基于基因编辑的低植物雌激素大豆
1.序列表
2.本技术包含序列表,该序列表已以ascii格式电子提交,并通过引用将其全部内容并入本文。创建于2020年9月18日的所述ascii副本,名为1702_042pct1_sl.txt,大小为33,318字节。
背景技术:
3.大豆是一种经济且丰富的供人类食用的蛋白质来源。来源于豆粕的蛋白质提取物具有良好的氨基酸分布和高蛋白质含量。大豆提取物以及整粒大豆被广泛用于各种消费品。大豆提取蛋白是婴儿配方食品的重要组成部分;例如,含有大豆蛋白的配方对乳糖不耐症婴儿很有价值。
4.植物产生多种次生化学物质,以及对正常植物发育和生理至关重要的基本分子。异黄酮是一类由豆科植物产生的具有多种功能的次生植物化学物质。异黄酮也被描述为植物雌激素,因为其是植物来源的类似雌激素的化合物。因此,已对异黄酮对人类荷尔蒙和发育系统的潜在影响进行研究。大豆蛋白提取物还含有植物次生代谢物,包括各种异黄酮(特别是染料木黄酮和黄豆苷元),其对诸如成熟女性的消费人群有益,在预防围绝经期综合征方面有潜在益处。然而,这些类似雌激素的异黄酮可能对包括婴儿和蛋白质补充剂的消费者的其他人群来说是不希望的。
5.异黄酮类化合物的生产代表了豆科植物苯丙素类途径的分支,该途径还包含几种其他重要植物化学物质(包括木质素、花青素类和芪类化合物)的生物合成途径。已知异黄酮参与与其他生物体的相互作用,包括根瘤菌的结瘤和病原体反应。
6.已经使用诸如靶向诱导基因组局部损伤(tilling)、转基因过表达或转基因抑制的技术来实现植物中异黄酮水平的改变(参见例如us 8,329,995、us 5,994,075、us 7,098,011)。然而,这些方法无法提供不含意外突变或转基因dna序列的产品,也无法满足消费者需求和公众接受度。
7.与现有品种相比,存在对异黄酮含量较低的大豆植物的需求。
技术实现要素:
8.本发明的方面涉及靶向诱变方法,特别是基因组编辑,并且排除了仅基于通过传统育种方法产生植物的实施方案(即,植物和植物材料产品(包括种子)不是专门通过本质上生物学的方法获得的,并且方法特征不限定本质上生物学的方法)。如本文所解释的,本文公开的大豆植物、植物部分和植物细胞中异黄酮含量较低或降低不是由于转基因的存在。
9.在一方面,本文提供了一种与未改变的大豆植物、植物部分或植物细胞相比具有降低的异黄酮含量的大豆植物、植物部分或植物细胞,其中所述大豆植物、植物部分或植物细胞在ifs1和ifs2基因之一或两者的编码区中包含一个或多个突变,并且其中所述植物、植物部分或植物细胞的特征在于ifs1和ifs2基因之一或两者的表达降低。
10.在另一方面,本文提供了一种与未改变的大豆植物、植物部分或植物细胞相比具有降低的异黄酮含量的大豆植物、植物部分或植物细胞,其中所述大豆植物、植物部分或植物细胞在myb29基因的编码区中的突变编码区中包含一个或多个突变,并且其中所述植物、植物部分或植物细胞的特征在于ifs2基因和ifs1基因之一或两者的表达降低表达降低。
11.在另一方面,本文提供了一种与未改变的大豆植物、植物部分或植物细胞相比具有降低的异黄酮含量的大豆植物、植物部分或植物细胞,其中所述大豆植物、植物部分或植物细胞在ifs1启动子或ifs2启动子的mybcore结合基序的编码区中包含一个或多个突变,并且其中所述植物、植物部分或植物细胞的特征在于ifs2基因和ifs1基因之一或两者的表达降低表达降低。
12.在本文公开的大豆植物、植物部分或植物细胞的一些实施方案中,所述一个或多个突变通过使用稀有切割核酸内切酶的靶向基因组修饰引入。在一些实施方案中,所述稀有切割核酸内切酶是转录激活因子样效应物核酸酶(talen)。在一些实施方案中,所述一个或多个突变通过使用包含与seq id no:1或seq id no:2内的序列结合的tal效应物的talen引入。在一些实施方案中,所述一个或多个突变通过使用包含与seq id no:3或seq id no:4内的序列结合的tal效应物的一种或多种talen引入。在一些实施方案中,所述一个或多个突变通过使用包含与seq id no:18内的序列结合的tal效应物的一种或多种talen引入。在一些实施方案中,所述一个或多个突变通过使用包含与seq id no:5-17和seq id no:19-35中的一个和多个结合的tal效应物的一种或多种talen引入。
