抗h5n1型禽流感病毒药物及其应用
1.技术领域
2.本发明属于抗h5n1型高致病性禽流感抗体技术领域,涉及一株抗h5n1型高致病性禽流感病毒全人源抗体组合pcag
‑
scfv 1a2
‑
gfp和igg1及其应用。
3.
背景技术:
4.禽流感是由a型流感病毒引起的禽类烈性传染病。近十年来,h5n1 型禽流感肆虐全球,不仅给世界养禽业造成损失,而且随着禽流感病毒基因组的重组或变异,使原来只在禽间传播的少数h5n1 型禽流感病毒获得了能在人体内繁殖的能力,从而具有感染人的能力,主要引起急性肺损伤、急性呼吸窘迫综合征(ards)、多脏器功能衰竭等多种并发症,虽然目前尚未有大范围的疫情爆发,但每年都有散发病例的出现;另一方面,由于人体内没有针对h5n1型禽流感病毒抗原的抗体,人感染高致病性禽流感病毒(hpaiv)后则会呈现高发病率和高死亡率的特征,因此,hpaiv 对人类健康和世界公共卫生安全也已构成重大威胁。
5.作为化学药物和疫苗的有效补充,由抗体介导的预防和临床治疗措施已显现良好的效果,其应用前景得到专家的认同。作为人体内一种最重要的抗病毒免疫介质,抗体分子可以通过阻断病毒颗粒与其受体的结合、激活巨噬细胞、nk 细胞等杀伤细胞,激活补体和t 细胞效应等多种机制来杀伤、清除病毒颗粒及受感染细胞。抗体不仅可以对流行区的高危易感人群进行紧急被动免疫,对于临床病人还可起到一定的治疗作用。
6.病毒是胞内寄居病原体,利用宿主细胞的核酸和蛋白装置来增殖,通过与宿主细胞表面受体的结合,可感染众多的靶细胞。病毒的生命形式存在两种方式:保持感染活性的细胞外形式和显示复制活性的细胞内形式。普通的抗体制剂,受其分子量限制,难以穿透细胞膜进入细胞内,因此,在本研究中,我们将前期研究获得的scfv 1a2基因组通过与高效表达载体连接后由脂质体介导转入细胞内,并进一步表达细胞内抗体,抑制已感染细胞内的病毒的组装,降低胞内病毒的复制速度,通过控制细胞内病毒数量以期待增加宿主细胞自身分泌的i型干扰素(ifn α\β)等细胞因子的分泌,进而促进特异性的ctl (cd8 t)细胞杀伤作用及相关细胞因子的转录水平;另一方面,将scfv 1a2抗体通过基因工程手段构建为含有fc段的全分子抗体igg1,使其能中和细胞外的病毒并有效的诱导机体的“抗体依赖细胞介导的细胞毒性作用”(antibody
‑
dependent cell
‑
mediated cytotoxicity,adcc)生物学效应。通过全人源细胞内抗体和细胞外抗体组合的治疗达到同时抑制细胞内病毒的增殖和细胞外病毒的感染,从而提高机体免疫水平,以期待提高hpaiv感染小鼠的生存率,为进一步探讨有针对性的hpaiv抗体治疗方案奠定基础。
7.
