线粒体呼吸链复合物v在特发性肺纤维化诊断中的应用
技术领域
背景技术:
1.特发性肺纤维化(idiopathic pulmonary fibrosis,ipf)是一种慢性、进行性和纤维化性间质性肺部疾病(lederer and martinez,2018)。ipf的预后很差,平均生存年限在2.5到5年之间,只有20%到30%的受试者再确诊5年后仍然活着(noble et al.,2011;raghu,2017;raghu et al.,2011)。到目前为止,ipf的确切原因仍然不清楚,仅有两种已批准的药物尼达尼布和吡非尼酮可用于治疗。这两种药物已被证明可以减缓病情发展,但不能阻止疾病(raghu,2017)。因此,如何在发病早期进行有效地诊断和治疗,一直是ipf疾病的问题关键。线粒体功能和行为是人类的生理核心,因此,线粒体功能障碍会导致各种疾病的发生(nunnari and suomalainen,2012)。近期研究表明,伴随衰老的生物能量学和线粒体功能的改变,可能参与了ipf的发病(bueno et al.,2020;morita et al.,2017)。
2.复合物v(complex v)是线粒体氧化磷酸化(oxphos)系统中的atp合成酶。它利用呼吸链复合物i-iv在线粒体膜上产生的质子电化学梯度,将质子流与adp和无机磷酸盐转化为atp(meulemans et al.,2010)。在人类细胞中,90%或更多的细胞atp是由复合物v(jonckheere et al.,2012)合成的。同时,有研究证明复合物v二聚化与线粒体接触位点和嵴组织系统(micos)协作建立了线粒体内膜曲率,启动了嵴的形成,从而决定了线粒体的形态(baker et al.,2019;cogliati et al.,2016;de los rios castillo et al.,2011;stephan et al.,2020)。因此,复合物v主要有两个功能,分别为atp合成酶和嵴组织者,在线粒体生物发生和细胞生物发生中起着决定性的作用。心血管疾病(kwong and molkentin,2015)、癌症(bonora et al.,2015)和神经退行性疾病(golpich et al.,2017)都曾报道过与复合物v相关的功能缺陷。因此,线粒体呼吸链复合物v在ipf病症中的功能机理研究具有重要意义。
技术实现要素:
3.本技术通过以下技术方案解决了本领域中存在的技术问题。
4.1.一种用于检测线粒体呼吸链复合物v活性的试剂在制备用于辅助诊断特发性肺纤维化的组合物或试剂盒中的用途。
5.2.项目1所述的用途,其中所述试剂为抗复合物v的抗体、其抗原结合片段、与编码复合物v的核酸结合的探针或引物、或适体。
6.3.项目2所述的用途,其中所述抗体为单克隆抗体。
7.4.用于辅助诊断发性肺纤维化的组合物,其包含用于检测线粒体呼吸链复合物v活性的试剂。
8.5.项目4所述的组合物,其中所述试剂为抗复合物v的抗体、其抗原结合片段、与编码复合物v的核酸结合的探针或引物、或适体。
9.6.项目5所述的组合物,其中所述抗体为单克隆抗体。
10.7.用于辅助诊断发性肺纤维化的试剂盒,其包含项目4-6任一项的组合物和使用说明书。
11.8.一种体外筛选治疗特发性肺纤维化的试剂的方法,其包括对tgf-β1诱导的特发性肺纤维化细胞模型施用所述试剂,并测定施用前后线粒体呼吸链复合物v活性,其中与施用所述试剂前的复合物v活性相比,施用所述试剂后复合物v活性升高表明所述试剂为特发性肺纤维化的潜在的治疗剂。
