biotechnol 21,635
‑
7,2003.,nielsen,c.b.等determinants of targeting by endogenous and exogenous micrornas and sirnas.rna 13,1894
‑
910,2007.,peek,a.s.和behlke,m.a.design of active small interfering rnas.curr opin mol ther 9,110
‑
8,2007.)。该副作用是一种被称为脱靶效应的现象。如上所述,sirna发挥具有剪切与sirna序列互补序列的mrna的作用。然而,sirna还可结合并剪切非目标mrna,其不与sirna的整个序列互补,而是仅与sirna序列的一部分互补。该现象被称为脱靶效应。
6.为了克服上述基于sirna的抗癌剂的技术缺陷,针对使用微小rna(以下称为“mirna”)作为治疗剂的研究正在进行(agostini,m.和knight,r.a.mir
‑
34:from bench to bedside.oncotarget 5,872
‑
81,2014.,van rooij,e.,purcell,a.l.和levin,a.a.developing micrornatherapeutics.circulation research 110,496
‑
507,2012.,burnett,j.c.和rossi,j.j.rna
‑
based therapeutics:current progress and future prospects.chem biol 19,60
‑
71,2012.,dangwal,s.和thum,t.microrna therapeutics in cardiovascular disease models.annu rev pharmacol toxicol 54,185
‑
203,2014.)。mirna是由16至27个核苷酸组成的rna,并且被分类为对抗翻译成蛋白质的信使rna(mrna)的蛋白质非编码rna(carthew,r.w.和sontheimer,e.j.origins and mechanisms of mirnas and sirnas.cell 136,642
‑
55,2009.,macfarlane,l.
‑
a.和murphy,p.r.microrna:biogenesis,function and role in cancer.current genomics 11,537
‑
561,2010.,bartel,d.p.micrornas:target recognition and regulatory functions.cell 136,215
‑
33,2009.)。mirna在较高等动物和植物细胞的基因组中发现,并且已知其发挥调节细胞新陈代谢和包括细胞产生、生长、分化和死亡的功能的重要作用。直到现在,在人类基因组中已经发现大约2000种mirna,并且相当数量的mirna的功能尚不清楚。
7.mirna通过称为pol ii的rna聚合酶从基因组转录到rna中,并且mirna的初始长度各种各样而不特定(carthew,r.w.和sontheimer,e.j.origins and mechanisms of mirnas and sirnas.cell 136,642
‑
55,2009.,brodersen,p.和voinnet,o.revisiting the principles of microrna target recognition and mode of action.nat rev mol cell biol 10,141
‑
148,2009)。这可归因于基因组中mirna的位置多样性。这是因为mirna以各种方式产生,其包括其中位于内含子(mrna非编码区)的mirna在同一时间点被转录为mrna产物的情况,以及其中位于基因组的基因间隔区的mirna被单独转录的情况(malone,c.d.&hannon,g.j.small rnas as guardians of the genome.cell 136,656
‑
68,2009.)。在上述初始阶段产生的mirna被称为初级微小rna(mir)。初级mir通过例如称为核内包涵体的核糖核酸酶(rnase)加工为前体mir(前体mirna,或前mir)(bartel,d.p.micrornas:target recognition and regulatory functions.cell 136,215
‑
33,2009)。前mir具有rna发卡结构并且由约70
‑
80个核苷酸组成。细胞核内的前mir通过输出蛋白等从细胞核输送到细胞溶质中,并且进一步通过另一种被称为dicer的核糖核酸酶在细胞溶质中进一步加工,从而产生由16
‑
27个核苷酸组成的双链成熟微小rna(在下文中,没有修饰语描述的mir指成熟的mir)。选择双链mir的一条rna链,通过结合核糖核蛋白复合体risc激活,并且基于mir的序列结合目标mrna。
8.通常,基于这些区域是否参与蛋白质编码,mrna大部分可被分为3个区域:含有蛋
白质编码翻译信息的编码区,和没有蛋白质编码信息的5'
‑
utr(非翻译区)和3'
‑
utr。尽管与其具有互补序列的诱导目标mrna剪切的sirna与mrna的5'
‑
utr、3'
‑
utr和编码区无关,mir主要结合3'
‑
utr(carthew,r.w.和sontheimer,e.j.origins and mechanisms of mirnas and sirnas.cell 136,642
‑
55,2009.,bartel,d.p.micrornas:target recognition and regulatory functions.cell 136,215
‑
33,2009.)。
9.除了结合mrna的位置不同之外,sirna和mirna之间的特征差异是sirna主要结合具有与整个sirna序列互补的序列的mrna,而mirna主要通过位于距mirna的5'端2
‑
8个核苷酸并且具有有限长度的种子区序列识别目标mrna。因此,甚至当整个mirna序列与目标序列的序列不完全互补并且含有非互补序列部分的时候,mirna的活性不受非互补序列部分影响(bartel,d.p.micrornas:target recognition and regulatory functions.cell 136,215
‑
33,2009)。因为种子区的长度为6
‑
8个核苷酸,所以具有各种mrna,其3'utr具有与种子区互补的序列,并且针对该区域,多种mrna可同时使用一种mirna控制。mirna的这种性质能够使mirna在控制许多细胞生理学方面,其包括细胞分裂、生长、分化和死亡,用作有效调节剂。此外,mirna用作调节剂在实现有效抗癌作用中提供了优势。这是因为mirna可同时抑制一些致癌基因的表达,而sirna旨在抑制单个基因的表达。
10.许多mrna的3'utr含有一个或多个mirna可结合的部分。根据生物信息学计算,已知所有mrna中约30%由mirna调节。
11.从mirna作为信号通路中的主要调节剂的事实可见,mirna在包括癌症的重大疾病中起重要作用(macfarlane,l.