技术实现要素:
8.解决的技术问题:本发明针对上述技术问题,提供一种抗h5n1型禽流感病毒药物
及其应用,利用抗h5n1型高致病性禽流感病毒全人源抗体组合pcag
‑
scfv 1a2
‑
gfp和igg1制备预防和治疗h5n1型禽流感病毒感染药物。
9.技术方案:抗h5n1型禽流感病毒药物,有效成分为pcag
‑
scfv 1a2
‑
gfp和igg1,所述pcag
‑
scfv 1a2
‑
gfp的轻链的氨基酸序列如seq id no:1所示,重链的氨基酸序列如seq id no:2所示;所述igg1的轻链的氨基酸序列如seq id no:5所示,重链的氨基酸序列如seq id no:6所示。
10.pcag
‑
scfv 1a2
‑
gfp轻链的核苷酸序列如seq id no:3所示,重链的核苷酸序列如seq id no:4所示。
11.igg1轻链的核苷酸序列如seq id no:7所示,重链的核苷酸序列如seq id no:8所示。
12.抗h5n1型高致病性禽流感全人源抗体pcag
‑
scfv 1a2
‑
gfp和igg1组合在制备抗h5n1型禽流感病毒药物中的应用,所述pcag
‑
scfv 1a2
‑
gfp的轻链的氨基酸序列如seq id no:1所示,重链的氨基酸序列如seq id no:2所示;所述igg1的轻链的氨基酸序列如seq id no:5所示,重链的氨基酸序列如seq id no:6所示。
13.有益效果:本发明以含有抗体scfv1a2 基因的pcomb3xss
‑
scfv1a2质粒为模板,pcr扩增pcomb3xss
‑
scfv 1a2 中的scfv 1a2 基因,并同时在片段的两侧添加真核载体的克隆位点序列,产物与高效真核表达载体pcag
‑
gfp 经酶切连接后筛选得到阳性重组子pcag
ꢀ–
scfv 1a2
‑
gfp,且证明细胞内抗体能够正确表达。本发明扩增抗h5n1 型高致病性禽流感病毒小分子scfv1a2抗体的可变区基因vl和vh,分别克隆于l链和h链的盒式真核表达载体pci
‑
rv220
‑
l 和pci
‑
rv220
‑
h 中,构建igg1 真核表达载体。初步筛选的稳定转染株进一步亚克隆,经过几轮筛选,能够获得每100ml产20mg的蛋白表达量,且全分子igg1和小分子scfv1a2的结合活性相同,未受fc片段的影响,亲和力较scfv1a2好。
14.细胞内和细胞外抗体及抗体组合的体内体外中和实验表明,全分子igg1抗体对h5n1型高致病性禽流感病毒的tcid50为10
‑3μg/ml;全分子igg1在15mg/kg用量时对100 tcid50 的a/jiangsu/1/2007(h5n1) 病毒感染小鼠的保护率可达到100%;全分子igg1在15mg/kg,并且辅助30μg pcag
‑
scfv
ꢀ‑
gfp 时对500 tcid50的a/jiangsu/1/2007 (h5n1) 病毒感染小鼠的保护率最高可达66.67%。抗h5n1型高致病性禽流感病毒全人源抗体组合pcag
‑
scfv 1a2
‑
gfp和igg1为抗禽流感病毒人源抗体运用中的一个创新思路。
15.附图说明