12.9.项目8所述的方法,其中所述方法还包括测定施用前后线粒体呼吸链复合物i的活性、亚基蛋白ndufa9的量、atp5a的量、ros含量、atp含量和/或线粒体膜电位,其中并测定施用前后线粒体呼吸链复合物v活性,其中分别与施用前复合物i的活性、亚基蛋白ndufa9的量、atp5a的量、ros含量、atp含量和线粒体膜电位相比,施用复合物i的活性升高、亚基蛋白ndufa9的量上调、atp5a的量上调、ros含量降低、atp含量升高和线粒体膜电位降低表明所述试剂为特发性肺纤维化的潜在的治疗剂。
13.10.项目8或9所述的方法,其中所述特发性肺纤维化细胞模型为用a549细胞构建的特发性肺纤维化细胞模型。
14.本文所述的用于检测线粒体呼吸链复合物v活性的试剂没有特别限制,只要其能够特异性检测到线粒体呼吸链复合物v活性水平以及该水平的变化即可。可以想到特异性结合复合物v的抗体或抗体抗原结合片段可用于检测线粒体呼吸链复合物v活性。
15.如本文所用,除非另有说明,否则术语“抗体”包括特异性结合其靶标的抗体或其抗原结合片段,并且是单克隆抗体、结构域抗体、单链、fab片段、f(ab')2片段、scfv、scab、dab、单结构域重链抗体和单结构域轻链抗体。在一些实施方案中,抗体是igg抗体。在一些实施方案中,抗体是igg1抗体。在一些实施方案中,抗体是igg4抗体。在一些实施方案中,抗体是scfv抗体。在一些实施方案中,结合其靶标的这种抗体或其免疫活性片段是小鼠、嵌合、人源化或完全人单克隆抗体。
16.适体指对于特别的靶分子具有结合活性的核酸分子。适体可以通过与特定靶分子结合,来抑制特定靶分子的活性。本发明的适体可以是rna、dna、经修饰的核酸或其混合物。本发明的适体还可以采取线性或环状形式。
17.本发明的适体可以如本文公开的且通过本领域本身已知的方法进行化学合成。适体以广泛多样的结合模式与靶分子结合,例如基于磷酸基的负电荷的离子键、基于核糖的疏水键和氢键、以及基于核酸碱基的氢键和堆积相互作用。特别地,以与构成核苷酸数目相同的数目存在的,基于磷酸基的负电荷的离子键很强,并且与蛋白质的正电荷表面上存在的赖氨酸和精氨酸结合。为此,不涉及与靶分子的直接结合的核酸碱基可以是被取代的。特别地,因为茎结构的区域已经形成碱基对,并且面对双螺旋结构的内部,所以核酸碱基不太可能与靶分子直接结合。因此,即使当碱基对替换为另一个碱基对时,适体的活性通常也不减少。在其中不形成碱基对的结构例如环结构中,如果核酸碱基不涉及与靶分子的直接结合,碱基取代是可能的。关于核糖的2'-位置的修饰,在核糖的2'-处的官能团很少与靶分子直接相互作用,但在许多情况下,他是无关的,并且可以由另一种经修饰的分子取代。因此,除非涉及与靶分子的直接结合的官能团被取代或缺失,适体通常保留其活性。总体三维结构不广泛改变也是重要的。
18.适体可以通过利用selex方法或其改善的版本(例如ellington等人(1990)
nature,346,818-822;tuerk等人(1990)science,249,505-510)进行制备。在selex方法中,通过增加轮数或使用竞争物质,浓缩且选择对于靶分子显示出更强的结合潜力的适体。因此,通过调整selex的轮数和/或改变竞争条件,在一些情况下,可以获得具有不同结合力的适体、具有不同结合模式的适体、以及具有相同结合力或结合模式但不同碱基序列的适体。selex方法包括通过pcr的扩增过程;通过在过程中经由使用锰离子等引起突变,能够以更高的多样性执行selex。
附图说明
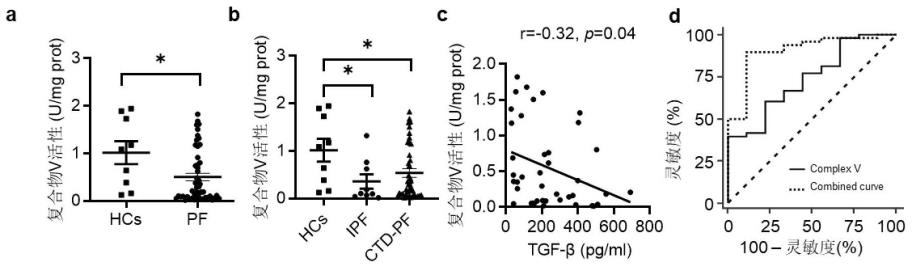
