1.本发明涉及药物新用途领域,尤其涉及可利霉素及其单体用于制备改善脂多糖和盲肠结扎穿孔诱导的败血症药物的用途。
背景技术:
2.败血症(septicemia)是指致病菌或条件致病菌侵入血循环,并在血中生长繁殖,产生毒素而发生的急性全身性感染。败血症患者面临的其中一个挑战是病原微生物的持续入侵。败血症相关病原体主要包括革兰氏阳性细菌,如金黄色葡萄球菌和肺炎链球菌,以及革兰氏阴性细菌大肠杆菌、克雷伯菌和铜绿假单胞菌。作为对病原体入侵的反应,身体可以通过模式识别受体(prrs)识别病原体相关分子模式(pamp),并通过toll样受体激活经典和非经典nf-κb途径。由于病原体的持续存在,免疫系统的持续激活可能导致体内免疫失衡,最终可能危及生命。因此,及时清除病原微生物尤为重要。液体净化已被批准作为控制败血症进展的治疗策略。败血症的病理生理学复杂,具有单一功能和靶点的免疫调节剂可能预后不佳。这些药物应该相互结合使用,但这可能会增加治疗的不确定性。此外,根据败血症和感染性休克的国际管理指南,强烈建议对早期败血症进行抗菌治疗。
3.可利霉素(ca),又称bitespiramycin和shengjimycin,是一种以异戊基螺旋霉素(isp)ⅰ、ⅱ和ⅲ为主要成分的16元大环内酯类抗生素。最近,研究表明ca也可以作为抗病毒和抗肿瘤剂发挥作用。cn113577087a公开了异戊酰螺旋霉素类化合物或其组合物在制备治疗脓毒症疾病药物中的应用,明确指出该脓毒症疾病由冠状病毒病诱导产生。但是并没有用于改善脂多糖(lps)和盲肠结扎穿孔(clp)诱导的小鼠败血症的研究。
技术实现要素:
4.本发明的目的在于提供一种改善脂多糖和盲肠结扎穿孔诱导的小鼠败血症的ca及其单体的新用途。
5.为实现上述目的,可利霉素及其单体用于制备改善脂多糖和盲肠结扎穿孔诱导的败血症药物的用途。
6.进一步,可利霉素及其单体通过干扰炎症信号传导来改善败血症症状。
7.进一步,可利霉素及其单体通过调节肠道微生物环境来改善败血症症状。
8.进一步,所述用途是指可利霉素及其单体具有抑制脂多糖诱发的炎症的作用。
9.进一步,所述用途是指可利霉素及其单体具有阻断nf-κb的免疫激活,体外通过调节nf-κb信号通路来减少lps诱导的炎症的作用。
10.进一步,所述用途是指可利霉素具有改善败血症小鼠肠道中益生菌和病原微生物的数量和丰度。
11.进一步,所述用途是指可利霉素及其单体具有调节脂多糖和盲肠结扎穿孔诱导的败血症患者肠道微生物环境的用途。
12.进一步,所述改善是指减少全身炎症反应、减轻肺和肝组织损伤以及抑制nf-κb信号通路。
13.进一步,所述单体是指可利霉素单体i,可利霉素单体iii。
14.本发明的实验验证了:ca不仅抑制lps诱导的巨噬细胞raw264.7和thp1细胞的炎症反应的表达,而且通过抑制lps和clp小鼠体内的炎症爆发来提高存活率。ca能有效改善lps和clp诱导的脓毒症小鼠的体温调节,这是治疗败血症的重要指标。值得注意的是,ca降低了lps和clp诱导的败血症小鼠外周血、肺和肝脏中il 6、il 1β和tnfα的表达,这表明ca治疗减轻了全身炎症,并对体内肝和肺具有保护作用。苏木精-伊红染色再次证实了ca对败血症小鼠肺和肝的保护作用。ca改变了肠道微生物群的丰度和多样性,包括减少病原体志贺菌和增加益生菌alloprevotella的比例。ca治疗改变了肠道微生物的代谢途径,增强了短链脂肪酸(scfas)的代谢通路,有利于缓解身体炎症。这些均证明ca对败血症诱导的炎症性疾病具有保护作用。
15.败血症的顽固性在于其复杂的发病机制,包括病原体入侵引发的炎症激活和持续炎症引起的免疫抑制。在败血症的早期阶段,模式识别受体(prrs),包括toll样受体、rig-i样受体、nod样受体和c型凝集素受体,触发细胞间信号级联反应,导致炎症。lps刺激的tlr4激活引起的nf-κb信号传导的激活是败血症早期炎症爆发的主要途径。在败血症早期阻断nf-κb通路有助于控制疾病进展。本发明还发现ca在体内外调节lps诱导的nf-κb信号传导,包括p-ikb-α和p-nf-κb,这表明ca通过干扰炎症信号传导来改善败血症。
16.免疫细胞表型的变化与败血症的发展密切相关。早期败血症患者的免疫细胞向炎症激活的方向发展。巨噬细胞和nk细胞可以激活tnf通路以应对病原体入侵,从而激活包括nf-κb通路在内的免疫反应,并通过级联反应诱导炎症。先前的研究表明,在败血症动物模型中,nk细胞的耗竭可降低全身炎症反应和体温过低,增强微生物清除率,恢复酸碱平衡,并提高存活率。然而,还需要更多的数据来证明ca对败血症中免疫细胞表达变化的影响。