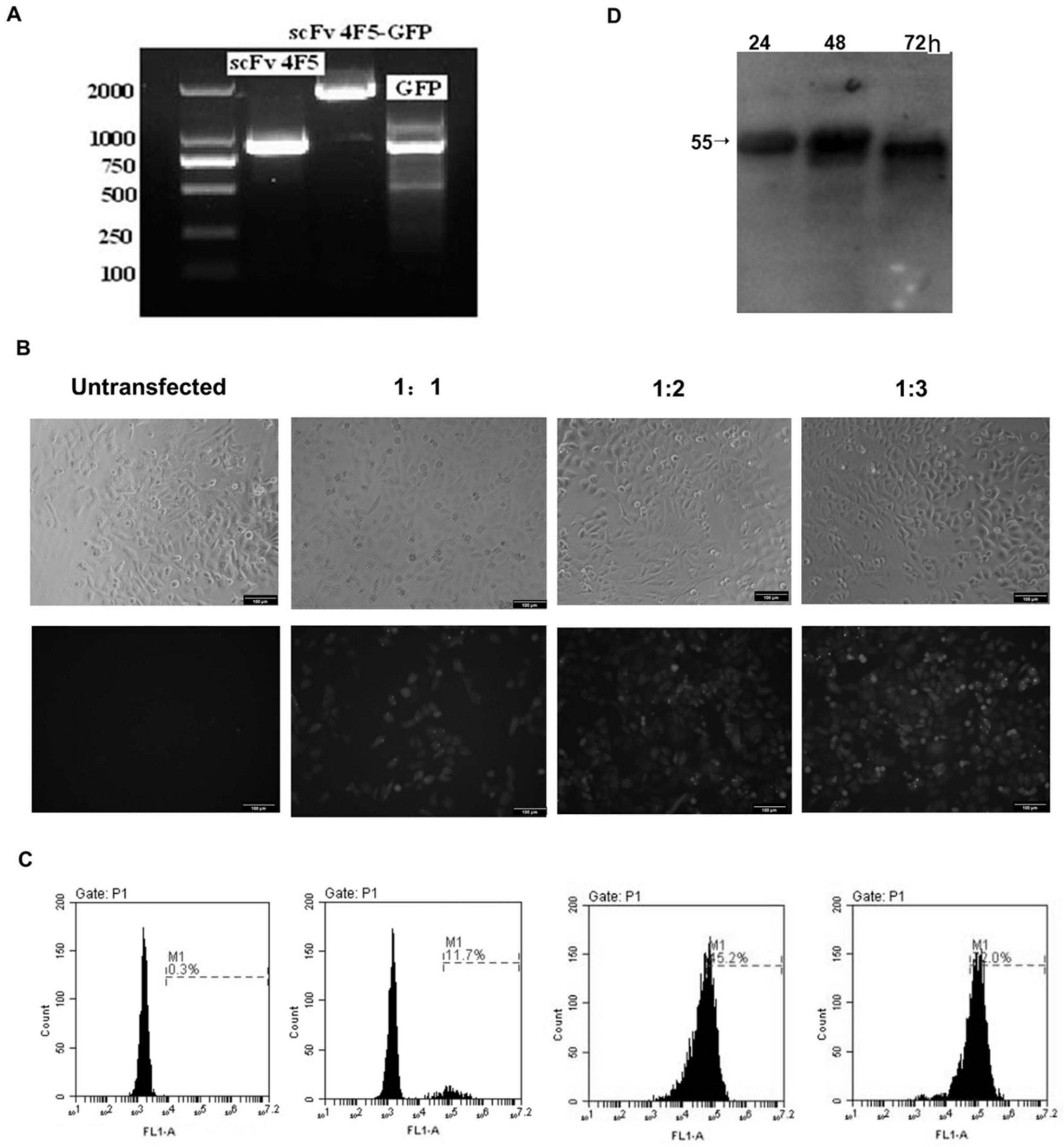
16.图1为重组真核表达载体pcag
‑
scfv1a2
‑
gfp的构建,细胞转染及细胞内抗体表达鉴定;其中a为空载体pcag
‑
gfp经pcr扩增后含有750bp左右的gfp条带,重组后的质粒pcagscfv4f5
‑
gfp经pcr扩增后含有1500bp大小的条带,b为荧光显微镜检测重组质粒的转染效率,重组质粒:lipofectamine
™ꢀ
2000=1:3,4μg/106细胞,转染条件下,荧光强度可达60%,c为转染后的细胞沉淀中均含有55kd大小的片段,转染48h后细胞中抗体表达量最高,d为western
ꢀ‑ꢀ
blot检测转染24,48,72h后细胞内scfv抗体的表达,转 染后的细胞沉淀中均含有55kd大小的片段,且转染48h后细胞中抗体表达量最高。
17.图2为全分子igg1重组质粒的构建,稳定表达细胞株的筛选,金分子抗体的表达纯
化及鉴定;其中a为重组全分子igg1质粒中的vl、vh、cl、ch1、vh2
‑
3、l和h基因片段的pcr鉴定,其中vl、vh、cl、ch1大小约为350bp,vh2
‑
3、l大小约为750bp,h大小为1500bp,b为2天后转染igg1质粒的细胞克隆(左),10天后能见到明显的克隆形成(右),c为western
‑
blot鉴定稳定表达细胞株中全分子igg1抗体蛋白的表达,细胞及培养上清中具有l和h抗体蛋白的表达,l大小为25kd,h大小为55kd,d为sds
‑
page鉴定亲和柱纯化后的抗体,l与h蛋白大小正确,且纯化后的蛋白纯度可达95%。
18.图3为全分子igg1抗体的特异性及亲和力鉴定;其中a为elisa鉴定igg1和scfv4f5病毒的结合活性,二者对病毒od450值相近,无明显的统计学差异,说明igg1和scfv4f5的结合活性相同,b为流式细胞术鉴定igg1对病毒的结合比例,可达95%(左),人igg抗体结合百分比为0.1%(右),c为biacorex100鉴定gg1抗体对病毒的亲和力常数kd值为4.47
×
10
‑
12
(左),scfv4f5为9.74
×
10
‑9(右)。
19.图4为细胞内和细胞外抗体及其组合对hpaiv感染小鼠的保护率;其中a为全分子igg1抗体对100tcid50a/jiangsu/1/2007(h5n1)感染小鼠的保护率,注射pbs组及2mg/kg组小鼠全部死亡,5mg/kg10mg/kg,15mg/kg存活率分别为16.67%、66.67%、100%,在15mg/kg用量时可达到100%的保护率,b为细胞内和细胞外抗体组合对500tcid50a/jiangsu/1/2007(h5n1)感染小鼠的保护率,当pcag
‑
scfv4f5
‑
gfp质粒用量0,5μg,10μg,20μg,30μg时,14天小鼠的存活率分别为16.67%、16.67%、33.33%、50.00%、66.67%。
20.图5为全人源抗体组合对病毒感染阻断后因子转录及表达水平的变化,病毒复制动力学的检测;其中a为elisa检测细胞因子ifn
‑
α,ifn
‑
β,ifn
‑
γ,il
‑
6,il
‑