19.图1.tgf-β升高与肺纤维化患者线粒体呼吸链复合物v活性降低相关。
20.图1a显示了肺纤维化患者与健康人(hc)的复合v活性。图1b显示了ipf、ctd-ild和健康人(hc)的复合物v活性。图1c显示了复合物v活性与血清tgf-β1水平的相关性分析。图1d显示了肺纤维化患者与健康人(hc)复合物v活性(实线)以及复合物v活性与血清tgf-β1联合水平(虚线)的roc曲线分析。
21.图2.tgf-β诱导a549细胞发生细胞形态和线粒体形态改变。
22.图2a显示了a549细胞用不同浓度的tgf-β1处理48h,分别用倒置荧光显微镜、透射电镜和omx模式结构光照明显微镜拍照。dic图像显示细胞在不同处理条件下的形态学变化,图像使用尼康ts2倒置显微镜10倍物镜,比例尺:100μm。tem图像显示了不同实验条件下线粒体嵴的变化,使用tecnai spirit(120kv)拍摄,比例尺:500nm。omx图像显示了不同处理条件下线粒体(白色)形态的3d变化,这些图像是使用sim-omx和60倍油浸物镜(比例尺:10μm)拍摄的。tgf-β1诱导的肺上皮细胞线粒体动态变化。图2b显示了image j软件中的mitochondria analyzer插件(默认参数)用于分析细胞3d图像中每个细胞中线粒体表面积、分支和分支节点,并进行数据统计,进行turkey多重比较检验的单因素方差分析。*:p《0.05。
23.图3.tgf-β诱导a549细胞发生线粒体功能障。
24.图3a显示了利用iga的方法检测已纯化线粒体蛋白中的呼吸链复合物ⅰ、复合物ⅱ、复合物ⅳ、复合物
ⅴ
的活性。图3b显示了通过蛋白免疫印迹对不同处理条件下a549中ndufa9、sdha、uqcrc2、cox4、atp5a、atp6、atp6v0d1、atp8、tom20和gapdh的蛋白表达水平进行检测。其中,所有泳道的蛋白上样量一致。图3c显示了通过tmre对不同tgf-β1浓度诱导的a549细胞中线粒体膜电位检测,图像是使用具60倍油浸物镜的olympus fv3000在相同的参数设置下拍摄的,比例尺:50μm。图3d显示了由image j统计图3c平均荧光强度,并计算线粒体膜电位变化倍数,利用graphpad作图。图3e、f显示了通过流式细胞仪测定tgf-β1诱导的a549细胞中dcf和mitosox的平均荧光强度,并统计a549细胞中ros和mitosox的倍数变化。图3d、e、f数据以95%ci的平均值表示,并进行turkey多重比较检验的单因素方差分析。*:p《0.05;**:p《0.01;***:p《0.001;****:p《0.0001。
25.图4.tgf-β诱导a549细胞纤维化在经pfd治疗后线粒体功能恢复。
26.图4a显示了a549细胞用不同的处理条件处理48小时,使用倒置荧光显微镜和激光共聚焦显微镜拍摄图像。不同处理条件:t=tgf-β15ng/ml,p1=pfd 250μg/ml,p2=pfd 500μg/ml。dic图像显示细胞在不同处理条件下的形态变化,图像使用尼康ts2倒置显微镜拍摄,比例尺:100μm。共聚焦图像显示了不同处理条件下线粒体(白色)形态的变化,图像使
用olympus fv1000 60x油浸物镜拍摄,比例尺:50μm,放大图像的比例尺为:20μm。图4b显示了利用iga方法检测已纯化的线粒体蛋白中线粒体呼吸链复合物ⅰ、复合物ⅱ、复合物ⅳ、复合物
ⅴ
