荔枝mads-box转录因子lcmads1及其在抑制植物器官脱落中的应用
技术领域
1.本发明涉及植物基因工程领域,尤其涉及荔枝mads-box转录因子lcmads1及其在抑制植物器官脱落中的应用。
背景技术:
2.荔枝(litchi chinensis sonn.)属于无患子科(sapindaceae)荔枝属(litchi),是我国特色果树,在生产中常出现“花而不实”、“满树花半树果,甚至颗粒无收”的场面,对荔枝产业的稳产、优质、健康、可持续生产带来困难。
3.mads-box是一类广泛分布于真核生物中的转录因子,在植物器官脱落过程中起着重要作用。过表达拟南芥mads-box家族成员agl15、agl18、atfyf都可以推迟拟南芥花器官脱落和衰老,而agl15/agl18双突变体花器官脱落早于野生型;stk是控制拟南芥珠柄发育和种子脱落的重要mads-box家族成员,stk突变体果荚与种子的体积变小,但珠柄细胞的分裂程度提高,珠柄与种子连接处的离区细胞特征丢失,成熟种子不能自由脱落。番茄mads-box家族成员j、mc和slmbp21可能是通过形成j-mc-slmbp21复合体来调控番茄花(果)柄离区的分化,例如在j突变体、mc反义转基因植株和slmbp21-rnai植株中,番茄花(果)柄离区均不能形成;而过表达slmbp21后番茄果柄近轴端变短并出现异位的类似离区的组织结构;将j、mc和slmbp21共转到野生型番茄后果柄明显变短并转变成类似离区的组织结构,没有明显的果柄近轴端、离区和远轴端的分化;j、mc和slmbp21蛋白之间可以两两发生互作,j和slmbp21蛋白自身也能够形成二聚体;基因表达分析表面三者有共同调控的下游基因,如lateral suppressor(ls),wuschel(lewus),goblet(gob),blind(bl);与拟南芥stk高度同源的番茄sl-agl11基因的过表达植株表现出果柄离区异常膨大且果实成熟时不脱落的现象。苹果的j同源基因mdjb异源超表达于番茄jointless突变体后可以恢复花(果)柄离区的结构和功能;
‘
库尔勒香梨’的j同源基因psjointless在番茄野生型中的异源超表达可导致花柄脱落率和纤维素酶活性显著提高,花柄变细,离区细胞数目增加且排列变得无序。可见,mads-box转录因子在植物器官脱落过程中发挥着重要的调控作用,关于荔枝mads-box转录因子的研究成果将为选育落果少的品种提供重要的理论指导,为产量的提高提供有效的育种手段,对农业生产实践具有重要意义。
技术实现要素:
4.本发明的首要目的在于克服现有技术的缺点与不足,提供一种荔枝mads-box转录因子lcmads1。
5.本发明的另一目的在于提供上述荔枝mads-box转录因子lcmads1在抑制植物器官脱落中的应用。
6.本发明的目的通过下述技术方案实现:
7.一种荔枝mads-box转录因子lcmads1,其氨基酸序列如seq id no:2所示。
8.上述荔枝mads-box转录因子lcmads1相关的生物材料,为下述生物材料中的任意一种或多种组合:
9.1)编码所述荔枝mads-box转录因子lcmads1的核酸分子;
10.2)含有1)中所述核酸分子的表达盒;
11.3)含有1)中所述核酸分子的重组载体,或含有2)中所述表达盒的重组载体;
12.4)含有1)中所述核酸分子的重组微生物,或含有2)中所述表达盒的重组微生物,或含有3)所述重组载体的重组微生物;
13.5)含有1)中所述核酸分子的转基因植物组织、或含有2)中所述表达盒的转基因植物组织。
14.进一步地,1)中所述核酸分子的序列如seq id no:1所示。
15.进一步地,3)中所述重组载体通过将seq id no:1所示序列构建到pcambia1302植物表达载体中得到。
16.进一步地,4)中所述重组微生物通过将3)中所述重组载体转化到农杆菌gv3101中得到。
17.上述荔枝mads-box转录因子lcmads1或荔枝mads-box转录因子lcmads1相关的生物材料在抑制植物器官脱落中的应用。
18.上述荔枝mads-box转录因子lcmads1或荔枝mads-box转录因子lcmads1相关的生物材料在培育器官脱落延迟的转基因植物中的应用。
19.进一步地,所述植物为双子叶植物或单子叶植物。
20.更进一步地,所述植物包括但不限于荔枝、拟南芥、番茄。
