1.本发明属于基因工程领域,涉及一种用于果蝇基因组无缝编辑的质粒载体及其构建方法和应用。
背景技术:
2.目前,对于果蝇基因组的编辑广泛使用第三代基因编辑技术crispr/cas9核酸内切酶进行,该酶可在sgrna的引导下作用于特定的基因组序列,产生dna双链断裂,利用同源定向修复机制可实现对靶基因的特异性敲除或敲入。如果在基因敲除的同时敲入attp位点,则后期可利用phic31同源重组酶介导attb位点与attp位点之间发生基因重组,进而在靶基因敲除位置引入不同基因突变。由于phic31同源重组酶介导的同源重组效率较高,因而该方法使用广泛。但是该方法在使用过程中会在敲入基因的两端引入50bp以上的外源序列,给实验结果带来不可预计的影响。为实现对果蝇基因组的无缝编辑,科研人员将crispr/cas9技术和piggybac转座酶联用,首先运用crispr/cas9技术,利用同源定向修复原理引入基因突变及筛选标记,然后利用piggybac转座酶将筛选标记精确移除,达到对基因组进行无缝编辑的效果。将这一原理运用在果蝇基因组的敲除过程中,可以利用杂交手段方便高效地移除筛选标记,具体方法见发明人于已发表的文章。该方法在运用crispr/cas9技术引入基因突变及筛选标记时,须将敲除基因两侧的同源序列精准地与载体ttaa序列连接,否则后续将无法被piggybac转座子酶识别并切除,因此无法利用限制性内切酶分别插入两侧同源臂,只能将两个同源臂、两侧带有itr的筛选标记dsred以同源重组的方式与载体进行多个片段的同源重组,但是多个片段的同源重组连接难度较大,其效率受限于片段的大小、浓度、及反应体系中不同片段的浓度,因此通常连接效率较低。
技术实现要素:
3.本发明的目的在于针对现有技术的上述不足,提供一种用于果蝇基因组无缝编辑的质粒载体。
4.本发明的另一目的是提供该质粒载体的构建方法。
5.本发明的又一目的是提供该质粒载体的应用。
6.本发明的目的可通过以下技术方案实现:
7.一种用于果蝇基因组无缝编辑的质粒载体pgx-dsred-itrs-ttaa,所述的pgx-dsred-itrs-ttaa含有3’itrs-dered-5’itrs元件,其中dsred基因不含有bbsi识别位点,dsred基因序列上、下游分别为pigggbac转座子3’和5'itrs,3’和5'itrs引入分别引入了bbsi内切酶位点,bsai内切酶位点,且紧邻ttaa序列。质粒载体pgx-dsred-itrs-ttaa主要元件的示意图如图1所示。
8.作为本发明的一种优选,所述的pgx-dsred-itrs-ttaa的骨架载体pgx-attp-dsred为以m5-δuast-rpr-frt_dsred质粒为基础,同义突变消除ampr基因中含有的bsai内
切酶识别位点所得,m5-δuast-rpr-frt_dsred质粒序列如seq id no.2所示。
9.作为本发明的进一步优选,所述的质粒载体pgx-dsred-itrs-ttaa序列如seq id no.1所示,质粒图谱见图2。
10.本发明所述的质粒载体pgx-dsred-itrs-ttaa的构建方法,包含以下步骤:
11.(1)通过同义突变消除骨架载体m5-δuast-rpr-frt_dsred中ampr基因包含的bsai内切酶识别位点获得pgx-attp-dsred;
12.(2)从载体pshd-dsred中扩增获得含3’tr-dered-5’tr序列的片段,含3’tr-dered-5’tr序列的片段中3’tr-dered-5’tr序列直接与ttaa相连接,并在上下游分别引入bsal和bbsl酶切位点以及骨架载体同源序列,3’tr-dered-5’tr序列中dsred基因上不存在bbsi识别位点;
13.(3)步骤(1)获得的pgx-attp-dsred载体经kpni-hf spei双酶切,获得的片段与步骤(2)获得的3’tr-dered-5’tr序列通过多片段同源重组酶进行连接获得质粒载体pgx-dsred-itrs-ttaa。
14.作为本发明的一种优选,步骤(1)通过overlap-pcr引入同义突变ggg
→