的蛋白活性。图4c显示了不同处理条件下a549蛋白提取物中ndufa9、sdha、uqcrc2、cox4、atp5a、tom20、β-actin和gapdh蛋白表达水平的免疫印迹实验结果。所有泳道蛋白上样量一致。图4d显示了不同处理条件下a549细胞中ros的倍数变化。通过流式细胞仪测定a549细胞中dcf的平均荧光强度。图4e显示了不同处理条件下a549细胞中的atp含量。图4f显示了不同处理条件下a549细胞tmre(线粒体膜电位)的变化,平均荧光强度由image j获得并进行统计。图4g显示了通过激光共聚焦利用tmre检测在不同处理条件下a549细胞中线粒体膜电位的变化。这些图像是使用具有60倍油浸物镜的olympus fv3000在相同的参数设置下拍摄的,比例尺:50μm。图4d、e、f数据以95%ci的平均值表示,并进行turkey多重比较检验的单因素方差分析。*:p《0.05;**:p《0.01;***:p《0.001;****:p《0.0001。
具体实施方式
27.为使本发明的目的、技术方案和优点更加清楚明白,以下结合具体实施例,并参照附图,对本发明作进一步的详细说明。
28.实施例1.tgf-β升高与肺纤维化患者线粒体呼吸链复合物v活性降低相关
29.为了研究ipf疾病与线粒体呼吸链复合物v之间的关系,我们收集了已诊断的ipf、cd-pf病人和hcs(健康人)的血液样本,通过生化试剂盒对线粒体呼吸链复合物i、复合物ii、复合物iii、复合物iv、复合物v进行了活性检测。图1表明线粒体呼吸链复合物v活性降低与肺纤维化患者tgf-β升高有关。图1a,b表明与健康人相比,肺纤维化患者的线粒体呼吸链复合物v活性显著降低。图1c表明线粒体呼吸链复合物v活性降低与tgf-β升高显著相关。图1d显示对ild患者与健康人的复合物v活性水平进行的受试者工作特征(roc)分析,结果显示曲线下面积(auc)值为74.44%。
30.具体方法
31.人类血液样本的获得
32.成人ipf患者(n=9)、结缔组织病相关pf(ctd-pf)患者(n=41)以及年龄和性别匹配的健康对照(hcs)(n=9)的血液样本从天津医科大学总医院收集获得。
33.人类血液样本线粒体呼吸链复合物的活性检测
34.对人类血液样本的线粒体呼吸链复合物的活性检测利用micro mitochondrial respiratory chain complex activity assay kit(索莱宝)按照其说明书进行。
35.实施例2.tgf-β诱导a549细胞发生细胞形态和线粒体形态改变
36.为了研究tgf-β与线粒体之间的联系,我们利用tgf-β诱导a549细胞构建了间质化细胞模型,并对线粒体形态,內嵴状况进行了观察。图2a是通过普通倒置荧光显微镜、透射电子显微镜(tem)以及结构光照明显微镜(sim,omx模式)分别对tgf-β诱导的a549细胞进行细胞形态、线粒体形态、內嵴状态观察,该结果表明tgf-β诱导a549细胞形态向间质化细胞形态转变,线粒体形态发生断裂,线粒体网状结构、內嵴结构受到破坏。图2b对sim拍摄的3d图片进行了细胞内线粒体表面积、分支节点的统计,反映了线粒体网络结构遭受破坏。结果表明随着tgf-β浓度的升高,线粒体结构破坏逐渐加重。
37.具体方法
38.细胞培养和分组处理
39.a549细胞采用含10%fbs,1%青霉素,1%链霉素的dmem培养基在5%co2,37℃培养箱中进行培养。
40.tgf-β1诱导处理方法:首先a549细胞在不含10%fbs的dmem培养基中培养24h,进行饥饿处理,之后分别添加tgf-β1至终浓度为0、1、2、5、10ng/ml,诱导培养48h.
