adk抑制剂在制备治疗脊髓损伤的药物中的应用
技术领域
1.本发明属于生物医药领域,涉及adk抑制剂在制备治疗脊髓损伤的药物中的应用。
背景技术:
2.脊髓损伤一直是一个全球性的难题,近年来,随着对脊髓损伤病理机制研究的不断深入,虽然取得了一定的进展,但是如何克服脊髓损伤后神经元突起再生仍然是人类的巨大挑战。最近越来越多的研究表明通过调控细胞骨架的运动,改变脊髓损伤的微环境,给予神经生长促进因子,或者给予干细胞的移植等可以部分的修复受损的神经元、促进轴突的再生、促进运动和感觉功能的恢复。但是,如何精细地调控神经元再生的时间、路径,如何有效控制再生区域的微环境促进神经元突起再生等等一系列问题都亟待解决。
3.目前有关脊髓损伤的治疗策略很多,从药物治疗、手术治疗到细胞治疗和康复训练,所有这些治疗措施在脊髓损伤的恢复过程中都起到了积极的作用。虽然医务工作人员竭尽全力对脊髓损伤患者进行治疗,使其死亡率从20世纪初期的50%降低到目前的6%左右,但脊髓损伤后的功能恢复仍不能令人满意。截瘫是一种比较常见的严重伤残,常引起各种并发症,严重影响患者的生活质量。急性脊髓损伤后,持续的机械压迫引起脊髓微循环障碍,导致局部缺血和损伤区域的进一步扩大。外科手术治疗主要通过硬膜外减压,解除骨性结构、椎间盘以及韧带对脊髓的压迫,减轻脊髓压力从而改善局部微环境,达到促进神经功能恢复的目的。
4.在脊髓损伤的临床前期试验中,观察到甲基强的松龙能够通过上调抗炎因子的释放,降低氧化应激反应程度,抑制细胞内钾离子的耗竭和脂质过氧化,从而发挥神经保护作用。但是,在一项大样本、多中心、随机对照研究中发现,大剂量甲基强的松龙疗法并不能使脊髓患者的临床症状得到改善,反而会增加胃肠道出血等不良事件的发生。在其他药物治疗方面,目前只有少数药物正在进行的临床试验的药物(利鲁唑、格列本脲、硫酸镁、尼莫地平和米诺环素)对脊髓损伤患者的神经具有保护能力。研究表明,微管(microtubulin,mt)稳定剂也可作为中枢神经系统(central nervous system,cns)各种疾病的候选治疗药物,从脑肿瘤到脊髓损伤,以及一些神经退行性疾病,包括阿尔茨海默氏症、帕金森氏症和肌萎缩性脊髓侧索硬化症。虽然这些药物已经被证明可以减少细胞死亡和减少损伤的进展,但它们并不能非常有效的促进神经再生和脊髓组织修复。所以,临床上也面临着对于脊髓损伤的患者无药可用的窘境。
5.adk( adenosine kinase,腺苷激酶)属于一种核苷激酶,其主要功能在于催化atp与adp之间的转变,adk催化底物的过程伴随adk从闭合状态到开放状态的蛋白质构象变化。adk广泛存在于各种动物中,并在神经系统中大量表达,其包括有两种剪接体,分为长型和短型,其中长型adk包含了核定位序列,主要分布于细胞核,而短型adk则定位于细胞质,其比长型adk少了20个氨基酸。
6.现有研究表明,adk功能障碍与多种疾病有关。adk-s对于控制癫痫发作很重要,而adk-l控制表观遗传功能,特别是dna甲基化,dna甲基化变化反过来又是调节大脑发育、成
熟、可塑性和细胞增殖的候选机制之一。在大脑发育过程中,adk经历了从神经元到星形胶质细胞的表达变化,因此在成人大脑中,adk主要表达于具有增殖潜力的可塑性细胞,如星形胶质细胞。在小鼠胚胎脑发育过程中发现随着时间的推移,adk-l/adk-s比值逐渐下降,adk-l对adks的表达占主导地位。出生后,adk-l/adk-s表达比率发生了转变,转变为adk-s相对adk-l来说为主体的表达模式,因此adk-l的表达与脑的生长发育有关。在出生后大脑发育的头两周,神经元adk-l转录迅速下降,adk-s转录上升。即在神经元和未成熟前体细胞的细胞核中表达的adk-l在胚胎和出生后早期的大脑发育中起着关键作用。在脑内缺乏adk表达的小鼠中,从胚胎第11天开始,显示出神经元可塑性改变导致应激诱导的癫痫发作,记忆和学习缺陷,而出生后adk在大脑大部分区域的表达中断并没有导致这些行为缺陷和癫痫发作。adk在心肌细胞的微管动态性、脑缺血、癫痫等疾病或症状中表现出密切的相关性,然而其在脊髓损伤中的功能作用尚不明晰。
