用于改进的单链可变片段的材料和方法
1.相关申请的交叉引用
2.本技术要求以下专利申请的权益:2019年12月11日提交的美国临时专利申请号62/946,897;2019年12月11日提交的美国临时专利申请号62/946,886;2019年12月11日提交的美国临时专利申请号62/946,882;2019年12月11日提交的美国临时专利申请号62/946,877;2019年12月11日提交的美国临时专利申请号62/946,865;2019年8月15日提交的美国临时专利申请号62/887,529;2019年8月15日提交的美国临时专利申请号62/887,527;2019年8月15日提交的美国临时专利申请号62/887,524;2019年8月15日提交的美国临时专利申请号62/887,519;以及2019年8月15日提交的美国临时专利申请号62/887,514,其各自以引用的方式全文并入本文。
3.序列表
4.本技术以引用的方式并入作为文本格式随本技术一起提交的序列表,该序列表的名称为“14620-227-228_sl.txt”,创建于2020年8月5日,大小为258,724字节。
1.技术领域
5.公开了用于改进的单链可变片段的材料和方法。
2.
背景技术:
6.抗原结合单链可变片段(scfv)是可以广泛用作治疗剂、成像剂、诊断剂或作为异源分子(诸如多特异性分子)的部分的模块。scfv的挑战之一是低稳定性和聚集趋势(综述于下列文献中:worn和pluckthun(2001)j mol biol 305:989-1010;rothlisberger等人,(2005)j mol biol 347:773-789;gross等人,(1989)transplant proc 21(1pt 1):127-130,porter等人,(2011)j cancer 2:331-332;porter等人,(2011)n engl j med 365:725-733)。因此,需要可以任选地结合到多特异性分子和异源分子中的改进的scfv设计。
3.
技术实现要素:
7.在一个方面,本公开提供了一种分离的单链可变片段(scfv),其包含重链可变区(vh)、接头(l)和轻链可变区(vl),其中该scfv包含
8.在结构保守的表面暴露的vh半胱氨酸(cys)与第一l cys之间的第一二硫键;
9.在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键;或者
10.在结构保守的表面暴露的vh cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键。
11.本公开还提供了一种分离的scfv,其包含vh、l和vl,其中
12.该vh包含在结构保守的表面暴露的vh框架残基位置处的vh cys,并且该l包含第一l cys;
13.该vl包含在结构保守的表面暴露的vl框架残基位置处的vl cys,并且该l包含第二l cys;或者
14.该vh包含在结构保守的表面暴露的vh框架残基位置处的vh cys,该vl包含在结构保守的表面暴露的vl框架残基位置处的vl cys,并且该l包含第一l cys和第二l cys,其中vh cys和第一l cys能够形成二硫键并且vl cys和第二l cys能够形成二硫键。
15.本公开还提供了一种scfv,其包含vh、l和vl,其中
16.该vh包含在h105处的cys;
17.该vl包含在l42处的cys;
18.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
19.该scfv处于vl-l-vh取向。
20.本公开还提供了一种scfv,其包含vh、l和vl,其中
21.该vh包含在h105处的cys;
22.该vl包含在l45处的cys;
23.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
24.该scfv处于vl-l-vh取向。
25.本公开还提供了一种scfv,其包含vh、l和vl,其中
26.该vh包含在h105处的cys;
27.该vl包含在l39处的cys;
28.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
29.该scfv处于vl-l-vh取向。
30.本公开还提供了一种scfv,其包含vh、l和vl,其中
31.该vh包含在h5处的cys;
32.该vl包含在l42处的cys;
33.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
34.该scfv处于vl-l-vh取向。
35.本公开还提供了一种scfv,其包含vh、l和vl,其中
36.该vh包含在h5处的cys;
37.该vl包含在l45处的cys;
38.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
39.该scfv处于vl-l-vh取向。
40.本公开还提供了一种scfv,其包含vh、l和vl,其中
41.该vh包含在h5处的cys;
42.该vl包含在l39处的cys;
43.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
44.该scfv处于vl-l-vh取向。
45.本公开还提供了一种scfv,其包含vh、l和vl,其中
46.该vh包含在h3处的cys;
47.该vl包含在l42处的cys;
48.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
49.该scfv处于vl-l-vh取向。
50.本公开还提供了一种scfv,其包含vh、l和vl,其中
51.该vh包含在h3处的cys;
52.该vl包含在l45处的cys;
53.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
54.该scfv处于vl-l-vh取向。
55.本公开还提供了一种scfv,其包含vh、l和vl,其中
56.该vh包含在h3处的cys;
57.该vl包含在l39处的cys;
58.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
59.该scfv处于vl-l-vh取向。
60.本公开还提供了一种scfv,其包含vh、l和vl,其中
61.该vh包含在h43处的cys;
62.该vl包含在l100处的cys;
63.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
64.该scfv处于vh-l-vl取向。
65.本公开还提供了一种scfv,其包含vh、l和vl,其中
66.该vh包含在h43处的cys;
67.该vl包含在l102处的cys;
68.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
69.该scfv处于vh-l-vl取向。
70.本公开还提供了一种scfv,其包含vh、l和vl,其中
71.该vh包含在h43处的cys;
72.该vl包含在l5处的cys;
73.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
74.该scfv处于vh-l-vl取向。
75.本公开还提供了一种scfv,其包含vh、l和vl,其中
76.该vh包含在h43处的cys;
77.该vl包含在l3处的cys;
78.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
79.该scfv处于vh-l-vl取向。
80.本公开还提供了一种scfv,其包含vh、l和vl,其中
81.该vh包含在h40处的cys;
82.该vl包含在l100处的cys;
83.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
84.该scfv处于vh-l-vl取向。
85.本公开还提供了一种scfv,其包含vh、l和vl,其中
86.该vh包含在h40处的cys;
87.该vl包含在l102处的cys;
88.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
89.该scfv处于vh-l-vl取向。
90.本公开还提供了一种scfv,其包含vh、l和vl,其中
91.该vh包含在h40处的cys;
92.该vl包含在l5处的cys;
93.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
94.该scfv处于vh-l-vl取向。
95.本公开还提供了一种scfv,其包含vh、l和vl,其中
96.该vh包含在h40处的cys;
97.该vl包含在l3处的cys;
98.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
99.该scfv处于vh-l-vl取向。
100.本公开还提供了一种scfv,其包含vh、l和vl,其中
101.该vh包含在h46处的cys;
102.该vl包含在l100处的cys;
103.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
104.该scfv处于vh-l-vl取向。
105.本公开还提供了一种scfv,其包含vh、l和vl,其中
106.该vh包含在h46处的cys;
107.该vl包含在l102处的cys;
108.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
109.该scfv处于vh-l-vl取向。
110.本公开还提供了一种scfv,其包含vh、l和vl,其中
111.该vh包含在h46处的cys;
112.该vl包含在l5处的cys;
113.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
114.该scfv处于vh-l-vl取向。
115.本公开还提供了一种scfv,其包含vh、l和vl,其中
116.该vh包含在h46处的cys;
117.该vl包含在l3处的cys;
118.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
119.该scfv处于vh-l-vl取向。
120.本公开还提供了一种包含本公开的spfv以及药学上可接受的载剂的药物组合物。
121.本公开还提供了一种编码本公开的spfv的多核苷酸。
122.本公开还提供了一种包含本公开的多核苷酸的载体。
123.本公开还提供了一种包含本公开的载体的宿主细胞。
124.本公开还提供了一种产生本公开的spfv的方法,包括在产生该spfv的条件下培养本公开的宿主细胞,以及纯化该spfv。
125.本公开还提供了一种与本公开的spfv结合的抗独特型抗体。
126.本公开还提供了一种包含本公开的spfv的试剂盒。
127.在另一方面,本公开提供了一种包含单链可变片段(scfv)的多特异性分子,该单
链可变片段包含重链可变区(vh)、接头(l)和轻链可变区(vl),其中该scfv包含
128.在结构保守的表面暴露的vh半胱氨酸(cys)与第一l cys之间的第一二硫键;
129.在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键;或者
130.在结构保守的表面暴露的vh cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键。
131.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
132.该vh包含在结构保守的表面暴露的vh框架残基位置处的vh cys,并且该l包含第一l cys;
133.该vl包含在结构保守的表面暴露的vl框架残基位置处的vl cys,并且该l包含第二l cys;或者
134.该vh包含在结构保守的表面暴露的vh框架残基位置处的vh cys,该vl包含在结构保守的表面暴露的vl框架残基位置处的vl cys,并且该l包含第一l cys和第二l cys,其中vh cys和第一l cys能够形成二硫键并且vl cys和第二l cys能够形成二硫键。
135.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
136.该vh包含在h105处的cys;
137.该vl包含在l42处的cys;
138.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
139.该scfv处于vl-l-vh取向。
140.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
141.该vh包含在h105处的cys;
142.该vl包含在l45处的cys;
143.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
144.该scfv处于vl-l-vh取向。
145.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
146.该vh包含在h105处的cys;
147.该vl包含在l39处的cys;
148.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
149.该scfv处于vl-l-vh取向。
150.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
151.该vh包含在h5处的cys;
152.该vl包含在l42处的cys;
153.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
154.该scfv处于vl-l-vh取向。
155.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
156.该vh包含在h5处的cys;
157.该vl包含在l45处的cys;
158.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
159.该scfv处于vl-l-vh取向。
160.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
161.该vh包含在h5处的cys;
162.该vl包含在l39处的cys;
163.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
164.该scfv处于vl-l-vh取向。
165.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
166.该vh包含在h3处的cys;
167.该vl包含在l42处的cys;
168.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
169.该scfv处于vl-l-vh取向。
170.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
171.该vh包含在h3处的cys;
172.该vl包含在l45处的cys;
173.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
174.该scfv处于vl-l-vh取向。
175.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
176.该vh包含在h3处的cys;
177.该vl包含在l39处的cys;
178.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
179.该scfv处于vl-l-vh取向。
180.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
181.该vh包含在h43处的cys;
182.该vl包含在l100处的cys;
183.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
184.该scfv处于vh-l-vl取向。
185.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
186.该vh包含在h43处的cys;
187.该vl包含在l102处的cys;
188.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
189.该scfv处于vh-l-vl取向。
190.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
191.该vh包含在h43处的cys;
192.该vl包含在l5处的cys;
193.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
194.该scfv处于vh-l-vl取向。
195.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
196.该vh包含在h43处的cys;
197.该vl包含在l3处的cys;
198.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
199.该scfv处于vh-l-vl取向。
200.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
201.该vh包含在h40处的cys;
202.该vl包含在l100处的cys;
203.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
204.该scfv处于vh-l-vl取向。
205.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
206.该vh包含在h40处的cys;
207.该vl包含在l102处的cys;
208.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
209.该scfv处于vh-l-vl取向。
210.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
211.该vh包含在h40处的cys;
212.该vl包含在l5处的cys;
213.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
214.该scfv处于vh-l-vl取向。
215.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
216.该vh包含在h40处的cys;
217.该vl包含在l3处的cys;
218.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
219.该scfv处于vh-l-vl取向。
220.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
221.该vh包含在h46处的cys;
222.该vl包含在l100处的cys;
223.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
224.该scfv处于vh-l-vl取向。
225.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
226.该vh包含在h46处的cys;
227.该vl包含在l102处的cys;
228.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
229.该scfv处于vh-l-vl取向。
230.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
231.该vh包含在h46处的cys;
232.该vl包含在l5处的cys;
233.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
234.该scfv处于vh-l-vl取向。
235.本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
236.该vh包含在h46处的cys;
237.该vl包含在l3处的cys;
238.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
239.该scfv处于vh-l-vl取向。
240.本公开还提供了一种包含本文所提供的多特异性分子以及药学上可接受的载剂的药物组合物。
241.在又一方面,本公开提供了一种包含单链可变片段(scfv)的异源分子,该单链可变片段包含重链可变区(vh)、接头(l)和轻链可变区(vl),其中该scfv包含
242.在结构保守的表面暴露的vh半胱氨酸(cys)与第一l cys之间的第一二硫键;
243.在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键;或者
244.在结构保守的表面暴露的vh cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键。
245.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
246.该vh包含在结构保守的表面暴露的vh框架残基位置处的vh cys,并且该l包含第一l cys;
247.该vl包含在结构保守的表面暴露的vl框架残基位置处的vl cys,并且该l包含第二l cys;或者
248.该vh包含在结构保守的表面暴露的vh框架残基位置处的vh cys,该vl包含在结构保守的表面暴露的vl框架残基位置处的vl cys,并且该l包含第一l cys和第二l cys,其中vh cys和第一l cys能够形成二硫键并且vl cys和第二l cys能够形成二硫键。
249.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
250.该vh包含在h105处的cys;
251.该vl包含在l42处的cys;
252.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
253.该scfv处于vl-l-vh取向。
254.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
255.该vh包含在h105处的cys;
256.该vl包含在l45处的cys;
257.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
258.该scfv处于vl-l-vh取向。
259.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
260.该vh包含在h105处的cys;
261.该vl包含在l39处的cys;
262.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
263.该scfv处于vl-l-vh取向。
264.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
265.该vh包含在h5处的cys;
266.该vl包含在l42处的cys;
267.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
268.该scfv处于vl-l-vh取向。
269.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
270.该vh包含在h5处的cys;
271.该vl包含在l45处的cys;
272.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
273.该scfv处于vl-l-vh取向。
274.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
275.该vh包含在h5处的cys;
276.该vl包含在l39处的cys;
277.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
278.该scfv处于vl-l-vh取向。
279.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
280.该vh包含在h3处的cys;
281.该vl包含在l42处的cys;
282.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
283.该scfv处于vl-l-vh取向。
284.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
285.该vh包含在h3处的cys;
286.该vl包含在l45处的cys;
287.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
288.该scfv处于vl-l-vh取向。
289.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
290.该vh包含在h3处的cys;
291.该vl包含在l39处的cys;
292.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
293.该scfv处于vl-l-vh取向。
294.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
295.该vh包含在h43处的cys;
296.该vl包含在l100处的cys;
297.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
298.该scfv处于vh-l-vl取向。
299.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
300.该vh包含在h43处的cys;
301.该vl包含在l102处的cys;
302.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
303.该scfv处于vh-l-vl取向。
304.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
305.该vh包含在h43处的cys;
306.该vl包含在l5处的cys;
307.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
308.该scfv处于vh-l-vl取向。
309.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
310.该vh包含在h43处的cys;
311.该vl包含在l3处的cys;
312.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
313.该scfv处于vh-l-vl取向。
314.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
315.该vh包含在h40处的cys;
316.该vl包含在l100处的cys;
317.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
318.该scfv处于vh-l-vl取向。
319.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
320.该vh包含在h40处的cys;
321.该vl包含在l102处的cys;
322.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
323.该scfv处于vh-l-vl取向。
324.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
325.该vh包含在h40处的cys;
326.该vl包含在l5处的cys;
327.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
328.该scfv处于vh-l-vl取向。
329.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
330.该vh包含在h40处的cys;
331.该vl包含在l3处的cys;
332.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
333.该scfv处于vh-l-vl取向。
334.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
335.该vh包含在h46处的cys;
336.该vl包含在l100处的cys;
337.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
338.该scfv处于vh-l-vl取向。
339.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
340.该vh包含在h46处的cys;
341.该vl包含在l102处的cys;
342.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
343.该scfv处于vh-l-vl取向。
344.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
345.该vh包含在h46处的cys;
346.该vl包含在l5处的cys;
347.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
348.该scfv处于vh-l-vl取向。
349.本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
350.该vh包含在h46处的cys;
351.该vl包含在l3处的cys;
352.该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
353.该scfv处于vh-l-vl取向。
354.本公开还提供了一种包含本公开的异源分子以及药学上可接受的载剂的药物组合物。
355.在又一方面,本公开提供了一种用于制备稳定化的scfv的方法,包括:
356.提供重链可变区(vh)和轻链可变区(vl),这两者形成抗原结合结构域;
357.提供接头(l),所述接头包含或被工程化为包含第一l cys;
358.工程化所述vh以在结构保守的表面暴露的vh框架残基位置处包含vh cys;以及
359.在所述vh cys与所述第一l cys之间形成二硫键,以制备所述稳定化的scfv。
360.本公开还提供了一种用于制备稳定化的scfv的方法,包括:
361.提供vh和vl,这两者形成抗原结合结构域;
362.提供l,所述l包含或被工程化为包含第二l cys;
363.工程化所述vl以在结构保守的表面暴露的vl框架残基位置处包含vl cys;以及
364.在所述vl cys与所述第二l cys之间形成二硫键,以制备所述稳定化的scfv。
365.本公开还提供了一种用于制备稳定化的scfv的方法,包括:
366.提供vh和vl,这两者形成抗原结合结构域;
367.提供l,所述l包含或被工程化为包含第一l cys和第二l cys;
368.工程化所述vh以在结构保守的表面暴露的vh框架残基位置处包含vh cys;
369.工程化所述vl以在结构保守的表面暴露的vl框架残基位置处包含vl cys;以及
370.在所述vh cys与所述第一l cys之间形成二硫键,并且在所述vl cys与所述第二l cys之间形成二硫键,以制备所述稳定化的scfv。
371.本公开还提供了一种用于制备稳定化的scfv的方法,包括
372.提供编码vh、l和vl的多核苷酸,其中
373.该vh包含在h105处的cys,并且该vl包含在l42处的cys;
374.该vh包含在h43处的cys,并且该vl包含在l100处的cys;
375.该vh包含在h3处的cys,并且该vl包含在l3处的cys;
376.该vh包含在h3处的cys,并且该vl包含在l5处的cys;
377.该vh包含在h3处的cys,并且该vl包含在l39处的cys;
378.该vh包含在h3处的cys,并且该vl包含在l42处的cys;
379.该vh包含在h3处的cys,并且该vl包含在l45处的cys;
380.该vh包含在h3处的cys,并且该vl包含在l100处的cys;
381.该vh包含在h3处的cys,并且该vl包含在l102处的cys;
382.该vh包含在h5处的cys,并且该vl包含在l3处的cys;
383.该vh包含在h5处的cys,并且该vl包含在l5处的cys;
384.该vh包含在h5处的cys,并且该vl包含在l39处的cys;
385.该vh包含在h5处的cys,并且该vl包含在l42处的cys;
386.该vh包含在h5处的cys,并且该vl包含在l45处的cys;
387.该vh包含在h5处的cys,并且该vl包含在l100处的cys;
388.该vh包含在h5处的cys,并且该vl包含在l102处的cys;
389.该vh包含在h40处的cys,并且该vl包含在l3处的cys;
390.该vh包含在h40处的cys,并且该vl包含在l5处的cys;
391.该vh包含在h40处的cys,并且该vl包含在l39处的cys;
392.该vh包含在h40处的cys,并且该vl包含在l42处的cys;
393.该vh包含在h40处的cys,并且该vl包含在l45处的cys;
394.该vh包含在h40处的cys,并且该vl包含在l100处的cys;
395.该vh包含在h40处的cys,并且该vl包含在l102处的cys;
396.该vh包含在h43处的cys,并且该vl包含在l3处的cys;
397.该vh包含在h43处的cys,并且该vl包含在l5处的cys;
398.该vh包含在h43处的cys,并且该vl包含在l39处的cys;
399.该vh包含在h43处的cys,并且该vl包含在l42处的cys;
400.该vh包含在h43处的cys,并且该vl包含在l45处的cys;
401.该vh包含在h43处的cys,并且该vl包含在l102处的cys;
402.该vh包含在h46处的cys,并且该vl包含在l3处的cys;
403.该vh包含在h46处的cys,并且该vl包含在l5处的cys;
404.该vh包含在h46处的cys,并且该vl包含在l39处的cys;
405.该vh包含在h46处的cys,并且该vl包含在l42处的cys;
406.该vh包含在h46处的cys,并且该vl包含在l45处的cys;
407.该vh包含在h46处的cys,并且该vl包含在l100处的cys;
408.该vh包含在h46处的cys,并且该vl包含在l102处的cys;
409.该vh包含在h105处的cys,并且该vl包含在l3处的cys;
410.该vh包含在h105处的cys,并且该vl包含在l5处的cys;
411.该vh包含在h105处的cys,并且该vl包含在l39处的cys;
412.该vh包含在h105处的cys,并且该vl包含在l45处的cys;
413.该vh包含在h105处的cys,并且该vl包含在l100处的cys;或者
414.该vh包含在h105处的cys,并且该vl包含在l102处的cys,其中残基根据chothia进行编号。
415.该l包含氨基酸序列seq id no:2、3、4、5、6或7;以及
416.在宿主细胞中表达所述多核苷酸,以产生所述稳定化的scfv。
417.在又一方面,本公开提供了一种分离的单链可变片段(scfv),其包含重链可变区(vh)、用于连接的构件(l)和轻链可变区(vl),其中该scfv包含:
418.在结构保守的表面暴露的vh半胱氨酸(cys)与第一l cys之间的第一二硫键;
419.在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键;或者
420.在结构保守的表面暴露的vh cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键。
421.本公开还提供了一种分离的单链可变片段(scfv),其包含用于抗原结合的构件、
接头(l)和轻链可变区(vl),其中该scfv包含
422.在结构保守的表面暴露的抗原结合构件半胱氨酸(cys)与第一l cys之间的第一二硫键;
423.在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键;或者
424.在结构保守的表面暴露的抗原结合构件cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键。
425.本公开还提供了一种分离的单链可变片段(scfv),其包含重链可变区(vh)、接头(l)和用于抗原结合的构件,其中该scfv包含
426.在结构保守的表面暴露的vh半胱氨酸(cys)与第一l cys之间的第一二硫键;
427.在结构保守的表面暴露的抗原结合构件cys与第二l cys之间的第二二硫键;或者
428.在结构保守的表面暴露的vh cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的抗原结合构件cys与第二l cys之间的第二二硫键。
429.本公开还提供了一种包含单链可变片段(scfv)的多特异性分子,该单链可变片段包含重链可变区(vh)、用于连接的构件(l)和轻链可变区(vl),其中该scfv包含
430.在结构保守的表面暴露的vh半胱氨酸(cys)与第一l cys之间的第一二硫键;
431.在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键;或者
432.在结构保守的表面暴露的vh cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键。
433.本公开还提供了一种包含单链可变片段(scfv)的多特异性分子,该单链可变片段包含用于抗原结合的构件、接头(l)和轻链可变区(vl),其中该scfv包含
434.在结构保守的表面暴露的抗原结合构件半胱氨酸(cys)与第一l cys之间的第一二硫键;
435.在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键;或者
436.在结构保守的表面暴露的抗原结合构件cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键。
437.本公开还提供了一种包含单链可变片段(scfv)的多特异性分子,该单链可变片段包含重链可变区(vh)、接头(l)和用于抗原结合的构件(vl),其中该scfv包含
438.在结构保守的表面暴露的vh半胱氨酸(cys)与第一l cys之间的第一二硫键;
439.在结构保守的表面暴露的抗原结合构件cys与第二l cys之间的第二二硫键;或者
440.在结构保守的表面暴露的vh cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的抗原结合构件cys与第二l cys之间的第二二硫键。