17.肠道微生物群在维持肠道屏障功能和调节免疫细胞功能方面发挥着重要作用[63],这与败血症的进展有关。ca作为一种已上市的抗菌药物,可能在肠道菌群的组成中发挥重要作用,这需要进一步分析。在这项研究中,ca治疗有效地改变了肠道微生物环境,使其朝着减少炎症的方向发展,包括病原体志贺菌的减少和益生菌异普雷沃氏菌的增加,以及scfas代谢途径的增加,这共同缓解了败血症的发展。这些结果表明,ca在败血症小鼠中具有强大的肠道微生物调节功能。
[0018]
总之,ca在体外和体内都能有效抑制炎症的发展。ca治疗抑制了lps诱导的促炎因子(il 6、il 1β和tnfα)的表达和释放以及nf-κb通路的激活。此外,ca对lps和clp诱导的小鼠败血症均有良好的治疗作用;它减缓了炎症的发展,并在体内对肺部和肝脏起到保护作用。此外,ca治疗在改善败血症小鼠肠道微生物群多样性和调节肠道微生物群代谢途径(增强短链脂肪酸的代谢途径)方面发挥着关键作用。这些结果表明,ca在治疗败血症炎症发展方面具有治疗潜力。
附图说明
[0019]
图1是ca及其主要成分ispⅰ(r=h)、ispⅱ(r=coch3)和ispⅲ(r=coch2ch3)的化学结构式。
[0020]
图2是lps(1μg/ml)刺激8小时后测量的细胞中il 6、il1β和tnfα的mrna表达图。
[0021]
图3是lps(1μg/ml)刺激12小时后,测量细胞培养基中il6、il1β和tnfα的含量图,其中a是il6的含量,b是il1β的含量,c是tnfα的含量。
[0022]
图4是热图显示29个富集基因在各组(mock、ca、lps和ca lps)之间的差异表达图。
[0023]
图5是可利霉素通过介导nf-κb途径影响lps诱导的炎症的ikb-α免疫荧光结果显示图和定量图。
[0024]
图6是可利霉素通过介导nf-κb途径影响lps诱导的炎症的nf-κb免疫荧光结果显示图和定量图。
[0025]
图7是对p-ikb-α(f)和p-nf-κb(g)的蛋白水平进行定量,并在蛋白质印迹中绘制图。
[0026]
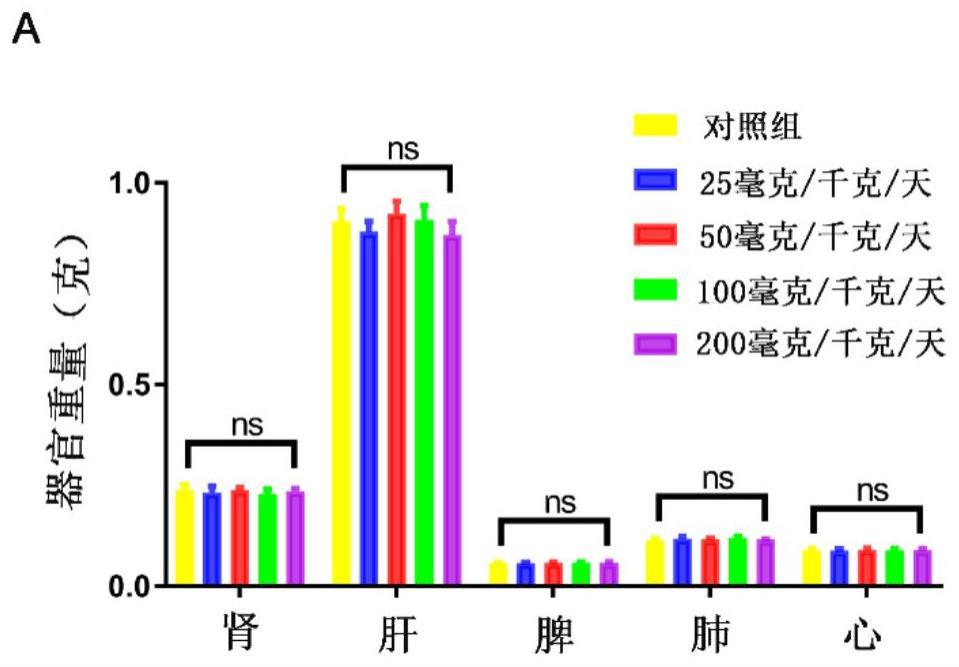
图8是连续灌胃给予不同浓度的ca(25-200mg/kg/天)6天对小鼠体重的影响结果图(n=5)。图9是不同浓度的ca(25-200mg/kg/天)灌胃对小鼠(n=5)心脏、肾脏、肺、肝脏和脾脏重量的影响结果图。
[0027]
图10是不同浓度ca的苏木精-伊红染色对肺和肝毒性的代表性图像以及半定量组织学评分(n=5)图。
[0028]
图11是可利霉素治疗lps诱导的小鼠炎症时,小鼠的治疗方案示意图。
[0029]
图12是可利霉素治疗lps诱导的小鼠炎症实验中,体温和存活率的变化图。
[0030]
图13是肝组织中il-6、il-1β和tnf-αmrna表达的变化(n=5)图)和肺部图。
[0031]
图14是肺组织中il-6、il-1β和tnf-αmrna表达的变化(n=5)图。
[0032]
图15是小鼠血清中il-6、il-1β和tnf-α水平的变化(n=5)图)。
[0033]
图16是蛋白质印迹显示ca对lps诱导的炎症小鼠肺中nf-κb信号通路蛋白的影响图以及定量图。
[0034]
图17是ca对clp小鼠炎症的影响实验中小鼠治疗方案示意图。
[0035]
图18是ca对clp小鼠炎症的影响实验中体温(n=5)和存活率(n=15)变化图。
[0036]
图19是ca对clp小鼠肝脏中il-6、il-1β和tnf-αmrna表达的影响(n=5)图。
[0037]
图20是ca对clp小鼠肺中il-6、il-1β和tnf-αmrna表达的影响(n=5)图。
[0038]
图21是ca对clp小鼠血清中il-6、il-1β和tnf-α水平的影响(n=5))。
[0039]
图22是从行为角度观察ca对clp小鼠的影响图。
[0040]
图23是ace指数、chao1指数、shannon指数和simpson指数图。
[0041]
图24是在属水平上不同类群之间肠道微生物的热图。
[0042]
图25是lef硒分析生成的分支图。
[0043]
图26是使用picrust功能基因预测肠道微生物群的clp和clp ca组之间的差异(数据库选择:kegg,p值阈值设置:0.05)图。
[0044]