13.在本文所述的大豆植物、植物部分或植物细胞的一些实施方案中,所述植物、植物部分或植物细胞不包含转基因。在一些实施方案中,所述植物部分是种子。
14.在一些实施方案中,所述植物、植物部分或植物细胞的异黄酮含量与相应的未改变的大豆植物、植物部分或植物细胞中的异黄酮含量相比降低了约10%、约20%、约30%、约50%、约60%、约70%、约80%或约90%。
15.在另一个方面,本文提供了一种使用靶向基因组修饰产生与未改变的植物、植物部分或植物细胞相比异黄酮含量降低的大豆植物、植物部分或植物细胞的方法,该方法包括在myb29基因的编码区、ifs2启动子的mybcore结合基序、ifs1启动子的mybcore结合基序、ifs2基因的编码区、ifs1基因的编码区或其组合中引入一个或多个突变。
16.在一些实施方案中,所述突变通过使用稀有切割核酸内切酶引入。在一些实施方案中,所述稀有切割核酸内切酶是talen。在一些实施方案中,所述一个或多个突变通过使用包含与seq id no:1或seq id no:2内的序列结合的tal效应物的talen引入。在一些实施方案中,所述一个或多个突变通过使用包含与seq id no:3或seq id no:4内的序列结合的tal效应物的talen引入。在一些实施方案中,所述一个或多个突变通过使用包含与seq id no:18内的序列结合的tal效应物的talen引入。在一些实施方案中,所述一个或多个突变通过使用包含与seq id no:5-17和seq id no:19-35中的一个和多个结合的tal效应物的talen引入。
17.在本文公开的方法的一些实施方案中,所述大豆植物、植物部分或植物细胞不包含转基因。
附图说明
18.由于通过参考以下详细描述并结合附图,可以更好地理解本发明的以上各方面和许多伴随的优点,由此使其得以更容易明白,在附图中:
19.图1a-b示出以下编码序列:(a)大豆ifs1(seq id no:1)和(b)大豆ifs2(seq id no:2)。
20.图2示出由位于靶序列侧翼的左半和右半talen靶向的ifs序列(分解图中示出了更大的分辨率)。图中分别按出现顺序公开了seq id nos 44-49。
21.图3是myb29转录因子序列和相关talen的图示。图中公开了seq id no:50。
22.图4示出ifs1启动子序列(seq.id no:3),其中突出显示了四个独特的mybcore基序(cngttr)。
23.图5是具有mybcore基序的ifs1启动子的图示。
24.图6示出具有鉴定的mybcore基序的ifs2启动子序列(seq.id no:4)。
25.图7是具有mybcore和talen结合位点的ifs2启动子的图示。图中公开了seq id no:51。
26.图8示出myb29的编码序列(seq.id no:18)。
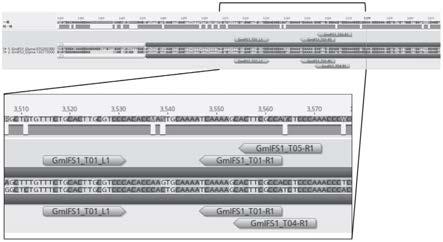
27.图9示出来自再生t0植物的已确认的突变谱(-25)的代表性ifs1 dna序列。图中公开了seq id no:36。
28.图10示出来自再生t0植物的已确认的突变谱(-34)的代表性ifs1 dna序列。图中公开了seq id no:37。
29.图11示出来自再生t0植物的已确认的突变谱(-4)的代表性ifs1 dna序列。图中公开了seq id no:38。
30.图12示出来自再生t0植物的已确认的突变谱(-4)的代表性ifs2 dna序列。图中公开了seq id no:39。
31.图13示出来自再生t0植物的已确认的突变谱(-38)的代表性ifs2 dna序列。图中公开了seq id no:40。
32.图14示出来自再生t0植物的已确认的突变谱(-4)的代表性ifs2 dna序列,命名为“ifs2.2”。“ifs2.2”名称表示独特的变体。在这种情况下,该缺失位点不同于图12中描述的-4nt缺失突变体。图中公开了seq id no:41。
33.图15示出来自再生t0植物的已确认的突变谱(-5)的代表性ifs2 dna序列。图中公开了seq id no:42。