441.本公开还提供了一种包含单链可变片段(scfv)的异源分子,该单链可变片段包含重链可变区(vh)、用于连接的构件(l)和轻链可变区(vl),其中该scfv包含
442.在结构保守的表面暴露的vh半胱氨酸(cys)与第一l cys之间的第一二硫键;
443.在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键;或者
444.在结构保守的表面暴露的vh cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键。
445.本公开还提供了一种包含单链可变片段(scfv)的异源分子,该单链可变片段包含用于抗原结合的构件、接头(l)和轻链可变区(vl),其中该scfv包含
446.在结构保守的表面暴露的抗原结合构件半胱氨酸(cys)与第一l cys之间的第一二硫键;
447.在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键;或者
448.在结构保守的表面暴露的抗原结合构件cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键。
449.本公开还提供了一种包含单链可变片段(scfv)的异源分子,该单链可变片段包含重链可变区(vh)、接头(l)和用于抗原结合的构件,其中该scfv包含
450.在结构保守的表面暴露的vh半胱氨酸(cys)与第一l cys之间的第一二硫键;
451.在结构保守的表面暴露的抗原结合构件cys与第二l cys之间的第二二硫键;或者
452.在结构保守的表面暴露的vh cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的抗原结合构件cys与第二l cys之间的第二二硫键。
453.本公开还提供了用于编码本文所提供的scfv的构件。
454.本公开还提供了用于复制本文所提供的载体的构件。
455.本公开还提供了一种组合物,其包含用于稳定scfv的构件。
456.本公开还提供了一种组合物,其包含用于增加scfv的热稳定性的构件。
457.本公开还提供了一种多特异性分子,其包含用于稳定scfv的构件。
458.本公开还提供了一种多特异性分子,其包含用于增加scfv的热稳定性的构件。
459.本公开还提供了一种异源分子,其包含用于稳定scfv的构件。
460.本公开还提供了一种异源分子,其包含用于增加scfv的热稳定性的构件。
4.附图说明
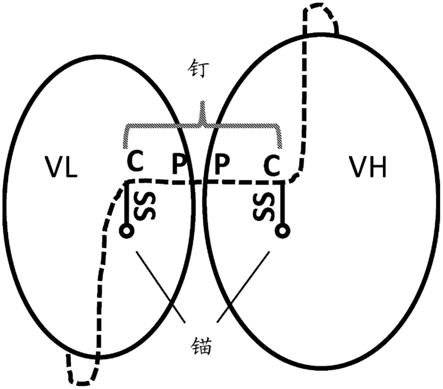
461.图1示出了稳定化的scfv(spfv)的示例性设计。vl和vh通过柔性接头连接,该柔性接头在图中以虚线示出,其含有钉序列cppc(seq id no:1),“ss”指示该接头中的钉序列与锚定点之间的二硫键。
462.图2示出了处于vl-接头-vh取向的spfv的锚定点选择的图解说明。种系人抗体的fv(pdb id 5i19,glk1)用于图形和说明性距离测量。以虚线示出的距离是残基的cβ原子之间的距离(以为单位)。选择具有所需距离的结构保守的框架位置作为突变成cys的锚定点。vl-接头-vh取向的锚定点是vl的chothia位置42(图中的k42)和vh的位置105(图中的q105)。还示出了c末端vl残基(k107)和n末端vh残基(q1)。
463.图3示出了处于vh-接头-vl取向的spfv的锚定点选择的图解说明。种系人抗体的fv(pdb id 5i19,glk1)用于图形和说明性距离测量。以虚线示出的距离是残基的cβ原子之间的距离(以为单位)。选择具有所需距离的结构保守的框架位置作为突变成cys的锚定点。vh-接头-vl取向的锚定点是vh的chothia位置43(图中的k43)和vl的位置100(图中的q100)。还示出了c末端vh残基(s114)和n末端vl残基(d1)。
464.图4示出了小鼠igg2a(pdb id 1igt)铰链cppc(seq id no:1)的两条重链中的两个cys残基之间的cβ(cys1)-cβ(cys2)距离的图解说明。距离在图中以埃为单位示出。
465.图5示出了人igg(pdb id 5dk3)铰链cppc(seq id no:1)的两条重链中的两个cys残基之间的cβ(cys1)-cβ(cys2)距离的图解说明。距离在图中以埃为单位示出。
466.图6示出了所选择的vl锚定点,其以灰色高亮显示并且在氨基酸比对下方编号为1
和2。vl序列根据chothia编号方案进行编号。vl锚定点1(chothia位置42)在处于vl-接头-vh取向的spfv中使用,并且vl锚定点2(chothia位置100)在处于vh-接头-vl取向的spfv中使用。glk1vl:seq id no:56,glk2vl:seq id no:57,cat2200vl:seq id no:58;cat2200bvl:seq id no:59。
467.图7示出了所选择的vh锚定点,其以灰色高亮显示并且在氨基酸比对下方编号为1和2。vh序列根据chothia编号方案进行编号。vh锚定点1(chothia位置105)在处于vl-接头-vh取向的spfv中使用,并且vh锚定点2(chothia位置43)在处于vh-接头-vl取向的spfv中使用。glk1vh:seq id no:60;glk2vh:seq id no:61,cat2200avh:seq id no:62。
468.图8示出了glk1 spfv vl-vh的结构。从该结构中明显看出在vh锚定点与接头之间以及vl锚定点与接头之间形成了钉。
469.图9示出了glk1 spfv vh-vl的结构。从该结构中明显看出在vh锚定点与接头之间以及vl锚定点与接头之间形成了钉。
470.图10示出了glk2 spfv vh-vl的结构。从该结构中明显看出在vh锚定点与接头之间以及vl锚定点与接头之间形成了钉。
471.图11示出了cat2200b spfv vh-vl的结构。从该结构中明显看出在vh锚定点与接头之间以及vl锚定点与接头之间形成了钉。
472.图12示出了未结合的cat2200b spfv vh-vl(上图)与结合至il-17a的cat2200a scfv vl-vh(下图)的比较。
473.图13示出了未结合的cat2200b spfv vh-vl(上图)与结合至il-17a的cat2200a spfv vl-vh(下图)的结构的前视图的比较。
474.图14示出了未结合的cat2200b spfv vh-vl(上图)与结合至il-17a的cat2200a spfv vl-vh(下图)的结构的后视图的比较。
475.图15a至图15m示出了与装订scfv融合的抗体的示意图。图15a至图15d示出了2:1异源二聚体,即与衍生自ltbrmab1的装订scfv融合的同种型对照抗体。图15e至图15h示出了2:1异源二聚体,即与衍生自ltbrmab1的装订scfv融合的edbmab1。图15i至图15j示出了2:1异源二聚体,即与衍生自ltbrmab1的较低亲和力变体的装订scfv融合的edbmab1。图15k至图15l示出了2:1异源二聚体,即与衍生自ltbrmab1的装订scfv融合的edbmab1或b21m,其在fc区中不含蛋白质a突变。图15m示出了2:1异源二聚体,即与衍生自ltbrmab1的装订scfv融合的mslnmab1。
476.图16a至图16c示出了证明使用2:1双特异性抗体进行a549 nf-κb报告基因测定的结果的曲线图。图16a:cova1456与cova1482、它们相应的对照分子cova1462和cova1486,以及重组人light引起的taa依赖性ltbr激活的比较;图16b:cova1482和含有ltbrmab1的较低亲和力变体的双特异性抗体cova14107和cova14108,以及cova1486引起的taa依赖性ltbr激活的比较;图16c:cova1482和cova14133(没有蛋白质a突变的构建体),以及它们相应的对照分子cova1486和cova14136引起的taa依赖性ltbr激活的比较。
477.图17示出了共培养实验后a375细胞上的icam-1的流式细胞术染色结果。将cova1482及其对照分子cova1486与重组人light进行比较。
478.图18a至图18d示出了证明与cova14136和cova1440相比,用抗edb/抗ltbr双特异性抗体cova14133处理的共培养物的上清液中的细胞因子的测量结果的曲线图。使用msd平
台进行测定。图18a:人rantes的浓度;图18b:人il-6的浓度;图18c:人il-8的浓度;图18d:人mip-3b的浓度。
479.图19a至图19b示出了在a549 nf-κb报告基因/chok1msln或a549 nf-κb报告基因/h226共培养细胞测定中,由msln/ltbr双特异性引起的ltbr激活。图19a:a549 nf-κb报告基因/h226共培养测定中的ltbr的激活。cova14146(2:1mslnmab1
×
ltbrmab1)与light以及同种型对照2:1构建体cova1486进行比较;图19b:在a549 nf-κb报告基因/h226共培养测定中,在ltbr激活后分泌的rantes的浓度。cova14146(2:1mslnmab1
×
ltbrmab1)与light以及同种型对照2:1构建体cova1486进行比较。
5.具体实施方式
480.通过结合形成本公开的一部分的附图参考以下具体实施方式,可以更容易地理解本发明所公开的方法。应当理解,本发明所公开的方法不限于本文所描述和/或示出的具体方法,并且本文所用的术语仅以举例的方式用于描述具体实施方案的目的,而并非旨在进行限制。
481.本文引用的所有专利、公布的专利申请和出版物均以引用方式并入本文,如同在本文中进行了充分阐述。
482.当提供一个列表时,除非另行指出,否则应当理解,该列表中的每个单独元素和该列表的每种组合都是单独的实施方案。例如,作为“a、b或c”呈现的实施方案的列表将被理解为包括实施方案“a”、“b”、“c”、“a或b”、“a或c”、“b或c”或者“a、b或c”。
483.如本说明书和所附权利要求中所用,除非内容另有明确说明,否则单数形式“一个”、“一种”、“该”和“所述”包括复数指代。因此,例如,对“一个细胞”的提及包括两个或更多个细胞的组合等等。
484.过渡术语“包括”、“基本上由
……
组成”和“由
……
组成”旨在暗示它们在专利用语中的公认含义;即,(i)“包括”与“包含”、“含有”或“其特征在于”同义,并且是包括端值在内或末端开放的,并且不排除附加的、未列出的要素或方法步骤;(ii)“由
……
组成”排除权利要求书未指定的任何要素、步骤或成分;以及(iii)“基本上由
……
组成”将权利要求的范围限制于指定的材料或步骤“以及本质上不影响受权利要求书保护的发明的基本及新颖特征的材料或步骤”。还提供了以短语“包括”(或其等同形式)描述的实施方案,如以“由
……
组成”和“基本上由
……
组成”独立描述的那些实施方案。
[0485]“约”是指处于如本领域的普通技术人员所确定的特定值的可接受误差范围之内,其将部分取决于该值是如何测量或测定的,即,测量系统的限制。在特定测定、结果或实施方案的上下文中,除非实施例或说明书其他地方内另有明确说明,否则“约”意指在根据本领域惯例的一个标准偏差之内、或多至5%的范围(无论哪个更大)。
[0486]“替代支架”是指包含与具有高构象耐受性的可变结构域相关联的结构化核心的单链蛋白质框架。可变结构域耐受待引入的变化而会不损害支架完整性,因此可工程化和选择可变结构域以与特异性抗原结合。
[0487]“抗体依赖性细胞的细胞毒性”、“抗体依赖性细胞介导的细胞毒性”或“adcc”是指诱导细胞死亡的机制,该机制依赖于抗体包被的靶细胞与具有裂解活性的效应细胞(诸如自然杀伤细胞(nk)、单核细胞、巨噬细胞和中性粒细胞)经由效应细胞上表达的fcγ受体
(fcγr)发生的相互作用。
[0488]“抗体依赖性细胞吞噬作用”或“adcp”是指通过吞噬细胞(诸如巨噬细胞或树突状细胞)的内化作用消除抗体包被的靶细胞的机制。
[0489]“抗原”是指能够介导免疫应答的任何分子(例如,蛋白质、肽、多糖、糖蛋白、糖脂、核酸、它们的部分,或它们的组合)。示例性免疫应答包括抗体产生和免疫细胞(诸如t细胞、b细胞或nk细胞)激活。
[0490]“抗原结合片段”或“抗原结合结构域”是指蛋白质的结合抗原的一部分。抗原结合片段可以是合成的、可酶促获得的或遗传工程化的多肽,并且包括:结合抗原的免疫球蛋白部分(诸如vh、vl、vh和vl、fab、fab’、f(ab’)2、fd和fv片段),由一个vh结构域或一个vl结构域、驼峰化vh结构域、vhh结构域组成的结构域抗体(dab),由模拟抗体cdr的氨基酸残基组成的最小识别单位(诸如fr3-cdr3-fr4部分、hcdr1、hcdr2和/或hcdr3和lcdr1、lcdr2和/或lcdr3),结合抗原的替代支架,以及包含抗原结合片段的多特异性蛋白质。抗原结合片段(诸如vh和vl)可以经由合成接头连接在一起以形成各种类型的单抗体设计,其中在vh结构域和vl结构域由单独的单链表达的情况下,vh/vl结构域可以在分子内或分子间配对,以形成单价抗原结合结构域,诸如单链fv(scfv)或双价抗体。抗原结合片段还可以缀合至其他抗体、蛋白质、抗原结合片段或替代支架,其可以是单特异性或多特异性的,以工程化双特异性蛋白质和多特异性蛋白质。
[0491]“抗体”广义上是指并且包括免疫球蛋白分子,具体包括单克隆抗体(包括鼠单克隆抗体、人单克隆抗体、人源化单克隆抗体和嵌合单克隆抗体),抗原结合片段,多特异性抗体诸如双特异性、三特异性、四特异性抗体等,二聚抗体、四聚抗体或多聚抗体,单链抗体,结构域抗体,以及包含具有所需特异性的抗原结合位点的免疫球蛋白分子的任何其他经修饰构型。“全长抗体”包含由二硫键互连的两条重链(hc)与两条轻链(lc),以及它们的多聚体(例如,igm)。每条重链由重链可变区(vh)和重链恒定区(由结构域ch1、铰链、ch2和ch3构成)构成。每条轻链由轻链可变区(vl)和轻链恒定区(cl)构成。vh区和vl区可进一步细分为超变区,该超变区称为互补决定区(cdr)并间插有框架区(fr)。各个vh和vl由三个cdr和四个fr片段构成,并按以下顺序从氨基端至羧基端排列:fr1、cdr1、fr2、cdr2、fr3、cdr3和fr4。免疫球蛋白可根据重链恒定结构域氨基酸序列被指定为五种主要种类,即iga、igd、ige、igg和igm。iga和igg进一步亚分类为同种型iga1、iga2、igg1、igg2、igg3和igg4。基于其恒定域的氨基酸序列,可将任何脊椎物种的抗体轻链指定为两种完全不同的类型即κ和λ中的一种。
[0492]“双特异性”是指特异性地结合两个不同抗原或同一抗原内的两个不同表位的分子(诸如抗体)。双特异性分子可能对其他相关的抗原具有交叉反应性,例如,对来自其他物种(同源物)(诸如人或猴,例如食蟹猕猴(macaca fascicularis)(cynomolgus,cyno)或黑猩猩(pan troglodytes))的相同抗原具有交叉反应性,或者可以结合两个或更多个不同抗原之间所共享的表位。
[0493]“嵌合抗原受体”或“car”是指将配体或抗原特异性移植到t细胞(例如初始t细胞、中枢记忆t细胞、效应记忆t细胞或它们的组合)上的工程化t细胞受体。car也被称为人造t细胞受体、嵌合t细胞受体或嵌合免疫受体。car包含能够结合到抗原的胞外结构域、跨膜结构域和至少一个胞内结构域。car胞内结构域包含已知用作传输信号以引起细胞中生物过
程的活化或抑制的结构域的多肽。跨膜结构域包含已知跨越细胞膜并且可起到连接胞外结构域和信号结构域的作用的任何肽或多肽。嵌合抗原受体可任选地包含铰链结构域,该铰链结构域充当胞外结构域与跨膜结构域之间的接头。
[0494]“补体依赖性细胞毒性”或“cdc”是指诱导细胞死亡的机制,其中靶结合蛋白质的fc效应结构域结合并激活补体成分c1q,c1q继而激活补体级联,从而导致靶细胞死亡。补体的激活还可以导致补体成分沉积在靶细胞表面上,这些补体成分通过结合白细胞上的补体受体(例如,cr3)来促进cdc。
[0495]“互补决定区”(cdr)是结合抗原的抗体区域。vh中存在三个cdr(hcdr1、hcdr2、hcdr3)并且vl中存在三个cdr(lcdr1、lcdr2、lcdr3)。cdr可以使用各种描绘诸如kabat(wu等人,(1970)j exp med 132:211-250;kabat等人,sequences of proteins of immunological interest,第5版,public health service,national institutes of health,bethesda,md.,1991)、chothia(chothia等人,(1987)j mol biol 196:901-17)、imgt(lefranc等人,(2003)dev comp immunol 27:55-77)和abm(martin和thornton(1996)j bmol biol 263:800-815)来限定。描述了各种描绘和可变区编号之间的对应关系(参见例如,lefranc等人,(2003)dev comp immunol 27:55-77;honegger和pluckthun,j mol biol(2001)309:657-670;国际免疫遗传学(imgt)数据库;web资源,http://www_imgt_org)。可用程序(诸如ucl business plc的abysis)可用于描绘cdr。除非说明书中另有明确地说明,否则如本文所用,术语“cdr”、“hcdr1”、“hcdr2”、“hcdr3”、“lcdr1”、“lcdr2”和“lcdr3”包括由任何上述方法(kabat、chothia、imgt或abm)定义的cdr。
[0496]“减少”、“减小”或“降低”通常是指测试分子介导与由对照或媒介物介导的应答相比降低的应答(即,下游效应)的能力。示例性应答包括蛋白质与其抗原或受体结合、与fcγr的结合增强或fc效应功能增强,诸如adcc、cdc和/或adcp增强。减少可以是测试分子与对照(或媒介物)之间的测量应答的统计上显著的差异,或者测量应答的减少,诸如减少到约1/1.1、1/1.2、1/1.5、1/2、1/3、1/4、1/5、1/6、1/7、1/8、1/9、1/10、1/15、1/20或1/30或者更小,诸如1/500、1/600、1/700、1/800、1/900或1/1000或者更小。
[0497]“增强”、“促进”或“增加”通常是指测试分子介导与由对照或媒介物介导的应答相比较大的应答(即,下游效应)的能力。示例性应答是蛋白质与其抗原或受体结合、与fcγr的结合增强或fc效应功能增强,诸如adcc、cdc和/或adcp增强。增强可以是测试分子与对照(或媒介物)之间的测量应答的统计上显著的差异,或者测量应答的增加,诸如增加到约1.1、1.2、1.5、2、3、4、5、6、7、8、9、10、15、20或30倍或者更多,诸如500、600、700、800、900或1000倍或者更多。
[0498]“表达载体”是指可用于在生物系统或再造生物系统中以指导由存在于表达载体中的多核苷酸序列所编码的多肽进行翻译的载体。
[0499]“异源”是指以下多肽或多核苷酸:其包括在本质上彼此没有相同的关系的两种或更多种多肽或者两种或更多种多核苷酸。
[0500]“异源多核苷酸”是指以下多核苷酸:其包括在本质上彼此没有相同的关系的两种或更多种多核苷酸。
[0501]“异源多肽”是指以下多肽:其包括在本质上彼此没有相同的关系的两种或更多种多肽。
[0502]“人抗体”是指当施用于人受试者时被优化以具有最小限度的免疫应答的抗体。人抗体的可变区来源于人免疫球蛋白序列。如果人抗体包含恒定区或恒定区的一部分,则该恒定区也来源于人免疫球蛋白序列。如果人抗体的可变区由使用人种系免疫球蛋白或重排免疫球蛋白基因的系统获得,则该人抗体包含“来源于”人起源序列的重链可变区和轻链可变区。此类示例性系统为在噬菌体上展示的人免疫球蛋白基因文库,以及转基因非人动物,诸如携带人免疫球蛋白基因座的小鼠或大鼠。由于用于获得人抗体和人免疫球蛋白基因座的系统之间的差异,体细胞突变的引入或有意将置换引入框架或cdr中或两者,因此“人抗体”与在人中表达的免疫球蛋白相比通常包含氨基酸差异。通常,“人抗体”的氨基酸序列与由人种系免疫球蛋白基因或重排免疫球蛋白基因编码的氨基酸序列具有至少约80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的同一性。在一些情况下,“人抗体”可以包含由人框架序列分析得出的共有框架序列(例如,如knappik等人,(2000)j mol biol 296:57-86中所述),或结合到展示在噬菌体上的人免疫球蛋白基因文库中的合成hcdr3(例如,如shi等人,(2010)j mol biol 397:385-396和国际专利公开号wo2009/085462中所述)。“人抗体”的定义中不包括至少一个cdr来源于非人物种的抗体。
[0503]“人源化抗体”是指其中至少一个cdr来源于非人物种并且至少一个框架来源于人免疫球蛋白序列的抗体。人源化抗体在框架中可包含置换,使得该框架可能不是表达的人免疫球蛋白或人免疫球蛋白种系基因序列的精确拷贝。
[0504]“分离的”是指已经与分子产生于其中的系统(例如重组细胞)中的其他组分基本上分离和/或从其中纯化出来的分子(诸如本公开的scfv或者含有本公开的scfv的异源蛋白)的同质群体,以及已经受至少一个纯化或分离步骤的蛋白质。“分离的”是指基本上不含其他细胞材料和/或化学物质的分子,并且涵盖分离至更高纯度(诸如80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%纯度)的分子。
[0505]“调节”是指测试分子介导与由对照或媒介物介导的应答相比增强或降低的应答(即,下游效应)的增强或降低的能力。
[0506]“单克隆抗体”是指从抗体分子的基本上同质的群体(即,除了可能熟知的改变(诸如从抗体重链移除c末端赖氨酸)或翻译后修饰(诸如氨基酸异构化或脱酰胺、甲硫氨酸氧化或者天冬酰胺或谷氨酰胺脱酰胺)之外,构成群体的各个抗体是相同的)获得的抗体。单克隆抗体通常结合一个抗原表位。双特异性单克隆抗体结合两个不同的抗原表位。单克隆抗体可在抗体群内具有异质糖基化。单克隆抗体可以是单特异性的或多特异性的,诸如双特异性的、单价的、二价的或多价的。
[0507]“多特异性”是指结合两个或更多个不同抗原或者同一抗原内的两个或更多个不同表位的分子。多特异性分子可能对其他相关的抗原具有交叉反应性,例如,对来自其他物种(同源)(诸如人或猴,例如食蟹猕猴或黑猩猩)的相同抗原具有交叉反应性,或者可以结合在两个或更多个不同抗原之间所共用的表位。
[0508]“多核苷酸”是指包含磷酸糖类主链共价连接的核苷酸链或其他等同共价化学物的分子。cdna是多核苷酸的典型示例。
[0509]“蛋白质”或“多肽”在本文中可互换使用,是指包含一种或多种多肽的分子,其中
每种多肽包含至少两个通过肽键连接的氨基酸残基。蛋白质可以是单体,或者可以是两个或更多个亚基的蛋白质复合物,所述亚基是相同或不同的。少于50个氨基酸的小多肽可以称作“肽”。蛋白质可以是异源融合蛋白、糖蛋白或通过翻译后修饰(诸如磷酸化、乙酰化、豆蔻酰化、棕榈酰化、糖基化、氧化、甲酰化、酰胺化、瓜氨酸化、聚谷氨酰化、adp核糖基化、聚乙二醇化或生物素化)进行修饰的蛋白质。
[0510]“重组体”是指通过重组手段制备、表达、创建或分离的多核苷酸、多肽、载体、病毒和其他大分子。
[0511]“单链fv”或“scfv”是指包含vh、vl以及vh与vl之间的接头的单链蛋白。该scfv可以具有处于任一取向的vl可变区和vh可变区,例如,相对于vh和vl的n末端至c末端顺序。因此,scfv可以处于vl-接头-vh或vh-接头-vl取向。scfv可以被工程化为在vh、vl与接头之间包含二硫键。
[0512]“特异性地结合”、“特异性结合”、或“结合”是指蛋白质(诸如scfv)以比针对其他抗原更高的亲和力结合至某个抗原或该抗原内的表位。通常,蛋白质(诸如scfv)以约1
×
10-6
m或更小、约1
×
10-7
m或更小、约5
×
10-8
m或更小、约1
×
10-8
m或更小、约1
×
10-9
m或更小、约1
×
10-10
m或更小、约1
×
10-11
m或更小、或约1
×
10-12
m或更小的平衡解离常数(kd),通常以其与非特异性抗原(例如bsa、酪蛋白)结合的kd的最多1/100的kd,与抗原或该抗原内的表位结合。
[0513]“装订单链fv”或“spfv”是指在vh与接头之间或者vl与接头之间包含一个或多个二硫键的scfv。通常,该spfv可以包含vh与接头之间的一个二硫键、vl与接头之间的一个二硫键,或者vh与接头之间以及vl与接头之间的两个二硫键。术语“spfv”不包括在vh与vl之间包含二硫键的scfv分子。
[0514]“受试者”包括任何人或非人动物。“非人动物”包括所有脊椎动物,例如,哺乳动物和非哺乳动物,诸如非人灵长类动物、绵羊、狗、猫、马、牛、鸡、两栖动物、爬行动物等。术语“受试者”和“患者”在本文中可以互换使用。
[0515]“治疗有效量”是指在必需的剂量下和必需的时间段内有效地实现期望的治疗结果的量。治疗有效量可根据以下因素变化:诸如个体的疾病状态、年龄、性别和重量,以及治疗剂或治疗剂组合在个体中引发期望的应答的能力。
[0516]
疾病或疾患的“治疗”是指实现以下各项中的一者或多者:降低疾患的严重程度和/或减少其持续时间,抑制所治疗疾患的特征性症状的恶化,限制或防止先前患有疾患的受试者中疾患的复发,或者限制或防止先前有疾患的症状的受试者中症状的复发。
[0517]“三特异性”是指特异性地结合三个不同抗原或同一抗原内的三个不同表位的分子(诸如抗体)。三特异性分子可能对其他相关的抗原具有交叉反应性,例如,对来自其他物种(同源物)(诸如人或猴,例如食蟹猕猴或黑猩猩)的相同抗原具有交叉反应性,或者可以结合三个或更多个不同抗原之间所共用的表位。
[0518]“变体”、“突变体”或“改变的”是指因一种或多种修饰(例如一个或多个置换、插入或缺失)而不同于参考多肽或参考多核苷酸的多肽或多核苷酸。
[0519]
除非另外明确指出,否则在整个说明书中,抗体恒定区中的氨基酸残基都根据如kabat等人,sequences of proteins of immunological interest,第5版,public health service,national institutes of health,bethesda,md.(1991)中所述的eu索引进行编
号。
[0520]
ig恒定区中的突变如下指出:l351y_f405a_y407v是指一个免疫球蛋白恒定区中的l351y、f405a和y407v突变。l351y_f405a_y407v/t394w是指该分子中存在的第一ig恒定区中的l351y、f405a和y407v突变,以及第二ig恒定区中的t394w突变。
[0521]
除非另外明确指出,否则可变区根据chothia进行编号。
[0522]“vh半胱氨酸”或“vh cys”是指位于vh框架中的cys残基。
[0523]“vl半胱氨酸”或“vl cys”是指位于vl框架中的cys残基。
[0524]“稳定化的”是指scfv与未加热的scfv样品相比保持相当的与hk2的结合,其被称为是热稳定化的。
[0525]“改进的稳定性”是指本公开的spfv与没有二硫键和cys残基被引入spfv中的亲本scfv相比具有升高的解链温度(tm)。升高的tm可以是升高2℃或更多,诸如3℃、4℃、5℃、6℃、7℃、8℃、9℃、10℃、11℃、12℃、13℃、14℃或15℃。
[0526]“锚定点”是指以下scfv vh或vl框架cys残基:其可以被诱变为cys而对总体scfv结构没有不利影响,并且能够与位于scfv接头中的cys形成二硫键。
[0527]“钉”是指含有一个或两个cys残基的scfv接头,所述cys残基能够与锚定点cys形成二硫键。
[0528]“表面暴露的”是指至少部分地暴露于蛋白质表面并且可接近溶剂(诸如可接近氘化)的氨基酸残基。基于一级序列或蛋白质预测残基的表面可接近性的算法是本领域熟知的。替代性地,表面暴露的残基可以从蛋白质的晶体结构中鉴定。
[0529]“ltbr”是指作为参与细胞凋亡和细胞因子释放的淋巴毒素的细胞表面受体的多肽,其是肿瘤坏死因子受体超家族的成员。ltbr也可以称为“肿瘤坏死因子受体超家族成员3(tnfrsf3)”。ltbr在许多细胞类型(包括上皮谱系和骨髓谱系的细胞)的表面上表达。ltbr可以特异性地结合淋巴毒素膜形式(淋巴毒素-α和淋巴毒素-β的复合物)。ltbr的激活可以经由traf3和traf5触发细胞凋亡,并且可以导致白介素8的释放。除非另外指明,否则优选地,ltbr为人ltbr。人ltbr氨基酸序列由uniprot编号p36941提供。
[0530]“edb”或“额外结构域b”是指基于纤连蛋白前mrna的剪接模式可以被包括在纤连蛋白分子中的纤连蛋白结构域。额外结构域b是包含91个氨基酸残基的完整纤连蛋白(fn)iii型重复序列。一般来讲,edb在正常成人组织中是检测不到的,但在胎儿组织和肿瘤组织中在细胞外基质中却表现出更高的表达,并且在血管生成过程期间在新血管系统周围累积,从而使得edb成为血管生成的潜在标志物和靶标。除非另外指明,否则优选地,edb为人edb。含有纤连蛋白同种型氨基酸序列的人edb由uniprot编号p02751提供。
[0531]“纤连蛋白”是指为细胞外基质的高分子量糖蛋白的多肽。纤连蛋白可以结合到跨膜受体蛋白(称为整合素)。纤连蛋白还可以结合其他细胞外基质蛋白,诸如胶原蛋白、纤维蛋白和硫酸乙酰肝素蛋白聚糖。纤连蛋白可以作为蛋白质二聚体存在,由通过一对二硫键连接的两个几乎相同的单体组成。纤连蛋白由单个基因产生,但是纤连蛋白前mrna分子的替代性剪接导致形成纤连蛋白的几种同种型,其中一种同种型是edb纤连蛋白。纤连蛋白可以在细胞粘附、生长、迁移和分化中起作用,并且对于诸如创伤愈合和胚胎发育等过程可能很重要。人纤连蛋白氨基酸序列由以下各项提供:uniprot编号p02751(其含有额外结构域b),以及ncbi登录号np_001263337(同种型b)、np_001263338(同种型c)、np_001263339(同
种型d)、np_001263340(同种型e)和np_001263341(同种型f),np_001293058(同种型8)、np_001293059(同种型9)、np_001293060(同种型10)、np_001293061(同种型11),以及np_002017(同种型3)。
[0532]
5.1物质组合物
[0533]
本公开提供稳定化的scfv分子(本文称为spfv(装订fv))、包含所述spfv的异源分子和多特异性分子、编码它们的多核苷酸、载体、宿主细胞,以及制备和使用它们的方法。本公开至少部分地基于对vh和/或vl(本文称为vh锚定点或vl锚定点)中以及柔性接头(本文称为钉)中的合适残基位置的鉴定,所述柔性接头可以被工程化为半胱氨酸残基,从而导致在该接头与scfv中的可变结构域之间形成二硫键。本文所述的“装订”策略广泛适用于所有vh/vl结构域和预先存在的scfv分子,从而提供与scfv的结构同一性,同时具有改善的稳定性。本文所述的spfv可以缀合到双特异性或多特异性形式的任何异源蛋白质中,包括嵌合抗原受体(car)、t细胞重定向分子、双多特异性分子和多特异性分子,并且可以用作治疗分子、诊断分子和检测分子。
[0534]
本公开的spfv
[0535]
本公开提供了一种分离的单链可变片段(scfv),其包含重链可变区(vh)、接头(l)和轻链可变区(vl),其中该scfv包含
[0536]
在结构保守的表面暴露的vh半胱氨酸(cys)与第一l cys之间的第一二硫键;
[0537]
在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键;或者
[0538]
在结构保守的表面暴露的vh cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键。
[0539]
本公开还提供了一种分离的scfv,其包含vh、l和vl,其中
[0540]
该vh包含在结构保守的表面暴露的vh框架残基位置处的vh cys,并且该l包含第一l cys;
[0541]
该vl包含在结构保守的表面暴露的vl框架残基位置处的vl cys,并且该l包含第二l cys;或者
[0542]
该vh包含在结构保守的表面暴露的vh框架残基位置处的vh cys,该vl包含在结构保守的表面暴露的vl框架残基位置处的vl cys,并且该l包含第一l cys和第二l cys,其中vh cys和第一l cys能够形成二硫键并且vl cys和第二l cys能够形成二硫键。二硫键通常在本公开的scfv的表达期间形成。
[0543]
尽管具体示例公开了具有两个二硫键的spfv,但是容易设想的是,可以制备并且利用具有在接头cys与vh cys或vl cys之间形成的一个二硫键的spfv,从而产生“半锚定”分子。在具有一个或两个二硫键的spfv中,锚定位置是相同的。半锚定分子中的接头cys位置可以变化,只要其满足与锚定点形成二硫键的距离和几何结构上的要求即可。可以预期,半锚定的spfv将抑制vl/vh的相对移动,类似于用两个二硫键稳定的vl/vh对,因此也将是稳定的。
[0544]
当与缺少二硫键的亲本scfv相比时,本公开的spfv表现出增加的热稳定性。一般来讲,当与缺少二硫键的亲本scfv相比时,无论该亲本scfv的tm是多少,spfv的tm均高约10℃。一般来讲,稳定性可以是热稳定性或机械稳定性。热稳定性可以使用差示量热法(dsc)评估,其中dsc扫描使用加热的蛋白质样品(诸如加热至60℃的样品)进行,之后使用两状态
转变或非两状态转变分析所得的热解链曲线。对于非两状态转变,记录两个转变(tm1和tm2),其分别对应于vl结构域和vh结构域的解链tm。
[0545]
在一些实施方案中,vh cys与vl cys之间的距离为约至约在一些实施方案中,vh cys与vl cys之间的距离为约在一些实施方案中,vh cys与vl cys之间的距离为约在一些实施方案中,vh cys与vl cys之间的距离为约
[0546]
在一些实施方案中,vh cys在h3、h5、h40、h43、h46或h105处,其中残基根据chothia进行编号。
[0547]
在一些实施方案中,vh cys在h3处。
[0548]
在一些实施方案中,vh cys在h5处。
[0549]
在一些实施方案中,vh cys在h40处。
[0550]
在一些实施方案中,vh cys在h43处。
[0551]
在一些实施方案中,vh cys在h46处。
[0552]
在一些实施方案中,vh cys在h105处。
[0553]
在一些实施方案中,vl cys在l3、l5、l39、l42、l45、l100或l102处,其中残基根据chothia进行编号。
[0554]
在一些实施方案中,vl cys在l3处。
[0555]
在一些实施方案中,vl cys在l5处。
[0556]
在一些实施方案中,vl cys在l39处。
[0557]
在一些实施方案中,vl cys在l42处。
[0558]
在一些实施方案中,vl cys在l45处。
[0559]
在一些实施方案中,vl cys在l100处。
[0560]
在一些实施方案中,vl cys在l102处。
[0561]
在一些实施方案中,vh cys在h105处并且vl cys在l42处。
[0562]
在一些实施方案中,vh cys在h43处并且vl cys在l100处。
[0563]
在一些实施方案中,vh cys在h3处并且vl cys在l3处。
[0564]
在一些实施方案中,vh cys在h3处并且vl cys在l5处。
[0565]
在一些实施方案中,vh cys在h3处并且vl cys在l39处。
[0566]
在一些实施方案中,vh cys在h3处并且vl cys在l42处。
[0567]
在一些实施方案中,vh cys在h3处并且vl cys在l45处。
[0568]
在一些实施方案中,vh cys在h3处并且vl cys在l100处。
[0569]
在一些实施方案中,vh cys在h3处并且vl cys在l102处。
[0570]
在一些实施方案中,vh cys在h5处并且vl cys在l3处。
[0571]
在一些实施方案中,vh cys在h5处并且vl cys在l5处。
[0572]
在一些实施方案中,vh cys在h5处并且vl cys在l39处。
[0573]
在一些实施方案中,vh cys在h5处并且vl cys在l42处。
[0574]
在一些实施方案中,vh cys在h5处并且vl cys在l45处。
[0575]
在一些实施方案中,vh cys在h5处并且vl cys在l100处。
[0576]
在一些实施方案中,vh cys在h5处并且vl cys在l102处。
[0577]
在一些实施方案中,vh cys在h40处并且vl cys在l3处。
[0578]
在一些实施方案中,vh cys在h40处并且vl cys在l5处。
[0579]
在一些实施方案中,vh cys在h40处并且vl cys在l39处。
[0580]
在一些实施方案中,vh cys在h40处并且vl cys在l42处。
[0581]
在一些实施方案中,vh cys在h40处并且vl cys在l45处。
[0582]
在一些实施方案中,vh cys在h40处并且vl cys在l100处。
[0583]
在一些实施方案中,vh cys在h40处并且vl cys在l102处。
[0584]
在一些实施方案中,vh cys在h43处并且vl cys在l3处。
[0585]
在一些实施方案中,vh cys在h43处并且vl cys在l5处。
[0586]
在一些实施方案中,vh cys在h43处并且vl cys在l39处。
[0587]
在一些实施方案中,vh cys在h43处并且vl cys在l42处。
[0588]
在一些实施方案中,vh cys在h43处并且vl cys在l45处。
[0589]
在一些实施方案中,vh cys在h43处并且vl cys在l102处。
[0590]
在一些实施方案中,vh cys在h46处并且vl cys在l3处。
[0591]
在一些实施方案中,vh cys在h46处并且vl cys在l5处。
[0592]
在一些实施方案中,vh cys在h46处并且vl cys在l39处。
[0593]
在一些实施方案中,vh cys在h46处并且vl cys在l42处。
[0594]
在一些实施方案中,vh cys在h46处并且vl cys在l45处。
[0595]
在一些实施方案中,vh cys在h46处并且vl cys在l100处。
[0596]
在一些实施方案中,vh cys在h46处并且vl cys在l102处。
[0597]
在一些实施方案中,vh cys在h105处并且vl cys在l3处。
[0598]
在一些实施方案中,vh cys在h105处并且vl cys在l5处。
[0599]
在一些实施方案中,vh cys在h105处并且vl cys在l39处。
[0600]
在一些实施方案中,vh cys在h105处并且vl cys在l45处。
[0601]
在一些实施方案中,vh cys在h105处并且vl cys在l100处。
[0602]
在一些实施方案中,vh cys在h105处并且vl cys在l102处。
[0603]
vh区和vl区的残基根据chothia进行编号。
[0604]
chothia编号是众所周知的。其他编号系统(诸如kabat或imgt编号)或者顺序编号可以用于对vh残基位置和vl残基位置进行编号。表1示出了示例性vh、glk1 vh(seq id no:60)的chothia编号、kabat编号和顺序编号之间的对应关系。表2示出了示例性vl、glk1 vl(seq id no:56)的chothia编号、kabat编号和顺序编号之间的对应关系。
[0605]
表1.