41.细胞形态观察:在tgf-β1处理后,利用普通倒置荧光显微镜在物镜10x,明场下及进行图片获取。
42.透射电子显微镜观察线粒体形态及內嵴结构:tgf-β1处理后的细胞经胰酶消化,pbs洗涤,离心收集细胞沉淀,加入适量的2.5%戊二醛固定细胞,之后使用3%四氧化锇重新固定细胞。将标本用乙醇分级脱水,之后epon树脂包埋,切片,然后用透射电子显微镜成像。
43.采用sim omx模式观察线粒体3d结构:a549细胞处理后,采用mitotracker green和dapi染色,pbs清洗后加入新鲜的培养基在sim上进行图像获取。
44.线粒体表面积、分支、分支节点统计:首先对sim获得的图像利用imaris软件构建线粒体表面,添加表面积的颜色条(color bar),并对线粒体表面积进行统计和数据输出。利用imagej软件对线粒体分支和分支节点进行统计。
45.实施例3、tgf-β诱导a549细胞发生线粒体功能障碍
46.为了进一步研究tgf-β对线粒体功能的影响,我们利用a549细胞模型对线粒体氧化磷酸化系统进行研究。图3a代表blue native page和in gel activity分析线粒体呼吸链各复合物,结果表明复合物i和复合物v的活性随tgf-β1处理浓度的增加而降低。图3b为线粒体各复合物亚基蛋白的western blot结果,ndufa9与atp5a量降低,其他没有显著量的变化。图3c,d结果表明线粒体膜电位(tmre)随tgf-β1处理浓度的增加而降低。图3e,f结果表明细胞ros含量和线粒体ros含量显著上调。结果表明tgf-β浓度的升高造成线粒体功能障碍。
47.具体方法
48.blue native page:收集已诱导处理的细胞(实施例2),加入200μl预冷pbs重悬细胞,然后加入等体积4mg/ml的digitonin(溶解于pbs),上下颠倒两次混匀,冰上孵育10min;之后,预冷pbs加至1.5ml,4℃,10000g离心10min;弃去上清,然后1ml pbs重悬沉淀,4℃,10000g离心10min离心去除残留的digitonin;加入50μl bn样品缓冲液,5μl 10%ddm,吹打10-20次重悬沉淀,注意不要吹起泡沫,冰上固定20min;4℃,20000g离心20min,取出上清至新的ep管,留5μl进行bca法蛋白定量,剩余的加入2μl的5%的考马斯亮蓝g-250,混匀。取10μg样品进行blue-native page分离线粒体复合物。
49.复合物i活性:将blue-native page分离胶孵育在50mm tris-hcl,ph 7.4缓冲液中(含有0.5mm nbt(nitroblue tetrazolium chloride,氯化硝基四氮唑),5mm nadh(nicotinamide adenine dinucleotide,烟酰胺腺嘌呤二核苷酸)),室温作用1小时。
50.复合物ii活性:将blue-native page分离胶孵育在50mm tris-hcl,ph 7.4缓冲液中(0.2mm pms(methyl phenazinium methyl sulfate,吩嗪甲基硫酸盐),84mm琥珀酸,50mm nbt),室温作用1小时。
51.复合物iv活性:将blue-native page分离胶孵育在50mm tris-hcl,ph 7.4缓冲液
中(含有0.1%二氨基联苯胺,24个单位/ml过氧化氢酶,0.1%细胞色素c),37度作用3-6小时。
52.复合物v活性:将blue-native page分离胶用清水漂洗10分钟,放入0.1m的甘氨酸缓冲液中(ph 8.6)作用1小时。再将分离胶放入含有下列成分的缓冲液中:35mm tris碱,270mm甘氨酸,14mm硫酸镁,5mm atp,0.2%硝酸银,37度作用3-6小时。
