之前具有按重量计约50%至约100%的纯度。在可以与任何前述实施方案组合的某些实施方案中,产品中的莱鲍迪苷v为莱鲍迪苷v多晶型物或非晶形莱鲍迪苷v。在可以与任何前述实施方案组合的某些实施方案中,莱鲍迪苷w在将其添加到产品中之前具有按重量计约50%至约100%的纯度。在可以与任何前述实施方案组合的某些实施方案中,产品中的莱鲍迪苷w为莱鲍迪苷w多晶型物或非晶形莱鲍迪苷w。
[0061]
附图简述
[0062]
本公开在考虑到以下对其的详述时将更好理解,并且优于以上所提到的那些的特征、方面和优势将变得显而易见。此详述参考以下附图,其中:
[0063]
图1描绘自甜菊醇开始的甜菊醇糖苷生物合成途径。
[0064]
图2描绘用箭头指示的sds
‑
page分析纯化的重组蛋白:a:hv1,b:ugt76g1,c:eugt11,d:atsus1,e:ugt76g1
‑
sus1(gs),f:eugt11
‑
sus1(eus)。
[0065]
图3描绘由甜茶苷生成莱鲍迪苷ka(“reb ka”)和莱鲍迪苷e(“reb e”)的hv1催化反应。a
‑
c:示出了甜茶苷(“rub”)、甜菊苷(“ste”)和莱鲍迪苷e(“reb e”)标准品的hplc保留时间。在6小时(d)、12小时(f)和24小时(h)通过单独的hv1酶促生成reb ka;在6小时(e)、12小时(g)和24小时(i)通过ugt
‑
sus(hv1
‑
atsus1)偶联体系酶促生成reb ka和reb e。
[0066]
图4描绘reb e通过hv1向莱鲍迪苷z的转化。(a):示出了莱鲍迪苷e(“reb e”)的hplc保留时间。在3小时(b)、7小时(c)、24小时(d)和44小时(e)在hv1
‑
atsus1偶联体系中通过hv1酶促生成莱鲍迪苷z(“reb z”)。
[0067]
图5描绘reb ka通过hv1向reb e的转化。(a
‑
b):示出了莱鲍迪苷ka(“reb ka”)和莱鲍迪苷e(“reb e”)标准品的hplc保留时间。在12小时(c)通过单独的hv1酶促生成reb e;在12小时(d)通过ugt
‑
sus(hv1
‑
atsus1)偶联体系酶促生成reb e。
[0068]
图6描绘由甜茶苷生成reb ka和甜菊苷的eugt11催化反应。(a
‑
f):示出了甜茶苷(“rub”)、甜菊苷(“ste”)、莱鲍迪苷g(“reb g”)、莱鲍迪苷e(“reb e”)、莱鲍迪苷d(“reb d”)和莱鲍迪苷d3(“reb d3”)标准品的hplc保留时间。在12小时(g)和48小时(j)通过单独的eugt11的酶促反应;在12小时(h)和48小时(k)通过ugt
‑
sus(eugt11
‑
atsus1)偶联体系的酶促反应;在12小时(i)和48小时(l)通过eus融合蛋白的酶促反应。
[0069]
图7描绘reb ka通过eugt11和eus融合蛋白向reb e和reb d3的转化。(a
‑
c):示出了莱鲍迪苷ka(“reb ka”)、莱鲍迪苷e(“reb e”)和莱鲍迪苷d3(“reb d3”)标准品的hplc保留时间。在12小时(d)和48小时(g)通过单独的eugt11的酶促反应;在12小时(e)和48小时
(h)通过ugt
‑
sus(eugt11
‑
atsus1)偶联体系的酶促反应;在12小时(f)和48小时(i)通过eus融合蛋白的酶促反应。
[0070]
图8描绘体外莱鲍迪苷g的ugt76g1生成。(a
‑
b):示出了甜茶苷(“rub”)和莱鲍迪苷g(“reb g”)标准品的hplc保留时间。在12小时(c)和24小时(f)通过单独的ugt76g1的酶促反应;在12小时(d)和24小时(g)通过ugt
‑
sus(eugt11
‑
atsus1)偶联体系的酶促反应;在12小时(e)和48小时(h)通过gs融合蛋白的酶促反应。
[0071]
图9描绘由莱鲍迪苷ka生成甜菊醇糖苷reb v和reb w的ugt76g1催化反应。(a
‑
d):示出了甜茶苷(“rub”)、莱鲍迪苷d(“reb d”)、莱鲍迪苷e(“reb e”)和莱鲍迪苷ka(“reb ka”)标准品的hplc保留时间。在6小时(e)和12小时(h)通过单独的ugt76g1的酶促反应;在6小时(f)和12小时(i)通过ugt
‑
sus(ugt76g1
‑
atsus1)偶联体系的酶促反应;在6小时(g)和12小时(j)通过gs融合蛋白的酶促反应。
[0072]
图10描绘体外reb v向reb w的ugt76g1转化。(a
‑
b):示出了reb v和reb w的hplc保留时间。(c):在6小时通过ugt76g1
‑
atsus1偶联体系的酶促反应。
[0073]
图11描绘reb g向reb v的hv1转化。(a
‑
c):示出了莱鲍迪苷g(“reb g”)、莱鲍迪苷a(“reba”)和莱鲍迪苷e(“reb e”)标准品的hplc保留时间。在12小时(d)和24小时(f)通过单独的hv1的酶促反应;在12小时(e)和24小时(g)通过ugt
‑
sus(hv1
‑
atsus1)偶联体系的酶促反应。
[0074]
图12描绘reb g向reb v的eugt11转化。(a
‑
d):示出了莱鲍迪苷g(“reb g”)、莱鲍迪苷a(“reba”)、莱鲍迪苷e(“reb e”)和莱鲍迪苷d(“reb d”)标准品的hplc保留时间。在12小时(e)和24小时(h)通过单独的eugt11的酶促反应;在12小时(f)和24小时(i)通过ugt
‑
sus(eugt11
‑
atsus1)偶联体系的酶促反应;在12小时(g)和24小时(j)通过eus融合酶的酶促反应。
[0075]
图13描绘通过重组hv1多肽、重组ugt76g1、gs融合酶和重组atsus1的组合催化由甜茶苷体外生成reb w。(a
‑
f):示出了甜茶苷(“rub”)、甜菊苷(“ste”)、莱鲍迪苷g(“reb g”)、莱鲍迪苷a(“reb a”)、莱鲍迪苷d(“reb d”)和莱鲍迪苷e(“reb e”)的标准品。在6小时(g)、12小时(i)和24小时(k)通过hv1、ugt76g1和atsus1酶促生成reb w;在6小时(h)、12小时(j)和24小时(l)通过hv1和gs融合蛋白酶促生成reb w。
[0076]
图14描绘通过重组eugt11多肽、重组ugt76g1、gs融合酶和重组atsus1的组合催化由甜茶苷体外生成reb w。(a
‑
e):示出了甜茶苷(“rub”)、甜菊苷(“ste”)、莱鲍迪苷g(“reb g”)、莱鲍迪苷e(“reb e”)和莱鲍迪苷d(“reb d”)的标准品。在12小时(f)和48小时(h)通过eugt11、ugt76g1和atsus1酶促生成reb w;在12小时(g)和48小时(i)通过eugt11和gs融合蛋白酶促生成reb w。
[0077]
图15描绘通过重组hv1多肽、重组ugt76g1、gs融合酶和重组atsus1的组合催化由reb g体外生成reb w。a
‑
d示出了莱鲍迪苷g(“reb g”)、莱鲍迪苷a(“reb a”)、莱鲍迪苷d(“reb d”)、莱鲍迪苷和莱鲍迪苷e(“reb e”)的标准品。在6小时(e)、12小时(g)和36小时(i)通过hv1、ugt76g1和atsus1酶促生成reb v和reb w;在6小时(f)、12小时(h)和36小时(j)通过hv1和gs融合蛋白酶促生成reb v和reb w。
[0078]
图16描绘通过重组eugt11多肽、重组ugt76g1、gs融合酶和重组atsus1的组合催化由reb g体外生成reb w。(a
‑
d):示出了莱鲍迪苷g(“reb g”)、莱鲍迪苷a(“reba”)、莱鲍迪
苷e(“reb e”)和莱鲍迪苷d(“reb d”)的标准品。在12小时(e)和48小时(g)通过eugt11、ugt76g1和atsus1酶促生成reb w;在12小时(f)和48小时(h)通过eugt11和gs融合蛋白酶促生成reb w。
[0079]
图17描绘reb v和reb g的结构。
[0080]
图18描绘reb v的关键tocsy和hmbc相关性。
[0081]
图19描绘reb w和reb v的结构。
[0082]
图20描绘reb w的关键tocsy和hmbc相关性。
[0083]
图21描绘甜菊醇糖苷的生物合成途径。
[0084]
图22描绘通过ugt76g1和gs融合酶催化由reb d体外生成reb m。(a
‑
b):示出了莱鲍迪苷d(“reb d”)和莱鲍迪苷m(“reb m”)标准品的hplc保留时间。在3小时(c)和6小时(f)通过单独的ugt76g1的酶促反应;在3小时(d)和6小时(g)通过ugt
‑
sus(ugt76g1
‑
atsus1)偶联体系的酶促反应;在3小时(e)和6小时(h)通过gs融合酶的酶促反应。
[0085]
图23描绘通过ugt76g1和gs融合酶催化由reb e体外生成reb d和reb m。(a
‑
c):示出了莱鲍迪苷e(“reb e”)、莱鲍迪苷d(“reb d”)和莱鲍迪苷m(“reb m”)标准品的hplc保留时间。在3小时(d)、12小时(g)和24小时(j)通过单独的ugt76g1的酶促反应;在3小时(e)、12小时(h)和24小时(k)通过ugt
‑
sus(ugt76g1
‑
atsus1)偶联体系的酶促反应;在3小时(f)、12小时(i)和24小时(l)通过gs融合酶的酶促反应。
[0086]
图24描绘通过重组hv1、重组ugt76g1、gs融合酶和/或重组atsus1的组合催化由甜菊苷体外生成reb d和reb m。(a
‑
d):示出了甜菊苷(“ste”)、莱鲍迪苷a(“reba”)、莱鲍迪苷d(“reb d”)和莱鲍迪苷m(“reb m”)标准品的hplc保留时间。在6小时(e)、12小时(h)和24小时(k)在ugt
‑
sus偶联体系中通过hv1和ugt76g1的酶促反应;在6小时(f)、12小时(i)和24小时(l)通过hv1和gs融合酶的酶促反应;在6小时(g)、12小时(j)和24小时(m)通过ugt76g1和hv1的酶促反应.
[0087]
图25描绘通过重组hv1、重组ugt76g1、gs融合酶和/或重组atsus1催化由莱鲍迪苷a体外生成reb d和reb m。(a
‑
c):示出了莱鲍迪苷a(“reba”)、莱鲍迪苷d(“reb d”)和莱鲍迪苷m(“reb m”)标准品的hplc保留时间。在6小时(d)、12小时(g)和24小时(j)在ugt
‑
sus偶联体系中通过hv1和ugt76g1的酶促反应;在6小时(e)、12小时(h)和24小时(k)通过hv1和gs融合酶的酶促反应。在6小时(f)、12小时(i)和24小时(j)通过ugt76g1和hv1的酶促反应。
[0088]
图26描绘reb m的结构。
[0089]
图27描绘reb m的关键tocsy和hmbc相关性。
[0090]
虽然本公开易受各种修改和替代性形式影响,但其具体实施方案已经在附图中以举例的方式示出并且本文在下面有详细描述。然而,应理解对具体实施方案的描述并非旨在限制本公开而包括属于所附权利要求书所定义的本公开的精神和范围内的所有修改、等效方案和替代方案。
[0091]
详述
[0092]
除非另有定义,否则本文使用的所有技术和科学术语具有本公开所属领域中普通技术人员通常所理解的相同含义。虽然在本公开的实践或试验中可使用与本文描述的那些类似或等效的任何方法和材料,但下面描述的是优选材料和方法。
[0093]
术语“互补”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不
限于用于描述能够相互杂交的核苷酸碱基之间的关系。例如,就dna而言,腺苷与胸腺嘧啶互补且胞嘧啶与鸟嘌呤互补。因此,主题技术还包括与所附序列表中所报告的完整序列互补的分离的核酸片段以及那些基本上相似的核酸序列。
[0094]
术语“核酸”和“核苷酸”根据本领域中普通技术人员所理解的其各自的普通和惯常含义使用,并且不限于用于指呈单链或双链形式的脱氧核糖核苷酸或核糖核苷酸及其聚合物。除非特别限制,该术语涵盖含有天然核苷酸的已知类似物,具有与参考核酸相似的结合性质并且以类似于天然存在的核苷酸的方式代谢的核酸。除非另有说明,否则特定核酸序列还涵盖其经保守性修饰的或简并性变体(例如简并密码子取代)和互补序列,以及明确指示的序列。
[0095]
术语“分离的”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且用于分离的核酸或分离的多肽的上下文中时,不限于用于指经人工,远离其天然环境存在且因此不是天然产物的核酸或多肽。分离的核酸或多肽可以呈纯化形式存在或者可以存在于非天然环境中,例如转基因宿主细胞中。
[0096]
如本文中所用的术语“温育”是指混合两种或更多种化学或生物实体(如化学化合物和酶)并允许其在利于生成甜菊醇糖苷组合物的条件下相互作用的过程。
[0097]
术语“简并性变体”是指具有与参考核酸序列的不同之处在于一个或多个简并密码子取代的残基序列的核酸序列。简并密码子取代可通过产生其中一个或多个选定(或所有)密码子的第三位被混合碱基和/或脱氧肌苷残基取代的序列来实现。核酸序列及其所有简并性变体将表达相同的氨基酸或多肽。
[0098]
术语“多肽”、“蛋白质”和“肽”根据本领域中普通技术人员所理解的其各自的普通和惯常含义使用;所述三个术语有时可互换使用,并且不限于用于指氨基酸聚合物或氨基酸类似物,而不管其大小或功能如何。虽然“蛋白质”常常在提到相对较大的多肽时使用,而“肽”常常在提到较小多肽时使用,但这些术语在本领域中的使用有重叠和变化。除非另外指出,否则如本文中所用的术语“多肽”是指肽、多肽和蛋白质。术语“蛋白质”、“多肽”和“肽”在提到多核苷酸产物时可互换使用。因此,示例性多肽包括多核苷酸产物、天然存在的蛋白质、同源物、直系同源物、旁系同源物、片段和其它等效物、前述物质的变体和类似物。
[0099]
术语“多肽片段”和“片段”在提到参考多肽使用时,根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指与参考多肽本身相比其中有氨基酸残基缺失,但是其中剩余的氨基酸序列通常与参考多肽中的相应位置相同的多肽。此类缺失可以出现在参考多肽的氨基末端或羧基末端,或可选地出现在两处。
[0100]
术语多肽或蛋白质的“功能片段”是指为全长多肽或蛋白质的一部分,并且具有与全长多肽或蛋白质基本上相同的生物活性,或执行基本上相同的功能(例如,执行相同的酶促反应)的肽片段。
[0101]
术语“变体多肽”、“经修饰的氨基酸序列”或“经修饰的多肽”,可互换使用,是指与参考多肽的不同之处在于一个或多个氨基酸,例如一个或多个氨基酸取代、缺失和/或添加的氨基酸序列。一方面,变体是保持参考多肽的一些或所有能力的“功能变体”。
[0102]
术语“功能变体”还包括经保守性取代的变体。术语“经保守性取代的变体”是指具有与参考肽的不同之处在于一个或多个保守性氨基酸取代并且保持参考肽的一些或所有活性的氨基酸序列的肽。“保守性氨基酸取代”是氨基酸残基被功能相似的残基取代。保守
性取代的实例包括一个非极性(疏水性)残基如异亮氨酸、缬氨酸、亮氨酸或甲硫氨酸对另一个的取代;一个带电的或极性(亲水性)残基对另一个的取代如精氨酸和赖氨酸之间,谷氨酰胺和天冬酰胺之间,苏氨酸和丝氨酸之间;一个碱性残基如赖氨酸或精氨酸对另一个的取代;或一个酸性残基如天冬氨酸或谷氨酸对另一个的取代;或一个芳香族残基,如苯丙氨酸、酪氨酸或色氨酸对另一个的取代。预计此类取代对蛋白质或多肽的表观分子量或等电点影响很小或没有影响。短语“经保守性取代的变体”还包括其中一个残基被化学衍生化残基置换的肽,条件是所得肽保持如本文所述的参考肽的一些或所有活性。