图27是ca和阿司匹林对小鼠肺组织中il-6、il-1β和tnf-αmrna表达的抑制作用(n=5)图。
[0045]
图28是ca和阿司匹林降低小鼠血清中il-6、il-1β和tnf-α水平的作用(n=5)图。
具体实施方式
[0046]
下面详细描述本发明的实施例,所述实施例的示例在附图中示出,其中自始至终
相同或类似的标号表示相同或类似的元件或具有相同或类似功能的元件。下面通过参考附图描述的实施例是示例性的,旨在用于解释本发明,而不能理解为对本发明的限制。实施例中未注明具体技术或条件者,按照本领域内的文献所描述的技术或条件或者按照产品说明书进行。所用试剂或仪器未注明生产厂商者,均为可以通过市购获得的常规产品。
[0047]
本发明中可利霉素称为ca,ispⅰ称为ca01或者可利霉素单体1,ispⅲ称为ca03或者可利霉素单体3。均可商购到。
[0048]
实施例
[0049]
以下实验的对照:细胞实验用的是同体积溶解可利霉素用的溶剂dmso当对照,小鼠实验是使用同体积溶解可利霉素的溶剂(80%玉米油 20%聚乙二醇400)当对照。
[0050]
1.实验
[0051]
1.1细胞培养和脂多糖刺激
[0052]
小鼠巨噬细胞系(raw264.7和raw lucia
tm isg),购自invivogen(美国加利福尼亚州),并与10%(v/v)胎牛血清(gibco,grand island,美国)和1%(v/v)青霉素-链霉素(besalmedia,中国上海)在37℃、恒温和5%二氧化碳的湿润培养箱中培养。thp1细胞(invivogen,california,usa)在与小鼠巨噬细胞系相同的条件下培养,不同之处在于灭活的胎牛血清(56℃,30分钟)。raw264.7,raw露西亚
tm
利用isg和thp1细胞研究ca的体外抗炎作用。用ca预处理细胞,然后用lps(cell signaling technology,boston,usa)处理以诱导炎症。
[0053]
1.2.细胞毒性测试
[0054]
使用购自medchemexpress(美国新泽西州)的细胞计数试剂盒-8(cck-8)测定ca的毒性。首先,将细胞接种在96孔板中,然后用不同浓度的ca处理24小时。将cck-8溶液添加到每个孔中,并在37℃下孵育2小时。在450nm处测量光密度(od)。
[0055]
1.3.定量pcr测试
[0056]
qrt-pcr检测用于测量ca处理的lps诱导的mrna表达的变化。使用trizol(takara,shiga county,japan)提取总rna,并使用cdna合成试剂盒(yeasen,shanghai,china)进行逆转录。rt-qpcr由quantstudio 6flex(thermo,massachusetts,usa)使用sybr green mix(yeasen,shanghai,china)进行。用于扩增靶基因的pcr引物见表1:
[0057]
表1引物序列表
[0058]
[0059][0060]
1.4.蛋白印染分析
[0061]
使用ripa缓冲液(beyotime,中国)提取总蛋白,并使用bca蛋白测定试剂盒(beyotime,江苏,中国)测定蛋白浓度。将等量的蛋白质进行sds-page,然后转移到用甲醇活化的pvdf膜上。将pvdf膜用immobilon信号增强剂(sigma-aldrich,密苏里州,美国)封闭1小时,并与指示的抗体在4℃下孵育过夜。在tbst中进行三次洗涤,然后用二级抗体孵育,重复洗涤步骤,然后使用红外分子成像系统odyssey c1(中国香港基因有限公司)进行开发。产品信息如下:预存蛋白标记10-180kd(p1018,北京lablead biotech,中国),抗nf-κb小鼠mab(6956,细胞信号技术,波士顿,美国),抗磷酸化nf-κb小鼠mab,抗磷酸化ikbα兔单克隆抗体(ap0707,abclone technology,中国武汉),抗gapdh小鼠单克隆抗体(97166,cell signaling technology,美国波士顿),驴抗小鼠igg二级抗体(925-32212,美国利科尔),驴抗兔igg二级抗(926-32213,美国利戈尔)。
[0062]
1.5.免疫荧光测试
[0063]