‑
a.和murphy,p.r.microrna:biogenesis,function and role in cancer.current genomics 11,537
‑
561.2010.,malone,c.d.和hannon,g.j.small rnas as guardians of the genome.cell 136,656
‑
68.2009.,nicoloso,m.s.,spizzo,r.,shimizu,m.,rossi,s.和calin,g.a.micrornas
‑‑
the micro steering wheel of tumour metastases.nat rev cancer 9,293
‑
302.2009.,landi,d.,gemignani,f.和landi,s.role of variations within microrna
‑
binding sites in cancer.mutagenesis 27,205
‑
10.2012.)。事实上,一些研究表明,癌细胞中mirna的表达模式与正常细胞中mirnas的表达模式大不相同。此外,已知mirna的表达模式根据癌症发生的原始器官大不相同。具体地,各种癌症,包括肺癌、肝癌、皮肤癌和血癌,根据原始器官显示出特有的mirna表达模式,其表示mirna在癌症生物学中发挥重要作用。具体地,已知癌细胞中mirna的表达水平通常低于它们在正常细胞中的表达水平。
12.基于mirna在癌症中的深度联系,近来已经尝试使用mirna作为抗癌治疗剂。例如,mirna,称为“mir
‑
34a”,正处于临床试验中,以证实其抑制癌细胞增殖和诱导癌细胞凋亡的能力(wiggins,j.f.等development of a lung cancer therapeutic based on the tumor suppressor microrna
‑
34.cancer res 70,5923
‑
30.2010.,bader,a.g.等mir
‑
34regulated genes and pathways as targets for therapeutic intervention.google patents,2009.,hermeking,h.the mir
‑
34family in cancer and apoptosis.cell death differ 17,193
‑
9.2010.,chang,t.c.等transactivation of mir
‑
34a by p53 broadly influences gene expression and promotes apoptosis.mol cell 26,745
‑
52.2007.)。
13.因此,本发明的发明人已经做出广泛努力以鉴定mirna,其具有比正处于临床试验的mir
‑
34a更好的抑制癌细胞增殖和诱导癌细胞凋亡的能力。因此,本发明的发明人已经鉴
定出具有优异抗癌功效的mir
‑
3670、mir
‑
8078和mir
‑
4477a,并且已经发现这些mirna通过有效抑制一些被称为致癌基因的基因的表达展现出抗癌效果,从而完成本发明。
技术实现要素:
14.技术问题
15.本发明的目的是鉴定具有优异的抑制癌细胞增殖和诱导癌细胞凋亡的能力的mirna,并且提供用于癌症治疗的药物组合物,其包含mirna作为活性成分。
16.技术方案
17.为了实现上述目的,本发明提供了用于治疗癌症的药物组合物,其包含一种或多种选自mir
‑
3670、mir
‑
8078和mir
‑
4477a的mirna作为活性成分。
18.附图简述
19.图1显示测量mir
‑
34a、mir
‑
100和mir
‑
125b的蛋白质表达抑制能力的结果。为了测量用于构建筛选文库的mirna是否具有活性,mir
‑
34a、mir
‑
100和mir
‑
125b通常选自整个筛选文库,并且将已知被这些mirna沉默的每个mrna3'utr插入到荧光素酶表达载体的3'utr中,并且测量蛋白质表达抑制能力。
20.图2显示通过使用由约1700种mirna组成的筛选文库处理肺癌nci
‑
h460细胞,使用刃天青(resazurin)试剂量化细胞的生长,以及将量化结果表示为相对生长值而获得的结果。
21.图3显示通过选择约50种对nci
‑
h460细胞系显示出优异功效的mirna,以及使用wst
‑
1试剂测量所选mirna的相对癌细胞生长抑制能力而获得的结果。
22.图4显示通过将mir
‑
34a、mir
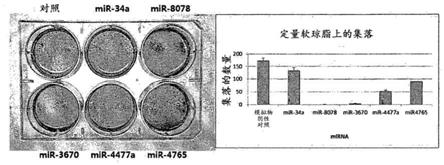
‑
3670、mir
‑
8078和mir
‑
4477a各自转染到肺癌细胞系中,使用fitc
‑
标记的annexin v染色细胞,以及通过流式细胞计数器分析染色的细胞以确定mirna对肺癌细胞系的凋亡效应而获得的结果。
23.图5显示通过将每个mirna转染到肺癌细胞系中,在软琼脂上培养细胞2周,以及测量mirna对抗集肺癌细胞系的集落形成能力的效果而获得的结果。
24.图6显示通过使用mirna目标预测软件选择目标候选物,以及将靶向所选候选物的sirna的细胞凋亡能力表示为z
‑
值而获得的结果。
25.图7显示进行qpcr以分析通过mirna抑制图6所示的基因表达程度的结果。
26.图8显示通过如图6所示将基因的3'utr克隆到荧光素酶的3'utr中,以及测量通过mirna抑制荧光素酶蛋白质的表达程度而获得的结果。
具体实施方式
27.除非另有定义,否则本文所使用的所有技术和科学术语具有与本发明所属领域普通技术人员通常理解的相同的含义。一般而言,本文所使用的命名法式本领域公知且常用的那些。
28.在本发明中,已经尝试鉴定具有比已知具有抗癌效果的mir
‑
34a更好功效的mirna,并评价所鉴定的mirna的抗癌效果。
29.在本发明中,合成由约1700种mirna组成的筛选文库(表1)。nci
‑
h460(肺癌细胞系)使用筛选文库处理,并且测量mirna抑制癌细胞生长的能力。因此,鉴定展现出比mir
‑
34a更好功效并且具有以下核苷酸序列的mir
‑
3670、mir
‑
8078和mir