[0103]
与主题技术的多肽有关的术语“变体”,还包括具有与参考多肽的氨基酸序列至少75%、至少76%、至少77%、至少78%、至少79%、至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%相同的氨基酸序列的功能活性多肽。
[0104]
术语“同源”在其所有语法形式和拼写变型中是指具有“共同进化起源”的多核苷酸或多肽,包括来自超家族的多核苷酸或多肽及来自不同物种的同源多核苷酸或蛋白质之间的关系(reeck等,cell 50:667,1987)。此类多核苷酸或多肽,不管是在百分比同一性还是保守性位置的特定氨基酸或基序的存在上,都具有其序列相似性所反映的序列同源性。例如,两个同源多肽可具有至少75%、至少76%、至少77%、至少78%、至少79%、至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%相同的氨基酸序列。
[0105]
关于主题技术的变体多肽序列的“百分比(%)氨基酸序列同一性”是指在比对序列并且在必要时引入空位以实现最大百分比序列同一性之后,而未将任何保守性取代视为序列同一性的一部分,候选序列中的氨基酸残基与参考多肽的氨基酸残基相同的百分比。
[0106]
为了测定百分比氨基酸序列同一性的比对可以按本领域技术范围内的各种方式实现,例如,使用可公用的计算机软件如blast、blast
‑
2、align、align
‑
2或megalign(dnastar)软件。本领域的技术人员可以测定适当的参数用于测量比对,包括在所比较的序列全长上实现最大限度比对所需的任何算法。例如,可使用序列比较程序ncbi
‑
blast2测定氨基酸序列同一性%。ncbi
‑
blast2序列比较程序可从ncbi.nlm.nih.gov下载。ncbi blast2使用几个搜索参数,其中那些搜索参数全部设为缺省值,包括例如,未遮掩是,链=全部,期望发生10,最小低复杂性长度=15/5,多程e值=0.01,多程常数=25,最终缺口比对的降低=25和评分矩阵=blosum62。在采用ncbi
‑
blast2进行氨基酸序列比较的情况下,给定氨基酸序列a对、与或针对给定氨基酸序列b的氨基酸序列同一性%(可以可选地表达为给定氨基酸序列a对、与或针对给定氨基酸序列b具有或包含某一%氨基酸序列同一性)计算如下:100
×
分数x/y,其中x是通过序列比对程序ncbi
‑
blast2在该程序对a和b的比对中评分为相同匹配的氨基酸残基的数量,并且其中y为b中的氨基酸残基的总数。将认识到氨基酸序列a的长度不等于氨基酸序列b的长度时,a对b的%氨基酸序列同一性将不等于b对a的%氨基酸序列同一性。
[0107]
在这个意义上,测定氨基酸序列“相似性”的技术是本领域公知的。一般而言,“相似性”是指在氨基酸相同或具有相似化学和/或物理性质如电荷或疏水性的适当位置对两
个或更多个多肽的氨基酸与氨基酸的精确比较。然后可测定所比较的多肽序列之间的所谓“百分比相似性”。测定核酸和氨基酸序列同一性的技术也是本领域公知的并且包括测定对于该基因而言mrna的核苷酸序列(通常经由cdna中间体)并测定其中编码的氨基酸序列,并且将这与第二氨基酸序列进行比较。一般而言,“同一性”是指分别两个多核苷酸或多肽序列的核苷酸与核苷酸或氨基酸与氨基酸的精确一致性。正如两个或更多个氨基酸序列一样,可以通过测定其“百分比同一性”来比较两个或更多个多核苷酸序列。wisconsin序列分析包第8版(可从genetics computer group,madison,wis.获得)中可用的程序,例如gap程序,能够分别计算两个多核苷酸之间的同一性及两个多肽序列之间的同一性和相似性。计算序列之间的同一性或相似性的其它程序为本领域技术人员已知。
[0108]
与参考位置“对应”的氨基酸位置是指通过比对氨基酸序列鉴定,与参考序列匹配的位置。此类比对可通过人工或通过使用公知的序列比对程序如clustalw2、blast 2等进行。
[0109]
除非另有说明,否则两个多肽或多核苷酸序列的百分比同一性是指在两个序列中较短序列的整个长度上相同氨基酸残基或核苷酸的百分比。
[0110]“编码序列”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指编码特定氨基酸序列的dna序列。
[0111]“合适调控序列”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指位于编码序列上游(5’非编码序列)、内部或下游(3’非编码序列)的核苷酸序列,并且其影响相关编码序列的转录、rna加工或稳定性或翻译。调控序列可包括启动子、翻译前导序列、内含子和聚腺苷酸化识别序列。
[0112]“启动子”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指能够控制编码序列或功能rna的表达的dna序列。一般而言,编码序列位于启动子序列的3’。启动子可整个源自天然基因,或由源自自然界发现的不同启动子的不同元件组成,或甚至包含合成dna片段。本领域的技术人员应理解,不同启动子可指导不同细胞类型中或不同发育阶段或响应于不同环境条件的基因的表达。使基因大多数时间在大多数细胞类型中表达的启动子,通常称为“组成型启动子”。还进一步认识到,因为在大多数情况下调控序列的确切边界尚未完全定义,所以不同长度的dna片段可具有相同的启动子活性。
[0113]
术语“可操作连接的”是指单个核酸片段上的核酸序列的缔合,使得一个核酸序列的功能受另一个影响。例如,启动子在能够影响编码序列的表达(即编码序列处于启动子的转录控制下)时,启动子与该编码序列可操作连接。编码序列可以在有义或反义方向上与调控序列可操作连接。
[0114]
如本文中所用的术语“表达”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指源自主题技术的核酸片段的有义(mrna)或反义rna的转录和稳定积聚。“过表达”是指转基因或重组生物体中基因产物的生成超过正常或非转化生物体中的生成水平。
[0115]“转化”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指多核苷酸向靶细胞中的转移。转移的多核苷酸可以并入到靶细胞的基因组或染色体dna中,产生基因稳定性遗传,或其可以独立于宿主染色体而复制。含有转化核酸片段的宿主生物体称为“转基因”或“重组”或“转化”生物体。
[0116]
术语“转化”、“转基因”和“重组”,当在本文中连同宿主细胞使用时,根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指已经向其中引入了异源核酸分子的宿主生物体的细胞,如植物或微生物细胞。核酸分子可以稳定整合到宿主细胞的基因组中,或核酸分子可以作为染色体外分子存在。此类染色体外分子可以自主复制。转化细胞、组织或受试者被理解为不但涵盖转化过程的最终产物,而且涵盖其转基因后代。
[0117]
术语“重组”、“异源”和“外源”,当在本文中连同多核苷酸使用时,根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指对特定宿主细胞来说外来来源的,或者若来自相同来源,则由其原始形式修饰的多核苷酸(例如,dna序列或基因)。因此,宿主细胞中的异源基因包括特定宿主细胞内源的但已经通过例如使用定点诱变或其它重组技术修饰的基因。所述术语还包括天然存在的dna序列的非天然存在的多个拷贝。因此,所述术语是指对细胞来说外来的或异源的,或与细胞同源但处于宿主细胞内一般未发现该元件的位置或形式的dna片段。
[0118]
类似地,术语“重组”、“异源”和“外源”,当在本文中连同多肽或氨基酸序列使用时,意指来源于对特定宿主细胞来说外来来源的或者,若来自相同来源,则由其原始形式修饰的多肽或氨基酸序列。因此,重组dna片段可以在宿主细胞中表达以生成重组多肽。
[0119]
术语“质粒”、“载体”和“盒”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指常常携带并非细胞中心代谢的一部分且通常呈环状双链dna分子形式的基因的染色体外元件。此类元件可为源自任何来源的单链或双链dna或rna的线性或环状自主复制序列、基因组整合序列、噬菌体或核苷酸序列,其中许多核苷酸序列已经连接或重组到能够连同适当的3’非翻译序列一起将选定基因产物的启动子片段和dna序列引入细胞中的独特构造中。“转化盒”是指含有外来基因且除该外来基因外还具有利于特定宿主细胞转化的元件的特定载体。“表达盒”是指含有外来基因且除该外来基因外还具有允许该基因在外来宿主中增强表达的元件的特定载体。
[0120]
本文中所用的标准重组dna和分子克隆技术是本领域公知的并且,例如,由sambrook,j.、fritsch,e.f.和maniatis,t.molecular cloning:alaboratory manual,第2版;cold spring harbor laboratory:cold spring harbor,n.y.,1989(下文称为“maniatis”);和由silhavy,t.j.、bennan,m.l.和enquist,l.w.experiments with gene fusions;cold spring harbor laboratory:cold spring harbor,n.y.,1984;和由ausubel,f.m.等,在1987年由greene publishing和wiley
‑
interscience出版的current protocols in molecular biology中有描述,其各自整体据此通过引用在与之一致的程度上并入本文。
[0121]
如本文中所用,“合成”或“有机合成”或“化学合成”用于指通过一系列化学反应制备化合物;这不包括提取化合物,例如,从天然来源提取。
[0122]
如本文中所用的术语“可口服消耗产品”是指与人或动物口腔接触的任何饮料、食物产品、膳食补充剂、营养制剂、药物组合物、口腔卫生组合物和美容产品,包括摄入口腔并随后由口腔排出的物质及饮用、嚼食、吞咽或以其它方式消化的物质;及在通常可接受的浓度范围内使用时对人或动物消耗而言是安全的物质。
[0123]
如本文中所用的术语“食物产品”是指水果、蔬菜、果汁、肉制品如火腿、培根(bacon)、香肠;蛋制品、水果浓缩物、明胶和明胶样产品如果酱、果冻、蜜饯等;乳制品如冰
激凌、酸奶油、酸奶和冻果汁露;糖霜、糖浆,包括糖蜜;玉米、小麦、黑麦、大豆、燕麦、水稻和大麦制品、谷类制品、坚果仁和坚果制品、蛋糕、曲奇、糖食(如糖果)、树胶、水果味硬糖(fruit flavored drop)和巧克力、口香糖、薄荷、奶油、糖霜、冰淇淋、馅饼和面包。“食物产品”还指调味品如药草、香料和佐料、增香剂,如谷氨酸一钠。“食物产品”进一步指还包括制备的包装产品,如饮食甜味剂、液体甜味剂、桌面调味剂、与水冲调得到非碳酸饮料的颗粒调味混合物、速食布丁混合物、速溶咖啡和茶、咖啡伴侣、麦乳精混合物、宠物食品、牲畜饲料、烟草,以及用于烘焙应用的材料,例如用于制备面包、曲奇、蛋糕、薄煎饼、甜甜圈等的粉状烘焙混合物。“食物产品”还指含有少量或不含蔗糖的减肥或低热量食品和饮料。
[0124]
如本文中所用,术语“立体异构体”是仅在其原子在空间中的取向上不同的独立分子的所有异构体的统称。“立体异构体”包括对映异构体和彼此非镜像的具有多于一个手性中心的化合物的异构体(非对映异构体)。
[0125]
如本文中所用,术语“非晶形莱鲍迪苷v”是指莱鲍迪苷v的非结晶固体形式。如本文中所用,术语“非晶形莱鲍迪苷w”是指莱鲍迪苷w的非结晶固体形式。
[0126]
如本文中所用,术语“甜味强度”是指由个体(例如人)所观察到的或经历的甜味感觉的相对强度,或由尝味者例如基于布里标度(brix scale)检测的甜味的程度或量。
[0127]
如本文中所用,术语“提高甜度”是指与不含莱鲍迪苷v和/或莱鲍迪苷w的相应可口服消耗产品相比,莱鲍迪苷v和/或莱鲍迪苷w增加、增大、增强、加重、放大和/或加强本公开的饮料产品或可消耗产品的一种或多种甜度特性的感官知觉而不改变其性质或质量的作用。
[0128]
如本文中所用,术语“异味”是指非典型或不常存在于本公开的饮料产品或可消耗产品中的味道的量或程度。例如,异味是对于消费者来说增甜消耗品不理想的味道,例如,苦味、类甘草味、金属味、令人反感的味道、涩味、延迟的甜味产生、绵长的甜回味及类似味道,等等。
[0129]
如本文中所用,术语“w/v
‑
%”是指对于每100ml含化合物如糖的本公开的液体可口服消耗产品而言此类化合物的重量(按克计)。如本文中所用,术语“w/w
‑
%”是指对于每克含化合物如糖的本公开的可口服消耗产品而言此类化合物的重量(按克计)。
[0130]
如本文中所用,术语“ppm”是指按重量计的百万分率,例如每千克含化合物的本公开的可口服消耗产品中此类化合物,如莱鲍迪苷v和/或莱鲍迪苷w的重量(按毫克计)(即,mg/kg)或每升含化合物的本公开的可口服消耗产品中此类化合物,如莱鲍迪苷v和/或莱鲍迪苷w的重量(按毫克计)(即,mg/l);或按体积计的百万分率,例如每升含化合物的本公开的可口服消耗产品中此类化合物,如莱鲍迪苷v和/或莱鲍迪苷w的重量(按毫升计)(即,ml/l)。
[0131]
根据本公开,公开了无热量甜味剂和合成无热量甜味剂的方法。同样根据本公开,公开了一种酶和使用该酶制备无热量甜味剂的方法。
[0132]
合成的无热量甜味剂:合成莱鲍迪苷v
[0133]
一方面,本公开涉及一种合成的无热量甜味剂。合成的无热量甜味剂是合成的莱鲍迪苷型甜菊醇糖苷并且已经命名为“莱鲍迪苷v”。莱鲍迪苷v(“reb v”)是在其结构中具有四个与糖苷配基甜菊醇连接的β
‑
d
‑
葡糖基单元的甜菊醇糖苷,甜菊醇糖苷配基部分在c
‑
13处具有呈醚键形式的glcβ1
‑3‑
glcβ1单元且在c
‑
19位置处具有另一个呈酯键形式的glcβ
1
‑2‑
glcβ1单元。
[0134]
基于大量的1d和2d nmr以及高分辨率质谱数据和水解研究,莱鲍迪苷v具有分子式c
44
h
70
o
23
和iupac名称,13
‑
[(3
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)氧基]对映
‑
贝壳杉
‑
16
‑
烯
‑
19
‑
烯酸
‑
(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)酯。
[0135]
合成的无热量甜味剂:合成莱鲍迪苷w
[0136]
一方面,本公开涉及一种合成的无热量甜味剂。合成的无热量甜味剂是合成的莱鲍迪苷型甜菊醇糖苷并且已经命名为“莱鲍迪苷w”。莱鲍迪苷w(“reb w”)是在其结构中具有五个与糖苷配基甜菊醇连接的β
‑
d
‑
葡糖基单元的甜菊醇糖苷,甜菊醇糖苷配基部分在c
‑
13处具有呈醚键形式的glcβ1
‑3‑
glcβ1单元且在c
‑
19位置处具有呈酯键形式的glcβ1
‑
2(glcβ1
‑
3)
‑
glcβ1单元。
[0137]
莱鲍迪苷w具有分子式c
50
h
80
o
28
和iupac名称,13
‑
[(3
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)氧基]对映
‑
贝壳杉
‑
16
‑
烯
‑
19
‑
烯酸
‑
[(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑3‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)酯。