细胞在玻璃底培养皿(nest biotechnology co.ltd.,wuxi,china)中培养,温度为37℃,含5%co2。药物治疗和刺激后,用4%多聚甲醛固定细胞10分钟,然后用triton-x100(beyotime,江苏,中国)透化15分钟,用5%bsa封闭1小时,并与1:200抗nf-κb(6956,cell signaling technology,boston,usa)或1:200抗ikb-α(a16929,abclone technology,wuhan,china)一级抗体在4℃下孵育过夜。在用1:1000alexa fluor 594山羊抗小鼠(thermo,massachusetts,usa)二级抗体孵育1小时后,用dapi(thermo)对细胞核进行染色2分钟。在上述每个步骤之间用pbs洗涤细胞。最后,在蔡司荧光显微镜(100
×
mirror,lsm780,carl zeiss,oberkochen,germany)下对细胞成像,并使用蔡司lsm 510软件进行分析。
[0064]
1.6.小鼠动物研究
[0065]
无特定病原体(spf)小鼠购自上海乌石动物有限公司(中国)。所有小鼠均在福建师范大学动物中心饲养,并在spf环境(50%湿度和24
±
1℃)中培养,光/暗周期为12小时,食物和水随意。所有动物实验均按照《实验动物护理与使用指南》进行,并经福建师范大学动物护理与利用委员会批准。将8只体重20
±
1g的10周龄雄性和雌性小鼠随机分为4组:对照组、ca组、败血症组和ca加败血症组。ca溶液是通过将其溶解在溶剂(80%玉米油 20%聚乙二醇400)中来制备的。从第一天到第三天,每天向小鼠灌胃100mg/kg的ca,并在第四天诱导败血症模型,如下所述。同时用等体积的溶剂灌胃给对照小鼠。此外,使用lps和clp小鼠模型来研究ca对败血症的治疗作用。小鼠腹膜内注射15mg/kg lps(cst,boston,usa)(溶于生理盐水)以诱导败血症。对照组腹膜内注射等量的生理盐水。为了产生clp诱导的败血症小鼠,用三溴乙醇(sigma-aldrich,密苏里州,美国)麻醉小鼠,并剃掉小鼠的腹部毛发并进行消毒。剖腹探查盲肠,用缝线结扎,并用18g针穿刺。轻轻挤出盲肠中的内容物,将盲肠与挤压的内容物一起放回腹腔,并将各层缝合回去[30]。随后,皮下注射1ml生理盐水进行复苏。对照组接受了假手术,除盲肠穿刺外,手术与clp组相同。
[0066]
1.7.组织病理学检查
[0067]
分离小鼠组织(肺和肝),用4%多聚甲醛固定,包埋在石蜡中,常规切片,并根据标准程序用苏木精和伊红染色。组织病理学变化的半定量分析:充血,1;水肿,2分;多形核白细胞和单核细胞浸润,3;坏死,4分。总得分计算为每个标准的得分总和。
[0068]
1.8.酶联免疫反应
[0069]
收集小鼠血清,并使用elisa(invitrogen,california,usa)测定小鼠外周血中il 6、il 1β和tnfα的水平。实验程序严格按照制造商的说明进行。
[0070]
1.9.rna测序
[0071]
提取raw264.7细胞的rna,并将其送往novogene(中国北京)进行ran-seq实验。收集clp小鼠的肠道粪便,并将其送往生物标记技术公司(bmk,中国北京)进行16s测序实验。
[0072]
1.10.统计分析
[0073]
所有实验结果均以平均值
±
标准差(sd)表示,所有实验至少独立进行三次。使用student t检验(graphpad软件)进行组间分析。p值《0.05被认为具有统计学意义。所进行的统计测试如图例所示:*p《0.05,**p《0.01,***p《0.001,***p《0.0001。
[0074]
2.结果
[0075]
2.1可利霉素强力地抑制脂多糖诱发的炎症
[0076]
可利霉素是一种多组分药物,以ispⅰ、ⅱ和ⅲ为主要成分(结构式见图1)。本实施例进行了细胞水平的研究,以验证ca的抗炎作用。
[0077]
通过cck测定不同浓度的ca、ca01和ca03的细胞毒性作用。所有3种药物(或成分)在高达10μg/ml时均未观察到细胞毒性。它们对lps诱导的ifnβ产生的影响也通过raw lucia中的萤光素酶测定进行了检测