技术实现要素:
7.本发明的目的在于解决现有技术中所存在的上述问题,从而针对脊髓损伤的相关机制进行了深入研究,揭示了通过调控目标靶点adk的活性从而有效促进细胞自噬,进而促进神经元的生长和脊髓损伤的修复,为脊髓损伤修复提供了新的治疗靶点,为临床干预和治疗脊髓损伤提供了切实的实验证据和科学依据。
8.为了解决上述技术问题,本发明是通过如下技术方案得以实现的。
9.本发明第一方面提供了adk抑制剂在制备用于脊髓损伤的药物中的应用。
10.作为优选地,所述adk抑制剂选自5-iodotubercidin(5-iod,cas no. 24386-93-4)、abt 702 dihydrochloride(abt702,cas no. 1188890-28-9)、基于adk设计的shrna中的一种或多种。
11.作为优选地,所述基于adk设计的shrna选自如下中的一种或多种:shrna1,序列为ugcugccgccaauuguuauaa;shrna2,序列为ccuugauaagtuauucucugaa;shrna3,序列为gcuuugagacuaaagacauua。
12.本发明第二方面提供了adk抑制剂在制备用于促进神经元生长的药物中的应用。
13.作为优选地,所述adk抑制剂选自adk抑制剂5-iodotubercidin(5-iod,cas no. 24386-93-4)、abt 702 dihydrochloride(abt702,cas no. 1188890-28-9)、基于adk设计的shrna中的一种或多种。
14.作为优选地,所述基于adk设计的shrna选自如下中的一种或多种:shrna1,序列为ugcugccgccaauuguuauaa;shrna2,序列为ccuugauaagtuauucucugaa;shrna3,序列为gcuuugagacuaaagacauua。
15.本发明第三方面提供了adk抑制剂在制备用于促进细胞自噬的药物中的应用。
16.作为优选地,所述adk抑制剂选自5-iodotubercidin(5-iod,cas no. 24386-93-4)、abt 702 dihydrochloride(abt702,cas no. 1188890-28-9)、基于adk设计的shrna中的一种或多种。
17.作为优选地,所述基于adk设计的shrna选自如下中的一种或多种:shrna1,序列为ugcugccgccaauuguuauaa;shrna2,序列为ccuugauaagtuauucucugaa;shrna3,序列为gcuuugagacuaaagacauua。
18.本发明第四方面提供了一种用于治疗脊髓损伤的药物组合物,包括adk抑制剂和atg101蛋白。
19.作为优选地,所述adk抑制剂选自5-iodotubercidin(5-iod,cas no. 24386-93-4)、abt 702 dihydrochloride(abt702,cas no. 1188890-28-9)、基于adk设计的shrna中的一种或多种。
20.作为优选地,所述基于adk设计的shrna选自如下中的一种或多种:shrna1,序列为ugcugccgccaauuguuauaa;shrna2,序列为ccuugauaagtuauucucugaa;shrna3,序列为gcuuugagacuaaagacauua。
21.本发明第五方面提供了一种用于促进神经元生长的药物组合物,包括adk抑制剂和atg101蛋白。
22.作为优选地,所述adk抑制剂选自5-iodotubercidin(5-iod,cas no. 24386-93-4)、abt 702 dihydrochloride(abt702,cas no. 1188890-28-9)、基于adk设计的shrna中的一种或多种。
23.作为优选地,所述基于adk设计的shrna选自如下中的一种或多种:shrna1,序列为ugcugccgccaauuguuauaa;shrna2,序列为ccuugauaagtuauucucugaa;shrna3,序列为gcuuugagacuaaagacauua。