[0138]
合成的无热量甜味剂:合成莱鲍迪苷ka
[0139]
一方面,本公开涉及一种合成的无热量甜味剂。合成的无热量甜味剂是合成的莱鲍迪苷型甜菊醇糖苷并且已经命名为“莱鲍迪苷ka”。莱鲍迪苷ka(“reb ka”)是在其结构中具有三个与糖苷配基甜菊醇连接的β
‑
d
‑
葡糖基单元的甜菊醇糖苷,甜菊醇糖苷配基部分在c
‑
13处具有呈醚键形式的glcβ1单元且在c
‑
19处具有呈醚键形式的glcβ1
‑2‑
glcβ1单元。基于大量的1d和2d nmr以及高分辨率质谱数据和水解研究,莱鲍迪苷ka具有分子式c
38
h
60
o
18
和iupac名称,13
‑
β
‑
d
‑
吡喃葡糖基氧基]对映
‑
贝壳杉
‑
16
‑
烯
‑
19
‑
烯酸
‑
(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)酯。
[0140]
合成的无热量甜味剂:合成莱鲍迪苷g
[0141]
一方面,本公开涉及一种合成的无热量甜味剂。合成的无热量甜味剂是合成的莱鲍迪苷型甜菊醇糖苷并且已经命名为“莱鲍迪苷g”。莱鲍迪苷g(“reb g”)是在其结构中具有三个与糖苷配基甜菊醇连接的β
‑
d
‑
葡糖基单元的甜菊醇糖苷,甜菊醇糖苷配基部分在c
‑
13处具有呈醚键形式的glcβ1
‑3‑
glcβ1单元且在c
‑
19处具有呈醚键形式的glcβ1单元。
[0142]
基于大量的1d和2d nmr以及高分辨率质谱数据和水解研究,莱鲍迪苷g具有分子式c
38
h
60
o
18
和iupac名称,13
‑
[(3
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)氧基]对映
‑
贝壳杉
‑
16
‑
烯
‑
19
‑
烯酸
‑
β
‑
d
‑
吡喃葡糖基)酯。
[0143]
合成的无热量甜味剂:合成莱鲍迪苷m
[0144]
一方面,本公开涉及一种合成的无热量甜味剂。合成的无热量甜味剂是合成的莱鲍迪苷型甜菊醇糖苷并且已经命名为“莱鲍迪苷m”。莱鲍迪苷m(“reb m”)是在其结构中具有六个与糖苷配基甜菊醇连接的β
‑
d
‑
葡糖基单元的甜菊醇糖苷,甜菊醇糖苷配基部分在c
‑
13位置处具有呈醚键形式的glcβ1
‑
2(glcβ1
‑
3)
‑
glcβ1单元且在c
‑
19位置处具有呈酯键形式的glcβ1
‑
2(glcβ1
‑
3)
‑
glcβ1单元。
[0145]
基于大量的1d和2d nmr以及高分辨率质谱数据和水解研究,莱鲍迪苷m具有分子式c
56
h
90
o
33
和iupac名称,13
‑
[(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑3‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)氧基]对映
‑
贝壳杉
‑
16
‑
烯
‑
19
‑
烯酸
‑
[(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑3‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)酯。
[0146]
合成甜菊醇糖苷的方法
[0147]
由莱鲍迪苷g生成莱鲍迪苷v的方法。另一方面,本公开涉及一种由莱鲍迪苷g合成莱鲍迪苷v的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷g;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;和hv1 udp
‑
糖基转移酶;有或无蔗糖合酶(sus)以及温育所述反应混合物足够时间以生成莱鲍迪苷v,其中葡萄糖共价偶联至莱鲍迪苷g以生成莱鲍迪苷v。
[0148]
由莱鲍迪苷g生成莱鲍迪苷v的方法。另一方面,本公开涉及一种由莱鲍迪苷g合成莱鲍迪苷v的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷g;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自eugt11、udp
‑
糖基转移酶
‑
蔗糖合酶(sus)融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶);有或无蔗糖合酶(sus)以及温育所述反应混合物足够时间以生成莱鲍迪苷v,其中葡萄糖共价偶联至莱鲍迪苷g以生成莱鲍迪苷v。
[0149]
由莱鲍迪苷ka生成莱鲍迪苷v的方法。
[0150]
另一方面,本公开涉及一种由莱鲍迪苷ka合成莱鲍迪苷v的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷ka;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自udp
‑
糖基转移酶(ugt76g1)和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶);有或无蔗糖合酶(sus)以及温育所述反应混合物足够时间以生成莱鲍迪苷v,其中葡萄糖共价偶联至莱鲍迪苷ka以生成莱鲍迪苷v。
[0151]
由甜茶苷生成莱鲍迪苷v的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷v的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自udp
‑
糖基转移酶(ugt76g1)、hv1和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶);有或无蔗糖合酶(sus)以及温育所述反应混合物足够时间以生成莱鲍迪苷v,其中葡萄糖共价偶联至甜茶苷以生成莱鲍迪苷ka。连续地,葡萄糖共价偶联至莱鲍迪苷ka以生成莱鲍迪苷v。
[0152]
由甜茶苷生成莱鲍迪苷v的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷a和莱鲍迪苷v的混合物的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自udp
‑
糖基转移酶(ugt76g1)、eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶);有或无蔗糖合酶(sus);以及温育所述反应混合物足够时间以生成莱鲍迪苷v,其中葡萄糖共价偶联至甜茶苷以生成莱鲍迪苷ka并且葡萄糖共价偶联至莱鲍迪苷ka以生成莱鲍迪苷v。葡萄糖共价偶联至甜茶苷以生成莱鲍迪苷g。连续地,葡萄糖共价偶联至莱鲍迪苷g以生成莱鲍迪苷v。
[0153]
由莱鲍迪苷v生成莱鲍迪苷w的方法。另一方面,本公开涉及一种由莱鲍迪苷v合成莱鲍迪苷w的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷v;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自udp
‑
糖基转移酶(ugt76g1)和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶);有或无蔗糖合酶(sus)以及温育所述反应混合物足够时间以生成莱鲍迪苷w,其中葡萄糖共价偶联至莱鲍迪苷v以生成莱鲍迪苷w。
[0154]
由莱鲍迪苷g生成莱鲍迪苷w的方法。另一方面,本公开涉及一种由莱鲍迪苷g合成莱鲍迪苷w的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷g;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自尿苷二磷酸糖基转移酶(ugt76g1)、udp
‑
糖基转移酶
‑
蔗糖合酶融合酶和hv1的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶);有或无蔗糖合酶(sus);以及温育所述反应混合物足够时间以生成莱鲍迪苷w,其中葡萄糖通过hv1共价偶联至莱鲍迪苷g以生成莱鲍迪苷v。连续地,葡萄糖通过ugt76g1共价偶联至莱鲍迪苷v以生成莱鲍迪苷w。
[0155]
由莱鲍迪苷g生成莱鲍迪苷w的方法。另一方面,本公开涉及一种由莱鲍迪苷g合成莱鲍迪苷w的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷g;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自ugt76g1、eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶);以及温育所述反应混合物足够时间以生成莱鲍迪苷w,其中葡萄糖通过eugt11共价偶联至莱鲍迪苷g以生成莱鲍迪苷v。连续地,葡萄糖通过ugt76g1共价偶联至莱鲍迪苷v以生成莱鲍迪苷w。
[0156]
由莱鲍迪苷ka生成莱鲍迪苷w的方法。另一方面,本公开涉及一种由莱鲍迪苷ka合成莱鲍迪苷w的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷ka;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自尿苷二磷酸糖基转移酶(ugt76g1)和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶(udp
‑
糖基转移酶);有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷w,其中葡萄糖共价偶联至莱鲍迪苷ka以生成莱鲍迪苷v。连续地,葡萄糖共价偶联至莱鲍迪苷v以生成莱鲍迪苷w。
[0157]
由甜茶苷生成莱鲍迪苷w的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷w的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自ugt76g1、hv1和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶;有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷w的混合物。
[0158]
由甜茶苷生成莱鲍迪苷w的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷w的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自ugt76g1、eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的尿苷二磷酸糖基转移酶;有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷w。
[0159]
由甜茶苷生成甜菊苷和莱鲍迪苷ka的混合物的方法。另一方面,本公开涉及一种由甜茶苷合成甜菊苷和莱鲍迪苷ka的混合物的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的udp
‑
糖基转移酶;有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成甜菊苷和莱鲍迪苷ka的混合物,其中葡萄糖共价偶联至甜茶苷的c2
’‑
19
‑
o
‑
葡萄糖以生成莱鲍迪苷ka;其中葡萄糖共价偶联至甜茶苷的c2
’‑
13
‑
o
‑
葡萄糖以生成甜菊苷。
[0160]
由甜茶苷生成莱鲍迪苷ka的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷ka的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷;选自蔗糖、尿苷
二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;和hv1 udp
‑
糖基转移酶;有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷ka,其中葡萄糖共价偶联至甜茶苷的c2
’‑
19
‑
o
‑
葡萄糖以生成莱鲍迪苷ka。
[0161]
由甜茶苷生成莱鲍迪苷g的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷g的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自ugt76g1和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的udp
‑
糖基转移酶;有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷g,其中葡萄糖共价偶联至甜茶苷的c3
’‑
13
‑
o
‑
葡萄糖以生成莱鲍迪苷g。
[0162]
由莱鲍迪苷ka生成莱鲍迪苷e的方法。另一方面,本公开涉及一种由莱鲍迪苷ka合成莱鲍迪苷e的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷ka;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;和hv1 udp
‑
糖基转移酶;有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷e,其中葡萄糖共价偶联至莱鲍迪苷ka的c2’13
‑
o
‑
葡萄糖以生成莱鲍迪苷e。
[0163]
由莱鲍迪苷ka生成莱鲍迪苷e的方法。另一方面,本公开涉及一种由莱鲍迪苷ka合成莱鲍迪苷e的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷ka;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的udp
‑
糖基转移酶;有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷e,其中葡萄糖共价偶联至莱鲍迪苷ka的c2’13
‑
o
‑
葡萄糖以生成莱鲍迪苷e。
[0164]
由甜茶苷生成莱鲍迪苷e的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷e的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;和选自eugt11和udp
‑
糖基转移酶
‑
蔗糖合成融合酶的udp
‑
糖基转移酶;有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷e,其中葡萄糖共价偶联至甜茶苷以生成莱鲍迪苷ka和甜菊苷的混合物。连续地,葡萄糖共价偶联至莱鲍迪苷ka和甜菊苷以生成莱鲍迪苷e。
[0165]
由甜茶苷生成莱鲍迪苷e的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷e的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;和hv1 udp
‑