tm isg细胞。数据显示,两种药物组合物均未对细胞产生明显的毒副作用,但均能抑制lps诱导的ifnβ的表达。进一步研究了10μg/ml ca、ca01和ca03对lps诱导的细胞因子产生的影响。结果表明,与ca01和ca03相比,ca在mrna表达上更有效地抑制了lps诱导的il 6、il 1β和tnfα(结果见图2,图2是lps(1μg/ml)刺激8小时后测量的细胞中il 6、il1β和tnfα的mrna表达图,其中a是细胞中tnfα的mrna表达,b是细胞中il 1β的mrna表达,c是细胞中il 6的mrna表达;因此在随后的实验中使用了ca。类似地,10μg/ml ca在thp1细胞中抑制lps诱导的il 6、il 1β和tnfα的mrna表达。10μg/ml ca还抑制lps在raw264.7细胞中诱导的il 6、il 1β和tnfα蛋白表达(图3,图3是lps(1μg/ml)刺激12小时后,测量细胞培养基中il6、il1β和tnfα的含量图,其中a是il6的含量,b是il1β的含量,c是tnfα的含量。总之,这些数据表明ca可以拮抗lps诱导的raw264.7和thp1细胞的炎症反应。
[0078]
使用rna测序来分析10μg/ml ca对lps诱导的炎症反应的遗传变化:数据显示,ca对lps诱导的炎症的抑制与多种免疫激活信号通路的变化有关,包括nf-κb信号通路。ca可抑制lps诱导的il 6、il 1β、nf-κb1等基因的表达(图4,图4是热图显示29个富集基因在各组(mock、ca、lps和ca lps)之间的差异表达图。(平均值
±
标准差,n=3;通过student t检验分析,**p《0.01,***p《0.001,*****p《0.0001))。用rt-qpcr验证rna测序显示的基因表达结果时,也观察到了同样的现象。
[0079]
图1-4是可利霉素抑制lps诱导的raw264.7细胞中炎症因子的表达。
[0080]
2.2可利霉素阻断nf-kb的免疫激活
[0081]
由于ca下调lps诱导的nf-κb mrna的表达,进一步研究了其对nf-κb信号通路中相
关蛋白变化的影响。lps可以激活ikk复合物,从而导致ikb磷酸化,从而释放nf-κb复合物来激活炎症反应。免疫荧光结果显示,ca保留了lps诱导的ikb-α的减少(见图5,图5是可利霉素通过介导nf-κb途径影响lps诱导的炎症的免疫荧光结果显示图,其中a是ikb-α的免疫荧光结果,b是荧光强度量化结果)并抑制了nf-κb核进入率(见图6,图6是可利霉素通过介导nf-κb途径影响lps诱导的炎症的免疫荧光结果显示图,其中a是nf-κb的免疫荧光结果,b是nf-κb入核率的统计图)。图5和图6说明当用lps(1μg/ml)刺激raw264.7细胞15分钟时,10μg/ml ca保留lps诱导的ikb-α的减少(a),并降低nf-κb核进入率(b)。总之,这些结果表明,ca可以在体外通过调节nf-κb信号通路来减少lps诱导的炎症。类似的,如蛋白质印迹所示,ca抑制lps诱导的p-ikb-α和p-nf-κb的蛋白表达(见图7,图7是对p-ikb-α(f)和p-nf-κb(g)的蛋白水平进行定量,并在蛋白质印迹中绘制图。其中a是各个基因的蛋白质印记图,b是p-ikb-α的定量图,c是p-nf-κb的定量图(平均值
±
标准差,n=3;通过student t检验分析,*p《0.05,**p《0.01,***p《0.001)。
[0082]
图5-7是可利霉素通过介导nf-κb途径影响lps诱导的炎症。
[0083]
2.3可利霉素在体内展现出低毒性
[0084]
为了研究ca在体内的作用,本实施例首先验证了它在c57bl/6小鼠中的毒性。用不同浓度的ca(25-200mg/kg
·
天)灌胃小鼠6天,以评估ca的药代动力学特征。结果发现,用测试的ca剂量,ca在6天后不会影响小鼠的体重(n=5,图8是连续灌胃给予不同浓度的ca(25-200mg/kg/天)6天对小鼠体重的影响结果图(n=5))。此外,通过提取小鼠的心、肝、脾、肺和肾,发现各种器官的重量也没有受到显著影响(n=5,图9是不同浓度的ca(25-200mg/kg/天)灌胃对小鼠(n=5)心脏、肾脏、肺、肝脏和脾脏重量的影响结果图)。此外,苏木精-伊红染色显示ca(25-200mg/kg
·
天)对小鼠的肺和肝没有明显的毒性和副作用(n=5),见图10,图10是不同浓度ca的苏木精-伊红染色对肺和肝毒性的代表性图像以及半定量组织学评分(n=5)图。其中a是肝组织的he染色图,b是肺组织的he染色图,c是肝组织的病理学评分,d是肺组织的病理学评分。
[0085]
图8-10是说明不同浓度ca对c57bl/6小鼠的影响。这些结果表明,使用的ca浓度在野生型小鼠中没有产生损伤。
[0086]
2.4可利霉素有效治疗脂多糖诱发的小鼠败血症
[0087]
参见图11,可利霉素治疗lps诱导的小鼠炎症时,小鼠的治疗方案示意图。