24.本发明第六方面提供了atg101蛋白在制备用于逆转基于adk引起的神经元再生障碍的药物中的应用。
25.应注意的是,在无特别说明的情况下,本发明上下文中所述“adk抑制剂”或类似表述是指能够特异性降低细胞和/或组织中adk表达水平的物质,所采用的5-iodotubercidin、abt 702 dihydrochloride等抑制剂仅用于对相关实验的开展和演示,以便于本领域技术人员更好地理解本发明的方案和构思,并不作为对本发明技术方案和保护范围的限制,本领域技术人员有能力基于adk蛋白序列或控制adk表达的核酸序列设计相关物质以调控adk表达水平,例如sirna、sgrna、shrna、抗adk抗体等,亦或可以直接购买市售产品,例如adk单克隆抗体、各类小分子抑制剂等。本发明中所使用的5-iodotubercidin、abt 702 dihydrochloride均购于美国sellect公司,其中5-iodotubercidin分子式为c
11h13
in4o4,分子量为392.15,纯度为99.60%;abt 702 dihydrochloride分子式为c
22h21
brcl2n6o,分子量为536.25,纯度为99.76%。
26.细胞自噬是细胞内的一种依赖溶酶体的细胞降解途径,是机体内存在的一种自我修复和维持生命的过程。细胞自噬可调节细胞骨架微管的动态性促进脊髓损伤修复,但是通过何种途径调控微管动态性促进脊髓损伤修复以及何种因素调控细胞自噬参与脊髓损伤修复等机制不明。那么,明确与自噬相关的关键基因、关键蛋白、代谢产物以及关键的信号通路至关重要,这些关键的基因及蛋白发挥促神经再生的机制也是值得探索的重要科学问题。
27.细胞自噬现象首先发现于酵母中,是广泛存在于真核细胞内的一种高度保守的溶酶体依赖性降解途径,是细胞自我保护的一种重要机制,对维持细胞存活、更新、物质再利用及内环境稳定具有重要的作用。自噬通过清除和再循环受损或不需要的蛋白质和细胞器来维持细胞稳态。自噬是在饥饿、细胞凋亡以及各种细胞系的发育过程中被诱导的。自噬是一种进化上保守的细胞成分降解和再循环过程,在组织内稳态和细胞生存中起着重要作
用。自噬是一个关键的细胞过程,越来越多的证据表明,干细胞的自我更新、多能性、分化和静止需要自噬激活,而功能失调的自噬可能与各种疾病有关。
28.现有研究发现在中枢神经系统缺血、缺氧实验中,大脑和小脑皮质大部分神经元细胞死亡,泛素聚集于神经元细胞周围,且随时间延长而增多,但蛋白酶体功能无明显改变,从而证明自噬对神经元细胞的存活具有保护作用。此外,另有研究表明,在成年大鼠脊髓半切模型中,损伤后4h beclin-1表达增高,并在3d达高峰,证实脊髓损伤后自噬被激活。
29.脊髓损伤是一种破坏性创伤,常常导致患者感觉、运动和自主神经功能丧失。由于脊髓损伤的修复机制并不十分清晰,所以脊髓损伤患者的治疗效果往往并不令人满意。细胞自噬是细胞内的一种依赖溶酶体的细胞降解途径,是机体内存在的一种自我修复和维持生命的过程。细胞自噬可调节细胞骨架微管的动态性促进脊髓损伤修复,但是通过何种途径调控微管动态性促进脊髓损伤修复以及何种因素调控细胞自噬参与脊髓损伤修复等机制不明。所以,阐明自噬在脊髓损伤的信号转导机制,对探寻脊髓损伤治疗的药物靶点具有特别重要的意义。因此,本发明通过脊髓损伤动物模型和神经元自噬模型的蛋白质组学联合分析发现adk同时参与了细胞自噬与脊髓损伤修复,进一步通过多种实验手段验证了adk通过自噬参与脊髓损伤修复。本发明也发现调控adk可促进损伤脊髓的修复。另外,相互作用蛋白质谱和分子实验显示adk可以与自噬相关蛋白101(atg101)相结合。
30.本发明相对于现有技术具有如下技术效果:本发明对脊髓损伤修复的具体机制进行了深入研究,发现adk同时参与了细胞自噬与脊髓损伤修复,进一步通过多种实验手段验证了adk通过自噬参与脊髓损伤修复;同时本发明也发现通过adk抑制剂干扰adk蛋白的表达可以明显增强细胞自噬。另外,本发明还明确了损伤神经元中共表达adk和atg101,adk和atg101可以共同调控损伤神经元的修复,过表达atg101可以逆转过表达adk对损伤神经元修复的抑制作用。本发明为脊髓损伤修复的药物开发提供了新的治疗靶点,为临床中脊髓损伤修复的干预和治疗提供了切实的实验证据和科学依据。