10的变化,b为real
‑
timepcr检测细胞因子转录水平的变化,c为western
‑
blot实验检测凋亡蛋白bim,caspase3蛋白表达水平的变化,d为mdck细胞培养术检测病毒复制动力学测定。
21.图6为免疫组化检测存活小鼠和死亡小鼠肺组织损伤病理学变化,其中a为iihc检测存活小鼠肺组织中h5n1病毒的表达,b为iihc检测死亡小鼠肺组织中h5n1病毒的表达.相对于死亡小鼠肺组织,存活小鼠中h5n1病毒颗粒较少。
22.具体实施方式
23.下面的实施例可使本专业技术人员更全面地理解本发明,但不以任何方式限制本发明。
24.实施例1重组真核表达载体pcag
–
scfv1a2
‑
gfp的构建以含有抗体scfv1a2基因的pcomb3xss
‑
scfv1a2质粒为模板,pcr扩增pcomb3xss
‑
scfv1a2中的scfv1a2基因大小为750bp(图1:a),并同时在片段的两侧添加真核载体的克隆位点序列,产物与高效真核表达载体pcag
‑
gfp经酶切连接后筛选阳性重组子pcag
–
scfv1a2
‑
gfp。
25.图1:a显示,空载体pcag
‑
gfp经pcr扩增后含有750bp左右的gfp条带,重组后的质粒pcag
‑
scfv1a2
‑
gfp经pcr扩增后含有1500bp大小的条带,证明构建的pcag
‑
scfv1a2
‑
gfp质粒正确。
26.将构建好的重组载体pcag
–
scfv1a2
‑
gfp与脂质体lipofectamine
™
2000混合后转染人肺泡上皮a549细胞,转染后24
‑
72h,荧光显微镜检测重组质粒的转染效率,重组质
粒:lipofectamine
™
2000=1:3,4μg/106细胞,转染条件下,荧光强度可达60%(图1:b)。
27.分别在转染24,48,72h后收集细胞,ripa细胞裂解液裂解细胞后,利用western
‑
blot检测细胞内scfv抗体的表达,如图1:c所示,转染后的细胞沉淀中均含有55kd大小的片段,转染48h后细胞中抗体表达量最高,证明细胞内抗体能够正确表达。
28.实施例2全分子igg1重组质粒的构建扩增抗h5n1型高致病性禽流感病毒小分子scfv1a2抗体的可变区基因vl和vh,分别克隆于l链和h链的盒式真核表达载体pci
‑
rv220
‑
l和pci
‑
rv220
‑
h中,构建igg1真核表达载体。
29.构建的全分子igg1真核表达载体共转染cho
‑
dg44(dhfr
‑
/
‑
)细胞并通过g418抗药抑制和ht组分缺失的选择筛选igg1抗体高效表达的稳定细胞株。
30.利用western
‑
blot鉴定稳定表达细胞株中全分子igg1抗体蛋白的表达,如图2:c,cho稳定表达细胞及培养上清中具有l和h抗体蛋白的表达,l大小为25kd,h大小为55kd。
31.利用proteing亲和柱纯化,sds
‑
page鉴定,l及h链大小与预期相符,且纯化后的蛋白纯度可达95%(图2:d)。
32.初步筛选的稳定转染株进一步亚克隆,继续筛选,重复3次亚克隆过程,在亚克隆工程中,不断优化表达条件的高压选择,得到既能稳定表达又能大量表达的igg1抗体;经过几轮筛选,能够获得每100ml产20mg的蛋白表达量。