21.进一步地,所述器官包括但不限于果实、花瓣、花萼、花丝、叶片。
22.本发明相对于现有技术具有如下的优点及效果:
23.本发明提供了一种具有抑制植物器官脱落作用的荔枝mads-box转录因子lcmads1,在促进落果处理的果柄离区中该基因的表达量显著低于对照和抑制落果处理,提示该基因与荔枝果实脱落密切相关。
24.本发明进一步实验表明荔枝lcmads1基因的异源超表达可显著抑制拟南芥花器官的脱落,明确了荔枝lcmads1基因在调控植物器官脱落过程中具有重要作用,因此本发明提供的荔枝lcmads1基因可为植物尤其是荔枝的品种改良和选育提供方向和技术支持。
附图说明
25.图1是荔枝果柄离区中lcmads1的基因表达水平分析图;其中,a为不同处理对
‘
妃子笑’荔枝幼果脱落的影响;b为通过qrt-pcr技术分析荔枝果柄离区中lcmads1的基因表达水平;gpd为环剥去叶处理,截断树体营养供应,诱导落果;eth为乙烯利处理,诱导落果;gpdd为环剥去叶 2,4-d处理,抑制落果;数据以平均值
±
标准差表示;用lsd检验,不同小写字母代表p《0.05水平上的显著性差异。
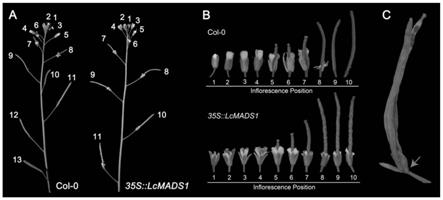
26.图2是异源超表达荔枝lcmads1基因抑制拟南芥花器官脱落观察结果图;其中,a为野生型(col-0)和异源超表达lcmads1(35s::lcmads1)株系花序;b为col-0与35s::lcmads1株系中花序的比较;c为35s::lcmads1株系成熟干枯开裂果荚;数字表示从花序顶端露出白色花瓣花朵开始,由上往下数花朵的位置;箭头指示未脱落花器官。
27.图3是col-0和35s::lcmads1株系花器官离区扫描电镜观察图;其中,a为col-0株系;b为35s::lcmads1株系;所观察花朵为p8花;箭头指示35s::lcmads1株系花器官强行拔下后的断裂面。
具体实施方式
28.下面结合实施例及附图对本发明作进一步详细的描述,但本发明的实施方式不限于此。
29.实施例1lcmads1基因的获得和在荔枝果实脱落过程中的表达模式分析
30.实验材料为
‘
妃子笑’荔枝。盛花后25d左右在植株不同方位选取长势基本一致的果穗。因为果实脱落与乙烯和营养供应密切相关,为此,我们设计了3个不同处理,每处理果穗6个,3个重复:(1)乙烯利处理(ethephon,eth),用250mg/l乙烯利浸泡果穗1min;(2)环剥去叶处理(girdling plus defoliation,gpd),在距离果穗基部10cm左右进行宽度约2cm、深度达木质部的环割,去除韧皮部,同时将环剥口以上的叶片摘除干净,阻断树体对幼果碳水化合物的供给;(3)环剥去叶 2,4-d处理(gpdd),环剥去叶处理后,用20mg/l 2,4-d浸泡果穗1min。以未做任何处理的果穗作为对照(control)。处理后每天记录落果数,连续观察3d。结果表明,eth和gpd处理均可显著地诱导荔枝幼果的大量脱落,gpdd可显著抑制荔枝幼果的脱落(图1a)。
31.于处理后0,1,2,3d采集荔枝果柄离区样品(即在果柄脱落部位上下0.2cm处切取材料),提取rna,用qrt-pcr对lcmads1基因的表达量进行检测,结果表明,在促进落果处理(eth和gpd)果柄离区中该基因的表达量显著低于对照(control)和抑制落果处理(gpdd)(图1b)。由此,我们猜测该基因与荔枝果实脱落密切相关。lcmads1基因在不同处理果柄离区中的表达量差异分析具体步骤如下:
32.1、荔枝rna的提取
33.取荔枝果柄通过柱式植物rnaout 2.0提取试剂盒(天恩泽)按照说明书提取总rna。提取得到的rna用1.2%琼脂糖凝胶电泳检测其完整性,并用紫外分光光度计(biospec-nano)检测rna浓度和od值。
34.2、反转录cdna
35.使用one-step gdna removal and cdna synthesis supermix试剂盒(transscript)将提取的rna等量反转录为cdna,稀释20倍使用。
36.3、qrt-pcr
37.利用primer 5.0软件设计lcmads1基因的特异性引物,用荔枝lcef-1α和lcgapdh作为内参基因。使用的荧光染料为gotaq qpcr master mix,使用仪器为bio-rad cfx96 pcr仪。扩增程序设定为:95℃预变性3min;95℃变性5s;55℃退火30s;72℃延伸30s;循环数40个。完成上述反应后,将数据导出,根据2