糖基转移酶;有或无蔗糖合酶;温育所述反应混合物足够时间以生成莱鲍迪苷e,其中葡萄糖共价偶联至甜茶苷以生成莱鲍迪苷ka;并且与hv1一起进一步温育莱鲍迪苷ka以生成莱鲍迪苷e。
[0166]
由甜茶苷生成莱鲍迪苷d3的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷d3的方法。该方法包括制备反应混合物,所述反应混合物包含甜茶苷;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;选自eugt11和udp
‑
糖基转移酶
‑
蔗糖合成融合酶的udp
‑
糖基转移酶;有或无蔗糖合酶;温育所述反应混合物足够时间以生成莱鲍迪苷d3,其中葡萄糖共价偶联至甜茶苷以生成甜菊苷和莱鲍迪苷ka的混合物;与eugt11一起进一步温育甜菊苷和莱鲍迪苷ka的混合物以生成莱鲍迪苷e,其中葡萄糖共价偶联至甜菊苷和莱鲍迪苷ka以生成莱鲍迪苷e;并且与eugt11一起进一步温育莱鲍迪苷e以生成莱鲍迪苷d3,其中葡萄糖共价偶联至莱鲍迪苷e以生成莱鲍迪苷d3。
[0167]
由莱鲍迪苷ka生成莱鲍迪苷d3的方法。另一方面,本公开涉及一种由莱鲍迪苷ka
合成莱鲍迪苷d3的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷ka,选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物,选自eugt11和udp
‑
糖基转移酶
‑
蔗糖合酶融合酶的udp
‑
糖基转移酶,有或无蔗糖合酶;温育所述反应混合物足够时间以生成莱鲍迪苷d3,其中葡萄糖共价偶联至莱鲍迪苷ka以生成莱鲍迪苷e;并且进一步温育莱鲍迪苷e与eugt11的混合物以生成莱鲍迪苷d3,其中葡萄糖共价偶联至莱鲍迪苷e以生成莱鲍迪苷d3。
[0168]
由莱鲍迪苷e生成莱鲍迪苷z的方法。另一方面,本公开涉及一种由莱鲍迪苷e合成莱鲍迪苷z的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷e;选自蔗糖、尿苷二磷酸(udp)和尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)的底物;和hv1 udp
‑
糖基转移酶;及蔗糖合酶;温育所述反应混合物足够时间以生成莱鲍迪苷z,其中葡萄糖共价偶联至莱鲍迪苷e以生成莱鲍迪苷z,其中葡萄糖共价偶联至莱鲍迪苷e的c2
’‑
13
‑
o
‑
葡萄糖以生成莱鲍迪苷z1。葡萄糖共价偶联至莱鲍迪苷e的c2
’‑
19
‑
o
‑
葡萄糖以生成莱鲍迪苷z2。
[0169]
由莱鲍迪苷d生成莱鲍迪苷m的方法。另一方面,本公开涉及一种由莱鲍迪苷d合成莱鲍迪苷m的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷d,选自蔗糖、尿苷二磷酸(udp)、尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)及其组合的底物,及选自ugt76g1、udp
‑
糖基转移酶
‑
蔗糖合酶融合酶及其组合的udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷m,其中葡萄糖共价偶联至莱鲍迪苷d以生成莱鲍迪苷m。
[0170]
由甜菊苷生成莱鲍迪苷d和莱鲍迪苷m的方法。另一方面,本公开涉及一种由甜菊苷合成莱鲍迪苷d和莱鲍迪苷m的方法。该方法包括制备反应混合物,所述反应混合物包含甜菊苷,选自蔗糖、尿苷二磷酸(udp)、尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)及其组合的底物,及选自hv1、ugt76g1、udp
‑
糖基转移酶
‑
蔗糖合酶融合酶及其组合的udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷d和/或莱鲍迪苷m。例如,在实施方案中,可温育所述反应混合物足够时间以生成莱鲍迪苷d,并进一步温育包含莱鲍迪苷d的反应混合物(例如,与ugt76g1和/或融合酶一起)以生成莱鲍迪苷m。在某些实施方案中,反应混合物将包含hv1和ugt76g1。在其它实施方案中,反应混合物将包含hv1和融合酶。
[0171]
在某些实施方案中,葡萄糖共价偶联至甜菊苷以生成莱鲍迪苷a和/或莱鲍迪苷e。例如,葡萄糖可通过ugt76g1或融合酶共价偶联至甜菊苷以生成莱鲍迪苷a和/或葡萄糖可通过hv1共价偶联至甜菊苷以生成莱鲍迪苷e。连续地,葡萄糖可通过hv1共价偶联至莱鲍迪苷a以生成莱鲍迪苷d和/或葡萄糖可通过ugt76g1或融合酶共价偶联至莱鲍迪苷e以生成莱鲍迪苷d。葡萄糖可进一步通过ugt76g1或融合酶共价偶联至莱鲍迪苷d以生成莱鲍迪苷m。
[0172]
由莱鲍迪苷a生成莱鲍迪苷d和莱鲍迪苷m的方法。另一方面,本公开涉及一种由莱鲍迪苷a合成莱鲍迪苷d和莱鲍迪苷m的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷a,选自蔗糖、尿苷二磷酸(udp)、尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)及其组合的底物,及选自hv1、ugt76g1、udp
‑
糖基转移酶
‑
蔗糖合酶融合酶及其组合的udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷d和/或莱鲍迪苷m。例如,在实施方案中,可温育反应混合物(例如,包含hv1)足够时间以生成莱鲍迪苷d,并进一步温育包含莱鲍迪苷d的反应混合物(例如,与ugt76g1和/或融合酶一起)以生成莱鲍
迪苷m。在某些实施方案中,反应混合物将包含hv1和ugt76g1。在其它实施方案中,反应混合物将包含hv1和融合酶。
[0173]
葡萄糖共价偶联至莱鲍迪苷a以生成莱鲍迪苷d。例如,葡萄糖可通过hv1共价偶联至莱鲍迪苷a以生成莱鲍迪苷d。连续地,葡萄糖可通过ugt76g1或融合酶共价偶联至莱鲍迪苷d以生成莱鲍迪苷m。
[0174]
由莱鲍迪苷e生成莱鲍迪苷d和莱鲍迪苷m的方法。另一方面,本公开涉及一种由莱鲍迪苷e合成莱鲍迪苷d和莱鲍迪苷m的方法。该方法包括制备反应混合物,所述反应混合物包含莱鲍迪苷e,选自蔗糖、尿苷二磷酸(udp)、尿苷二磷酸
‑
葡萄糖(udp
‑
葡萄糖)及其组合的底物,及选自ugt76g1、udp
‑
糖基转移酶
‑
蔗糖合酶融合酶及其组合的udp
‑
糖基转移酶,有或无蔗糖合酶;以及温育所述反应混合物足够时间以生成莱鲍迪苷d和/或莱鲍迪苷m。例如,在实施方案中,可温育所述反应混合物(例如,包含ugt76g1和/或融合酶)足够时间以生成莱鲍迪苷d,并进一步温育包含莱鲍迪苷d的反应混合物以生成莱鲍迪苷m。
[0175]
葡萄糖共价偶联至莱鲍迪苷e以生成莱鲍迪苷d。例如,葡萄糖可通过ugt76g1或融合酶共价偶联至莱鲍迪苷e以生成莱鲍迪苷d。连续地,葡萄糖可通过ugt76g1或融合酶共价偶联至莱鲍迪苷d以生成莱鲍迪苷m。
[0176]
大多数甜菊醇糖苷是使用尿苷5
’‑
二磷酸葡萄糖(udp
‑
葡萄糖)作为糖部分的供体,通过通常由udp
‑
糖基转移酶(ugt)催化的甜菊醇的几个糖基化反应形成的。在植物中,ugt是酶的非常不同的类,其将葡萄糖残基从udp
‑
葡萄糖转移到甜菊醇。
[0177]
尿苷二磷酸糖基转移酶(ugt76g1)是具有生成有关糖苷(莱鲍迪苷a和d)的1,3
‑
13
‑
o
‑
葡萄糖糖基化活性的ugt。令人惊讶且出乎意料的是,发现ugt76g1也具有由甜茶苷生成莱鲍迪苷g及由莱鲍迪苷d生成莱鲍迪苷m的1,3
‑
19
‑
o
‑
葡萄糖糖基化活性。ugt76g1可以将莱鲍迪苷ka转化为reb v并继续形成reb w。特别合适的ugt76g1具有seq id no:1的氨基酸序列。
[0178]
eugt11(在wo 2013022989中有描述)是具有1,2
‑
19
‑
o
‑
葡萄糖和1,2
‑
13
‑
o
‑
葡萄糖糖基化活性的ugt。已知eugt11会催化甜菊苷生成莱鲍迪苷e和莱鲍迪苷a生成莱鲍迪苷d。令人惊讶且出乎意料的是,发现eugt11可在体外用于通过新的酶活性(β1,6
‑
13
‑
o
‑
葡萄糖糖基化活性)由莱鲍迪苷e合成莱鲍迪苷d3(转让给conagen,inc.的美国专利申请序列号14/269,435)。eugt11具有由甜茶苷生成莱鲍迪苷ka的1,2
‑
19
‑
o
‑
葡萄糖糖基化活性。特别合适的eugt11具有seq id no:3的氨基酸序列。
[0179]
hv1是具有生成有关甜菊醇糖苷(莱鲍迪苷e、d和z)的1,2
‑
19
‑
o
‑
葡萄糖糖基化活性的ugt。令人惊讶且出乎意料的是,发现hv1也具有由甜茶苷生成莱鲍迪苷ka的1,2
‑
19
‑
o
‑
葡萄糖糖基化活性。hv1也可以将reb g转化为reb v和将reb ka转化为reb e。特别合适的hv1具有seq id no:5的氨基酸序列。
[0180]
所述方法还可包括向含有尿苷二磷酸(udp)糖基转移酶的反应混合物中添加蔗糖合酶。蔗糖合酶催化ndp
‑
葡萄糖和d
‑
果糖之间的化学反应以生成ndp和蔗糖。蔗糖合酶为糖基转移酶。这种酶类的系统名称为ndp
‑
葡萄糖:d
‑
果糖2
‑
α
‑
d
‑
葡糖基转移酶。常用的其它名称包括udp葡萄糖
‑
果糖葡糖基转移酶、蔗糖合成酶、蔗糖
‑
udp葡糖基转移酶、蔗糖
‑
尿苷二磷酸葡糖基转移酶和尿苷二磷酸葡萄糖
‑
果糖葡糖基转移酶。向包括尿苷二磷酸糖基转移酶的反应混合物中添加蔗糖合酶产生“ugt
‑
sus偶联体系”。在ugt
‑
sus偶联体系中,可由udp
和蔗糖再生udp
‑
葡萄糖,这允许省略向反应混合物中添加额外的udp
‑
葡萄糖或在反应混合物中使用udp。合适的蔗糖合酶可以是例如,拟南芥(arabidopsis)蔗糖合酶1;拟南芥蔗糖合酶3;和绿豆(vigna radiate)蔗糖合酶。特别合适的蔗糖合酶可以是,例如,拟南芥蔗糖合酶1。特别合适的拟南芥蔗糖合酶1是具有seq id no:7的氨基酸序列的阿拉伯芥(arabidopsis thaliana)蔗糖合酶1(atsus1)。
[0181]
另一方面,尿苷二磷酸糖基转移酶融合酶可用于所述方法中。特别合适的尿苷二磷酸糖基转移酶融合酶可以是ugt
‑
sus1融合酶。udp
‑
糖基转移酶可以是包括偶联至蔗糖合酶结构域的尿苷二磷酸糖基转移酶结构域的udp
‑
糖基转移酶融合酶。具体而言,udp
‑
糖基转移酶融合酶包括偶联至蔗糖合酶结构域的尿苷二磷酸糖基转移酶结构域。另外,ugt
‑
sus1融合酶具有蔗糖合酶活性,并且因此,可以由udp和蔗糖再生udp
‑
葡萄糖。特别合适的ugt
‑
sus1融合酶可以是,例如,具有seq id no:9的氨基酸序列的ugt76g1
‑
atsus1融合酶(命名为:“gs”)。另一种特别合适的ugt
‑
sus1融合酶可以是,例如,具有seq id no:11的氨基酸序列的eugt11
‑
sus1融合酶(命名为:“eus”)。
[0182]
合适的蔗糖合酶结构域可以是例如,拟南芥蔗糖合酶1;拟南芥蔗糖合酶3;和绿豆蔗糖合酶。特别合适的蔗糖合酶结构域可以是,例如,拟南芥蔗糖合酶1。特别合适的拟南芥蔗糖合酶1是具有seq id no:7的氨基酸序列的阿拉伯芥蔗糖合酶1(atsus1)。
[0183]
ugt76g1
‑
atsus1(“gs”)融合酶可具有与seq id no:9中列出的氨基酸序列至少70%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%相同的多肽序列。适当地,ugt
‑
atsus1融合酶的氨基酸序列与seq id no:9具有至少80%同一性。更适当地,ugt
‑
atsus1融合酶的氨基酸序列与seq id no:9中列出的氨基酸序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%的氨基酸序列同一性。
[0184]
分离的核酸可包括编码ugt
‑
atsus1融合酶的多肽的核苷酸序列,所述分离的核酸具有与seq id no:10中列出的核酸序列具有至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同源性的核酸序列。适当地,分离的核酸包括编码udp
‑
糖基转移酶融合酶的多肽的核苷酸序列,所述udp
‑
糖基转移酶融合酶具有与seq id no:9中列出的氨基酸序列具有至少80%序列同一性的氨基酸序列。更适当地,分离的核酸包括编码udp
‑
糖基转移酶融合酶的多肽的核苷酸序列,所述udp
‑
糖基转移酶融合酶具有与seq id no:9中列出的氨基酸序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的氨基酸序列。分离的核酸因此包括编码seq id no:10的功能片段、seq id no:9的功能变体或与seq id no:9具有(例如)至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的其它同源多肽的那些核苷酸序列。正如本领域技术人员所知,编码udp
‑
糖基转移酶的核酸序列可以经密码子优化以在合适的宿主生物体,例如细菌和酵母中表达。
[0185]
eugt11
‑
sus1(“eus”)融合酶可具有与seq id no:11中列出的氨基酸序列至少70%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、
至少96%、至少97%、至少98%、至少99%和甚至100%相同的多肽序列。适当地,eugt11
‑
sus1融合酶的氨基酸序列与seq id no:11具有至少80%同一性。更适当地,eugt11
‑
sus1融合酶的氨基酸序列与seq id no:11中列出的氨基酸序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%的氨基酸序列同一性。
[0186]
分离的核酸可包括编码eugt11
‑
sus1融合酶的多肽的核苷酸序列,所述分离的核酸具有与seq id no:12中列出的核酸序列具有至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同源性的核酸序列。适当地,分离的核酸包括编码具有的氨基酸序列与seq id no:11中列出的氨基酸序列具有至少80%序列同一性的eugt11
‑
sus1融合酶的多肽的核苷酸序列。更适当地,分离的核酸包括编码eugt11
‑
sus1融合酶的多肽的核苷酸序列,所述eugt11
‑
sus1融合酶具有与seq id no:11中列出的氨基酸序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的氨基酸序列。分离的核酸因此包括编码seq id no:11的功能片段、seq id no:11的功能变体或与seq id no:11具有(例如)至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的其它同源多肽的那些核苷酸序列。正如本领域技术人员所知,编码eugt11
‑
sus1的核酸序列可以经密码子优化以在合适的宿主生物体,例如细菌和酵母中表达。
[0187]
可口服消耗产品
[0188]