‑
4477a(表3)。发现所鉴定的mirna具有优异的抗癌效果(图4至8)。
30.因此,本发明针对用于治疗癌症的药物组合物,其包含一种或多种选自mir
‑
3670、mir
‑
8078和mir
‑
4477a的mirna作为活性成分。
31.在本发明中,mir
‑
3670可包含具有核苷酸序列seq id no:35和seq id no:36;或seq id no:35和seq id no:67的双链rna作为活性成分。
32.在本发明中,mir
‑
4477a可包含具有核苷酸序列seq id no:43和seq id no:44;或seq id no:43和seq id no:68的双链rna作为活性成分。
33.在本发明中,mir
‑
8078可包含具有核苷酸序列seq id no:65和seq id no:66;或seq id no:65和seq id no:69的双链rna作为活性成分。
34.另一方面,本发明的mir
‑
3670模板链可由seq id no:35表示。
35.5'
‑
agagcucacagcuguccuucucua
‑
3'(mimat0018093.seq id no:35)
36.虽然在体内有活性的mirna是单链的,但是它应当以双链形式和与其具有相似长度的核苷酸序列一起递送到细胞中,以便它结合于risc。在此,结合序列的反义序列与活性序列互补。互补序列可以是完美互补序列或内源性序列。位于每个双链序列的3'末端或一个链的3'末端的核苷酸不可能与反义序列的核苷酸结合,并且所述核苷酸被称为3'
‑
悬垂(overhang)。双链序列可包含3'
‑
悬垂。
37.换言之,mir
‑
3670的完美互补序列可由seq id no:36表示。
38.5'
‑
gagaaggacagcugugagcucuuu
‑
3'(seq id no:36)
39.此外,mir
‑
3670的内源性互补序列可由seq id no:67表示。
40.5'
‑
gacugguauagcugcuuuuggagccuca
‑
3'(seq id no:67)
41.如在上述
背景技术:
部分所描述的,从mirna的活性序列的第二个核苷酸开始计数的第8
‑
9个核苷酸对应的种子序列是活性的主要因素。包含种子区域的长双链序列可用于制备双链rna。
42.与mir
‑
3670相似,mir
‑
4477a和mir
‑
8078的活性序列和用于与其形成双链序列的互补序列可由以下序列表示。这些双链序列可包含如上所述的3'
‑
悬垂以形成长双链序列,并且还可包含种子区域以形成长双链序列。
43.mir
‑
4477a
44.5'
‑
cuauuaaggacauuugugauuc
‑
3'(mimat0019004 seq id no:43)
45.mir
‑
4477a的完美互补序列:
46.5'
‑
aucacaaauguccuuaauaguu
‑
3'(seq id no:44)
47.mir
‑
4477a的内源性互补序列:
48.5'
‑
aucacaaauguccuuaauggca
‑
3'(seq id no:68)
49.mir
‑
8078
50.5'
‑
ggucuaggcccggugagagacuc(mimat0031005,seq id no:65)
51.mir
‑
8078的完美互补序列:
52.5'
‑
gucucucaccgggccuagaccuu(seq id no:66)
53.mir
‑
8078的内源性互补序列:
54.5'
‑
cuccaccgggcugaccggccug
‑
3'(seq id no:69)
55.在本发明中,发现了通过文库筛选鉴定的mirna通过控制通常已知在癌细胞的诱导、产生和生长中起主要作用的基因而展现出抗癌效果。mirna的特征在于一种mirna可同时控制一些mrna在同一时间的表达。在本发明中也发现了这种性质,并且用于开发新的基于低聚物的抗癌药物。
56.发现了本发明的mir
‑
3670同时抑制cbx4、nras、casr、txlna、snip1、hnf1a、fzd4、trib1、adma19和ckap5的表达,mir
‑
8078抑制greb1、hectd3和ripk4的表达,以及mir
‑
4477a同时抑制stil、kif11、akap11和fam120a的表达(图6)。
57.已知通过本发明的mirna抑制的目标基因具有以下功能。
58.已知cbx4(多梳色素框4)涉及肿瘤血管生成并刺激肿瘤转移,并且nras在肿瘤生长和细胞分化中起重要作用(orouji,e.等map kinase pathway gene copy alterations in nras/braf wild
‑
type advanced melanoma.int j cancer(2015);zheng,c.等microrna
‑
195functions as a tumor suppressor by inhibiting cbx4 in hepatocellular carcinoma.oncol rep 33,1115
‑
22(2015);jiao,h.k.等prognostic significance of cbx4 expression and its beneficial effect for transarterial chemoembolization in hepatocellular carcinoma.cell death dis 6,e1689(2015);ohashi,k.等characteristics of lung cancers harboring nras mutations.clin cancer res 19,2584
‑
91(2013))。
59.casr被发现在肿瘤中过表达,并且是肿瘤转移所必需的,并且已知txlna与肿瘤生长和转移相关。报道的临床结果表明显示txlna高表达水平的患者组生存率低(mashidori,t.,shirataki,h.,kamai,t.,nakamura,f.&yoshida,k.increased alpha
‑