‑△△
ct
法计算基因的表达量。引物序列见表1。
38.表1lcmads1 qrt-pcr引物序列
[0039][0040]
获得的lcmads1基因的cds序列如seq id no:1所示,其开放阅读框为834bp,编码277个氨基酸,如seq id no:2所示,该基因命名为lcmads1:
[0041]
lcmads1 cds序列(seq id no:1):
[0042]
atgcaacttattattaggtacttcaagatgggaagagggaagatcgagatcaagagaatcgaaaacacgacgaatcgtcaggtcaccttctgcaagaggagaaatggactcttgaagaaagcttatgaactatcagttctttgtgatgctgaagttgccctcatcgtcttctccagccgcggtcgcctctatgagtactctaataacaacaacataaggtcaaccatagagaagtacaagaaggcagcctctgataatacaaatgcaacatctgttacagaaatcaatgctcagtattatcagcaggaatcagctaagctgcgtcaacagattcaaatgttgacgaattccaacaggcacttaatgggagattccctgagctctttgtctgtgaaggagctgaagcagctggagaataggcttgaacgaggcattactagaatcagatcaaagaagcatgaaatgttgctagctgaaattgagtacttgcagaaaagggagatggagctggaaaatgaaagtgtgactcttagatcaaggattgcggacattgagaggtttcagcagtctaacatggtaacaggcccagagctgaatgcaattcatgcgttggcttctcgaaatttcttcaatccagctttcattgaggatgcaagtggaagctcatattcccatcctgacaagaaactgctccatcttgggtattttctctacatatttcatttcattgcatttatatatttccatagttatttattggaggctaatcgggtgtgggacaaaatttcgtggggaatcagatttagtatcattacaaaaccaaccgtcagatga
[0043]
lcmads1编码的氨基酸序列(seq id no:2):
[0044]
mqliiryfkmgrgkieikrienttnrqvtfckrrngllkkayelsvlcdaevalivfssrgrlyeysnnnnirstiekykkaasdntnatsvteinaqyyqqesaklrqqiqmltnsnrhlmgdslsslsvkelkqlenrlergitrirskkhemllaeieylqkremelenesvtlrsriadierfqqsnmvtgpelnaihalasrnffnpafiedasgssyshpdkkllhlgyflyifhfiafiyfhsylleanrvwdkiswgirfsiitkptvr
[0045]
实施例2拟南芥异源超表达鉴定荔枝lcmads1基因的功能
[0046]
1、lcmads1基因全长扩增引物设计
[0047]
采用pcr方法用kod-plus-neo酶进行序列扩增,所用cdna为上述步骤中荔枝果柄样品。利用primer 5.0软件设计全长扩增引物,根据lcmads1的cds序列,分别在起始密码子atg和终止密码子处设计含有适合酶切位点引物,所用酶切位点为nco i。引物序列见表2,方框部位为酶切位点。
[0048]
表2 lcmads1全长扩增引物序列
[0049][0050]
2、基因克隆的pcr反应体系、程序和产物检测
[0051]
(1)pcr反应体系,如表3所示:
[0052]
表3 pcr反应体系
[0053][0054][0055]
(2)pcr反应程序
[0056]
94℃预变性2min,94℃变性15s,58℃退火30s,68℃延伸2min,35个循环,68℃延伸5min,12℃保存。
[0057]
(3)pcr产物的检测与回收
[0058]