另一方面,本公开涉及一种选自饮料产品和可消耗产品,具有增甜量的莱鲍迪苷v的可口服消耗产品。另一方面,本公开涉及一种选自饮料产品和可消耗产品,具有增甜量的莱鲍迪苷w的可口服消耗产品。另一方面,本公开涉及一种选自饮料产品和可消耗产品,具有增甜量的莱鲍迪苷ka的可口服消耗产品。另一方面,本公开涉及一种选自饮料产品和可消耗产品,具有增甜量的莱鲍迪苷g的可口服消耗产品。另一方面,本公开涉及一种选自饮料产品和可消耗产品,具有增甜量的莱鲍迪苷m的可口服消耗产品。
[0189]
可口服消耗产品可具有相当于约1%(w/v
‑
%)至约4%(w/v
‑
%)蔗糖溶液的甜味强度。
[0190]
可口服消耗产品可具有约5ppm至约100ppm莱鲍迪苷v。可口服消耗产品可具有约5ppm至约100ppm莱鲍迪苷w。可口服消耗产品可具有约5ppm至约100ppm莱鲍迪苷ka。可口服消耗产品可具有约5ppm至约100ppm莱鲍迪苷g。可口服消耗产品可具有约5ppm至约100ppm莱鲍迪苷m。
[0191]
莱鲍迪苷v可以是可口服消耗产品中唯一的甜味剂。莱鲍迪苷w可以是可口服消耗产品中唯一的甜味剂。莱鲍迪苷ka可以是可口服消耗产品中唯一的甜味剂。莱鲍迪苷g可以是可口服消耗产品中唯一的甜味剂。莱鲍迪苷m可以是可口服消耗产品中唯一的甜味剂。
[0192]
可口服消耗产品也可具有至少一种附加甜味剂。所述至少一种附加甜味剂可为例如,天然高强度甜味剂。附加甜味剂可选自甜叶菊提取物、甜菊醇糖苷、甜菊苷、莱鲍迪苷a、莱鲍迪苷b、莱鲍迪苷c、莱鲍迪苷d、莱鲍迪苷d3、莱鲍迪苷e、莱鲍迪苷f、杜尔可苷a、甜茶苷、甜菊醇二糖苷、蔗糖、高果糖玉米糖浆、果糖、葡萄糖、木糖、阿拉伯糖、鼠李糖、赤藓糖
醇、木糖醇、甘露糖醇、山梨糖醇、肌醇、acek、阿斯巴甜、纽甜、三氯蔗糖、糖精、柚皮苷二氢查耳酮(nardhc)、新橙皮苷二氢查耳酮(ndhc)、甜茶苷、罗汉果苷iv、赛门苷i、罗汉果苷v、莫那甜、索马甜、莫奈林、布拉奇因、l
‑
丙氨酸、甘氨酸、罗汉果、赫南德辛、叶甜素、三叶苷及其组合。
[0193]
可口服消耗产品也可具有至少一种添加剂。添加剂可为,例如,碳水化合物、多元醇、氨基酸或其盐、聚氨基酸或其盐、糖酸或其盐、核苷酸、有机酸、无机酸、有机盐、有机酸式盐、有机碱式盐、无机盐、苦味化合物、食用香料、调味成分、涩味化合物、蛋白质、蛋白质水解物、表面活性剂、乳化剂、类黄酮、醇、聚合物及其组合。
[0194]
一方面,本公开涉及一种包含增甜量的莱鲍迪苷v的饮料产品。一方面,本公开涉及一种包含增甜量的莱鲍迪苷w的饮料产品。一方面,本公开涉及一种包含增甜量的莱鲍迪苷ka的饮料产品。一方面,本公开涉及一种包含增甜量的莱鲍迪苷g的饮料产品。一方面,本公开涉及一种包含增甜量的莱鲍迪苷m的饮料产品。
[0195]
饮料产品可以是,例如,碳酸饮料产品和非碳酸饮料产品。饮料产品也可以是,例如,软饮料、泉水饮料、冷冻饮料;即饮饮料;冷冻即饮饮料、咖啡、茶、乳品饮料、粉状软饮料、液体浓缩物、调味水、强化水、果汁、果汁味饮料、运动饮料和能量饮料。
[0196]
在一些实施方案中,本公开的饮料产品可以包括一种或多种饮料成分(例如,酸化剂、果汁和/或蔬菜汁、果肉等)、调味剂、着色剂、防腐剂、维生素、矿物质、电解质、赤藓糖醇、塔格糖、甘油和二氧化碳。此类饮料产品可呈任何合适的形式提供,如饮料浓缩物和碳酸、即饮饮料。
[0197]
在某些实施方案中,本公开的饮料产品可以具有许多不同的特定配方或组成中的任一种。本公开的饮料产品的配方可以在某种程度上根据诸如产品的预期细分市场、其所需营养特性、风味特性等因素改变。例如,在某些实施方案中,向特定饮料产品的配方中添加另外的成分通常可以是一种选择。例如,可向任何此类配方中添加附加(即,更多和/或其它)甜味剂,通常可添加调味剂、电解质、维生素、果汁或其它水果产品、促味剂、掩蔽剂等、增味剂和/或碳酸化剂,以改变味道、口感、营养特性等。在实施方案中,饮料产品可以是含有水、约5ppm至约100ppm莱鲍迪苷v、酸化剂和调味剂的可乐饮料。在实施方案中,饮料产品可以是含有水、约5ppm至约100ppm莱鲍迪苷w、酸化剂和调味剂的可乐饮料。在实施方案中,饮料产品可以是含有水、约5ppm至约100ppm莱鲍迪苷m、酸化剂和调味剂的可乐饮料。示例性调味剂可以是,例如,可乐调味剂、柑橘调味剂和香料调味剂。在一些实施方案中,可添加呈二氧化碳形式的碳酸化剂用于起泡。在其它实施方案中,根据其它成分、生产技术、所需保质期等,可添加防腐剂。在某些实施方案中,可添加咖啡因。在一些实施方案中,饮料产品可以是可乐味碳酸饮料,典型地含有苏打水、甜味剂、可乐果提取物和/或其它调味剂、焦糖色素、一种或多种酸和任选的其它成分。
[0198]
饮料产品中存在的莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g的合适量可以为,例如,约5ppm至约100ppm。在一些实施方案中,低浓度的莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g,例如,低于100ppm,并且具有相当于浓度介于10,000ppm至30,000ppm之间的蔗糖溶液的甜度。最终浓度范围为约5ppm至约100ppm、约5ppm至约95ppm、约5ppm至约90ppm、约5ppm至约85ppm、约5ppm至约80ppm、约5ppm至约75ppm、约5ppm至约70ppm、约5ppm至约65ppm、约5ppm至约60ppm、约5ppm至约55ppm、约5ppm至约
50ppm、约5ppm至约45ppm、约5ppm至约40ppm、约5ppm至约35ppm、约5ppm至约30ppm、约5ppm至约25ppm、约5ppm至约20ppm、约5ppm至约15ppm或约5ppm至约10ppm。可选地,莱鲍迪苷v或莱鲍迪苷w可以按以下范围的最终浓度存在于本公开的饮料产品中:约5ppm至约100ppm、约10ppm至约100ppm、约15ppm至约100ppm、约20ppm至约100ppm、约25ppm至约100ppm、约30ppm至约100ppm、约35ppm至约100ppm、约40ppm至约100ppm、约45ppm至约100ppm、约50ppm至约100ppm、约55ppm至约100ppm、约60ppm至约100ppm、约65ppm至约100ppm、约70ppm至约100ppm、约75ppm至约100ppm、约80ppm至约100ppm、约85ppm至约100ppm、约90ppm至约100ppm或约95ppm至约100ppm。
[0199]
另一方面,本公开涉及一种包含增甜量的莱鲍迪苷v的消耗品。另一方面,本公开涉及一种包含增甜量的莱鲍迪苷w的消耗品。另一方面,本公开涉及一种包含增甜量的莱鲍迪苷ka的消耗品。另一方面,本公开涉及一种包含增甜量的莱鲍迪苷g的消耗品。另一方面,本公开涉及一种包含增甜量的莱鲍迪苷m的消耗品。该消耗品可以是,例如,食物产品、营养制剂、药剂、膳食补充剂、口腔卫生组合物、可食用的凝胶组合物、美容产物和桌面调味剂。
[0200]
如本文中所用,“膳食补充剂”是指意在补充膳食和提供营养素(如维生素、矿物质、纤维、脂肪酸、氨基酸等)的化合物,所述化合物在膳食中可能缺失或不被大量消耗。可使用本领域已知的任何合适的膳食补充剂。合适的膳食补充剂的实例可以是,例如,营养素、维生素、矿物质、纤维、脂肪酸、药草、植物性药材、氨基酸和代谢物。
[0201]
如本文中所用,“营养制剂”是指如下化合物,其包括可提供药用或健康益处的任何食物或食物部分,所述益处包括预防和/或治疗疾病或病症(例如,疲劳、失眠、老化作用、健忘、情绪障碍、心血管疾病和血液中的高胆固醇水平、糖尿病、骨质疏松症、炎症、自身免疫性病症等)。可使用本领域已知的任何合适的营养制剂。在一些实施方案中,营养制剂可用作食品和饮料的补充剂及用作肠内或肠胃外应用的药物制剂,其可以是固体制剂,如胶囊或片剂,或液体制剂,如溶液或混悬液。
[0202]
在一些实施方案中,膳食补充剂和营养制剂可进一步含有保护性水胶体(如树胶、蛋白质、改性淀粉)、粘合剂、成膜剂、封装剂/材料、壁/壳材料、基质化合物、包衣、乳化剂、表面活性剂、增溶剂(油、脂肪、蜡、卵磷脂等)、吸附剂、载体、填料、共化合物、分散剂、润湿剂、加工助剂(溶剂)、流化剂、掩味剂、增重剂、成胶剂、胶凝剂、抗氧化剂和抗菌剂。
[0203]
如本文中所用,“凝胶”是指胶体体系,该体系中颗粒网络横跨液体介质的体积。虽然凝胶主要由液体组成,并因此展现出类似液体的密度,但由于横跨液体介质的颗粒网络,因而凝胶具有固体的结构连贯性。出于这个原因,凝胶通常表现为固体、胶状材料。凝胶可用于许多应用。例如,凝胶可用于食物、涂料和粘合剂中。可食用的凝胶被称为“可食用凝胶组合物”。可食用凝胶组合物通常作为小吃、作为甜品、作为主食的一部分或与主食一起被食用。合适的可食用凝胶组合物的实例可以是,例如,凝胶甜品、布丁、果酱、果冻、浆糊、松糕、肉冻、棉花糖、橡皮糖等。在一些实施方案中,可食用凝胶混合物通常是粉状或颗粒状固体,可向其中添加流体以形成可食用凝胶组合物。合适的流体实例可以是,例如,水、乳品流体、乳品类似物流体、果汁、酒精、酒精饮料及其组合。合适的乳品流体的实例可以是,例如,牛奶、发酵乳、奶油、液体乳清及其混合物。合适的乳品类似物流体的实例可以是,例如,豆奶和非乳品咖啡伴侣。
[0204]
如本文中所用,术语“胶凝成分”是指可以在液体介质中形成胶体体系的任何材
料。合适的胶凝成分的实例可以是,例如,明胶、藻酸盐、鹿角菜胶、树胶、果胶、魔芋、琼脂、食用酸、凝乳酵素(rennet)、淀粉、淀粉衍生物及其组合。本领域普通技术人员熟知用于可食用凝胶混合物或可食用凝胶组合物的胶凝成分的量可根据许多因素而显著变化,所述因素例如使用的特定胶凝成分、使用的特定液基以及所需的凝胶性质。
[0205]
本公开的凝胶混合物和凝胶组合物可通过本领域已知的任何合适的方法制备。在一些实施方案中,除胶凝剂外,可使用其它成分制备本公开的可食用凝胶混合物和可食用凝胶组合物。其它合适成分的实例可以是,例如,食用酸、食用酸的盐、缓冲体系、膨胀剂、螯合剂、交联剂、一种或多种调料、一种或多种着色剂及其组合。
[0206]
本领域中已知的任何合适的药物组合物均可使用。在某些实施方案中,本公开的药物组合物可含有约5ppm至约100ppm的莱鲍迪苷v和一种或多种药学上可接受的赋形剂。在某些实施方案中,本公开的药物组合物可含有约5ppm至约100ppm的莱鲍迪苷w和一种或多种药学上可接受的赋形剂。在某些实施方案中,本公开的药物组合物可含有约5ppm至约100ppm的莱鲍迪苷ka和一种或多种药学上可接受的赋形剂。在某些实施方案中,本公开的药物组合物可含有约5ppm至约100ppm的莱鲍迪苷g和一种或多种药学上可接受的赋形剂。在某些实施方案中,本公开的药物组合物可含有约5ppm至约100ppm的莱鲍迪苷m和一种或多种药学上可接受的赋形剂。在一些实施方案中,本公开的药物组合物可用于配制含有一种或多种发挥生物作用的活性剂的药物。因此,在一些实施方案中,本公开的药物组合物可含有一种或多种发挥生物作用的活性剂。合适的活性剂是本领域(例如,医师案头参考)中公知的。此类组合物可根据本领域中公知的程序制备,例如,如remington’s pharmaceutical sciences,mack publishing co.,easton,pa.,usa中所描述那样。
[0207]
莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g可以与本领域中已知的任何合适的牙科和口腔卫生组合物一起使用。合适的牙科和口腔卫生组合物的实例可以是,例如,牙膏、牙齿抛光剂(tooth polish)、牙线、漱口水、口腔清洗剂、洁齿剂(dentrifices)、口腔喷雾、口腔清新剂、牙菌斑清洗剂、牙镇痛药等。
[0208]
消耗品中存在的莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g的合适量可为,例如,约百万分之5(ppm)至约百万分之100(ppm)。在一些实施方案中,低浓度的莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g,例如,低于100ppm,具有相当于浓度介于10,000ppm至30,000ppm之间的蔗糖溶液的甜度。最终浓度范围为约5ppm至约100ppm、约5ppm至约95ppm、约5ppm至约90ppm、约5ppm至约85ppm、约5ppm至约80ppm、约5ppm至约75ppm、约5ppm至约70ppm、约5ppm至约65ppm、约5ppm至约60ppm、约5ppm至约55ppm、约5ppm至约50ppm、约5ppm至约45ppm、约5ppm至约40ppm、约5ppm至约35ppm、约5ppm至约30ppm、约5ppm至约25ppm、约5ppm至约20ppm、约5ppm至约15ppm或约5ppm至约10ppm。可选地,莱鲍迪苷v或莱鲍迪苷w可按以下范围的最终浓度存在于本公开的可消耗产品中:约5ppm至约100ppm、约10ppm至约100ppm、约15ppm至约100ppm、约20ppm至约100ppm、约25ppm至约100ppm、约30ppm至约100ppm、约35ppm至约100ppm、约40ppm至约100ppm、约45ppm至约100ppm、约50ppm至约100ppm、约55ppm至约100ppm、约60ppm至约100ppm、约65ppm至约100ppm、约70ppm至约100ppm、约75ppm至约100ppm、约80ppm至约100ppm、约85ppm至约100ppm、约90ppm至约100ppm或约95ppm至约100ppm。
[0209]
在某些实施方案中,在食物产品组合物中存在约5ppm至约100ppm莱鲍迪苷v、莱鲍
迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g。如本文中所用,“食物产品组合物”是指可以但不必具有营养价值且旨在用于人和动物消耗的任何固体或液体可摄入材料。
[0210]
合适的食物产品组合物的实例可以是,例如,糖食组合物,如糖果、薄荷、水果味硬糖、可可制品、巧克力等;调味品,如番茄酱、芥末、蛋黄酱等;口香糖;谷物组合物;烘焙物,如面包、蛋糕、馅饼、曲奇等;乳制品,例如牛奶、奶酪、奶油、冰淇淋、酸奶油、酸奶、冰冻果子露等;桌面甜味剂组合物;汤;炖菜;方便食品;肉,如火腿、培根、香肠、肉干等;明胶和明胶样产品如果酱、果冻、蜜饯等;水果;蔬菜;蛋制品;糖霜;糖浆,包括糖蜜;零食;坚果仁和坚果制品;以及动物饲料。
[0211]
食物产品组合物也可以是药草、香料和佐料、天然和合成的调料,以及增香剂,如谷氨酸一钠。在一些实施方案中,食物产品组合物可以是,例如,制备的包装产品,如饮食甜味剂、液体甜味剂、颗粒调味混合物、宠物食品、牲畜饲料、烟草,以及用于烘焙应用的材料,例如用于制备面包、曲奇、蛋糕、薄煎饼、甜甜圈等的粉状烘焙混合物。在其它实施方案中,食物产品组合物也可以是含有少量或不含蔗糖的减肥或低热量食品和饮料。
[0212]
在可以与任何前述实施方案组合的某些实施方案中,莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g可以是唯一的甜味剂,并且所述产品具有相当于约1%至约4%(w/v
‑
%)蔗糖溶液的甜味强度。在可以与任何前述实施方案组合的某些实施方案中,可消耗产品和饮料产品还可包括附加甜味剂,其中所述产品具有相当于约1%至约10%(w/v
‑