[0606]
[0607]
[0608][0609]
表2.
[0610]
[0611][0612]
在一些实施方案中,该l包含衍生自免疫球蛋白(ig)铰链区的连续氨基酸序列。
[0613]
在一些实施方案中,ig铰链区衍生自人ig铰链区或非人ig铰链区。示例性非人ig铰链区是来自小鼠、大鼠、狗、鸡和非人灵长类动物(诸如猴)的那些。
[0614]
在一些实施方案中,ig铰链区衍生自人ig铰链区。
[0615]
在一些实施方案中,人ig铰链区是igg1、igg2、igg3、igg4、igm、iga或ige同种型。
[0616]
一般来讲,ig铰链区被定义为包含人igg的残基216并且终止于人igg的残基230处,其中残基根据eu索引进行编号。在一些情况下,还可以将从约残基231至约残基237的下铰链区包含到该铰链中。因此,igg1铰链区可以包含氨基酸序列epkscdkthtcppcp(seq id no:63),或者在包含下铰链时,包含氨基酸序列epkscdkthtcppcpapellgg(seq id no:64)。其他ig同种型的铰链区是众所周知的,并且其氨基酸序列可以例如在immunogenetics网站处获得。例如,igg2铰链包含氨基酸序列erkccvecppcp(seq id no:65)。
[0617]
在l包含ig铰链区氨基酸序列的至少一部分或工程化ig铰链区的至少一部分的情况下,该l包含“衍生自”ig铰链区的连续氨基酸序列。当与野生型ig铰链相比时,工程化ig铰链区包含一个或多个突变。可以引入的示例性突变是cys残基的置换(以将l中cys的数目减少至一个或两个)、pro残基的置换,或者任何保守修饰,诸如保守置换。
[0618]“保守修饰”是指不会显著影响或改变含有氨基酸修饰的抗体的结合特征的氨基酸修饰。保守修饰包括氨基酸置换、添加和缺失。“保守氨基酸置换”是其中氨基酸被具有相似侧链的氨基酸残基替换的置换。具有相似侧链的氨基酸残基家族是有严格规定的,包括具有以下侧链的氨基酸:酸性侧链(例如,天冬氨酸、谷氨酸)、碱性侧链(例如,赖氨酸、精氨酸、组氨酸)、非极性侧链(例如,丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、甲硫氨酸)、不带电极性侧链(例如,甘氨酸、天冬酰胺、谷氨酰胺、半胱氨酸、丝氨酸、苏氨酸、酪氨酸、色氨酸)、芳族侧链(例如,苯丙氨酸、色氨酸、组氨酸、酪氨酸)、脂族侧链(例如,甘氨酸、丙氨酸、缬氨酸、亮氨酸、异亮氨酸、丝氨酸、苏氨酸)、酰胺(例如,天冬酰胺、谷氨酰胺)、β-分支侧链(例如,苏氨酸、缬氨酸、异亮氨酸)和含硫侧链(半胱氨酸、甲硫氨酸)。此外,多肽中的任何天然残基还可以用丙氨酸来置换,如先前针对丙氨酸扫描诱变所述(maclennan等人,(1988)acta physiol scand suppl 643:55-67;sasaki等人,(1988)adv biophys 35:1-24)。氨基酸置换可以通过已知的方法进行,例如通过pcr诱变(美国专利号4,683,195)进行。所得的变体铰链可以结合到本公开的spfv构建体中,并且使用已知的测定法和本文所述的测定法测试其特性,诸如稳定性以及与抗原的结合。
[0619]
在一些实施方案中,该l包含氨基酸序列c(x)yc(seq id no:23),其中x为甘氨酸(gly)、丝氨酸(ser)、脯氨酸(pro)、丙氨酸(ala)、精氨酸(arg)、天冬酰胺(asn)、天冬氨酸(asp)、谷氨酸(glu)、谷氨酰胺(gln)、组氨酸(his)、异亮氨酸(ile)、亮氨酸(leu)、赖氨酸(lys)、苯丙氨酸(phe)、苏氨酸(thr)、色氨酸(trp)或酪氨酸(tyr),并且y是1至3的整数。可以将pro包含到该接头中以提供刚性。可以将gly包含到该接头中以允许最大的柔性。还可以在该l中使用除cys和met之外的任何其他氨基酸。
[0620]
在一些实施方案中,该l包含氨基酸序列c(x)yc(seq id no:24),其中x是gly、ser或pro,并且y是1至3的整数。
[0621]
在一些实施方案中,该l包含氨基酸序列cpc、cgc、csc、cppc(seq id no:1)、cgpc(seq id no:28)、cpgc(seq id no:29)、cggc(seq id no:30)、cspg(seq id no:31)、cpsc(seq id no:32)、cssc(seq id no:33)、cgsc(seq id no:34)、csgc(seq id no:35)、cpppc(seq id no:36)、cgppc(seq id no:37)、cpgpc(seq id no:38)、cppgc(seq id no:39)、cggpc(seq id no:40)、cpggc(seq id no:41)、cgggc(seq id no:42)、csppc(seq id no:43)、cpspc(seq id no:44)、cppsc(seq id no:45)、csspc(seq id no:46)、cpssc(seq id no:47)、csssc(seq id no:48)、cgspc(seq id no:49)、cpgsc(seq id no:50)、csgpc(seq id no:51)或cpsgc(seq id no:52)。
[0622]
在一些实施方案中,该l包含氨基酸序列cpc。
[0623]
在一些实施方案中,该l包含氨基酸序列cgc。
[0624]
在一些实施方案中,该l包含氨基酸序列csc。
[0625]
在一些实施方案中,该l包含氨基酸序列cppc(seq id no:1)。
[0626]
在一些实施方案中,该l包含氨基酸序列cgpc(seq id no:28)。
[0627]
在一些实施方案中,该l包含氨基酸序列cpgc(seq id no:29)。
[0628]
在一些实施方案中,该l包含氨基酸序列cggc(seq id no:30)。
[0629]
在一些实施方案中,该l包含氨基酸序列cspg(seq id no:31)。
[0630]
在一些实施方案中,该l包含氨基酸序列cpsc(seq id no:32)。
[0631]
在一些实施方案中,该l包含氨基酸序列cssc(seq id no:33)。
[0632]
在一些实施方案中,该l包含氨基酸序列cgsc(seq id no:34)。
[0633]
在一些实施方案中,该l包含氨基酸序列csgc(seq id no:35)。
[0634]
在一些实施方案中,该l包含氨基酸序列cpppc(seq id no:36)。
[0635]
在一些实施方案中,该l包含氨基酸序列cgppc(seq id no:37)。
[0636]
在一些实施方案中,该l包含氨基酸序列cpgpc(seq id no:38)。
[0637]
在一些实施方案中,该l包含氨基酸序列cppgc(seq id no:39)。
[0638]
在一些实施方案中,该l包含氨基酸序列cggpc(seq id no:40)。
[0639]
在一些实施方案中,该l包含氨基酸序列cpggc(seq id no:41)。
[0640]
在一些实施方案中,该l包含氨基酸序列cgggc(seq id no:42)。
[0641]
在一些实施方案中,该l包含氨基酸序列csppc(seq id no:43)。
[0642]
在一些实施方案中,该l包含氨基酸序列cpspc(seq id no:44)。
[0643]
在一些实施方案中,该l包含氨基酸序列cppsc(seq id no:45)。
[0644]
在一些实施方案中,该l包含氨基酸序列csspc(seq id no:46)。
[0645]
在一些实施方案中,该l包含氨基酸序列cpssc(seq id no:47)。
[0646]
在一些实施方案中,该l包含氨基酸序列csssc(seq id no:48)。
[0647]
在一些实施方案中,该l包含氨基酸序列cgspc(seq id no:49)。
[0648]
在一些实施方案中,该l包含氨基酸序列cpgsc(seq id no:50)。
[0649]
在一些实施方案中,该l包含氨基酸序列csgpc(seq id no:51)。
[0650]
在一些实施方案中,该l包含氨基酸序列cpsgc(seq id no:52)。
[0651]
在一些实施方案中,该l包含约14个至约19个氨基酸。
[0652]
在一些实施方案中,该l包含约14个氨基酸。
[0653]
在一些实施方案中,该l包含约15个氨基酸。
[0654]
在一些实施方案中,该l包含约16个氨基酸。
[0655]
在一些实施方案中,该l包含约17个氨基酸。
[0656]
在一些实施方案中,该l包含约18个氨基酸。
[0657]
在一些实施方案中,该l包含约19个氨基酸。
[0658]
在一些实施方案中,该l包含氨基酸序列(x)mc(x)yc(x)n(seq id no:25),其中x是gly、ser、pro、ala、arg、asn、asp、glu、gln、his、ile、leu、lys、phe、thr、trp或tyr,m是6至9的整数,y是1至3的整数,并且n是4至6的整数。
[0659]
在一些实施方案中,该l包含氨基酸序列(x)mc(x)yc(x)n(seq id no:26),其中x是gly、ser、pro、ala、arg、asn、asp、glu、gln、his、ile、leu、lys、thr或tyr,m是6至9的整数,y是1至3的整数,并且n是4至6的整数。
[0660]
在一些实施方案中,该l包含氨基酸序列(x)mc(x)yc(x)n(seq id no:27);其中x是gly或pro,m是6至9的整数,y是1至3的整数,并且n是4至6的整数。
[0661]
在一些实施方案中,该l包含氨基酸序列seq id no:2、3、4、5、6或7。
[0662]
在一些实施方案中,该l包含氨基酸序列seq id no:2。
[0663]
在一些实施方案中,该l包含氨基酸序列seq id no:3。
[0664]
在一些实施方案中,该l包含氨基酸序列seq id no:4。
[0665]
在一些实施方案中,该l包含氨基酸序列seq id no:5。
[0666]
在一些实施方案中,该l包含氨基酸序列seq id no:6。
[0667]
在一些实施方案中,该l包含氨基酸序列seq id no:7。
[0668]
在一些实施方案中,本公开的spfv处于vl-l-vh取向。
[0669]
在一些实施方案中,本公开的spfv处于vh-l-vl取向。
[0670]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0671]
该vh包含在h105处的cys;
[0672]
该vl包含在l42处的cys;
[0673]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0674]
该scfv处于vl-l-vh取向。
[0675]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0676]
该vh包含在h105处的cys;
[0677]
该vl包含在l45处的cys;
[0678]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0679]
该scfv处于vl-l-vh取向。
[0680]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0681]
该vh包含在h105处的cys;
[0682]
该vl包含在l39处的cys;
[0683]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0684]
该scfv处于vl-l-vh取向。
[0685]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0686]
该vh包含在h5处的cys;
[0687]
该vl包含在l42处的cys;
[0688]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0689]
该scfv处于vl-l-vh取向。
[0690]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0691]
该vh包含在h5处的cys;
[0692]
该vl包含在l45处的cys;
[0693]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0694]
该scfv处于vl-l-vh取向。
[0695]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0696]
该vh包含在h5处的cys;
[0697]
该vl包含在l39处的cys;
[0698]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0699]
该scfv处于vl-l-vh取向。
[0700]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0701]
该vh包含在h3处的cys;
[0702]
该vl包含在l42处的cys;
[0703]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0704]
该scfv处于vl-l-vh取向。
[0705]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0706]
该vh包含在h3处的cys;
[0707]
该vl包含在l45处的cys;
[0708]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0709]
该scfv处于vl-l-vh取向。
[0710]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0711]
该vh包含在h3处的cys;
[0712]
该vl包含在l39处的cys;
[0713]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0714]
该scfv处于vl-l-vh取向。
[0715]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0716]
该vh包含在h43处的cys;
[0717]
该vl包含在l100处的cys;
[0718]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0719]
该scfv处于vh-l-vl取向。
[0720]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0721]
该vh包含在h43处的cys;
[0722]
该vl包含在l102处的cys;
[0723]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0724]
该scfv处于vh-l-vl取向。
[0725]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0726]
该vh包含在h43处的cys;
[0727]
该vl包含在l5处的cys;
[0728]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0729]
该scfv处于vh-l-vl取向。
[0730]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0731]
该vh包含在h43处的cys;
[0732]
该vl包含在l3处的cys;
[0733]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0734]
该scfv处于vh-l-vl取向。
[0735]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0736]
该vh包含在h40处的cys;
[0737]
该vl包含在l100处的cys;
[0738]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0739]
该scfv处于vh-l-vl取向。
[0740]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0741]
该vh包含在h40处的cys;
[0742]
该vl包含在l102处的cys;
[0743]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0744]
该scfv处于vh-l-vl取向。
[0745]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0746]
该vh包含在h40处的cys;
[0747]
该vl包含在l5处的cys;
[0748]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0749]
该scfv处于vh-l-vl取向。
[0750]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0751]
该vh包含在h40处的cys;
[0752]
该vl包含在l3处的cys;
[0753]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0754]
该scfv处于vh-l-vl取向。
[0755]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0756]
该vh包含在h46处的cys;
[0757]
该vl包含在l100处的cys;
[0758]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0759]
该scfv处于vh-l-vl取向。
[0760]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0761]
该vh包含在h46处的cys;
[0762]
该vl包含在l102处的cys;
[0763]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0764]
该scfv处于vh-l-vl取向。
[0765]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0766]
该vh包含在h46处的cys;
[0767]
该vl包含在l5处的cys;
[0768]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0769]
该scfv处于vh-l-vl取向。
[0770]
本公开还提供了一种scfv,其包含vh、l和vl,其中
[0771]
该vh包含在h46处的cys;
[0772]
该vl包含在l3处的cys;
[0773]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0774]
该scfv处于vh-l-vl取向。
[0775]
在一些实施方案中,该l包含氨基酸序列seq id no:3。
[0776]
在一些实施方案中,该l包含氨基酸序列seq id no:4。
[0777]
在一些实施方案中,该l包含氨基酸序列seq id no:5。
[0778]
在一些实施方案中,该l包含氨基酸序列seq id no:6。
[0779]
在一些实施方案中,该l包含氨基酸序列seq id no:7。
[0780]
包含本公开的spfv的异源分子
[0781]
本公开的spfv可以类似于本领域已知的缺少二硫键的非稳定化scfv那样与第二分子缀合。示例性的第二分子在本文中公开,并且包括半衰期延长部分、成像剂、治疗剂、具有各种抗体形式的抗体及其片段、抗原结合结构域、fc区、免疫球蛋白重/轻链或其片段、多特异性分子和嵌合抗原受体(car)。
[0782]
本公开还提供了一种包含单链可变片段(scfv)的异源分子,该单链可变片段包含重链可变区(vh)、接头(l)和轻链可变区(vl),其中该scfv包含
[0783]
在结构保守的表面暴露的vh半胱氨酸(cys)与第一l cys之间的第一二硫键;
[0784]
在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键;或者
[0785]
在结构保守的表面暴露的vh cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键。
[0786]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0787]
该vh包含在结构保守的表面暴露的vh框架残基位置处的vh cys,并且该l包含第一l cys;
[0788]
该vl包含在结构保守的表面暴露的vl框架残基位置处的vl cys,并且该l包含第二l cys;或者
[0789]
该vh包含在结构保守的表面暴露的vh框架残基位置处的vh cys,该vl包含在结构保守的表面暴露的vl框架残基位置处的vl cys,并且该l包含第一l cys和第二l cys,其中vh cys和第一l cys能够形成二硫键并且vl cys和第二l cys能够形成二硫键。
[0790]
在一些实施方案中,vh cys与vl cys之间的距离为约至约
[0791]
在一些实施方案中,vh cys在h3、h5、h40、h43、h46或h105处,其中残基根据chothia进行编号。
[0792]
在一些实施方案中,vl cys在l3、l5、l39、l42、l45、l100或l102处,其中残基根据chothia进行编号。
[0793]
在一些实施方案中,vh cys在h105处并且vl cys在l42处。
[0794]
在一些实施方案中,vh cys在h43处并且vl cys在l100处。
[0795]
在一些实施方案中,vh cys在h3处并且vl cys在l3处。
[0796]
在一些实施方案中,vh cys在h3处并且vl cys在l5处。
[0797]
在一些实施方案中,vh cys在h3处并且vl cys在l39处。
[0798]
在一些实施方案中,vh cys在h3处并且vl cys在l42处。
[0799]
在一些实施方案中,vh cys在h3处并且vl cys在l45处。
[0800]
在一些实施方案中,vh cys在h3处并且vl cys在l100处。
[0801]
在一些实施方案中,vh cys在h3处并且vl cys在l102处。
[0802]
在一些实施方案中,vh cys在h5处并且vl cys在l3处。
[0803]
在一些实施方案中,vh cys在h5处并且vl cys在l5处。
[0804]
在一些实施方案中,vh cys在h5处并且vl cys在l39处。
[0805]
在一些实施方案中,vh cys在h5处并且vl cys在l42处。
[0806]
在一些实施方案中,vh cys在h5处并且vl cys在l45处。
[0807]
在一些实施方案中,vh cys在h5处并且vl cys在l100处。
[0808]
在一些实施方案中,vh cys在h5处并且vl cys在l102处。
[0809]
在一些实施方案中,vh cys在h40处并且vl cys在l3处。
[0810]
在一些实施方案中,vh cys在h40处并且vl cys在l5处。
[0811]
在一些实施方案中,vh cys在h40处并且vl cys在l39处。
[0812]
在一些实施方案中,vh cys在h40处并且vl cys在l42处。
[0813]
在一些实施方案中,vh cys在h40处并且vl cys在l45处。
[0814]
在一些实施方案中,vh cys在h40处并且vl cys在l100处。
[0815]
在一些实施方案中,vh cys在h40处并且vl cys在l102处。
[0816]
在一些实施方案中,vh cys在h43处并且vl cys在l3处。
[0817]
在一些实施方案中,vh cys在h43处并且vl cys在l5处。
[0818]
在一些实施方案中,vh cys在h43处并且vl cys在l39处。
[0819]
在一些实施方案中,vh cys在h43处并且vl cys在l42处。
[0820]
在一些实施方案中,vh cys在h43处并且vl cys在l45处。
[0821]
在一些实施方案中,vh cys在h43处并且vl cys在l100处。
[0822]
在一些实施方案中,vh cys在h43处并且vl cys在l102处。
[0823]
在一些实施方案中,vh cys在h46处并且vl cys在l3处。
[0824]
在一些实施方案中,vh cys在h46处并且vl cys在l5处。
[0825]
在一些实施方案中,vh cys在h46处并且vl cys在l39处。
[0826]
在一些实施方案中,vh cys在h46处并且vl cys在l42处。
[0827]
在一些实施方案中,vh cys在h46处并且vl cys在l45处。
[0828]
在一些实施方案中,vh cys在h46处并且vl cys在l100处。
[0829]
在一些实施方案中,vh cys在h46处并且vl cys在l102处。
[0830]
在一些实施方案中,vh cys在h105处并且vl cys在l3处。
[0831]
在一些实施方案中,vh cys在h105处并且vl cys在l5处。
[0832]
在一些实施方案中,vh cys在h105处并且vl cys在l39处。
[0833]
在一些实施方案中,vh cys在h105处并且vl cys在l42处。
[0834]
在一些实施方案中,vh cys在h105处并且vl cys在l45处。
[0835]
在一些实施方案中,vh cys在h105处并且vl cys在l100处。
[0836]
在一些实施方案中,vh cys在h105处并且vl cys在l102处。
[0837]
vh区和vl区的残基根据chothia进行编号。
[0838]
在一些实施方案中,该l包含衍生自免疫球蛋白(ig)铰链区的连续氨基酸序列。
[0839]
在一些实施方案中,ig铰链区衍生自人ig铰链区或非人ig铰链区。示例性非人ig铰链区是来自小鼠、大鼠、狗、鸡和非人灵长类动物(诸如猴)的那些。
[0840]
在一些实施方案中,ig铰链区衍生自人ig铰链区。
[0841]
在一些实施方案中,人ig铰链区是igg1、igg2、igg3、igg4、igm、iga或ige同种型。
[0842]
在一些实施方案中,该l包含氨基酸序列c(x)yc(seq id no:23),其中x是gly、ser、pro、ala、arg、asn、asp、glu、gln、his、ile、leu、lys、phe、thr、trp或tyr,并且y是1至3
的整数。可以将pro包含到该接头中以提供刚性。可以将gly包含到该接头中以允许最大的柔性。还可以在该l中使用除cys和met之外的任何其他氨基酸。
[0843]
在一些实施方案中,该l包含氨基酸序列c(x)yc(seq id no:24),其中x是gly、ser或pro,并且y是1至3的整数。
[0844]
在一些实施方案中,该l包含氨基酸序列cpc、cgc、csc、cppc(seq id no:1)、cgpc(seq id no:28)、cpgc(seq id no:29)、cggc(seq id no:30)、cspg(seq id no:31)、cpsc(seq id no:32)、cssc(seq id no:33)、cgsc(seq id no:34)、csgc(seq id no:35)、cpppc(seq id no:36)、cgppc(seq id no:37)、cpgpc(seq id no:38)、cppgc(seq id no:39)、cggpc(seq id no:40)、cpggc(seq id no:41)、cgggc(seq id no:42)、csppc(seq id no:43)、cpspc(seq id no:44)、cppsc(seq id no:45)、csspc(seq id no:46)、cpssc(seq id no:47)、csssc(seq id no:48)、cgspc(seq id no:49)、cpgsc(seq id no:50)、csgpc(seq id no:51)或cpsgc(seq id no:52)。
[0845]
在一些实施方案中,该l包含约14个至约19个氨基酸。
[0846]
在一些实施方案中,该l包含约14个氨基酸。
[0847]
在一些实施方案中,该l包含约15个氨基酸。
[0848]
在一些实施方案中,该l包含约16个氨基酸。
[0849]
在一些实施方案中,该l包含约17个氨基酸。
[0850]
在一些实施方案中,该l包含约18个氨基酸。
[0851]
在一些实施方案中,该l包含约19个氨基酸。
[0852]
在一些实施方案中,l包含氨基酸序列(x)mc(x)yc(x)n(seq id no:25);其中x是gly、ser、pro、ala、arg、asn、asp、glu、gln、his、ile、leu、lys、phe、thr、trp或tyr,m是6至9的整数,y是1至3的整数,并且n是4至6的整数。
[0853]
在一些实施方案中,该l包含氨基酸序列(x)mc(x)yc(x)n(seq id no:26);其中x是gly、ser或pro、ala、arg、asn、asp、glu、gln、his、ile、leu、lys、thr或tyr,m是6至9的整数,y是1至3的整数,并且n是4至6的整数。
[0854]
在一些实施方案中,该l包含氨基酸序列(x)mc(x)yc(x)n(seq id no:27);其中x是gly或pro,m是6至9的整数,y是1至3的整数,并且n是4至6的整数。
[0855]
在一些实施方案中,该l包含氨基酸序列seq id no:2、3、4、5、6或7。
[0856]
在一些实施方案中,本公开的spfv处于vl-l-vh取向。
[0857]
在一些实施方案中,本公开的spfv处于vh-l-vl取向。
[0858]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0859]
该vh包含在h105处的cys;
[0860]
该vl包含在l42处的cys;
[0861]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0862]
该scfv处于vl-l-vh取向。
[0863]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0864]
该vh包含在h105处的cys;
[0865]
该vl包含在l45处的cys;
[0866]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0867]
该scfv处于vl-l-vh取向。
[0868]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0869]
该vh包含在h105处的cys;
[0870]
该vl包含在l39处的cys;
[0871]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0872]
该scfv处于vl-l-vh取向。
[0873]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0874]
该vh包含在h5处的cys;
[0875]
该vl包含在l42处的cys;
[0876]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0877]
该scfv处于vl-l-vh取向。
[0878]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0879]
该vh包含在h5处的cys;
[0880]
该vl包含在l45处的cys;
[0881]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0882]
该scfv处于vl-l-vh取向。
[0883]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0884]
该vh包含在h5处的cys;
[0885]
该vl包含在l39处的cys;
[0886]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0887]
该scfv处于vl-l-vh取向。
[0888]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0889]
该vh包含在h3处的cys;
[0890]
该vl包含在l42处的cys;
[0891]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0892]
该scfv处于vl-l-vh取向。
[0893]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0894]
该vh包含在h3处的cys;
[0895]
该vl包含在l45处的cys;
[0896]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0897]
该scfv处于vl-l-vh取向。
[0898]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0899]
该vh包含在h3处的cys;
[0900]
该vl包含在l39处的cys;
[0901]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0902]
该scfv处于vl-l-vh取向。
[0903]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0904]
该vh包含在h43处的cys;
[0905]
该vl包含在l100处的cys;
[0906]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0907]
该scfv处于vh-l-vl取向。
[0908]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0909]
该vh包含在h43处的cys;
[0910]
该vl包含在l102处的cys;
[0911]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0912]
该scfv处于vh-l-vl取向。
[0913]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0914]
该vh包含在h43处的cys;
[0915]
该vl包含在l5处的cys;
[0916]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0917]
该scfv处于vh-l-vl取向。
[0918]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0919]
该vh包含在h43处的cys;
[0920]
该vl包含在l3处的cys;
[0921]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0922]
该scfv处于vh-l-vl取向。
[0923]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0924]
该vh包含在h40处的cys;
[0925]
该vl包含在l100处的cys;