ggt,通过上下游hindiii和scai酶切位点切除原载体骨架m5-δuast-rpr-frt_dsred中包含bsai酶切位点的片段,将线性骨架片段与点突变的片段通过单片段同源重组酶连接。
15.作为本发明的一种优选,步骤(1)包含以下步骤:
16.(i)hindiii-hf scai-hf双酶切m5-δuast-rpr-frt_dsred载体,回收长度为3318bp的长片段,命名为a;
17.(ii)获得含同义点突变的ampr片段:以m5-δuast-rpr-frt_dsred载体为模板,分别以x33 x28和x29 x35为引物,使用q5高保真酶扩增获得长度为1220bp的片段b和长度为451bp的片段c;以等摩尔数混合的片段b和c为模板,以x33 x35为引物,使用q5高保真酶扩增获得长度为1634bp的片段d;其中引物x33序列为atttaagtgtatacttcggt,引物x28序列为tggagccggtgagcgtggttctcgcggtatcattgca,引物x29序列为tgcaatgataccgcgagaaccacgctcaccggctcca,引物x35序列为ttctcagaatgacttggttg;
18.(iii)连接获得含同义点突变ampr序列的骨架载体pgx-attp-dsred:将片段a和d在10ul单片段同源重组酶体系中连接,并验证获得载体pgx-attp-dsred。
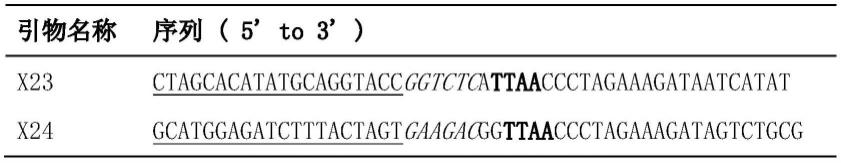
19.作为本发明的一种优选,步骤(2)包含以下步骤:以pshd-dsred载体为模板,分别以x23 x26和x24 x25为引物,使用q5高保真酶扩增获得长度为926bp的片段f和长度为866bp的片段g;其中引物x23序列为ctagcacatatgcaggtaccggtctcattaaccctagaaagataatcatat,引物x26序列为ctcccagcccatagtctttttctgcattacggggccg,引物x24序列为gcatggagatctttactagtgaagacggttaaccctagaaagatagtctgcg,引物x25序列为cggccccgtaatgcagaaaaagactatgggctgggag;所述的pshd-dsred载体序列如seq id no.3所示。
20.作为本发明的一种优选,步骤(3)包含以下步骤:
21.(i)将步骤1获得的新载体pgx-attp-dsred经kpni-hf spei双酶切,回收长度为3133bp的长片段,命名为e;
22.(ii)将所述的片段f、g和e在多片段同源重组酶体系中连接获得含同义点突变dsred序列的载体pgx-dsred-itrs-ttaa。
23.本发明所述的质粒载体pgx-dsred-itrs-ttaa在构建果蝇基因组无缝编辑所需同
源臂供体质粒中的应用。
24.一种构建果蝇基因组无缝编辑供体质粒的方法,包含以下步骤:
25.(1)pcr扩增获得5’和3’同源臂,引物序列包含与骨架载体同源的互补序列-ttaa-与目的基因基本互补且引物同义突变的序列;5’和3’同源臂中至少有一条不含有限制性内切酶bbsi、bsai的酶切位点;
26.(2)pgx-dsred-itrs-ttaa载体经bbsi xhoi双酶切,回收长度为4784bp的长片段;
27.(3)将一测同源臂和步骤(2)中获得的长度为4784bp的载体片段连接后再插入另一侧同源臂;当有一侧同源臂含有限制性内切酶bbsi或bsai的酶切位点时,首先插入不含此2种酶切位点的同源臂,成功以后插入另一侧同源臂;若上下游同源臂均不包含这2种酶切位点,则可任意插入一侧同源臂,成功以后插入另一侧同源臂;同源臂与载体片段连接的方式选自同源重组或t4连接酶连接。同源臂与酶切后的载体连接时,推荐使用同源重组的方法进行连接,以保证连接方向正确。若想采用t4连接酶进行连接,则需在同源臂两侧设计正确的序列,使酶切得到的粘性末端与载体相同,由于同源臂两侧的粘性末端相同,因此插入方向会有正反向两种,需进行pcr鉴定获得方向正确的插入。无论通过何种方式连接同源臂与载体,请务必通过测序确定插入序列正确。
28.本发明详述