附图说明
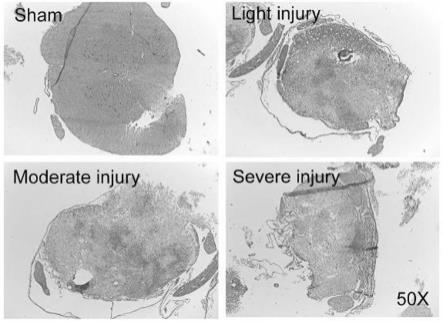
31.图1为脊髓损伤后72h不同程度损伤的脊髓组织横断片he染色图,放大倍数为50
×
。
32.图2为脊髓损伤72h大鼠关注后取出不同程度损伤的脊髓实物图,scale bar为5mm。
33.图3为脊髓损伤后72h取出不同程度损伤的脊髓组织横切面的免疫荧光染色结果示意图,scale bar为200μm。
34.图4为不同损伤程度的脊髓组织中相关自噬蛋白表达水平结果示意图。
35.图5为假手术组和损伤脊髓的脊髓组织横断面的免疫荧光染色结果示意图,scale bar为200μm。
36.图6为不同处理方式下对海马神经元中相关自噬蛋白表达情况以及突起总长度和分支数目的影响结果示意图。
37.图7为不同处理方式下cos7细胞荧光染色结果示意图。
38.图8为不同处理方式下c0s7细胞中t-tub/a-tub比值结果示意图。
39.图9为不同损伤程度的脊髓组织中蛋白质谱结果显示adk蛋白表达量。
40.图10为不同损伤程度的脊髓组织中adk蛋白的表达情况结果示意图。
41.图11为假手术组和损伤脊髓组织横切面的免疫荧光染色结果示意图,scale bar为200μm。
42.图12为不同处理条件下对海马神经元的影响结果示意图。
43.图13为不同处理条件下对神经元突起总分支数目、一级和二级分支数目、神经元突起总长度、一级和二级分支突起总长度分析结果示意图。
44.图14为adk抑制剂对脊髓损伤大鼠体重以及下肢功能恢复的影响结果示意图。
45.图15为不同shrna对sh-sy-5y细胞内adk抑制作用结果示意图。
46.图16为不同shrna对sh-sy-5y细胞内p62和lc3蛋白表达情况影响结果示意图。
47.图17为不同shrna对sh-sy-5y细胞内p62和lc3蛋白表达情况影响定量结果示意图。
48.图18为adk抑制剂处理下sh-sy-5y细胞内p62和lc3蛋白表达情况结果示意图。
49.图19为adk抑制剂处理下sh-sy-5y细胞内p62和lc3蛋白表达情况定量结果示意图。
50.图20为体外纯化gst-adk蛋白、gst-adk蛋白与脊髓组织蛋白进行pull down后的蛋白跑胶后进行考马斯亮蓝染色结果示意图。
51.图21为对回收的蛋白进行蛋白质谱鉴定和分析的结果示意图。
52.图22为体外纯化gst-atg101蛋白跑胶后进行考马斯亮蓝染色以及gst或gst-atg101与脊髓组织蛋白裂解液进行pull down后用adk抗体检测结果示意图。
53.图23为对培养3天的海马神经元以及损伤脊髓组织切片的免疫荧光染色结果示意图。
54.图24为不同处理条件下对体外海马神经元生长影响结果示意图。
55.图25为不同处理条件下对体外海马神经元生长定量影响结果示意图。
具体实施方式
56.为使本发明的目的、技术方案及效果更加清楚、明确,以下参照实施例对本发明作进一步详细说明。应当理解,此处所描述的具体实施例仅用以解释本发明,并不用于限定本发明。
57.在无特别说明的情况下,本发明上下文中所列出的包括sh-sy-5y、cos7等细胞均购自于中国科学院上海生科院细胞资源中心,并根据细胞生物学中的常规方法进行培养。所有细胞系均通过中国典型培养物保藏中心(武汉)的短串联重复分析鉴定,并使用pcr检测试剂盒(上海biothrive sci)验证是否存在支原体污染,同时在液氮中冷冻保存并用于后续实验。本发明所使用的试剂中,均通过市售获得。本发明所使用的实验方法和技术,例如sh-sy-5y、cos7细胞培养、western blot、分子克隆、pcr、蛋白质谱分析、免疫荧光染色、激光共聚焦、流式细胞术及动物实验等均为本领域的常规方法和技术。
58.生物学实验重复中选择具有代表性的结果呈现在上下文附图中,数据按照图示中规定的以mean
±
sd和mean
±