[0926]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0927]
该scfv处于vh-l-vl取向。
[0928]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0929]
该vh包含在h40处的cys;
[0930]
该vl包含在l102处的cys;
[0931]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0932]
该scfv处于vh-l-vl取向。
[0933]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0934]
该vh包含在h40处的cys;
[0935]
该vl包含在l5处的cys;
[0936]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0937]
该scfv处于vh-l-vl取向。
[0938]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0939]
该vh包含在h40处的cys;
[0940]
该vl包含在l3处的cys;
[0941]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0942]
该scfv处于vh-l-vl取向。
[0943]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0944]
该vh包含在h46处的cys;
[0945]
该vl包含在l100处的cys;
[0946]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0947]
该scfv处于vh-l-vl取向。
[0948]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0949]
该vh包含在h46处的cys;
[0950]
该vl包含在l102处的cys;
[0951]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0952]
该scfv处于vh-l-vl取向。
[0953]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0954]
该vh包含在h46处的cys;
[0955]
该vl包含在l5处的cys;
[0956]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0957]
该scfv处于vh-l-vl取向。
[0958]
本公开还提供了一种包含scfv的异源分子,该scfv包含vh、l和vl,其中
[0959]
该vh包含在h46处的cys;
[0960]
该vl包含在l3处的cys;
[0961]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[0962]
该scfv处于vh-l-vl取向。
[0963]
在一些实施方案中,该l包含氨基酸序列seq id no:3。
[0964]
在一些实施方案中,该l包含氨基酸序列seq id no:4。
[0965]
在一些实施方案中,该l包含氨基酸序列seq id no:5。
[0966]
在一些实施方案中,该l包含氨基酸序列seq id no:6。
[0967]
在一些实施方案中,该l包含氨基酸序列seq id no:7。
[0968]
在一些实施方案中,本公开的scfv与第二蛋白质、多核苷酸、治疗剂、细胞毒素剂或可检测标签缀合。
[0969]
在一些实施方案中,第二蛋白质是半衰期延长部分。
[0970]
在一些实施方案中,第二蛋白质是抗体或其片段。
[0971]
在一些实施方案中,第二蛋白质是抗原结合片段。
[0972]
在一些实施方案中,第二蛋白质是治疗性分子。
[0973]
包含本公开的spfv和半衰期延长部分的异源分子
[0974]
在一些实施方案中,本公开的spfv与半衰期延长部分缀合。
[0975]
示例性的半衰期延长部分是免疫球蛋白(ig)、ig的片段、ig恒定区、ig恒定区的片段、fc区、转铁蛋白、白蛋白、白蛋白变体、白蛋白结合结构域或聚乙二醇。人ig的氨基酸序列是众所周知的,并且包括igg1、igg2、igg3、igg4、igm、iga和ige。
[0976]
在一些实施方案中,本公开的spfv与ig或ig的片段缀合。
[0977]
在一些实施方案中,本公开的spfv与fc区缀合。
[0978]
在一些实施方案中,本公开的spfv与转铁蛋白缀合。
[0979]
在一些实施方案中,本公开的spfv与白蛋白缀合。
[0980]
在一些实施方案中,本公开的spfv与白蛋白结合蛋白缀合。
[0981]
在一些实施方案中,本公开的spfv与聚乙二醇(peg)缀合。示例性peg分子是peg5000或peg20,000。
[0982]
在一些实施方案中,本公开的spfv与脂肪酸或脂肪酸酯缀合。示例性的脂肪酸和脂肪酸酯是用于获得期望特性的月桂酸酯、肉豆蔻酸酯、硬脂酸酯、花生酸酯、山嵛酸酯、油酸酯、花生四烯酸酯、辛二酸、十四烷二酸、十八烷二酸、二十二烷二酸等、聚赖氨酸、辛烷、碳水化合物(葡聚糖、纤维素、寡糖或多糖)。
[0983]
半衰期延长部分可以是与本公开的spfv直接融合的,并且可以通过标准克隆和表达技术产生。替代性地,众所周知的化学偶联方法可以用于将所述部分附接到本公开的重组产生的spfv。
[0984]
包含本公开的spfv和细胞毒素剂或可检测标签的异源分子
[0985]
本公开还提供了包含本公开的spfv的异源分子,其中本公开的spfv与第二蛋白质、多核苷酸、治疗剂、细胞毒素剂或可检测标签缀合。
[0986]
包含本公开的spfv的异源分子可以用于在体外或在体内指导治疗、介导杀伤、可视化、鉴定或纯化表达该spfv所结合的抗原的细胞。
[0987]
在一些实施方案中,可检测标签也是细胞毒素剂。
[0988]
可检测标签包括当缀合至本公开的spfv时使得该spfv能够经由光谱、光化学、生物化学、免疫化学或化学手段检测的组合物。
[0989]
示例性的可检测标签包括放射性同位素、磁珠、金属珠、胶体粒子、荧光染料、电子高密度试剂、酶(例如,如elisa中常用的)、生物素、地高辛、半抗原、发光分子、化学发光分子、荧光染料、荧光团、荧光淬灭剂、有色分子、放射性同位素、闪烁体、抗生物素蛋白、链霉抗生物素蛋白、蛋白质a、蛋白质g、抗体或其片段、多组氨酸、ni
2
、flag标签、myc标签、重金属、酶、碱性磷酸酶、过氧化物酶、荧光素酶、电子供体/受体、吖啶酯和比色底物。
[0990]
可检测标签可自发地发出信号,例如当可检测标签为放射性同位素时。在其他情况下,可检测标签由于受到外场的刺激而发出信号。
[0991]
示例性放射性同位素可以是发出γ、发出auger、发出β、发出α或发出正电子的放射性同位素。示例性放射性同位素包括3h、
11
c、
13
c、
15
n、
18
f、
19
f、
55
co、
57
co、
60
co、
61
cu、
62
cu、
64
cu、
67
cu、
68
ga、
72
as、
75
br、
86
y、
89
zr、
90
sr、
94m
tc、
99m
tc、
115
in、
123
1、
124
1、
125
i、
131
1、
211
at、
212
bi、
213
bi、
223
ra、
226
ra、
225
ac和
227
ac。
[0992]
示例性金属原子是原子序数大于20的金属,诸如钙、钪、钛、钒、铬、锰、铁、钴、镍、铜、锌、镓、锗、砷、硒、溴、氪、铷、锶、钇、锆、铌、钼、锝、钌、铑、钯、银、镉、铟、锡、锑、碲、碘、氙、铯、钡、镧、铪、钽、钨、铼、锇、铱、铂、金、汞、铊、铅、铋、钫、镭、锕、铈、镨、钕、钷、钐、铕、钆、铽、镝、钬、铒、铥、镱、镥、钍、镤、铀、镎、钚、镅、锔、锫、锎、锿、镄、钔、锘或铹原子。
[0993]
在一些实施方案中,金属原子可以是原子序数大于二十的碱土金属。
[0994]
在一些实施方案中,金属原子可以是镧系元素。
[0995]
在一些实施方案中,金属原子可以是锕系元素。
[0996]
在一些实施方案中,金属原子可以是过渡金属。
[0997]
在一些实施方案中,金属原子可以是贫金属。
[0998]
在一些实施方案中,金属原子可以是金原子、铋原子、钽原子和钆原子。
[0999]
在一些实施方案中,金属原子可以是原子序数为53(即碘)至83(即铋)的金属。
[1000]
在一些实施方案中,金属原子可以是适用于磁共振成像的原子。
[1001]
金属原子可以是 1、 2或 3氧化态形式的金属离子,诸如ba
2
、bi
3
、cs
、ca
2
、cr
2
、cr
3
、cr
6
、co
2
、co
3
、cu
、cu
2
、cu
3
、ga
3
、gd
3
、au
、au
3
、fe
2
、fe
3
、f
3
、pb
2
、mn
2
、mn
3
、mn
4
、mn
7
、hg
2
、ni
2
、ni
3
、ag
、sr
2
、sn
2
、sn
4
和zn
2
。金属原子可包括金属氧化物,诸如氧化铁、氧化锰或氧化钆。
[1002]
合适的染料包括任何可商购染料,诸如例如5(6)-羧基荧光素、irdye680rd马来酰亚胺或irdye 800cw、钌多吡啶染料等。
[1003]
合适的荧光团是异硫氰酸荧光素(fitc)、氨基硫脲荧光素、罗丹明、德克萨斯红、cydye(例如,cy3、cy5、cy5.5)、alexa fluor(例如,alexa488、alexa555、alexa594;alexa647)、近红外(nir)(700-900nm)荧光染料、以及碳菁和氨基苯乙烯基染料。
[1004]
包含与可检测标签缀合的本公开的scfv的异源分子可以用作成像剂。
[1005]
在一些实施方案中,细胞毒素剂是化疗剂、药物、生长抑制剂、毒素(例如,细菌、真菌、植物或动物来源的酶活性毒素,或其片段)或放射性同位素(即,放射性缀合物)。
[1006]
在一些实施方案中,细胞毒素剂是道诺霉素、阿霉素、甲氨蝶呤、长春地辛、细菌毒素(诸如,白喉毒素)、蓖麻毒素、格尔德霉素、美登木素生物碱或卡里奇霉素。细胞毒素剂可通过包括微管蛋白结合、dna结合或拓扑异构酶抑制在内的机制引发其细胞毒性和细胞抑制效应。
[1007]
在一些实施方案中,细胞毒素剂是酶促活性毒素,诸如白喉a链、白喉毒素的非结合活性片段、外毒素a链(来自铜绿假单胞菌(pseudomonas aeruginosa))、蓖麻毒素a链、相思豆毒素a链、蒴莲根毒素a链、α-八叠球菌素、油桐(aleurites fordii)蛋白质、石竹素蛋白质、美洲商陆(phytolaca americana)蛋白质(papi、papii和pap-s)、苦瓜(momordica charantia)抑制剂、麻疯树毒素、巴豆毒素、肥阜草(sapaonaria officinalis)抑制剂、白树毒素、迈托毒素、局限曲霉素、酚霉素、伊诺霉素和单端孢霉烯族化合物。
[1008]
在一些实施方案中,细胞毒素剂是放射性核素,诸如
212
bi、
131
i、
131
in、
90
y和
186
re。
[1009]
在一些实施方案中,细胞毒素剂是多拉司他汀或多拉司他汀肽类似物和衍生物、奥瑞司他汀或单甲基奥瑞司他汀苯丙氨酸。示例性分子在美国专利5,635,483和5,780,588中公开。多拉司他汀和奥瑞司他汀已被证实可干扰微管动力学、gtp水解以及细胞核和细胞分裂,并且具有抗癌和抗真菌活性。多拉司他汀或奥瑞司他汀药物部分可以通过肽药物部分的n(氨基)末端或c(羧基)末端(参见例如wo02/088172)或经由工程化到蛋白质中的任何半胱氨酸附接到本发明的抗体。
[1010]
可以使用已知方法进行与可检测标签的缀合。
[1011]
在一些实施方案中,可检测标签与螯合剂复合。
[1012]
在一些实施方案中,可检测标签经由接头与本公开的spfv缀合。
[1013]
可检测标签或细胞毒性部分可以使用已知方法直接或间接地连接到本公开的spfv。合适的接头是本领域已知的,包括例如辅基、非酚类接头(n-琥珀酰亚胺基-苯甲酸酯的衍生物;十二硼酸酯)、大环螯合剂和无环螯合剂两者的螯合部分,诸如1,4,7,10-四氮杂环十二烷-1,4,7,10-四乙酸(dota)的衍生物、二亚乙基三胺五乙酸(dtpa)的衍生物、s-2-(4-异硫氰酸基苄基)-1,4,7-三氮杂环壬烷-1,4,7-三乙酸(nota)的衍生物和1,4,8,11-四氮杂环十二烷-1,4,8,11-四乙酸(teta)的衍生物、n-琥珀酰亚胺基-3-(2-吡啶基二硫醇)
丙酸酯(spdp)、亚氨基噻吩(it)、亚胺酸酯的双官能衍生物(诸如己二亚胺酸二甲酯hcl)、活性酯(诸如双琥珀酰亚胺辛二酸酯)、醛(诸如戊二醛)、双叠氮基化合物(诸如双(对叠氮基苯甲酰基)己二胺)、双重氮衍生物(诸如双-(对重氮苯甲酰基)-乙二胺)、二异氰酸酯(诸如甲苯2,6-二异氰酸酯)和双活性氟化合物(诸如1,5-二氟-2,4-二硝基苯)以及其他螯合部分。合适的肽接头是公知的。
[1014]
包含本公开的spfv和免疫球蛋白(ig)恒定区或其片段的异源分子
[1015]
本公开的spfv可以与ig恒定区或ig恒定区的片段缀合,以赋予抗体样特性,包括fc效应子功能c1q结合、补体依赖性细胞毒性(cdc)、fc受体结合、抗体依赖性细胞介导的细胞毒性(adcc)、细胞表面受体(例如,b细胞受体;)bcr)的吞噬作用或下调。ig恒定区或ig恒定区的片段也作为如本文所述的半衰期延长部分起作用。还可以使用标准方法将本公开的spfv工程化到全长抗体中。包含本公开的spfv的全长抗体可以如本文所述进一步工程化。
[1016]
免疫球蛋白重链恒定区包含亚结构域ch1、铰链、ch2和ch3。在该重链上,ch1结构域跨残基118-215、ch2结构域跨残基231-340,并且ch3结构域跨残基341-447,残基根据eu索引进行编号。在一些情况下,残基341被称为ch2结构域残基。铰链通常被限定为包含人igg1的残基216并且终止于人igg1的残基230处,但是还可以包含从约残基231至约残基237的下铰链区,如本文所述。ig fc区至少包含ig恒定区的ch2结构域和ch3结构域,因此至少包含ig重链恒定区的从约231至447的区域。
[1017]
本发明还提供了与免疫球蛋白(ig)恒定区或ig恒定区的片段缀合的本公开的spfv。
[1018]
在一些实施方案中,ig恒定区是重链恒定区。
[1019]
在一些实施方案中,ig恒定区是轻链恒定区。
[1020]
在一些实施方案中,ig恒定区的片段包括fc区。
[1021]
在一些实施方案中,ig恒定区的片段包括ch2结构域。
[1022]
在一些实施方案中,ig恒定区的片段包括ch3结构域。
[1023]
在一些实施方案中,ig恒定区的片段包括ch2结构域和ch3结构域。
[1024]
在一些实施方案中,ig恒定区的片段包括铰链、ch2结构域和ch3结构域的至少一部分。铰链的部分是指ig铰链的一个或多个氨基酸残基。
[1025]
在一些实施方案中,ig恒定区的片段包括铰链、ch2结构域和ch3结构域。
[1026]
在一些实施方案中,本公开的spfv与ig恒定区或ig恒定区的片段的n末端缀合。
[1027]
在一些实施方案中,本公开的spfv与ig恒定区或ig恒定区的片段的c末端缀合。
[1028]
可以使用几种已知的测定法来评价与ig恒定区或ig恒定区的片段缀合的本公开的spfv的功能。可以使用本文所述的方法来评价与靶抗原的结合。可以在fc受体结合测定法中使用可溶形式的受体(诸如fcγri、fcγrii、fcγriii或fcrn),或使用基于细胞的测量例如adcc、cdc或adcp的测定法,来测定ig恒定结构域或ig恒定区的片段(诸如fc区)赋予的改变的特性。
[1029]
可以使用表达本公开的spfv所结合的抗原的细胞作为靶细胞并且使用nk细胞作为效应细胞,使用体外测定法来评价adcc。可以通过从裂解的细胞释放标签(例如,放射性底物、荧光染料或天然细胞内蛋白质)来检测细胞溶解。在示例性测定法中,按1个靶细胞比4个效应细胞的比率使用靶细胞。将靶细胞用batda预标记并与效应细胞和测试抗体混合。
将样本温育2小时,并且通过测量释放到上清液中的batda来测量细胞裂解率。使数据针对使用0.67%triton x-100(sigma aldrich)的最大细胞毒性和通过在不存在任何抗体的情况下来自靶细胞的batda自发释放测定的最小对照标准化。
[1030]
adcp可以通过使用单核细胞衍生的巨噬细胞作为效应细胞并且使用表达本公开的spfv所结合的抗原的任何细胞作为靶细胞来评估,所述靶细胞被工程化以表达gfp或其他标记分子。在示例性测定中,效应细胞:靶细胞比率可为例如4:1。可在含或不含本发明抗体的情况下,将效应细胞与靶细胞一起温育4小时。在温育后,可使用细胞消化液(accutase)分离细胞。可使用偶联至荧光标记的抗-cd11b抗体和抗-cd14抗体鉴定巨噬细胞,并且可使用标准方法基于cd11
cd14
巨噬细胞中的gfp荧光%确定吞噬百分比。
[1031]
可以例如通过以下方式测量细胞的cdc:将daudi细胞以1
×
105个细胞/孔(50μl/孔)铺板到rpmi-b(补充有1%bsa的rpmi)中,将50μl测试蛋白质以介于0μg/ml至100μg/ml之间的最终浓度添加到孔中,将反应在室温处温育15分钟,将11μl混合人血清添加到孔中,然后将反应在37℃下温育45分钟。可使用标准方法在facs测定法中检测碘化丙啶染色细胞%作为裂解细胞百分比(%)。
[1032]
包含本公开的spfv和嵌合抗原受体(car)或其片段的异源分子
[1033]
本公开的spfv可以与嵌合抗原受体(car)或car的片段缀合。包含本公开的spfv的car因此可以是单特异性的或多特异性的,包含本公开的一个或多个scfv分子作为其胞外结构域。
[1034]
嵌合抗原受体(car)是遗传工程化的受体。这些工程化受体可根据本领域已知的技术容易地插入包括t细胞在内的免疫细胞中并由其表达。对于car,单个受体可被编程为识别特异性抗原,并且当与该抗原结合时,激活免疫细胞以攻击和破坏携带该抗原的细胞。当这些抗原存在于靶细胞上时,表达car的免疫细胞可以靶向并且杀伤该靶细胞。
[1035]
car通常包含结合抗原的胞外结构域、任选的接头、跨膜结构域,以及包含共刺激结构域和/或信号传导结构域的胞质结构域。
[1036]
car的胞外结构域可以含有结合所需抗原(诸如本公开的scfv)的任何多肽。car也可被工程化以结合两个或更多个所需抗原,所述抗原可串联排列并由接头序列隔开。例如,本公开的一个或多个scfv、结构域抗体、美洲驼vhh抗体或其他仅vh抗体片段可以经由接头串联组织,以产生双特异性或多特异性car。
[1037]
car的跨膜结构域可来源于以下项的跨膜结构域:cd8、t细胞受体的α、β或ζ链、cd28、cd3ε、cd45、cd4、cd5、cd8、cd9、cd16、cd22、cd33、cd37、cd64、cd80、cd86、cd134、cd137、cd154、kirds2、ox40、cd2、cd27、lfa-1(cdi la、cd18)、icos(cd278)、4-1bb(cd137)、4-1bbl、gitr、cd40、baffr、hvem(lightr)、slamf7、nkp80(klrfi)、cd160、cd19、il2r β、il2r γ、il7r a、itga1、vla1、cd49a、itga4、ia4、cd49d、itga6、vla-6、cd49f、itgad、cdi id、itgae、cd103、itgal、cdi la、lfa-1、itgam、cdi lb、itgax、cdi lc、itgb1、cd29、itgb2、cd18、lfa-1、itgb7、tnfr2、dnam1(cd226)、slamf4(cd244、2b4)、cd84、cd96(触觉)、ceacam1、crt am、ly9(cd229)、cd160(by55)、psgl1、cd100(sema4d)、slamf6(ntb-a、lyl08)、slam(slamf1、cd150、ipo-3)、blame(slamf8)、selplg(cd162)、ltbr、pag/cbp、nkp44、nkp30、nkp46、nkg2d和/或nkg2c。
[1038]
car的细胞内共刺激结构域可来源于一种或多种共刺激分子的胞内结构域。共刺
激分子是熟知的除抗原受体或fc受体之外的细胞表面分子,其提供t淋巴细胞在与抗原结合时的有效活化和功能所需的第二信号。可用于car的示例性共刺激结构域是4-1bb、cd2、cd7、cd27、cd28、cd30、cd40、cd54(icam)、cd83、cd134(ox40)、cd150(slamf1)、cd152(ctla4)、cd223(lag3)、cd270(hvem)、cd278(icos)、dap10、lat、nkd2c slp76、trim和zap70的胞内结构域。
[1039]
car的细胞内信号传导结构域可以衍生自例如cd3ζ、cd3ε、cd22、cd79a、cd66d或cd39的信号传导结构域。“细胞内信号传导结构域”是指car多肽参与将有效car与靶抗原结合的消息转导到免疫效应细胞内部以引发效应细胞功能的部分,所述效应细胞功能例如激活、细胞因子产生、增殖和细胞毒性活性,包括细胞毒性因子向car结合的靶细胞的释放,或者在抗原结合到胞外car结构域后引发的其他细胞应答。
[1040]
定位在胞外结构域与跨膜结构域之间的car内的任选接头可以是长度为约2个至100个氨基酸的多肽。接头可包括柔性残基(诸如甘氨酸和丝氨酸)或由所述柔性残基构成,使得相邻的蛋白质结构域相对于彼此自由移动。当希望确保两个相邻结构域不会在空间上彼此干扰时,可使用较长的接头。接头可以是可裂解的或不可裂解的。示例性可裂解接头包括2a。
[1041]
示例性car包含本公开的scfv、cd8跨膜结构域和cd3ζ信号传导结构域。另一种示例性car包含本公开的scfv、cd8或cd28跨膜结构域、cd28、41bb或ox40共刺激结构域和cd3ζ信号传导结构域。
[1042]
car通过标准分子生物学技术来生成。
[1043]
本公开的spfv可以直接与第二分子缀合或经由接头与第二分子缀合。示例性接头包括免疫球蛋白铰链区的部分、衍生自任何免疫球蛋白重链或轻链同种型的cl或ch1、富含gly的接头、含有gly和ser的接头、含有gly和ala的接头、含有ala和ser的接头,以及含有pro的接头。可以被包含到接头中的示例性氨基酸为gly、ser、pro、thr、glu、lys、arg、ile、leu和his。替代性地,多种非蛋白质聚合物(包括聚乙二醇(peg)、聚丙二醇、聚氧化烯或聚乙二醇和聚丙二醇的共聚物)可以用作接头。示例性接头在例如国际专利公布号wo2019/060695中有所描述。
[1044]
在一些实施方案中,该异源分子是单特异性的。
[1045]
在一些实施方案中,该异源分子是多特异性的。
[1046]
在一些实施方案中,该异源分子是双特异性的。
[1047]
在一些实施方案中,该异源分子是三特异性的。
[1048]
在一些实施方案中,该异源分子是四特异性的。
[1049]
包含本公开的spfv的多特异性分子
[1050]
本公开还提供了一种包含单链可变片段(scfv)的多特异性分子,该单链可变片段包含重链可变区(vh)、接头(l)和轻链可变区(vl),其中该scfv包含
[1051]
在结构保守的表面暴露的vh半胱氨酸(cys)与第一l cys之间的第一二硫键;
[1052]
在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键;或者
[1053]
在结构保守的表面暴露的vh cys与第一l cys之间的第一二硫键,以及在结构保守的表面暴露的vl cys与第二l cys之间的第二二硫键。
[1054]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1055]
该vh包含在结构保守的表面暴露的vh框架残基位置处的vh cys,并且该l包含第一l cys;
[1056]
该vl包含在结构保守的表面暴露的vl框架残基位置处的vl cys,并且该l包含第二l cys;或者
[1057]
该vh包含在结构保守的表面暴露的vh框架残基位置处的vh cys,该vl包含在结构保守的表面暴露的vl框架残基位置处的vl cys,并且该l包含第一l cys和第二l cys,其中vh cys和第一l cys能够形成二硫键并且vl cys和第二l cys能够形成二硫键。
[1058]
在一些实施方案中,vh cys与vl cys之间的距离为约至约
[1059]
在一些实施方案中,vh cys在h3、h5、h40、h43、h46或h105处,其中残基根据chothia进行编号。
[1060]
在一些实施方案中,vl cys在l3、l5、l39、l42、l45、l100或l102处,其中残基根据chothia进行编号。
[1061]
在一些实施方案中,
[1062]
vh cys在h105处并且vl cys在l42处;
[1063]
vh cys在h43处并且vl cys在l100处;
[1064]
vh cys在h3处并且vl cys在l3处;
[1065]
vh cys在h3处并且vl cys在l5处;
[1066]
vh cys在h3处并且vl cys在l39处;
[1067]
vh cys在h3处并且vl cys在l42处;
[1068]
vh cys在h3处并且vl cys在l45处;
[1069]
vh cys在h3处并且vl cys在l100处;
[1070]
vh cys在h3处并且vl cys在l102处;
[1071]
vh cys在h5处并且vl cys在l3处;
[1072]
vh cys在h5处并且vl cys在l5处;
[1073]
vh cys在h5处并且vl cys在l39处;
[1074]
vh cys在h5处并且vl cys在l42处;
[1075]
vh cys在h5处并且vl cys在l45处;
[1076]
vh cys在h5处并且vl cys在l100处;
[1077]
vh cys在h5处并且vl cys在l102处;
[1078]
vh cys在h40处并且vl cys在l3处;
[1079]
vh cys在h40处并且vl cys在l5处;
[1080]
vh cys在h40处并且vl cys在l39处;
[1081]
vh cys在h40处并且vl cys在l42处;
[1082]
vh cys在h40处并且vl cys在l45处;
[1083]
vh cys在h40处并且vl cys在l100处;
[1084]
vh cys在h40处并且vl cys在l102处;
[1085]
vh cys在h43处并且vl cys在l3处;
[1086]
vh cys在h43处并且vl cys在l5处;
[1087]
vh cys在h43处并且vl cys在l39处;
[1088]
vh cys在h43处并且vl cys在l42处;
[1089]
vh cys在h43处并且vl cys在l45处;
[1090]
vh cys在h43处并且vl cys在l102处;
[1091]
vh cys在h46处并且vl cys在l3处;
[1092]
vh cys在h46处并且vl cys在l5处;
[1093]
vh cys在h46处并且vl cys在l39处;
[1094]
vh cys在h46处并且vl cys在l42处;
[1095]
vh cys在h46处并且vl cys在l45处;
[1096]
vh cys在h46处并且vl cys在l100处;
[1097]
vh cys在h46处并且vl cys在l102处;
[1098]
vh cys在h105处并且vl cys在l3处;
[1099]
vh cys在h105处并且vl cys在l5处;
[1100]
vh cys在h105处并且vl cys在l39处;
[1101]
vh cys在h105处并且vl cys在l45处;
[1102]
vh cys在h105处并且vl cys在l100处;或者
[1103]
vh cys在h105处并且vl cys在l102处;
[1104]
本文的残基根据chothia进行编号。
[1105]
在一些实施方案中,该l包含衍生自免疫球蛋白(ig)铰链区的连续氨基酸序列。
[1106]
在一些实施方案中,ig铰链区衍生自人ig铰链区或非人ig铰链区。
[1107]
在一些实施方案中,ig铰链区衍生自人ig铰链区。
[1108]
在一些实施方案中,人ig铰链区为igg1、igg2、igg3或igg4同种型。
[1109]
在一些实施方案中,该l包含氨基酸序列c(x)yc(seq id no:23),其中x是gly、ser、pro、ala、arg、asn、asp、glu、gln、his、ile、leu、lys、phe、thr、trp或tyr,并且y是1至3的整数。
[1110]
在一些实施方案中,该l包含氨基酸序列c(x)yc(seq id no:24),其中x是gly、ser或pro,并且y是1至3的整数。
[1111]
在一些实施方案中,该l包含氨基酸序列cpc、cgc、csc、cppc(seq id no:1)、cgpc(seq id no:28)、cpgc(seq id no:29)、cggc(seq id no:30)、cspg(seq id no:31)、cpsc(seq id no:32)、cssc(seq id no:33)、cgsc(seq id no:34)、csgc(seq id no:35)、cpppc(seq id no:36)、cgppc(seq id no:37)、cpgpc(seq id no:38)、cppgc(seq id no:39)、cggpc(seq id no:40)、cpggc(seq id no:41)、cgggc(seq id no:42)、csppc(seq id no:43)、cpspc(seq id no:44)、cppsc(seq id no:45)、csspc(seq id no:46)、cpssc(seq id no:47)、csssc(seq id no:48)、cgspc(seq id no:49)、cpgsc(seq id no:50)、csgpc(seq id no:51)或cpsgc(seq id no:52)。
[1112]
在一些实施方案中,该l包含约14个至约19个氨基酸,诸如约14个、约15个、约16个、约17个、约18个或约19个氨基酸。
[1113]
在一些实施方案中,l包含氨基酸序列(x)mc(x)yc(x)n(seq id no:25);其中x是gly、ser、pro、ala、arg、asn、asp、glu、gln、his、ile、leu、lys、phe、thr、trp或tyr,m是6至9的整数,y是1至3的整数,并且n是4至6的整数。
[1114]
在一些实施方案中,该l包含氨基酸序列(x)mc(x)yc(x)n(seq id no:26);其中x是gly、ser、pro、ala、arg、asn、asp、glu、gln、his、ile、leu、lys、thr或tyr,m是6至9的整数,y是1至3的整数,并且n是4至6的整数。
[1115]
在一些实施方案中,该l包含氨基酸序列(x)mc(x)yc(x)n(seq id no:27);其中x是gly或pro,m是6至9的整数,y是1至3的整数,并且n是4至6的整数。
[1116]
在一些实施方案中,该l包含氨基酸序列seq id no:2、3、4、5、6或7。
[1117]
在一些实施方案中,本公开的spfv处于vl-l-vh取向。
[1118]
在一些实施方案中,本公开的spfv处于vh-l-vl取向。
[1119]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1120]
该vh包含在h105处的cys;
[1121]
该vl包含在l42处的cys;
[1122]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1123]
该scfv处于vl-l-vh取向。
[1124]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1125]
该vh包含在h105处的cys;
[1126]
该vl包含在l45处的cys;
[1127]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1128]
该scfv处于vl-l-vh取向。
[1129]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1130]
该vh包含在h105处的cys;
[1131]
该vl包含在l39处的cys;
[1132]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1133]
该scfv处于vl-l-vh取向。
[1134]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1135]
该vh包含在h5处的cys;
[1136]
该vl包含在l42处的cys;
[1137]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1138]
该scfv处于vl-l-vh取向。
[1139]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1140]
该vh包含在h5处的cys;
[1141]
该vl包含在l45处的cys;
[1142]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1143]
该scfv处于vl-l-vh取向。
[1144]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1145]
该vh包含在h5处的cys;
[1146]
该vl包含在l39处的cys;
[1147]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1148]
该scfv处于vl-l-vh取向。
[1149]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1150]
该vh包含在h3处的cys;
[1151]
该vl包含在l42处的cys;
[1152]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1153]
该scfv处于vl-l-vh取向。
[1154]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1155]
该vh包含在h3处的cys;
[1156]
该vl包含在l45处的cys;
[1157]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1158]
该scfv处于vl-l-vh取向。
[1159]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1160]
该vh包含在h3处的cys;
[1161]
该vl包含在l39处的cys;
[1162]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1163]
该scfv处于vl-l-vh取向。
[1164]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1165]
该vh包含在h43处的cys;
[1166]
该vl包含在l100处的cys;
[1167]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1168]
该scfv处于vh-l-vl取向。
[1169]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1170]
该vh包含在h43处的cys;
[1171]
该vl包含在l102处的cys;
[1172]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1173]