%)蔗糖溶液的甜味强度。在可以与任何前述实施方案组合的某些实施方案中,产品中的每种增甜成分均可为高强度甜味剂。在可以与任何前述实施方案组合的某些实施方案中,产品中的每种增甜成分均可为天然高强度甜味剂。在可以与任何前述实施方案组合的某些实施方案中,附加甜味剂含有选自以下的一种或多种甜味剂:甜叶菊提取物、甜菊醇糖苷、甜菊苷、莱鲍迪苷a、莱鲍迪苷b、莱鲍迪苷c、莱鲍迪苷d、莱鲍迪苷d3、莱鲍迪苷f、杜尔可苷a、甜茶苷、甜菊醇二糖苷、蔗糖、高果糖玉米糖浆、果糖、葡萄糖、木糖、阿拉伯糖、鼠李糖、赤藓糖醇、木糖醇、甘露糖醇、山梨糖醇、肌醇、acek、阿斯巴甜、纽甜、三氯蔗糖、糖精、柚皮苷二氢查耳酮(nardhc)、新橙皮苷二氢查耳酮(ndhc)、甜茶苷、罗汉果苷iv、赛门苷i、罗汉果苷v、莫那甜、索马甜、莫奈林、布拉奇因、l
‑
丙氨酸、甘氨酸、罗汉果、赫南德辛、叶甜素、三叶苷及其组合。在可以与任何前述实施方案组合的某些实施方案中,可消耗产品和饮料产品还可包括一种或多种选自以下的添加剂:碳水化合物、多元醇、氨基酸或其盐、聚氨基酸或其盐、糖酸或其盐、核苷酸、有机酸、无机酸、有机盐、有机酸式盐、有机碱式盐、无机盐、苦味化合物、食用香料、调味成分、涩味化合物、蛋白质、蛋白质水解物、表面活性剂、乳化剂、类黄酮、醇、聚合物及其组合。在可以与任何前述实施方案组合的某些实施方案中,莱鲍迪苷d3在将其添加到产品中之前具有按重量计约50%至约100%的纯度。
[0213]
甜味剂
[0214]
另一方面,本公开涉及一种由以下化学结构组成的甜味剂:
[0215][0216]
另一方面,本公开涉及一种由以下化学结构组成的甜味剂:
[0217][0218]
另一方面,本公开涉及一种由以下化学结构组成的甜味剂:
[0219][0220]
另一方面,本公开涉及一种由以下化学结构组成的甜味剂:
[0221][0222]
另一方面,本公开涉及一种由以下化学结构组成的甜味剂:
[0223][0224]
在某些实施方案中,甜味剂还可包括填料、膨胀剂和抗结块剂中的至少一种。合适的填料、膨胀剂和抗结块剂是本领域中已知的。
[0225]
在某些实施方案中,可按足以使可消耗产品和饮料产品变甜和/或增强其甜度的最终浓度包括和/或添加莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g甜味剂。莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g的“最终浓度”是指最终可消耗产品和饮料产品中(即,已经添加了所有成分和/或化合物而生产可消耗产品和饮料产品后)存在的莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g的浓度。因此,在某些实施方案中,将莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g包括和/或添加到用于制备可消耗产品和饮料产品的化合物或成分中。莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g可存在于单一化合物或成分,或多种化合物和成分中。在其它实施方案中,莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g可包括和/或添加到可消耗产品和饮料产品中。在某些优选实施方案中,按以下范围的最终浓度包括和/或添加莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g:约5ppm至约100ppm、约5ppm至约95ppm、约5ppm至约90ppm、约5ppm至约85ppm、约5ppm至约80ppm、约5ppm至约75ppm、约5ppm至约70ppm、约5ppm至约65ppm、约5ppm至约60ppm、约5ppm至约55ppm、约5ppm至约50ppm、约5ppm至约45ppm、约5ppm至约40ppm、约5ppm至约35ppm、约5ppm至约30ppm、约5ppm至约25ppm、约5ppm至约20ppm、约5ppm至约15ppm或约5ppm至约10ppm。可选地,按以下范围的最终浓度包括和/或添加莱鲍迪苷v或莱鲍迪苷w:约5ppm至约100ppm、约10ppm至约100ppm、约15ppm至约100ppm、约20ppm至约100ppm、约25ppm至约100ppm、约30ppm至约100ppm、约35ppm至约100ppm、约40ppm至约100ppm、约45ppm至约100ppm、约50ppm至约100ppm、约55ppm至约100ppm、约60ppm至约100ppm、约65ppm至约100ppm、约70ppm至约100ppm、约75ppm至约100ppm、约80ppm至约100ppm、约85ppm至约100ppm、约90ppm至约100ppm或约95ppm至约100ppm。例如,可按以下最终浓度包括和/或添加莱鲍迪苷v或莱鲍迪苷w:约5ppm、约10ppm、约15ppm、约20ppm、约25ppm、约30ppm、约35ppm、约40ppm、约45ppm、约
50ppm、约55ppm、约60ppm、约65ppm、约70ppm、约75ppm、约80ppm、约85ppm、约90ppm、约95ppm或约100ppm,包括介于这些值之间的任何范围。
[0226]
在某些实施方案中,莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g是包括和/或添加到可消耗产品和饮料产品中的唯一甜味剂。在此类实施方案中,可消耗产品和饮料产品具有相当于约1%至约4%(w/v
‑
%)蔗糖溶液、约1%至约3%(w/v
‑
%)蔗糖溶液或约1%至约2%(w/v
‑
%)蔗糖溶液的甜味强度。可选地,可消耗产品和饮料产品具有相当于约1%至约4%(w/v
‑
%)蔗糖溶液、约2%至约4%(w/v
‑
%)蔗糖溶液、约3%至约4%(w/v
‑
%)蔗糖溶液或约4%蔗糖溶液的甜味强度。例如,可消耗产品和饮料产品可具有相当于约1%、约2%、约3%或约4%(w/v
‑
%)蔗糖溶液,包括介于这些值之间的任何范围的甜味强度。
[0227]
本公开的可消耗产品和饮料产品可包括以足以达到所需甜味强度、营养特性、味道特性、口感或其它感官因素的比率存在的莱鲍迪苷v、莱鲍迪苷w、莱鲍迪苷ka、莱鲍迪苷m或莱鲍迪苷g和一种或多种本公开的甜味剂的混合物。
[0228]
在考虑了以下非限制性实施例后本公开将得到更全面的理解。
实施例
[0229]
实施例1
[0230]
在该实施例中,合成了所有候选ugt基因的全长dna片段。
[0231]
特别地,密码子优化cdna用于大肠杆菌(e.coli)表达(genscript,piscataway,nj)。将合成的dna克隆到细菌表达载体petite n
‑
his sumo kan vector(lucigen)中。对于编码udp
‑
糖基转移酶融合酶(ugt76g1
‑
atsus1和eugt11
‑
atsus1)的核苷酸序列而言,在介于来自阿拉伯芥(atsus1)的编码尿苷二磷酸糖基转移酶结构域的核苷酸序列和编码蔗糖合酶1的核苷酸序列之间的框内插入gsg
‑
接头(由核苷酸序列:ggttctggt编码)。表2总结了蛋白质和序列标识号。
[0232]
表2.序列标识号。
[0233]
[0234][0235]
将每个表达构建体转化到大肠杆菌bl21(de3)中,随后使其在37℃下在含有50μg/ml卡那霉素(kanamycin)的lb培养基中生长,直至达到0.8
‑
1.0的od
600
。通过添加1mm异丙基β
‑
d
‑1‑
硫代半乳糖吡喃糖苷(iptg)诱导蛋白质表达并且使培养物在16℃下进一步生长22小时。通过离心(3,000x g;10分钟;4℃)收获细胞。收集细胞团块并立即使用或储存在
‑
80℃下。
[0236]
使细胞团块重新悬浮在裂解缓冲液(50mm磷酸钾缓冲液(ph 7.2)、25μg/ml溶菌酶、5μg/ml dna酶i、20mm咪唑、500mm nacl、10%甘油和0.4%triton x
‑
100)中。在4℃下通过超声处理破坏细胞,并且通过离心(18,000x g;30分钟)使细胞碎片澄清。将上清液加载至平衡的(平衡缓冲液:50mm磷酸钾缓冲液(ph 7.2)、20mm咪唑、500mm nacl、10%甘油)ni
‑
nta(qiagen)亲和柱上。加载蛋白质样品之后,用平衡缓冲液洗涤该柱以去除未结合的污染蛋白质。用含有250mm咪唑的平衡缓冲液洗脱his标记的ugt重组多肽。图2中示出了纯化hv1(61.4kd)、ugt76g1(65.4kd)、atsus1(106.3kd)、eugt11(62kd)、ugt76g1
‑
sus1(gs)(157.25kd)和eugt11
‑
atsus1(155kd)融合蛋白。
[0237]
实施例2
[0238]
在该实施例中,通过使用受试甜菊醇糖苷作为底物测定候选ugt重组多肽的糖基转移酶活性。
[0239]
通常,以200μl在体外反应体系中测试重组多肽(10μg)。该反应体系含50mm磷酸钾缓冲液(ph 7.2)、3mm mgcl2、1mg/ml甜菊醇糖苷底物、1mm udp
‑
葡萄糖。反应在30℃下进行并通过添加200μl 1
‑
丁醇而终止。用200μl 1
‑
丁醇萃取样品三次。汇合级分经干燥并溶于70μl 80%甲醇中以进行高效液相色谱(hplc)分析。使用甜茶苷(99%,blue california,ca)、纯化的reb g(98.8%)、reb ka(98.4%)和reb v(80%)在体外反应中用作底物。
[0240]
ugt催化的糖基化反应与蔗糖合酶(如atsus1)催化的udp
‑
葡萄糖生成反应联合。在这种方法中,由蔗糖和udp生成udp
‑
葡萄糖,使得可以省略添加额外的udp
‑
葡萄糖。在所述测定中,在ugt反应体系中添加重组atsus1并且由udp再生udp
‑
葡萄糖。合成atsus1序列(bieniawska等,plant j.2007,49:810
‑
828)并插入细菌表达载体中。表达重组atsus1蛋白并通过亲和色谱法纯化。
[0241]
使用dionex uplc ultimate 3000系统(sunnyvale,ca)进行hplc分析,所述系统包括四元泵、温控柱室、自动取样器和紫外吸光度检测器。使用phenomenex lunanh2、luna c18或synergi hydro
‑
rp与保护柱进行甜菊醇糖苷表征。于水中或于na3po4缓冲液中的乙腈用于hplc分析中的等度洗脱。检测波长为210nm。
[0242]
实施例3
[0243]
在该实施例中,分析重组hv1多肽,在有或无atsus1的所有反应条件下将糖部分转移至甜茶苷以生成莱鲍迪苷ka(“minor diterpene glycosides from the leaves of stevia rebaudiana”.journal of natural products(2014),77(5),1231
‑
1235)。
[0244]
如图3所示,在有或无atsus1的所有反应条件下,重组hv1多肽将糖部分转移至甜
茶苷以生成reb ka。在ugt
‑
sus偶联反应体系(g、i)中甜茶苷被重组hv1完全转化为reb ka和reb e。然而,24小时后(h)仅部分甜茶苷被未偶联至atsus1的单独的重组hv1多肽转化为reb ka,表明atsus1提高了ugt
‑
sus偶联体系中的转化效率。在hv1
‑
atsus1偶联反应体系中,生成的reb ka可连续地转化为reb e。
[0245]
实施例4
[0246]
在该实施例中,使用reb e作为底物分析hv1活性。
[0247]
reb e底物(0.5mg/ml)与重组hv1多肽(20μg)和atsus1(20μg)一起在ugt
‑
sus偶联反应体系(200μl)中在类似于以上实施例中所用的条件下温育。如图4所示,由重组hv1多肽和atsus1的组合生成reb z。这些结果表明hv1可以将葡萄糖部分转移至reb e以形成rz。图4显示可由莱鲍迪苷e(“reb e”),经重组hv1多肽和重组atsus1催化在hv1
‑
atsus1偶联反应体系中生成莱鲍迪苷z(“reb z”)。hv1可以将葡萄糖转移至reb e以生成reb z,比率介于60:40与70:30之间的reb z1和reb z2的混合物(转让给conagen inc.的美国临时申请第61/898,571号)。
[0248]
实施例5
[0249]
在该实施例中,为确认reb ka转化为reb e,将纯化reb ka底物与重组hv1一起温育,有或无atsus1。如图5所示,在两种反应条件下由重组hv1多肽生成reb e。然而,ugt
‑
sus偶联反应体系中的atsus1多肽可以提高反应效率。在ugt
‑
sus偶联体系(d)中所有reb ka底物可完全转化为reb e。
[0250]
实施例6
[0251]
在该实施例中,使用甜茶苷作为底物分析eugt11活性。
[0252]
如图6所示,在有或无atsus1的所有反应条件下eugt11均可以将糖部分转移至甜茶苷以生成reb ka和甜菊苷。在ugt
‑
sus偶联体系中atsus1提高了转化效率。在hv1
‑
atsus1偶联反应体系中,reb e可以被eugt11连续转化。eus融合蛋白在相同反应条件下表现出更高活性。在48小时所有生成的reb ka和甜菊苷被eus完全转化为reb e。reb e可以连续转化为reb d3。
[0253]
实施例7
[0254]
在该实施例中,使用reb ka作为底物分析eugt11活性。
[0255]
eugt11是具有生成有关甜菊醇糖苷的1,2
‑
19
‑
o
‑
葡萄糖糖基化活性的ugt(转让给evolva sa的pct公布申请wo2013/022989)。例如,eugt11可以催化由甜菊苷生成reb e的反应。eugt11还具有可以将葡萄糖分子转移至莱鲍迪苷e以形成莱鲍迪苷d3的1,6
‑
13
‑
o
‑
葡萄糖糖基化活性(转让给conagen,inc.的美国专利申请序列号14/269,435)。在实验中,我们发现eugt11可以将葡萄糖残基转移至reb ka以形成reb e。如图7所示,在有atsus1(e、h)或无atsus1(d、g)的所有反应条件下,eugt11可以将糖部分转移至reb ka以生成reb e。atsus1提高了ugt
‑
sus偶联体系(e、h)中的转化效率。在eugt11
‑
atsus1偶联反应体系(e、h)和eus融合反应体系(f、i)中,所有reb ka完全转化并且生成的reb e可以连续转化为reb d3。
[0256]
实施例8
[0257]
在该实施例中,使用甜茶苷作为底物分析ugt76g1活性。
[0258]
ugt76g1具有可以将葡萄糖分子转移至甜菊苷以形成莱鲍迪苷a和转移至reb e以
形成莱鲍迪苷d的1,3
‑
13
‑
o
‑
葡萄糖糖基化活性。在该实施例中,我们发现ugt76g1可以将葡萄糖残基转移至甜茶苷以形成莱鲍迪苷g。
[0259]
如图8所示,在有atsus1(d、g)或无atsus1(c、f)的所有反应条件下,ugt76g1可以将糖部分转移至甜茶苷以生成reb g。atsus1提高了ugt
‑
sus偶联体系中的转化效率。gs融合蛋白在相同反应条件(e、h)下表现出更高活性。在12小时所有甜茶苷完全转化为reb g(e)。
[0260]
实施例9
[0261]
在该实施例中,使用莱鲍迪苷ka作为底物分析ugt76g1活性。
[0262]
为进一步鉴定ugt76g1的酶活性,使用莱鲍迪苷ka作为底物进行体外测定。令人惊讶地,在早期时间点生成新型甜菊醇糖苷(莱鲍迪苷v“reb v”)。在以后的时间点,反应中生成的reb v转化为另一种新型甜菊醇糖苷(莱鲍迪苷w“reb w”)。
[0263]
如图9所示,在有atsus1(f、i)或无atsus1(e、h)的所有反应条件下,ugt76g1可以将糖部分转移至reb ka以生成reb v。atsus1提高了ugt