taxilin protein expression is associated with the metastatic and invasive potential of renal cell cancer.biomed res 32,103
‑
10(2011);tennakoon,s.,aggarwal,a.&kallay,e.the calcium
‑
sensing receptor and the hallmarks of cancer.biochim biophys acta(2015);ohtomo,n.等expression of alpha
‑
taxilin in hepatocellular carcinoma correlates with growth activity and malignant potential of the tumor.int j oncol 37,1417
‑
23(2010))。
60.称为转录辅激活因子的snip1刺激细胞生长和分化所必需的细胞周期蛋白(cyclin)d1的表达。已知显示snip1高表达水平的患者组预后不良。此外,已知snip1结合起细胞增殖主要调节子作用的c
‑
myc,从而刺激肿瘤生长(li,q.等snip1:a new activator of hse signaling pathway.mol cell biochem 362,1
‑
6(2012);fujii,m.等snip1 is a candidate modifier of the transcriptional activity of c
‑
myc on e box
‑
dependent target genes.mol cell 24,771
‑
83(2006);roche,k.c.,rocha,s.,bracken,c.p.和perkins,n.d.regulation of atr
‑
dependent pathways by the fha domain containing protein snip1.oncogene 26,4523
‑
30(2007);jeon,h.s.等high expression of snip1correlates with poor prognosis in non
‑
small cell lung cancer and snip1 interferes with the recruitment of hdac1 to rb in vitro.lung cancer 82,24
‑
30(2013);liang,x.等hypoxia
‑
inducible factor
‑
1alpha,in association with twist2 and snip1,is a critical prognostic factor in patients with tongue squamous cell carcinoma.oral oncol 47,92
‑
7(2011))。
61.hnf1a和fzd4是wnt信号传导通路的组分,其深入涉及肿瘤生长和存活。wnt信号传导通路在肿瘤生物学中已经被深入研究,并且广泛已知其重要性。已知trib1在抑制肿瘤细胞生长、转移和凋亡,以及调节作为肿瘤生长的主要信号传导通路的mapk信号通路中发挥作用(pecina
‑
slaus,n.等wnt signaling transcription factors tcf
‑
1 and lef
‑
1are upregulated inmalignant astrocytic brain tumors.histol histopathol 29,1557
‑
64(2014);ueno,k.等tumor suppressor microrna
‑
493decreases cell motility and migration ability in human bladder cancer cells by downregulating rhoc and fzd4.mol cancer ther 11,244
‑
53(2012);lin,z.y.等microrna
‑
224inhibits progression of human prostate cancer by downregulating trib1.int j cancer 135,541
‑
50(2014);soubeyrand,s.,naing,t.,martinuk,a.和mcpherson,r.erk1/2regulates hepatocyte trib1 in response to mitochondrial dysfunction.biochim biophys acta 1833,3405
‑
14(2013))。
62.adam19是分布在细胞膜中的蛋白质,并且已知进行各种生物学功能,其包括细胞
‑
细胞相互作用和细胞
‑
细胞外基质相互作用。如在肿瘤生物学中已知的,adam19与肿瘤的生长和转移深入相关。在本发明中发现通过基因功能筛选发现在肿瘤存活中发挥重要作用的ckap5基因由mirna控制。greb1与响应激素的组织或肿瘤的信号传导通路有关,并且已知其在各种肿瘤细胞中过度表达以刺激细胞的生长(zhang,q.等role of microrna
‑
30ctargeting adam19 in colorectal cancer.plos one 10,e0120698(2015);shan,n.,shen,l.,wang,j.,he,d.&duan,c.mir
‑
153inhibits migration and invasion of human non
‑
small
‑
cell lung cancer by targetin;g adam19.biochem biophys res commun 456,385
‑
91(2015);martens
‑
de kemp,s.r.等functional genetic screens identify genes essential for tumor cell survival in head and neck and lung cancer.clin cancer res 19,1994