34.图16示出来自再生t0植物的已确认的突变谱(-5)的代表性ifs2 dna序列,命名为“ifs2.2”。该缺失位点不同于图15中描述的-5nt缺失突变体。图中公开了seq id no:43。
35.图17示出植物雌激素分析的结果,该分析对从三个突变植物系(指定为#1-#3)的种子中提取的黄豆苷、黄豆黄苷和染料木苷的量与野生型种子提取物(对照)中它们的量进行了比较,所述量以干植物材料的μg/mg表示。
36.图18是图17中提供的值的图形表示。
37.图19示出植物雌激素含量分析的结果,该分析对从三个突变植物系(指定为#1-#3)的种子中提取的总植物雌激素量与野生型种子提取物(对照)中的该量进行了比较,所述量以干植物材料的μg/mg表示。
38.图20示出对照提取物(上)和样品#1提取物(下)中异黄酮组分的代表性hplc色谱图。加框区域示出在对照中分别与黄豆苷、黄豆黄苷和染料木苷相关的吸光度峰明显大于样品#1。
具体实施方式
39.本公开集中于选择性靶向负责产生异黄酮的最终生物合成酶异黄酮合酶(ifsl和ifs2)以及调节ifs1和ifs2的表达的基因(例如myb29)和调节序列(例如启动子)。
40.本公开满足了消费者对具有较低异黄酮含量的大豆蛋白和油产品的需求。本公开进一步满足了消费者对具有较低异黄酮含量的非转基因大豆蛋白和油产品的需求。
41.因此,在一方面,本文提供了与未改变的大豆植物、植物部分或植物细胞相比具有降低的异黄酮含量的大豆植物、植物部分和植物细胞,本公开的大豆植物、植物部分和植物细胞在异黄酮合成酶(ifs)基因ifs1和ifs2的编码区中包括一个或多个突变。本文公开的大豆植物、植物部分和植物细胞无转基因,并且在与靶标ifs1和ifs2同源的基因中无修饰。
42.在另一方面,本文提供了一种与未改变的大豆植物、植物部分或植物细胞相比具有降低的异黄酮含量的大豆植物、植物部分和植物细胞,本公开的的大豆植物、植物部分和植物细胞在基因myb29中包括一个或多个突变,myb29是在ifs1和ifs2的受控表达中重要的转录调节因子。
43.在另一方面,本文提供了一种与未改变的大豆植物、植物部分或植物细胞相比具有降低的异黄酮含量的大豆植物、植物部分或植物细胞,其在ifsl和/或ifs2的调控区中包含一个或多个突变。在一些实施方案中,调控区是启动子区。
44.在另一方面,本文提供了一种与未改变的大豆植物、植物部分或植物细胞相比具有降低的异黄酮含量的大豆植物、植物部分或植物细胞的生产方法,该方法包括通过在ifs1的编码区、ifs2的编码区、ifs1基因的调控区、ifs2基因的调控区、myb29基因或其组合中引入一个或多个突变来进行靶向基因编辑,其中ifs1和/或ifs2基因的表达被消除。在一些实施方案中,基因编辑使用基因的talen编辑来进行,以消除大豆种子中的ifs1和/或ifs2基因的表达而不破坏大豆根中的表达,其中异黄酮对大豆有益。
45.发明人发现,ifs1和ifs2基因序列具有可以同时被单个稀有切割核酸内切酶如talen靶向的区域。发明人进一步分析了来自几个大豆品种的异黄酮合成酶基因序列,并鉴定了保守区域以进一步聚焦稀有切割核酸内切酶诸如talen的靶位点。因此,经验证用于ifs1和ifs2基因编辑的稀有切割核酸内切酶诸如talen可用于多种商业大豆品系。talen编辑下游产生的植物与在类似条件下生长的标准大豆品种相比,异黄酮明显减少。在一些实施方案中,本文公开的大豆植物、植物部分或植物细胞的异黄酮含量与相应的未改变的大豆植物、植物部分或植物细胞的异黄酮含量相比,减少约10%、约20%、约30%、约50%、约60%、约70%、约80%、约90%、约95%、约98%或约99%。如本文所用,“未改变的大豆”是指未被基因编辑技术例如本文公开的talen编辑改变的大豆。未改变的植物包括野生型植物。在一些实施方案中,大豆植物部分是大豆种子。
46.因此,本公开提供了一种以前未知的用于在多个大豆品种中特异性靶向异黄酮合成酶基因的方法,该方法以不会导致整个基因组的其他位置发生突变的方式进行,并且不会在最终的选择编辑的品系中引入外来物质。此外,可以将源自基因编辑技术的植物与抗