sem展示。所有实验至少重复三次。数据采用graphpad prism 5.0或spss 20.0软件进行分析。采用t检验或方差分析比较两组或两组以上的平均值差异。
p<0.05被认为是一个显著的差异。
59.实施例1脊髓损伤(sci)模型的构建,所选用的实验动物为220-250g雌性大鼠(购自于广东省动物中心(spf级)),单独饲养在25
±
3℃的spf级设施中,提供常规食物和水,具体包括如下步骤:(1)腹腔注射10%水合氯醛(0.35ml/100g体重)用于麻醉sd大鼠,此后进行2.5厘米的纵向背侧切口,露出t9-11脊突和椎板。
60.(2)切除整个t10椎板,并且脊柱的暴露区域约为2.5mm
×
3mm。
61.(3)用稳定器将t10切面双边固定,控制冲击头的氮气罐设置为18psi或124kpa。将带有大鼠的u型稳定器加载到路易斯维尔损伤系统设备(lisa)的平台上,并直接在撞击器下方调节硬脑膜/脊髓高度,同时通过激光束进行监测。
62.(4)将碰撞深度调整为不同的损坏级别,对于轻度(light injury),中度(moderate injury)和重度(severe injury)伤害,碰撞深度设置为0.6mm,1.0mm或1.8mm,时间设置为0.5s。
63.(5)诱发损伤后,将稳定器从平台上拆下,将大鼠从稳定器中移出,评估受伤部位,并抑制出血,最后使用3-0丝线缝合大鼠的肌肉和皮肤。
64.目标sci模型动物符合以下伤害标准:麻痹性麻痹,尾巴摇摆反射,身体和腿部轻弹,脊髓缺血和伤口部位周围的水肿。假手术(sham)的动物接受了t10全椎板切除术,但是脊髓没有受伤。所有大鼠均接受2000u/天的庆大霉素治疗,每8小时手动挤压每只大鼠的膀胱以帮助排尿,直到观察到自发性排尿。
65.应用上述构建的sci大鼠模型,在损伤72h后选取各组大鼠脊髓t10(脊髓损伤造模处)为中心长约0.8cm的脊髓,将取下的脊髓组织浸泡于4%多聚甲醛中进行固定处理,随后进行切片和染色观察脊髓组织的变化,结果如图1-3所示。其中he染色显示,在脊髓损伤组中,大鼠脊髓组织呈现水肿,炎性细胞浸润,脊髓核固缩等现象,而假手术组的脊髓组织显示无明显异常(参见图1)。从组织形态上来看,脊髓损伤组的脊髓表面呈现不平整以及充血的状态,而假手术组的脊髓光滑平整无充血(参见图2)。通过对相应阶段脊髓进行免疫荧光染色发现,现在假手术组中gfap及mbp的表达强烈,而脊髓损伤组的手术中央区域的结构高度紊乱,gfap及mbp的表达下调(参见图3),这表明与假手术组相比,脊髓损伤组受损节段呈现胶质细胞增多,神经细胞减少的情况。
66.随后,分别取各组小鼠脊髓组织进行自噬相关蛋白检测,具体步骤如下:(1)将玻璃板清洗干净,配好分离胶和浓缩胶,室温静置30min以保证完全凝胶;在buffer中拔掉胶板上的梳子,将变性的蛋白样品加入梳子孔中。
67.(3)上样完毕,恒压65-80v,当溴酚蓝指示带被压缩成一条线进入分离胶后,恒压120-140v,直至溴酚蓝指示带跑至凝胶末端处即停止电泳。
68.(4)电泳结束后,将pvdf膜泡在甲醇中2-4s,然后浸泡在1
×
transfer buffer中,同时将海绵、滤纸浸泡;打开电转印夹,黑色朝下,铺上一层浸泡好的海绵和滤纸,将凝胶平铺上,用镊子小心夹起pvdf膜平放在凝胶上,附上一层滤纸,左手固定住滤纸上下角,右手拿小玻璃管驱赶气泡,注意不可将手指压到膜的中间,盖上海绵,电转印夹插入电转槽,连接好电极,恒压100v,75min。
69.(5)转膜完成后,将膜放入1
×
tbs中漂洗,放入5%脱脂牛奶的封闭液中,置于摇床上,室温封闭90min。
70.(6)将封闭后的转印膜放入1
×
tbs中漂洗,一抗(lc3 i、lc3 ii、p62、gapdh)1:1000稀释,用封闭液配制1ml稀释的抗体,将抗体稀释液滴在自制的湿盒薄膜上,将转印膜倒贴在上面,确认膜下没有气泡,用保鲜膜封住湿盒以免膜干掉,4℃孵育过夜或室温(20-25℃)孵育3h;用1
×
tbst洗膜3次,每次15min,同样方法配制二抗(1:15000稀释),湿盒里室温孵育2h;用1
×
tbst洗膜3次,每次15min。