[0088]
就ca对lps诱导的炎症的治疗作用而言,发现ca(100mg/kg,三次灌胃,每两天一次)减轻了lps诱导的小鼠(15mg/kg)体温过低(n=10),并延长了lps小鼠的存活时间(n=10)。见图12,图12是可利霉素治疗lps诱导的小鼠炎症实验中,体温和存活率的变化图。其中a是体温变化图,b是存活率变化图。在败血症中,持续的炎症风暴会导致器官损伤和衰竭。ca降低了lps诱导的败血症小鼠(n=5)肝脏中il 6、il 1β和tnfα的mrna表达(图13是肝组织中il-6、il-1β和tnf-αmrna表达的变化(n=5)图,其中a是il-6的mrna表达含量,b是il-1β的mrna表达含量,c是tnf-α的mrna表达含量)和肺部(图14是肺组织中il-6、il-1β和tnf-αmrna表达的变化(n=5)图其中a是il-6的mrna表达含量,b是il-1β的mrna表达含量,c是tnf-α的mrna表达含量)。苏木精-伊红染色显示,ca减轻了lps诱导的败血症小鼠的肺部和肝脏组织中的病变,并减少了中性粒细胞的浸润(n=5)。值得注意的是,中性粒细胞浸润增加与感染性呼吸衰竭和肝脏损伤有关。ca在lps小鼠组织和器官中的保护作用可能归因
于ca优异的药代动力学特性,如高组织亲和力和长半衰期。此外,通过检测小鼠血液中的炎症因子,发现ca处理的lps小鼠外周血中il 6、il 1β和tnfα的含量显著降低,正如预期的那样(n=5),图15是小鼠血清中il-6、il-1β和tnf-α水平的变化(n=5)图,其中a是il-6的蛋白表达含量,b是il-1β的蛋白表达含量,c是tnf-α的蛋白表达含量。western印迹显示,ca可以抑制小鼠肺组织中p-ikb-α和p-nf-κb的表达(图16是蛋白质印迹显示ca对lps诱导的炎症小鼠肺中nf-κb信号通路蛋白的影响图以及定量图。其中a是各个基因的蛋白印记图,b是p-ikb-α的定量图,c是p-nf-κb的定量图,对照是玉米油灌胃(lps剂量为15mg/kg,平均值
±
标准差,n=5;通过student t检验分析,*p《0.05,**p《0.01)),而阻断nf-κb信号传导可以降低炎症因子的表达。
[0089]
图12-16是可利霉素治疗lps诱导的小鼠炎症系列图。
[0090]
这些数据提供了大量证据来支持ca对lps诱导的败血症小鼠全身炎症的缓解作用,包括减少全身炎症反应、减轻肺和肝组织损伤以及抑制nf-κb信号通路。
[0091]
2.5可利霉素治疗clp诱发的小鼠败血症
[0092]
clp诱导的败血症模型是败血症模型的“金标准”。因此本实验建立了clp小鼠模型来验证ca对败血症诱导的炎症的治疗作用。参见图17,ca对clp小鼠炎症的影响实验中小鼠治疗方案示意图。
[0093]
通过行为观察进行的初步实验表明,clp小鼠的活动在手术后显著降低。小鼠蜷缩、颤抖,并从眼眶分泌白色粘液,这些症状在败血症小鼠中常见,在ca治疗后减弱(100mg/kg,灌胃三次,两天一次,n=5)。ca可以有效延长clp小鼠的存活率(n=15)并保持稳定的体温(n=5)见图18,图18是ca对clp小鼠炎症的影响实验中体温(n=5)和存活率(n=15)变化图,其中a是体温变化图,b是存活率图。ca治疗显著降低了clp小鼠(n=5)肝脏中炎症因子的mrna表达,包括il 6、il 1β和tnfα(图19是ca对clp小鼠肝脏中il6、il1β和tnfαmrna表达的影响(n=5)图,其中a是il6的mrna表达含量,b是il1β的mrna表达含量,c是tnfα的mrna表达含量)和肺部(图20是ca对clp小鼠肺中il6、il1β和tnfαmrna表达的影响(n=5)图,其中a是il6的mrna表达含量,b是il1β的mrna表达含量,c是tnfα的mrna表达含量)。也就是说ca治疗显著降低了clp小鼠血清中il 6、il 1β和tnfα的含量(n=5,图21是ca对clp小鼠血清中il6、il1β和tnfα水平的影响(n=5),其中a是il6的蛋白表达含量,b是il1β的蛋白表达含量,c是tnfα的蛋白表达含量)。另外小鼠处理后的状态见图22,图22是从行为角度观察ca对clp小鼠的影响(n=5)图(平均值
±
标准差;通过student t检验分析,*p《0.05,**p《0.01,***p《0.001,*****p《0.0001)。
[0094]
图17-22验证了ca对clp小鼠炎症的影响系列图。