29.本发明质粒改造的目的是:获得dsred序列上下游分别为pigggbac转座子3’和5'itrs,3’和5'itrs含bsal和bbsl酶切位点,且紧邻ttaa序列的载体,目的载体主要元件的示意图如图1。
30.解决的问题和技术手段:
31.3和5'itrs均需与ttaa直接相连,两者之间不能存在其碱基,才能被pigggbac转座子酶识别并准确移除,因此需要选用切割位点位于识别位点以外的限制性内切酶,使得确切后遗留ttaa序列,3'和5'itrs分别选用bsal和bbsl酶,这两个内切酶的切割位点位于识别位点下游,将切割位点设计为ttaa序列,则可在酶切后产生ttaa粘性末端。
[0032]3’
tr-dered-5’tr序列可从载体pshd-dsred中扩增获得,为了使该段序列直接与ttaa相连接,并同时在上下游分别引入bsal和bbsl酶切位点,在设计引物时5’端增添这部分序列,为了扩增后的片段可利用同源重组的方法连接至骨架载体m5-δuast-rpr-frt_dsred载体,引物中还需要添加与该载体同源的序列,以下为扩增时所用引物的序列:
[0033][0034]
注:粗体为ttaa序列,斜体为bsal和bbsl内切酶识别位点,下划线字体为与骨架载体同源序列
[0035]
经测序发现质粒载体pshd-dsred上的dsred基因上存在bbsi识别位点gaagac,如图5所示。为了防止使用过程中将该基因切断,可对其对应的氨基酸做同义突变aag
→
aaa,将该酶切位点消除,密码子依然编码赖氨酸,不影响蛋白质功能,方法为通过overlap-pcr引入突变,使用到的引物序列如下:
[0036][0037]
注:加粗下划线字体为引入的同义突变
[0038]
图3为本发明所用的初始骨架载体m5-δuast-rpr-frt_dsred,经测序发现该质粒ampr基因上存在bsal识别位点ggtctc,如图6所示。为了防止使用过程中将该基因切断,可对其对应的氨基酸做同义突变可对其对应的氨基酸做同义突变ggg
→
ggt,将该酶切位点消除,密码子依然编码甘氨酸,不影响蛋白质功能,方法为通过overlap-pcr引入突变,通过上下游hindiii和scai酶切位点切除原载体骨架中包含bsai酶切位点的片段,将线性骨架片段与点突变的片段通过单片段同源重组酶连接,使用到的引物序列如下:
[0039][0040][0041]
注:加粗下划线字体为引入的同义突变
[0042]
有益效果:
[0043]
dsred上下游酶切位点分别采用了切割位点位于识别位点下游的两种限制性内切酶bbsi和bsai,保证了酶切产生的粘性末端为ttaa序列,从而在同源重组之后得到载体中dsred上下游转座子元件5’tr和3
‘
tr均紧邻ttaa序列。本发明在piggybac转座子5
‘
tr和3
‘
tr分别引入了bbsi内切酶位点和bsai内切酶位点,方便分别插入同源重组所需的上下游同源臂。通过这样的设计方案,在构建果蝇基因组无缝编辑所需同源臂供体质粒时,可先通过单酶切在载体中引入一侧同源臂,测序鉴定正确之后,再通过另一个内切酶引入剩余一侧同源臂。由于单片段同源重组的远高于多片段同源重组,因此本发明降低了获得无缝编辑所需供体质粒的难度。
附图说明
[0044]
图1质粒载体pgx-dsred-itrs-ttaa主要元件的示意图。
[0045]
图2pgx-dsred-itrs-ttaa质粒图谱。
[0046]
图3m5-δuast-rpr-frt_dsred质粒图谱。
[0047]
图4pshd-dsred质粒图谱,本发明中使用到的dsred序列及两侧piggybac转座子的来源质粒,由于dsred基因上存在bbsi识别位点gaagac,因此需要对其相应的氨基酸做同义突变,将该酶切位点消除。
[0048]
图5pshd-dsred质粒中dsred基因上存在bbsi识别位点示意图。
[0049]
图6骨架载体m5-δuast-rpr-frt_dsred中ampr基因上存在bsal识别位点ggtctc示意图。
[0050]
图7m5-δuast-rpr-frt_dsred载体hindiii-hf scai-hf双酶切图。