71.(7)ecl法检测:首先将发光液a和发光液b等体积混合,然后将洗好的转印膜倒贴在混好的发光液上,避光室温孵育2min进行显影。
72.检测结果图4-5所示。结果显示,自噬相关蛋白lc3 i和lc3 ii蛋白表达明显上调,而p62蛋白表达显著下降,以上结果表明细胞自噬参与了脊髓损伤修复的过程,由此可见明确自噬与脊髓损伤的修复密切相关。
73.既往研究表明自噬可通过稳定细胞骨架微管促进神经元轴突再生,为了验证这一结论,通过调控神经元自噬水平观察自噬对细胞骨架微管动态性和神经元生长的影响。对此,在体外合成了tat-scramble(对照)和tat-beclin1肽段,且纯度达到98%以上;tat-beclin1是一种特异的自噬诱导肽,该肽可用于增强体内和体外的自噬。随后将这两种诱导肽处理体外培养的神经元,通过western blot检测p62与lc3的表达情况,gapdh为内参,结果发现tat-beclin1明显增强神经元自噬水平,同时它还明显促进突起的延伸(参见图6)。
74.进一步地,研究自噬是如何影响细胞骨架微管的动态性促进神经生长:将培养的cos7细胞用tat-scramble和tat-beclin1预处理24h后用浓度为0.1mm的nocodazole(微管解聚药物)处理15min,去除nocodazole的同时加入tat-scramble和tat-beclin1,统计突起总长度和分支数目,观察微管重新聚合的情况,计算t-tub(不稳定微管)/a-tub(稳定微管)的比值,n=30/组。结果表明,tat-beclin1通过增强细胞自噬促进微管的稳定性(参见图7-8),其中**表示p<0.01,***表示p<0.001。
75.综上可知,自噬可以通过增加微管稳定性促进脊髓损伤修复,但是调控自噬的关键蛋白并不明确。为了探寻同时参与脊髓损伤修复和细胞自噬的蛋白,通过构建了脊髓损伤模型和神经元自噬模型,将两种模型的差异表达蛋白进行维恩图分析,结果发现adk是同时与脊髓损伤和神经元自噬密切相关的唯一蛋白。
76.实施例2根据实施例1中所述反复进行大鼠脊髓损伤模型的制备,在损伤72h后选取各组大鼠脊髓t10(脊髓损伤造模处)为中心长约0.8cm的脊髓样品进行蛋白质谱分析,具体步骤如下:(1)将各组样品进行充分的蛋白酶解,随后进行除盐处理,得到蛋白样品。
77.(2)将各组蛋白样品用20-30ul 0.1% fa(金水配制)溶解,稀释5倍后测浓度,根据所测浓度调整需再加的0.1% fa的量,使样品终浓度为0.5ug/ul。
78.(3)将各组样品各取15μl 至新ep管离心12000g,20min。取上清12μl至新离心管12000g离心20min。取9.5μl上清至新离心管,加入0.5μl标肽(10
×
irt),涡旋混匀,离心12000g 10min后即可上样。
79.(4)将质谱数据进行搜库,获得的蛋白质信息进行蛋白注释,进而进行信号通路富
集性分析。
80.分析结果如图9所示。结果显示,在脊髓损伤后,脊髓内adk的表达水平呈现明显的下降趋势,且该下降的水平在一定程度上与脊髓的损伤程度呈现正相关。
81.随后,分别取各组小鼠脊髓组织利用western blot进行adk蛋白表达水平检测,具体步骤如下:(1)将玻璃板清洗干净,配好分离胶和浓缩胶,室温静置30min以保证完全凝胶;在buffer中拔掉胶板上的梳子,将变性的蛋白样品加入梳子孔中。
82.(3)上样完毕,恒压65-80v,当溴酚蓝指示带被压缩成一条线进入分离胶后,恒压120-140v,直至溴酚蓝指示带跑至凝胶末端处即停止电泳。
83.(4)电泳结束后,将pvdf膜泡在甲醇中2-4s,然后浸泡在1
×
transfer buffer中,同时将海绵、滤纸浸泡;打开电转印夹,黑色朝下,铺上一层浸泡好的海绵和滤纸,将凝胶平铺上,用镊子小心夹起pvdf膜平放在凝胶上,附上一层滤纸,左手固定住滤纸上下角,右手拿小玻璃管驱赶气泡,注意不可将手指压到膜的中间,盖上海绵,电转印夹插入电转槽,连接好电极,恒压100v,75min。
84.(5)转膜完成后,将膜放入1
×