33.实施例3全分子igg1抗体特异性和亲和力的鉴定(1)利用elisa、细胞流式技术分析igg1抗体的特性以h5n1hapiv病毒包板,分别鉴定igg1和scfv1a2病毒的结合活性,图3:a说明igg1和scfv1a2对病毒od450值相近,无明显的统计学差异,说明全分子igg1和小分子scfv1a2的结合活性相同,未受fc片段的影响。
34.a/jiangsu/1/2007(h5n1)感染mdck细胞72小时后,分别加入igg1及商品化的人igg抗体,4℃避光孵育1小时,以含1%bsa的pbs洗涤3次后加入pe标记的抗人荧光二抗,4℃避光孵育30min;以含1%bsa的pbs洗涤4次后,细胞重悬于0.2ml4%多聚甲醛固定液中进行流式细胞仪检测。图3:b可见igg1对病毒的结合比例可达95%,商品化的人igg抗体结合百分比仅为0.1%,说明igg1结合活性较好。
35.(2)全分子igg1抗体的亲和力测定利用biacorex100测定抗体与h5n1型高致病性禽流感病毒糖蛋白ha结合的亲和力,在芯片上包被抗原ha1(杆状病毒
‑
昆虫表达系统表达并纯化),加不同浓度的抗体,测定亲和力,igg1抗体kd值为4.47
×
10
‑
12
,而scfv1a2仅为9.74
×
10
‑9(图3:c),说明igg1的亲和力较scfv1a2好。
36.实施例4全人源抗体组合pcag
‑
scfv1a2
‑
gfp和igg1在h5n1型禽流感病毒感染小鼠试验中的治疗作用(1)mdck细胞中和试验检测全分子igg1抗体对h5n1型hpaiv的中和作用利用reed
‑
muench方法测定病毒对mdck细胞的组织细胞半数感染量(tcid50),全分子igg1抗体倍比稀释(从2μg/ml开始2倍稀释,稀释12个梯度)后与100tcid50病毒混合,37℃孵育2h,然后加入到mdck细胞单层中,置37℃、5%co2培养箱培养3
‑
5d,细胞单层经充分洗涤、丙酮固定后,加入鼠源抗禽流感病毒核蛋白np单抗,弃上清,洗去未结合的抗体,加
hrp标记的抗鼠igg二抗,读取od
450
值,计算tcid50, 结果显示全分子igg1抗体对h5n1 型高致病性禽流感病毒的tcid50为10
‑3μg/ml,低于scfv1a2的0.01μg/ml。
37.(2)全人源中和性细胞内和细胞外抗体对hpaiv 感染小鼠的阻断作用6 周龄balb/c小鼠,随机分组,每组6只,鼻腔接种100 tcid50高致病性禽流感病毒a/jiangsu/1/2007(h5n1),实验组感染后12h、24 h、48h在小鼠腹腔注射不同浓度梯度的全分子igg1抗体,浓度梯度为2mg/kg,5mg/kg,10mg/kg,15mg/kg,对照组只在相同时间相同部位注射相同体积的pbs,每天观察、记录小鼠死亡情况,共14天,绘制小鼠生存率曲线。结果显示:当全分子igg1抗体用量从2mg/kg增加至15mg/kg时,对a/jiangsu/08
‑