拒细胞生物学再生的优良品系杂交。
47.ifs1和ifs2基因的表达已显示至少部分受myb29转录因子的调节(chu s.et al.,plos genet.13(5)(2017))。myb29是转录激活因子,其结合启动子区域中的mybcore序列基序并上调下游基因例如ifs1/ifs2的表达。因此,本文公开了通过在myb29基因中引入突变从而减少或消除ifs1和ifs2基因中的一种或多种的表达来产生大豆植物、植物部分或植物细胞的方法。此外,本文公开了一种大豆植物、植物部分或植物细胞,其在myb29基因中包含一种或多种突变,其中大豆植物、植物部分或植物细胞的ifs1和ifs2基因中的一种或多种的表达减少或消除。
48.发明人鉴定了ifs1和ifs2的启动子区域中的mybcore序列,以及设计用于切除这些结合基序的talen。来自ifs1和/或ifs2启动子的mybcore结合基序的切除特别令人感兴趣,因为myb29基因可以保持不变并具有完全功能,能够调节其他靶基因,同时消除其对ifs1和/或ifs2的调节。
49.在本文公开的方法、植物、植物部分和植物细胞的一些实施方案中,一个或多个突变通过使用稀有切割核酸内切酶的靶向基因组修饰引入。本文中的术语“稀有切割核酸内切酶”是指具有针对长度约12-40bp(例如,长度14-40bp)的具有识别序列(靶序列)的核酸序列的核酸内切酶活性的天然或工程蛋白质。典型的稀有切割核酸内切酶在其识别位点内引起切割,留下4个核苷酸交错切口,带有3'oh或5'oh突出端。这些稀有切割核酸内切酶可以是兆核酸酶,诸如归巢核酸内切酶的野生型或变体蛋白,更特别地属于十二肽家族(参见wo 2004/067736),或可能来自将dna结合结构域与具有切割活性的催化结构域相连接的融合蛋白。tal效应物核酸内切酶和锌指核酸酶(zfn)是dna结合结构域与核酸内切酶foki的催化结构域融合的例子。有关稀有切割核酸内切酶的评述,请参见baker,nature methods 9:23-26,2012。
[0050]“稀有切割核酸内切酶”也涵盖可用于本文提供的方法中的rna引导系统。例如,成簇的规则间隔短回文重复/crispr相关(crispr/cas)系统使用rna来引导dna切割(参见例如belahj et al.,plant methods 9:39,2013)。这些系统由cas9核酸内切酶和指导rna(crispr rna(crrna)和反式激活crrna(tracrrna)之间的复合物,或crrna的3'端和tracrrna的5'端之间的合成融合)。指导rna将cas9结合和dna切割引导至与原型间隔区相邻基序(pam)相邻的序列,例如,用于来自化脓链球菌(streptococcus pyogenes)的cas9的ngg。一旦到达目标dna序列,cas9在从与目标序列互补的crrna序列的3'端的三个核苷酸的位置产生dna双链断裂。除了cas9之外,来自普雷沃菌属(prevotella)和弗朗西斯氏菌属(francisella)1(cpf1)的crispr系统可用于引入突变(参见例如zetsche et al.,cell 163:759-771(2015))。
[0051]
在本文公开的方法、植物、植物部分和植物细胞的一些实施方案中,所述一个或多个突变通过使用包含与seq id no:1(图1a)内的序列结合的tal效应物的talen引入。在本文公开的方法、植物、植物部分和植物细胞的一些实施方案中,所述一个或多个突变通过使用包含与seq id no:2(图1b)内的序列结合的tal效应物的talen引入。在本文公开的方法、植物、植物部分和植物细胞中,所述一个或多个突变通过使用包含与seq id no:3(图4)内的序列结合的tal效应物的talen引入。在本文公开的方法、植物、植物部分和植物细胞中,所述一个或多个突变通过使用包含与seq id no:4(图6)内的序列结合的tal效应物的
talen引入。在本文公开的方法、植物、植物部分和植物细胞中,所述一个或多个突变通过使用包含与seq id no:18(图8)内的序列结合的tal效应物的talen引入。在一些实施方案中,所述一个或多个突变通过使用包含与选自下表1中所示的seq id no:5-17和19-35的序列结合的tal效应物的一种或多种talen引入。