‑
2003(2013);rae,j.m.等greb1 is a novel androgen
‑
regulated gene required for prostate cancer growth.prostate 66,886
‑
94(2006);zhang,l.等development of transcriptomic biomarker signature in human saliva to detect lung cancer.cell mol life sci 69,3341
‑
50(2012);laviolette,l.a.,hodgkinson,k.m.,minhas,n.,perez
‑
iratxeta,c.和vanderhyden,b.c.17beta
‑
estradiol upregulates greb1 and accelerates ovarian tumor progression in vivo.int j cancer 135,1072
‑
84(2014))。
63.hectd3是e3泛素连接酶,并且已知其将多聚泛素附接于引发凋亡的半胱天冬酶
‑
8以诱导胱天冬酶
‑
8的降解,从而抑制肿瘤细胞凋亡。并且,已知hectd3稳定malt1蛋白从而增加对顺铂抗癌药物的抗性。ripk4是受体相互作用蛋白激酶4,并且已知其诱导细胞生长信号传导因子β
‑
连环蛋白的积累并激活wnt信号传导通路。已知人工移除ripk4在肿瘤动物模型中可抑制肿瘤生长(li,y.等the hectd3 e3 ubiquitin ligase facilitates cancer cell survival by promoting k63
‑
linked polyubiquitination of caspase
‑
8.cell death dis 4,e935(2013);li,y.等the hectd3 e3 ubiquitin ligase suppresses cisplatin
‑
induced apoptosis via stabilizing malt1.neoplasia 15,39
‑
48(2013);huang,x.等phosphorylation of dishevelled by protein kinase ripk4 regulates wnt signaling.science 339,1441
‑
5(2013))。
64.已知stil基因,是细胞周期中g2期到m期过渡的必要因子,在各种细胞中高度表达,并且对肿瘤生长和存活是必需的。已知kif11对肿瘤细胞生长和转移也是必需的,并且抑制kif11活性可抑制肿瘤生长(erez,a.等sil overexpression in lung cancer characterizes tumors with increased mitotic activity.oncogene 23,5371
‑
7(2004);erez,a.等the sil gene is essential for mitotic entry and survival of cancer cells.cancer res 67,4022
‑
7(2007);tang,y.,orth,j.d.,xie,t.和mitchison,t.j.rapid induction of apoptosis during kinesin
‑
5 inhibitor
‑
induced mitotic arrest in hl60 cells.cancer lett 310,15
‑
24(2011);venere,m.等the mitotic kinesin kif11 is a driver of invasion,proliferation,and self
‑
renewal in glioblastoma.sci transl med 7,304ra143(2015))。
65.结合蛋白激酶a(pka)以增加pka活性的akap11也结合gsk
‑
3β以促进pka介导的gsk
‑
3的磷酸化。磷酸化的gsk
‑
3β失去其活性,其是在细胞中被认为是诱导肿瘤生长的生长刺激信号的主要机制。将肿瘤细胞暴露于各种应激条件下,诸如酸性条件、缺氧条件等,并且即使在如此恶劣的条件下也保持肿瘤细胞的死亡被抑制的机制。例如,已知fam120a是rna结合蛋白激活激酶,诸如src,抑制细胞凋亡并增加药物抗性(logue,j.s.等akap220protein organizes signaling elements that impact cell migration.j biol chem 286,39269
‑
81(2011);whiting,j.l.等protein kinase aopposes the phosphorylation
‑
dependent recruitment of glycogen synthase kinase 3beta to a
‑
kinase anchoring protein 220.j biol chem 290,19445
‑
57(2015);tanji,c.等a
‑
kinase anchoring protein akap220 binds to glycogen synthase kinase
‑
3beta(gsk
‑
3beta)and mediates protein kinase a
‑
dependent inhibition of gsk
‑
3beta.j biol chem 277,36955
‑
61(2002);tanaka,m.等anovel rna
‑
binding protein,ossa/c9orf10,regulates activity of src kinases to protect cells from oxidative stress
‑
induced apoptosis.mol cell biol 29,402
‑