6(h5n1)病毒感染小鼠的治疗保护率呈上升趋势,14天后注射pbs组及2mg/kg组小鼠全部死亡,5mg/kg,10mg/kg,15mg/kg存活率分别为16.67%、66.67%、100%;随着抗体用量的增加,对a/jiangsu/1/2007(h5n1) 病毒感染小鼠的保护率呈上升趋势,在15mg/kg用量时可达到100%的保护率(图4:a)。
38.(3)全人源中和性细胞内和细胞外抗体组合对hpaiv 感染小鼠的阻断作用6 周龄balb/c小鼠,随机分组,每组6只,鼻腔接种500 tcid50高致病性禽流感病毒a/jiangsu/1/2007(h5n1),感染后12h、 24 h、48h从腹腔注射15mg/kg的全分子igg1 抗体,同时从小鼠前肢注射不同浓度梯度的质粒pcag
‑
scfv1a2
‑
gfp 与 lipofectamine
™ꢀ
2000 混合液(v:v=1:3),实验组浓度梯度为5μg,10μg,20μg,30μg,对照组只在相同时间相同部位注射相同体积的pbs,每天观察、记录小鼠死亡情况,共14 天,绘制小鼠生存率曲线。结果显示:当pcag
‑
scfv1a2
‑
gfp 质粒用量0,5μg,10μg,20μg,30μg时,对a/jiangsu/1/2007(h5n1)病毒感染小鼠的治疗保护率呈上升趋势,14天小鼠的存活率分别为16.67%、16.67%、33.33%、50.00%、66.67%(图4:b), 当pcag
‑
scfv1a2
‑
gfp 质粒继续增加至50μg,对a/jiangsu/1/2007(h5n1) 病毒感染鸡胚的保护率并无上升趋势。
39.实施例5全人源抗体组合pcag
‑
scfv 1a2
‑
gfp和igg1在h5n1型禽流感病毒感染小鼠试验中的安全性(1) 细胞因子的检测:6 周龄balb/c 小鼠,随机分组,分别鼻腔接种500 tcid50高致病性禽流感病 毒a/jiangsu/1/2007(h5n1),感染后12h、24 h、48h 从腹腔注射15mg/kg的全分子 igg1 抗体,同时从小鼠前肢注射30μg的质粒pcag
‑
scfv
‑
gfp 与 lipofectamine
™ꢀ
2000 混合液(v:v=1:3),分别在首次注射抗体后2d,4d,8d,12d 及14d 收集存活的小鼠,每组随机抽取3只,收集小鼠血清,利用elisa法检测小鼠血清中ifn
‑
α,ifn
‑
β,ifn
‑
γ, il
‑
6,il
‑
10 等细胞因子的变化。图5:a可见:ifn
‑
α,ifn
‑
β随时间变化无明显差 异,ifn
‑
γ在第4天有升高,第8天达到最高点,12天后呈下调趋势。il
‑
6,il
‑
10 变化趋势与ifn
‑
γ相同,第4天有升高,第8天达到最高点,12天后呈下调趋势。说明抗体组合对ifn
‑
γ,il
‑
6,il
‑
10有一定的抑制作用。
40.(2) real
‑
time pcr 检测相关基因转录水平的表达变化:6 周龄balb/c 小鼠,随机分组,分别鼻腔接种500 tcid50高致病性禽流感病 毒a/jiangsu/1/2007(h5n1),感染后12h、24 h、48h 从腹腔注射15mg/kg的全分子 igg1 抗体,同时从小鼠前肢注射30μg的质粒pcag
‑
scfv
‑