[0095]
总之,这些结果表明,ca治疗在减少clp诱导的败血症小鼠的全身炎症方面具有良好的治疗效果。
[0096]
2.6可利霉素对clp小鼠肠道微生物群分布的影响
[0097]
败血症易感性与肠道微生物群的变化有关,其发病机制包括肠道病原体的扩张、引发免疫炎症反应和肠道益生菌产品的减少。ca具有良好的抗生素功能,ca治疗无疑可以调节败血症小鼠的肠道菌群。因此,利用16s rrna下一代测序技术分析了败血症小鼠ca治疗后肠道菌群的变化,以阐明ca(100mg/kg,灌胃三次,两天一次)的治疗效果与clp败血症小鼠肠道菌群变化之间的关系。检测mock、ca、clp和clp ca组的肠道微生物群。
[0098]
实验结果表明,各组的shannon指数曲线平缓,表明测序数据饱和;特征类型不再随着测序量的增加而增加,这增加了测序结果的可靠性。维恩图显示了组之间共同或独特特征的数量。clp ca组特有的操作分类单元(otu)的数量(46个otu)高于其他组(mock 5otu、ca 3otu和clp 5otu,显示了otu水平上不同组之间特征的一致性)。主坐标分析(pcoa)的3d图显示了各组微生物区系α多样性的差异;群体之间的距离反映了群体之间物种多样性的差异,表明这些群落在组成结构方面是不同的。
[0099]
ace指数、chao1指数、shannon指数和simpson指数可以反映微生物群落的α多样性。clp组的shannon和simpson指数均显著低于mock组(p《0.05,p《0.001)。与clp组相比,clp ca组的ace、chao1、shannon和simpson指数均显著增加(p《0.001,p《0.001,p=0.001,p《0.001)),表明ca可以显著上调clp小鼠肠道微生物的α多样性。图23是ace指数、chao1指数、shannon指数和simpson指数图,其中a是ace指数图,b是chao1指数,c是shannon指数,d是simpson指数。此外,肠道微生物α多样性降低与炎症性肠病有关,而结肠炎则显著恶化了败血症相关的结果。
[0100]
为了确认哪种细菌被ca治疗改变,进而影响clp诱导的败血症的疾病进展,按百分比计数了前十种细菌,并绘制了属水平的热图(图24是属水平的肠道微生物群的组成。(一)属水平上不同类群之间肠道微生物的热图,其中a是相对丰度图,b是热图。(平均值
±
标准差;通过student t检验分析,*p《0.05,***p《0.001))。前十个富集的微生物物种是:escherichia_shigela、akkermansia、拟杆菌类、未分类的muribaculaceae、副杆菌类、利吉拉杆菌属、梭杆菌属、[梭状芽孢杆菌]_innocuum_group、克雷伯菌属和异丙菌属。
[0101]
图23-24验证了ca对clp小鼠粪便微生物群的影响系列图。
[0102]
根据结果,与野生型小鼠(5.5%,p《0.0001)相比,clp诱导的败血症小鼠中志贺菌的比例显著增加(53.4%)。表明志贺菌可能是clp诱导的败血症小鼠的主要病原体。志贺菌是一种致病细菌,可导致小鼠胱天蛋白酶11活化,导致il 1β表达和致命的感染性休克。此外,几项研究表明志贺菌感染与败血症的发展有关。检测到ca治疗显著降低了clp小鼠肠道中escherichia_shigela的数量(从53.3%降至35.3%,p《0.0001),这可能是ca诱导clp小鼠治疗效果的关键因素。ca降低了clp小鼠中克雷伯菌的比例(从0.7%降至0.4%,p《0.01),这是一种与败血症进展相关的机会性和耐药性共生细菌。在ca治疗的clp小鼠中,alloprevotella的丰度增加(从3.8%增加到8.7%,p《0.0001)。益生菌alloprevetella有助于加速肠道微生物群多样性的恢复,这对预防败血症进展很重要。
[0103]
总之,这些结果表明,ca可以影响败血症小鼠肠道中益生菌和病原微生物的数量和丰度,这可能是ca对clp败血症鼠治疗效果的关键。
[0104]
2.7可利霉素治疗对clp小鼠肠道微生物群影响的差异分析和预测
[0105]