[0051]
图8获得含同义点突变的ampr片段的扩增片段跑胶结果。
[0052]
图9pgx-attp-dsred测序结果片段。
[0053]
图10扩增含同义点突变的dsred片段跑胶结果。
[0054]
图11pgx-attp-dsred载体kpni-hf spei双酶切结果。
[0055]
图12pgx-dsred-itrs-ttaa载体测序结果片段。
[0056]
图13kis-2455-5’arm和kis-2455-3’arm扩增产物的跑胶结果。
[0057]
图14pgx-dsred-itrs-ttaa载体bbsi xhoi双酶切结果。
[0058]
图15pgx-dsred-itrs-ttaa-3’arm载体测序结果片段。
[0059]
图16pgx-dsred-itrs-ttaa-3’arm载体nhei bsai双酶切结果。
[0060]
图17成功插入了同源臂kis-2455-3’arm和kis-2455-5’arm的供体质粒测序结果片段。
具体实施方式
[0061]
实施例1质粒pgx-dsred-itrs-ttaa构建过程:
[0062]
1.消除骨架载体m5-δuast-rpr-frt_dsred中ampr基因包含的bsai内切酶识别位点
[0063]
1)m5-δuast-rpr-frt_dsred载体双酶切:hindiii-hf scai-hf(neb,buffer cutsmart,50ul体系),图7左图为软件预测的酶切条带,3318bp 1590bp,图7右图为实际酶切后获得的片段,回收长度为3318bp的长片段(红色箭头所指),命名为a。
[0064]
2)获得含同义点突变的ampr片段:以m5-δuast-rpr-frt_dsred载体为模板,分别以x33 x28和x29 x35为引物,使用q5高保真酶扩增获得长度为1220bp的片段b和长度为451bp的片段c,图8左图为扩增片段跑胶结果,回收两片段(红色箭头所指)。以等摩尔数混合的片段b和c为模板,以x33 x35为引物,使用q5高保真酶扩增获得长度为1634bp的片段d,大于1.5kb的片段(图8右图红色箭头所指)。引物x33序列为atttaagtgtatacttcggt,引物x28序列为tggagccggtgagcgtggttctcgcggtatcattgca,引物x29序列为tgcaatgataccgcgagaaccacgctcaccggctcca,引物x35序列为ttctcagaatgacttggttg。
[0065]
3)连接获得含同义点突变ampr序列的新载体,并将其命名为pgx-attp-dsred:将片段a和d在10ul单片段同源重组酶(clone expressii one step cloning kit_c112_诺唯赞)体系中连接,连接产物转化dh5a感受态细胞,挑选4个菌落pcr阳性的菌液测序,测序结果显示4个质粒均含设计突变(图9,红框标记),并且其他位置没有非特异性点突变。
[0066]
2.获得含同义点突变的dsred片段:以pshd-dsred载体为模板,分别以x23 x26、x24 x25为引物,使用q5高保真酶扩增,x23 x26的扩增产物为926bp,x24 x25的扩增产物为866bp,图10为扩增片段跑胶结果,回收两片段(红色箭头所指),分别命名为f和g。引物x23序列为ctagcacatatgcaggtaccggtctcattaaccctagaaagataatcatat,引物x26序列为ctcccagcccatagtctttttctgcattacggggccg,引物x24序列为gcatggagatctttactagtgaagacggttaaccctagaaagatagtctgcg,引物x25序列为cggccccgtaatgcagaaaaagactatgggctgggag。
[0067]
3.获得含同义点突变dsred序列的目的载体:
[0068]
1)将步骤1获得的新载体pgx-attp-dsred双酶切:kpni-hf spei(neb,buffer cutsmart,50ul体系),图11左图为软件预测的酶切条带,3133bp 1775bp,图11右图为实际酶切后获得的片段,回收长度为3133bp的长片段(红色箭头所指),命名为e。