该scfv处于vh-l-vl取向。
[1174]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1175]
该vh包含在h43处的cys;
[1176]
该vl包含在l5处的cys;
[1177]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1178]
该scfv处于vh-l-vl取向。
[1179]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1180]
该vh包含在h43处的cys;
[1181]
该vl包含在l3处的cys;
[1182]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1183]
该scfv处于vh-l-vl取向。
[1184]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1185]
该vh包含在h40处的cys;
[1186]
该vl包含在l100处的cys;
[1187]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1188]
该scfv处于vh-l-vl取向。
[1189]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1190]
该vh包含在h40处的cys;
[1191]
该vl包含在l102处的cys;
[1192]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1193]
该scfv处于vh-l-vl取向。
[1194]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1195]
该vh包含在h40处的cys;
[1196]
该vl包含在l5处的cys;
[1197]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1198]
该scfv处于vh-l-vl取向。
[1199]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1200]
该vh包含在h40处的cys;
[1201]
该vl包含在l3处的cys;
[1202]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1203]
该scfv处于vh-l-vl取向。
[1204]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1205]
该vh包含在h46处的cys;
[1206]
该vl包含在l100处的cys;
[1207]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1208]
该scfv处于vh-l-vl取向。
[1209]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1210]
该vh包含在h46处的cys;
[1211]
该vl包含在l102处的cys;
[1212]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1213]
该scfv处于vh-l-vl取向。
[1214]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1215]
该vh包含在h46处的cys;
[1216]
该vl包含在l5处的cys;
[1217]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1218]
该scfv处于vh-l-vl取向。
[1219]
本公开还提供了一种包含scfv的多特异性分子,该scfv包含vh、l和vl,其中
[1220]
该vh包含在h46处的cys;
[1221]
该vl包含在l3处的cys;
[1222]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1223]
该scfv处于vh-l-vl取向。
[1224]
在一些实施方案中,该l包含氨基酸序列seq id no:3。
[1225]
在一些实施方案中,该l包含氨基酸序列seq id no:6。
[1226]
在一些实施方案中,该l包含氨基酸序列seq id no:7。
[1227]
在一些实施方案中,该多特异性分子包括抗体或抗体片段。
[1228]
在一些实施方案中,该多特异性蛋白质包括ig恒定区或ig恒定区的片段。
[1229]
在一些实施方案中,ig恒定区包括fc区。
[1230]
在一些实施方案中,ig恒定区包括ch2结构域。
[1231]
在一些实施方案中,ig恒定区的片段包括ch3结构域。
[1232]
在一些实施方案中,ig恒定区的片段包括ch2结构域和ch3结构域。
[1233]
在一些实施方案中,ig恒定区的片段包括铰链、ch2结构域和ch3结构域的至少一部分。
[1234]
在一些实施方案中,ig恒定区的片段包括铰链、ch2结构域和ch3结构域。
[1235]
在一些实施方案中,本公开的spfv与ig恒定区的n末端或ig恒定区的片段的n末端缀合。
[1236]
在一些实施方案中,本公开的spfv与ig恒定区的c末端或ig恒定区的片段的c末端缀合。
[1237]
在一些实施方案中,ig恒定区或ig恒定区的片段是igg1、igg2、以及igg3或igg4同种型。
[1238]
在一些实施方案中,ig恒定区或ig恒定区的片段包含导致多特异性分子与fcγr的结合降低的至少一个突变。
[1239]
在一些实施方案中,导致多特异性分子与fcγr的结合降低的至少一个突变选自由以下项组成的组:f234a/l235a、l234a/l235a、l234a/l235a/d265s、v234a/g237a/p238s/h268a/v309l/a330s/p331s、f234a/l235a、s228p/f234a/l235a、n297a、v234a/g237a、k214t/e233p/l234v/l235a/g236缺失/a327g/p331a/d365e/l358m、h268q/v309l/a330s/p331s、s267e/l328f、l234f/l235e/d265a、l234a/l235a/g237a/p238s/h268a/a330s/p331s、s228p/f234a/l235a/g237a/p238s和s228p/f234a/l235a/g236缺失/g237a/p238s,其中残基根据eu索引进行编号。
[1240]
在一些实施方案中,ig恒定区或ig恒定区的片段包含导致多特异性分子与fcγr的结合增强的至少一个突变。
[1241]
在一些实施方案中,导致多特异性分子与fcγr的结合增强的至少一个突变选自由以下项组成的组:s239d/i332e、s298a/e333a/k334a、f243l/r292p/y300l、f243l/r292p/y300l/p396l、f243l/r292p/y300l/v305i/p396l和g236a/s239d/i332e,其中残基根据eu索引进行编号。
[1242]
在一些实施方案中,fcγr为fcγri、fcγriia、fcγriib或fcγriii。
[1243]
在一些实施方案中,ig恒定区或ig恒定区的片段包含调节多特异性分子的半衰期的至少一个突变。
[1244]
在一些实施方案中,调节多特异性分子的半衰期的至少一个突变选自由以下项组成的组:h435a、p257i/n434h、d376v/n434h、m252y/s254t/t256e/h433k/n434f、t308p/n434a和h435r,其中残基根据eu索引进行编号。
[1245]
在一些实施方案中,ig恒定区或ig恒定区的片段在ch3结构域中包含至少一个突变。
[1246]
在一些实施方案中,ch3结构域中的至少一个突变选自由以下项组成的组:t350v、l351y、f405a、y407v、t366y、t366w、f405w、t394w、t394s、y407t、y407a、t366s/l368a/
y407v、l351y/f405a/y407v、t366i/k392m/t394w、f405a/y407v、t366l/k392m/t394w、l351y/y407a、t366a/k409f、l351y/y407a、t366v/k409f、t366a/k409f、t350v/l351y/f405a/y407v和t350v/t366l/k392l/t394w,其中残基根据eu索引进行编号。
[1247]
在一些实施方案中,该多特异性分子是双特异性的。
[1248]
在一些实施方案中,该多特异性分子是三特异性的。
[1249]
在一些实施方案中,该多特异性分子是四特异性的。
[1250]
5.2产生包含本公开的spfv的多特异性蛋白质
[1251]
本公开的spfv可以使用已知的重组技术、表达和纯化方案工程化到任何已知形式的多特异性分子中。
[1252]
本公开的spfv可以被工程化到在ch3结构域中具有一个或多个突变的全长多特异性抗体中,所述突变促进两个半分子的稳定性。这些多特异性抗体可以使用fab臂交换或通过各种链的共表达在体外产生。对于体外fab臂交换,将两个单特异性二价抗体工程化为在ch3结构域中具有一个或多个置换,将所述抗体在足以允许铰链区中的半胱氨酸经历二硫键异构化的还原条件下一起温育;从而通过fab臂交换产生双特异性抗体。温育条件最理想地可恢复到非还原条件。可使用的示例性还原剂为2-巯基乙胺(2-mea)、二硫苏糖醇(dtt)、二硫赤藓糖醇(dte)、谷胱甘肽、三(2-羧乙基)膦(tcep)、l-半胱氨酸和β-巯基乙醇,优选地为选自由2-巯基乙胺、二硫苏糖醇和三(2-羧乙基)膦组成的组的还原剂。例如,可使用如下条件:在至少25mm 2-mea的存在下或至少0.5mm二硫苏糖醇的存在下,在5-8的ph例如ph7.0或ph7.4,至少20℃的温度下,温育至少90分钟。
[1253]
可以使用的ch3突变包括诸如杵臼突变(genentech)、静电匹配突变(chugai,amgen,novonordisk,oncomed)、链交换工程化结构域主体(seedbody)(emd serono)、突变(genmab)以及其他非对称突变(例如,zymeworks)的技术。
[1254]
旋钮空穴突变公开于例如wo1996/027011中,并且包括ch3区域的界面上的突变,其中具有小侧链(空穴)的氨基酸被引入第一ch3区域,而具有大侧链(旋钮)的氨基酸被引入第二ch3区域,导致第一ch3区域和第二ch3区域之间的优先相互作用。形成旋钮和空穴的示例性ch3区域突变是t366y/f405a、t366w/f405w、f405w/y407a、t394w/y407t、t394s/y407a、t366w/t394s、f405w/t394s和t366w/t366s_l368a_y407v。
[1255]
重链异源二聚体的形成可通过使用静电相互作用,通过置换第一ch3区域上的带正电残基和第二ch3区域上的带负电残基来促进,如us2010/0015133、us2009/0182127、us2010/028637或us2011/0123532中所述。
[1256]
可用于促进重链异源二聚化的其他非对称突变是l351y_f405a_y407v/t394w、t366i_k392m_t394w/f405a_y407v、t366l_k392m_t394w/f405a_y407v、l351y_y407a/t366a_k409f、l351y_y407a/t366v_k409f、y407a/t366a_k409f或t350v_l351y_f405a_y407v/t350v_t366l_k392l_t394w,如us2012/0149876或us2013/0195849(zymeworks)中所述。
[1257]
seedbody突变涉及将选择的igg残基置换为iga残基以促进重链异源二聚化,如us20070287170中所述。
[1258]
可以使用的其他示例性突变是r409d_k370e/d399k_e357k、s354c_t366w/y349c_t366s_l368a_y407v、y349c_t366w/s354c_t366s_l368a_y407v、t366k/l351d、l351k/
y349e、l351k/y349d、l351k/l368e、l351y_y407a/t366a_k409f、l351y_y407a/t366v_k409f、k392d/d399k、k392d/e356k、k253e_d282k_k322d/d239k_e240k_k292d、k392d_k409d/d356k_d399k,如wo2007/147901、wo2011/143545、wo2013/157954、wo2013/096291和us2018/0118849中所述。
[1259]
突变(genmab)公开于例如us2014/0303356中,并且包括突变f405l/k409r、野生型/f405l_r409k、t350i_k370t_f405l/k409r、k370w/k409r、d399afghilmnrstvwy/k409r、t366adefghilmqvy/k409r、l368adeghnrstvq/k409agrh、d399fhkrq/k409agrh、f405iklstvw/k409agrh和y407lwq/k409agrh。
[1260]
本公开的spfv可以结合到其中的另外的双特异性或多特异性结构包括:双可变结构域免疫球蛋白(dvd)(国际专利公布号wo2009/134776;dvd是全长抗体,其包含具有vh1-接头-vh2-ch结构的重链和具有vl1-接头-vl2-cl结构的轻链;接头是任选的)、包括多种二聚化结构域以连接具有不同特异性的两个抗体臂的结构诸如亮氨酸拉链或胶原二聚化结构域(国际专利公布wo2012/022811、美国专利5,932,448;美国专利6,833,441)、缀合在一起的两个或更多个结构域抗体(dab)、双价抗体、仅重链抗体诸如骆驼科抗体和工程化骆驼科抗体、双重靶向(dt)-ig(gsk/domantis)、二合一抗体(genentech)、交联mab(karmanos cancer center)、mab2(f-star)和covx-主体(covx/pfizer)、igg样双特异性抗体(innclone/eli lilly)、ts2ab(medimmune/az)和bsab(zymogenetics)、hercules(biogen idec)和tvab(roche)、scfv/fc融合体(academic institution)、scorpion(emergent biosolutions/trubion,zymogenetics/bms)、双亲和重靶向技术(fc-dart)(macrogenics)和双(scfv)
2-fab(national research center for antibody medicine
‑‑
china)、双功能或bis-fab(genentech)、对接锁定(dnl)(immunomedics)、二价双特异性(biotecnol)和fab-fv(ucb-celltech)。基于scfv的、基于双价抗体的结构域抗体包括但不限于双特异性t细胞衔接器(bite)(micromet)、串联双价抗体(tandab)(affimed)、双亲和重靶向技术(dart)(macrogenics)、单链双价抗体(academic)、tcr样抗体(ait,receptorlogics)、人血清白蛋白scfv融合体(merrimack)和combody(epigen biotech)、双重靶向纳米抗体(ablynx)、双重靶向仅重链结构域抗体。
[1261]
本公开的scfv还可以被工程化到包含三条多肽链的多特异性蛋白质中。在此类设计中,至少一个抗原结合结构域为本公开的scfv的形式。示例性设计包括(其中“1”表示第一抗原结合结构域,“2”表示第二抗原结合结构域,并且“3”表示第三抗原结合结构域:
[1262]
设计1:链a)scfv1-ch2-ch3;链b)vl2-cl;链c)vh2-ch1-铰链-ch2-ch3
[1263]
设计2:链a)scfv1-铰链-ch2-ch3;链b)vl2-cl;链c)vh2-ch1-铰链-ch2-ch3
[1264]
设计3:链a)scfv1-ch1-铰链-ch2-ch3;链b)vl2-cl;链c)vh2-ch1-铰链-ch2-ch3
[1265]
设计4:链a)ch2-ch3-scfv1;链b)vl2-cl;链c)vh2-ch1-铰链-ch2-ch3
[1266]
ch3工程化可以结合到设计1至4,诸如,如us2012/0149876或us2013/0195849(zymeworks)中所述的突变l351y_f405a_y407v/t394w、t366i_k392m_t394w/f405a_y407v、t366l_k392m_t394w/f405a_y407v、l351y_y407a/t366a_k409f、l351y_y407a/t366v_k409f、y407a/t366a_k409f或t350v_l351y_f405a_y407v/t350v_t366l_k392l_t394w。
[1267]
5.3同种型、同种异型和fc工程化
[1268]
本公开的多特异性分子或异源分子中存在的ig恒定区或ig恒定区的片段(诸如fc
区)可以是任何同种异型或同种型。
[1269]
在一些实施方案中,ig恒定区或ig恒定区的片段是igg1同种型。
[1270]
在一些实施方案中,ig恒定区或ig恒定区的片段是igg2同种型。
[1271]
在一些实施方案中,ig恒定区或ig恒定区的片段是igg3同种型。
[1272]
在一些实施方案中,ig恒定区或ig恒定区的片段是igg4同种型。
[1273]
ig恒定区或ig恒定区的片段可以是任何同种异型。预期同种异型对ig恒定区的特性(诸如结合或fc介导的效应功能)没有影响。包含ig恒定区或其片段的治疗性蛋白质的免疫原性与输注反应风险的增加和治疗应答持续时间的减少相关联(baert等人,(2003)n engl j med 348:602-608)。包含ig恒定区或其片段的治疗性蛋白质在宿主中诱导免疫应答的程度可以部分地由ig恒定区的同种异型来确定(stickler等人,(2011)genes and immunity 12:213-221)。ig恒定区同种异型与抗体的恒定区序列中特定位置处的氨基酸序列变异相关。表3示出选择的igg1、igg2和igg4同种异型。
[1274]
表3.
[1275][1276]
c末端赖氨酸(ctl)可以通过血流中的内源性循环羧肽酶从ig恒定区除去(cai等人,(2011)biotechnol bioeng 108:404-412)。在制备期间,可通过控制细胞外zn
2
、edta或edta
–
fe
3
的浓度,将ctl移除控制到小于最大水平,如美国专利公开号us2014/0273092中所述。蛋白质的ctl含量可以使用已知方法测量。
[1277]
在一些实施方案中,与ig恒定区缀合的本公开的spfv具有约10%至约90%的c末端赖氨酸含量。在一些实施方案中,c末端赖氨酸含量为约20%至约80%。在一些实施方案中,c末端赖氨酸含量为约40%至约70%。在一些实施方案中,c末端赖氨酸含量为约55%至约70%。在一些实施方案中,c末端赖氨酸含量为约60%。
[1278]
可以对本公开的包含ig恒定区或ig恒定区的片段的多特异性分子或异源分子进行fc区突变,以调节它们的效应功能,诸如adcc、adcp和/或adcp和/或药物动力学特性。这可以通过向fc中引入突变来实现,所述突变调节突变的fc与激活fcγr(fcγri、fcγriia、fcγriii)、抑制性fcγriib和/或fcrn的结合。
[1279]
在一些实施方案中,本公开的多特异性分子或异源分子在ig恒定区或ig恒定区的片段中包含至少一个突变。
[1280]
在一些实施方案中,所述至少一个突变在fc区中。
[1281]
在一些实施方案中,本公开的多特异性分子或异源分子在fc区中包含至少一个、两个、三个、四个、五个、六个、七个、八个、九个、十个、十一个、十二个、十三个、十四个或十五个突变。
[1282]
在一些实施方案中,本公开的多特异性分子或异源分子在fc区中包含调节抗体与fcrn的结合的至少一个突变。
[1283]
可以被突变以调节半衰期(例如,与fcrn结合)的fc位置包括位置250、252、253、254、256、257、307、376、380、428、434和435。可以单独或组合进行的示例性突变为突变t250q、m252y、i253a、s254t、t256e、p257i、t307a、d376v、e380a、m428l、h433k、n434s、n434a、n434h、n434f、h435a和h435r。可以进行以增加半衰期的示例性的单独突变或组合突变为突变m428l/n434s、m252y/s254t/t256e、t250q/m428l、n434a和t307a/e380a/n434a。可以进行以缩短半衰期的示例性的单独突变或组合突变为突变h435a、p257i/n434h、d376v/n434h、m252y/s254t/t256e/h433k/n434f、t308p/n434a和h435r。
[1284]
在一些实施方案中,本公开的多特异性分子或异源分子在fc区中包含m252y/s254t/t256e突变。
[1285]
在一些实施方案中,本公开的多特异性分子或异源分子在fc区中包含至少一个突变,其降低蛋白质与激活fcγ受体(fcγr)的结合并且/或者降低fc效应功能,诸如c1q结合、补体依赖性细胞毒性(cdc)、抗体依赖性细胞介导的细胞毒性(adcc)或吞噬作用(adcp)。
[1286]
可以被突变以降低本公开的多特异性分子或异源分子与激活fcγr的结合并且随后降低效应功能的fc位置包括位置214、233、234、235、236、237、238、265、267、268、270、295、297、309、327、328、329、330、331和365。可以单独或组合进行的示例性突变为igg1、igg2、igg3或igg4中的突变k214t、e233p、l234v、l234a、g236缺失、v234a、f234a、l235a、g237a、p238a、p238s、d265a、d265s、s267e、h268a、h268q、q268a、n297a、a327q、p329a、d270a、q295a、v309l、a327s、l328f、a330s和p331s。导致本公开的多特异性分子或异源分子具有降低的adcc的示例性组合突变为以下突变:igg1上的l234a/l235a、igg1上的l234a/l235a/d265s、igg2上的v234a/g237a/p238s/h268a/v309l/a330s/p331s、igg4上的f234a/l235a、igg4上的s228p/f234a/l235a、所有ig同种型上的n297a、igg2上的v234a/g237a、igg1上的k214t/e233p/l234v/l235a/g236缺失/a327g/p331a/d365e/l358m、igg2上的h268q/v309l/a330s/p331s、igg1上的s267e/l328f、igg1上的l234f/l235e/d265a、igg1上的l234a/l235a/g237a/p238s/h268a/a330s/p331s、igg4上的s228p/f234a/l235a/g237a/p238s,以及igg4上的s228p/f234a/l235a/g236缺失/g237a/p238s。还可使用杂合igg2/4fc域,例如具有来自igg2的残基117-260和来自igg4的残基261-447的fc。
[1287]
导致本公开的多特异性分子或异源分子具有降低的cdc的示例性突变为k322a突变。
[1288]
众所周知的s228p突变可以在igg4中进行,以增强igg4稳定性。
[1289]
在一些实施方案中,本公开的多特异性分子或异源分子在fc区中包含选自由以下项组成的组的至少一个突变:k214t、e233p、l234v、l234a、g236缺失、v234a、f234a、l235a、g237a、p238a、p238s、d265a、s267e、h268a、h268q、q268a、n297a、a327q、p329a、d270a、q295a、v309l、a327s、l328f、a330s和p331s。
[1290]
在一些实施方案中,本公开的多特异性分子或异源分子在fc区中包含l234a/l235a/d265s突变。
[1291]
在一些实施方案中,本公开的多特异性分子或异源分子在fc区中包含l234a/
l235a突变。
[1292]
在一些实施方案中,本公开的多特异性分子或异源分子在fc区中包含至少一个突变,其增强本公开的多特异性分子或异源分子与fcγr的结合并且/或者增强fc效应功能,诸如c1q结合、补体依赖性细胞毒性(cdc)、抗体依赖性细胞介导的细胞毒性(adcc)和/或吞噬作用(adcp)。
[1293]
可以被突变以增加本公开的多特异性分子或异源分子与激活fcγr的结合并且/或者增强fc效应功能的fc位置包括位置236、239、243、256,290,292、298、300、305、312、326、330、332、333、334、345、360、339、378、396或430(残基根据eu索引进行编号)。可以单独或组合进行的示例性突变为g236a、s239d、f243l、t256a、k290a、r292p、s298a、y300l、v305l、k326a、a330k、i332e、e333a、k334a、a339t和p396l。导致蛋白质具有增加的adcc或adcp的示例性组合突变为s239d/i332e、s298a/e333a/k334a、f243l/r292p/y300l、f243l/r292p/y300l/p396l、f243l/r292p/y300l/v305i/p396l和g236a/s239d/i332e。
[1294]
可以被突变以增强cdc的fc位置包括位置267、268、324、326、333、345和430。可以单独或组合进行的示例性突变为s267e、f1268f、s324t、k326a、k326w、e333a、e345k、e345q、e345r、e345y、e430s、e430f和e430t。导致本公开的多特异性分子或异源分子具有增加的cdc的示例性组合突变为k326a/e333a、k326w/e333a、h268f/s324t、s267e/h268f、s267e/s324t和s267e/h268f/s324t。
[1295]
本文所述的特异性突变是分别与igg1、igg2和igg4野生型氨基酸序列seq id no:66、67和68相比时的突变。
[1296]
seq id no:66,野生型igg1
[1297]
astkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsrdeltknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1298]
seq id no:67;野生型igg2
[1299]
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssnfgtqtytcnvdhkpsntkvdktverkccvecppcpappvagpsvflfppkpkdtlmisrtpevtcvvvdvshedpevqfnwyvdgvevhnaktkpreeqfnstfrvvsvltvvhqdwlngkeykckvsnkglpapiektisktkgqprepqvytlppsreemtknqvsltclvkgfypsdisvewesngqpennykttppmldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1300]
seq id no: 68;野生型igg4
[1301]
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcpscpapeflggpsvflfppkpkdtlmisrtpevtcvvvdvsqedpevqfnwyvdgvevhnaktkpreeqfnstyrvvsvltvlhqdwlngkeykckvsnkglpssiektiskakgqprepqvytlppsqeemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqegnvfscsvmhealhnhytqkslslslgk
[1302]
可以在被工程化以表达每种受体的细胞上使用流式细胞术来评价本公开的多特异性分子或异源分子与fcγr或fcrn的结合。在示例性结合测定中,将2
×
105个细胞/孔接
种到96孔板中并在bsa染色缓冲液(美国圣何塞的bd biosciences(bd biosciences,san jose,usa))中于4℃封闭30分钟。将细胞与本公开的测试多特异性分子或测试异源分子在冰上于4℃下温育1.5小时。在用bsa染色缓冲液洗涤两次之后,将细胞与r-pe标记的抗人igg二级抗体(jackson immunoresearch laboratories)于4℃一起温育45分钟。将细胞在染色缓冲液中洗涤两次,然后重悬于150μl的含有1:200稀释的draq7活/死染色剂的染色缓冲液(cell signaling technology,danvers,usa)中。通过miltenyi macsquant流式细胞仪(miltenyi biotec,auburn,usa),分别使用b2和b4通道来检测经染色细胞的pe信号和draq7信号。活细胞根据draq7排除法进行门控,并且测定所收集的至少10,000个活事件的几何平均荧光信号。使用flowjo软件(tree star)进行分析。将数据绘制为抗体浓度对平均荧光信号的对数。进行非线性回归分析。
[1303]
5.4糖工程化
[1304]
通过工程化ig恒定区或ig恒定区低聚糖组分的片段,可以增强与ig恒定区或ig恒定区的片段缀合的本公开的多特异性分子或异源分子介导adcc的能力。人igg1或igg3在asn297处被熟知的双分枝g0、g0f、g1、g1f、g2或g2f形式的大多数聚糖n-糖基化。含有ig恒定区的蛋白质可以由通常具有至少约85%聚糖岩藻糖含量的非工程化cho细胞产生。从附接到ig恒定区或ig恒定区的片段的双触角复合物型低聚糖去除核心岩藻糖,经由改善的fcγriiia结合且不改变抗原结合或cdc活性,增强了本公开的多特异性分子或异源分子的adcc。此类多特异性分子或异源分子可以使用不同的方法来获得,所述方法据报道导致成功地表达携带双触角复合物型fc低聚糖的相对高脱岩藻糖基化免疫球蛋白,诸如控制培养物的重量克分子渗透压浓度(konno等人,(2012)cytotechnology 64:249-265)、应用变体cho细胞系lec13作为宿主细胞系(shields等人,(2002)j biol chem 277:26733-26740)、应用变体cho细胞系eb66作为宿主细胞系(olivier等人,(2010)mabs;2:405-415)、应用大鼠杂交瘤细胞系yb2/0作为宿主细胞系(shinkawa等人,(2003)j biol chem 278:3466-3473)、引入特异性地针对1,6-岩藻糖基转移酶(fut8)基因的小干扰rna(mori等人,(2004)biotechnol bioeng 88:901-908),或者共表达β-1,4-n-乙酰葡糖胺基转移酶iii和高尔基体α-甘露糖苷酶ii或强效α-甘露糖苷酶i抑制剂、几夫碱(ferrara等人,(2006)j biol chem 281:5032-5036)。
[1305]
在一些实施方案中,包含ig恒定区或ig恒定区的片段的本公开的多特异性分子或异源分子具有双触角聚糖结构,其岩藻糖含量介于约1%至约15%之间,例如约15%、14%、13%、12%、11%、10%、9%、8%、7%、6%、5%、4%、3%、2%或1%。在一些实施方案中,包含ig恒定区或ig恒定区的片段的本公开的多特异性分子或异源分子具有聚糖结构,其岩藻糖含量为约50%、40%、45%、40%、35%、30%、25%或20%。
[1306]“岩藻糖含量”是指糖链内asn297处的岩藻糖单糖的量。岩藻糖的相对量为含岩藻糖的结构相对于所有糖结构的百分比。这些可以通过多种方法进行表征和定量,例如:1)使用经n-糖苷酶f处理的样品(例如,复合物、杂合物,以及低聚甘露糖结构和高甘露糖结构)的maldi-tof,如国际专利公布号wo2008/077546中所述;2)通过酶促释放asn297聚糖,随后衍生化并通过具有荧光检测的hplc(uplc)和/或hplc-ms(uplc-ms)进行检测/定量;3)在用或不用endo s或其它酶处理asn297聚糖的情况下,对天然或还原后的mab进行完整蛋白分析,所述其它酶在第一glcnac单糖和第二glcnac单糖之间进行裂解,留下连接至第一
glcnac的岩藻糖;4)通过酶消化(例如,胰蛋白酶或肽链内切酶lys-c)将mab消化为成分肽,随后通过hplc-ms(uplc-ms)进行分离、检测和定量;5)通过用pngase f在asn 297处进行特异性酶促去糖基化将mab低聚糖与mab蛋白质分离。这样释放的低聚糖可以通过各种互补技术用荧光团标记、分离和鉴定,这些技术允许:采用基质辅助激光解吸电离(maldi)质谱法,通过比较实验质量与理论质量来对聚糖结构进行精细表征;通过离子交换hplc(glycosep c)确定唾液酸化程度;通过正相hplc(glycosep n),根据亲水性标准分离和定量低聚糖形式;以及通过高效毛细管电泳激光诱导荧光(hpce-lif)分离和定量低聚糖。
[1307]“低岩藻糖”或“低岩藻糖含量”是指岩藻糖含量约介于1%至15%之间的包含ig恒定区或ig恒定区的片段的本公开的多特异性分子或异源分子。
[1308]“正常岩藻糖”或“正常岩藻糖含量”是指岩藻糖含量约超过50%,通常约超过80%或超过85%的包含ig恒定区或ig恒定区的片段的本公开的多特异性分子或异源分子。
[1309]
5.5抗独特型抗体
[1310]
抗独特型抗体是与本公开的spfv特异性结合的抗体。
[1311]
本发明还提供了与本公开的spfv特异性结合的抗独特型抗体。
[1312]
在一些实施方案中,该抗独特型抗体与本公开的spfv中的二硫键结合。
[1313]
在一些实施方案中,该抗独特型抗体与本公开的spfv的抗原结合结构域结合。
[1314]
5.6多核苷酸、载体、宿主细胞
[1315]
本公开还提供了一种编码本公开的spfv的分离的多核苷酸。
[1316]
本公开还提供了一种包含本公开的多核苷酸的载体。
[1317]
在一些实施方案中,载体是表达载体。表达载体可以是质粒载体、病毒载体、用于杆状病毒表达的载体、用于原核表达的载体、用于真核表达的载体、基于转座子的载体,或者适于将本公开的多核苷酸引入给定的细胞或生物体中的任何其他载体。编码本公开的spfv的多核苷酸可以可操作地连接到表达载体中促进spfv表达的控制序列。此类调节元件可包含转录启动子、编码合适的mrna核糖体结合位点的序列以及控制转录和翻译的终止的序列。表达载体还可以包括一个或多个非转录元件,诸如复制起点、其他5’或3’侧接非转录序列、5’或3’非翻译序列(诸如必要的核糖体结合位点)、剪接供体和受体位点,或者选择标志物。多核苷酸可为cdna。驱动spfv表达的启动子可以是强启动子、弱启动子、组织特异性启动子、诱导型启动子或发育特异性启动子。可以使用的示例性启动子为次黄嘌呤磷酸核糖基转移酶(hprt)、腺苷脱氨酶、丙酮酸激酶、β-肌动蛋白、人肌球蛋白、人血红蛋白、人肌肉肌酸,及其他。此外,许多病毒启动子在真核细胞中组成性地发挥功能,并适合与所述实施方案一起使用。此类病毒启动子包括巨细胞病毒(cmv)立即早期启动子、sv40的早期启动子和晚期启动子、小鼠乳腺肿瘤病毒(mmtv)启动子、莫洛尼白血病病毒的长末端重复序列(ltr)、人免疫缺陷病毒(hiv)、eb病毒(ebv)、劳氏肉瘤病毒(rsv)和其他逆转录病毒,以及单纯疱疹病毒的胸苷激酶启动子。诱导型启动子诸如金属硫蛋白启动子、四环素诱导型启动子、强力霉素诱导型启动子、含有一个或多个干扰素刺激应答元件(isre)的启动子,诸如蛋白激酶r 2’,5