gfp 与 lipofectamine
™ꢀ
2000 混合液(v:v=1:3),分别在首次注射抗体后2d,4d,8 d ,12d 及14d 收集存活的小鼠,每组随机抽取3只,收集小鼠外周抗凝血,利用淋巴细胞分离液,从小鼠外周抗凝血中分离外周血单核细胞(pmbc),提取rna,反转录为 cdna
后,real
‑
time pcr 检测ifn
‑
α,ifn
‑
β,ifn
‑
γ,il
‑
6,il
‑
10,绘制曲线, 记录基因表达水平的变化。 图5:b可见:基因转录水平的变化趋势与细胞因子检测相同,ifn
‑
α,ifn
‑
β随时 间变化无明显差异,ifn
‑
γ在第4天有升高,第8天达到最高点,12天后呈下调趋 势。il
‑
6,il
‑
10变化趋势与ifn
‑
γ相同,第4天有升高,第8天达到最高点,12天后呈下调趋势。说明抗体组合对ifn
‑
γ,il
‑
6,il
‑
10有一定的抑制作用。
41.western
‑
blot实验检测凋亡蛋白bim,caspase 3 蛋白表达水平的变化。如图5:c,bim蛋白在感染的第12,14天均有较高的表达水平,cleave的caspase 3蛋白则在 第4天表达最高,8及以后则无明显升高,说明抗体组合对细胞的凋亡有一定的抑制作用。
42.(3)病毒复制动力学测定:6周龄balb/c小鼠,随机分组,分别鼻腔接种500tcid50 高致病性禽流感病毒a/jiangsu/1/2007(h5n1),感染后12h、24h、48h从腹腔注射15mg/kg 的全分子igg1抗体,同时从小鼠前肢注射30μg的质粒pcag
‑
scfv
‑
gfp与lipofectamine
™
2000混合液(v:v=1:3),分别在首次注射抗体后2d,4d,8d,12d及14d收集存活的小 鼠,每组随机抽取3只,收集死亡和存活小鼠的肺组织。对不同感染时期小鼠肺脏,加入细 胞培养液,破碎离心后取上清液分别接种于mdck单层细胞,加盖营养琼脂,一周后结晶 紫染色,计数病毒斑,计算病毒滴度(log pfu/g),绘制曲线,观察病毒复制速度变化。如 图5:d病毒在注射抗体后24小时最高,可达到108,在重复给药后病毒滴度有所下降,第 14天后可降至102,说明抗体组合对病毒的复制增至有较好的抑制作用。
43.(4)cd8 t细胞活 力及凋亡的检测:处死小鼠,取新鲜脾脏,网筛磨成单个细胞,cfse标记,加入特异性的 pro5 mhc i pentamers,流式检测cd8 t细胞增殖信号,流式分选并测定cd8 t的凋亡比 例,绘制曲线,记录cd8 t细胞增殖及凋亡比例的变化,cd8 t第8天表达最高,12天及14天表达均有降低。
44.(5) 利用免疫组化检测存活小鼠和死亡小鼠肺组织损伤病理学变化。分别取存活小鼠和死亡小鼠肺组织,石蜡包埋,切片,脱蜡和水化后,抗原进行修复,每张切片加入抗 np 单克隆抗体及二抗后,苏木精复染,冲洗反蓝。切片经过梯度酒精脱水干燥,中性树胶封固,免疫组化染色结果及分析见图 6:相对于死亡小鼠肺组织(图6:b),存活小鼠中 h5n1 病毒颗粒较少(图6:a)。
再多了解一些
本文用于企业家、创业者技术爱好者查询,结果仅供参考。