13(2009);bartolome,r.a.等il13 receptor alpha2 signaling requires a scaffold protein,fam120a,to activate the fak and pi3k pathways in colon cancer metastasis.cancer res 75,2434
‑
44(2015))。
66.在本发明中,显示当使用sirna减少上述基因的细胞内含量时,细胞生长被抑制,如使用mir
‑
3670、mir4477a或mir
‑
8078时(图6)。此外,qpcr结果表明当mir
‑
3670、mir
‑
4477a或mir
‑
8078被递送至肺癌细胞中时,基因的mrna表达水平降低(图7)。此外,荧光素酶测定结果表明上述基因是通过mir
‑
3670、mir
‑
4477a或mir
‑
8078直接靶向的基因(图8)。这表明mir
‑
3670、mir
‑
4477a或mir
‑
8078直接并同时抑制在肿瘤细胞生长和存活中重要的各种基因的表达,从而诱导肿瘤细胞凋亡。
67.在本发明中,癌症可以是一种或多种选自下组的癌症:原发性癌症,诸如肺癌、肝癌、胃癌、结肠直肠癌、胰腺癌、胆囊和胆管癌、乳腺癌、白血病、食道癌、非霍奇金淋巴瘤、甲状腺癌、子宫颈癌或皮肤癌;由原发性癌症起源部位转移到其他器官引起的转移性癌;以及通过促进异常过度的细胞分裂而造成的赘生性细胞疾病,但不限于此。
68.mirna的序列(根据本发明,其可用作用于治疗癌症的药物组合物的活性成分)是来源于人基因组的序列,但是可以是从其他动物基因组获得的mirna序列,而不将源于mirna的基因组局限于人类基因组。
69.在本发明中,mirna可用作各种产生生物学等效效果的mirna模拟物。例如,可使用包含含有相同种子区域的mirna序列的经修饰的mirna。在此,可减少seq id no:1或seq id no:2的长度,并且还可使用由15个核苷酸组成的长度短的mirna模拟物。
70.在本发明中,mirna的mirna模拟物可部分地包含硫代磷酸酯结构,其中,rna磷酸酯骨架结构取代有另一种元素诸如硫。此外,还可使用通过使用dna、pna(肽核酸)或lna(锁核酸)分子全部或部分取代rna而获得的那些。此外,也可使用通过使用各种功能性结构取代rna糖的2’羟基基团而获得那些。所述修饰包括,但不限于,甲基化、甲氧基化、氟化等。
71.在本发明中,mirna不限于成熟的mirna和由此衍生的mirna模拟物的双链rna,但是可以mirna前体的形式使用。mirna前体还可通过取代上述rna磷酸酯骨架结构,使用dna、pna或lna分子全部或部分取代rna核酸,或修饰rna糖的2’羟基基团而获得。
72.在本发明中,mirna可以mirna前体或初级mirna(pri
‑
mirna)的形式使用,并且其可通过化学方法合成或以质粒的形式递送至细胞以便表达。
73.将mirna递送到在培养皿中培养的细胞的方法的实例,其可用于本发明,包括使用mirna和阳离子脂质的混合物的方法,通过电刺激将mirna递送到细胞的方法,和使用病毒的方法,但不限于此。
74.用于治疗癌症的药物组合物,其包含本发明的mirna作为活性成分,可进一步包含药学上可接受的载体,并且可与载体一起配制。
75.如本文所使用的术语“药学上可接受的载体”是指在不刺激有机体的情况下不损害所施用化合物的生物活性和特征的载体或稀释剂。作为配制为液体溶液的组合物中药学上可接受的载体,使用无菌和生物相容性载体。药学上可接受的载体可以是生理盐水、无菌水、林格氏溶液、缓冲盐水、白蛋白注射溶液、葡萄糖溶液、麦芽糊精溶液、甘油、乙醇或它们当中两种或更多种的混合物。此外,必要时,本发明的组合物包含其他常规添加剂,其包括抗氧化剂、缓冲剂和抑菌剂。此外,本发明的组合物可借助稀释剂、分散剂、表面活性剂、粘合剂和润滑剂,配制为可注射的形式,诸如水溶液、混悬液或乳液。此外,本发明的组合物可以丸剂、胶囊、颗粒或片剂的形式配制。
76.预防或治疗癌症的组合物,其包含本发明的mirna和药学上可接受的载体,可用作任一包含其作为活性成分的制剂,并可制备为口服或肠胃外制剂。本发明的药物制剂包括适合于口服、经直肠、经鼻、局部(包括经颊和舌下)、皮下、经阴道或肠胃外(包括肌内、皮下和静脉内)施用的那些或适合于通过吸入或吹入施用的那些。
77.包含本发明的组合物作为活性成分的口服制剂的实例包括片剂、锭剂(troches)、糖锭(lozenges)、水性混悬液或乳化混悬液、粉剂、颗粒剂、乳剂、硬胶囊或软胶囊、糖浆或酏剂。制剂诸如片剂或胶囊可包含粘合剂,诸如乳糖、蔗糖、山梨醇、甘露醇、淀粉、支链淀粉、纤维素或明胶;赋形剂,诸如磷酸二钙;崩解剂,诸如玉米淀粉或甘薯淀粉;以及润滑剂,诸如硬脂酸镁、硬脂酸钙、硬脂酰富马酸钠或聚乙二醇蜡。除上述提及的物质之外,胶囊制剂还可包含液相载体,诸如脂肪油。
78.包含本发明的组合物作为活性成分的肠胃外制剂可包括皮下、静脉或肌内注射的可注射形式,栓剂或经由呼吸器官可吸入的喷雾,诸如喷雾剂。可注射的制剂可通过使本发明的组合物与稳定剂或缓冲液在水中混合制备溶液或混悬液来制备,并将溶液或悬浮液加载到安瓿或小瓶中以制备单位剂型。栓剂制剂包括栓剂,或例如含有常规栓剂基质的保留
灌肠,诸如可可脂或其他甘油酯。对于喷雾制剂,诸如喷雾剂,用于喷洒水分散浓缩物或湿粉末的抛射剂可与添加剂组合使用。
79.实施例
80.在下文中,将参考实施例进一步详细描述本发明。对本领域的普通技术人员显而易见的是,这些实施例仅用于说明目的,并且不被解释为限制本发明的范围。
81.实施例1:构建mirna筛选文库
82.基于茎环结构,将从mirna数据库mirbase(www.mirbase.org)提供的21版人mirna序列(由大约1700种mirna组成的筛选文库)通过用于常规低聚物合成的固相合成法合成为双链mirna序列。合成的mirna的每个链通过使用c18树脂反相分离而纯化。所有合成的mirna链通过maldi
‑
tof ms质谱法分析以确定是否合成所需序列。为了合成双链mirna,在95℃在盐的存在下加热互补链2分钟,然后缓慢冷却以制备双链。
83.表1:mirna序列