’‑
寡腺苷酸合成酶、mx基因和adar1。本公开的载体还可以含有一个或多个内部核糖体进入位点(ires)。ires序列包含在融合载体中可能有利于增强一些蛋白质的表达。本公开的载体可以是圆形的或线性的。可将它们制备成包含在原核或真核宿主细胞中起作用的复制系统。复制系统可以衍生自例如cole1、sv40、2μ质粒、λ、牛乳头状瘤病毒,等
等。可以将这些表达载体设计用于瞬时表达、用于稳定表达,或用于这两者。可以将这些表达载体制备用于组成型表达或用于诱导型表达。
[1318]
可使用的示例性载体为细菌:pbs、phagescript、psix174、pbluescript sk、pbs ks、pnh8a、pnh16a、pnh18a、pnh46a(stratagene,la jolla,calif.,usa);ptrc99a、pkk223-3、pkk233-3、pdr540和prit5(pharmacia,uppsala,sweden)。真核:pwlneo、psv2cat、pog44、pxr1、psg(stratagene)、psvk3、pbpv、pmsg和psvl(pharmacia)、pee6.4(lonza)和pee12.4(lonza)。附加载体包括puc系列(fermentas life sciences,glen burnie,md.)、pbluescript系列(stratagene,lajolla,calif.)、pet系列(novagen,madison,wis.)、pgex系列(pharmacia biotech,uppsala,sweden)和pex系列(clontech,palo alto,calif.)。可以使用噬菌体载体,诸如λgt10、λgt11、λembl4和λnm1149、λzapii(stratagene)。示例性植物表达载体包括pbi01、pbi01.2、pbi121、pbi101.3和pbin19(clontech)。示例性动物表达载体包括peuk-cl、pmam和pmamneo(clontech)。该表达载体可以是病毒载体,例如逆转录病毒载体,例如γ逆转录病毒载体。
[1319]
本公开还提供了一种包含本公开的载体的宿主细胞。
[1320]“宿主细胞”是指已引入载体的细胞。应该理解,术语宿主细胞不仅旨在指特定的主体细胞,还指此类细胞的子代,并且也指特定主体细胞所产生的稳定细胞系。因为由于突变或者环境影响,在后代中可能发生某些修饰,所以此类子代可能与亲代细胞不同,但是仍包括在如本文所用的术语“宿主细胞”的范围内。此类宿主细胞可以是真核细胞、原核细胞、植物细胞或古菌细胞。原核宿主细胞的示例是大肠杆菌(escherichia coli)、杆菌属(bacilli)诸如枯草芽孢杆菌(bacillus subtilis)和其它肠杆菌科(enterobacteriaceae)诸如沙门氏菌(salmonella)、沙雷氏菌(serratia)以及各种假单胞菌属(pseudomonas)物种。其他微生物诸如酵母也可用于表达。酵母属(saccharomyce)(例如,酿酒酵母(s.cerevisiae))和毕赤酵母属(pichia)是合适的酵母宿主细胞的示例。示例性真核细胞可以是哺乳动物、昆虫、禽类或其它动物来源。哺乳动物真核细胞包括无限增殖化细胞系,诸如杂交瘤或骨髓瘤细胞系,诸如sp2/0(美国典型培养物保藏中心(atcc),manassas,va,crl-1581)、ns0(欧洲细胞培养物保藏中心(ecacc),salisbury,wiltshire,uk,ecacc no.85110503)、fo(atcc crl-1646)和ag653(atcc crl-1580)鼠细胞系。一种示例性人骨髓瘤细胞系是u266(attc crl-tib-196)。其它可用的细胞系包括衍生自中国仓鼠卵巢(cho)细胞的那些细胞系,诸如cho-k1sv(lonza biologics,walkersville,md)、cho-k1(atcc crl-61)或dg44。
[1321]
本公开还提供了一种产生本公开的spfv的方法,包括在产生该spfv的条件下培养本公开的宿主细胞,以及回收由该宿主细胞产生的spfv。制备scfv和纯化这些scfv的方法是已知的。一旦被合成(以化学方式或重组方式),就可以根据标准程序纯化本公开的spfv,所述标准程序包括硫酸铵沉淀、亲和柱、柱层析法、高效液相色谱法(hplc)纯化、凝胶电泳等等(一般参见scopes,protein purification(springer-verlag,n.y.,(1982))。本公开的scfv可以是基本上纯的,例如,至少约80%至85%纯、至少约85%至90%纯、至少约90%至95%纯、或者至少约98%至99%纯,或者更加纯,例如,不含污染物(诸如细胞碎片、除主题蛋白质之外的大分子,等等)。
[1322]
可以使用标准分子生物学方法将编码本公开的scfv的多核苷酸结合到载体中。使
cys之间形成二硫键,以制备所述稳定化的scfv。
[1343]
二硫键通常在scfv的表达期间形成。
[1344]
可以将形成抗原结合结构域的任何已知的scfv vh/vl对工程化到本公开的稳定化的scfv中。替代性地,可以使用已知方法从头鉴定感兴趣的抗原结合vh/vl对,并且可以将所得的vh/vl对工程化为spfv形式。
[1345]
例如,kohler和milstein的杂交瘤方法可以用于鉴定结合感兴趣抗原的vh/vl对,并且可以将所得的vh/vl对工程化为spfv。替代性地,在其基因组中携带人免疫球蛋白(ig)基因座的转基因动物(诸如小鼠、大鼠或鸡)可以用于生成抗原结合片段,并且在例如美国专利号6,150,584、国际专利公布号wo1999/45962、国际专利公布号wo2002/066630、wo2002/43478、wo2002/043478和wo1990/04036中有所描述。可破坏此类动物中的内源性免疫球蛋白基因座或使该基因座缺失,并且可使用转染色体或微小基因,通过同源或非同源重组将至少一种完整或部分的人免疫球蛋白基因座插入动物的基因组中。可以邀请诸如regeneron(http://_www_regeneron_com)、harbour antibodies(http://_www_harbourantibodies_com)、open monoclonal technology,inc.(omt)(http://_www_omtinc_net)、kymab(http://_www_kymab_com)、trianni(http://_www_trianni_com)和ablexis(http://_www_ablexis_com)等公司使用如上所述的技术来提供针对选定抗原的人抗体。噬菌体展示也可以用于产生可以被工程化为spfv的抗原结合片段。
[1346]
在一些实施方案中,本公开的spfv是人源化的。在一些实施方案中,本公开的spfv是人的。在一些实施方案中,本公开的spfv是非人的。
[1347]
在一些实施方案中,在稳定化的scfv中,vh cys与vl cys之间的距离为约至约
[1348]
在一些实施方案中,vh cys在h3、h5、h40、h43、h46或h105处,其中残基根据chothia进行编号。
[1349]
在一些实施方案中,vl cys在l3、l5、l39、l42、l45、l100或l102处,其中残基根据chothia进行编号。
[1350]
在一些实施方案中,
[1351]
vh cys在h105处并且vl cys在l42处;
[1352]
vh cys在h43处并且vl cys在l100处;
[1353]
vh cys在h3处并且vl cys在l3处;
[1354]
vh cys在h3处并且vl cys在l5处;
[1355]
vh cys在h3处并且vl cys在l39处;
[1356]
vh cys在h3处并且vl cys在l42处;
[1357]
vh cys在h3处并且vl cys在l45处;
[1358]
vh cys在h3处并且vl cys在l100处;
[1359]
vh cys在h3处并且vl cys在l102处;
[1360]
vh cys在h5处并且vl cys在l3处;
[1361]
vh cys在h5处并且vl cys在l5处;
[1362]
vh cys在h5处并且vl cys在l39处;
[1363]
vh cys在h5处并且vl cys在l42处;
[1364]
vh cys在h5处并且vl cys在l45处;
[1365]
vh cys在h5处并且vl cys在l100处;
[1366]
vh cys在h5处并且vl cys在l102处;
[1367]
vh cys在h40处并且vl cys在l3处;
[1368]
vh cys在h40处并且vl cys在l5处;
[1369]
vh cys在h40处并且vl cys在l39处;
[1370]
vh cys在h40处并且vl cys在l42处;
[1371]
vh cys在h40处并且vl cys在l45处;
[1372]
vh cys在h40处并且vl cys在l100处;
[1373]
vh cys在h40处并且vl cys在l102处;
[1374]
vh cys在h43处并且vl cys在l3处;
[1375]
vh cys在h43处并且vl cys在l5处;
[1376]
vh cys在h43处并且vl cys在l39处;
[1377]
vh cys在h43处并且vl cys在l42处;
[1378]
vh cys在h43处并且vl cys在l45处;
[1379]
vh cys在h43处并且vl cys在l102处;
[1380]
vh cys在h46处并且vl cys在l3处;
[1381]
vh cys在h46处并且vl cys在l5处;
[1382]
vh cys在h46处并且vl cys在l39处;
[1383]
vh cys在h46处并且vl cys在l42处;
[1384]
vh cys在h46处并且vl cys在l45处;
[1385]
vh cys在h46处并且vl cys在l100处;
[1386]
vh cys在h46处并且vl cys在l102处;
[1387]
vh cys在h105处并且vl cys在l3处;
[1388]
vh cys在h105处并且vl cys在l5处;
[1389]
vh cys在h105处并且vl cys在l39处;
[1390]
vh cys在h105处并且vl cys在l45处;
[1391]
vh cys在h105处并且vl cys在l100处;或者
[1392]
vh cys在h105处并且vl cys在l102处,其中残基根据chothia进行编号。
[1393]
在一些实施方案中,该l包含衍生自免疫球蛋白(ig)铰链区的连续氨基酸序列。
[1394]
在一些实施方案中,ig铰链区衍生自人ig铰链区或非人ig铰链区。
[1395]
在一些实施方案中,ig铰链区衍生自人ig铰链区。
[1396]
在一些实施方案中,人ig铰链区为igg1、igg2、igg3或igg4同种型。
[1397]
在一些实施方案中,该l包含氨基酸序列c(x)yc(seq id no:23),其中x是gly、ser、pro、ala、arg、asn、asp、glu、gln、his、ile、leu、lys、phe、thr、trp或tyr,并且y是1至3的整数。
[1398]
在一些实施方案中,该l包含氨基酸序列c(x)yc(seq id no:24),其中x是gly、ser或pro,并且y是1至3的整数。
[1399]
在一些实施方案中,该l包含氨基酸序列cpc、cgc、csc、cppc(seq id no:1)、cgpc(seq id no:28)、cpgc(seq id no:29)、cggc(seq id no:30)、cspg(seq id no:31)、cpsc(seq id no:32)、cssc(seq id no:33)、cgsc(seq id no:34)、csgc(seq id no:35)、cpppc(seq id no:36)、cgppc(seq id no:37)、cpgpc(seq id no:38)、cppgc(seq id no:39)、cggpc(seq id no:40)、cpggc(seq id no:41)、cgggc(seq id no:42)、csppc(seq id no:43)、cpspc(seq id no:44)、cppsc(seq id no:45)、csspc(seq id no:46)、cpssc(seq id no:47)、csssc(seq id no:48)、cgspc(seq id no:49)、cpgsc(seq id no:50)、csgpc(seq id no:51)或cpsgc(seq id no:52)。
[1400]
在一些实施方案中,该l包含约14个至约19个氨基酸,诸如约14个、约15个、约16个、约17个、约18个或约19个氨基酸。
[1401]
在一些实施方案中,l包含氨基酸序列(x)mc(x)yc(x)n(seq id no:25);其中x是gly、ser、pro、ala、arg、asn、asp、glu、gln、his、ile、leu、lys、phe、thr、trp或tyr,m是6至9的整数,y是1至3的整数,并且n是4至6的整数。
[1402]
在一些实施方案中,该l包含氨基酸序列(x)mc(x)yc(x)n(seq id no:26);其中x是gly、ser、pro、ala、arg、asn、asp、glu、gln、his、ile、leu、lys、thr或tyr,m是6至9的整数,y是1至3的整数,并且n是4至6的整数。
[1403]
在一些实施方案中,该l包含氨基酸序列(x)mc(x)yc(x)n(seq id no:27);其中x是gly或pro,m是6至9的整数,y是1至3的整数,并且n是4至6的整数。
[1404]
在一些实施方案中,该l包含氨基酸序列seq id no:2、3、4、5、6或7。
[1405]
在一些实施方案中,本公开的稳定化的spfv处于vl-l-vh取向。
[1406]
在一些实施方案中,本公开的稳定化的spfv处于vh-l-vl取向。
[1407]
在一些实施方案中,
[1408]
该vh包含在h105处的cys;
[1409]
该vl包含在l42处的cys;
[1410]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1411]
该scfv处于vl-l-vh取向。
[1412]
在一些实施方案中,
[1413]
该vh包含在h105处的cys;
[1414]
该vl包含在l45处的cys;
[1415]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1416]
该scfv处于vl-l-vh取向。
[1417]
在一些实施方案中,
[1418]
该vh包含在h105处的cys;
[1419]
该vl包含在l39处的cys;
[1420]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1421]
该scfv处于vl-l-vh取向。
[1422]
在一些实施方案中,
[1423]
该vh包含在h5处的cys;
[1424]
该vl包含在l42处的cys;
[1425]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1426]
该scfv处于vl-l-vh取向。
[1427]
在一些实施方案中,
[1428]
该vh包含在h5处的cys;
[1429]
该vl包含在l45处的cys;
[1430]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1431]
该scfv处于vl-l-vh取向。
[1432]
在一些实施方案中,
[1433]
该vh包含在h5处的cys;
[1434]
该vl包含在l39处的cys;
[1435]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1436]
该scfv处于vl-l-vh取向。
[1437]
在一些实施方案中,
[1438]
该vh包含在h3处的cys;
[1439]
该vl包含在l42处的cys;
[1440]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1441]
该scfv处于vl-l-vh取向。
[1442]
在一些实施方案中,
[1443]
该vh包含在h3处的cys;
[1444]
该vl包含在l45处的cys;
[1445]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1446]
该scfv处于vl-l-vh取向。
[1447]
在一些实施方案中,
[1448]
该vh包含在h3处的cys;
[1449]
该vl包含在l39处的cys;
[1450]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1451]
该scfv处于vl-l-vh取向。
[1452]
在一些实施方案中,
[1453]
该vh包含在h43处的cys;
[1454]
该vl包含在l100处的cys;
[1455]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1456]
该scfv处于vh-l-vl取向。
[1457]
在一些实施方案中,
[1458]
该vh包含在h43处的cys;
[1459]
该vl包含在l102处的cys;
[1460]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1461]
该scfv处于vh-l-vl取向。
[1462]
在一些实施方案中,
[1463]
该vh包含在h43处的cys;
[1464]
该vl包含在l5处的cys;
[1465]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1466]
该scfv处于vh-l-vl取向。
[1467]
在一些实施方案中,
[1468]
该vh包含在h43处的cys;
[1469]
该vl包含在l3处的cys;
[1470]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1471]
该scfv处于vh-l-vl取向。
[1472]
在一些实施方案中,
[1473]
该vh包含在h40处的cys;
[1474]
该vl包含在l100处的cys;
[1475]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1476]
该scfv处于vh-l-vl取向。
[1477]
在一些实施方案中,
[1478]
该vh包含在h40处的cys;
[1479]
该vl包含在l102处的cys;
[1480]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1481]
该scfv处于vh-l-vl取向。
[1482]
在一些实施方案中,
[1483]
该vh包含在h40处的cys;
[1484]
该vl包含在l5处的cys;
[1485]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1486]
该scfv处于vh-l-vl取向。
[1487]
在一些实施方案中,
[1488]
该vh包含在h40处的cys;
[1489]
该vl包含在l3处的cys;
[1490]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1491]
该scfv处于vh-l-vl取向。
[1492]
在一些实施方案中,
[1493]
该vh包含在h46处的cys;
[1494]
该vl包含在l100处的cys;
[1495]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1496]
该scfv处于vh-l-vl取向。
[1497]
在一些实施方案中,
[1498]
该vh包含在h46处的cys;
[1499]
该vl包含在l102处的cys;
[1500]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1501]
该scfv处于vh-l-vl取向。
[1502]
在一些实施方案中,
[1503]
该vh包含在h46处的cys;
[1504]
该vl包含在l5处的cys;
[1505]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1506]
该scfv处于vh-l-vl取向。
[1507]
在一些实施方案中,
[1508]
该vh包含在h46处的cys;
[1509]
该vl包含在l3处的cys;
[1510]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;并且
[1511]
该scfv处于vh-l-vl取向。
[1512]
在一些实施方案中,该l包含氨基酸序列seq id no:3。
[1513]
在一些实施方案中,该l包含氨基酸序列seq id no:6。
[1514]
在一些实施方案中,该l包含氨基酸序列seq id no:7。
[1515]
在一些实施方案中,当与缺少二硫键的亲本scfv相比时,本公开的稳定化的spfv以相当的亲和力结合抗原。
[1516]
本公开还提供了一种用于制备稳定化的scfv的方法,包括
[1517]
提供编码vh、l和vl的多核苷酸,其中
[1518]
该vh包含在h105处的cys,并且该vl包含在l42处的cys;
[1519]
该vh包含在h43处的cys,并且该vl包含在l100处的cys;
[1520]
该vh包含在h3处的cys,并且该vl包含在l3处的cys;
[1521]
该vh包含在h3处的cys,并且该vl包含在l5处的cys;
[1522]
该vh包含在h3处的cys,并且该vl包含在l39处的cys;
[1523]
该vh包含在h3处的cys,并且该vl包含在l42处的cys;
[1524]
该vh包含在h3处的cys,并且该vl包含在l45处的cys;
[1525]
该vh包含在h3处的cys,并且该vl包含在l100处的cys;
[1526]
该vh包含在h3处的cys,并且该vl包含在l102处的cys;
[1527]
该vh包含在h5处的cys,并且该vl包含在l3处的cys;
[1528]
该vh包含在h5处的cys,并且该vl包含在l5处的cys;
[1529]
该vh包含在h5处的cys,并且该vl包含在l39处的cys;
[1530]
该vh包含在h5处的cys,并且该vl包含在l42处的cys;
[1531]
该vh包含在h5处的cys,并且该vl包含在l45处的cys;
[1532]
该vh包含在h5处的cys,并且该vl包含在l100处的cys;
[1533]
该vh包含在h5处的cys,并且该vl包含在l102处的cys;
[1534]
该vh包含在h40处的cys,并且该vl包含在l3处的cys;
[1535]
该vh包含在h40处的cys,并且该vl包含在l5处的cys;
[1536]
该vh包含在h40处的cys,并且该vl包含在l39处的cys;
[1537]
该vh包含在h40处的cys,并且该vl包含在l42处的cys;
[1538]
该vh包含在h40处的cys,并且该vl包含在l45处的cys;
[1539]
该vh包含在h40处的cys,并且该vl包含在l100处的cys;
[1540]
该vh包含在h40处的cys,并且该vl包含在l102处的cys;
[1541]
该vh包含在h43处的cys,并且该vl包含在l3处的cys;
[1542]
该vh包含在h43处的cys,并且该vl包含在l5处的cys;
[1543]
该vh包含在h43处的cys,并且该vl包含在l39处的cys;
[1544]
该vh包含在h43处的cys,并且该vl包含在l42处的cys;
[1545]
该vh包含在h43处的cys,并且该vl包含在l45处的cys;
[1546]
该vh包含在h43处的cys,并且该vl包含在l102处的cys;
[1547]
该vh包含在h46处的cys,并且该vl包含在l3处的cys;
[1548]
该vh包含在h46处的cys,并且该vl包含在l5处的cys;
[1549]
该vh包含在h46处的cys,并且该vl包含在l39处的cys;
[1550]
该vh包含在h46处的cys,并且该vl包含在l42处的cys;
[1551]
该vh包含在h46处的cys,并且该vl包含在l45处的cys;
[1552]
该vh包含在h46处的cys,并且该vl包含在l100处的cys;
[1553]
该vh包含在h46处的cys,并且该vl包含在l102处的cys;
[1554]
该vh包含在h105处的cys,并且该vl包含在l3处的cys;
[1555]
该vh包含在h105处的cys,并且该vl包含在l5处的cys;
[1556]
该vh包含在h105处的cys,并且该vl包含在l39处的cys;
[1557]
该vh包含在h105处的cys,并且该vl包含在l45处的cys;
[1558]
该vh包含在h105处的cys,并且该vl包含在l100处的cys;或者
[1559]
该vh包含在h105处的cys,并且该vl包含在l102处的cys,其中残基根据chothia进行编号;
[1560]
该l包含氨基酸序列seq id no:2、3、4、5、6或7;以及
[1561]
在宿主细胞中表达所述多核苷酸,以产生所述稳定化的scfv。
[1562]
在一些实施方案中,宿主细胞是原核细胞。
[1563]
在一些实施方案中,宿主细胞是真核细胞。
[1564]
提供以下实施例以进一步描述本文所公开的实施方案中的一些。这些实施例旨在说明而非限制本发明所公开的实施方案。
[1565]
6.实施例
[1566]
6.1实施例1:稳定化scfv的设计
[1567]
单克隆抗体(mab)通过两个可变结构域vl和vh识别其靶抗原。首先由bird等人(1988)science 242:423-426(1988)将单链fv(scfv)设计为vl和vh与柔性接头的基因融合物,其处于vl-接头-vh取向或vh-接头-vl取向。柔性接头通常是甘氨酸-丝氨酸接头的三或四个重复,诸如(ggggs)n;n=1-4(seq id no:2、seq id no:53、seq id no:54、seq id no:55)。scfv重演了其亲本mab的抗原结合特异性,并且在很大程度上重演了其亲本mab的亲和力。这些scfv分子已广泛用作检测/诊断试剂或者用作用于制备更复杂的分子(诸如双特异性治疗剂、多特异性治疗剂)的砌块(brinkmann和kontermann(2017)mabs 9:182-212)或用于car-t疗法中(gross等人,(1989),transplant proc 21(第1期,第1部分):127-130;porter等人,(2011)j cancer 2:331-332;porter等人,(2011)n engl j med 365:725-733)。
[1568]
scfv分子的挑战之一是低稳定性和聚集趋势(综述于下列文献中:worn和
pluckthun(2001)j mol biol 305:989-1010;rothlisberger等人,(2005)j mol biol 347:773-789)。已尝试了许多策略来改善其特性(arnd等人,(2001)j mol biol 312:221-228;monsellier等人,(2006)j mol biol 362:580-593;zhao等人,(2010)int j mol sci 12:1-11;perchiacca和tessier(2012)annu rev chem biomol eng 3:263-286;asial等人,(2013)nat commmun 4:2901;gil和schrum(2013)adv biosci biteccchnol 4:73-84;tiller和tessier(2015)annu rev biomed eng 17:191-216)。这些策略包括在vl/vh结构域之间引入二硫键、改善vl/vh结构域稳定性和/或界面相互作用、使用不同的实验方法、使用附加的二聚化基序,等等。关键的难点是这些策略中的大多数通常对vh/vl对具有特异性,因此不能容易地转移到其他vh/vl对。有时,工程化可能对vl/vh结构和scfv特性具有负面影响。最近,zhang等人在抗黄曲霉毒素b1scfv(h4)的vh位置44与vl位置100之间引入了二硫化物并且成功地实现该scfv的显著稳定化,同时保持其结合亲和力(zhao等人,(2010)int j mol sci 12:1-11)。然而,由于所选择的两个位置之间的距离和角度约束,vl/vh间二硫化物如果被应用于其他vl/vh对,则可能限制/扭曲这两个结构域之间的相对取向,而该相对取向通常是结合所需的。
[1569]
fab片段的重链与轻链之间的界面包括vh/vl相互作用和ch1/cl相互作用。这两组独立的相互作用提供协同稳定化作用。此外,v/c结合部还有助于一些稳定化效果。相比之下,在scfv中,vh/vl界面仅由vh/vl相互作用维持。该接头被设计成柔性和非限制性的,除了在长度被设计成如此短以促进scfv间相互作用以形成二聚体和低聚体的情况下,仅将两者松散地偶联在一起。已知该接头的长度和性质在足够长时对scfv的稳定性贡献很小。
[1570]
6.1.1“装订”设计
[1571]
该工作的目的是通过抑制但不负面影响形成scfv的vh与vl之间的相对运动,来设计和产生稳定化的scfv。这是通过在vh与接头之间以及vl与接头之间工程化二硫键来稳定scfv而实现的。当适当定位时,约束(即,二硫键)将起到由上文讨论的ch1/cl和v/c相互作用所提供的协同效应的作用。为此,鉴定了两个结构保守的表面暴露的框架位置(锚定点),vh和vl上各一个,它们与典型的预测抗原结合位点不重叠,并且被突变为半胱氨酸(cys)残基。随后在柔性接头中选择两个位置用于cys位置。当以促进接头cys与每个锚定点之间形成二硫键的方式设计接头cys残基之间的距离和位置时,与不存在二硫键时的拴系相比,vh和vl将更紧密地拴系在一起。该方案在图1中用含有cppc序列(seq id no:1)的示例性接头来描绘。在柔性接头与锚定点之间形成二硫键的概念在本文中被称为“装订”。所得的“装订”scfv分子在本文中被称为spfv(“装订fv”)。
[1572]
6.1.2选择锚定点,设计钉序列和接头
[1573]
对于广泛适用的装订方案,重要的是锚定点在结构上是保守的、暴露在vl和vh两者的表面上,并且其突变为cys残基不会影响vl和vh的折叠或者与抗原的结合。锚定点与vl结构域和vh结构域的n末端和c末端的距离和几何结构也是正确形成二硫化物的重要考虑因素。
[1574]
分别针对处于vl-接头-vh和vh-接头-vl取向的spfv选择锚定点。对于vl-接头-vh取向,选择vl中的chothia位置42和vh中的chothia位置105作为锚定点。图2示出了在人种系抗体(pdb id 5i19,下文的glk1)的fv内,处于vl-接头-vh取向的spfv的所选择锚定点的图解说明。在glk1中,vl chothia位置42是赖氨酸(k),并且vh chothia位置105是谷氨酰胺
(q)。对于vh-接头-vl取向,选择vl中的chothia位置100和vh中的chothia位置43作为锚定点。图2示出了在人种系抗体(pdb id 5i19,glk1)的fv内,处于vh-接头-vl取向的spfv的所选择锚定点的图解说明。在glk1中,vl chothia位置100是谷氨酰胺(q),并且vh chothia位置43是赖氨酸(k)。所选择的锚定点在结构上是保守的,并且几何结构在含有κ或λ轻链的抗体中非常相似。锚定点对之间的距离在从大约(对于vl-接头-vh取向)至大约(对于vh-接头-vl取向)的范围内。
[1575]
嵌入连接vh和vl的接头内的钉序列被设计成与spfv中的锚定点之间的距离具有相似的长度。作为钉序列的初始示例,cppc(seq id no:1)被选择作为可能的钉序列,部分原因是该序列天然地存在于人igg1铰链以及一些啮齿动物igg中。人和小鼠igg分子的铰链的结构证明,小鼠igg铰链(图4)和人igg(图5)中的cβ(cys1)-cβ(cys2)距离在约至的范围内。由于该范围与处于vl-接头-vh和vh-接头-vl这两种取向的两个锚定点之间的距离非常相似,所以cppc(seq id no:1)钉序列具有提供用于高效且正确地装订(即,形成适当的二硫键)到锚定点的正确几何结构的潜力。一般来讲,该钉序列被设计成具有两个cys残基。为了正确装订,该钉序列的n末端cys与spfv n末端结构域锚定点形成二硫键,并且该钉序列的c末端cys与spfv c末端结构域锚定点形成二硫键。
[1576]
因此,将连接vh和vl的接头以及将序列的n末端和c末端两者连接到钉序列的接头设计成包含钉序列,以延伸接头,从而提供足够大的接头长度,以允许vh和vl的链内折叠,并且以促进钉序列的正确定位。
[1577]
在vl-接头-vh设计中,vl锚定点(k42)、vh锚定点(q105)、vl的c末端(k107)和vh的n末端(q1)之间的距离在图2中示出。在vh-接头-vl设计中,vl锚定点(q100)、vh锚定点(k43)、vh的c末端(s114)和vl的n末端(d1)之间的距离在图3中示出。建模表明,这些距离可以由约14至19个残基的接头长度跨越,其中4个残基的钉序列侧接约6至9个残基的n末端接头延伸和约4至6个残基的c末端接头延伸。设计的接头长度因此可以表示为n 4 m,其中n=6-9个残基并且m=4-6个残基,4指示cppc(seq id no:1)钉序列的长度。n个残基和m个残基可以是甘氨酸或丝氨酸,或者其他氨基酸残基。预期这些接头长度足够长且柔性足够大以允许装订,但是又短到不允许加扰。
[1578]
6.2实施例2:产生并表征spfv
[1579]
为了评价装订设计,选择了三种人抗体来产生scfv和对应的spfv:来自合成噬菌体抗体文库(shi等人,(2010)j mol biol 397:385-396)的具有κ轻链的两种抗体(glk1和glk2),以及获自出版物(gerhardt等人,(2009)j mol biol 394:905-921)的含λ抗体(cat2200)。对于cat2200,在亲本vh中引入t28g突变以产生变体(cat2200a),以降低其与靶标il-17的一些相互作用。此外,将s42q突变(chothia)工程化到亲本cat2200 vl中并且与t28g vh配对以产生cat2200b。glk1、glk2、cat2200a和cat2200b的vl结构域和vh结构域的氨基酸序列分别在图6和图7中示出。vh结构域氨基酸序列在bat2200a与cat2200b之间是相同的。glk1vh最接近人ighv2-23*01,glk2vh最接近人ighv5-51,cat2200vh最接近人ighv2-23*01。glk1vl最接近人igkv1-39*01,glk2vl最接近人igkv3-20*01,并且cat2200vl最接近人iglv6-57*01。
[1580]
产生所有的scfv分子和spfv分子,并且以vl-接头-vh和vh-接头-vl这两种取向表
达。对于scfv构建体,使用标准(ggggs)4(seq id no;2)接头。对于spfv,使用上述n和m范围内的不同接头长度。对于glk1 spfv,将9-4-5接头用于两种取向。对于glk2 spfv,将9-4-5接头长度和6-4-6接头长度分别用于vl-vh取向和vh-vl取向。对于cat2200a spfv,vl-vh分子分别用8-4-4接头和9-4-4接头制成,并且cat2200b spfv vh-vl用9-4-4接头制成。表4示出了所产生的分子及其接头序列。表5示出了氨基酸序列或产生的分子。
[1581]
表4.
[1582][1583]
表5.