tbs中漂洗,放入5%脱脂牛奶的封闭液中,置于摇床上,室温封闭90min。
85.(6)将封闭后的转印膜放入1
×
tbs中漂洗,一抗(adk、gapdh)1:1000稀释,用封闭液配制1ml稀释的抗体,将抗体稀释液滴在自制的湿盒薄膜上,将转印膜倒贴在上面,确认膜下没有气泡,用保鲜膜封住湿盒以免膜干掉,4℃孵育过夜或室温(20-25℃)孵育3h;用1
×
tbst洗膜3次,每次15min,同样方法配制二抗(1:15000稀释),湿盒里室温孵育2h;用1
×
tbst洗膜3次,每次15min。
86.(7)ecl法检测:首先将发光液a和发光液b等体积混合,然后将洗好的转印膜倒贴在混好的发光液上,避光室温孵育2min进行显影。
87.检测结果图10所示。结果显示,与上述蛋白质谱分析结果相一致,在脊髓损伤后,脊髓内adk的表达水平呈现明显的下降趋势,且该下降的水平在一定程度上与脊髓的损伤程度呈现正相关,其中*表示p<0.05,**表示p<0.01,***表示p<0.001,vs sham组。进一步地,取上述各组脊髓损伤模型样品进行免疫荧光染色分析,结果同样显示脊髓损伤后adk的表达呈现明显的下降趋势(参见图11)。由此可以明确,adk与脊髓损伤修复过程存在密切相关性。
88.为了验证adk如何影响神经元的生长和脊髓损伤修复,采用adk的抑制剂(abt 702或5-iodotubercidin)来处理神经元和脊髓损伤的动物,具体步骤如下:(1)体外培养海马神经元细胞,24h后分别用5-iodotubercidin、bafilomycin a1(自噬抑制剂)、torin(自噬诱导剂)对海马神经元细胞进行处理;(2)继续培养48h后,对海马神经元细胞进行固定并拍照,标尺为40μm;同时统计各组突起总分支、一级和二级分支数目以及各组神经元突起总长度、一级和二级分支突起总长度,n=30/组。
89.结果如图12-13所示。结果显示,5-iodotubercidin、自噬抑制剂(bafilomycin a1)和自噬诱导剂(torin)分别处理体外培养的海马神经元,torin处理的神经突起长度和二级分支数目较对照组明显增加,说明增强自噬可促进神经元突起的生长;且抑制adk的生
(ulk 1/2)复合物的重要组成部分。在自噬的起始阶段,ulk 1/2复合物负责招募下游atg蛋白并促进自噬前体的形成。atg101是机体调控自噬的关键蛋白,它可与atg13发生相互作用共同调控细胞自噬。
101.接下来,通过pull down的方法进一步验证atg101是否与adk蛋白发生相互作用。结果如图22所示,结果发现,将纯化后的gst-atg101蛋白与脊髓蛋白裂解液进行相互作用后,atg101可与脊髓中adk蛋白相结合。随后,通过对体外培养3天的海马神经元细胞进行免疫荧光染色分析发现, atg101与adk存在共定位的现象;而对大鼠脊髓组织切片的免疫荧光染色同样显示atg101与adk也存在共同表达的情况(参见图23),上述结果进一步证实了两者蛋白可能通过相互作用参与脊髓损伤修复。
102.为了揭示adk和atg101如何共同调控损伤神经元再生,通过在神经损伤模型中运用过表达的方式观察adk和atg101对突起生长的调节:在海马神经元培养的第二天,同时过表达gfp flag(control),gfp-adk flag,gfp flag-atg101和 gfp-adk flag-atg101。在转染后48h固定细胞并进行免疫荧光染色,测量神经元突起的长度及分支数量。
103.结果如图24-25所示。结果显示,与对照组相比,过表达adk可抑制损伤神经元修复,无论是一级分支、二级分支和突起总分支的伸展长度,adk都是发挥抑制的作用;同时,adk还可抑制神经元突起的分支。如果损伤神经元共表达adk和atg101,那么atg101可逆转atg对损伤神经元修复的抑制作用,其中*表示p<0.05,**表示p<0.01,***表示p<0.001,vs control组。综上所述,这些结果表明,adk和atg101可以共同调控损伤神经元的修复,过表达atg101可以逆转过表达adk对损伤神经元修复的抑制效果。