[0069]
2)连接获得含同义点突变dsred序列的新载体,并将其命名为pgx-dsred-itrs-ttaa:将片段f、g和e在10ul多片段同源重组酶(cloneexpress muleis one step cloning kit_c113_诺唯赞)体系中连接,连接产物转化dh5a感受态细胞,挑选2个菌落pcr阳性的菌液测序,测序结果(图12)显示2个质粒均含设计突变(红框标记),并且其他位置没有非特异性点突变。
[0070]
实施例2使用质粒pgx-dsred-itrs-ttaa构建无缝编辑所需同源臂质粒以下案例为使用本专利发明的质粒pgx-dsred-itrs-ttaa构建kismet基因点突变gln2455term所需同源臂质粒的实验过程实验目的:将含点突变的kismet同源臂克隆至质粒载体pgx-dsred-itrs-ttaa中。
[0071]
实验过程:
[0072]
1.设计构建方案及所需引物序列,pcr扩增获得5’和3’同源臂,引物序列如下:
[0073][0074]
注:粗体为ttaa序列,斜体字体为与骨架载体同源序列,下划线字体为引入的同义突变
[0075]
(1)以胚胎注射所用基因型果蝇基因组dna为模板,分别扩增5’和3’同源臂:kis-2455-5’arm扩增引物:x74 x75=1624bp;kis-2455-3’arm扩增引物:x76 x77=1844bp,扩增产物跑胶结果如下,回收两片段(红色箭头所指),分别命名为kis-2455-5’arm和kis-2455-3’arm(图13)。
[0076]
2.pgx-dsred-itrs-ttaa载体双酶切:bbsi xhoi(neb,buffer 2.1,50ul体系),软件预测的酶切条带,4784bp 66bp,图14为实际酶切后获得的片段,回收长度为4784bp的长片段(红色箭头所指),命名为pgx-dsred-itrs-ttaa_b x。
[0077]
3.连接含同义点突变的kis-2455-3’arm至pgx-dsred-itrs-ttaa质粒载体,并将其命名为pgx-dsred-itrs-ttaa-3’arm:将片段kis-2455-3’arm和pgx-dsred-itrs-ttaa_b x在10ul单片段同源重组酶(clone express ii onestep cloning kit_c112_诺唯赞)体系中连接,连接产物转化dh5a感受态细胞,挑选2个菌落pcr阳性的菌液测序,测序结果显示2个质粒均含设计突变(红框标记),并且其他位置没有非特异性点突变(图15)
[0078]
4.pgx-dsred-itrs-ttaa-3’arm载体双酶切:nhei bsai(neb,buffercutsmart,
50ul体系),图16为酶切后获得的片段,回收长度为6602bp的长片段(红色箭头所指),命名为pgx-dsred-itrs-ttaa-3’arm_n b。
[0079]
5.kis-2455-5’arm至pgx-dsred-itrs-ttaa质粒载体:将片段kis-2455-5’arm和pgx-dsred-itrs-ttaa-3’arm_n b在10ul单片段同源重组酶(clone express ii one step cloning kit_c112_诺唯赞)体系中连接,连接产物转化dh5a感受态细胞,挑选2个菌落pcr阳性的菌液测序,测序结果显示2个质粒均成功插入了kis-2455-5’arm,并且其他位置没有非特异性点突变(图17)。
[0080]
经过以上步骤我们成功获得了可用于构建kismet-gln2455term点突变果蝇所需的同源臂供体载体,后续构建转基因果蝇的步骤参见发明人于2019年发表在《遗传》杂志上的文章(王珏,黄娟,许蕊*:利用crispr/cas9和piggybac实现果蝇基因组无缝编辑,遗传,2019,41(5):422-29.)。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。