[1584]
[1585]
[1586][1587]
将除cat2200a scfv vl-vh之外的所有scfv和spfv分子克隆到cmv启动子驱动的哺乳动物表达载体中。使用制造商的方案将这些构建体转染到expi293细胞中,并且将细胞培养5天。在1ml his-trap hp柱(ge healthcare)上经由aktaxpress系统(ge healthcare)从澄清的上清液中纯化每种蛋白质。用0%至100%洗脱缓冲液(洗涤缓冲液:50mm tris,ph 7.5,500mm nacl,20mm咪唑;洗脱缓冲液:50mm tris,ph 7.5,500mm nacl,500mm咪唑)的梯度制备柱,以去除松散结合的镍,然后在dpbs中重新平衡。首先将澄清的上清液调节至50mm tris,ph 7.5和20mm咪唑,然后在4℃下以0.8ml/min上样到1ml histrap hp柱上。然后用pbs洗涤该柱,直到获得稳定基线。然后将该柱用20cv的洗涤缓冲液进一步洗涤,用洗脱缓冲液洗脱到单个进样环中,并且在1x dpbs中用26/10hiprep脱盐柱脱盐并收集级分。然后将含有经纯化蛋白质的级分汇集并浓缩。将glk2 scfv和spfv蛋白质透析到dpbs中以进行热稳定性测量(dsc和nanodsf),并且透析到25mm tris,ph 7.5和100mm nacl中用于其他研究。在25mm mes,ph 6.0和100mm nacl中透析其他的scfv蛋白质和spfv蛋白质。
[1588]
cat2200a scfv vl-vh购自供应商。dpbs,ph 7.2中的浓度为0.77mg/ml。il-17(具有k70q a132q c106s突变的12-132,下文简称为il-17(seq id no:22))的突变体购自accelagen(ca)。将蛋白质从大肠杆菌(e.coli)包含体遵循其专有的重折叠方案重折叠,并且在20mm nacl,20mm mes,ph 6.0中以1.50mg/ml提供。
[1589]
seqid no: 22(il-17a突变体)
[1590]
mnsedknfprtvmvnlnihnrntntnpkrssdyynrstspwnlhrnedperypsviweaqcrhlgcinadgnvdyhmnsvpiqqeilvlrrepphspnsfrlekilvsvgctcvtpivhhvq
[1591]
6.2.1产生的scfv和spfv分子的热稳定性
[1592]
通过差示量热法(dsc)研究scfv分子和spfv分子的热稳定性。将scfv蛋白质和spfv蛋白质针对glk1用1x dpbs(gibco)透析过夜,并且针对glk2用cat2200a/cat2200b或mes(25mm mes,ph 6.0,100mm nacl)透析过夜。然后0.22微米过滤透析缓冲液,用作参考溶液,并且用于dsc实验中的缓冲液-缓冲液空白。在过滤缓冲液中将蛋白质稀释至约0.5mg/ml,将400μl的每种蛋白质或缓冲液样品装载到96深孔板(microliter analytical supplies,07-2100)中,并且在实验过程中在自动进样器抽屉中保持在4℃。使用具有自动进样器的microcal毛细管dsc(malvern)进行dsc实验。dsc扫描在25℃至95℃下以60℃/h的扫描速率进行,没有对样品重新扫描。没有选择反馈,将过滤周期设定为15s。在每个样品之后,用10%contrad-70溶液清洗细胞,并且运行缓冲液-缓冲液空白。用microcal vp-毛细
管dsc自动分析附加装置(malvern),使用origin 7.0进行数据分析。手动选择基线范围和类型,然后减去。从样品曲线中减去先前的缓冲液空白,之后进行浓缩依赖性归一化。使用两状态转变和非两状态转变两者来分析热解链曲线。两状态拟合(一个转变)与实验曲线的一致性差。因此,通过手工进行非两状态拟合来计算两个转变(tm1和tm2)。tm数据报告于表6中。所有scfv蛋白质和spfv蛋白质的dsc曲线均表现出仅能够与非两状态转变匹配的偏度。因此,对于每个scfv或spfv,报告两个转变(tm1和tm2)(表6)。最可能的是,这两个转变分别对应于vl结构域和vh结构域的解链tm。一般来讲,在比较时,无论是对于tm1还是tm2,scfv与spfv之间的差异通过装订都大致增加了10℃,而与起始scfv的tm无关。只有一个例外,即,在约7℃下,glk2 scfv和spfv(vh-vl取向)差异的情况。这可能是由于较短的6 4 6接头可能已在装订几何结构中引起轻微的应变。δtm1(vl)和δtm2(vh)几乎相同的事实表明,除了增强vl/vh相互作用之外,装订还导致结构域自身的稳定化。替代性地,较强的vh/vl相互作用将稳定化效应传递到vl/vh结构域的稳定化中。概括地说,如本工作中所述的装订显著增加了scfv的稳定性。
[1593]
表6.
[1594]
[1595]
测试cat2200 spfv与il-17的结合。当与cat2200 scfv比较时,该结合是相当的。
[1596]
6.3实施例3:通过产生的scfv分子和spfv分子的结晶来验证正确的装订
[1597]
将蛋白质浓缩在它们各自的缓冲液中:glk1 spfv vl-vh,在25mm mes,ph 6.0,100mm nacl中浓缩至8.67mg/ml;glk1 spfv vh-vl,在25mm mes,ph 6.0,100mm nacl中浓缩至5mg/ml;glk2 spfv vh-vl,在25mm tris,ph 7.5,100mm nacl中浓缩至8.66mg/ml;cat2200b spfv vh-vl,在25mm mes,ph 6.0,100mm nacl中浓缩。使用mosquito机器人在corning 3550晶体托盘中以坐滴形式对每种蛋白质进行结晶。每个孔装有100nl蛋白质和100nl池液,并且在20℃下用70μl池液进行温育。池液是ih1和ih2定制条件,以及peg ion screen ht(hampton research)。一些初始条件通过在优化尝试中改变池液组分来精修。获得了一些scfv蛋白质和spfv蛋白质的衍射质量晶体。表7示出所使用条件的汇总。将晶体在补充有20%甘油的母液中浸泡数秒,并且在液氮中快速冷冻。在argonne national lab的imca-cat beamline 17id处收集x射线数据。
[1598]
表7.
[1599][1600]
6.3.1使cat2200a scfv vl-vh和cat2200a spfv vl-vh与il-17复合结晶
[1601]
产生il-17/cat2200a scfv vl-vh复合物,方式为:将333μl的il17(seq id no:22)(1.5mg/ml)与1.74ml的cat2200a scfv(0.69mg/ml)混合,并且在4℃下温育3小时。用10kda截止值的amicon ultra浓缩器将混合物浓缩至约400μl,并且上样到在250mm nacl,20mm hepes,ph 7.5中平衡的superdex75柱上。将对应于该复合物的级分合并,并且浓缩至150μl的体积。将样品稀释并浓缩4倍:添加350μl 50mm nacl,20mm hepes,ph 7.5,然后浓缩至刚好低于150μl。将体积调至约105μl,测得浓度为2.69mg/ml。使用mosquito结晶机器人以坐滴形式建立结晶,该机器人在corning3550板中具有150nl蛋白质 150nl池液并且用80μl池液(是一组室内预配制的缓冲液和沉淀剂条件)进行结晶。将板在20℃下温育。条件
之一(乙酸钠,ph 4.5,25%peg 3k,0.2m乙酸铵)产生非常小的晶体。收获这些晶体,在hampton seed bead管中,在100μl 27%peg 3350、200mm乙酸铵、100mm乙酸钠(ph 4.5)中使用hampton seed bead将其转变为结晶晶种。
[1602]
通过相同的程序获得衍射质量晶体,不同的是添加上述晶种:150nl蛋白质 100nl池液 50μl晶种。从0.1m tris 8.5、18%peg3k、0.2m liso4生长晶体,并且将其转移到合成母液(0.1m tris,ph 8.5,10%peg 3350,0.2m liso4和20%甘油)中,然后在液氮中快速冷冻。在argonne national laboratory的imca-cat id17处收集x射线衍射数据。
[1603]
通过将167μl il-17(250μg)与154μl mscw274(467μg,溶于250mm nacl、20mm mes(ph 6.5)中)混合产生il-17/cat2200a spfv vl-vh复合物,并且在4℃下温育过夜。将混合物在10kda分子量截止值(mwco)的amicon ultra 0.5ml浓缩器中浓缩至约100μl,然后重复稀释并浓缩5倍:浓缩至约150μl,并且添加350μl 50mm nacl,20mm hepes,ph 7.5。最终体积为100μl,并且复合物的浓度被确定为6.0mg/ml。使用mosquito机器人,以与就scfv/il-17复合物而言类似的方式以坐滴形式建立结晶。坐滴由150nl蛋白质 120nl池液 30nl晶种(上述的scfv/il-17)组成。池液是一组具有不同的peg 3350浓度和盐的条件。将结晶板在20℃下温育。从15.5%peg 3350、0.4m nah2po4获得小晶体。将晶体转移到16%peg 3350、0.2m nah2po4、20%甘油和快速冷冻的ln2中。在argonne national laboratory的imca-cat id17处收集x射线衍射数据。
[1604]
所有x射线衍射数据均用xds(kabsch等人,(2010)acta crystallogr d biol crystallogr 66(第2部分):125-132;monsellier和bedouelle(2006)j mol biol 362:580-593)和ccp4(collaborative computational project,n.(1994)acta crystallogr d biol crystallogr 53:240-255)处理。使用phaser,利用在moe(montreal,canada)中产生的同源性模型,通过分子置换(mr)求解所有晶体结构(read(2001)acta crystallogr d biol crystallogr 57(第10部分):1373-1382),不同的是对于scfv cat2200a scfv vl-vh/il-17复合物,将其结构pdb id 2vxs(gerhardt等人,(2009)j mol biol 394:905-921)用作搜索模型。在phenix中对结构模型进行了精修(adams等人,(2004)j synchrotron radiat 11(第1部分):53-55)并且在coot中进行手动调整(emsley等人,(2010)acta crystallogr d biol crystallogr 66(第4部分):486-501)。分子图形在pymol(www_schrodinger_com)中生成。
[1605]
6.3.2结构
[1606]
未结合的scfv分子和spfv分子的结构在图8、图9、图10和图11中示出。图8示出了glk1 spfv vl-vh的结构。图9示出了glk1 spfv vh-vl的结构。图10示出了glk2 spfv vh-vl的结构。图11示出了cat2200b spfv vh-vl的结构。这些结构与vl和vh这两种结构域彼此抵靠堆积的典型fv结构一致。一般来讲,大多数接头残基在电子密度图中是有序的并且得到解析。钉与锚定点之间的二硫键通常在两种vl-vh取向上均井然有序。除了未结合的scfv结构和spfv结构之外,我们还试图揭示对抗原结合的任何结构上的影响。将cat2200 scfv和spfv变体分子与其同源靶标il-17复合结晶。对于结晶的cat2200变体的scfv和spfv,结构在有和没有结合靶标的情况下几乎相同(图12,图13,图14)。图12示出了未结合的cat2200b spfv vh-vl与结合至il-17的cat2200a scfv vl-vh的比较。图13示出了未结合的cat2200b spfv vh-vl与结合至il-17的cat2200a spfv vl-vh的结构的前视图的比较。
图14示出了未结合的cat2200b spfv vh-vl与结合至il-17的cat2200a spfv vl-vh的结构的后视图的比较。无论钉是何取向或者存在与否,结构也是相同的。结构对之间所有匹配的cα原子的rmsd均非常小:在未结合的spfv-vh-vl与抗原结合的scfv-vl-vh之间为(图12),在未结合的spfv-vh-vl与spfv-vl-vh之间为(分别为图13和图14),并且在结合的scfv与结合的spfv之间为结构证据表明,装订如所设计的那样起作用。而且,装订不影响vl和vh的结构域结构或者相对的vl/vh填充。
[1607]
6.4实施例4:附加锚定点的设计
[1608]
使用实施例1中所述的方法来识别用于装订的任何附加锚定点。识别了以下锚定点:
[1609]
对于vl-接头-vh取向:vl chothia位置42、45和39,以及vh chothia位置105、5和3。在图6中,glk1vl上的vl残基为k42、k45、k39,并且glk1上的vh残基为q105、l5和q3。钉在所指示的任何位置之间形成。
[1610]
对于vh-接头-vl取向:vh chothia位置43、40和46,vl chothia位置102、5和3,钉在任何所述位置之间形成。
[1611]
克隆、表达具有本实施例中所述锚定点的spfv,并且使用本文和实施例2中所述的测定法来测试钉的形成和它们的热稳定性。
[1612]
6.5实施例5:半装订
[1613]
产生并且表达在vh与接头之间或vl与接头之间含有一个钉的构建体。使用本文所述的方法来表达、纯化和分析所产生的构建体。
[1614]
6.6实施例6:包含spfv的多特异性构建体
[1615]
在本实施例中构建并且测试了结合本文所提供的spfv结构的示例性多特异性结合分子。具体地讲,将衍生自表8中所示的靶结合序列的双特异性抗体和对照分子在无血清/无动物成分的培养基中的cho悬浮培养物中瞬时表达,并且通过蛋白质a亲和色谱法纯化,之后使用pure仪器(ge healthcare)在superdex 200 10/300gl柱(ge healthcare)上运用制备型尺寸排阻色谱法(sec)。重链含有杵臼(kih)突变以促进异源二聚化(ridgway等人,protein eng.9(7):617-21(1996);atwell等人,j.mol.biol.270(1):26-35(1997);merchant等人,nat.biotechnol.16(7):677-81(1998))。当与野生型igg1相比时,抗体含有包含一组七个fc突变(l234a、l235a、g237a、p238s、h268a、a330s和p331s)的igg1σ fc,以降低fc受体相互作用(tam等人,antibodies(2017))。
[1616]
产生具有igg1σ突变和kih突变的双特异性抗体。
[1617]
表8:用于实施例6的构建体的靶结合序列。
[1618][1619]
*:本文所用的edbmab1(wo9745544)是已经在临床上测试过的抗ed-b抗体,结合至ed-b或相邻结构域的其他抗体先前已经描述过(carnemolla等人,int.j.cancer 68:397-405(1996))
[1620]
表8中的序列如下:
[1621]
seq id no:69(vh bha10)
[1622]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgqglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvss
[1623]
seq id no:70(vl bha10)
[1624]
diqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgqgtkveik
[1625]
seq id no:71(vh l19)
[1626]
evqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvss
[1627]
seq id no:72(vl l19)
[1628]
eivltqspgtlslspgeratlscrasqsvsssflawyqqkpgqaprlliyyassratgipdrfsgsgsgtdftltisrlepedfavyycqqtgripptfgqgtkveik
[1629]
seq id no:73(vh b21m)
[1630]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvss
[1631]
seq id no:74(vl b21m)
[1632]
divmtqspdslavslgeratincrasqsvdyngisymhwyqqkpgqppklliyaasnpesgvpdrfsgsgsgtdftltisslqaedvavyycqqiiedpwtfgqgtkveik
[1633]
seq id no:75(vh mslnmab1)
[1634]
qvqlqqsgpelekpgasvkisckasgysftgytmnwvkqshgkslewiglitpyngassynqkfrgkatltvdkssstaymdllsltsedsavyfcarggydgrgfdywgsgtpvtvss
[1635]
seq id no:76(vl mslnmab1)
[1636]
dieltqspaimsaspgekvtmtcsasssvsymhwyqqksgtspkrwiydtsklasgvpgrfsgsgsgnsysltissveaeddatyycqqwskhpltfgsgtkveik
[1637]
通过在280nm(od280)处的吸光度测量确定蛋白质浓度,并且确定纯化产量。在thermo vanquish hplc系统上使用bio sec-5柱(agilent,5μm粒径,)进行分析性
sec。将10μl经纯化的蛋白质上样到柱上,并且通过od280记录洗脱。
[1638]
表9示出了本实施例中描述的双特异性抗体和对照分子的结构特性概述。以粗体显示的分子是根据本发明的示例性分子,而其他分子是不同方面的对照。表10示出了另一种比较用双特异性抗体的结构特性,该抗体靶向ltbr和间皮素(不存在于细胞外基质中的肿瘤相关抗原),如实施例9中所讨论。
[1639][1640]
表10:msln/ltbr双特异性抗体的结构特性概述
[1641][1642]
上文列出的具有2:1化学计量比的非对称抗体(都是igg1σ,都具有kih突变)如下产生:
[1643]
i.产生cova1484,方式为将承载n末端装订的scfv bha10(vh-vl取向seq id no:77)融合(seq id no:78,包含seq id no:79)的抗rsv b21m抗体重链与抗rsv b21m抗体(图15a)的重链(hc;seq id no:80)和轻链(lc;seq id no:81)共表达。
[1644]
seq id no:77[装订的scfv bha10(vh-vl)]
[1645]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1646]
seq id no:78(hc b21m n末端装订的bha10(vh-vl),igg1,杵,具有pa突变)
[1647]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveikggggsggggsggggsggggsggggsqitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1648]
seq id no:79(hc b21m(rsv)igg1杵,具有pa突变)
[1649]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqyn
styrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1650]
seq id no:80(hc b21m(rsv)igg1臼)
[1651]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1652]
seq id no:81[lc b21m(rsv)]
[1653]
divmtqspdslavslgeratincrasqsvdyngisymhwyqqkpgqppklliyaasnpesgvpdrfsgsgsgtdftltisslqaedvavyycqqiiedpwtfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1654]
ii.产生cova1485,方式为将承载n末端装订的scfv bha10(vl-vh取向seq id no:82)融合(seq id no:83,包含seq id no:79)的抗rsv b21m抗体重链与抗rsv b21m抗体(图15b)的重链(hc;seq id no:80)和轻链(lc;seq id no:81)共表达。
[1655]
seq id no:82[装订的scfv bha10(vl-vh)]
[1656]
diqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgcapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgqgtkveikggsggsggcppcgsggqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgqglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgcgttvtvss
[1657]
seq id no:83(hcb21m末端装订的bha10(vl-vh),igg1,杵,具有pa突变)
[1658]
diqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgcapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgqgtkveikggsggsggcppcgsggqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgqglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgcgttvtvssggggsggggsggggsggggsggggsqitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1659]
seq id no:79(hcb21m(rsv)igg1杵,具有pa突变)
[1660]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1661]
seq id no:80(hc b21m(rsv)igg1臼)
[1662]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1663]
seq id no:81[lc b21m(rsv)]
[1664]
divmtqspdslavslgeratincrasqsvdyngisymhwyqqkpgqppklliyaasnpesgvpdrfsgsgsgtdftltisslqaedvavyycqqiiedpwtfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1665]
iii.产生cova1486,方式为将承载c末端装订的scfv bha10(vh-vl取向seq id no:77)融合(seq id no:84,包含seq id no:79)的抗rsv b21m抗体重链与抗rsv b21m抗体(图15c)的重链(hc;seq id no:80)和轻链(lc;seq id no:81)共表达。
[1666]
seq id no:77[装订的scfv bha10(vh-vl)]
[1667]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1668]
seq id no:84(hc b21m c末端装订的bha(vh-vl),igg1,杵,具有pa突变)
[1669]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgkggggsggggsggggsqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1670]
seq id no:79(hc b21m(rsv)igg1杵,具有pa突变)
[1671]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1672]
seq id no:80(hc b21m(rsv)igg1臼)
[1673]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrl
titkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1674]
seq id no:81[lc b21m(rsv)]
[1675]
divmtqspdslavslgeratincrasqsvdyngisymhwyqqkpgqppklliyaasnpesgvpdrfsgsgsgtdftltisslqaedvavyycqqiiedpwtfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1676]
iv.产生cova1487,方式为将承载c末端装订的scfv bha10(vl-vh取向seq id no:82)融合(seq id no:85,包含seq id no:79)的抗rsv b21m抗体重链与抗rsv b21m抗体(图15d)的重链(hc;seq id no:80)和轻链(lc;seq id no:81)共表达。
[1677]
seq id no:82[装订的scfv bha10(vl-vh)]
[1678]
diqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgcapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgqgtkveikggsggsggcppcgsggqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgqglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgcgttvtvss
[1679]
seq id no:85(hc b21m c末端装订的bha(vl-vh),igg1,杵,具有pa突变)
[1680]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgkggggsggggsggggsdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgcapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgqgtkveikggsggsggcppcgsggqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgqglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgcgttvtvss
[1681]
seq id no:79(hcb21m(rsv)igg1杵,具有pa突变)
[1682]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1683]
seq id no:80(hc b21m(rsv)igg1臼)
[1684]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepksc
dkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1685]
seq id no:81[lc b21m(rsv)]
[1686]
divmtqspdslavslgeratincrasqsvdyngisymhwyqqkpgqppklliyaasnpesgvpdrfsgsgsgtdftltisslqaedvavyycqqiiedpwtfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1687]
v.产生cova1480,方式为将承载n末端装订的scfv bha10(vh-vl取向seq id no:77)融合(seq id no:86,包含seq id no:87)的抗edb抗体edbmab1重链与抗edb抗体edbmab1(图15e)的重链(hc;seq id no:88)和轻链(lc;seq id no:89)共表达。
[1688]
seq id no:77[装订的scfv bha10(vh-vl)]
[1689]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1690]
seq id no:86(hc l19 n末端装订的bha10(vh-vl),igg1,杵,具有pa突变)
[1691]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveikggggsggggsggggsggggsggggsevqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1692]
seq id no:87(hc l19 igg1杵,具有pa突变)
[1693]
vqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1694]
seq id no:88(hc l19 igg1臼)
[1695]
evqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdia
vewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1696]
seq id no:89(lc l19)
[1697]
eivltqspgtlslspgeratlscrasqsvsssflawyqqkpgqaprlliyyassratgipdrfsgsgsgtdftltisrlepedfavyycqqtgripptfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1698]
vi.产生cova1481,方式为将承载n末端装订的scfv bha10(vl-vh取向seq id no:82)融合(seq id no:90,包含seq id no:87)的抗edb抗体edbmab1重链与抗edb抗体edbmab1(图15f)的重链(hc;seq id no:88)和轻链(lc;seq id no:89)共表达。
[1699]
seq id no:82[装订的scfv bha10(vl-vh)]
[1700]
diqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgcapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgqgtkveikggsggsggcppcgsggqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgqglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgcgttvtvss
[1701]
seq id no:90(hc l19 n末端装订的bha10(vl-vh),igg1,杵,具有pa突变)
[1702]
diqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgcapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgqgtkveikggsggsggcppcgsggqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgqglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgcgttvtvssggggsggggsggggsggggsggggsevqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1703]
seq id no:87(hc l19 igg1杵,具有pa突变)
[1704]
vqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1705]
seq id no:88(hc l19 igg1臼)
[1706]
evqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1707]
seq id no:89(lc l19)
[1708]
eivltqspgtlslspgeratlscrasqsvsssflawyqqkpgqaprlliyyassratgipdrfsgsgsgtdftltisrlepedfavyycqqtgripptfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1709]
vii.产生cova1482,方式为将承载c末端装订的scfv bha10(vh-vl取向seq id no:77)融合(seq id no:91,包含seq id no:87)的抗edb抗体edbmab1重链与抗edb抗体edbmab1(图15g)的重链(hc;seq id no:88)和轻链(lc;seq id no:89)共表达。
[1710]
seq id no:77[装订的scfv bha10(vh-vl)]
[1711]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1712]
seq id no:91(hc l19 c末端装订的bha10(vh-vl),igg1,杵,具有pa突变)
[1713]
evqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgkggggsggggsggggsqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1714]
seq id no:87(hc l19 igg1杵,具有pa突变)
[1715]
vqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1716]
seq id no:88(hc l19 igg1臼)
[1717]
evqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1718]
seq id no:89(lc l19)
[1719]
eivltqspgtlslspgeratlscrasqsvsssflawyqqkpgqaprlliyyassratgipdrfsgsgsgtdftltisrlepedfavyycqqtgripptfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreak
y36f_s49y_f87y seq id no:93)融合(seq id no:94,包含seq id no:87)的抗edb抗体edbmab1重链与抗edb抗体edbmab1(图15i)的重链(hc;seq id no:88)和轻链(lc;seq id no:89)共表达。
[1732]
seq id no:93[装订的scfv(vl3_y36f_s49y_f87y)bha10(vh-vl)]
[1733]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawfqqkpgkapksliysasyrysgvpsrfsgsgsgtdftltisslqpedfatyycqqydtypftfgcgtkveik
[1734]
seq id no:94(hc l19 c末端装订的(vl3_y36f_s49y_f87y)bha(vh-vl),igg1,杵,具有pa突变)
[1735]
evqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgkggggsggggsggggsqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawfqqkpgkapksliysasyrysgvpsrfsgsgsgtdftltisslqpedfatyycqqydtypftfgcgtkveik
[1736]
seq id no:87(hc l19 igg1杵,具有pa突变)
[1737]
vqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1738]
seq id no:88(hc l19 igg1臼)
[1739]
evqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1740]
seq id no:89(lc l19)
[1741]
eivltqspgtlslspgeratlscrasqsvsssflawyqqkpgqaprlliyyassratgipdrfsgsgsgtdftltisrlepedfavyycqqtgripptfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1742]
x.产生cova14108,方式为将承载c末端装订的scfv bha10(vh-vl取向,vh_cdr1_
y33a seq id no:95)融合(seq id no:96,包含seq id no:87)的抗edb抗体edbmab1重链与抗edb抗体edbmab1(图15j)的重链(hc;seq id no:88)和轻链(lc;seq id no:89)共表达。
[1743]
seq id no:95[装订的scfv(vh_cdr1_y33a)bha10(vh-vl)]
[1744]
qvqlvqsgaevkkpgssvkvsckasgytfttyalhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1745]
seq id no:96(hc l19 c末端装订的(vh_cdr1_y33a)bha10(vh-vl),igg1,杵,具有pa突变)
[1746]
evqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgkggggsggggsggggsqvqlvqsgaevkkpgssvkvsckasgytfttyalhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1747]
seq id no:87(hc l19 igg1杵,具有pa突变)
[1748]
vqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1749]
seq id no:88(hc l19 igg1臼)
[1750]
evqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1751]
seq id no:89(lc l19)
[1752]
eivltqspgtlslspgeratlscrasqsvsssflawyqqkpgqaprlliyyassratgipdrfsgsgsgtdftltisrlepedfavyycqqtgripptfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1753]
xi.产生cova14133,方式为将承载c末端装订的scfv bha10(vh-vl取向seq id no:77)融合(seq id no:97,包含seq id no:98)的抗edb抗体edbmab1重链与抗edb抗体
edbmab1(图15k)的重链(hc;seq id no:88)和轻链(lc;seq id no:89)共表达。
[1754]
seq id no:77[装订的scfv bha10(vh-vl)]
[1755]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1756]
seq id no:97(hc l19 c末端装订的bha10(vh-vl),igg1,杵,无pa突变)
[1757]
evqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgkggggsggggsggggsqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1758]
seq id no:98(hc l19 igg1杵)
[1759]
evqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1760]
seq id no:88(hc l19 igg1臼)
[1761]
evqllesggglvqpggslrlscaasgftfssfsmswvrqapgkglewvssisgssgttyyadsvkgrftisrdnskntlylqmnslraedtavyycakpfpyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1762]
seq id no:89(lc l19)
[1763]
eivltqspgtlslspgeratlscrasqsvsssflawyqqkpgqaprlliyyassratgipdrfsgsgsgtdftltisrlepedfavyycqqtgripptfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1764]
xii.产生cova14136,方式为将承载c末端装订的scfv bha10(vh-vl取向seq id no:77)融合(seq id no:99,包含seq id no:100)的抗rsv b21m抗体重链与抗edb抗体edbmab1(图15l)的重链(hc;seq id no:80)和轻链(lc;seq id no:81)共表达。
[1765]
seq id no:77[装订的scfv bha10(vh-vl)]
[1766]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1767]
seq id no:99(hc b21m c末端装订的bha10(vh-vl),igg1,杵,无pa突变)
[1768]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgkggggsggggsggggsqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1769]
seq id no:100(hc b21m(rsv)igg1杵)
[1770]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1771]
seq id no:80(hc b21m(rsv)igg1臼)
[1772]
qitlkesgptlvkptqtltltctfsgfslstsgmgvswirqppgkalewlahiywdddkrynpslksrltitkdtsknqvvltmtnmdpvdtatyycarlygftygfaywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1773]
seq id no:81[lc b21m(rsv)]
[1774]
divmtqspdslavslgeratincrasqsvdyngisymhwyqqkpgqppklliyaasnpesgvpdrfsgsgsgtdftltisslqaedvavyycqqiiedpwtfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1775]
xiii.产生cova14146,方式为将承载c末端装订的scfv bha10(vh-vl取向seq id no:77)融合(seq id no:101,包含seq id no:102)的抗间皮素抗体mslnmab1重链与抗间皮素抗体mslnmab1(图15m)的重链(hc;seq id no:103)和轻链(lc;seq id no:104)共表达。
[1776]
seq id no:77[装订的scfv bha10(vh-vl)]
[1777]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsa
svgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1778]
seq id no:101(mslnmab1 hc c末端装订的bha10(vh-vl),igg1,杵,具有pa突变)
[1779]
qvqlqqsgpelekpgasvkisckasgysftgytmnwvkqshgkslewiglitpyngassynqkfrgkatltvdkssstaymdllsltsedsavyfcarggydgrgfdywgsgtpvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgkggggsggggsggggsqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1780]
seq id no:102(mslnmab1 hc,igg1,杵,具有pa突变)
[1781]
qvqlqqsgpelekpgasvkisckasgysftgytmnwvkqshgkslewiglitpyngassynqkfrgkatltvdkssstaymdllsltsedsavyfcarggydgrgfdywgsgtpvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnrftqkslslspgk
[1782]
seq id no:103(hc mslnmab1 igg1臼)
[1783]
qvqlqqsgpelekpgasvkisckasgysftgytmnwvkqshgkslewiglitpyngassynqkfrgkatltvdkssstaymdllsltsedsavyfcarggydgrgfdywgsgtpvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1784]
seq id no:104(lc mlsnmab1)
[1785]
dieltqspaimsaspgekvtmtcsasssvsymhwyqqksgtspkrwiydtsklasgvpgrfsgsgsgnsysltissveaeddatyycqqwskhpltfgsgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1786]
上文所述的所有构建体均能够以高产量和高纯度表达和纯化(参见下表11),表明结合本文所提供的spfv的双特异性构建体具有良好的生物物理特性。
[1787]
表11:所选择的ebd/ltbr双特异性抗体的产量和纯度
[1788][1789]
6.7实施例7:edb依赖性体外ltbr激活
–
nf-κb荧光素酶报告基因测定
[1790]
为了证明edb/ltbr双特异性抗体能够以edb依赖性方式激活ltbr,在存在或不存在含有edb的纤连蛋白(edb 纤连蛋白)的情况下,在a549细胞nf-κb荧光素酶报告基因测定中测试了化合物的活性。nf-κb信号传导在调节细胞发育和免疫稳态中起到关键作用。通过肿瘤坏死因子受体(tnfr)或tnfr超家族成员(例如ltbr)实现的nf-κb激活在与它们各自的配体接合时发生。a549肺上皮细胞系天然地表达ltbr,并且nf-κb荧光素酶报告基因构建体稳定地整合到a549肺上皮细胞系的基因组中。在被刺激物激活后,内源性nf-κb转录因子与dna应答元件结合以诱导荧光素酶基因的转录。
[1791]
为了证明ltbr的edb依赖性激活,将高结合96孔μclear平底板(greiner;monroe,nc)用150ng/孔人重组edb 纤连蛋白结构域7-b-8-9(edb ;seq id no:105)或150ng/孔人重组纤连蛋白结构域7-8-9(edb-;seq id no:106)包被过夜,所述序列在下面列出。
[1792]
seq id no:105(纤连蛋白结构域7b89)
[1793]
plspptnlhleanpdtgvltvswersttpditgyritttptngqqgnsleevvhadqssctfdnlspgleynvsvytvkddkesvpisdtiipevpqltdlsfvditdssiglrwtplnsstiigyritvvaagegipifedfvdssvgyytvtglepgidydisvitlinggesapttltqqtavppptdlrftnigpdtmrvtwapppsidltnflvryspvkneedvaelsispsdnavvltnllpgteyvvsvssvyeqhestplrgrqktgldsptgidfsditansftvhwiapratitgyrirhhpehfsgrpredrvphsrnsitltnltpgteyvvsivalngreesplligqqsthhhhhh
[1794]
seq id no:106(纤连蛋白结构域789)
[1795]
plspptnlhleanpdtgvltvswersttpditgyritttptngqqgnsleevvhadqssctfdnlspgleynvsvytvkddkesvpisdtiipavppptdlrftnigpdtmrvtwapppsidltnflvryspvkneedvaelsispsdnavvltnllpgteyvvsvssvyeqhestplrgrqktgldsptgidfsditansftvhwiapratitgyrirhhpehfsgrpredrvphsrnsitltnltpgteyvvsivalngreesplligqqsthhhhhh
[1796]
在温育过夜之后,将经包被的板用pbs洗涤,并且在37℃下用测定培养基(dmem 10%加热灭活的fbs)封闭2小时。在测定培养基中制备待测试化合物的1:5稀释系列,作为2倍浓度的储备液(测试的最终浓度在200nm至2.6pm的范围内)。在通过抽吸去除封闭溶液之后,将50μl经稀释的化合物添加到预先封闭的板中。将50μl a549细胞悬浮液(细胞悬浮液浓度=40万个细胞/ml测定培养基)添加到每个孔中(20,000个细胞/孔)。预先通过使用accutase/edta将a549细胞从细胞培养瓶中分离出来,然后转移到测定培养基中。将细胞与化合物在37℃/5%co2下温育18小时至20小时。
[1797]
温育18小时之后,将bio-glo荧光素酶测定系统(promega;madison,wi)用于检测
荧光素酶活性。使用tecan m1000 pro仪器以500毫秒的积分时间测量发光。根据所得的相对光单位(rlu),ltbr信号传导的诱导倍数计算如下:诱导倍数=rlu
受刺激
细胞/平均rlu
未受刺激
细胞(在每个所测试的板中包含未受刺激的细胞作为对照)。
[1798]
使用graphpad prism绘制剂量响应曲线(包括标准偏差),并且如果适用的话,应用非线性拟合(log(激动剂)对比响应(可变斜率-三个参数))。为了拟合数据,使用graphpad prism的x=log(x)函数转化x值(化合物的浓度)。
[1799]
在相同的a549 nf-κb报告基因测定中将cova1482与cova1456进行比较。cova1482仅在用于scfv的稳定化方法中不同于cova1456。使用本文所述的装订平台稳定化(即,经由vh/接头和vl接头二硫键稳定化)也衍生自ltbrmab1的cova1482中的scfv,其中cova1456在vh与vl之间通过二硫化物稳定化(即,经由vh/vl二硫键稳定化)。图16a示出cova1482和cova1456均以edb依赖性方式强效地激活ltbr。对应的同种型对照cova1486和cova1462未激活ltbr(图16a)。这些结果证明,将spfv结合到多特异性分子中对该多特异性分子的活性没有负面影响。在这种报告基因测定中,2:1双特异性edb/ltbr抗体(cova1482或cova1456)在诱导nf-κb信号传导方面示出增加的效力。在使用相同实验设置的几个测定中针对cova1482计算的平均ec
50
为约30pm
±
10pm,而对照1:1异源二聚体构建体示出该测定中的ec
50
为约3nm(这里未示出数据),这表明2:1双特异性抗体的效力可以是1:1双特异性抗体的100倍。这可以通过用2个taa结合位点实现的ltbr结合位点的聚集增加来解释。
[1800]
为了研究对ltbr的亲和力对此类双特异性抗体以taa依赖性方式激活ltbr的能力的影响,产生了衍生自ltbrmab1的scfv片段的较低亲和力变体(seq id no:107,kd≈60nm和seq id no:108,kd≈600nm)并且用于构建2:1双特异性抗体cova14107和cova14108(参见表9)。
[1801]
seq id no:107[装订的scfv(vl3_y36f_s49y_f87y)bha10(vh-vl)]
[1802]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawfqqkpgkapksliysasyrysgvpsrfsgsgsgtdftltisslqpedfatyycqqydtypftfgcgtkveik
[1803]
seq id no:108[装订的scfv(vh_cdr1_y33a)bha10(vh-vl)]
[1804]
qvqlvqsgaevkkpgssvkvsckasgytfttyalhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1805]
在a549 nf-κb报告基因测定中测试所产生的双特异性,以查看亲和力对激活ltbr的影响。图16b示出在该测定中对ltbr的较低亲和力对应于双特异性抗体以taa依赖性方式激活ltbr的较低能力。数据还证明,将spfv结合到多特异性分子中对该分子的活性没有负面影响。
[1806]
如实施例6中所提到的,在一些构建体的fc中引入消除与蛋白质a的结合(用于纯化抗体)的突变(wo2010151792),以便促进所需异源二聚体的纯化。产生没有这些突变的cova14133,并且将其活性与cova1482比较,以证明fc区中的突变不影响双特异性抗体的活性。在a549 nf-κb报告基因测定中比较cova14133和cova1482以及它们相应的同种型对照
cova14136和cova1486。图16c示出cova14133以taa依赖性方式激活ltbr,其效率与cova1482相似,证明fc中的突变不影响双特异性抗体激活ltbr的能力,并且对spfv的功能没有影响。
[1807]
总而言之,证明cova14133具有以taa依赖性方式激活ltbr的优异能力。
[1808]
6.8实施例8:edb依赖性体外ltbr激活
–
a375/wi38va亚系2ra共培养细胞测定
[1809]
进行a375/wi38va亚系2ra共培养测定,以验证在edb 纤连蛋白(由wi38va细胞产生并且沉积在细胞外基质中(zardi,l.等人,embo j,6,2337-42(1987)))存在下的ltbr激活是否导致细胞因子和趋化因子的释放以及a375细胞上的粘附分子icam-1的上调,并且验证结合spfv对活性没有负面影响。将wi38va亚系2ra(ccl75.1
tm
)细胞以5000个细胞/孔的密度接种在96孔板中,并且在其生长培养基(没有谷氨酰胺的mem 10%加热灭活的fbs 0.1mm neaa 2mm l-gln 1mm丙酮酸钠)中在37℃/5%co2下温育48小时。在测定培养基(dmem 10%加热灭活的fbs)中制备待测试化合物的1:5稀释系列,作为2倍浓度的储备液(测试的最终浓度在40nm至0.5pm的范围内)。在与wi38va亚系2ra细胞共培养温育之前,将a375细胞(crl-1619
tm
)用celltrace紫(ctv,invitrogen;carlsbad,ca)标记。为了标记,将浓度为10
×
106个细胞/ml并且具有在5%fbs的pbs溶液中的2.5μm ctv的细胞悬浮液在避光的同时在室温下温育5分钟。然后洗涤细胞,并且以0.4
×
106个细胞/ml的密度重悬在测定培养基中。从含有48小时wi38va亚系2ra培养物的板中小心去除培养基,然后在每个孔中添加50μl a375细胞悬浮液(20,000个细胞/孔;ctv 或ctv-)。将50μl的系列稀释化合物(每个孔的最终体积为100μl)添加到细胞中并且在37℃/5%co2下温育24小时。
[1810]
在温育24小时之后,通过离心澄清上清液,并且储存用于使用msd测定来测量细胞因子和趋化因子。通过流式细胞术进一步处理细胞以进行icam-1测量。
[1811]
6.8.1通过流式细胞术检测icam-1
[1812]
小心地去除留在96孔板中的任何培养基,将细胞与accutase分离,转移到深孔96孔板(一式三份合并在1个孔中),洗涤,重悬在100μlfacs缓冲液(pbs 1%fbs 0.1%nan3)中并且转移至圆底96孔板。将抗体(即,标记的抗人icam-1pe(克隆1h4,thermo;waltham,ma)或标记的同种型对照抗体pe(mpc-11,biolegend;san diego,ca)和live/dead可固定近红外染色剂(invitrogen)(单一染色或组合染色)如表12所示进行稀释。
[1813]
表12:facs缓冲液中的单一染色或组合染色的稀释方案
[1814][1815]
将细胞在4℃下以400
×
g离心4分钟,弃去上清液,并且如表12中所述制备50μl抗体溶液。将细胞和抗体在黑暗中在4℃下温育30分钟。温育之后,在每个孔中添加120μl,然后将细胞在4℃下以400
×
g离心4分钟。将细胞用facs缓冲液洗涤一次,离心并且重悬在90μl facs缓冲液中。然后通过添加90μl 3.7%福尔马林的pbs溶液来固定细胞,并且在黑暗中在冰上温育15分钟。固定之后,将细胞在4℃下以400
×
g离心4分钟,并且重悬在100μl facs
缓冲液中。使用macs quant仪器以筛选模式在高流速下测量细胞,获得49μl/孔。使用flowlogics软件(版本700.2a)分析数据并且用graphpad prism绘图。
[1816]
6.8.2使用msd平台对经处理细胞的上清液中的细胞因子进行测量
[1817]
使用msd平台和多重msd板测量已知处于nf-κb信号传导控制下的几种细胞因子。这里列出了测量的细胞因子的一些示例:
[1818]
·
rantes:使用r-plex抗体集人rantes(msd);
[1819]
·
i-tac、ip-10、mip-3b:使用3-plex细胞因子释放测定(msd);
[1820]
·
il-8、ip-10、mip-3b:使用3-plex细胞因子释放测定(msd);以及
[1821]
·
il-12p70、il-6、tnf-a、mip-3a、sdf-1a:使用5-plex细胞因子释放测定(msd)
[1822]
使用msd平台按照制造商的说明书测量经处理细胞的上清液中的细胞因子的浓度。简言之,该方案涉及以下步骤:
[1823]
(1)板的制备涉及用接头偶联的捕获抗体包被所提供的板。将板在2℃至8℃下振荡温育过夜。第二天,使用洗板机(biotek;winooski,vt)用pbst(pbs加0.05%tween-20)洗涤板;
[1824]
(2)制备校准品标准品和检测抗体溶液;
[1825]
(3)根据材料的可用性,将上清液按1:3或1:5稀释。
[1826]
6.8.3测定方案:
[1827]
ο步骤1:将样品或校准品标准品添加到板中,并且将板在室温下在振荡的同时温育1小时;
[1828]
ο步骤2:洗涤板,并且添加检测抗体。将板在室温下振荡温育1小时。
[1829]
ο步骤3:洗涤板,并且添加2
×
读数缓冲液t。在msd仪器上分析板。
[1830]
使用mesoscale软件(msd discovery work bench程序4.0.12.1版)分析数据,并且使用graphpad prism绘图。
[1831]
6.8.4通过流式细胞术检测icam-1
[1832]
先前已证明nf-κb信号传导可以导致细胞表面上的icam-1上调(da silva antunes等人,front immunol,9:576,(2018))。因此,测量了与edb/ltbr双特异性抗体共培养温育后a375细胞表面上的icam-1表达的水平。例如,图17示出在与edb/ltbr双特异性cova1482一起温育后icam-1的上调,证明了ltbr结合spfv的功能性。同种型对照分子cova1486未引起icam-1上调。这些发现表明,经由与edb结合而聚集ltbr scfv的能力是ltbr激活(以及因此引起的icam-1上调)的先决条件。
[1833]
6.8.5结果-在经处理的细胞的上清液中的细胞因子测量结果
[1834]
在用如上所述的edb/ltbr双特异性抗体和对照分子处理的共培养物的上清液中测量作为ltbr激活的结果表达的几种细胞因子和趋化因子。图18a至图18d示出了通过用cova14133激活ltbr而上调的4个细胞因子代表性示例(图18a:rantes,图18b:il-6,图18c:il-8和图18d:mip-3b)。cova14136中的非靶向ltbrmab1衍生的scfv没有将ltbr激活,因此,上清液中的细胞因子的浓度未增加到高于背景。背景由用b21m抗体或cova1440(2(mab)b21m同种型对照mab)实现的水平(在曲线图中作为单一浓度示出)表示。结果证明,ltbr结合spfv在体外是有功能的。
[1835]
总的来说,ltbr激活后的icam-1上调和细胞因子分泌证实了ltbr激活可能对细胞
具有的预期作用。
[1836]
在该实施例中,证明本文所提供的分子实现了有效的肿瘤相关抗原(此处是含有edb的纤连蛋白)依赖性ltbr激活。
[1837]
6.9实施例9:间皮素依赖性体外ltbr激活
–
与a549 nf-κb报告基因细胞和chok1-人msln或h226的共培养细胞测定
[1838]
在实施例7和实施例8中,证明包含本文所提供的spfv结构且靶向edb(细胞外基质中的肿瘤相关抗原)和ltbr的双特异性抗体以肿瘤抗原依赖性方式非常有效地激活ltbr。为了验证该发现是否适用于任何肿瘤抗原,而不管其位置在何处(沉积在细胞外基质中或肿瘤细胞的细胞表面上),如实施例6中所述设计并且生产了靶向间皮素(msln)的双特异性2:1抗体、在不同类型的肿瘤上表达的肿瘤相关抗原(hassan和ho,eur.j.cancer,44:46-53(2008))和ltbr。cova14146是2:1msln/ltbr双特异性抗体,其由与衍生自ltbrmab1的spfv片段融合的抗间皮素抗体(mslnmab1)组成。要证明靶向ltbr和肿瘤细胞的细胞表面上存在的肿瘤相关抗原(例如,间皮素)的ltbr双特异性抗体是否能够以肿瘤依赖性方式有效地激活ltbr,使用了共培养细胞测定。所使用的共培养测定是a549细胞nf-κb荧光素酶报告基因细胞测定(实施例7中所述)和已知表达间皮素(fan等人,mol.canc.ther.1:595
–
600(2002))和ltbr的h226细胞(间皮瘤细胞系;crl-5826)。
[1839]
6.9.1制备h226细胞
[1840]
将h226细胞(表达间皮素的约200,000个拷贝和ltbr的10,000个拷贝)悬浮液以10,000个细胞/孔(在75μl测定培养基中:dmem 10%fbs-hi)接种在96孔组织培养板中,并且在37℃/5%co2下在其生长培养基(分别为mem 2mm谷氨酰胺 10%fbs-hi 10μg/ml嘌呤霉素和rpmi-1640 10%fbs 1mm丙酮酸钠)中温育6小时,以允许细胞附着到板上。
[1841]
6.9.2制备化合物
[1842]
在从100nm降低至1.3pm的浓度范围内测试化合物。在测定培养基(dmem 10%fbs-hi)中制备化合物的4倍1/5系列稀释液,并且在4℃下储存直到使用。
[1843]
6.9.3制备并添加a549报告基因细胞
[1844]
将a549报告基因细胞从具有accutase/edta的细胞培养瓶中分离出来,然后转移到测定培养基(dmem 10%fbs-hi)中。在向装有h226细胞的板中添加每孔总共20,000个a549报告基因细胞之后,将50μl预稀释化合物添加到每个孔中并且在37℃/5%co2下温育20小时。
[1845]
6.9.4测量经处理的共培养物中的发光
[1846]
温育20小时之后,将bio-glo荧光素酶测定系统(promega;madison,wi)根据制造商的说明书用于检测荧光素酶活性。使用tecan m1000 pro仪器以500毫秒的积分时间测量发光。根据所得的相对光单位(rlu),ltbr信号传导的诱导倍数计算如下:诱导倍数=rlu
受刺激
细胞/平均rlu
未受刺激
细胞(在每个所测试的板中包含未受刺激的细胞作为对照)。
[1847]
使用graphpad prism绘制剂量响应曲线(包括标准偏差),并且如果适用的话,应用非线性拟合(log(激动剂)对比响应(可变斜率-三个参数))。为了拟合数据,使用graphpad prism的x=log(x)函数转化x值(化合物的浓度)。
[1848]
6.9.5使用msd平台对经处理细胞的上清液中的细胞因子进行测量
[1849]
可以使用msd平台和多重msd板测量已知处于nf-κb信号传导控制下的几种细胞因
子。例如,本文描述了使用r-plex抗体集人rantes(msd)测量rantes的方法。
[1850]
使用msd平台按照制造商的说明书测量经处理细胞的上清液中的rantes的浓度。简言之,该方案涉及以下步骤:
[1851]
(1)板的制备涉及用接头偶联的捕获抗体包被所提供的板。将板在2℃至8℃下振荡温育过夜。第二天,使用洗板机(biotek;winooski,vt)用pbst(pbs加0.05%tween-20)洗涤板;
[1852]
(2)制备校准品标准品和检测抗体溶液;
[1853]
(3)根据材料的可用性,将上清液按1:3或1:5稀释。
[1854]
6.9.6测定方案:
[1855]
ο步骤1:将样品或校准品标准品添加到板中,并且将板在室温下在振荡的同时温育1小时;
[1856]
ο步骤2:洗涤板,并且添加检测抗体。将板在室温下振荡温育1小时。
[1857]
ο步骤3:洗涤板,并且添加2
×
读数缓冲液t。在msd仪器上分析板。
[1858]
使用mesoscale软件(msd discovery work bench程序4.0.12.1版)分析数据,并且使用graphpad prism绘图。
[1859]
6.9.7结果
–
a549报告基因细胞/h226共培养测定中的ltbr的间皮素依赖性激活
[1860]
进行使用a549报告基因细胞和h226细胞的共培养测定,以验证cova14146是否能够在以下具有更强生理性的系统中激活ltbr:在该系统中,由于cova14146广泛表达(lukashev等人,cancer res.,66(19):9617-24(2006)),预期ltbr和间皮素(以及肿瘤细胞的细胞表面上的其他肿瘤相关抗原,例如egfr)将在肿瘤细胞的细胞表面上共表达。在图19a中,证明在这些条件下,cova14146未有效地激活ltbr。测量经处理细胞的上清液中分泌的rantes的浓度,以证实cova14146不能有效地激活ltbr。如所预期的,图19b示出rantes由用cova14146处理的细胞分泌,分泌程度与用同种型对照分子cova1486处理的细胞相同,从而证实ltbr在这些条件下不能被激活。
[1861]
6.10实施例10:特异性地结合ltbr和存在于细胞外基质中的其他taa的双特异性抗体
[1862]
以上实施例证明,靶向ltbr和细胞外基质中表达的taa(在该情况下为纤连蛋白的edb)的双特异性抗体能够在表达edb的肿瘤组织中选择性地激活ltbr。该实施例证明,通过提供另外两个此类taa的示例,即腱生蛋白c的结构域a2和纤连蛋白的额外结构域a,这更普遍地适用于靶向ltbr和存在于细胞外基质中的另一种taa的双特异性抗体。
[1863]
通常按照上述实施例中描述的方法,制备了与ltbr和存在于细胞外基质中的其他taa(即,腱生蛋白c的结构域a2(tnca2)或纤连蛋白的额外结构域a(eda))结合的另外的双特异性抗体。这些taa的序列已经例如描述于以下各项中:
[1864]-tnca2:uniprot登录号p24821.3;
[1865]-eda:uniprot登录号p02751。
[1866]
此外,针对这些taa的抗体已经例如先前描述于以下各项中:
[1867]-tnca2:wo2011/020783;
[1868]-eda:ep2142567。
[1869]
用于本实施例(通过与ltbr scfv组合以产生本发明的多特异性结合分子)的针对
这些靶标的结合结构域的示例性序列被提供为:
[1870]-tnca2:vh seq id no:109和vl seq id no:110;
[1871]-eda:vh seq id no:111和vl seq id no:112。
[1872]
这些序列如下:
[1873]
seq id no:109(抗tnca2 ab 2b10的vh)
[1874]
qvqlvqsgaevkkpgssvkvsckasggtfssyaiswvrqapgqglewmggiipifgtanyaqkfqgrvtitadkststaymelsslrsedtavyycarlygyayygafdywgqgttvtvss
[1875]
seq id no:110(抗tnca2 ab 2b10的vl)
[1876]
diqmtqspsslsasvgdrvtitcrasqgirndlgwyqqkpgkapkrliyaasslqsgvpsrfsgggsgteftltisslqpedfatyyclqnglqpatfgqgtkveik
[1877]
seq id no:111(抗eda ab f8的vh)
[1878]
evqllesggglvqpggslrlscaasgftfslftmswvrqapgkglewvsaisgsggstyyadsvkgrftisrdnskntlylqmnslraedtavyycaksthlylfdywgqgtlvtvss
[1879]
seq id no:112(抗eda ab f8的vl)
[1880]
eivltqspgtlslspgeratlscrasqsvsmpflawyqqkpgqaprlliygassratgipdrfsgsgsgtdftltisrlepedfavyycqqmrgrpptfgqgtkveik
[1881]
制备与ltbr和这些taa结合的双特异性抗体,并且按照如实施例7中所述的类似程序,在taa依赖性体外ltbr激活
–
nf-κb荧光素酶报告基因测定中测试ltbr的taa特异性激活。
[1882]
如下产生具有2:1化学计量比的非对称抗体:
[1883]
产生cova14198,方式为将承载c末端装订的scfv bha10(vh-vl取向seq id no:77)融合(seq id no:113,包含seq id no:114)的抗eda抗体重链与抗eda抗体的重链(hc;seq id no:115)和轻链(lc;seq id no:116)共表达。这些序列在下面列出。
[1884]
seq id no:77[装订的scfv bha10(vh-vl)]
[1885]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1886]
seq id no:113(f8 hc,具有bha10装订的(vh-vl)scfv c末端融合的igg1σ,杵突变)
[1887]
evqllesggglvqpggslrlscaasgftfslftmswvrqapgkglewvsaisgsggstyyadsvkgrftisrdnskntlylqmnslraedtavyycaksthlylfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgkggggsggggsggggsqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcq
qydtypftfgcgtkveik
[1888]
seq id no:114(f8 hc,igg1σ,杵突变)
[1889]
evqllesggglvqpggslrlscaasgftfslftmswvrqapgkglewvsaisgsggstyyadsvkgrftisrdnskntlylqmnslraedtavyycaksthlylfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1890]
seq id no:115(f8 hc,igg1σ和臼突变)
[1891]
evqllesggglvqpggslrlscaasgftfslftmswvrqapgkglewvsaisgsggstyyadsvkgrftisrdnskntlylqmnslraedtavyycaksthlylfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1892]
seq id no:116(f8 lc)
[1893]
eivltqspgtlslspgeratlscrasqsvsmpflawyqqkpgqaprlliygassratgipdrfsgsgsgtdftltisrlepedfavyycqqmrgrpptfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1894]
产生cova14202,方式为将承载c末端装订的scfv bha10(vh-vl取向seq id no:77)融合(seq id no:117,包含seq id no:118)的腱生蛋白c抗体重链的抗结构域a2与腱生蛋白c抗体的抗结构域a2的重链(hc;seq id no:119)和轻链(lc;seq id no:120)共表达。这些序列在下面列出。
[1895]
seq id no:77[装订的scfv bha10(vh-vl)]
[1896]
qvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfatyfcqqydtypftfgcgtkveik
[1897]
seq id no:117(2b10 hc,具有bha10装订的(vh-vl)scfv c末端融合的igg1σ,杵突变)
[1898]
qvqlvqsgaevkkpgssvkvsckasggtfssyaiswvrqapgqglewmggiipifgtanyaqkfqgrvtitadkststaymelsslrsedtavyycarlygyayygafdywgqgttvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgkggggsggggsggggsqvqlvqsgaevkkpgssvkvsckasgytfttyylhwvrqapgcglewmgwiypgnvhaqynekfkgrvtitadkststaymelsslrsedtavyycarswegfpywgqgttvtvssgggsgggsgcppcggggdiqmtqspsslsasvgdrvtitckasqnvginvawyqqkpgkapkslissasyrysgvpsrfsgsgsgtdftltisslqpedfaty
fcqqydtypftfgcgtkveik
[1899]
seq id no:118(2b10 hc,igg1σ,杵突变)
[1900]
qvqlvqsgaevkkpgssvkvsckasggtfssyaiswvrqapgqglewmggiipifgtanyaqkfqgrvtitadkststaymelsslrsedtavyycarlygyayygafdywgqgttvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvytlppcreemtknqvslwclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1901]
seq id no:119(2b10 hc,igg1σ和臼突变)
[1902]
qvqlvqsgaevkkpgssvkvsckasggtfssyaiswvrqapgqglewmggiipifgtanyaqkfqgrvtitadkststaymelsslrsedtavyycarlygyayygafdywgqgttvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapeaagassvflfppkpkdtlmisrtpevtcvvvdvsaedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpssiektiskakgqprepqvctlppsreemtknqvslscavkgfypsdiavewesngqpennykttppvldsdgsfflvskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
[1903]
seq id no:120(2b10 lc)
[1904]
diqmtqspsslsasvgdrvtitcrasqgirndlgwyqqkpgkapkrliyaasslqsgvpsrfsgggsgteftltisslqpedfatyyclqnglqpatfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
[1905]
为了证明ltbr的taa依赖性激活,将高结合96孔μclear平底板(greiner;monroe,nc)用以下物质包被过夜:
[1906]-150ng/孔的含有重组结构域a2(a2
)的腱生蛋白c结构域a1-a2-a3(tnca2
;seq id no:121)或150ng/孔的人重组腱生蛋白结构域a1-a3(即,缺少结构域a2,表示为a2-)(tnca2-;seq id no:122);或者
[1907]-150ng/孔的含有重组eda(eda
)的纤连蛋白结构域11-a-12(eda
;seq id no:123)或150ng/孔的人重组纤连蛋白结构域11-12(即,不含eda,表示为eda-)(eda-;seq id no:124)。
[1908]
上文提到的序列如下:
[1909]
seq id no:121(人腱生蛋白c结构域a1-a2-a3,包括用于纯化的(his)
6-标签)
[1910]
eqapelenltvtevgwdglrlnwtaadqayehfiiqvqeankveaarnltvpgslravdipglkaatpytvsiygviqgyrtpvlsaeastgetpnlgevvvaevgwdalklnwtapegayeyffiqvqeadtveaaqnltvpgglrstdlpglkaathytitirgvtqdfsttplsvevlteevpdmgnltvtevswdalrlnwttpdgtydqftiqvqeadqveeahnltvpgslrsmeipglragtpytvtlhgevrghstrplavevvthhhhhh
[1911]
seq id no:122(人腱生蛋白c结构域a1-a3,包括用于纯化的(his)
6-标签)
[1912]
eqapelenltvtevgwdglrlnwtaadqayehfiiqvqeankveaarnltvpgslravdipglkaatpytvsiygviqgyrtpvlsaeeevpdmgnltvtevswdalrlnwttpdgtydqftiqvqeadqveeahnltvpgslrsmeipglragtpytvtlhgevrghstrplavevvthhhhhh
[1913]
seq id no:123(人纤连蛋白结构域11-a-12,包括用于纯化的(his)
6-标签)
[1914]
eidkpsqmqvtdvqdnsisvkwlpssspvtgyrvtttpkngpgptktktagpdqtemtieglqptveyv
vsvyaqnpsgesqplvqtavtnidrpkglaftdvdvdsikiawespqgqvsryrvtysspedgihelfpapdgeedtaelqglrpgseytvsvvalhddmesqpligtqstaipaptdlkftqvtptslsaqwtppnvqltgyrvrvtpkektgpmkeinlapdsssvvvsglmvatkyevsvyalkdtltsrpaqgvvttlehhhhhh
[1915]
seq id no:124(人纤连蛋白结构域11-12,包括用于纯化的(his)
6-标签)
[1916]
eidkpsqmqvtdvqdnsisvkwlpssspvtgyrvtttpkngpgptktktagpdqtemtieglqptveyvvsvyaqnpsgesqplvqtavttipaptdlkftqvtptslsaqwtppnvqltgyrvrvtpkektgpmkeinlapdsssvvvsglmvatkyevsvyalkdtltsrpaqgvvttlehhhhhh
[1917]
这些测定的结果与上文针对edb/ltbr双特异性抗体所示的结果一致,即,它们证明ltbr的激活主要在由相应的双特异性结合分子所靶向的细胞外基质的肿瘤抗原的存在下发生(表13)。
[1918]
表13:在存在或不存在eda或tnca2的情况下,分别针对cova14198和cova14202的nf-κb信号传导的最大诱导倍数。
[1919][1920]
特别地,tnca2/ltbr双特异性结合分子可以在tnca2抗原的存在下强烈地激活ltbr,并且因此触发nf-κb信号传导,并且eda/ltbr双特异性结合分子可以在eda抗原的存在下强烈地激活ltbr,并且因此触发nf-κb信号传导。在不存在相应taa的情况下,这些双特异性抗体不激活或仅最小程度地激活ltbr。
[1921]
该实施例证明,经由本文所提供的多特异性结合分子激活ltbr的方法(例如,与ltbr和与细胞外基质相关联的肿瘤相关抗原两者结合)对迄今为止已测试过的细胞外基质的全部三种不同的肿瘤相关抗原起作用,因此普遍地适用于存在于细胞外基质中的肿瘤相关抗原。该实施例还证明,结合spfv的分子是有功能的。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。