用1.2%琼脂糖凝胶电泳检测pcr产物,电压为135v,电流110a,电泳30min。随后将琼脂糖凝胶放入凝胶成像系统进行成像,确认是否含有目的基因的片段,若有亮带出现且大小正确即可使用手术刀在紫外切胶仪下快速切割含有目的片段的胶块,再用dna凝胶回收试剂盒(fastpure gel dna extraction mini kit,诺唯赞)进行回收,方法参照试剂盒说明书进行,回收产物保存在-20℃冰箱中。
[0059]
3、lcmads1-pcambia1302植物表达载体构建
[0060]
通过in-fusion连接试剂盒(clonexpress ii one step cloning kit,诺唯赞)将lcmads1克隆片段插入到pcambia1302载体(启动子为camv35s,转化植物的筛选标记为潮霉素)中,连接体系如表4所示,混匀,37℃反应30min。
[0061]
表4 in-fusion连接体系
[0062][0063]
反应完成后,转大肠杆菌dh5α,通过pcr鉴定筛选阳性克隆,将验证正确的单克隆送到广州艾基生物技术有限公司测序。
[0064]
4、转化农杆菌
[0065]
经测序鉴定后,将重组质粒转化于农杆菌gv3101中,通过pcr鉴定筛选阳性克隆。
[0066]
5、农杆菌介导的拟南芥遗传转化(蘸花法)及阳性转基因株系的筛选
[0067]
将获得的含重组质粒的农杆菌在lb液体培养基(加有100mg/l kan和50mg/l rif)中培养至od=0.8,室温5000rpm离心10min收集菌体,向菌体沉淀缓慢加入已经配置好的侵染液(5%蔗糖,0.05%(v/v)silwet l-77),菌液od约为1。将野生型拟南芥col-0花序置于侵染液中1min,侵染后,拟南芥植株避光条件下培养过夜,第二天早上取出于正常条件下继续培养。为了提高侵染效率需要对以上的拟南芥每隔4-5d侵染一次,侵染3-4次后正常培养直至种子成熟。待成熟后,收获拟南芥种子,即为转基因的t0代种子。
[0068]
6、转基因拟南芥植株的表型鉴定
[0069]
成熟t0代种子干燥后,经0.5%naclo3消毒后播种于含潮霉素b(hyg,50mg/l)抗生素的ms筛选培养基上,置于4℃中春化2-3d后移至光照培养箱中培养,待其长出根和2片子叶时筛选阳性苗,将其转移至营养土中继续培养(假阳性苗子在hyg抗性存在条件下不长根,子叶黄化或者不生长)。待阳性植株幼苗长出8片叶子左右时,收集叶片进行qrt-pcr检测。
[0070]
每一代的植株都要进行阳性株系的检测,直至繁殖至t3代,获得纯合转基因拟南芥株系。
[0071]
在同等条件下种植转基因t3代株系和野生型株系,表型观察发现野生型拟南芥col-0植株从第9个花/果荚(p9)开始花器官(花萼、花瓣、雄蕊)已全部脱落,而35s::lcmads1株系的花器官并未发现明显的脱落现象,表明lcmads1的异源超表达可抑制花器官的脱落,而且花萼没有表现出明显衰老的现象,甚至果荚成熟时仍保持鲜绿,没有变黄现象,当果荚干枯后,花萼仍未脱落(图2)。
[0072]
实施例3拟南芥花器官离区表面扫描电镜观察
[0073]
分别采集col-0和35s::lcmads1株系第8位置上的花/果荚(p8),用镊子去除花器官后经faa固定液(50%无水乙醇、5%冰醋酸、1%甲醛)固定,抽气后4℃保存。样品经临界点干燥后喷金镀膜,在扫描电镜下观察并拍照。结果表明(图3),35s::lcmads1株系花器官(p8花/果荚)拔下困难,强行拔掉后离区部位会留下明显的细胞破损现象(明显断裂痕,箭头所示),而col-0株系p8位置花器官离区细胞表面光滑,表面细胞间早已松动,一碰就会掉落。
[0074]
综上,荔枝lcmads1基因在果柄离区中的表达随着荔枝果实脱落的加剧而明显下调,通过遗传转化手段将lcmads1基因进行异源超表达可显著抑制拟南芥花器官的脱落,推测,荔枝lcmads1基因具有抑制植物器官脱落的功能。
[0075]
上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受上述实施例的限制,其他的任何未背离本发明的精神实质与原理下所作的改变、修饰、替代、组合、简化,均应为等效的置换方式,都包含在本发明的保护范围之内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。