‑
sus偶联体系(f、i)中的转化效率。在ugt76g1
‑
atsus1偶联反应体系(i)和gs融合反应体系(j)中,在12小时生成的reb v完全转化为reb w。
[0264]
实施例10
[0265]
在该实施例中,使用reb v作为底物分析ugt76g1活性。
[0266]
将作为底物的纯化reb v引入到反应体系中。如图10c所示,在6小时在ugt
‑
sus1偶联体系中reb v惊人地被ugt76g1重组多肽完全转化为reb w。
[0267]
实施例11
[0268]
在该实施例中,使用reb g作为底物分析hv1活性。
[0269]
如图11所示,在有或无atsus1的所有反应条件下,重组hv1多肽将糖部分转移至莱鲍迪苷g以生成reb v。在ugt
‑
sus偶联反应体系(e、g)中reb g被重组hv1完全转化为reb v。然而,24小时后(f)仅部分reb g被未偶联至atsus1的单独的重组hv1多肽转化为reb v,表明atsus1提高了ugt
‑
sus偶联体系中的转化效率。
[0270]
实施例12
[0271]
在该实施例中,使用reb g作为底物分析eugt11活性。
[0272]
如图12所示,在有atsus1(f、i)或无atsus1(e、h)的所有反应条件下,重组eugt11多肽将糖部分转移至莱鲍迪苷g以生成reb v。在ugt
‑
sus偶联反应体系(f、i)中更多的reb g被重组eugt11转化为reb v。然而,仅部分reb g被未偶联至atsus1的单独的重组eugt11多肽转化为reb v(e、h),表明atsus1提高了ugt
‑
sus偶联体系中的转化效率。eus融合蛋白在相同反应条件下表现出更高活性(g、j)。24小时(j)该反应体系中的所有reb g被eus完全转化为reb v。
[0273]
实施例13
[0274]
在该实施例中,使用甜茶苷作为底物分析hv1与ugt76g1组合的活性。
[0275]
甜茶苷底物与重组hv1多肽、ugt76g1和atsus1一起在ugt
‑
sus偶联反应体系中在类似于以上实施例中所用的条件下温育。通过hplc分析产物。如图13所示,由重组hv1多肽、ugt76g1和atsus1的组合生成reb v和reb w。因此,显示出1,2
‑
19
‑
o
‑
葡萄糖和1,2
‑
13
‑
o
‑
葡萄糖糖基化活性的重组hv1多肽,可以与其它ugt酶(如ugt76g1,其显示出1,3
‑
13
‑
o
‑
葡萄糖
和1,3
‑
19
‑
o
‑
葡萄糖糖基化活性)组合使用,用于甜菊醇糖苷的复杂、多步生物合成。如果hv1重组蛋白与gs融合蛋白在反应体系中组合,也由这些ugt酶生成reb v和reb w,表明可由gs融合蛋白产生ugt
‑
sus偶联反应。
[0276]
实施例14
[0277]
在该实施例中,使用甜茶苷作为底物分析eugt11与ugt76g1组合的活性。
[0278]
甜茶苷底物与重组eugt11多肽、ugt76g1和atsus1一起在ugt
‑
sus偶联反应体系中在类似于以上实施例中所用的条件下温育。通过hplc分析产物。如图14所示,由重组eugt11多肽、ugt76g1和atsus1的组合生成reb w。因此,显示出1,2
‑
19
‑
o
‑
葡萄糖和1,2
‑
13
‑
o
‑
葡萄糖糖基化活性的重组eugt11多肽,可以与其它ugt酶(如ugt76g1,其显示出1,3
‑
13
‑
o
‑
葡萄糖和1,3
‑
19
‑
o
‑
葡萄糖糖基化活性)组合使用,用于甜菊醇糖苷的复杂、多步生物合成。如果eugt11重组蛋白与gs融合蛋白在反应体系中组合,也由这些ugt酶生成reb w,表明可由gs融合蛋白产生ugt
‑
sus偶联反应。
[0279]
实施例15
[0280]
在该实施例中,使用reb g作为底物分析hv1与ugt76g1组合的活性。
[0281]
reb g底物与重组hv1多肽、ugt76g1和atsus1一起在ugt
‑
sus偶联反应体系中在类似于以上实施例中所用的条件下温育。通过hplc分析产物。如图15所示,由重组hv1多肽、ugt76g1和atsus1的组合生成reb v和reb w。在12小时后,所有甜茶苷底物转化为reb v,并且在36小时后,所有生成的reb v转化为reb w。因此,显示出1,2
‑
19
‑
o
‑
葡萄糖和1,2
‑
13
‑
o
‑
葡萄糖糖基化活性的重组hv1多肽,可以与其它ugt酶(如ugt76g1,其显示出1,3
‑
13
‑
o
‑
葡萄糖和1,3
‑
19
‑
o
‑
葡萄糖糖基化活性)组合使用,用于甜菊醇糖苷的复杂、多步生物合成。如果hv1重组蛋白与gs融合蛋白在反应体系中组合,也由这些ugt酶生成reb v和reb w,表明可由gs融合蛋白产生ugt
‑
sus偶联反应。
[0282]
实施例16
[0283]
在该实施例中,使用reb g作为底物分析eugt11与ugt76g1组合的活性。
[0284]
reb g底物与重组eugt11多肽、ugt76g1和atsus1一起在ugt
‑
sus偶联反应体系中在类似于以上实施例中所用的条件下温育。通过hplc分析产物。如图16所示,由重组eugt11多肽、ugt76g1和atsus1的组合生成和reb w。因此,显示出1,2
‑
19
‑
o
‑
葡萄糖和1,2
‑
13
‑
o
‑
葡萄糖糖基化活性的重组eugt11多肽,可以与其它ugt酶(如ugt76g1,其显示出1,3
‑
13
‑
o
‑
葡萄糖和1,3
‑
19
‑
o
‑
葡萄糖糖基化活性)组合使用,用于甜菊醇糖苷的复杂、多步生物合成。如果eugt11重组蛋白与gs融合蛋白在反应体系中组合,也由这些ugt酶生成reb w,表明可由gs融合蛋白产生ugt
‑
sus偶联反应。
[0285]
实施例17
[0286]
在该实施例中,使用reb d作为底物分析ugt76g1和gs融合酶活性。
[0287]
reb d底物与重组ugt76g1一起在类似于以上实施例中所用的条件下温育。通过hplc分析产物。如图22所示,在反应中由有atsus1(图22d和g)或无atsus1(图22c和f)的ugt76g1生成reb m。因此,显示出1,3
‑
19
‑
o
‑
葡萄糖糖基化活性的重组ugt76g1多肽,可用于莱鲍迪苷m的生物合成。在ugt
‑
sus偶联反应体系(图22g)中,reb d被重组ugt76g1完全转化为reb m。然而,6小时后(f)仅部分reb d被未偶联至atsus1的单独的重组ugt76g1多肽转化为reb m,表明atsus1提高了ugt
‑
sus偶联体系中的转化效率。gs融合蛋白在相同反应条件
下(e、h)表现出与ugt76g1
‑
atsus1偶联反应相似的活性。在6小时(h)所有reb d被gs完全转化为reb m,表明可由gs融合蛋白产生ugt
‑
sus偶联反应。
[0288]
实施例18
[0289]
在该实施例中,使用reb e作为底物分析ugt76g1和gs融合酶活性。
[0290]
reb e底物与重组ugt76g1或gs融合酶一起在类似于以上实施例中所用的条件下温育。通过hplc分析产物。如图23所示,在反应中由有atsus1(图23e、h和k)或无atsus1(图22d、g和j)的ugt76g1和gs融合酶(图23f、i和l)生成reb d。此外,由反应中生成的reb d形成reb m。因此,显示出1,3
‑
13
‑
o
‑
葡萄糖和1,3
‑
19
‑
o
‑
葡萄糖糖基化活性的重组ugt76g1多肽,可用于莱鲍迪苷d和莱鲍迪苷m的生物合成。24小时后在ugt
‑
sus偶联反应体系中,reb e被重组ugt76g1完全转化为reb m(图23k)。然而,24小时后(j)仅reb d被未偶联至atsus1的单独的重组ugt76g1多肽从reb e转化,表明atsus1通过连续的udpg生成而提高了ugt
‑
sus偶联体系中的转化效率。gs融合蛋白在相同反应条件下(图23f、i和l)表现出与ugt76g1
‑
atsus1偶联反应相似的活性,表明可由gs融合蛋白产生ugt
‑
sus偶联反应。
[0291]
实施例19
[0292]
在该实施例中,使用甜菊苷作为底物分析hv1与ugt76g1组合的活性。
[0293]
甜茶苷底物与重组hv1多肽和ugt76g1或gs融合酶一起在类似于以上实施例中所用的条件下温育。通过hplc分析产物。如图24所示,在所有反应中由重组hv1多肽和ugt76g1的组合生成reb a。此外,在反应中使用重组hv1多肽、ugt76g1多肽和atsus1的组合(图24e、h和k)或重组gs融合酶和hv1多肽的组合(图24f、i和l)检测到reb d和reb m。显示出1,2
‑
19
‑
o
‑
葡萄糖糖基化活性的重组hv1多肽,可以与其它ugt酶(如ugt76g1,其显示出1,3
‑
13
‑
o
‑
葡萄糖和1,3
‑
19
‑
o
‑
葡萄糖糖基化活性)组合使用,用于莱鲍迪苷d和莱鲍迪苷m的复杂、多步生物合成。结果还显示atsus1通过连续的udpg生成提高了ugt
‑
sus偶联体系中的转化效率(图24e、h和k)。gs融合蛋白在相同反应条件下(图24f、i和l)表现出与ugt76g1
‑
atsus1偶联反应相似的活性,表明可由gs融合蛋白产生ugt
‑
sus偶联反应。
[0294]
实施例20
[0295]
在该实施例中,使用reb a作为底物分析hv1与ugt76g1组合的活性。
[0296]
reba底物与重组hv1多肽和ugt76g1或gs融合酶一起在类似于以上实施例中所用的条件下温育。通过hplc分析产物。如图25所示,在所有反应中由重组hv1多肽和ugt76g1的组合生成reb d。此外,在反应中使用重组hv1多肽、ugt76g1多肽和atsus1的组合(图25d、g和j)或重组gs融合酶和hv1多肽的组合(图25e、h和k)检测到reb m。显示出1,2
‑
19
‑
o
‑
葡萄糖糖基化活性的重组hv1多肽,可以与其它ugt酶(如ugt76g1,其显示出1,3
‑
19
‑
o
‑
葡萄糖糖基化活性)组合使用,用于莱鲍迪苷d和莱鲍迪苷m的复杂、多步生物合成。结果还显示atsus1通过连续的udpg生成提高了ugt
‑
sus偶联体系中的转化效率(图25d、g和j)。gs融合蛋白在相同反应条件下(图25e、h和k)表现出与ugt76g1
‑
atsus1偶联反应相似的活性,表明可由gs融合蛋白产生ugt
‑
sus偶联反应。
[0297]
实施例21
[0298]
在该实施例中,通过nmr分析reb v的结构。
[0299]
用于reb v表征的材料通过使用reb g的酶促转化生成并通过hplc纯化。用将其分辨率设为30k的ltq orbitrap discoveryhrms仪器生成hrms数据。在正离子电喷射模式下
扫描数据从m/z 150至1500。针电压设为4kv;其它源条件为鞘气=25,辅助气=0,吹扫气=5(为任意单位的所有气流),毛细管电压=30v,毛细管温度=300℃和管透镜电压=75。样品用2:2:1乙腈:甲醇:水(与输注洗脱液相同)稀释并注射50微升。在brukeravance drx 500mhz或varian inova600mhz仪器上使用标准脉冲序列获得nmr波谱。1d(1h和
13
c)和2d(tocsy、hmqc和hmbc)nmr波谱在c5d5n中进行。
[0300]
已基于其在m/z 989.4198处显示出对应于[m na]
的加合离子的正电高分辨率(hr)质谱将reb v的分子式推导为c
44
h
70
o
23
;这种组成受
13
c nmr波谱数据支持。reb v的1h nmr波谱数据显示在δ0.97和1.40存在两个甲基单线态,在环外双键的δ5.06和5.71存在两个为单线态的烯烃质子,在δ0.74
‑
2.72存在九个sp3亚甲基和两个sp3次甲基质子,这是早期分离自甜叶菊属的对映
‑
贝壳杉烷二萜的特点。对映
‑
贝壳杉烷二萜的基本骨架受到cosy和tocsy研究支持,该研究显示出关键相关性:h
‑
1/h
‑
2;h
‑
2/h
‑
3;h
‑
5/h
‑
6;h
‑
6/h
‑
7;h
‑
9/h
‑
11;h
‑
11/h
‑
12。reb v的1h nmr波谱还显示存在四个在δ5.08、5.38、5.57和6.23共振的异头质子;表明在其结构中存在四个糖单元。用5%h2so4酸水解reb v提供了通过tlc与真实样品直接比较所鉴定的d
‑
葡萄糖。酶水解reb v供给了通过与标准化合物的1h nmr和co
‑
tlc的比较鉴定为甜菊醇的糖苷配基。在δ5.08(d,j=7.8hz)、5.38(d,j=8.1hz)、5.57(d,j=8.0hz)和6.23(d,j=7.8hz)对于葡萄糖部分的四个异头质子所观察到的大偶联常数,表明其β
‑
取向与对甜菊醇糖苷所报告的相同。基于tocsy、hmqc和hmbc数据分配reb v的1h和
13
c nmr值并且于表3中给出。
[0301]
表3.reb v和reb g的1h和
13
c nmr波谱数据(化学位移和偶联常数)
a
‑
c
[0302]
[0303]
[0304][0305]
a
基于tocsy、hmqc和hmbc相关性进行的分配;
b
化学位移值按δ(ppm)计;
c
偶联常数按hz计。
[0306]
基于来自reb v的nmr波谱数据和水解实验的结果,推断在其结构中存在四个与糖苷配基甜菊醇连接的β
‑
d
‑
葡糖基单元。对reb v与reb g的1h和
13
c nmr值的严密比较表明存在在c
‑
13处具有呈醚键形式的3
‑
o
‑
β
‑
d
‑
吡喃葡糖基单元且在c
‑
19位置处具有另一个呈酯键形式的β
‑
d
‑
葡糖基单元的甜菊醇糖苷配基,剩下第四个β
‑
d
‑
葡糖基部分的分配(图17)。在β
‑
d
‑
葡糖基部分的糖i的2
‑
位的1h和
13
c化学位移的低场位移支持在该位置存在β
‑
d
‑
葡糖基单元。该结构还受到如图18所示的关键tocsy和hmbc相关性的支持。基于nmr和质谱数据以及水解研究的结果,将通过reb g的酶促转化生成的reb v的结构推导为13
‑
[(3
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)氧基]对映
‑
贝壳杉
‑
16
‑
烯
‑
19
‑
烯酸
‑
(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)酯。
[0307]
reb v的酸水解。向reb v(5mg)于meoh(10ml)中的溶液添加3ml的5%h2so4并且使混合物回流24小时。然后用饱和碳酸钠中和反应混合物并用乙酸乙酯(etoac)(2x 25ml)萃取以得到含糖的水性级分及含糖苷配基部分的etoac级分。浓缩水相并使用tlc体系etoac/正丁醇/水(2:7:1)和ch2cl2/meoh/水(10:6:1)与标准糖进行比较;将糖鉴定为d
‑
葡萄糖。
[0308]
reb v的酶水解。将reb v(1mg)溶于10ml的0.1m乙酸钠缓冲液(ph 4.5)中并添加来自黑曲霉(aspergillus niger)(50ul,sigma
‑
aldrich,p2736)的粗果胶酶。在50℃下搅拌混合物96小时。反应期间由1的水解沉淀出来的产物通过比较其与标准化合物的co
‑
tlc和1hnmr波谱数据被鉴定为甜菊醇。基于大量的1d和2d nmr以及高分辨率质谱数据和水解研究,确认命名为reb v的化合物为13
‑
[(3