[0052]
表1:tal效应物核酸内切酶靶序列
[0053][0054][0055]
在一些实施方案中,talen包含tal效应物,其结合本文公开的序列,例如seq id no:1-4和18,的功能变体内的序列。如本文所用的术语“核酸或蛋白质序列的功能变体”,例如以seq id no:1-4和18作为参考,是指保留非变体基因的生物学功能的变体基因序列或基因序列的一部分。功能变体也包括编码多肽的目的基因的变体,其具有不影响所得蛋白质功能的序列改变,例如在非保守残基中。也包含在内的有与未改变的例如野生型的本文
所示的基因的核酸序列基本相同的变体,即,仅具有一些序列变异,例如在非保守残基中,并且其具有生物活性。如本文所用,根据本发明的各个方面的特定核苷酸或氨基酸序列的变体可具有与本文公开的基因的特定非变异核苷酸序列,例如seq id no:1-4和18之一的至少约50%至至少约99%,例如至少约75%、至少约85%、至少约86%、至少约87%、至少约88%、至少约89%、至少约90%、至少约92%、至少约94%、至少约95%、至少约96%、至少约97%、至少约98%,或至少约99%或更多的序列同一性。序列同一性可以使用序列比对程序确定,该程序是本领域中众所周知的。
[0056]
talen基因编辑基于对含有dna识别结构域和在质粒中编码的foki核酸内切酶结构域的两个半talen蛋白的基因特异性靶向,质粒包括适当的启动子、终止子和在受体植物细胞中表达所必需的其他非编码序列。foki仅在二聚体组合中具有活性,并且需要两个半talen来启用talen基因编辑所必要的核酸内切酶dna切割。talen引入到细胞,例如大豆(glycine max)细胞,可以通过本领域技术人员已知的多种方式来完成。引入基因编辑talen的常用方法包括用携带包含talen基因盒的质粒的农杆菌属物种转染、用编码talen序列盒的质粒或mrna进行基因枪轰击、用编码talen序列盒的质粒或mrna进行peg介导的转化,以及通过包括peg介导的转染的多种方法引入talen蛋白。
[0057]
因此,与来自未改变植物的相应植物部分或产品相比,本文提供的方法可用于生产异黄酮含量降低的植物部分(例如种子)或植物产品(例如粗粉、油)。本发明将在以下实施例中进一步描述,其并不限制权利要求中描述的本发明的范围。
[0058]
在一些实施方案中,在ifs1和ifs2基因中具有内切核酸酶诱导的突变的大豆植物的再生可以如下完成。在一些实施方案中,通过递送设计成靶向ifs1和ifs2两者的一种talen(对)可以敲除ifs1和ifs2基因。为了实现这一点,将ifs1和ifs2 talen克隆到合适的dna载体中。可以通过农杆菌介导的转化或使用基因枪将这些载体递送至植物细胞。可以使用标准转化方法产生表达tal效应物核酸内切酶的转基因大豆植物。将植物转移到土壤中,在大约4周的生长后,从每株植物收获一片小叶子,用于dna提取和基因分型。首先使用pcr筛选每个dna样品中是否存在t-dna。对ifs1和ifs2基因的进一步基于pcr的表征来评估是否发生突变。
[0059]
在一些实施方案中,具有内切核酸酶介导的ifs1或ifs2启动子中mybcore元件缺失的大豆植物的再生可以如下完成。对ifs1和ifs2的上游区域进行序列分析以鉴定具有序列cngttr的mybcore元件,其中n是任何核苷酸,r是嘌呤(g或a)。对于ifs1启动子,例如,鉴定了四个mybcore基序并设计了talen对(图5)。对于ifs2启动子,例如,鉴定了一个mybcore基序并设计了talen对(图7)。将靶向ifs1启动子和ifs2启动子或这两个启动子二者的一对或多对talen克隆到合适的dna载体中,递送至植物细胞,该植物细胞再生为转基因小植株。在土壤中生长大约4周后,从每株植物中收获一片小叶子进行dna提取和基因分型。首先使用pcr筛选每个dna样本中是否存在t-dna。对ifs1和ifs2基因的启动子区域的进一步基于pcr的表征来评估是否发生突变。
[0060]