为了进一步确定ca治疗改变了哪些细菌,进而影响clp诱导的败血症的疾病进展,使用效应大小的线性判别分析(lefse)进行高维类别比较,检测到细菌群落优势的显著差异。lefse分析用于评估肠道微生物群,以确定其丰度在门和物种水平上的统计差异,并得出分支图(图25是lef硒分析生成的分支图),lda得分临界值》4.0。根据分析结果,正如预期的那样,志贺菌是clp诱导的败血症小鼠的关键细菌类型。拟杆菌门(包括拟杆菌纲、拟杆菌目、普雷沃菌科、allowprevotella属和副拟杆菌属)在clp小鼠中ca处理后显著富集。allowprevotella是一种益生菌,一项研究表明,拟杆菌作为益生菌调节肺部炎症。此外,拟
杆菌门和厚壁菌门产生短链脂肪酸(scfa),包括丁酸、丙酸和乙酸,它们通过作用于肠内分泌细胞表达的g蛋白偶联受体来影响宿主代谢。因此,clp小鼠的ca治疗可能是通过调节肠道微生物群(主要是拟杆菌门)及其代谢产物的丰度来介导的。
[0106]
此外,为了研究ca处理如何发展clp小鼠肠道微生物群的功能活性,通过重建未观察状态(picrust)对群落进行系统发育研究,对细菌群落进行系统发育研究,将16s序列映射到这些细菌种群中可能存在的基因功能和途径(数据库选择:kegg,p值阈值设置:0.05)。picrust分析揭示了不同的宏基因组功能内容,并显示存在26个核心预测类别(图26是使用picrust功能基因预测肠道微生物群的clp和clp ca组之间的差异(数据库选择:kegg,p值阈值设置:0.05)图)。根据kegg途径类别(iii级)的分析,许多代谢相关途径显著富集,包括“次级代谢产物的生物合成”(clp组7.11%,clp ca组7.29%,p《0.0001)、“氨基酸的生物合成”,“嘌呤代谢”(clp组为1.94%,clp ca组为1.97%,p《0.01)、“糖酵解/葡萄糖新生”(clp小组为1.09%,clp ca小组为1.17%,p《0.0001)等,而ca治疗上调了clp小鼠肠道微生物中的这些代谢途径。短链脂肪酸,特别是乙酸盐、丙酸盐和丁酸盐,主要由肠道微生物的厌氧发酵产生,而ca处理会上调厌氧发酵。此外,短链脂肪酸缺乏与炎症性疾病有关,如炎症性肠病和银屑病关节炎。scfas含量的变化可能与ca治疗对clp小鼠炎症的缓解有关。此外,ca治疗显著上调了“抗生素的生物合成”(clp组为5.26%,clp ca组为5.43%,p《0.0001),这也有助于改善clp小鼠的肠道微生物环境。这些数据表明,ca治疗clp小鼠主要通过影响肠道微生物的代谢途径来调节代谢产物的分泌。
[0107]
图25-26是各组肠道微生物组分的差异分析和基因功能预测系列图。
[0108]
总之,ca治疗改变了肠道微生物群的组成,包括致病菌志贺菌的含量降低和益生菌拟杆菌的含量增加,并通过影响肠道微生物群代谢途径改变了代谢产物的分泌(scfas表达增加)。这表明ca治疗在调节肠道微生物环境以治疗clp败血症小鼠方面是有效的。
[0109]
2.8可利霉素在体内抑制lps诱导的炎症方面比阿司匹林表现更好
[0110]
阿司匹林(asp)是一种经典的抗炎药,最近被推荐用于败血症的治疗。多项研究表明,阿司匹林可抑制lps诱导的nf-κb通路的炎症激活。因此,本发明比较了ca和阿司匹林治疗lps诱导的小鼠败血症的疗效。结果发现,相同剂量(100mg/kg)的ca比阿司匹林更好地抑制lps小鼠肺组织中il 6的mrna表达(n=5,图27是ca和阿司匹林对小鼠肺组织中il6、il1β和tnfαmrna表达的抑制作用(n=5)图,其中a是il6的mrna表达含量,b是il1β的mrna表达含量,c是tnfα的mrna表达含量)。当检测血清中的炎症因子(il 6和tnfα)时,获得了一致的结果(n=5,图28是ca和阿司匹林降低小鼠血清中il6、il1β和tnfα水平的作用(n=5)图,其中a是il6的蛋白表达含量,b是il1β的蛋白表达含量,c是tnfα的蛋白表达含量)。图27和28中,ca和阿司匹林的剂量均为100mg/kg,给药三次,每两天一次,其中ca通过灌胃给药,阿司匹林通过腹膜内注射)(平均值
±
sd;通过student t检验分析,*p《0.05,**p《0.01,***p《0.001,***p《0.0001)。
[0111]
简而言之,这些结果表明,在体内对lps诱导的炎症激活的反应中,ca的抗炎能力优于阿司匹林。
[0112]
尽管上面已经示出和描述了本发明的实施例,可以理解的是,上述实施例是示例性的,不能理解为对本发明的限制,本领域的普通技术人员在不脱离本发明的原理和宗旨的情况下在本发明的范围内可以对上述实施例进行变化、修改、替换和变型。
再多了解一些
本文用于创业者技术爱好者查询,仅供学习研究,如用于商业用途,请联系技术所有人。