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)氧基]对映
‑
贝壳杉
‑
16
‑
烯
‑
19
‑
烯酸
‑
(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)酯。
[0309]
实施例22
[0310]
在该实施例中,通过nmr分析reb w的结构。
[0311]
用于reb w表征的材料通过使用reb v的酶促转化生成并通过hplc纯化。用将其分辨率设为30k的ltq orbitrap discoveryhrms仪器生成hrms数据。在正离子电喷射模式下扫描数据从m/z 150至1500。针电压设为4kv;其它源条件为鞘气=25,辅助气=0,吹扫气=5(为任意单位的所有气流),毛细管电压=30v,毛细管温度=300c和管透镜电压=75。样品用2:2:1乙腈:甲醇:水(与输注洗脱液相同)稀释并注射50微升。在brukeravance drx 500mhz或varian inova600mhz仪器上使用标准脉冲序列获得nmr波谱。1d(1h和
13
c)和2d(tocsy、hmqc和hmbc)nmr波谱在c5d5n中进行。
[0312]
已基于其在m/z 1151.4708处显示出对应于[m na]
的加合离子的正电高分辨率(hr)质谱将reb w的分子式推导为c
50
h
80
o
28
;这种组成受
13
c nmr波谱数据支持。reb w的1h nmr波谱数据显示在δ0.92和1.39存在两个甲基单线态,在环外双键的δ5.10和5.73存在两个为单线态的烯烃质子,在δ0.72
‑
2.72存在九个sp3亚甲基和两个sp3次甲基质子,这是早期分离自甜叶菊属的对映
‑
贝壳杉烷二萜的特点。对映
‑
贝壳杉烷二萜的基本骨架受到tocsy研究支持,该研究显示出关键相关性:h
‑
1/h
‑
2;h
‑
2/h
‑
3;h
‑
5/h
‑
6;h
‑
6/h
‑
7;h
‑
9/h
‑
11;h
‑
11/h
‑
12。reb w的1hnmr波谱还显示存在五个在δ5.10、5.34、5.41、5.81和6.14共振的
异头质子;表明在其结构中存在五个糖单元。用5%h2so4酸水解reb w提供了通过tlc与真实样品直接比较所鉴定的d
‑
葡萄糖。酶水解reb w供给了通过与标准化合物的1hnmr和co
‑
tlc的比较鉴定为甜菊醇的糖苷配基。在δ5.10(d,j=7.4hz)、5.34(d,j=7.9hz)、5.41(d,j=7.9hz)、5.89(d,j=7.9hz)和6.14(d,j=7.9hz)对于葡萄糖部分的五个异头质子所观察到的大偶联常数,表明其β
‑
取向与对甜菊醇糖苷所报告的相同[1
‑
5,9
‑
13]。基于tocsy、hmqc和hmbc数据分配reb w的1h和
13
c nmr值并且于表4中给出。
[0313]
表4.reb w和reb v的1h和
13
c nmr波谱数据(化学位移和偶联常数)
a
‑
c
[0314]
[0315][0316]
a
基于tocsy、hmqc和hmbc相关性进行的分配;
b
化学位移值按δ(ppm)计;
c
偶联常数按hz计。
[0317]
基于来自reb w的nmr波谱数据和水解实验的结果,推断在其结构中存在五个与糖苷配基甜菊醇连接的β
‑
d
‑
葡糖基单元。对reb w与reb v的1h和
13
c nmr值的严密比较表明存在在c
‑
13处具有呈醚键形式的3
‑
o
‑
β
‑
d
‑
吡喃葡糖基单元且在c
‑
19位置处具有呈酯键形式的2
‑
o
‑
β
‑
d
‑
吡喃葡糖基单元的甜菊醇糖苷配基,剩下第五个β
‑
d
‑
葡糖基部分的分配(图19)。在β
‑
d
‑
葡糖基部分的糖i的3
‑
位的1h和
13
c化学位移的低场位移支持在该位置存在β
‑
d
‑
葡糖基单元。该结构还受到如图20所示的关键tocsy和hmbc相关性的支持。基于nmr和质谱数据以及水解研究的结果,将通过reb v的酶促转化生成的reb w的结构推导为13
‑
[(3
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)氧基]对映
‑
贝壳杉
‑
16
‑
烯
‑
19
‑
烯酸
‑
[(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑3‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)酯。
[0318]
reb w的酸水解。向reb w(5mg)于meoh(10ml)中的溶液添加3ml的5%h2so4并且使混合物回流24小时。然后用饱和碳酸钠中和反应混合物并用乙酸乙酯(etoac)(2x 25ml)萃取以得到含糖的水性级分及含糖苷配基部分的etoac级分。浓缩水相并使用tlc体系etoac/正丁醇/水(2:7:1)和ch2cl2/meoh/水(10:6:1)与标准糖进行比较;将糖鉴定为d
‑
葡萄糖。
[0319]
reb w的酶水解。将reb w(1mg)溶于10ml的0.1m乙酸钠缓冲液(ph 4.5)中并添加来自黑曲霉(50ul,sigma
‑
aldrich,p2736)的粗果胶酶。在50℃下搅拌混合物96小时。产物在反应期间沉淀出来并且经过滤,然后结晶。从reb w水解获得的所得产物通过比较其与标准化合物的co
‑
tlc和1hnmr波谱数据被鉴定为甜菊醇。基于大量的1d和2d nmr以及高分辨率质谱数据和水解研究,确认命名为reb w的化合物为13
‑
[(3
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)氧基]对映
‑
贝壳杉
‑
16
‑
烯
‑
19
‑
烯酸
‑
[(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑3‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)酯。
[0320]
nmr分析之后,将reb v和reb w的结构鉴定为新型甜菊醇糖苷。以上结果进一步证明,ugt76g1不但具有1,3
‑
13
‑
o
‑
葡萄糖糖基化活性,而且具有1,3
‑
19
‑
o
‑
葡萄糖糖基化活性。
[0321]
实施例23
[0322]
在该实施例中,通过nmr分析reb m的结构。
[0323]
用于reb m表征的材料通过使用reb d的酶促转化生成并通过hplc纯化。用将其分辨率设为30k的ltq orbitrap discoveryhrms仪器生成hrms数据。在正离子电喷射模式下扫描数据从m/z 150至1500。针电压设为4kv;其它源条件为鞘气=25,辅助气=0,吹扫气=5(为任意单位的所有气流),毛细管电压=30v,毛细管温度=300c和管透镜电压=75。样品用2:2:1乙腈:甲醇:水(与输注洗脱液相同)稀释并注射50微升。
[0324]
在bruker avance drx 500mhz或varian inova 600mhz仪器上使用标准脉冲序列获得nmr波谱。1d(1h和
13
c)和2d(tocsy、hmqc和hmbc)nmr波谱在c5d5n中进行。
[0325]
已基于其在m/z 1349.5964处显示出[m nh4 ch3cn]
离子的正电高分辨率(hr)质谱将reb m的分子式推导为c
56
h
90
o
33
;这种组成受
13
c nmr波谱数据支持。reb m的1h nmr波谱显示在δ1.35和1.42存在两个甲基单线态,在环外双键的δ4.92和5.65存在两个为单线态的烯烃质子,在δ0.77
‑
2.77存在九个亚甲基和两个次甲基质子,这是早期分离自甜叶菊属的对映
‑
贝壳杉烷二萜的特点。对映
‑
贝壳杉烷二萜的基本骨架受到cosy(h
‑
1/h
‑
2;h
‑
2/h
‑
3;h
‑
5/h
‑
6;h
‑
6/h
‑
7;h
‑
9/h
‑
11;h
‑
11/h
‑
12)和hmbc(h
‑
1/c
‑
2、c
‑
10;h
‑
3/c
‑
1、c
‑
2、c
‑
4、c
‑
5、c
‑
18、c
‑
19;h
‑
5/c
‑
4、c
‑
6、c
‑
7、c
‑
9、c
‑
10、c
‑
18、c
‑
19、c
‑
20;h
‑
9/c
‑
8、c
‑
10、c
‑
11、c
‑
12、c
‑
14、c
‑
15;h
‑
14/c
‑
8、c
‑
9、c
‑
13、c
‑
15、c
‑
16和h
‑
17/c
‑
13、c
‑
15、c
‑
16)相关性的支持。reb m的1hnmr波谱还显示存在在δ5.33、5.47、5.50、5.52、5.85和6.43共振的异头质子;表明在其结构中存在六个糖单元。酶水解reb m供给了通过与标准化合物的co
‑
tlc的比较鉴定为甜菊醇的糖苷配基。用5%h2so4酸水解reb m提供了通过tlc与真实样品直接比较所鉴定的葡萄糖。基于tocsy、hmqc和hmbc相关性分配reb m中选定的质子和碳的1h和
13
c nmr值(表5)。
[0326]
基于来自reb m的nmr波谱数据的结果,推断在其结构中存在六个葡糖基单元(图26)。对reb m与莱鲍迪苷d的1h和
13
c nmr波谱的严密比较表明reb m也是甜菊醇糖苷,其具有三个附连在c
‑
13羟基处作为2,3
‑
分支吡喃葡糖基取代基的葡萄糖残基和在c
‑
19处呈酯形式的2
‑
取代的吡喃葡糖基部分,剩下另一个葡糖基部分的分配。图27所示的关键tocsy和hmbc相关性表明第六个葡糖基部分置于糖i的c
‑
3位置。在δ5.33(d,j=8.4hz)、5.47(d,j=7.8hz)、5.50(d,j=7.4hz)、5.52(d,j=7.4hz)、5.85(d,j=7.4hz)和6.43(d,j=7.8hz)对于葡萄糖部分的六个异头质子所观察到的大偶联常数,表明其β
‑
取向与对甜菊醇糖苷所报告的相同。基于nmr和质谱研究的结果及在与从文献中报告的莱鲍迪苷m的波谱值的比较中,将通过酶促反应生成的reb m的结构指定为13
‑
[(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑3‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)氧基]对映
‑
贝壳杉
‑
16
‑
烯
‑
19
‑
烯酸
‑
[(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑3‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)酯。
[0327]
表5.通过酶促反应生成的reb m的1h和
13
c nmr波谱数据(化学位移和偶联常数)
a
‑
c
。
[0328]
[0329][0330]
[0331]
a
基于tocsy、hsqc和hmbc相关性进行的分配;
b
化学位移值按δ(ppm)计;
c
偶联常数按hz计。
[0332]
化合物1的酸水解:向生成的reb m(5mg)于meoh(10ml)中的溶液添加3ml的5%h2so4并且使混合物回流24小时。然后用饱和碳酸钠中和反应混合物并用乙酸乙酯(etoac)(2x 25ml)萃取以得到含糖的水性级分及含糖苷配基部分的etoac级分。浓缩水相并使用tlc体系etoac/正丁醇/水(2:7:1)和ch2cl2/meoh/水(10:6:1)与标准糖进行比较;将糖鉴定为d
‑
葡萄糖。
[0333]
化合物的酶水解:将生成的reb m(1mg)溶于10ml的0.1m乙酸钠缓冲液(ph 4.5)中并添加来自黑曲霉(50ul,sigma
‑
aldrich,p2736)的粗果胶酶。在50℃下搅拌混合物96小时。反应期间由1的水解沉淀出来的产物通过比较其与标准化合物的co
‑
tlc和1hnmr波谱数据被鉴定为甜菊醇。
[0334]
获得的命名为莱鲍迪苷m(reb m)的化合物通过生物转化而生成。对莱鲍迪苷m(reb m)的完全1h和
13
c nmr波谱分配基于大量的1d和2d nmr以及高分辨率质谱数据进行,其表明结构为13
‑
[(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑3‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)氧基]对映
‑
贝壳杉
‑
16
‑
烯
‑
19
‑
烯酸
‑
[(2
‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑3‑
o
‑
β
‑
d
‑
吡喃葡糖基
‑
β
‑
d
‑
吡喃葡糖基)酯。
[0335]
实施例24
[0336]
在该实施例中,讨论甜菊醇糖苷的生物合成途径。
[0337]
图21是说明由甜茶苷生物合成甜菊醇糖苷的新型途径的图解。如本文所述,重组hv1多肽(“hv1”)含有将第二葡糖苷部分转移至甜茶苷19
‑
o
‑
葡萄糖的c
‑2’
以生成莱鲍迪苷ka(“reb ka”)的1,2
‑
o
‑
葡萄糖糖基化活性;重组eugt11多肽(“eugt11”)含有将第二葡萄糖部分转移至甜茶苷19
‑
o
‑
葡萄糖的c
‑2’
以生成莱鲍迪苷ka;或将第二葡萄糖部分转移至甜茶苷的13
‑
o
‑
葡萄糖的c
‑2’
以生成甜菊苷的1,2
‑
o
‑
葡萄糖糖基化活性;重组ugt76g1酶(“ugt76g1”)含有将第二葡萄糖部分转移至甜茶苷的13
‑
o
‑
葡萄糖的c
‑3’
以生成莱鲍迪苷g(“reb g”)的1,3
‑
o
‑
葡萄糖糖基化活性。hv1和eugt11两者将第二糖部分转移至莱鲍迪苷g的19
‑
o
‑
葡萄糖的c
‑2’
以生成莱鲍迪苷v(“reb v”),或将第二葡萄糖部分转移至莱鲍迪苷ka的13
‑
o
‑
葡萄糖的c
‑2’
以生成莱鲍迪苷e(“reb e”)。图21还显示,重组ugt76g1酶催化第三糖部分转移至莱鲍迪苷v的c
‑
19
‑
o
‑
葡萄糖的c
‑3’
以生成莱鲍迪苷w(“reb w”)的反应并且eugt11可以连续地将第三葡萄糖部分转移至莱鲍迪苷e的c
‑
13
‑
o
‑
葡萄糖的c
‑6’
以生成莱鲍迪苷d3。hv1可以将第三葡萄糖部分转移至莱鲍迪苷e的c
‑
13
‑
o
‑
葡萄糖的c
‑2’
以生成莱鲍迪苷z1(“reb z1”),并且可以将第三葡萄糖部分转移至莱鲍迪苷e的c
‑
19
‑
o
‑
葡萄糖的c
‑2’
以生成莱鲍迪苷z2(“reb z2”)。hv1和eugt11两者可以催化甜菊苷转化为reb e及莱鲍迪苷a(“reb a”)转化为莱鲍迪苷d(“reb d”)。ugt76g1可以将第三葡萄糖部分转移至莱鲍迪苷e(“reb e”)的c
‑
13
‑
o
‑
葡萄糖的c
‑3’
以形成莱鲍迪苷d(“reb d”)。ugt76g1也催化甜菊苷转化为莱鲍迪苷(“reba”)及莱鲍迪苷d(“reb d”)转化为莱鲍迪苷m(“reb m”)。
[0338]
鉴于以上所述,将会看到实现了本公开的几个优点并获得了其它有利结果。因为在不背离本公开范围的前提下可以在以上方法和体系上进行各种变化,所以意图是应将以上描述中所含的及附图中所示的所有问题均解释为说明性而非限制性意义。
[0339]
在介绍本公开或其型式、实施方案或方面的要素时,冠词“一种”、“一个”、“该”和
“
所述”旨在意指存在一个或多个要素。术语“包含”、“包括”和“具有”旨在为包括性且意指可能存在除所列要素之外的附加要素。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。