53.蛋白免疫印迹(western blot):收集已诱导处理的细胞(实施例2),加入200μl含蛋白酶抑制剂混合物(roche)的ripa裂解液,冰上放置15分钟,14000rpm离心15分钟,上清用bca试剂盒测定蛋白浓度。配制15%sds-page胶,将已经测好蛋白浓度的样品,加上一定体积的蛋白上样缓冲液,95度加热15分钟,取一定量蛋白加入到12%sds-page胶中,120v恒压,电泳2小时左右。将胶上的蛋白样品转移到pvdf膜上,5%脱脂奶粉室温封闭3小时,然后不同一抗按照说明书稀释,孵育过夜。tbst(150mm nacl,20mm tris-hcl,ph7.4,tween-200.05%)洗膜3次,每次10分钟。二抗1:3000稀释,室温孵育3小时。tbst洗膜三次后加入超敏ecl化学发光试剂(碧云天公司),放入暗盒中曝光。
54.线粒体膜电位检测:将a549细胞在接种至共聚焦培养皿中进行tgf-β1诱导处理后,用pbs稀释tmre至500nm,37℃避光孵育10min,pbs洗涤后加入新鲜培养基,同过共聚焦显微镜在同一条件下对不同处理细胞进行拍摄,通过imagej软件统计荧光强度。
55.细胞ros检测:处理后的细胞经胰酶消化收集,按照活性氧检测试剂盒(碧云天)的操作说明进行处理,通过流式细胞仪进行检测。
56.线粒体ros检测:处理后的细胞经胰酶消化收集,按照mitosox
tm red mitochondrial superoxide indicator(invitrogen)的操作说明进行处理,通过流式细胞仪进行检测。
57.实施例4.tgf-β诱导a549细胞纤维化经pfd治疗后线粒体功能恢复
58.为了研究目前已有ipf治疗药物pfd对线粒体功能障碍的治疗和阻止tgf-β诱导纤维化的效果,我们利用a549细胞模型对此进行研究。图4a为pfd可以阻止tgf-β诱导的细胞形态间质化转变和线粒体形态结构的破坏。图4b代表blue native page和in gel activity分析线粒体呼吸链各复合物,结果表明复合物i和复合物v的活性在加入pfd治疗后相较tgf-β诱导组明显上调。图4c为线粒体各复合物亚基蛋白的western blot结果,亚基蛋白ndufa9与atp5a量在加入pfd治疗后相较tgf-β诱导组明显上调,其他没有显著量的变化。图4d结果表明细胞ros含量在加入pfd后得到明显抑制。图4e为pfd治疗后atp的检测结果,atp含量与tgf-β诱导组相比加入pfd后有所恢复。图4f,h结果表明线粒体膜电位(tmre)在加入pfd治疗后相较tgf-β诱导组明显降低。结果表明tgf-β浓度的升高造成线粒体功能障碍。
59.具体方法
60.细胞处理:首先对a549细胞进行去血清饥饿处理24h,之后分别加入5ng/ml tgf-β1(t组),5ng/ml tgf-β1 250μg/ml pfd(t-p1组),5ng/ml tgf-β1 500μg/ml pfd(t-p2组)和500μg/ml pfd(p2组)进行48h诱导处理,设置正常培养的细胞为对照组。
61.细胞形态观察、blue native page、western blot、ros、线粒体膜电位检测的具体方法与上述实施例2和实施例3中的操作方法相同。
62.atp检测:处理后的细胞经胰酶消化收集,按照atp检测试剂盒(碧云天)的操作说
明进行,通过流式细胞仪进行检测。
63.参考文献
64.baker,n.,patel,j.,and khacho,m.(2019).linking mitochondrial dynamics,cristae remodeling and supercomplex formation:how mitochondrial structure can regulate bioenergetics.mitochondrion 49,259-268.
65.bonora,m.,wieckowski,m.r.,chinopoulos,c.,kepp,o.,kroemer,g.,galluzzi,l.,and pinton,p.(2015).molecular mechanisms of cell death:central implication of atp synthase in mitochondrial permeability transition.oncogene 34,1475-1486.