84.85.86.87.88.89.90.91.92.93.94.95.96.97.98.99.100.101.102.103.104.105.106.107.108.109.110.111.112.113.114.115.116.117.118.119.120.121.122.123.124.125.[0126][0127]
实施例2:测量mirna文库样本的活性
[0128]
为了确定在实施例中合成的双链mirna是否将有活性,从约1700种mirna中选择mir
‑
34a、mir
‑
100和mir
‑
125b。选择所选的mirna,因为具有许多针对目标mrna的研究,其功能和表达被控制并且在结合mrna3'utr的区域。已知表达通过mir
‑
34a、mir
‑
100和mir
‑
125b各自控制的bcl2、mtor和lin28b mrna各自的3'utr区域,被替换成萤火虫荧光素酶载体的3'utr,从而构建对应于每种mirna的载体。每种载体和mir对照,或者对应于每种载体的mir
‑
34a、mir
‑
100或mir
‑
125b,通过使用作为细胞内低聚物递送试剂的lipofectamine 2000(lipofectamine 2000,invitrogen)被共同转染到hek
‑
293t细胞中(一式三份样品的制备),并且将细胞在37℃和5%(v/v)co2的条件下培养24小时。荧光素酶活性通过照度计(thermo scientific)测量以确定所合成的mirna的活性(图1)。
[0129]
实施例3:用于鉴定抑制肺癌细胞增殖的微小核糖核酸(microrna)的筛选
[0130]
将3,000~10,000个nci
‑
h460细胞分散在96孔板的各孔中并在37℃和5%(v/v)co2的条件下培养24小时,然后选自mirna文库的每种mirna通过使用rnaimax试剂(invitrogen)转染到细胞中达到100nm的最终浓度。因为每种mirna被转染三次,因此,每种mirna制备三块96孔板。在与上述相同的条件下进一步培养细胞24小时,然后使用荧光计(tecan)测量通过添加刃天青试剂(promega)产生的荧光值。为了比较性评价mirna的细胞生长抑制能力,计算针对96孔板测量的96个值的平均值和标准偏差,并且使用以下等式计算针对每个孔从平均值测量的值的标准偏差倍数(z值):
[0131][0132]
其中,x
i
是在每个孔中测量的值,μ是针对平板的所有孔测量的值的平均值。每个
孔的标准偏差倍数z
i
是针对一式三份平板获得的值的平均值,并且选择显示z
i
值小于
‑
2的约第一候选mirna(图2和表2)。
[0133]
表2:第一次选择的mirna序列
[0134]
[0135]
[0136][0137]
实施例4:鉴定抑制肺癌细胞增殖的mirna的第二次筛选
[0138]
使用第一次筛选中获得的约50个mirna候选物,使用增加的测量精确度进行第二次筛选。除了使用wst
‑
1试剂(roche)替代刃天青试剂来测量细胞的增殖潜力之外,在与第一次筛选相同的条件下进行实验。wst
‑
1相对于刃天青的优点在于信号强度可更量化地测量。通过测量每种mirna的细胞增殖抑制能力获得的值表示为与对照相关的值,并且结果示于图3。mir
‑
34a用作阳性对照。
[0139]
实施例5:分析诱导细胞凋亡的能力
[0140]
用于筛选的方法是一种通过定量测量细胞数量而测量细胞增殖的相对抑制的方法。抑制细胞增殖的机制包括降低细胞分化速率的方法和诱导细胞凋亡的方法。为了分析本发明中发现的mirna抑制细胞增殖的机制,使用荧光激活细胞分选器(facs)分析细胞凋亡的程度。具体地,将细胞分散于6孔板中,并且通过使用rnaimax试剂将mirna转染到细胞中,然后在与上述相同的条件下培养细胞48小时。接着,使用fitc
‑
标记的annexin v处理细
胞并使用荧光激活细胞分选器(facs)进行分析(图4)。分析的结果表明使用mir
‑
3670、mir
‑
4477a和mir
‑
8078各自处理的细胞大部分死亡。这表明筛选中鉴定的mirna的癌细胞生长抑制效果由诱导凋亡引起(表3)。
[0141]
表3:最终选择的mirna序列
[0142]
seq id no:35hsa
‑
mir
‑
3670链1agagcucacagcuguccuucucuaseq id no:36hsa
‑
mir
‑
3670链2gagaaggacagcugugagcucuuuseq id no:67mir
‑
3670
‑
icicgacugguauagcugcuuuuggagccucaseq id no:43hsa
‑
mir
‑
4477a链1cuauuaaggacauuugugauucseq id no:44hsa
‑
mir
‑
4477a链2aucacaaauguccuuaauaguuseq id no:68mir
‑
4477a
‑
icicaucacaaauguccuuaauggcaseq id no:65hsa
‑
mir
‑
8078链1ggucuaggcccggugagagacucseq id no:66hsa
‑
mir
‑
8078链2gucucucaccgggccuagaccuuseq id no:69mir
‑
8078
‑
iciccuccaccgggcugaccggccug
[0143]
实施例6:在软琼脂上通过mirna抑制肿瘤细胞生长
[0144]
当使用软琼脂培养肿瘤细胞时,可测量肿瘤细胞的特征。正常细胞需要支撑物诸如用于生长的培养皿,而肿瘤细胞的特征在于,甚至在没有物理强大支撑诸如软琼脂的环境下生长。使用这种肿瘤特异性特征,检查软琼脂上的细胞集落形成能力。nci
‑
h460肺癌细胞各自使用对照mirna、mir
‑
34a、mir
‑
8078、mir
‑
3670、mir
‑
4477a和mir
‑
4765处理并培养24小时。接着,将细胞与软琼脂混合,并在6孔板中培养2周。使用结晶紫将细胞染色,并测量每个组的集落数量(图5)。因此,可见使用mir
‑
8078或mir
‑
3670处理的细胞形成很少的集落或不形成集落,而使用mir
‑
4477a处理的细胞显示出等于对照组的约30%的集落形成能力。
[0145]
实施例7:在敲低目标mrna时测量mirna的细胞增殖抑制作用
[0146]