在一些实施方案中,在myb29中具有内切核酸酶诱导的突变的大豆植物的再生可以如下完成。分析myb29的编码序列并且设计talen以诱导功能缺失的损失,其将阻止myb29诱导ifs1或ifs2表达。将设计成与myb29的外显子2中的靶序列(例如,在seq id no:18内)结合的talen对被递送至大豆细胞并再生为含有所需突变的植物。
[0061]
本发明将在以下实施例中进一步描述,其并不限制权利要求中描述的本发明的范围。
[0062]
实施例
[0063]
进行实验以产生在ifs1和ifs2中具有突变的大豆植物。为了实现这一点,将gmifs1_t01 tal效应物核酸内切酶对(seq id no:5和6(表1))克隆到细菌载体中,其中由胭脂碱合酶(nos)启动子驱动tal效应物核酸内切酶表达,并通过农杆菌介导的转化或使用基因枪递送到大豆植物细胞。
[0064]
使用标准转化方法产生表达tal效应物核酸内切酶的转基因大豆植物。用编码tal效应物核酸内切酶对的序列转化野生型大豆半子叶后,再生假定的转基因植物。将植物转移到土壤中,大约生长4周后,从每株植物中收获一片小叶子用于dna提取和基因分型。产生多个事件。然后对所有t0转基因阳性植物进行分析,以鉴定在tal效应物核酸内切酶识别位点发生突变的植物。
[0065]
然后将阳性植物的ifs1和ifs2 pcr扩增子插入克隆载体并进行sanger测序以确认和表征突变体谱。然后将所得读数与野生型序列比对以确定等位基因类型。代表性序列示于图9-16(seq id no:36-43)。
[0066]
为了确定由在大豆ifs1和ifs2基因中tal效应物核酸内切酶诱导的突变产生的种子上的植物雌激素谱,使大豆品系生长至成熟并使其自体受精,从而产生在ifs1和ifs2基因中为纯合突变体的种子。通过测定总植物雌激素含量和黄豆苷、黄豆黄苷和染料木苷的量来分析这些种子的植物雌激素含量。简而言之,将来自三个突变植物系(#1-#3)和野生型植物系(对照)的各个大豆种子用研钵和研杵粉碎,用80%甲醇从粉碎的组织中提取植物雌激素。突变株系的提取物呈现强烈的黄色色调,而野生型种子提取物呈奶油色(未示出)。对提取物进行离心并过滤上清液。水解过滤的样品,使用定量hplc分析(260nm作为标准吸收波长)确定非酯化植物雌激素形式。代表性色谱图如图20所示。与对照种子相比,分析结果显示每个突变系产生的种子的总植物雌激素显著降低(总植物雌激素变化55-100倍(图17和19)),并且黄豆苷、黄豆黄苷和染料木苷的每一种(图17和18)也显著降低。例如,与野生型种子相比,样品#3的种子显示出黄豆苷降低98.87%,黄豆黄苷降低97.56%,染料木苷降低99.30%(图17)。总植物雌激素在样品#1中降低98.25%,在样品#2中降低98.57%,在样品#3中降低99.07%(图19)。
[0067]
除非另有说明,如本文所用的术语“约”表示相关值可以被修改正负百分之五( /-5%)并且保持在所公开的实施方案的范围内。
[0068]
本文公开的本发明的替代要素或实施方案的分组不应被解释为限制。每个组成员可以单独或与组的其他成员或本文发现的其他要素的任何组合被提及和要求保护。出于方便和/或可专利性的原因,预计一个组的一个或多个成员可能被包含在一个组中或从组中删除。当任何此类包含或删除发生时,本说明书被视为包含修改后的组,从而满足所附权利要求中使用的所有马库什组的书面描述。
[0069]
除非另有说明,核酸或寡核苷酸以5'到3'方向从左到右书写。
[0070]
除非另有说明,否则当在本技术中使用时,词语“本文”、“上文”和“下文”以及类似含义的词语是指本技术的整体而不是本技术的任何特定部分。如本文所用的词语“约”和“左右”包括围绕所述值的微小变化,通常在标准误差范围内,例如在所述值的10%内。
[0071]
在此提及的所引用的专利、专利申请和科学文献通过引用的方式整体并入本文,就好像每个单独的出版物、专利或专利申请被具体地和单独地指示以引用的方式并入一样。
[0072]
虽然已经说明和描述了本发明的优选实施方案,但是应当理解,在不背离本发明的精神和范围的情况下可以在其中做出各种改变。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。