66.bueno,m.,calyeca,j.,rojas,m.,and mora,a.l.(2020).mitochondria dysfunction and metabolic reprogramming as drivers of idiopathic pulmonary fibrosis.redox biol 33,101509.
67.cogliati,s.,enriquez,j.a.,and scorrano,l.(2016).mitochondrial cristae:where beauty meets functionality.trends biochem sci 41,261-273.
68.de los rios castillo,d.,zarco-zavala,m.,olvera-sanchez,s.,pardo,j.p.,juarez,o.,martinez,f.,mendoza-hernandez,g.,garcia-trejo,j.j.,and flores-herrera,o.(2011).atypical cristae morphology of human syncytiotrophoblast mitochondria:role for complex v.j biol chem 286,23911-23919.
69.golpich,m.,amini,e.,mohamed,z.,azman ali,r.,mohamed ibrahim,n.,and ahmadiani,a.(2017).mitochondrial dysfunction and biogenesis in neurodegenerative diseases:pathogenesis and treatment.cns neuroscience&therapeutics 23,5-22.
70.jonckheere,a.i.,smeitink,j.a.,and rodenburg,r.j.(2012).mitochondrial atp synthase:architecture,function and pathology.journal of inherited metabolic disease 35,211-225.
71.kwong,j.q.,and molkentin,j.d.(2015).physiological and pathological roles of the mitochondrial permeability transition pore in the heart.cell metab21,206-214.
72.lederer,d.j.,and martinez,f.j.(2018).idiopathic pulmonary fibrosis.the new england journal of medicine 378,1811-1823.
73.meulemans,a.,seneca,s.,pribyl,t.,smet,j.,alderweirldt,v.,waeytens,a.,lissens,w.,van coster,r.,de meirleir,l.,di rago,j.p.,et al.(2010).defining the pathogenesis of the human atp12p w94r mutation using a saccharomyces cerevisiae yeast model.j biol chem 285,4099-4109.
74.morita,m.,prudent,j.,basu,k.,goyon,v.,katsumura,s.,hulea,l.,pearl,d.,siddiqui,n.,strack,s.,mcguirk,s.,et al.(2017).mtor controls mitochondrial dynamics and cell survival via mtfp1.mol cell 67,922-935e925.
75.noble,p.w.,albera,c.,bradford,w.z.,costabel,u.,glassberg,m.k.,kardatzke,d.,king,t.e.,jr.,lancaster,l.,sahn,s.a.,szwarcberg,j.,et al.(2011)
.pirfenidone in patients with idiopathic pulmonary fibrosis(capacity):two randomised trials.lancet(london,england)377,1760-1769.
76.nunnari,j.,and suomalainen,a.(2012).mitochondria:in sickness and in health.cell 148,1145-1159.
77.raghu,g.(2017).idiopathic pulmonary fibrosis:lessons from clinical trials over the past 25 years.the european respiratory journal 50.
78.raghu,g.,collard,h.r.,egan,j.j.,martinez,f.j.,behr,j.,brown,k.k.,colby,t.v.,cordier,j.f.,flaherty,k.r.,lasky,j.a.,et al.(2011).an official ats/ers/jrs/alat statement:idiopathic pulmonary fibrosis:evidence-based guidelines for diagnosis and management.american journal of respiratory and critical care medicine 183,788-824.
79.stephan,t.,bruser,c.,deckers,m.,steyer,a.m.,balzarotti,f.,barbot,m.,behr,t.s.,heim,g.,hubner,w.,ilgen,p.,et al.(2020).micos assembly controls mitochondrial inner membrane remodeling and crista junction redistribution to mediate cristae formation.the embo journal 39,e104105.
80.以上所述的具体实施例,对本发明的目的、技术方案和有益效果进行了进一步详细说明,应理解的是,以上所述仅为本发明的具体实施例而已,并不用于限制本发明,凡在本发明的精神和原则之内,所做的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。