表达由mirna控制的目标mrna具有与部分mirna序列的互补的序列。mirna的种子区域序列在抑制mrna中尤其重要,因为种子区域序列结合在此具有互补序列的mrna以抑制基因表达。然而,由于种子区域序列相对较短(8
‑
9个核苷酸),使用软件预测由mirna靶向的mrna。然而,已知甚至当使用软件时,仅一些预测的目标是重要目标。为了克服该缺点,使用sirna减少通过软件预测的目标基因的细胞内含量,然后确定细胞的生长是否被抑制,从而选择目标基因。为了预测由mirna靶向的目标mrna,通常用于本领域的targetscan用作目标预测软件,并且选择由mir
‑
3670、mir
‑
4477a或mir
‑
8078靶向的约600种基因。对于每种所选基因,合成3种sirna,并且使用合成的sirna,实验以与实施例3中描述的相同方式进行。具体地,将细胞分散在96孔板中,各自使用sirna处理,然后培养48小时,之后使用刃天青测量细胞增殖的能力。以与实施例3中描述的相同方式,从针对总共1800个样本(约600个基因
×
三种不同sirna)测量的值的平均值计算每个基因的z值。结果示于图6。
[0147]
实施例8:分析由mirna靶向的mrna
[0148]
mirna起抑制蛋白质从mrna产生的作用,并且起诱导由此靶向的大部分mrna降解的作用。因此,将mirna转染到细胞中,并且通过qpcr分析由mirna靶向的mrna的细胞内含量,以确定含量是否将降低,从而确定mirna是否将靶向mrna。为了确定实施例7中发现的目标基因的细胞内含量在将mirna递送到细胞内时是否将大量降低,将mir
‑
3670、mir
‑
4477a和mir
‑
8078各自转染到肺癌细胞中,然后将其培养48小时,之后从每个细胞中提取rna,并
且定量测量细胞中rna的含量(图7)。因此,可见预测由mir
‑
3670、mir
‑
4477a和mir
‑
8078各自靶向的mrna的含量显著降低。
[0149]
实施例9:通过荧光素酶测定检查目标mrna
[0150]
mirna结合目标mrna的3'utr(未翻译区)以抑制蛋白质从目标mrna产生。由于此原因,荧光素酶测定法通常用来直接测量mirna和目标mrna之间的关系。在targetscan软件下,提供含有mirna结合序列的3'utr序列。如以上实施例2中所述,将3'utr序列克隆到萤火虫荧光素酶中以构建载体。将所构建的载体和mirna共同转染到人类胚胎肾(hek)细胞中,并且测量载体的荧光素酶表达水平。为了校正转染效率,还转染了海肾荧光素酶(renilla luciferase)。在共同转染mirna、萤火虫荧光素酶和海肾荧光素酶后,将细胞培养48小时,并且通过照度计测量细胞的荧光素酶活性(图9)。因此,可见目标mrna通过相应的mirna直接控制。
[0151]
工业实用性
[0152]
如上所述,本发明的用于治疗癌症的药物组合物含有一种或多种选自mir
‑
3670、mir
‑
4477a和mir
‑
8078的mirna作为活性成分。与目前正处于临床试验以确定作为抗癌剂的适用性的mir
‑
34a和含有其他mirna作为活性成分的用于治疗癌症的其他药物组合物相比,本发明的药物组合物展现出改善的抗癌效果。因此,本发明的药物组合物可广泛地用作抗癌组合物。
[0153]
尽管已经参考具体特征详细描述了本发明,对本领域技术人员显而易见的是,本说明书仅用于优选的实施方案,并且不限制本发明的范围。因此,本发明的实质范围将由所附权利要求及其等同物来限定。
[0154]
综上所述,本技术包括但不限于以下各项:
[0155]
1.一种用于治疗癌症的药物组合物,其包含一种或多种选自mir
‑
3670、mir
‑
8078和mir
‑
4477a的mirna作为活性成分。
[0156]
2.项1的药物组合物,其中mir
‑
3670包含由核苷酸序列seq id no:35和seq id no:36;或seq id no:35和seq id no:67表示的双链rna作为活性成分。
[0157]
3.项1的药物组合物,其中mir
‑
4477a包含由核苷酸序列seq id no:43和seq id no:44;或seq id no:43和seq id no:68表示的双链rna作为活性成分。
[0158]
4.项1的药物组合物,其中mir
‑
8078包含由核苷酸序列seq id no:65和seq id no:66;或seq id no:65和seq id no:69表示的双链rna作为活性成分。
[0159]
5.项1的药物组合物,其中所述mirna诱导癌细胞凋亡以治疗癌症。
[0160]
6.项1的药物组合物,其中所述癌症是一种或多种选自下组的癌症:原发性癌症,诸如肺癌、肝癌、胃癌、结肠直肠癌、胰腺癌、胆囊和胆管癌、乳腺癌、白血病、食道癌、非霍奇金淋巴瘤、甲状腺癌、子宫颈癌或皮肤癌;由原发性癌症起源部位转移到其他器官引起的转移性癌;以及通过促进异常过度的细胞分裂而造成的赘生性细胞疾病。
[0161]
7.项1至5中任一项的药物组合物,其中所述mirna是mirna模拟物。
[0162]
8.项7的药物组合物,其中所述mirna模拟物选自下组:部分包含硫代磷酸酯结构的mirna模拟物,其中rna磷酸酯骨架结构取代有另一种元素诸如硫;其中rna完全或部分取代有dna、pna(肽核酸)或lna(锁核酸)分子的mirna模拟物;以及其中rna糖的2’羟基基团取代有各种功能性结构诸如甲基化、甲氧基化或氟化的mirna模拟物。
[0163]
9.项7的药物组合物,所述mirna模拟物由mirna前体以初级mirna或前体mirna的形式组成,或由mirna前体以质粒的形式组成。
[0164]
10.项9的药物组合物,其中所述mirna前体选自下组:部分包含硫代磷酸酯结构的mirna前体,其中rna磷酸酯骨架结构取代有另一种元素诸如硫;其中rna完全或部分取代有dna、pna(肽核酸)或lna(锁核酸)分子的mirna前体;以及其中rna糖的2’羟基基团取代有各种功能性结构诸如甲基化、甲氧基化或氟化的mirna前体。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。