104.adk是一种进化保守的磷酸转移酶,可将嘌呤核糖核苷腺苷转化为5'-腺苷一磷酸。这种酶促反应在确定腺苷基调方面起着基本作用,其在所有生命系统中均起着稳态和代谢调节剂的作用。有文献报道在急性脑损伤后adk的短暂下调可保护大脑免受癫痫发作和细胞死亡的影响。所以,脊髓损伤后adk的下调,是否也能保护损伤的脊髓,这值得进行深入研究。
105.adk是否以及如何影响神经元的生长和脊髓损伤修复的并不清楚。abt-702是一种非核苷类adk抑制剂,不仅在动物疼痛模型中显示镇痛作用,而且对脊髓损伤后肢体的运动功能的恢复也有一定的促进作用。5-iodotubercidin亦是一种adk抑制剂,其能够促进神经干细胞的增殖。因此,为了研究adk对神经元的生长和脊髓损伤修复,本发明采用了adk抑制剂abt-702和5-iodotubercidin来处理神经元和脊髓损伤的动物。结果表明脊髓损伤中,adk的表达明显下降,在利用adk抑制剂抑制adk的生物学效应后,可以明显增加神经元突起长度及神经元突起数量;另一方面,运用adk抑制剂处理脊髓损伤的大鼠模型后,发现adk抑制剂在不影响动物体重的前提下,adk抑制剂显著增强脊髓损伤动物下肢功能的修复。
106.抑制adk与促进自噬对神经元生长的促进作用是一致的,然而adk是否参与调控自噬促进神经元生长则并不明确。lc3-ii定位于前自噬体和自噬体,是自噬体的标志分子,随自噬体膜的增多而增加。经常应用lc3-ii/lc3-i用于监测自噬囊泡的水平。当自噬被抑制时自噬体积累,p62水平升高。通过检测lc3 i和lc3 ii等自噬标志蛋白的表达发现,敲低adk表达可引起p62蛋白的表达量显著下降,同时lc3 ii/ lc3 i的比值明显升高,说明通过干扰adk蛋白的表达可以明显增强细胞自噬;此外,本发明通过adk蛋白激酶的抑制剂5-iodotubercidin处理细胞,结果显示5-iodotubercidin抑制adk活性可明显增强细胞自噬
水平,这个结果与干扰细胞中adk表达的结果相类似,以上结果表明adk的确参与调控细胞自噬。为了揭示adk和atg101如何共同调控损伤神经元再生,发明利用神经损伤模型运用过表达的方式观察adk和atg101对突起生长的调节。并发现损伤神经元共表达adk和atg101,且atg101可逆转atg对损伤神经元修复的抑制作用。上述结果表明,adk和atg101可以共同调控损伤神经元的修复,过表达atg101可以逆转过表达adk对损伤神经元修复的抑制效果。
107.脊髓损伤是一种破坏性创伤,常常导致患者感觉、运动和自主神经功能丧失。由于脊髓损伤的修复机制并不十分清晰,所以脊髓损伤患者的治疗效果往往并不令人满意。细胞自噬是细胞内的一种依赖溶酶体的细胞降解途径,是机体内存在的一种自我修复和维持生命的过程。细胞自噬可调节细胞骨架微管的动态性促进脊髓损伤修复,但是通过何种途径调控微管动态性促进脊髓损伤修复以及何种因素调控细胞自噬参与脊髓损伤修复等机制不明。所以,阐明自噬在脊髓损伤的信号转导机制,对探寻脊髓损伤治疗的药物靶点具有特别重要的意义。因此,本发明通过脊髓损伤动物模型和神经元自噬模型的蛋白质组学联合分析发现adk同时参与了细胞自噬与脊髓损伤修复,进一步通过多种实验手段验证了adk通过自噬参与脊髓损伤修复。本发明也发现调控adk可促进损伤脊髓的修复。另外,相互作用蛋白质谱和分子实验显示adk可以与自噬相关蛋白101(atg101)相结合。综上所述,本发明将阐明了adk通过atg101调控自噬参与脊髓损伤修复的机制,为确立adk作为脊髓损伤治疗的新靶点提供充分的科学依据,具有广阔的临床应用前景。
108.以上具体实施方式部分对本发明所涉及的分析方法进行了具体的介绍。应当注意的是,上述介绍仅是为了帮助本领域技术人员更好地理解本发明的方法及思路,而不是对相关内容的限制。在不脱离本发明原理的情况下,本领域技术人员还可以对本发明进行适当的调整或修改,上述调整和修改也应当属于本发明的保护范围。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。